 Mingshu Xiao
Mingshu Xiao Yueli Shi
Yueli Shi Sujing Jiang
Sujing Jiang Mengqing Cao
Mengqing Cao Weiyu Chen
Weiyu Chen Yun Xu
Yun Xu Zhiyong Xu
Zhiyong Xu Kai Wang
Kai Wang
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Oncol. , 29 November 2022
Sec. Cancer Imaging and Image-directed Interventions
Volume 12 - 2022 | https://doi.org/10.3389/fonc.2022.1039378
This article is part of the Research Topic Tumor Microenvironment Targeted Nanomedicine: A Feasible Strategy for Cancer Imaging and Theranostics View all 6 articles
Anti-angiogenesis therapy and immunotherapy are the first-line therapeutic strategies for various tumor treatments in the clinic, bringing significant advantages for tumor patients. Recent studies have shown that anti-angiogenic therapy can potentiate immunotherapy, with many clinical trials conducted based on the combination of anti-angiogenic agents and immune checkpoint inhibitors (ICIs). However, currently available clinical dosing strategies and tools are limited, emphasizing the need for more improvements. Although significant progress has been achieved, several big questions remained, such as how to achieve cell-specific targeting in the tumor microenvironment? How to improve drug delivery efficiency in tumors? Can nanotechnology be used to potentiate existing clinical drugs and achieve synergistic sensitization effects? Over the recent few years, nanomedicines have shown unique advantages in antitumor research, including cell-specific targeting, improved delivery potentiation, and photothermal effects. Given that the applications of nanomaterials in tumor immunotherapy have been widely reported, this review provides a comprehensive overview of research advances on nanomaterials in anti-angiogenesis therapy, mainly focusing on the immunosuppressive effects of abnormal tumor vessels in the tumor immune microenvironment, the targets and strategies of anti-angiogenesis nanomedicines, and the potential synergistic effects and molecular mechanisms of anti-angiogenic nanomedicines in combination with immunotherapy, ultimately providing new perspectives on the nanomedicine-based synergy between anti-angiogenic and immunotherapy.
Angiogenesis is the process of neovascularization from existing peripheral blood vessels, providing essential nutrients for solid tumor growth and a pathway for malignant metastasis, considered one of the hallmarks of cancers (1). This process involves the joint participation of endothelial cells, other stromal cells and the extracellular matrix (ECM) (2–4). Unlike normal tissue vasculature, the tumor vasculature exhibits abnormal structure and function, including a disordered vascular network, narrowed lumen, increased permeability and other defects, which leads to internal tumor hypoxia and low PH, inefficient drug delivery and reduced sensitivity to radiotherapy in the clinic (5–7).
Inhibition of angiogenesis is widely acknowledged as a promising antitumor strategy, exhibiting antitumor effects in various solid cancers. However, excessive inhibition of tumor angiogenesis tends to cause vascular degeneration, reduce drug delivery, and induce drug resistance (8, 9). In addition, treatment with single anti-angiogenic drugs tends to cause compensatory expression of other pro-angiogenic factors or pathway activation, leading to drug resistance (10–13). An increasing body of evidence from recently published studies suggests that short-term inhibition of tumor growth can induce normalization of tumor vascular structure and function, increase tumor vascular perfusion capacity while alleviating intratumor hypoxia levels, and improve the tumor suppressive immune microenvironment, which has the potential to potentiate the efficacy of radiotherapy and immunotherapy (14–16).
In recent years, nanomedicine has achieved significant progress, especially for targeted drug delivery and precision interventions (17, 18). Nanomedicines have unique advantages in terms of precision and multi-targeted interventions while showing enhanced effects when combined with other antitumor treatment strategies (19–21). This review provides a comprehensive overview of the characteristics and molecular basis of tumor vascular structure and function and describes the impact of tumor vascular abnormalities on the tumor immune microenvironment. Furthermore, we summarize the latest research advances of current anti-angiogenic nanomedicines and their effects on the immune microenvironment and immunotherapy.
Tumor angiogenesis is the biological process by which tumor cells and tumor microenvironment (TME) induce microvascular growth and establish blood circulation for tumor tissue (22). Tumor vasculature provides oxygen and nutrients for tumor tissues while carrying away metabolic wastes and carbon dioxide, promoting tumor growth and providing a pathway for tumor metastasis (23). It has been established that almost all solid tumors cannot grow beyond the size of 1-2 mm3 without the support of the vascular system, which indicates that neovascularization is a prerequisite for the sustained growth of tumors (24). Sprouting angiogenesis is the main and classical way of tumor angiogenesis and is predominantly discussed in this paper. This process refers to the generation of microvessels by endothelial cells in a “budding” manner based on the original blood vessels. The main steps include: ① degradation of vascular basement membrane and activation of endothelial cells; ② endothelial cell migration and proliferation; ③ endothelial cell formation of luminal structures and capillaries; ④ generation of new basement membrane and recruitment of pericytes to build vessel wall, formation of mature vessels and extension into solid tumors (25, 26).
Tumor angiogenesis is a complex process involving multiple mediators, mainly composed of tumor cells, tumor stromal cells (including endothelial cells, inflammatory cells, pericytes, fibroblasts, smooth muscle cells, etc.), extracellular matrix (ECM), and a variety of cytokines (27–29). All these effectors constitute the regulatory network of tumor angiogenesis. Current evidence suggests that the imbalance between pro- and anti-angiogenic factors is the key to initiating angiogenesis (30). Under physiological conditions, pro- and anti-angiogenic factors are in balance, and blood vessels remain quiescent and rarely form new branches. During tumor angiogenesis, the pro-angiogenic signals is hyperactivating, disrupting the balance and inducing angiogenesis (24, 31). Many factors contribute to the disruption of homeostasis, such as carcinogenic mutations, hypoxia, low pH, tumor-associated inflammation, recruitment of immune cells, and nutritional deficiencies. Targeted intervention of these factors is the key strategy for nano-targeted anti-vascular therapy.
Angiogenesis-stimulating factors represent a group of mediators, including growth factors, bioactive lipids, ECM degrading enzymes, cytokines, adhesion molecules and a variety of small molecule nucleic acids (Table 1). In recent years, the role of adhesion molecules in tumor angiogenesis has received increasing attention (32). Among the five major classes of adhesion molecules identified so far, it has been thought that the integrin family are the most important molecules for the process of angiogenesis by mediate cell-ECM adhesion (33).However, several other adhesion proteins, including vascular endothelial cell calcium adhesion protein (VE-cadherin), endothelial cell adhesion molecule 1 (PECAM1), and intercellular adhesion molecule 1 (ICAM1), vascular cell adhesion molecule-1 (VCAM-1), have been reported to mediating the adhesion of immune cells in tumor blood vessels, which is the key process for tumor immune cells to enter the stroma of tumor tissue and performing immunocidal functions (34–36). Understanding the mutual relationship among these regulatory molecules and their role in promoting angiogenesis is crucial for elucidating the mechanism of tumor angiogenesis and developing effective anti-angiogenic therapies.
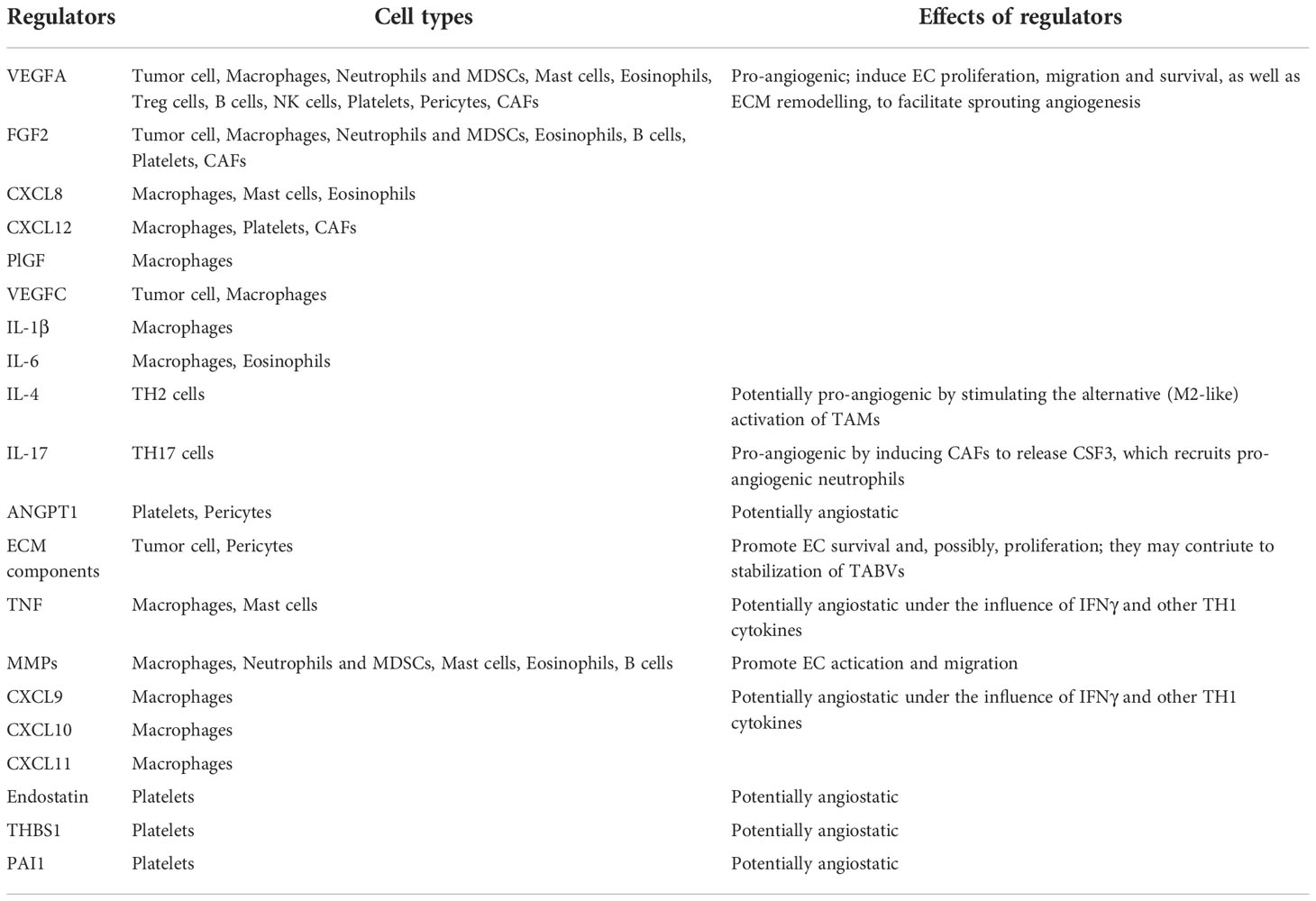
Table 1 The sources and effects of the sources tumor vasculogenesis regulators.
Unlike physiological angiogenesis, tumor angiogenesis is a rapid and aberrant process, resulting in newly formed vessels that are often immature and have abnormal structures and functions (5). Tumor vessels are often hyperplastic, tortuous, dilated, overlapping, and exhibit disordered branching. The newly formed tumor microvessels lack normal structures, and are sinusoidal, striped, and thin-walled, composed of only one layer of endothelial cells. In addition, tumor vessels often exhibit high heterogeneity and distribution (Figure 1) (6, 14, 37).
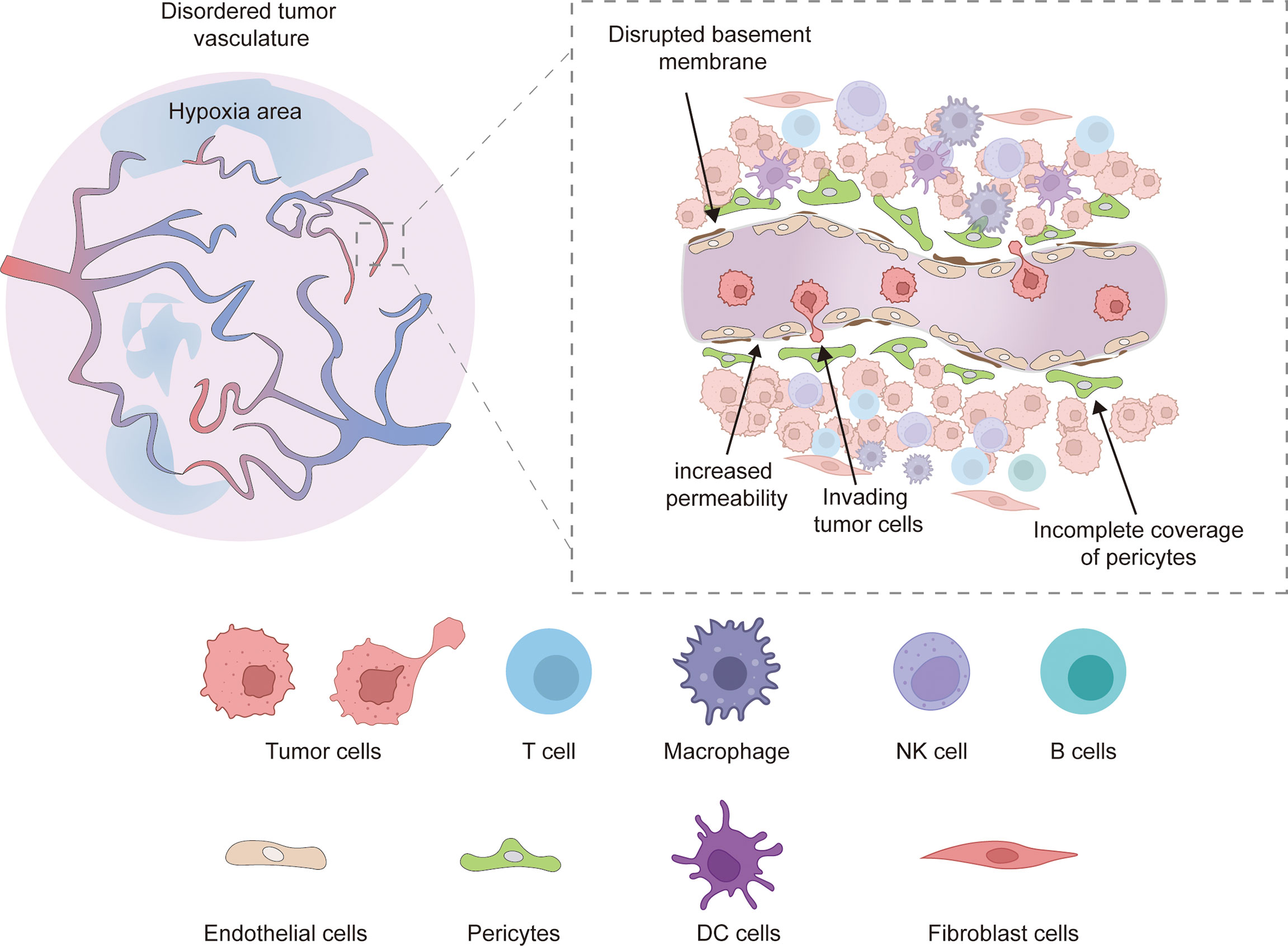
Figure 1 Abnormal tumor vessels are characterized as hyperplastic, tortuous, dilated, overlapping, and exhibit disordered branching, which leads to local tissue hypoxia. Besides, Defects in vascular branching, pericyte coverage, and basement membrane coverage were also observed in disordered tumor vasculature. The abnormal vessel structure and components increase vessel permeability, leading to increased tumor metastasis, and the tumor endothelial cells showed decreased adhesion molecules expression (ICAM1, VCAM1), resulting in reduced adhesion of immune cells on the surface of tumor vessels and infiltration in the tumor microenvironment. Hypoxia and low PH conditions also severely reduce the normal immune function of tumor immune cells.
It is widely thought that pericytes overlying endothelial cells play a regulatory role in the developmental stability and maturation of blood vessels. In tumor vessels, pericytes are detached from ECs, resulting in dysregulated flow characteristics and increased permeability. An incomplete vascular basement membrane (BM) is a feature of tumor vasculature. The BM of tumor vessels exhibits heterogeneity in thickness, fracture or absence and is loosely connected to endothelial cells and pericytes (15). In addition, TME leads to abnormal EC morphology and function. Compared with normal ECs, tumor-associated ECs change from a quiescent state to an activated state, showing high proliferation and migration, overlapping growth, loss of polarity, loose connections, and enlarged cell space (38). Meanwhile, the absence of a smooth muscle layer is also an abnormal indication of tumor vasculature (39). These structural and cellular abnormalities lead to abnormal vascular function; the neoplastic vessels exhibit lower blood flow, susceptibility to leakage, impaired vessel perfusion and increased vascular permeability, which lead to an abnormal TME: higher interstitial fluid pressure, hypoxia, acidosis and necrosis (40).In addition, restricted blood flow and high tumor interstitial pressure result in impaired drug delivery and reduced infiltration of immune effector cells (14). Meanwhile, hypoxia and acidic microenvironments impair the antitumor function of infiltrating immune cells (41). The abnormal vascular function makes tumors more susceptible to growth and metastasis and hinders the implementation of efficient tumor therapy.
Immunotherapies such as immune checkpoint blockade are nowadays increasingly used for tumor treatment. The key to efficient immunotherapy is alleviating tumor immunosuppression and restoring the immune response (42). Tumor vascular abnormality is a major reason for the immunosuppressive TEM. As mentioned above, the abnormal structure and function of tumor blood vessels lead to interstitial hypertension and hypertonia and form a physical barrier, resulting in a reduced recruitment of immune effector cells (43). Besides, tumor-associated ECs express lower levels of adhesion molecules, such as intercellular adhesion molecule 1 (ICAM1), which prevents immune cells from adhering to endothelial cells, interferes with the trafficking of immune cells, and leads to a decrease in immune cell infiltration (especially T cell) (44, 45). Besides, upregulation of immune checkpoints such as PD-L1/2 in tumor-associated ECs leads to decreased recruitment and activation of cytotoxic T lymphocytes (CTLs) (46). It has been shown that Fas ligand (FasL), highly expressed in tumor-associated ECs, selectively mediates CTL but not Treg apoptosis (47).
Hypoxia is also a major driver that favors an immunosuppressive TME. Hypoxia causes immunosuppression via five mechanisms: (1) promoting the accumulation of immunosuppressive cells in tumor tissues, such as Treg, myeloid-derived suppressor cells (MDSCs), tumor-associated macrophages (TAMs), etc.; (2) promoting the production and release of immunosuppressive factors such as TGFβ, IL-10, Arg1, VEGFA (3) upregulating the expression of immunosuppressive molecules, such as PD-L1 on tumor cells, macrophages; PD-1 on CD8 + T cells; TIM and CTLA4 on MDSCs, Tregs, DCS and TAMs (4) inducing TAM to immunosuppressive M2 polarization phenotype which inhibits the maturation of DC cells and leads to the accumulation of immunosuppressive metabolites such as adenosine and lactate by changing the metabolism of tumor cells. (5) Attenuating survival, cytotoxicity and migratory activity of immune effector cells such as CD8 + cytotoxic T cells, CD4 + T cells and natural killer NK cells (48, 49). In addition, acidosis is a trigger for immunosuppression, as it reduces IFN-γ secretion by M1 macrophages and increases IL-1β secretion, inhibits T cell proliferation, promotes NK and T cell apoptosis, and inhibits CD8+ T cell and NK cell production of effector molecules (granzyme B, perforin, etc.). It also stimulates neovascularization by delaying the apoptosis of neutrophils and promoting infection (50, 51).
Excessive amounts of pro-angiogenic factors also contribute to the immunosuppressive TEM. For example, high-level VEGF promotes CTL depletion by upregulating the expression of immune checkpoint molecules (PD-1/PD-L1, CTLA4, LAG3, TIM3) (52). VEGF has also been associated with immune escape by inhibiting the differentiation and function of NK cells. In addition, VEGF triggers tumor evasion from the immune system by inducing Tregs and MDSCs proliferation (53). Growing evidence suggests that VEGF can promote M2 polarization of TAMs, inhibit the differentiation and maturation of DC cells as well as the antigen-presenting ability of mature DCs and reduce T cell infiltration (54, 55). As mentioned above, there is a complex interaction between the tumor vasculature and the tumor immune microenvironment. Normalizing the tumor vascular system increases immune cell infiltration and restores the antitumor capacity of immune cells, suggesting it is a potentially effective anti-cancer strategy (56).
Angiogenesis represents an attractive target for cancer therapies because of the vital effects of tumor vessels in tumor progression. Due to the lack of specific biomarkers and resistance associated with a combination of the traditional approach with anti-angiogenic drugs, the efficacy during clinical practice remains unclear. Nanotechnology provides a new approach whereby nanomaterials bind to the potential target and release anti-angiogenic drugs at specific sites (57, 58). The nano drug delivery systems can reduce adverse effects on non-target healthy tissue, provide accumulation into the target tissue and protect nucleic acids, including small interfering RNA (siRNA) and micro RNA (miRNA), from rapid degradation in the body (59). Mechanisms and targets of nanomaterial-based anti-angiogenic therapy are as follows (Table 2).
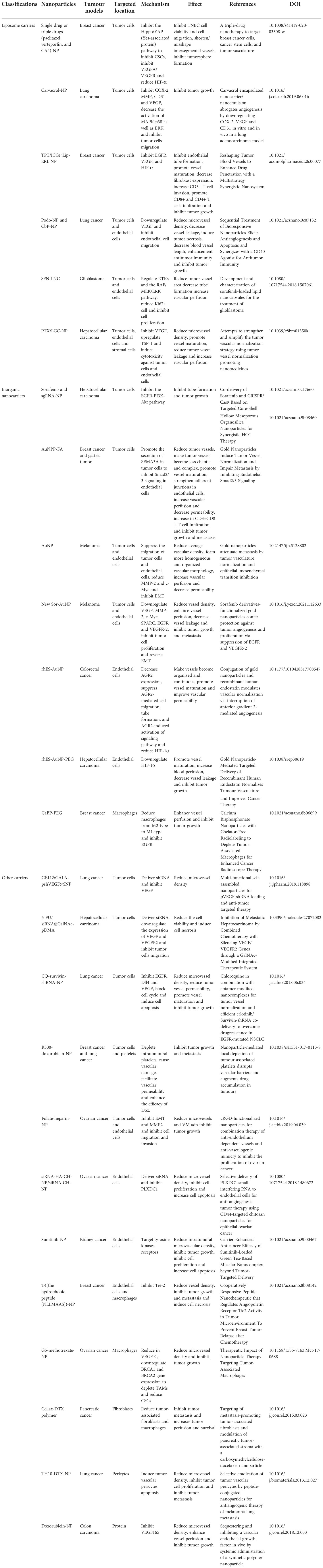
Table 2 Mechanisms and targets of nanomaterial-based anti-angiogenic therapy.
Vascular endothelial growth factor A (VEGFA) is a crucial regulator of angiogenesis progression. It binds to VEGFR2 in vascular endothelial cells to stimulate the proliferation and trigger endothelial cell migration through the RAS–RAF–MAPK (mitogen-activated protein kinase)–ERK (extracellular signal-regulated protein kinase) signaling pathway (27). Accordingly, VEGF gene expression regulation has attracted significant interest in anti-angiogenic cancer therapy. In vitro and in vivo experiments showed that plasmid DNA containing a small hairpin RNA (shRNA) expression cassette with epidermal growth factor receptor (EGFR)-specific binding ligand GE11 and pH-sensitive fusogenic peptide GALA could target tumor cells and induce endocytosis, whereby nanoparticles (GE11&GALA-pshVEGF@SNPs) entered tumor cells to inhibit VEGF expression, reduce tumor angiogenesis, and inhibit tumor growth (60). The RNA interference (RNAi) system based on gene silencing mechanisms is a powerful tool in antitumor angiogenic therapy. SiRNA has also been applied for the regulation of VEGF expression. Asialoglycoprotein receptor (ASGPR) is a mammalian lectin specifically expressed on the hepatocytes. N-acetyl-galactosamine (GalNAc) residues can mediate the specific cellular uptake of the nanoparticle by targeting ASGPR-expressing liver cells as one of the specific ligands for ASGPR. Due to the nanoparticles’ high compatibility and low cytotoxicity, they were modified with GalNAc to enter hepatocellular carcinoma, and siRNA was used to regulate VEGF transcription (61). However, the limitations of non-specific absorption and low efficiency of cellular uptake reportedly hamper their clinical applications. Fortunately, a method was proposed to tackle this problem by generating siRNA in situ through enzyme-free DNA amplification (62). Moreover, a new system containing superparamagnetic iron oxide nanoparticles, calcium phosphate (CaP) and PEG-polyanion block copolymers could improve the efficiency of siRNA transportation and promote its aggregation (63). In addition, blocking VEGF rather than influencing gene expression in vivo can be an anti-angiogenic approach. VEGFA165 is thought to be the most frequently expressed isoform in tumors and is the most physiologically relevant VEGFA isoform (27). Synthetic copolymer nanoparticles (NPs) were synthesized as the protein affinity reagents (PARs) that bind with vascular endothelial growth factor (VEGF165) to inhibit angiogenesis in vivo (64)
However, anti-angiogenic therapy through inhibition of the VEGF pathway has its limitations. In this respect, no Phase III clinical trial has shown anti-angiogenic benefits during glioblastoma therapy. Clavreul et al. considered it might be attributed to VEGF-independent angiogenesis, induction of tumor invasion and inefficient antiangiogenic factor delivery to the tumor (65). CD11b+Gr1+ cells, including neutrophils, macrophages, and myeloid-derived suppressor cells, promote growth factor granulocyte colony-stimulating factor (G-CSF) and promote myeloid cell-dependent angiogenesis via secreting protein Bv8. This angiogenic pathway bypassing VEGF represents the response mechanism to anti-VEGF therapy (65). Besides, cancer stem cells (CSCs) can drive tumor drug resistance. Interestingly, CSCs are a small subpopulation of cells with stem cell-like characteristics that can self-renew, divide asymmetrically to give rise to daughter cells and modulate the vascularization of the tumor by promoting the expression of HIF-1, VEGF and SDF-1/CXCL12 (66). Antivascular therapies are known to drive CSCs propagation and lead to the appearance of resistance mechanisms (67). Therefore, reducing CSCs in antivascular therapy can be applied in nanomaterial-based anti-angiogenic therapy. In triple-negative breast cancer (TNBC), encapsulation of paclitaxel, verteporfin, and combretastatin (CA4) significantly reduces CSCs and gene expression of VEGFA as well as the VEGF receptor (68). In ovarian cancer, tumor-associated macrophages can be selected by binding G5-MTX nanoparticles with folate receptor-2(FOLR2) which is highly expressed in TAMs. These nanoparticles can overcome resistance to anti-VEGF-A therapy inhibition of VEGF-C and BRCA1 gene expression for depletion of TAMs and the reduction of CSCs. Also, through coculture of TAMs and tumor cells, a mild increase in sphere number of tumor cells suggested that TAMs promoted stemness. In other words, depleting TAMs could reduce stemness (69).
Besides the VEGF pathway, multiple signaling pathways are related to nanomaterial-based anti-angiogenic therapy. It has been reported that the EGFR-PI3K-Akt pathway can be regulated at the gene level through CRISPR/Cas9 systems with a chitosan nanodelivery system to recognize the HepG2 cell adhesion molecules (CAMs) during the treatment of hepatocellular carcinoma. Tube formation of HUVEC cells was significantly inhibited after NPs treatment in vivo (70). In addition, it has been reported that Tyrosine kinase with immunoglobulin and epidermal growth factor homology-2(Tie-2) is expressed in endothelial cells, and tie-2 positive macrophages (TPM) promote tumor vascular reconstruction and chemoresistance through the angiopoietin (ANG)/Tie2 signaling pathway. Researchers designed dual-responsive amphiphilic peptide (mPEG1000-K(DEAP)-AAN-NLLMAAS) to modify the small peptide T4 (NLLMAAS) to protect T4 and target the acidic tumor microenvironment. T4 released by the ultimate nanoformulation (P-T4) could interact with tie-2 to inhibit the phosphorylation of downstream molecule focal adhesion kinase, which is relevant to migration and tubule formation (71).
As mentioned above, tumor endothelial cells (TEC) have characteristics that lead to leakiness and chaotic blood flow of tumor vessels. There are different markers, including EGFR, Pax2 and CXCR7 in TEC, which can bind with numerous ligands, such as peptides, nucleic acids aptamers, sugars and cationic charged materials. These markers can be harnessed to distinguish from normal endothelial cells and target TEC (72, 73). Nanomaterials can be loaded with antiangiogenic drugs with different effects. Accordingly, target tumor vessel therapies can be classified into two categories: anti-angiogenic agents (AAs) and vascular disrupting agents (VDAs) (74).
As for the target of vascular endothelial cells, the VEGF signaling pathway has drawn much attention. Tumor endothelial marker 7(TEM7), referred to as PLXDC1, has a plexin-semaphorin-integrin (PSI) domain. The PSI domain exists in semaphorin, and its receptor acts as a vascular endothelial growth factor (VEGF) receptor in endothelial cells (75). In a recent study, tumor endothelial cell CD44 receptor was specifically targeted by hyaluronic acid (HA), and angiogenic gene PLXDC1 was silenced by siRNA by combining with chitosan (CH) for endocytosis. Thus HA-chitosan- nanoparticle/siRNA system provides a new approach for anti-angiogenesis tumor therapy at the transcriptional level (76).
Besides, interleukin-8 (IL-8) is another pro-angiogenic factor activated by the VEGF pathway. It plays a role in the tube formation of endothelial cells. Poly (ethylene glycol)-conjugated epigallocatechin-3-O-gallate (PEG-EGCG) loaded with sunitinib has been reported to gather in tumor tissue. Moreover, Sunitinib-loaded micellar nanocomplex (SU-MNC) inhibits the mRNA and protein expression of IL-8 in endothelial cells to inhibit proliferation and migration (77).
Multimodal antivascular nanodevice of AAs and VDAs for cancer therapy has been reported to maximize the therapeutic potential of antivascular therapeutics. Some nanodevices have additional anti-angiogenic capabilities, such as photothermal therapy (PTT), which create local high-temperature conditions via gold nanorod and photodynamic therapy (PDT), producing toxic reactive oxygen species via photosensitizers under near-Infrared light conditions (78, 79). Moreover, the precise localization characteristics of nanoparticles promote the efficiency of PTT and PDT. The ferritin nanocage (Fn) targeting CGKRK peptides in the tumor loaded with “556-Ph”, a new type of metalla-aromatics complex of NIR-absorbing organic agent, yielded a better performance with PTT and PDT (80). Furthermore, combining PTT, PDT and VDA to destroy blood vessels can achieve better results. Nanotechnology can be used to develop a vascular disrupting agent that can be released before anti-angiogenic agents. VDAs can decrease the number of vessels by destroying the tumor’s neo-formed vasculature and ensure nanoparticles can more efficiently target specific regions (67).
Matrix metalloproteinases (MMPs), abundant in the tumor extracellular matrix (ECM), can decompose the extracellular matrix and disrupt the tumor microenvironment to promote the metastasis of cancer cells and induce angiogenesis. Moreover, they can biologically activate VEGF and induce endothelial cell motility (81). Therefore, MMPs are also regarded as another potential therapeutic target. Paclitaxel (PTX) nanocrystals were prepared with MMP-sensitive β-casein/marimastat (MATT) and PTX to treat breast cancer metastasis. These complexes could target tumor cells and the local tumor microenvironment and suppress the activities of MMP-2 and MMP-9 in vivo (82). In lung cancer, integrating chemotherapy drug carriers with Versatile polypeptide-LinTT1-Pvgli-Tat (LPT) increased tumor-penetrating capacity, reduced MMPs, inhibited tumor growth and decreased blood vessel density (83). A new nanomaterial called carvacrol nanoemulsion (CN) yielded anti-angiogenesis effects at multiple stages. CN influenced COX-2 to decrease MMPs and VEGF levels and could bind with the glycine residue of VEGF and the allosteric area of CD31 to disrupt adhesion between endothelial cells. In addition, CN could block MAPK and NF-κb pathways to interfere with the effect of VEGF and suppress inflammation to reduce angiogenesis, respectively (84).
Platelet aggregation is induced by tumor cells, tumor-infiltrating macrophages, and tumor endothelial cells by activating the coagulation pathway. Tumor-derived ADP, cathepsin B, and matrix metalloproteinases also play a crucial role. Furthermore, planets promote angiogenesis through microRNA, lipids, platelet-derived microparticles, and surface receptors (85). Recent reports suggest that platelets act as a membrane in nanoparticles and prevent tumor-specific immunological recognition (86). On the other hand, nanoparticles loaded with platelet inhibitors have been designed for tumor therapy. For example, a new nanoparticle coated with MMP2-cleavable peptides for targeted aggregation of MMP2 in tumor tissue contains R300 that can bind to platelet receptors and deplete platelets (87). Moreover, it can be used to increase drug permeability. However, more emphasis should be placed on bleeding complications in blocking specific tumor-platelet interaction sites (88).
Pericytes, perivascular cells associated with resistance to anti-VEGF therapeutics, play a critical role in tumor angiogenesis. Guan et al. developed new nanoparticles (TH10-DTX-NP) conjugated with TH10 peptide (TAASGVRSMH) and loaded with docetaxel. TH10 peptide could bind with NG2 receptors, highly expressed on the pericytes, and DTX could induce pericyte apoptosis. Consequently, it was shown that TH10-DTX-NP-induced pericyte apoptosis could decrease microvessel density and inhibit metastases in melanoma therapy (89).
Over recent years, it has been found that vasculogenic mimicry (VM) of tumor cells, an alternative circulatory system, plays a crucial role in tumor angiogenesis. Highly aggressive tumor cells can form vessel-like structures by mimicking endothelial cells. These structures can also provide sufficient blood supply and nutrients to tumors (90), providing an effective nanomaterial-based anti-angiogenic strategy. Moreover, CRGD was conjugated with heparin and folic acid to form two nanoparticles (CRGD-NPS1 and CRGD-NPS2). CRGD-NPS2 could regulate the MMP2/Laminin 5c2 signal pathway to inhibit endothelium-dependent vessels (EDV) and tumor cell-mediated VM in selected ovarian cancer cells capable of forming VM channels (91).
Normalization of tumor vasculature reportedly improves the effect of antitumor therapy by correcting structural abnormalities of tumor vessels and restoring normal function to a certain extent. This finding suggests that sustaining the balance between pro-angiogenic and anti-angiogenic factors is essential to decreasing micro-vessel density, improving pericyte coverage and regulating vascular permeability and perfusion. As a result, normal vessels relieve hypoxia to inhibit tumor invasion and recurrence (Figure 2). Moreover, vascular normalization is conducive to subsequent treatment, including chemotherapy (92), radiotherapy (93) and immunotherapy (94). Importantly, nanotechnology offers an excellent platform to improve the bioavailability and stability of drugs, reduce side effects and achieve combination therapy. Hence, various nanomedicines have been applied to target abnormal tumor vessels for more efficient therapy (95).
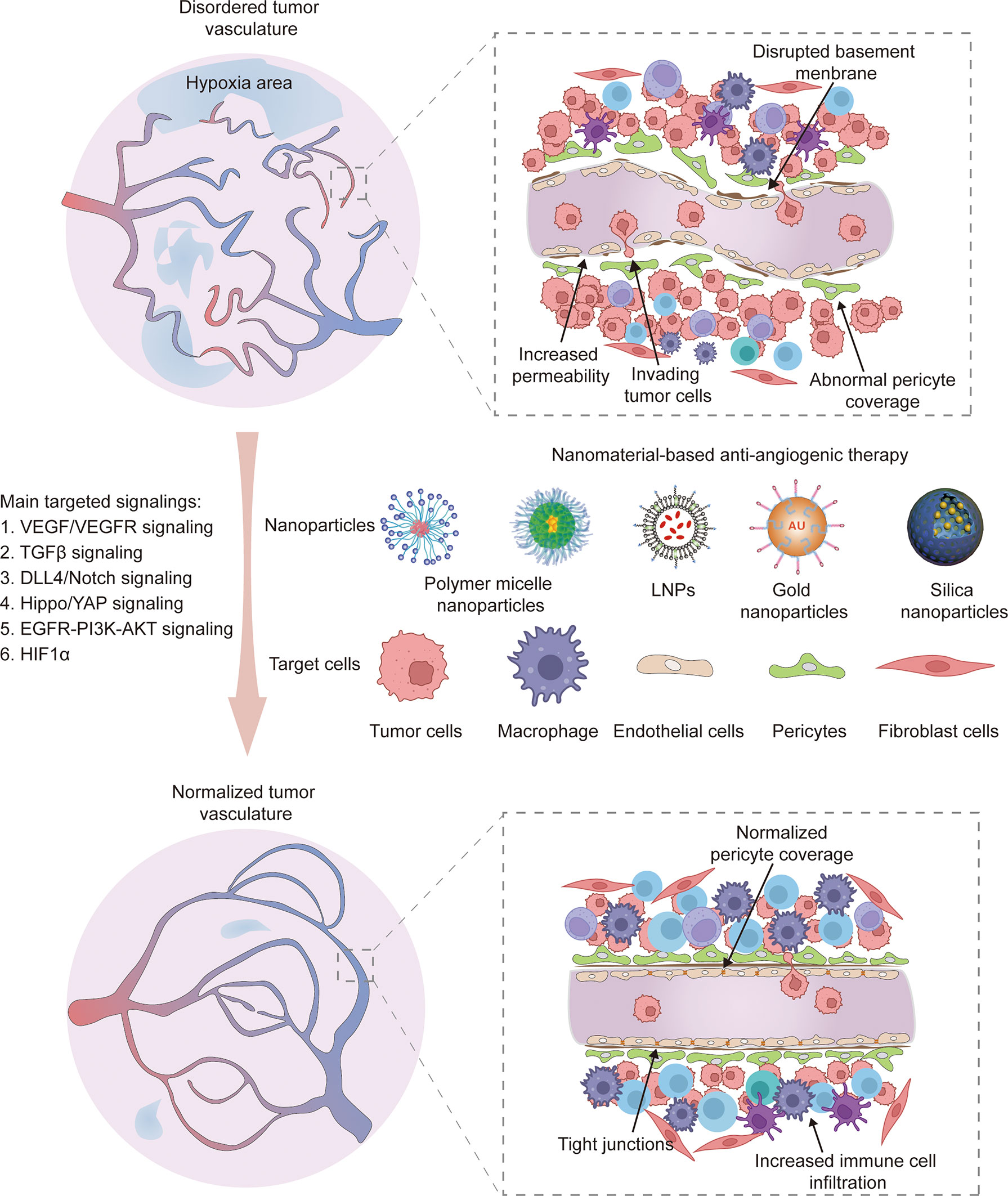
Figure 2 The normalization of tumor vasculature restores normal vessel function like decreasing micro-vessel density, improving pericyte coverage, increasing endothelium tight junctions and promoting immune cell infiltration. Various nanoparticle carriers including polymer micelle nanoparticles, lipid nanoparticles (LNPs), gold nanoparticles and silica nanoparticles have been applied to target various cell types, such as tumor cells, macrophages, endothelial cells, pericytes and fibroblast cells. The main targeted signaling are VEGF/VEGFR, TGFβ-Smads, DLL4/Notch, Hippo/YAP, EGFR-PI3K-AKT and HIF1α.
Epithelial-mesenchymal transition (EMT) is a process by which epithelial cells lose their adhesiveness and gain increased motility, contributing to VM formation and tumor cell metastasis. It can be stimulated by many factors, including c-myc, MMP-2 and HIF-α (96). Over the years, gold nanoparticles(AuNPs) have been applied in nanomaterial-based vascular normalization therapy of breast cancer (97), melanoma (96, 98), liver cancer (99) and metastatic colorectal cancer (100). Intriguingly, AuNPs could inhibit MMP-2 and c-Myc to suppress EMT and the migration of HUVECs (96). Fan Pan et al. conjugated gold nanoparticles and recombinant human endostatin to modulate vascular normalization in metastatic colorectal cancer. They found that NPs inhibited EMT via interruption of anterior gradient 2 (AGR2)-mediated angiogenesis, a latent tumor angiogenesis factor regulated by hypoxia-induced factor-1 (HIF-1) (100). Changes in tumor vessels after AuNPs vascular normalization therapy include a decrease in vascular density, permeability and hypoxic area vascular density and increased vascular perfusion, pericyte coverage of tumor and α -smooth muscle actin (α -SMA), an index of vessel maturation (96–100).
It is well-established that TAMs play a pivotal role in tumor angiogenesis through VEGF and other pathways, which leads to abnormal tumor vasculatures (101). TAMs are generally classified into immunosuppressive M2-type and immunosupportive M1-type. M2-type macrophages represent the dominant type of TAMs related to tumor growth, progress and metastasis. Furthermore, myeloid cell infiltration and differentiation are triggered by a hypoxic microenvironment; thus, more TAMs are recruited to the tumor site, which leads to poor therapeutic outcomes (102). Tian et al. designed calcium bisphosphonates (CaBP-PEG) nanoparticles that gather in tumor tissue through enhanced permeability and retention (EPR). In breast cancer treatment, NPs reduced macrophages from M2-type to M1-type and blocked the TAMs-stimulated angiogenesis signaling pathway. The dilation of tumor blood vessels and saturated oxygen indicated normalization of tumor vasculature and relief from hypoxia (103).
Current evidence suggests that tortuous and highly permeable tumor vessels cause accumulation of vascular macromolecules in tumor tissues and high interstitial fluid pressure (IFP), which prevents drug penetration (104). Furthermore, high IFP can aggravate the abnormal vascular network and hypoxia environment (105). Sirin Yonucu et al. demonstrated that tumor vascular normalization could restore intravascular pressure gradients and prune non-functional vessels by building a mathematical model and analyzing it with cases. Peripheral aggregation of drugs as micelles, nanoprobes and liposomes was changed into aggregation in the tumor center (104). Hence, vascular normalization and regulation of IFP decrease tumor repopulation caused by heterogeneous drug accumulation and increase the probability of treatment success. Hyaluronic acid (HA), a key component in ECM, is highly expressed in tumor tissue. Hyaluronidase (HAase) has been used to decompose HA and decrease IFP to enhance chemotherapy drug penetration for many years (106). Jingjing Jiao et al. designed the nanoparticle (PM@HAase-mPEG), combining a porphyrin-based metallacage, HAase and DSPE-mPEG2000 to normalize blood vessels, relieve hypoxia and promote the cellular accumulation of drugs. The results indicated that PM@HAase-mPEG treatment could increase the density of blood vessels, blood perfusion and oxygen content. In addition, normalizing tumor vasculature via HAase could enhance the efficacy of PDT therapy for breast cancer (105).
Tumor-associated fibroblasts (TAFs), derived from the “activation” of tumor stromal cells such as fibroblasts, can create tension in matrix components such as collagen. Mechanical vascular compression contributes to the abnormal structure of tumor vessels and hindrance of drug penetration (107). Moreover, depletion of TAFs has been shown to decompress vessels and improve perfusion (108).
Interestingly, nanotechnology can be harnessed to promote long-term blood circulation and high accumulation of drugs in tumor tissue. Mark J. Ernsting et al. designed Cellax-DTX polymer, a conjugate of docetaxel (DTX), polyethylene glycol (PEG), and acetylated carboxymethylcellulose. Cellax-DTX reduced CAF content and increased duct luminal area and blood perfusion during the treatment of pancreatic ductal adenocarcinomas (109). Furthermore, TAF-targeted normalization of tumor vasculature could improve nanomaterial-based tumor drug delivery. Celecoxib, a selective COX-2 inhibitor, was designed with PTX-loaded micelles to reduce TAFs and normalize tumor vessels. The mechanism of TAF depletion involved suppression of fibroblast proliferation and activation stimulated by FGF-2 and transforming growth factor beta-1 (TGF-beta1) by inhibiting ERK1/2 phosphorylation, inhibition of the CXCL12/CXCR4 axis, induction of G1-S cell cycle arrest and apoptosis of TAF and reprogramming of TAF into normal fibroblasts. Consequently, microvessel density and increased pericyte coverage on endothelial cells was reduced after lung cancer treatment. Furthermore, it increased the in vivo delivery of micelles and improved the therapeutic benefits of PTX-loaded micelles (110). The interaction of nanomaterials and vascular normalization demonstrates that their combination may have great potential to improve tumor treatment.
The abnormal structure and function of tumor vessels disrupt the tumor immune microenvironment. On the one hand, abnormal tumor vessels decrease antitumor infiltrating lymphocytes via high IFP, downregulation of adhesion molecules and hypoxia, which activates inhibitory signaling pathways for antitumor immune response. On the other hand, the hypoxic tumor microenvironment can recruit Treg and promote TAM polarization to immunosuppressive M2-like phenotype to participate in immune evasion (94). Moreover, normalization of tumor vasculature is beneficial in rescuing these immunosuppressive outcomes. Changes in immune cell infiltration in nanomaterial-induced tumor vascular normalization include increased levels of tumor-infiltrating CD4+ and CD8+ T lymphocytes, natural killer cells and dendritic cells, decrease in myeloid-derived suppressor cells and polarization of TAMs from M2-to-M1 type (97, 111–114). Immune regulatory changes may be attributed to the direct effects on immune cells, including depletion of M2-type macrophages, indirect effects when they induce changes in protein expression on endothelial cells or tumor cells and rescue hypoxia and physical effects through the morphology of vascular normalization or the reduction of ECM (115). Lenvatinib, an inhibitor of VEGFRs, was loaded in Bi/Se nanoparticles to promote tumor vascular remodeling. The anti-angiogenic drug Lenvatinib was reported to enhance memory T-cell recruitment and CD8+ T lymphocyte, which can be potentiated by Bi/Se nanoparticles (111). Furthermore, the expression of TGF-β genes involved in the ECM components synthesis was inhibited by tranilast combined with Doxil nanomedicine to induce tumor vessel normalization. The distances between cancer-associated fibroblasts and CD3+ T cells were measured to reflect collagen barrier size. This finding represents another way to shape the tumor microenvironment and restrict T cell infiltration (114).
The interactions between anti-angiogenic therapies and immunotherapies could be considered a ‘two-way street’. A series of preclinical and clinical studies indicated the mutually improved effect of anti-angiogenic and immune-checkpoint inhibitors (94, 115). Of course, only if the action of anti-angiogenic drugs are limited directly to the tumor with little influence on normal tissues will disrupting tumor vasculature be beneficial for immunotherapy. Therefore, nanotechnology is needed to prevent negative effects on healthy non-target tissue while providing accumulation into the target tissue (116). In addition, the nanomedicine-mediated PDT and PTT can promote dendritic cell maturation and immune cell activation. It is reported that PDT and PTT synergetic with immune therapy for cancer treatment (117, 118). Furthermore, nanoparticles can be created to enhance cooperative and coordinated therapeutic effects by co-delivery and targeting of many therapeutic agents, as well as through immunomodulation. Nanoparticles have the potential to enhance treatment results by assuming these functions in combination therapy techniques (119). It is known that the immune checkpoint molecule programmed cell death protein 1(PD-1) can bind with its ligand (PD-L1) to protect tumor cells from immune surveillance via T cell exhaustion. CD8+ T cell infiltration plays a key role in anti-PD-1/PD-L1 therapy (112). Combining tumor vascular normalization with anti-PD-L1 therapy in breast cancer (97, 114) and hepatocellular carcinoma (112) has attracted significant interest in recent years. As a result, combination therapies could increase CD3+CD8+T cells, inhibit tumor proliferation and prevent tumor metastasis, providing a promising strategy to enhance immune-checkpoint inhibitor efficacy and the antitumor immunity of highly metastatic tumors.
Vascular normalization can even regulate “cold” immune microenvironment and provide a platform for sequential immunotherapy. The CD40 agonist stimulates antigen-presenting cells to process tumor-associated antigens produced by dying tumor cells, reversing immunosuppressive tumor microenvironments. However, CD40 may not have significant anticancer efficacy as a single treatment in patients with immunologically “cold” tumor (120). Xiang Ling et al. created the combination of anti-angiogenesis bioresponsive nanoparticles and chemotherapy regimens with CD40 agonist in a lung cancer mouse model. Compared with single treatment, combination therapies can reverse immunosuppression to elicit strong systemic antitumor immunity (113). For immunosuppressed tumors, nanomedicine induced vascular normalization provides a new idea for the application of immunotherapy.
Since the antitumor angiogenesis therapy was first proposed, the structure and function of the tumor microvasculature have gradually been revealed in the past two decades, along with advances in research tools and imaging techniques, and significant progress has been made in understanding the key mechanisms of angiogenesis and vascular remodeling in solid tumors, especially those involved in tumor angiogenesis and tumor vascular network maturation, and the effects of these structures on other components of the TME. The close interactions between these structures and other components of the TME are essential for the development of novel anti-angiogenic drugs and better guidance of combination antitumor therapy.
In recent years, immunotherapy has been widely studied and yielded promising therapeutic effects in preclinical studies. However, clinically, some patients do not respond to immune checkpoint therapy or are resistant to long-term use. It should be borne in mind that long-term use of immunotherapy treatment also presents serious cardiovascular side effects, etc. Achieving more efficient targeted delivery and improving suppressive TME while reducing toxic side effects warrants further research. Indeed, the past decade has witnessed unprecedented inroads in nanomedicine with the development of effective delivery systems that are not limited to targeting tumor cells but also normalize tumor vascular structure and function, reduce intratumor hypoxia, and increase blood perfusion while increasing immune cell infiltration and activating immune cells to achieve better immune functions in the TME.
Overall, the abnormalities of tumor vascular structure and function and its effects on the tumor microenvironment were summarized in this review, and the application of nano-strategies in combination with multi-targets and multi-combination nano-drugs showed significant anti-cancer effects in a wide range of solid tumors with low systemic toxicities, while exhibiting more low-dose and efficient antitumor effects when combined with other chemotherapeutic agents and immunotherapy.
However, several important issues remain to be addressed before anti-angiogenic nanodrugs are translated into clinical applications. First, the efficacy and safety of anti-angiogenic nanodrugs have only been evaluated over a relatively short period (a few days or weeks) in most current studies. Indeed, long follow-up observations and toxicological experiments are needed to ensure the absence of local or systemic toxic side effects of specific nanodrugs and other components before clinical translation. Currently available functional identification methods for tumor vascular normalization are only applicable in animals, and the establishment of reliable biomarkers and assays for vascular normalization for clinical practice is of great significance to refining the applicability of nanomedicines to cancer patients. Most current nanomedicines have been evaluated using allogeneic transplantation tumor models, and the application of spontaneous tumor models or PDX models would be more beneficial for evaluating the translational application value. Besides, quality control and the dose and mode of administration of nanomedicines warrant further evaluation in clinical trials. In addition, based on the series of issues mentioned above, almost all nanomedicines have failed to achieve the expected efficacy in antitumor clinical trials and clinical applications. More than 100 antitumor clinical trials of nanomedicines have been registered in clinical trials(https://clinicaltrials.gov/), including those based on LNP, Polysiloxane Gd-Chelates based nanoparticles (NCT04881032), Ceramide NanoLiposome (KNAN2001), SNB-101 (NCT04640480) and others. However, until now, only paclitaxel albumin-stabilized nanoparticle formulation has shown increased percentage of patients with response in clinical trials, but it also showed increased serious and other adverse events, such as hemoglobin decreased, neutrophil count decreased (NCT00626405). Therefore, there are still many issues that need to be thought and solved to achieve clinical translation of nanomedicines.
Despite these limitations, it is widely expected that the rapid development of nanomedicine will solve and greatly improve these problems in the next 5-10 years, and nanomedicines are expected to become the first or second line of standardized treatment for cancer patients.
All authors contributed to writing and revising the manuscript content.
This work is supported by grants from the National Natural Science Foundation of China (81871874 to KW), the Zhejiang Provincial Department of Science and Technology of China (2020C03027 to KW), the Key Science and Technology Projects of Jinhua Science and Technology Bureau(2022-3-047 to ZX).
We are grateful to all the participants who have made this research possible. And we would like to thank Freescience for English language editing.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.

1. Hanahan D. Hallmarks of cancer: New dimensions. Cancer Discovery (2022) 12(1):31–46. doi: 10.1158/2159-8290.CD-21-1059
2. Rivera LB, Bergers G. Cancer. tumor angiogenesis, from foe to friend. Science (2015) 349(6249):694–5. doi: 10.1126/science.aad0862
3. De Palma M, Biziato D, Petrova TV. Microenvironmental regulation of tumour angiogenesis. Nat Rev Cancer (2017) 17(8):457–74. doi: 10.1038/nrc.2017.51
4. Jain RK. Molecular regulation of vessel maturation. Nat Med (2003) 9(6):685–93. doi: 10.1038/nm0603-685
5. Jain RK. Normalization of tumor vasculature: An emerging concept in antiangiogenic therapy. Science (2005) 307(5706):58–62. doi: 10.1126/science.1104819
6. Carmeliet P, Jain RK. Principles and mechanisms of vessel normalization for cancer and other angiogenic diseases. Nat Rev Drug Discovery (2011) 10(6):417–27. doi: 10.1038/nrd3455
7. Goel S, Duda DG, Xu L, Munn LL, Boucher Y, Fukumura D, et al. Normalization of the vasculature for treatment of cancer and other diseases. Physiol Rev (2011) 91(3):1071–121. doi: 10.1152/physrev.00038.2010
8. Itatani Y, Kawada K, Yamamoto T, Sakai Y. Resistance to anti-angiogenic therapy in cancer-alterations to anti-vegf pathway. Int J Mol Sci (2018) 19(4):1232. doi: 10.3390/ijms19041232
9. Huinen ZR, Huijbers EJM, van Beijnum JR, Nowak-Sliwinska P, Griffioen AW. Anti-angiogenic agents - overcoming tumour endothelial cell anergy and improving immunotherapy outcomes. Nat Rev Clin Oncol (2021) 18(8):527–40. doi: 10.1038/s41571-021-00496-y
10. Cascone T, Xu L, Lin HY, Liu W, Tran HT, Liu Y, et al. The Hgf/C-met pathway is a driver and biomarker of vegfr-inhibitor resistance and vascular remodeling in non-small cell lung cancer. Clin Cancer Res (2017) 23(18):5489–501. doi: 10.1158/1078-0432.CCR-16-3216
11. Sennino B, McDonald DM. Controlling escape from angiogenesis inhibitors. Nat Rev Cancer (2012) 12(10):699–709. doi: 10.1038/nrc3366
12. Mitamura T, Pradeep S, McGuire M, Wu SY, Ma S, Hatakeyama H, et al. Induction of anti-vegf therapy resistance by upregulated expression of microseminoprotein (Msmp). Oncogene (2018) 37(6):722–31. doi: 10.1038/onc.2017.348
13. Chen M, Lei X, Shi C, Huang M, Li X, Wu B, et al. Pericyte-targeting prodrug overcomes tumor resistance to vascular disrupting agents. J Clin Invest (2017) 127(10):3689–701. doi: 10.1172/JCI94258
14. Viallard C, Larrivee B. Tumor angiogenesis and vascular normalization: Alternative therapeutic targets. Angiogenesis (2017) 20(4):409–26. doi: 10.1007/s10456-017-9562-9
15. Martin JD, Seano G, Jain RK. Normalizing function of tumor vessels: Progress, opportunities, and challenges. Annu Rev Physiol (2019) 81:505–34. doi: 10.1146/annurev-physiol-020518-114700
16. Lee WS, Yang H, Chon HJ, Kim C. Combination of anti-angiogenic therapy and immune checkpoint blockade normalizes vascular-immune crosstalk to potentiate cancer immunity. Exp Mol Med (2020) 52(9):1475–85. doi: 10.1038/s12276-020-00500-y
17. Peer D, Karp JM, Hong S, Farokhzad OC, Margalit R, Langer R. Nanocarriers as an emerging platform for cancer therapy. Nat Nanotechnol (2007) 2(12):751–60. doi: 10.1038/nnano.2007.387
18. Shi J, Kantoff PW, Wooster R, Farokhzad OC. Cancer nanomedicine: Progress, challenges and opportunities. Nat Rev Cancer (2017) 17(1):20–37. doi: 10.1038/nrc.2016.108
19. Jahanban-Esfahlan R, Seidi K, Banimohamad-Shotorbani B, Jahanban-Esfahlan A, Yousefi B. Combination of nanotechnology with vascular targeting agents for effective cancer therapy. J Cell Physiol (2018) 233(4):2982–92. doi: 10.1002/jcp.26051
20. Mukherjee A, Madamsetty VS, Paul MK, Mukherjee S. Recent advancements of nanomedicine towards antiangiogenic therapy in cancer. Int J Mol Sci (2020) 21(2):455. doi: 10.3390/ijms21020455
21. Zhang Y, He J. Tumor vasculature-targeting nanomedicines. Acta Biomater (2021) 134:1–12. doi: 10.1016/j.actbio.2021.07.015
22. Kahn BM, Lucas A, Alur RG, Wengyn MD, Schwartz GW, Li J, et al. The vascular landscape of human cancer. J Clin Invest (2021) 131(2):e136655. doi: 10.1172/JCI136655
23. Zerbini G, Lorenzi M, Palini A. Tumor angiogenesis. N Engl J Med (2008) 359(7):763. doi: 10.1056/NEJMc081278
24. Hanahan D, Folkman J. Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell (1996) 86(3):353–64. doi: 10.1016/s0092-8674(00)80108-7
25. Marcelo KL, Goldie LC, Hirschi KK. Regulation of endothelial cell differentiation and specification. Circ Res (2013) 112(9):1272–87. doi: 10.1161/CIRCRESAHA.113.300506
26. Yang H, Lee S, Lee S, Kim K, Yang Y, Kim JH, et al. Sox17 promotes tumor angiogenesis and destabilizes tumor vessels in mice. J Clin Invest (2013) 123(1):418–31. doi: 10.1172/JCI64547
27. Ferrara N, Adamis AP. Ten years of anti-vascular endothelial growth factor therapy. Nat Rev Drug Discovery (2016) 15(6):385–403. doi: 10.1038/nrd.2015.17
28. Kerbel RS. Tumor angiogenesis: Past, present and the near future. Carcinogenesis (2000) 21(3):505–15. doi: 10.1093/carcin/21.3.505
29. Pietrangeli P, Seguella L, Annunziata G, Casano F, Capuano R, Pesce M, et al. Lathyrus sativus diamine oxidase counteracts histamine-induced cell proliferation, migration and pro-angiogenic mediators release in human colon adenocarcinoma cell line caco-2. Phytother Res (2019) 33(7):1878–87. doi: 10.1002/ptr.6378
30. Bergers G, Benjamin LE. Tumorigenesis and the angiogenic switch. Nat Rev Cancer (2003) 3(6):401–10. doi: 10.1038/nrc1093
31. Ribatti D, Nico B, Crivellato E, Roccaro AM, Vacca A. The history of the angiogenic switch concept. Leukemia (2007) 21(1):44–52. doi: 10.1038/sj.leu.2404402
32. Francavilla C, Maddaluno L, Cavallaro U. The functional role of cell adhesion molecules in tumor angiogenesis. Semin Cancer Biol (2009) 19(5):298–309. doi: 10.1016/j.semcancer.2009.05.004
33. Giancotti FG, Ruoslahti E. Integrin signaling. Science (1999) 285(5430):1028–32. doi: 10.1126/science.285.5430.1028
34. van Beijnum JR, Huijbers EJM, van Loon K, Blanas A, Akbari P, Roos A, et al. Extracellular vimentin mimics vegf and is a target for anti-angiogenic immunotherapy. Nat Commun (2022) 13(1):2842. doi: 10.1038/s41467-022-30063-7
35. Yang Y, McCloskey JE, Yang H, Puc J, Alcaina Y, Vedvyas Y, et al. Bispecific car T cells against epcam and inducible icam-1 overcome antigen heterogeneity and generate superior antitumor responses. Cancer Immunol Res (2021) 9(10):1158–74. doi: 10.1158/2326-6066.CIR-21-0062
36. Huijbers EJM, Khan KA, Kerbel RS, Griffioen AW. Tumors resurrect an embryonic vascular program to escape immunity. Sci Immunol (2022) 7(67):eabm6388. doi: 10.1126/sciimmunol.abm6388
37. Xu Z, Guo C, Ye Q, Shi Y, Sun Y, Zhang J, et al. Endothelial deletion of Shp2 suppresses tumor angiogenesis and promotes vascular normalization. Nat Commun (2021) 12(1):6310. doi: 10.1038/s41467-021-26697-8
38. Aird WC. Endothelial cell heterogeneity. Cold Spring Harb Perspect Med (2012) 2(1):a006429. doi: 10.1101/cshperspect.a006429
39. Armulik A, Abramsson A, Betsholtz C. Endothelial/Pericyte interactions. Circ Res (2005) 97(6):512–23. doi: 10.1161/01.RES.0000182903.16652.d7
40. Jain RK. Normalizing tumor vasculature with anti-angiogenic therapy: A new paradigm for combination therapy. Nat Med (2001) 7(9):987–9. doi: 10.1038/nm0901-987
41. Vaupel P, Kallinowski F, Okunieff P. Blood flow, oxygen and nutrient supply, and metabolic microenvironment of human tumors: A review. Cancer Res (1989) 49(23):6449–65.
42. Murciano-Goroff YR, Warner AB, Wolchok JD. The future of cancer immunotherapy: Microenvironment-targeting combinations. Cell Res (2020) 30(6):507–19. doi: 10.1038/s41422-020-0337-2
43. Lanitis E, Irving M, Coukos G. Targeting the tumor vasculature to enhance T cell activity. Curr Opin Immunol (2015) 33:55–63. doi: 10.1016/j.coi.2015.01.011
44. Buckanovich RJ, Facciabene A, Kim S, Benencia F, Sasaroli D, Balint K, et al. Endothelin b receptor mediates the endothelial barrier to T cell homing to tumors and disables immune therapy. Nat Med (2008) 14(1):28–36. doi: 10.1038/nm1699
45. Muller WA. Mechanisms of leukocyte transendothelial migration. Annu Rev Pathol (2011) 6:323–44. doi: 10.1146/annurev-pathol-011110-130224
46. Rodig N, Ryan T, Allen JA, Pang H, Grabie N, Chernova T, et al. Endothelial expression of pd-L1 and pd-L2 down-regulates Cd8+ T cell activation and cytolysis. Eur J Immunol (2003) 33(11):3117–26. doi: 10.1002/eji.200324270
47. Motz GT, Santoro SP, Wang LP, Garrabrant T, Lastra RR, Hagemann IS, et al. Tumor endothelium fasl establishes a selective immune barrier promoting tolerance in tumors. Nat Med (2014) 20(6):607–15. doi: 10.1038/nm.3541
48. Taylor CT, Colgan SP. Regulation of immunity and inflammation by hypoxia in immunological niches. Nat Rev Immunol (2017) 17(12):774–85. doi: 10.1038/nri.2017.103
49. Barsoum IB, Smallwood CA, Siemens DR, Graham CH. A mechanism of hypoxia-mediated escape from adaptive immunity in cancer cells. Cancer Res (2014) 74(3):665–74. doi: 10.1158/0008-5472.CAN-13-0992
50. Roma-Rodrigues C, Mendes R, Baptista PV, Fernandes AR. Targeting tumor microenvironment for cancer therapy. Int J Mol Sci (2019) 20(4):840. doi: 10.3390/ijms20040840
51. Damgaci S, Ibrahim-Hashim A, Enriquez-Navas PM, Pilon-Thomas S, Guvenis A, Gillies RJ. Hypoxia and acidosis: Immune suppressors and therapeutic targets. Immunology (2018) 154(3):354–62. doi: 10.1111/imm.12917
52. Voron T, Colussi O, Marcheteau E, Pernot S, Nizard M, Pointet AL, et al. Vegf-a modulates expression of inhibitory checkpoints on Cd8+ T cells in tumors. J Exp Med (2015) 212(2):139–48. doi: 10.1084/jem.20140559
53. Terme M, Pernot S, Marcheteau E, Sandoval F, Benhamouda N, Colussi O, et al. Vegfa-vegfr pathway blockade inhibits tumor-induced regulatory T-cell proliferation in colorectal cancer. Cancer Res (2013) 73(2):539–49. doi: 10.1158/0008-5472.CAN-12-2325
54. Huang Y, Chen X, Dikov MM, Novitskiy SV, Mosse CA, Yang L, et al. Distinct roles of vegfr-1 and vegfr-2 in the aberrant hematopoiesis associated with elevated levels of vegf. Blood (2007) 110(2):624–31. doi: 10.1182/blood-2007-01-065714
55. Varney ML, Johansson SL, Singh RK. Tumour-associated macrophage infiltration, neovascularization and aggressiveness in malignant melanoma: Role of monocyte chemotactic protein-1 and vascular endothelial growth factor-a. Melanoma Res (2005) 15(5):417–25. doi: 10.1097/00008390-200510000-00010
56. Li SJ, Chen JX, Sun ZJ. Improving antitumor immunity using antiangiogenic agents: Mechanistic insights, current progress, and clinical challenges. Cancer Commun (Lond) (2021) 41(9):830–50. doi: 10.1002/cac2.12183
57. Teleanu RI, Chircov C, Grumezescu AM, Teleanu DM. Tumor angiogenesis and anti-angiogenic strategies for cancer treatment. J Clin Med (2019) 9(1):84. doi: 10.3390/jcm9010084
58. Bianchini F, De Santis A, Portioli E, Russo Krauss I, Battistini L, Curti C, et al. Integrin-targeted amprgd sunitinib liposomes as integrated antiangiogenic tools. Nanomedicine (2019) 18:135–45. doi: 10.1016/j.nano.2019.02.015
59. Catuogno S, Esposito CL, Condorelli G, de Franciscis V. Nucleic acids delivering nucleic acids. Adv Drug Delivery Rev (2018) 134:79–93. doi: 10.1016/j.addr.2018.04.006
60. Lu S, Bao X, Hai W, Shi S, Chen Y, Yu Q, et al. Multi-functional self-assembled nanoparticles for pvegf-shrna loading and anti-tumor targeted therapy. Int J Pharm (2020) 575:118898. doi: 10.1016/j.ijpharm.2019.118898
61. Li X, Wang X, Liu N, Wang Q, Hu J. Inhibition of metastatic hepatocarcinoma by combined chemotherapy with silencing Vegf/Vegfr2 genes through a galnac-modified integrated therapeutic system. Molecules (2022) 27(7):2082. doi: 10.3390/molecules27072082
62. Jiang Q, Yue S, Yu K, Tian T, Zhang J, Chu H, et al. Endogenous microrna triggered enzyme-free DNA logic self-assembly for amplified bioimaging and enhanced gene therapy Via in situ generation of sirnas. J Nanobiotechnol (2021) 19(1):288. doi: 10.1186/s12951-021-01040-x
63. Dalmina M, Pittella F, Sierra JA, Souza GRR, Silva AH, Pasa AA, et al. Magnetically responsive hybrid nanoparticles for in vitro sirna delivery to breast cancer cells. Mater Sci Eng C Mater Biol Appl (2019) 99:1182–90. doi: 10.1016/j.msec.2019.02.026
64. Koide H, Yoshimatsu K, Hoshino Y, Ariizumi S, Okishima A, Ide T, et al. Sequestering and inhibiting a vascular endothelial growth factor in vivo by systemic administration of a synthetic polymer nanoparticle. J Control Release (2019) 295:13–20. doi: 10.1016/j.jconrel.2018.12.033
65. Clavreul A, Pourbaghi-Masouleh M, Roger E, Menei P. Nanocarriers and nonviral methods for delivering antiangiogenic factors for glioblastoma therapy: The story so far. Int J Nanomed (2019) 14:2497–513. doi: 10.2147/IJN.S194858
66. Lopez de Andres J, Grinan-Lison C, Jimenez G, Marchal JA. Cancer stem cell secretome in the tumor microenvironment: A key point for an effective personalized cancer treatment. J Hematol Oncol (2020) 13(1):136. doi: 10.1186/s13045-020-00966-3
67. Paris JL, Villaverde G, Gomez-Grana S, Vallet-Regi M. Nanoparticles for multimodal antivascular therapeutics: Dual drug release, photothermal and photodynamic therapy. Acta Biomater (2020) 101:459–68. doi: 10.1016/j.actbio.2019.11.004
68. El-Sahli S, Hua K, Sulaiman A, Chambers J, Li L, Farah E, et al. A triple-drug nanotherapy to target breast cancer cells, cancer stem cells, and tumor vasculature. Cell Death Dis (2021) 12(1):8. doi: 10.1038/s41419-020-03308-w
69. Penn CA, Yang K, Zong H, Lim JY, Cole A, Yang D, et al. Therapeutic impact of nanoparticle therapy targeting tumor-associated macrophages. Mol Cancer Ther (2018) 17(1):96–106. doi: 10.1158/1535-7163.MCT-17-0688
70. Zhang BC, Luo BY, Zou JJ, Wu PY, Jiang JL, Le JQ, et al. Co-Delivery of sorafenib and Crispr/Cas9 based on targeted core-shell hollow mesoporous organosilica nanoparticles for synergistic hcc therapy. ACS Appl Mater Interfaces (2020) 12(51):57362–72. doi: 10.1021/acsami.0c17660
71. Zhang L, Qi Y, Min H, Ni C, Wang F, Wang B, et al. Cooperatively responsive peptide nanotherapeutic that regulates angiopoietin receptor Tie2 activity in tumor microenvironment to prevent breast tumor relapse after chemotherapy. ACS Nano (2019) 13(5):5091–102. doi: 10.1021/acsnano.8b08142
72. Hida K, Maishi N, Sakurai Y, Hida Y, Harashima H. Heterogeneity of tumor endothelial cells and drug delivery. Adv Drug Delivery Rev (2016) 99(Pt B):140–7. doi: 10.1016/j.addr.2015.11.008
73. Sakurai Y, Akita H, Harashima H. Targeting tumor endothelial cells with nanoparticles. Int J Mol Sci (2019) 20(23):5819. doi: 10.3390/ijms20235819
74. Siemann DW, Chaplin DJ, Horsman MR. Realizing the potential of vascular targeted therapy: The rationale for combining vascular disrupting agents and anti-angiogenic agents to treat cancer. Cancer Invest (2017) 35(8):519–34. doi: 10.1080/07357907.2017.1364745
75. Lee HK, Bae HR, Park HK, Seo IA, Lee EY, Suh DJ, et al. Cloning, characterization and neuronal expression profiles of tumor endothelial marker 7 in the rat brain. Brain Res Mol Brain Res (2005) 136(1-2):189–98. doi: 10.1016/j.molbrainres.2005.02.010
76. Kim GH, Won JE, Byeon Y, Kim MG, Wi TI, Lee JM, et al. Selective delivery of Plxdc1 small interfering rna to endothelial cells for anti-angiogenesis tumor therapy using Cd44-targeted chitosan nanoparticles for epithelial ovarian cancer. Drug Delivery (2018) 25(1):1394–402. doi: 10.1080/10717544.2018.1480672
77. Yongvongsoontorn N, Chung JE, Gao SJ, Bae KH, Yamashita A, Tan MH, et al. Carrier-enhanced anticancer efficacy of sunitinib-loaded green tea-based micellar nanocomplex beyond tumor-targeted delivery. ACS Nano (2019) 13(7):7591–602. doi: 10.1021/acsnano.9b00467
78. Cabral AS, Leonel ECR, Candido NM, Piva HL, de Melo MT, Taboga SR, et al. Combined photodynamic therapy with chloroaluminum phthalocyanine and doxorubicin nanoemulsions in breast cancer model. J Photochem Photobiol B (2021) 218:112181. doi: 10.1016/j.jphotobiol.2021.112181
79. Sun C, Gao S, Tan Y, Zhang Z, Xu H. Side-chain selenium-grafted polymers combining antiangiogenesis treatment with photodynamic therapy and chemotherapy. ACS Biomater Sci Eng (2021) 7(7):3201–8. doi: 10.1021/acsbiomaterials.1c00254
80. Zhang J, Zeng Y, Su M, Yu M, Zhang Y, Cheng H, et al. Multifunctional ferritin nanoparticles as theranostics for imaging-guided tumor phototherapy. J BioMed Nanotechnol (2019) 15(7):1546–55. doi: 10.1166/jbn.2019.2788
81. Belotti D, Paganoni P, Manenti L, Garofalo A, Marchini S, Taraboletti G, et al. Matrix metalloproteinases (Mmp9 and Mmp2) induce the release of vascular endothelial growth factor (Vegf) by ovarian carcinoma cells: Implications for ascites formation. Cancer Res (2003) 63(17):5224–9.
82. Xiao Q, Zhu X, Yuan Y, Yin L, He W. A drug-Delivering-Drug strategy for combined treatment of metastatic breast cancer. Nanomedicine (2018) 14(8):2678–88. doi: 10.1016/j.nano.2018.06.012
83. Sun Z, Li R, Sun J, Peng Y, Xiao L, Zhang X, et al. Matrix metalloproteinase cleavable nanoparticles for tumor microenvironment and tumor cell dual-targeting drug delivery. ACS Appl Mater Interfaces (2017) 9(46):40614–27. doi: 10.1021/acsami.7b11614
84. Khan I, Bhardwaj M, Shukla S, Lee H, Oh MH, Bajpai VK, et al. Carvacrol encapsulated Nanocarrier/Nanoemulsion abrogates angiogenesis by downregulating cox-2, vegf and Cd31 in vitro and in vivo in a lung adenocarcinoma model. Colloids Surf B Biointerfaces (2019) 181:612–22. doi: 10.1016/j.colsurfb.2019.06.016
85. Wojtukiewicz MZ, Sierko E, Hempel D, Tucker SC, Honn KV. Platelets and cancer angiogenesis nexus. Cancer Metastasis Rev (2017) 36(2):249–62. doi: 10.1007/s10555-017-9673-1
86. Jing L, Qu H, Wu D, Zhu C, Yang Y, Jin X, et al. Platelet-camouflaged nanococktail: Simultaneous inhibition of drug-resistant tumor growth and metastasis Via a cancer cells and tumor vasculature dual-targeting strategy. Theranostics (2018) 8(10):2683–95. doi: 10.7150/thno.23654
87. Li S, Zhang Y, Wang J, Zhao Y, Ji T, Zhao X, et al. Nanoparticle-mediated local depletion of tumour-associated platelets disrupts vascular barriers and augments drug accumulation in tumours. Nat BioMed Eng (2017) 1(8):667–79. doi: 10.1038/s41551-017-0115-8
88. Zhang Y, Liu G, Wei J, Nie G. Platelet membrane-based and tumor-associated platelettargeted drug delivery systems for cancer therapy. Front Med (2018) 12(6):667–77. doi: 10.1007/s11684-017-0583-y
89. Guan YY, Luan X, Xu JR, Liu YR, Lu Q, Wang C, et al. Selective eradication of tumor vascular pericytes by peptide-conjugated nanoparticles for antiangiogenic therapy of melanoma lung metastasis. Biomaterials (2014) 35(9):3060–70. doi: 10.1016/j.biomaterials.2013.12.027
90. Lugano R, Ramachandran M, Dimberg A. Tumor angiogenesis: Causes, consequences, challenges and opportunities. Cell Mol Life Sci (2020) 77(9):1745–70. doi: 10.1007/s00018-019-03351-7
91. Wang Y, Tong L, Wang J, Luo J, Tang J, Zhong L, et al. Crgd-functionalized nanoparticles for combination therapy of anti-endothelium dependent vessels and anti-vasculogenic mimicry to inhibit the proliferation of ovarian cancer. Acta Biomater (2019) 94:495–504. doi: 10.1016/j.actbio.2019.06.039
92. Du S, Xiong H, Xu C, Lu Y, Yao J. Attempts to strengthen and simplify the tumor vascular normalization strategy using tumor vessel normalization promoting nanomedicines. Biomater Sci (2019) 7(3):1147–60. doi: 10.1039/c8bm01350k
93. Wang X, Niu X, Sha W, Feng X, Yu L, Zhang Z, et al. An oxidation responsive nano-radiosensitizer increases radiotherapy efficacy by remolding tumor vasculature. Biomater Sci (2021) 9(18):6308–24. doi: 10.1039/d1bm00834j
94. Yi M, Jiao D, Qin S, Chu Q, Wu K, Li A. Synergistic effect of immune checkpoint blockade and anti-angiogenesis in cancer treatment. Mol Cancer (2019) 18(1):60. doi: 10.1186/s12943-019-0974-6
95. Mattheolabakis G, Mikelis CM. Nanoparticle delivery and tumor vascular normalization: The chicken or the egg? Front Oncol (2019) 9:1227. doi: 10.3389/fonc.2019.01227
96. Li W, Li X, Liu S, Yang W, Pan F, Yang XY, et al. Gold nanoparticles attenuate metastasis by tumor vasculature normalization and epithelial-mesenchymal transition inhibition. Int J Nanomed (2017) 12:3509–20. doi: 10.2147/IJN.S128802
97. Huang N, Liu Y, Fang Y, Zheng S, Wu J, Wang M, et al. Gold nanoparticles induce tumor vessel normalization and impair metastasis by inhibiting endothelial Smad2/3 signaling. ACS Nano (2020) 14(7):7940–58. doi: 10.1021/acsnano.9b08460
98. Huang W, Xing Y, Zhu L, Zhuo J, Cai M. Sorafenib derivatives-functionalized gold nanoparticles confer protection against tumor angiogenesis and proliferation Via suppression of egfr and vegfr-2. Exp Cell Res (2021) 406(1):112633. doi: 10.1016/j.yexcr.2021.112633
99. Li W, Zhao X, Du B, Li X, Liu S, Yang XY, et al. Gold nanoparticle-mediated targeted delivery of recombinant human endostatin normalizes tumour vasculature and improves cancer therapy. Sci Rep (2016) 6:30619. doi: 10.1038/srep30619
100. Pan F, Yang W, Li W, Yang XY, Liu S, Li X, et al. Conjugation of gold nanoparticles and recombinant human endostatin modulates vascular normalization Via interruption of anterior gradient 2-mediated angiogenesis. Tumour Biol (2017) 39(7):1010428317708547. doi: 10.1177/1010428317708547
101. Raposo TP, Pires I, Carvalho MI, Prada J, Argyle DJ, Queiroga FL. Tumour-associated macrophages are associated with vascular endothelial growth factor expression in canine mammary tumours. Vet Comp Oncol (2015) 13(4):464–74. doi: 10.1111/vco.12067
102. Corzo CA, Condamine T, Lu L, Cotter MJ, Youn JI, Cheng P, et al. Hif-1alpha regulates function and differentiation of myeloid-derived suppressor cells in the tumor microenvironment. J Exp Med (2010) 207(11):2439–53. doi: 10.1084/jem.20100587
103. Tian L, Yi X, Dong Z, Xu J, Liang C, Chao Y, et al. Calcium bisphosphonate nanoparticles with chelator-free radiolabeling to deplete tumor-associated macrophages for enhanced cancer radioisotope therapy. ACS Nano (2018) 12(11):11541–51. doi: 10.1021/acsnano.8b06699
104. Yonucu S, Yiotalmaz D, Phipps C, Unlu MB, Kohandel M. Quantifying the effects of antiangiogenic and chemotherapy drug combinations on drug delivery and treatment efficacy. PloS Comput Biol (2017) 13(9):e1005724. doi: 10.1371/journal.pcbi.1005724
105. Jiao J, He J, Li M, Yang J, Yang H, Wang X, et al. A porphyrin-based metallacage for enhanced photodynamic therapy. Nanoscale (2022) 14(17):6373–83. doi: 10.1039/d1nr08293k
106. Gong H, Chao Y, Xiang J, Han X, Song G, Feng L, et al. Hyaluronidase to enhance nanoparticle-based photodynamic tumor therapy. Nano Lett (2016) 16(4):2512–21. doi: 10.1021/acs.nanolett.6b00068
107. Stylianopoulos T, Martin JD, Chauhan VP, Jain SR, Diop-Frimpong B, Bardeesy N, et al. Causes, consequences, and remedies for growth-induced solid stress in murine and human tumors. Proc Natl Acad Sci U.S.A. (2012) 109(38):15101–8. doi: 10.1073/pnas.1213353109
108. Stylianopoulos T, Munn LL, Jain RK. Reengineering the physical microenvironment of tumors to improve drug delivery and efficacy: From mathematical modeling to bench to bedside. Trends Cancer (2018) 4(4):292–319. doi: 10.1016/j.trecan.2018.02.005
109. Ernsting MJ, Hoang B, Lohse I, Undzys E, Cao P, Do T, et al. Targeting of metastasis-promoting tumor-associated fibroblasts and modulation of pancreatic tumor-associated stroma with a carboxymethylcellulose-docetaxel nanoparticle. J Control Release (2015) 206:122–30. doi: 10.1016/j.jconrel.2015.03.023
110. Zhang B, Jin K, Jiang T, Wang L, Shen S, Luo Z, et al. Celecoxib normalizes the tumor microenvironment and enhances small nanotherapeutics delivery to A549 tumors in nude mice. Sci Rep (2017) 7(1):10071. doi: 10.1038/s41598-017-09520-7
111. Liu J, Chen J, Liu H, Zhang K, Zeng Q, Yang S, et al. Bi/Se-based nanotherapeutics sensitize ct image-guided stereotactic body radiotherapy through reprogramming the microenvironment of hepatocellular carcinoma. ACS Appl Mater Interfaces (2021) 13(36):42473–85. doi: 10.1021/acsami.1c11763
112. Bao X, Shen N, Lou Y, Yu H, Wang Y, Liu L, et al. Enhanced anti-Pd-1 therapy in hepatocellular carcinoma by tumor vascular disruption and normalization dependent on combretastatin A4 nanoparticles and Dc101. Theranostics (2021) 11(12):5955–69. doi: 10.7150/thno.58164
113. Ling X, Jiang X, Li Y, Han W, Rodriguez M, Xu Z, et al. Sequential treatment of bioresponsive nanoparticles elicits antiangiogenesis and apoptosis and synergizes with a Cd40 agonist for antitumor immunity. ACS Nano (2021) 15(1):765–80. doi: 10.1021/acsnano.0c07132
114. Panagi M, Voutouri C, Mpekris F, Papageorgis P, Martin MR, Martin JD, et al. Tgf-beta inhibition combined with cytotoxic nanomedicine normalizes triple negative breast cancer microenvironment towards anti-tumor immunity. Theranostics (2020) 10(4):1910–22. doi: 10.7150/thno.36936
115. Khan KA, Kerbel RS. Improving immunotherapy outcomes with anti-angiogenic treatments and vice versa. Nat Rev Clin Oncol (2018) 15(5):310–24. doi: 10.1038/nrclinonc.2018.9
116. Gupta J, Safdari HA, Hoque M. Nanoparticle mediated cancer immunotherapy. Semin Cancer Biol (2021) 69:307–24. doi: 10.1016/j.semcancer.2020.03.015
117. Sun H, Yu T, Li X, Lei Y, Li J, Wang X, et al. Second near-infrared photothermal-amplified immunotherapy using photoactivatable composite nanostimulators. J Nanobiotechnol (2021) 19(1):433. doi: 10.1186/s12951-021-01197-5
118. He T, Luo Y, Zhang Q, Men Z, Su T, Fan L, et al. Hyalase-mediated cascade degradation of a matrix barrier and immune cell penetration by a photothermal microneedle for efficient anticancer therapy. ACS Appl Mater Interfaces (2021) 13(23):26790–9. doi: 10.1021/acsami.1c06725
119. Boone CE, Wang L, Gautam A, Newton IG, Steinmetz NF. Combining nanomedicine and immune checkpoint therapy for cancer immunotherapy. Wiley Interdiscip Rev Nanomed Nanobiotechnol (2022) 14(1):e1739. doi: 10.1002/wnan.1739
Keywords: cancer, anti-angiogenesis therapy, immunotherapy, tumor immune microenvironment (TIM), nanomedicine
Citation: Xiao M, Shi Y, Jiang S, Cao M, Chen W, Xu Y, Xu Z and Wang K (2022) Recent advances of nanomaterial-based anti-angiogenic therapy in tumor vascular normalization and immunotherapy. Front. Oncol. 12:1039378. doi: 10.3389/fonc.2022.1039378
Received: 08 September 2022; Accepted: 14 November 2022;
Published: 29 November 2022.
Edited by:
Yi Wei, Sichuan University, ChinaReviewed by:
Phei Er Saw, Sun Yat-sen Memorial Hospital, ChinaCopyright © 2022 Xiao, Shi, Jiang, Cao, Chen, Xu, Xu and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kai Wang, S2Fpd0B6anUuZWR1LmNu; Zhiyong Xu, eHV6aGl5b25nQHpqdS5lZHUuY24=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.