
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Oncol. , 05 November 2021
Sec. Genitourinary Oncology
Volume 11 - 2021 | https://doi.org/10.3389/fonc.2021.781414
This article is part of the Research Topic Advances in Molecular Targeted Therapies of Urologic Cancers View all 10 articles
 Mohammad Taheri1
Mohammad Taheri1 Sajad Najafi2
Sajad Najafi2 Abbas Basiri3Bashdar Mahmud Hussen4
Abbas Basiri3Bashdar Mahmud Hussen4 Aria Baniahmad5Elena Jamali6
Aria Baniahmad5Elena Jamali6 Soudeh Ghafouri-Fard7*
Soudeh Ghafouri-Fard7*Globally, prostate cancer (PCa) is the second most commonly diagnosed cancer in men globally. Early diagnosis may help in promoting survival in the affected patients. Circular RNAs (circRNAs) are a novel class of non-coding RNAs (ncRNAs) which have been found to show extensive dysregulation in a handful of human diseases including cancers. Progressions in RNA identification techniques have provided a vast number of circRNAs exhibiting either up-regulation or down-regulation in PCa tissues compared to normal adjacent tissues. The mechanism of action is not clear for most of dysregulated circRNAs. Among them, function of a number of newly identified dysregulated circRNAs have been assessed in PCa cells. Increase in cell proliferation, migration, invasion, and metastasis have been reported for up-regulated circRNAs which suggest their role as oncogenes. On the other hand, down-regulated circRNAs have shown tumor suppressing actions in experimental studies. Furthermore, in a majority of studies, circRNAs have been found to sponge microRNAs (miRNAs), negatively regulating expression or activity of the downstream miRNAs. Additionally, they have been identified in interaction with regulatory proteins. This axis consequently regulates a signaling pathway, a tumor suppressor, or an oncogene. Easy, quick, and reliable detection of circRNAs in human body fluids also suggests their potentials as biomarker candidates for diagnosis and prediction of prognosis in PCa patients. In this review, we have discussed the role and potentials of a number of dysregulated circRNAs in PCa.
Prostate cancer (PCa) accounts for more than 1 in 5 new cancer cases in men (1). High age is the main risk factor for PCa. Race, environmental and genetic factors are known as other predisposing factors (2). Majority of PCa cases are diagnosed with diseases of low to intermediate risk, and a minority of 30% experience shorter survival in case of distant metastasis of the malignancy (3). Gleason score is the most commonly used grading system for prediction of outcomes in PCa patients (4). The scores range from 6-10 with high scores corresponding to malignant PCa cells and lower survival in PCa patients. The main biomarker currently used for diagnosis of PCa is prostate-specific antigen (PSA) which harbors good diagnostic values, however cannot distinguish PCa from resembling milder prostate disorders like benign prostate hyperplasia (BPH) and prostatitis (5). Non-coding RNAs (ncRNAs) have been studied in PCa and their role in development, progression, and metastasis of malignancy has been evaluated in a handful of in vitro and in vivo experiments. Among ncRNAs, microRNAs (miRNAs) and lncRNAs [see more at (6, 7)] have been studied more in PCa compared with circRNAs and their roles and mechanisms in development and progression of PCa have been clarified due to the historical priority of discovery and facility of detection in research studies.
A large proportion of eukaryotic genome encodes no protein which is termed as non-coding RNAs (ncRNAs). These transcripts have been primarily described as junk DNA and now known to have essential regulatory roles. Circular RNAs (circRNAs) are covalently closed RNA transcripts usually belonging to a novel type of ncRNAs, namely long non-coding category (lncRNAs). CircRNAs have been primarily reported in viroids (8). Hsu and Coca-Prados (9) reported the first circRNAs in eukaryotes in 1979 via electron microscopy in HeLa cells. Compared to linear types of regulatory RNAs and even among ncRNAs, less attention has been paid to circRNAs. However, increasing evidence demonstrates their biological functions. CircRNAs are believed to be formed via back-splicing from pre-messenger RNA (pre-mRNA) or originated from differentially spliced transcripts (10). The main characteristic of circRNAs as their names suggest is their determinant circular form which develops via covalent linkages between the 5′ and 3′ ends and sometimes 5′-2′ phosphodiester bonds. CircRNAs exhibit dissimilarities to conventional linear RNAs such as mRNAs and transfer RNAs (tRNAs) which include lacking capping and polyadenylated (poly A) tail at their 5′ and 3′ ends, respectively. Lack of open ends makes RNA loops resistant to RNA degrading enzyme RNase R which facilitates biochemical characterization of circRNAs (11). Several approaches based on non-polyadenylation characteristic of circRNAs, rRNA-depletion, and RNase R-resistance have been developed for detection of circRNAs (12). The qRT-PCR analysis following RNase R treatment is the most common approach used for validation of circRNAs (13). Also, fast and easy detection of circRNAs can be conducted through rolling-circle amplification (RCA) (14). This technique does not require any advanced equipment or fluorescent probes and is performed just using qRT-PCR machine and gel electrophoresis. First, the circRNAs using first-generation primers is reverse transcribed generating a multimeric cDNA through RCA method compared to a monomeric cDNA for linear RNA template. Then using second-generation primer, the circRNAs-specific ligation site is spanned and subsequently can be seen on gel electrophoresis (15). High-throughput RNA sequencing (RNA-seq) along with bioinformatics tools [e.g., CircMiner (16)] and confirmatory techniques like quantitative real-time polymerase chain reaction (qRT-PCR, where the junction/fusion site is known) and Fluorescence in situ hybridization (FISH) have helped substantial progression in identification of differentially expressed circRNAs in cancerous tissues. Although qRT-PCR is the most common approach for experimental detection of circRNAs, however, currently, no easy, quick, and cheap technique is available for diagnostics to detect a specific circRNA, and so it is suggested that we have a long way to bring circRNAs to clinical setting (17).
CircRNAs have been identified in large quantities and have been revealed to be expressed widely in animal cells. Thousands of circRNAs are transcribed in considerable fractions from a large number of human genes (18). The number of identified unique circRNAs are more than twice of the linear counterparts (19). However, they are usually found in lesser quantities compare with their linear counterparts (10). In mammals, circRNAs show conservation in their sequences among different species, are mainly found in cytoplasm, and demonstrate specific tissue, cellular and developmental stage distribution (20, 21), even more specific compared to corresponding mRNA isoforms (22). Precise functions of circRNAs have not been clarified, but regulatory roles have been described for an increasing number of them. The first circRNA, which its function has been characterized, was CDR1as (21). CDR1as was shown to play role in gene expression at posttranscriptional stage via binding to and sponging miR-7. This circRNA is involved in brain development (21). Aberrant expression of circRNAs have been associated with pathological conditions such as cardiovascular diseases (23), sudden cardiac death (SCD) (24), neurodegenerative and psychiatric disorders (25, 26), kidney diseases (27), inflammation (28), autoimmune diseases (29) and particularly various types of cancer. Cellular studies have shown a vast number of circRNAs to be dysregulated in cancer tissues compared to normal tissues and this imbalance can enhance tumor development and progression via affecting cell cycle. Aberrant expression includes either up- or down-regulation in levels of circRNAs compared to those in normal cells. Up-regulated circRNAs in cancer are known as oncogenes. These oncogenic circRNAs such as circMBOAT2 (30) and circFOXO3 (31) accelerate tumor cell proliferation, migration, invasion, and metastasis, while suppressing apoptosis. On the other hand, down-regulated circRNAs are considered as tumor suppressors. CircRNAs can be detected in high abundance due to their stability in body fluids like serum and urine, and can also be specifically expressed in extracellular vesicles (32, 33). Therefore, their detection provides easy, rapid, reliable, and minimally invasive diagnostic routes for many types of cancers and other pathological conditions. The expression levels of circRNAs in a majority of studies have been significantly correlated with clinicopathological features in cancerous patients, accordingly they can help in prediction of the disease prognosis. Most importantly, targeting oncogenic circRNAs or reversing intermediates using RNA interference (RNAi) and antisense oligonucleotides (ASO) or enhancing expression of circRNA with tumor suppressing functions has suggested new therapeutical potentials in fighting against malignancies. Thus, circRNAs have been introduced as novel diagnostic and prognostic biomarkers and therapeutical targets particularly in cancer.
In this review, we focus on a number of circRNAs being dysregulated in prostate cancer (PCa), with an overview of the studies assessed the tumorigenic and anti-carcinogenic roles of them in PCa tissues and finally, their potential applications in diagnosis and prediction of prognosis in PCa patients.
In various studies, thousands of circRNAs have been found to show aberrant expression in PCa tissues compared to normal adjacent tissue (NAT) or also between several stages of malignancy including primary and metastatic PCa (34, 35). Some of these circRNAs promote PCa cell tumorigenicity enhancing cell proliferation, invasion and metastasis. An increased resistance to chemotherapy agents is another impact of oncogenic circRNAs, which can affect the survival in PCa patients (36). Regulatory effects on expression of androgens or their receptors and resistance to them or reverse interactions which play role in development of PCa have also been reported for several circRNAs [e.g., circRNA-17 (37), circSMARCA5 (38), and circRNA-51217 (39)]. For some other circRNAs such as circ-ITCH (40) and circMBOAT2 (30), a correlation has been recognized between circRNA expression levels and prognosis in PCa patients. Precise mechanism of action for circRNAs has been poorly understood. In bioinformatics-constructed regulatory networks such as Gene Ontology (GO), Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway, and competing endogenous RNA (ceRNA) network analyses to predict the function of circRNAs and also, RNA-pull down assays, an interaction between circRNAs and targeted miRNAs is frequently reported. Along with main miRNAs sponging activity, circRNAs also have been shown to exert their regulatory functions through interactions with proteins particularly RNA-binding proteins, which play role in RNAs maturation and influence various cellular functions (41). RNA-binding proteins can interplay with circRNA junctions and participate in splicing, processing, folding, stabilization, and localization of circRNAs (42). An example of circRNAs interacting with RNA-binding proteins are has_circ_0000020 (interacting with HuR, FMRP and EIF4A3) (43).
Also, interaction of circRNA with other ncRNAs particularly miRNAs, based on their complementary sequences has been reported in a set of experiments. Additionally, circRNAs can regulate RNA-binding proteins, and linear protein-coding mRNAs (44). These studies suggest an axis through which the circRNA affects (mainly reverses) expression or activity [see review in (45)] of a mediator miRNA as a target, which itself impacts a specific target signaling pathway, a tumor suppressor or an oncogene. Thus, it is thought that an oncogenic circRNA exerts its function through a signaling axis eventually activates an oncogene or through a signaling pathway accelerates cell cycle. Based on tissue, cancer or malignancy stage-specific expression of a circRNA in PCa, diagnostic analyses have unveiled reliability of a set of circRNAs as potential biomarkers in distinguishing PCa from any other condition or among cancer stages. Association between expression level of a circRNA and clinicopathological features in PCa patients like tumor stage, grade, lymph node metastasis in addition to prognosis, also has been found in a number of studies. For instance, Greene et al. (35) have reported that circRNAs not only demonstrate differential expression in PCa tissues, but also are expressed aberrantly according to the androgen dependency, which is known to play role in the pathogenesis of the disease. Furthermore, targeting aberrantly expressed circRNAs in PCa has exhibited hopeful results in a number of studies decreasing aggressiveness and tumorigenesis in cell and in vivo studies.
Up-regulated circRNAs, as discussed above, act as oncogenic ncRNAs promoting tumorigenic features of tumor cell lines and also, increase tumor progression in vivo. CircRNA knockdown using specific small interfering RNA (siRNA) or overexpression by encoding vectors have been employed in functional analysis. Enhanced tumor cell proliferation, invasion and migration have been reported following over-expression of these circRNAs, in cell viability and colony formation, migration and invasion assays, respectively. Suppressed apoptosis is also reported in functional analysis of up-regulated circRNAs. Furthermore, oncogenic circRNAs could decrease chemosensitivity and radiosensitivity of the cancer cell lines to the current therapeutical approaches of PCa (46, 47). As a result, decreased effectiveness of the cancer therapeutics and eventually adverse consequences such as shortened survival is predicted for the patients.
For instance, Zhang et al. (48) have identified 89 circRNAs which showed aberrant expression in PCa tissues, among them 32 circRNAs showed increased expression and remaining 57 demonstrated to be down-regulated. In further investigations, 5 prominently overexpressed circRNAs in comparison to their corresponding mRNAs in PCa tissues including hsa_circ_0006754, hsa_circ_0005848, hsa_circ_0006410, hsa_circ_0003970, and hsa_circ_AKAP7 were recognized. Interaction networks revealed 215 linkages between 5 selected circRNAs and corresponding miRNAs. Several miRNAs including miR-204-5p, miR-3160-5p, and miR-548 were identified as the most prominent targets of associated circRNAs which play role in suppression or promotion of carcinogenesis or via enhancing apoptosis, inhibiting cell proliferation or PI3K/AKT signaling pathway, respectively. MAPK signaling pathway was known as the most important signaling pathway affected by the selected circRNAs, while other pathways like hormone-associated and lipid metabolism-related were also involved in the carcinogenic axes of the highly expressed circRNAs. Survival rate analysis in PCa patients by the Kaplan-Meier curve unveiled a positive correlation between higher expression of cognate genes corresponding to anti-carcinogenic circRNAs (hsa_circ_0006410, hsa_circ_AKAP7 and hsa_circ_0005848) and better overall survival (OS) in PCa patients. Also, Yu et al. (49) identified 13 circRNAs in association with resistance to enzalutamide as an androgen deprivation therapy (ADP) drug used against PCa. Six miRNAs, 167 mRNAs, and 10 hub genes were identified as targets of the circRNAs. Among them, 8 prognostic-associated mRNAs were shown to be associated with survival rates in PCa patients, also with an AUC of 0.816 confirming the accuracy of miRNAs signature in detection of PCa. Additionally, knockdown experiments revealed that circRNA hsa_circ_0047641 promote PCa cell proliferation, migration, and invasion. CircMBOAT2 is another circRNA participating in the pathogenesis of PCa through increasing cell proliferation, migration, and invasion of malignant cells. This circRNA significantly up-regulates mTOR expression through sequestering miR-1271-5p, leading to the activation of the PI3K/Akt cascade (50).
CircSLC19A1 is another up-regulated circRNA in PCa tissues. CircSLC19A1 knock down has suppressed viability of PCa cells and their proliferation through modulation of miR-326/MAPK1 axis (51). CircABCC4 is an example of up-regulated circRNAs in PCa tissues and cell lines which enhances expression of FOXP4 through sequestering miR-1182. CircABCC4 silencing has inhibited proliferation of PCa as well as their migratory potential and invasiveness. Besides, circABCC4 knock down has attenuated growth of PCa in vivo. Cumulatively, circABCC4 accelerates malignant behavior of PCa (52).
Table 1 summarizes the studies on a set of up-regulated circRNAs in PCa.

Table 1 Up-regulated circRNAs in PCa.
Decreased expression in high throughput RNA analyses of PCa tissues compared to paired NATs draws attention to a second group of dysregulated circRNAs. Down-regulated circRNAs exhibit anti-oncogenic behaviors in experimental analyses inhibiting proliferation, migration, invasion, and metastasis of PCa cells. Sponge activity is seen for a majority of circRNAs which mainly show reverse regulation on downstream miRNAs. Inhibited miRNAs are mainly oncogenic RNAs which activate their corresponding oncogenes or inactivate related tumor suppressors. Mediator miRNAs, predominantly act via a signaling axis which firstly regulate expression a downstream protein, and itself makes changes to a signaling pathway. Affected known signaling pathways in PCa like MEK/ERK and Wnt/β-Catenin have been reported in a handful of studies on down-regulated circRNAs (see Table 2).
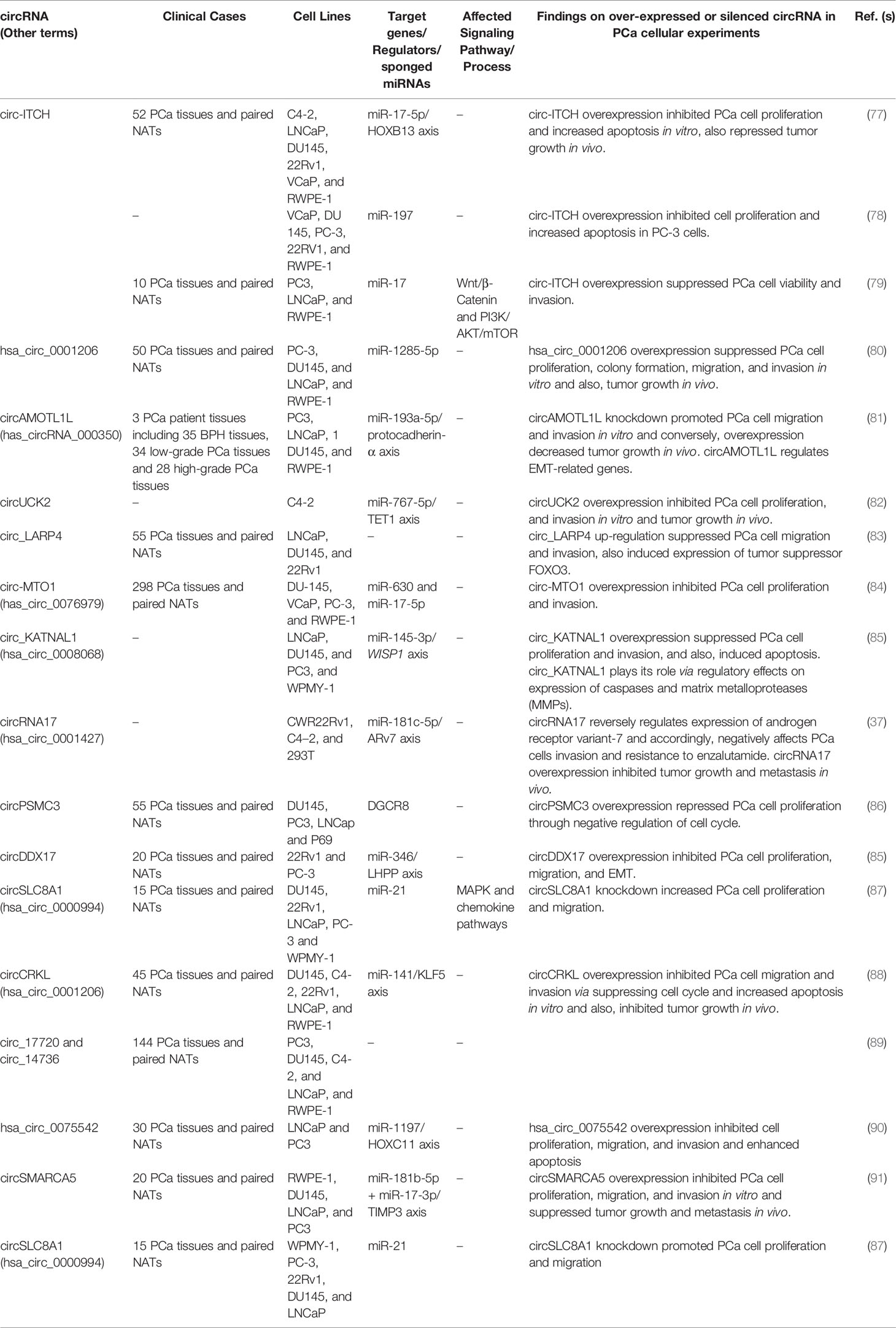
Table 2 Down-regulated circRNAs in PCa.
Circular RNA Itchy E3 ubiquitin protein ligase (circ-ITCH) is an example for down-regulated circRNAs in PCa which has been studied in four distinct experiments (77–79). Wang et al. (92) demonstrated that circ-ITCH down-regulation increases PCa cell proliferation and decreases apoptosis in vitro, while its up-regulation decreases cell proliferation and in vivo tumor growth. Luciferase assay showed direct interaction of circ-ITCH with microRNA miR-17-5p and reverse relationship between their expression levels, which reveals that circ-ITCH acts as sponge for downstream miR-17-5p. miR-17-5p, itself negatively regulates expression of HOXB13, which is known as a tumor suppressor gene being involved in development and progression of PCa. Yuan et al. (78) showed the same experimental results about consequences of circ-ITCH down-regulation, although miR-197 was identified as target miRNA for circ-ITCH. Also, miR-17 was detected as target of circ-ITCH in a study by Li et al. (79). Furthermore, they demonstrated that down-regulation of circ-ITCH is associated with up-regulation of expression of proteins involved in β-catenin, p-AKT, and p-mTOR signaling pathways indicating that circ-ITCH negatively regulates these pathways which have role in the progression of various tumors like PCa. In another study, Huang et al. (40) assessed the correlation between circ-ITCH expression and clinicopathological features, survival and prognosis in PCa. Direct association between low circ-ITCH levels and more aggressive clinicopathological features, poor survival, and unfavorable prognosis confirmed the experimental studies identifying circ-ITCH as a tumor suppressor circRNA in PCa. circAMOTL1L is another down-regulated circRNA in PCa. Its down-regulation has promoted PCa cell migration and invasion in vitro, while its overexpression has decreased tumor growth in vivo. circAMOTL1L has been shown to regulate expression of EMT-related genes (81).
Figure 1 illustrates the role of several circRNAs in PCa via modulating the PI3K/AKT/mTOR and MAPK/ERK pathways.
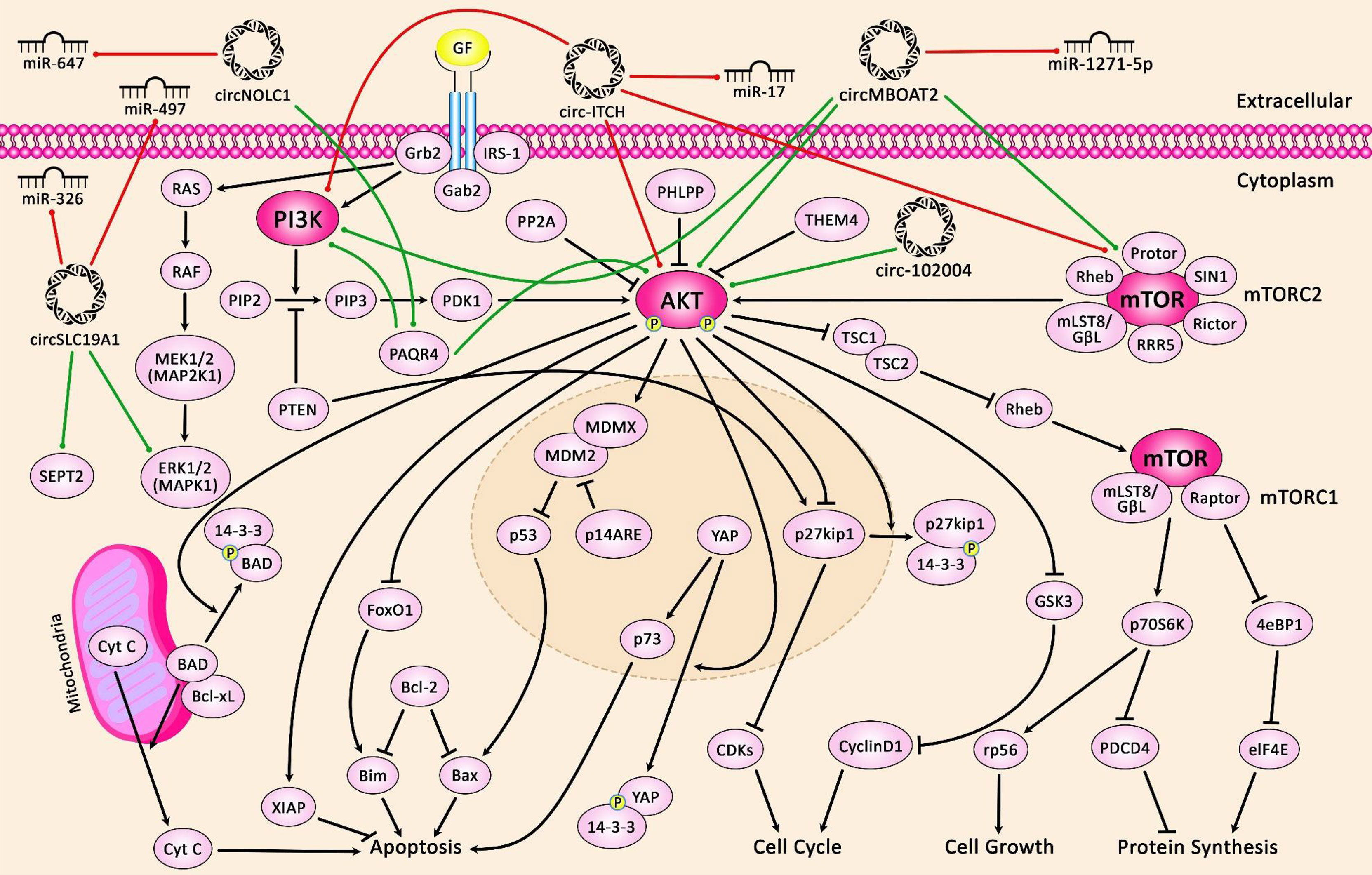
Figure 1 A schematic diagram of the crosstalk between circRNAs and PI3K/AKT/mTOR and MAPK/ERK signaling pathways in prostate cancer. The figure suggests that extracellular circRNAs enter cells.
PI3K signaling cascade linking RTK signaling results in downstream activation of PI3K/AKT/mTOR, elevating cell proliferation and survival. Besides, the MAPK-ERK signaling cascade also called the Raf/MEK/ERK pathway, is the main signal pathway of the MAPK signal cascade. The main MAPK/ERK kinase kinase (MEKK) components are the Raf family members Raf-1, A-Raf and B-Raf. Activated Raf activates MEK-1/2 by phosphorylating serine residues. Moreover, MEK-1/2 upregulates ERK-1/2 through phosphorylating the threonine and tyrosine residues of ERK-1/2. Activated ERK can regulate the phosphorylation of some nuclear transcription factors that are directly involved in the modulation of cell proliferation and differentiation. A recent study has demonstrated that overexpression of circMBOAT2 significantly upregulates mTOR expression via sponging miR-1271-5p, leading to the activation of the PI3K/Akt cascade, eventually elevating the cell proliferation, migration, and invasion of prostate cancer (50). Another research has denoted that circSLC19A1 elevates the expression level of MAPK1 by downregulating the miR-326 expression, thereby promoting prostate cancer cell proliferation, migration and invasion (51). Green arrows indicate upregulation of target genes modulated via circRNAs; red arrows depict inhibition by them.
As discussed above, circRNAs are found in extracellular vesicles extracted from plasma, exhibit high resistance to degradation and so, are detected in high concentration in easily received liquid biopsies suggesting potential application as biomarkers in diagnosis of various types of cancer (93). Early, quick, and minimal or non-invasive diagnosis based on detection of dysregulated circRNAs in human bio-specimens like blood and urine (94) makes it possible to provide scheduled and real time monitoring of the responses to treatment and prediction of prognosis in PCa patients. Accordingly, early treatment improves patient survival and so, good prognosis could be predicted for the patients. Both classes of dysregulated circRNAs including up- and down-regulated types can be detected in PCa patient samples and used as clinical biomarkers.
Wang et al. (89) have developed the first bioinformatics-based prognosis model for prediction of biochemical recurrence (BCR) in PCa which used a signature comprised of 8 circRNAs. Among them, two circRNAs including circ_17720 and circ_14736 were detected in exosomes extracted from plasma samples of PCa patients. Furthermore, both exhibited down-regulation in PCa tissues compared to NATs. Experimental analyses revealed that they repress PCa cell proliferation. Survival analysis via Kaplan-Meier for eight circRNAs showed that up-regulated circRNA correlated with unfavorable BCR-free survival and those with down-regulation changes were associated with less BCR. The area under curve (AUC) in Receiver-operating characteristic (ROC) for the signature was reported to be 0.799.
Greene et al. (95) profiled circRNAs in enzalutamide-chemoresistance in LNCaP PCa cell lines using high-throughput RNA sequencing. In bioinformatics analyses, five aberrantly expressed circRNAs were identified in enzalutamide-resistant LNCaP cells. Among them, hsa_circ_0004870 showed diminished expression in cells with high levels of androgen receptor (AR) compared to low AR-expressing cells and also, in malignant cells related to benign LNCaP cells. The corresponding BMP39 gene was also showed down-regulation in enzalutamide-resistant cells. Chen et al. (96) showed that a circRNA signature not only can distinguish PCa tissues from healthy prostate tissues, but also help distinguishment of PCa subtypes. Several circRNAs have shown dysregulation in accordance with Gleason score or correlated with advancement of clinicopathological features in PCa patients (see Table 3). Mao et al. (98) demonstrated that increased circPDHX expression levels in PCa tissues is correlated with malignant clinicopathological features in PCa patients. Kaplan-Meier analysis to assess the association between circPDHX expression and prognosis revealed that PCa patients with elevated circPDHX levels had poorer survival relative to patients with low levels. Univariate and multivariate regression analyses also showed that circPDHX high expression level along with advanced Gleason score act as independent prognostic factors for PCa patients predicting poorer survival. In diagnostic analyses, acceptable values of AOC in ROC curve, sensitivity, and specificity of 0.64, 80.0%, and 58.7%, respectively, were found for circPDHX showing promising results especially in distinguishment of PCa from healthy people. He et al. (100) evaluated expression of circrNAs in urinary extracellular vesicles. Their study has indicated the accuracy of a urine circRNA classifier (Ccirc) composed of circPLXDC2, circSCAF8, circPDLIM5, circCCNT2, and circSCAMP1 in detection of PCa. Their results demonstrated higher accuracy of Ccirc compared to that for two care risk calculators. Also, the Ccirc showed better value in prediction of high grades of PCa in combination with risk calculators relative to that of standards alone. In another study, Zhong et al. (97) identified 160 autophagy-related circRNAs, then constructed a circRNA signature containing five circRNAs hsa_circ_0001747, hsa_circ_0002100, hsa_circ_0000280, hsa_circ_0000437, and hsa_circ_0001085 with aberrant expression between high risk and low risk groups of PCa patients. Univariate and multivariate Cox regression analyses demonstrated the signature as an independent prognostic indicator in PCa patients. Also, ROC curve showed higher AUC values for the signature compared to conventional indicators like PSA, age, clinicopathological T stage, and Gleason score. Among the signature circRNAs, hsa_circ_0001747 was identified in association with a higher number of autophagy-related genes and its knockdown in experimental validation promoted PCa cell proliferation in vitro and in vivo through autophagy augmentation.
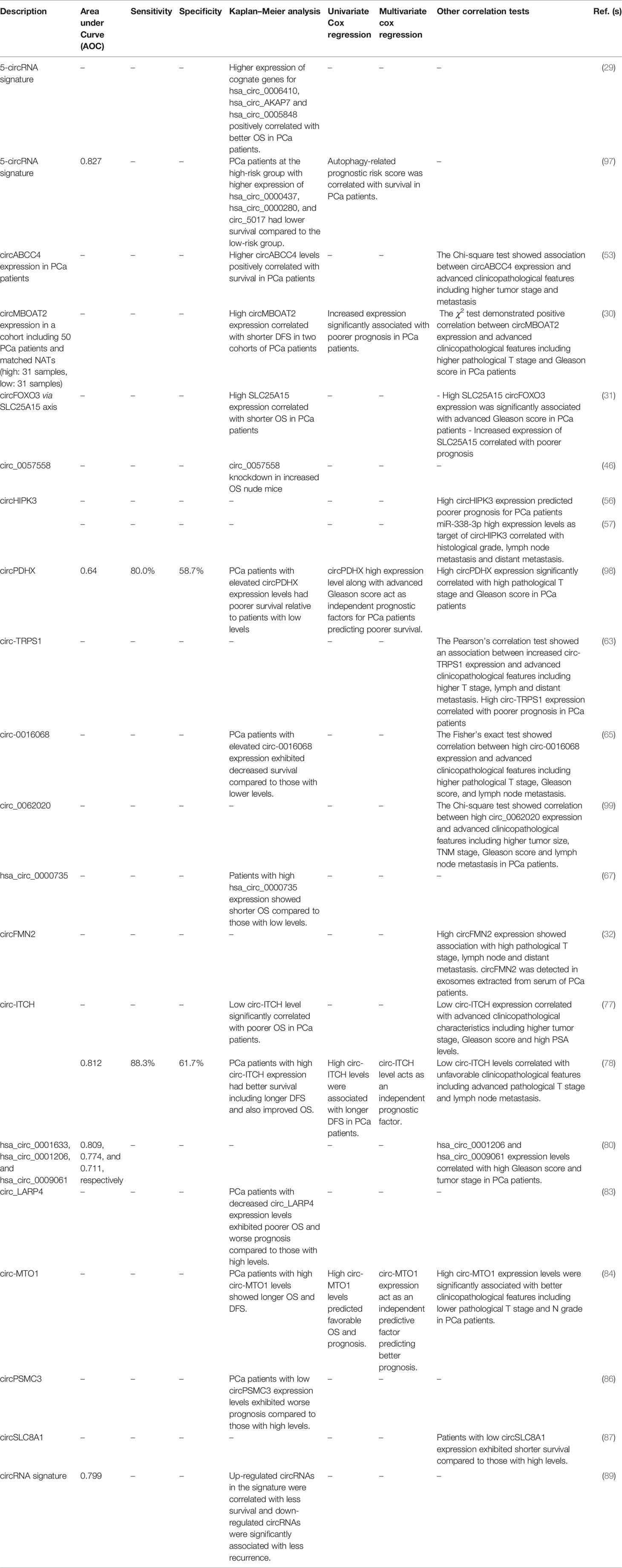
Table 3 circRNAs with diagnostic or prognostic values in PCa.
The prognostic value of circRNAs has also assessed in PCa. For instance, expression levels of hsa_circ_0000437, hsa_circ_0000280, and circ_5017 have been correlated with poor survival of patients (97). Moreover, expression of circABCC4 has been associated with advanced clinicopathological features including higher tumor stage, metastasis and poor clinical outcomes (53). Over-expression of circMBOAT2 has also indicated shorted disease-free survival in two independent cohorts of PCa patients (30). Similarly, over-expression of circFOXO3 has been associated with advanced Gleason score and shorter overall survival of PCa patients (31).
Taken together, these results suggest circRNAs as ideal candidates to be used as biomarkers for diagnosis, prediction of prognosis and also provide therapeutical targets in treatment of PCa. Further studies are required to bring the circRNAs to clinical settings as useful tools with diagnostic, prognostic and therapeutical applications. Table 3 shows the studies which have assessed the diagnostic, prognostic or clinical significance values of circRNAs in PCa.
CircRNAs are a novel type of ncRNAs for which some regulatory functions are known. Changes in their expression have been found in several disorders especially cancer. High-throughput technologies have helped identification of a vast number of circRNAs which exhibit dysregulation including down- or up-regulation in cancer tissues compared to NATs. Experimental and functional analyses have shown up-regulated circRNAs act as oncogenes, which promote tumorigenicity in cell studies. Conversely, down-regulated circRNAs play role as tumor suppressors and inhibit tumorigenic behaviors of cancer cell lines. In the majority of circRNAs, existence of interactions between them and miRNAs has revealed a mechanism through which circRNAs exert their roles and regulate cellular processes especially cell cycle. Precise understanding of action mechanisms may help finding therapeutic targets for cancer therapy. Clinical assessments, also have unveiled circRNAs as ideal candidates for diagnostic and prognostic applications. Similar to many cancers, a number of circRNAs have been identified to be dysregulated in PCa. In this review, we assessed the preeminent studies on the role of circRNAs in PCa in two categories of down- (Table 1) and up-regulated (Table 2) circRNAs focusing on functional experiments. Oncogenic circRNAs promote tumorigenicity via increasing cell proliferation, migration, and invasion in vitro and tumor growth and metastasis in vivo. Sponged miRNAs have been recognized in the majority of studies, through them circRNAs exert their roles via an axis which finally affects expression or activity of oncogenes or tumor suppressors, or directly influence the cell cycle.
miR-204-5p, miR-3160-5p, miR-548, miR-1182, miR-1271-5p, miR‐29a‐3p, miR-206, miR-185-3p, miR-29a, miR-193a-3p, miR-338-3p, miR-1299, miR-501-3p, miR-378a-3p, miR-124-3p, miR‐152‐3p, miR-647, miR-515-5p, miR-330-3p, miR-30b-5p, miR-615-5p, miR-326, miR-497, miR-7, miR-1238 and miR-646 are among the most important cancer-related miRNAs being sponged by circRNAs. The sponging effects of circRNAs on these miRNAs have a crucial role in the regulation of activity of cancer-related pathways. CircFOXO3/miR-29a‐3p, circ_0057558/miR-206, circ_0088233/miR-185-3p, circMYLK/miR-29a, circHIPK3/miR-193a-3p, circHIPK3/miR-338-3p, circHIPK3/miR-212, circ_0006404/miR-1299, circ-ZNF609/miR-501-3p, circPDHX/miR-378a-3p, circ-TRPS1/miR-124-3p, circNOLC1/miR-647 are examples of circRNAs/miRNAs axes with crucial roles in the pathogenesis of PCa.
Furthermore, diagnostic and prognostic values of circRNAs have been reviewed. Acceptable values have been reported for a set of circRNAs in PCa which suggest diagnostic and prognostic potentials of circRNAs. Some studies have proposed the role of circRNAs for easy, quick and less invasive diagnosis and prediction of prognosis of PCa patients based on their expression levels in liquid biopsies. However, based on the heterogeneous pattern of expression of circRNAs among patients, multi-gene panels are more promising than individual circRNAs. In addition, they may have therapeutic potentials, however further studies are required to utilize the potentials of circRNAs in clinical settings. It is also necessary to appraise expression of circRNAs in different settings to find possible factors that affect their expression in various cellular contexts.
SN, ArB, and SG-F wrote the draft and revised it. MT designed and supervised the study. AbB, BMH, and EJ collected the data and designed the figures and tables. All authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA: Cancer J Clin (2021) 71(3):209–49. doi: 10.3322/caac.21660
2. Rawla P. Epidemiology of Prostate Cancer. World J Oncol (2019) 10(2):63–89. doi: 10.14740/wjon1191
3. Schymura MJ, Sun L, Percy-Laurry A. Prostate Cancer Collaborative Stage Data Items–Their Definitions, Quality, Usage, and Clinical Implications: A Review of SEER Data for 2004-2010. Cancer (2014) 120 Suppl 23:3758–70. doi: 10.1002/cncr.29052
4. Gleason DF. Histologic Grading of Prostate Cancer: A Perspective. Hum Pathol (1992) 23(3):273–9. doi: 10.1016/0046-8177(92)90108-F
5. Carter HB. Prostate-Specific Antigen (PSA) Screening for Prostate Cancer: Revisiting the Evidence. JAMA (2018) 319(18):1866–8. doi: 10.1001/jama.2018.4914
6. Malik B, Feng FY. Long Noncoding RNAs in Prostate Cancer: Overview and Clinical Implications. Asian J Androl (2016) 18(4):568–74. doi: 10.4103/1008-682X.177123
7. Hasegawa T, Lewis H, Esquela-Kerscher A. Chapter 12 - The Role of Noncoding RNAs in Prostate Cancer. In: Laurence J, editor. Translating MicroRNAs to the Clinic. Boston: Academic Press (2017). p. 329–69.
8. Sanger HL, Klotz G, Riesner D, Gross HJ, Kleinschmidt AK. Viroids are Single-Stranded Covalently Closed Circular RNA Molecules Existing as Highly Base-Paired Rod-Like Structures. Proc Natl Acad Sci USA (1976) 73(11):3852–6. doi: 10.1073/pnas.73.11.3852
9. Hsu M-T, Coca-Prados M. Electron Microscopic Evidence for the Circular Form of RNA in the Cytoplasm of Eukaryotic Cells. Nature (1979) 280(5720):339–40. doi: 10.1038/280339a0
10. Chen L-L. The Biogenesis and Emerging Roles of Circular RNAs. Nat Rev Mol Cell Biol (2016) 17(4):205–11. doi: 10.1038/nrm.2015.32
11. Suzuki H, Zuo Y, Wang J, Zhang MQ, Malhotra A, Mayeda A. Characterization of RNase R-Digested Cellular RNA Source That Consists of Lariat and Circular RNAs From pre-mRNA Splicing. Nucleic Acids Res (2006) 34(8):e63–e. doi: 10.1093/nar/gkl151
12. López-Jiménez E, Rojas AM, Andrés-León E. Xiao J, editor. Circular RNAs: Biogenesis and Functions. Singapore: Springer Singapore (2018). p. 17–33.
13. Szabo L, Salzman J. Detecting Circular RNAs: Bioinformatic and Experimental Challenges. Nat Rev Genet (2016) 17(11):679–92. doi: 10.1038/nrg.2016.114
14. Boss M, Arenz C. A Fast and Easy Method for Specific Detection of Circular RNA by Rolling-Circle Amplification. Chembiochem Eur J Chem Biol (2020) 21(6):793–6. doi: 10.1002/cbic.201900514
15. Zeng X, Lin W, Guo M, Zou Q. A Comprehensive Overview and Evaluation of Circular RNA Detection Tools. PLoS Comput Biol (2017) 13(6):e1005420. doi: 10.1371/journal.pcbi.1005420
16. Asghari H, Lin YY, Xu Y, Haghshenas E, Collins CC, Hach F. CircMiner: Accurate and Rapid Detection of Circular RNA Through Splice-Aware Pseudo-Alignment Scheme. Bioinf (Oxford England) (2020) 36(12):3703–11. doi: 10.1093/bioinformatics/btaa232
17. Jiang F, Hong F, Shah MW, Shen X. Circular RNAs as Diagnostic Biomarkers in Gastric Cancer: A Meta-Analysis Review. Pathol Res Pract (2019) 215(6):152419. doi: 10.1016/j.prp.2019.04.011
18. Salzman J, Gawad C, Wang PL, Lacayo N, Brown PO. Circular RNAs are the Predominant Transcript Isoform From Hundreds of Human Genes in Diverse Cell Types. PLoS One (2012) 7(2):e30733. doi: 10.1371/journal.pone.0030733
19. Hua JT, Chen S, He HH. Landscape of Noncoding RNA in Prostate Cancer. Trends Genet (2019) 35(11):840–51. doi: 10.1016/j.tig.2019.08.004
20. Salzman J, Chen RE, Olsen MN, Wang PL, Brown PO. Cell-Type Specific Features of Circular RNA Expression. PLoS Genet (2013) 9(9):e1003777. doi: 10.1371/journal.pgen.1003777
21. Memczak S, Jens M, Elefsinioti A, Torti F, Krueger J, Rybak A, et al. Circular RNAs are a Large Class of Animal RNAs With Regulatory Potency. Nature (2013) 495(7441):333–8. doi: 10.1038/nature11928
22. Guo JU, Agarwal V, Guo H, Bartel DP. Expanded Identification and Characterization of Mammalian Circular RNAs. Genome Biol (2014) 15(7):409. doi: 10.1186/s13059-014-0409-z
23. Ding Y, Lu C, Zhang W, Wang Y, Li Y, Zhu Y, et al. The Emerging Role of Circular RNAs in Cardiovascular Diseases. J Physiol Biochem (2021) 77:343–53. doi: 10.1007/s13105-021-00807-y
24. Tian M, Cao Z, Pang H. Circular RNAs in Sudden Cardiac Death Related Diseases: Novel Biomarker for Clinical and Forensic Diagnosis. Mol (Basel Switzerland) (2021) 26(4). doi: 10.3390/molecules26041155
25. Chen D, Hao S, Xu J. Revisiting the Relationship Between Alzheimer's Disease and Cancer With a circRNA Perspective. Front Cell Dev Biol (2021) 9:647197. doi: 10.3389/fcell.2021.647197
26. Mahmoudi E, Green MJ, Cairns MJ. Dysregulation of circRNA Expression in the Peripheral Blood of Individuals With Schizophrenia and Bipolar Disorder. J Mol Med (Berlin Germany) (2021) 99:981–91. doi: 10.21203/rs.3.rs-231116/v1
27. Chen XT, Li ZW, Zhao X, Li ML, Hou PF, Chu SF, et al. Role of Circular RNA in Kidney-Related Diseases. Front Pharmacol (2021) 12:615882. doi: 10.3389/fphar.2021.615882
28. Shi L, Zhang H, Sun J, Gao X, Liu C. CircSEC24A Promotes IL-1β-Induced Apoptosis and Inflammation in Chondrocytes by Regulating miR-142-5p/SOX5 Axis. Biotechnol Appl Biochem (2021). doi: 10.1002/bab.2145
29. Zhai X, Zhang Y, Xin S, Cao P, Lu J. Insights Into the Involvement of Circular RNAs in Autoimmune Diseases. Front Immunol (2021) 12:622316. doi: 10.3389/fimmu.2021.622316
30. Shi J, Liu C, Chen C, Guo K, Tang Z, Luo Y, et al. Circular RNA Circmboat2 Promotes Prostate Cancer Progression via a miR-1271-5p/mTOR Axis. Aging (Albany NY) (2020) 12(13):13255–80. doi: 10.18632/aging.103432
31. Kong Z, Wan X, Lu Y, Zhang Y, Huang Y, Xu Y, et al. Circular RNA Circfoxo3 Promotes Prostate Cancer Progression Through Sponging miR-29a-3p. J Cell Mol Med (2020) 24(1):799–813. doi: 10.1111/jcmm.14791
32. Shan G, Shao B, Liu Q, Zeng Y, Fu C, Chen A, et al. Circfmn2 Sponges miR-1238 to Promote the Expression of LIM-Homeobox Gene 2 in Prostate Cancer Cells. Mol Ther Nucleic Acids (2020), 21:133–46. doi: 10.1016/j.omtn.2020.05.008
33. Zheng Y, Li JX, Chen CJ, Lin ZY, Liu JX, Lin FJ. Extracellular Vesicle-Derived Circ_SLC19A1 Promotes Prostate Cancer Cell Growth and Invasion Through the miR-497/Septin 2 Pathway. Cell Biol Int (2020) 44(4):1037–45. doi: 10.1002/cbin.11303
34. Xia Q, Ding T, Zhang G, Li Z, Zeng L, Zhu Y, et al. Circular RNA Expression Profiling Identifies Prostate Cancer- Specific circRNAs in Prostate Cancer. Cell Physiol Biochem (2018) 50(5):1903–15. doi: 10.1159/000494870
35. Greene J, Baird A-M, Lim M, Flynn J, McNevin C, Brady L, et al. Differential CircRNA Expression Signatures May Serve as Potential Novel Biomarkers in Prostate Cancer. Front Cell Dev Biol (2021) 9:605686–. doi: 10.3389/fcell.2021.605686
36. Chen Q, Shen H, Zhu X, Liu Y, Yang H, Chen H, et al. A Nuclear lncRNA Linc00839 as a Myc Target to Promote Breast Cancer Chemoresistance via PI3K/AKT Signaling Pathway. Cancer Sci (2020) 111(9):3279. doi: 10.1111/cas.14555
37. Wu G, Sun Y, Xiang Z, Wang K, Liu B, Xiao G, et al. Preclinical Study Using Circular RNA 17 and Micro RNA 181c-5p to Suppress the Enzalutamide-Resistant Prostate Cancer Progression. Cell Death Dis (2019) 10(2):37–. doi: 10.1038/s41419-018-1048-1
38. Kong Z, Wan X, Zhang Y, Zhang P, Zhang Y, Zhang X, et al. Androgen-Responsive Circular RNA Circsmarca5 is Up-Regulated and Promotes Cell Proliferation in Prostate Cancer. Biochem Biophys Res Commun (2017) 493(3):1217–23. doi: 10.1016/j.bbrc.2017.07.162
39. Xu H, Sun Y, You B, Huang C-P, Ye D, Chang C. Androgen Receptor Reverses the Oncometabolite R-2-Hydroxyglutarate-Induced Prostate Cancer Cell Invasion via Suppressing the circRNA-51217/miRNA-646/Tgfβ1/P-Smad2/3 Signaling. Cancer Lett (2020) 472:151–64. doi: 10.1016/j.canlet.2019.12.014
40. Huang E, Chen X, Yuan Y. Downregulated Circular RNA Itchy E3 Ubiquitin Protein Ligase Correlates With Advanced Pathologic T Stage, High Lymph Node Metastasis Risk and Poor Survivals in Prostate Cancer Patients. Cancer Biomarkers Section A Dis Markers (2019) 26(1):41–50. doi: 10.3233/CBM-182111
41. Huang A, Zheng H, Wu Z, Chen M, Huang Y. Circular RNA-Protein Interactions: Functions, Mechanisms, and Identification. Theranostics (2020) 10(8):3503–17. doi: 10.7150/thno.42174
42. Janas T, Janas MM, Sapoń K, Janas T. Mechanisms of RNA Loading Into Exosomes. FEBS Lett (2015) 589(13):1391–8. doi: 10.1016/j.febslet.2015.04.036
43. Dudekula DB, Panda AC, Grammatikakis I, De S, Abdelmohsen K, Gorospe M. CircInteractome: A Web Tool for Exploring Circular RNAs and Their Interacting Proteins and microRNAs. RNA Biol (2016) 13(1):34–42. doi: 10.1080/15476286.2015.1128065
44. López-Jiménez E, Rojas AM, Andrés-León E. RNA Sequencing and Prediction Tools for Circular RNAs Analysis. Adv Exp Med Biol (2018) 1087:17–33. doi: 10.1007/978-981-13-1426-1_2
45. Panda AC. Circular RNAs Act as miRNA Sponges. Adv Exp Med Biol (2018) 1087:67–79. doi: 10.1007/978-981-13-1426-1_6
46. Ding T, Zhu Y, Jin H, Zhang P, Guo J, Zheng J. Circular RNA Circ_0057558 Controls Prostate Cancer Cell Proliferation Through Regulating miR-206/USP33/c-Myc Axis. Front Cell Dev Biol (2021) 9:644397–. doi: 10.3389/fcell.2021.644397
47. Du S, Zhang P, Ren W, Yang F, Du C. Circ-ZNF609 Accelerates the Radioresistance of Prostate Cancer Cells by Promoting the Glycolytic Metabolism Through miR-501-3p/HK2 Axis. Cancer Manag Res (2020) 12:7487–99. doi: 10.2147/CMAR.S257441
48. Zhang L, Zhang W, Li H, Tang X, Xu S, Wu M, et al. Five Circular RNAs in Metabolism Pathways Related to Prostate Cancer. Front Genet (2021) 12:636419–. doi: 10.3389/fgene.2021.636419
49. Yu J, Sun S, Mao W, Xu B, Chen M. Identification of Enzalutamide Resistance-Related circRNA-miRNA-mRNA Regulatory Networks in Patients With Prostate Cancer. Onco Targets Ther (2021) 14:3833–48. doi: 10.2147/OTT.S309917
50. Shi J, Liu C, Chen C, Guo K, Tang Z, Luo Y, et al. Circular RNA Circmboat2 Promotes Prostate Cancer Progression via a miR-1271-5p/mTOR Axis. Aging (Albany NY) (2020) 12(13):13255. doi: 10.18632/aging.103432
51. Huang B, Zhou D, Huang X, Xu X, Xu Z. Silencing Circslc19a1 Inhibits Prostate Cancer Cell Proliferation, Migration and Invasion Through Regulating miR-326/MAPK1 Axis. Cancer Manag Res (2020) 12:11883–95. doi: 10.2147/CMAR.S267927
52. Huang C, Deng H, Wang Y, Jiang H, Xu R, Zhu X, et al. Circular RNA Circabcc4 as the ceRNA of miR-1182 Facilitates Prostate Cancer Progression by Promoting FOXP4 Expression. J Cell Mol Med (2019) 23(9):6112–9. doi: 10.1111/jcmm.14477
53. Huang C, Deng H, Wang Y, Jiang H, Xu R, Zhu X, et al. Circular RNA Circabcc4 as the ceRNA of miR-1182 Facilitates Prostate Cancer Progression by Promoting FOXP4 Expression. J Cell Mol Med (2019) 23(9):6112–9. doi: 10.1111/jcmm.14477
54. Deng ZH, Yu GS, Deng KL, Feng ZH, Huang Q, Pan B, et al. Hsa_circ_0088233 Alleviates Proliferation, Migration, and Invasion of Prostate Cancer by Targeting hsa-miR-185-3p. Front Cell Dev Biol (2020) 8:528155. doi: 10.3389/fcell.2020.528155
55. Dai Y, Li D, Chen X, Tan X, Gu J, Chen M, et al. Circular RNA Myosin Light Chain Kinase (MYLK) Promotes Prostate Cancer Progression Through Modulating Mir-29a Expression. Med Sci Monitor Int Med J Exp Clin Res (2018) 24:3462–71. doi: 10.12659/MSM.908009
56. Li C, Zheng H, Hou W, Bao H, Xiong J, Che W, et al. Long non-Coding RNA Linc00645 Promotes TGF-β-Induced Epithelial–Mesenchymal Transition by Regulating miR-205-3p-ZEB1 Axis in Glioma. Cell Death Dis (2019) 10(10):717. doi: 10.1038/s41419-019-1948-8
57. Cai C, Zhi Y, Wang K, Zhang P, Ji Z, Xie C, et al. CircHIPK3 Overexpression Accelerates the Proliferation and Invasion of Prostate Cancer Cells Through Regulating miRNA-338-3p. Onco Targets Ther (2019) 12:3363–72. doi: 10.2147/OTT.S196931
58. Tang Y, Liu J, Li X, Wang W. Exosomal circRNA HIPK3 Knockdown Inhibited Cell Proliferation and Metastasis in Prostate Cancer by Regulating miR-212/BMI-1 Pathway. J Biosci (2021) 46.
59. Chen Y, Yang F, Fang E, Xiao W, Mei H, Li H, et al. Circular RNA Circago2 Drives Cancer Progression Through Facilitating HuR-Repressed Functions of AGO2-miRNA Complexes. Cell Death Differ (2019) 26(7):1346–64. doi: 10.1038/s41418-018-0220-6
60. Li P, Wang Z, Li S, Wang L. Circ_0006404 Accelerates Prostate Cancer Progression Through Regulating miR-1299/CFL2 Signaling. Onco Targets Ther (2021) 14:83–95. doi: 10.2147/OTT.S277831
61. Si-Tu J, Cai Y, Feng T, Yang D, Yuan S, Yang X, et al. Upregulated Circular RNA Circ-102004 That Promotes Cell Proliferation in Prostate Cancer. Int J Biol Macromol. (2019) 122:1235–43. doi: 10.1016/j.ijbiomac.2018.09.076
62. Chen W, Cen S, Zhou X, Yang T, Wu K, Zou L, et al. Circular RNA CircNOLC1, Upregulated by NF-KappaB, Promotes the Progression of Prostate Cancer via miR-647/PAQR4 Axis. Front Cell Dev Biol (2021) 8:624764–. doi: 10.3389/fcell.2020.624764
63. Sha J, Xia L, Han Q, Chi C, Zhu Y, Pan J, et al. Downregulation of Circ-TRPS1 Suppressed Prostatic Cancer Prognoses by Regulating miR-124-3p/EZH2 Axis-Mediated Stemness. Am J Cancer Res (2020) 10(12):4372–85. doi: 10.21203/rs.3.rs-48783/v1
64. Zhang Y, Shi Z, Li Z, Wang X, Zheng P, Li H. Circ_0057553/miR-515-5p Regulates Prostate Cancer Cell Proliferation, Apoptosis, Migration, Invasion and Aerobic Glycolysis by Targeting Yes1. OncoTargets Ther (2020) 13:11289–99. doi: 10.2147/OTT.S272294
65. Li Q, Wang W, Zhang M, Sun W, Shi W, Li F. Circular RNA Circ-0016068 Promotes the Growth, Migration, and Invasion of Prostate Cancer Cells by Regulating the miR-330-3p/BMI-1 Axis as a Competing Endogenous RNA. Front Cell Dev Biol (2020) 8:827–. doi: 10.3389/fcell.2020.00827
66. Cai F, Li J, Zhang J, Huang S. Knockdown of Circ_CCNB2 Sensitizes Prostate Cancer to Radiation Through Repressing Autophagy by the miR-30b-5p/KIF18A Axis. Cancer Biother Radiopharmaceut (2020). doi: 10.1089/cbr.2019.3538
67. Gao Y, Liu J, Huan J, Che F. Downregulation of Circular RNA Hsa_Circ_0000735 Boosts Prostate Cancer Sensitivity to Docetaxel via Sponging miR-7. Cancer Cell Int (2020) 20:334–. doi: 10.1186/s12935-020-01421-6
68. Umemori M, Kurata M, Yamamoto A, Yamamoto K, Ishibashi S, Ikeda M, et al. The Expression of MYC is Strongly Dependent on the Circular PVT1 Expression in Pure Gleason Pattern 4 of Prostatic Cancer. Med Mol Morphol (2020) 53(3):156–67. doi: 10.1007/s00795-020-00243-9
69. Jiang H, Lv DJ, Song XL, Wang C, Yu YZ, Zhao SC. Upregulated Circzmiz1 Promotes the Proliferation of Prostate Cancer Cells and is a Valuable Marker in Plasma. Neoplasma (2020) 67(1):68–77. doi: 10.4149/neo_2019_190213N116
70. Gong L, Tang Y, Jiang L, Tang W, Luo S. Regulation of Circgolph3 and its Binding Protein CBX7 on the Proliferation and Apoptosis of Prostate Cancer Cells. Biosci Rep (2020) 40(12):BSR20200936. doi: 10.1042/BSR20200936
71. Zeng L, Liu Y-M, Yang N, Zhang T, Xie H. Hsa_circRNA_100146 Promotes Prostate Cancer Progression by Upregulating TRIP13 via Sponging miR-615-5p. Front Mol Biosci (2021) 8:693477–. doi: 10.3389/fmolb.2021.693477
72. Chen H, Zhang P, Yu B, Liu J. The Circular RNA Circxpo1 Promotes Tumor Growth via Sponging MicroRNA-23a in Prostate Carcinoma. Front Oncol (2021) 11(2925). doi: 10.3389/fonc.2021.712145
73. Xu S, Lian Z, Zhang S, Xu Y, Zhang H. CircGNG4 Promotes the Progression of Prostate Cancer by Sponging miR-223 to Enhance EYA3/c-Myc Expression. Front Cell Dev Biol (2021) 9:684125. doi: 10.3389/fcell.2021.684125
74. Zhang H, Li M, Zhang J, Shen Y, Gui Q. Exosomal Circ-XIAP Promotes Docetaxel Resistance in Prostate Cancer by Regulating miR-1182/TPD52 Axis. Drug Des Devel Ther (2021) 15:1835–49. doi: 10.2147/DDDT.S300376
75. Yang D, Yang B, Zhu Y, Xia Q, Zhang Y, Zhu X, et al. Circular RNA-DPP4 Serves an Oncogenic Role in Prostate Cancer Progression Through Regulating miR-195/Cyclin D1 Axis. Cancer Cell Int (2021) 21(1):379. doi: 10.1186/s12935-021-02062-z
76. Pan J, Liu Z, Yang Z, Liang E, Fang C, Zhang D, et al. Circ_0001686 Promotes Prostate Cancer Progression by Up-Regulating SMAD3/TGFBR2 via miR-411-5p. World J Mens Health (2021) 0:39. doi: 10.5534/wjmh.200204
77. Zhu Q, Li Y, Guo Y, Hu L, Xiao Z, Liu X, et al. Long non-Coding RNA SNHG16 Promotes Proliferation and Inhibits Apoptosis of Diffuse Large B-Cell Lymphoma Cells by Targeting miR-497-5p/PIM1 Axis. J Cell Mol Med (2019) 23(11):7395–405. doi: 10.1111/jcmm.14601
78. Yuan Y, Chen X, Huang E. Upregulation of Circular RNA Itchy E3 Ubiquitin Protein Ligase Inhibits Cell Proliferation and Promotes Cell Apoptosis Through Targeting MiR-197 in Prostate Cancer. Technol Cancer Res Treat (2019) 18:1533033819886867. doi: 10.1177/1533033819886867
79. Li S, Yu C, Zhang Y, Liu J, Jia Y, Sun F, et al. Circular RNA Cir-ITCH Is a Potential Therapeutic Target for the Treatment of Castration-Resistant Prostate Cancer. BioMed Res Int (2020) 2020:7586521. doi: 10.1155/2020/7586521
80. Liu X, Sun N, Mo N, Lu S, Song E, Ren C, et al. Quercetin Inhibits Kidney Fibrosis and the Epithelial to Mesenchymal Transition of the Renal Tubular System Involving Suppression of the Sonic Hedgehog Signaling Pathway. Food Funct (2019) 10(6):3782–97. doi: 10.1039/C9FO00373H
81. Yang Z, Qu C-B, Zhang Y, Zhang W-F, Wang D-D, Gao C-C, et al. Dysregulation of P53-RBM25-Mediated Circamotl1l Biogenesis Contributes to Prostate Cancer Progression Through the Circamotl1l-miR-193a-5p-Pcdha Pathway. Oncogene (2019) 38(14):2516–32. doi: 10.1038/s41388-018-0602-8
82. Xiang Z, Xu C, Wu G, Liu B, Wu D. CircRNA-UCK2 Increased TET1 Inhibits Proliferation and Invasion of Prostate Cancer Cells Via Sponge MiRNA-767-5p. Open Med (Wars) (2019) 14:833–42. doi: 10.1515/med-2019-0097
83. Weng XD, Yan T, Liu CL. Circular RNA_LARP4 Inhibits Cell Migration and Invasion of Prostate Cancer by Targeting FOXO3A. Eur Rev Med Pharmacol Sci (2020) 24(10):5303–9. doi: 10.18632/aging.103432
84. Hu Y, Guo B. Circ-MTO1 Correlates With Favorable Prognosis and Inhibits Cell Proliferation, Invasion as Well as miR-17-5p Expression in Prostate Cancer. J Clin Lab Anal (2020) 34(3):e23086. doi: 10.1002/jcla.23086
85. Zheng Y, Chen CJ, Lin ZY, Li JX, Liu J, Lin FJ, et al. Circ_KATNAL1 Regulates Prostate Cancer Cell Growth and Invasiveness Through the miR-145-3p/WISP1 Pathway. Biochem Cell Biol Biochim Biol Cellulaire (2020) 98(3):396–404. doi: 10.1139/bcb-2019-0211
86. Dong JS, Wu B, Chen XH. Circ PSMC3 Inhibits Prostate Cancer Cell Proliferation by Downregulating DGCR8. Eur Rev Med Pharmacol Sci (2020) 24(5):2264–70. doi: 10.26355/eurrev_202003_20492
87. Wang D, Yan S, Wang L, Li Y, Qiao B. Circslc8a1 Acts as a Tumor Suppressor in Prostate Cancer via Sponging miR-21. BioMed Res Int (2021) 2021:6614591.
88. Nan C, Wang Y, Yang S, Chen Y. circCRKL Suppresses the Progression of Prostate Cancer Cells by Regulating the miR-141/KLF5 Axis. Pathol Res Practice (2020) 216(11):153182. doi: 10.1016/j.prp.2020.153182
89. Wang S, Su W, Zhong C, Yang T, Chen W, Chen G, et al. An Eight-CircRNA Assessment Model for Predicting Biochemical Recurrence in Prostate Cancer. Front Cell Dev Biol (2020) 8:599494. doi: 10.3389/fcell.2020.599494
90. Han Y, Wen X, Li X, Chen D, Peng L, Lai B, et al. Circular RNA Hsa_Circ_0075542 Acts as a Sponge for microRNA-1197 to Suppress Malignant Characteristics and Promote Apoptosis in Prostate Cancer Cells. Bioengineered (2021) 12(1):5620–31. doi: 10.1080/21655979.2021.1967064
91. Xie X, Sun F-K, Huang X, Wang C-H, Dai J, Zhao J-P, et al. Circsmarca5, Inhibits Prostate Cancer Proliferative, Migrative, and Invasive Capabilities via the miR-181b-5p/miR-17-3p-TIMP3 Axis. Aging (Albany NY) (2021) 13(15):19908–19. doi: 10.18632/aging.203408
92. Wang X, Wang R, Wu Z, Bai P. Circular RNA ITCH Suppressed Prostate Cancer Progression by Increasing HOXB13 Expression via Spongy miR-17-5p. Cancer Cell Int (2019) 19(1):328. doi: 10.1186/s12935-019-0994-8
93. Wang S, Zhang K, Tan S, Xin J, Yuan Q, Xu H, et al. Circular RNAs in Body Fluids as Cancer Biomarkers: The New Frontier of Liquid Biopsies. Mol Cancer (2021) 20(1):1–10. doi: 10.1186/s12943-020-01298-z
94. Bahn JH, Zhang Q, Li F, Chan TM, Lin X, Kim Y, et al. The Landscape of microRNA, Piwi-Interacting RNA, and Circular RNA in Human Saliva. Clin Chem (2015) 61(1):221–30. doi: 10.1373/clinchem.2014.230433
95. Greene J, Baird A-M, Casey O, Brady L, Blackshields G, Lim M, et al. Circular RNAs are Differentially Expressed in Prostate Cancer and are Potentially Associated With Resistance to Enzalutamide. Sci Rep (2019) 9(1):10739–. doi: 10.1038/s41598-019-47189-2
96. Chen S, Huang V, Xu X, Livingstone J, Soares F, Jeon J, et al. Widespread and Functional RNA Circularization in Localized Prostate Cancer. Cell (2019) 176(4):831–43.e22. doi: 10.1016/j.cell.2019.01.025
97. Zhong C, Wu K, Wang S, Long Z, Yang T, Zhong W, et al. Autophagy-Related circRNA Evaluation Reveals Hsa_Circ_0001747 as a Potential Favorable Prognostic Factor for Biochemical Recurrence in Patients With Prostate Cancer. Cell Death Dis (2021) 12(8):726. doi: 10.1038/s41419-021-04015-w
98. Mao Y, Li W, Hua B, Gu X, Pan W, Chen Q, et al. Circular RNA_PDHX Promotes the Proliferation and Invasion of Prostate Cancer by Sponging MiR-378a-3p. Front Cell Dev Biol (2021) 8:602707–. doi: 10.3389/fcell.2020.602707
99. Li H, Zhi Y, Ma C, Shen Q, Sun F, Cai C. Circ_0062020 Knockdown Strengthens the Radiosensitivity of Prostate Cancer Cells. Cancer Manag Res (2020) 12:11701–12. doi: 10.2147/CMAR.S273826
Keywords: circular RNAs, prostate cancer, diagnosis, prognosis, biomarker
Citation: Taheri M, Najafi S, Basiri A, Hussen BM, Baniahmad A, Jamali E and Ghafouri-Fard S (2021) The Role and Clinical Potentials of Circular RNAs in Prostate Cancer. Front. Oncol. 11:781414. doi: 10.3389/fonc.2021.781414
Received: 22 September 2021; Accepted: 20 October 2021;
Published: 05 November 2021.
Edited by:
Bianca Nitzsche, Charité – Universitätsmedizin Berlin, GermanyReviewed by:
Soichiro Yamamura, University of California, San Francisco, United StatesCopyright © 2021 Taheri, Najafi, Basiri, Hussen, Baniahmad, Jamali and Ghafouri-Fard. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Soudeh Ghafouri-Fard, cy5naGFmb3VyaWZhcmRAc2JtdS5hYy5pcg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.