 Zitong Yang
Zitong Yang Qinchen Li
Qinchen Li Xiangyi Zheng
Xiangyi Zheng Liping Xie
Liping Xie- Department of Urology, The First Affiliated Hospital, Zhejiang University School of Medicine, Hangzhou, China
The incidence of urological cancer has been gradually increasing in the last few decades. However, current diagnostic tools and treatment strategies continue to have limitations. Substantial evidence shows that long noncoding RNAs (lncRNAs) play essential roles in carcinogenesis and the progression, treatment response and prognosis of multiple human cancers, including urological cancers, gastrointestinal tumours, reproductive cancers and respiratory neoplasms. LncRNA small nucleolar RNA host genes (SNHGs), a subgroup of lncRNAs, have been found to be dysregulated in tumour cell biology. In this review, we summarize the impacts of lncRNA SNHGs in urological malignancies and the underlying mechanisms.
Introduction
Urological cancer is one of the most common malignancies in humans. Among the urological cancers, prostate cancer (PCa), bladder cancer (BCa) and kidney cancer are the three most common (1). Although treatment strategies have improved significantly, the recurrence rate remains high, and advanced tumour status tends to be associated with unfavourable outcomes (2–4). Therefore, much research in the last one to two decades has been dedicated to exploring potential biomarkers and identifying promising options for patients with urological malignancies.
Noncoding RNAs (ncRNAs) represent a large group of RNAs that lack the capacity to encode proteins (5). Instead, ncRNAs play important roles in epigenetic modification, transcription and translation, and the majority of ncRNAs act as regulatory factors in gene expression (6). LncRNA is a type of ncRNA longer than 200 nucleotides. lncRNAs are involved in the intracellular transportation of macromolecular substances, such as proteins, and ensure that they are delivered to appropriate functional regions and are activated or deactivated correctly (7). Recent literature rejected the former definition of lncRNA as transcription noise and revealed its multiple roles in the regulation of cell behaviour, especially during tumorigenesis and progression (8). Small nucleolar RNAs (snoRNAs) are a class of ncRNAs that predominantly exist in the nucleolus and are approximately 60-300 nucleotides in length (9). In humans, more than half of snoRNAs are transcribed from host genes, of which SNHGs account for nearly four-fifths (10). The aberrant expression of SNHGs, including lncRNA SNHGs, has been shown to be associated with the origin and development of several types of cancers (11).
Although correlations between lncRNA SNHGs and urological cancers have been reported, the roles that lncRNA SNHGs play in urological cancers are diverse and complex. We aim to comprehensively review this issue in this article.
lncRNA SNHGs in Prostate Cancer
In men, PCa has long had the highest incidence of all tumours, and its incidence continues to grow globally each year (12) (Figure 1). Relative to the progression of other malignant tumours, the progression of PCa is slow, and many patients can achieve long-term tumour-bearing survival. However, when the disease advances to the terminal stage, i.e., castration-resistant prostate cancer (CRPC) or metastatic castration-resistant prostate cancer (mCRPC), the prognosis is unsatisfactory, and most mortality occurs in this stage (13). Thus, it is imperative to clarify the mechanism of tumorigenesis and tumour progression at the gene level and to develop novel strategies for treatment.
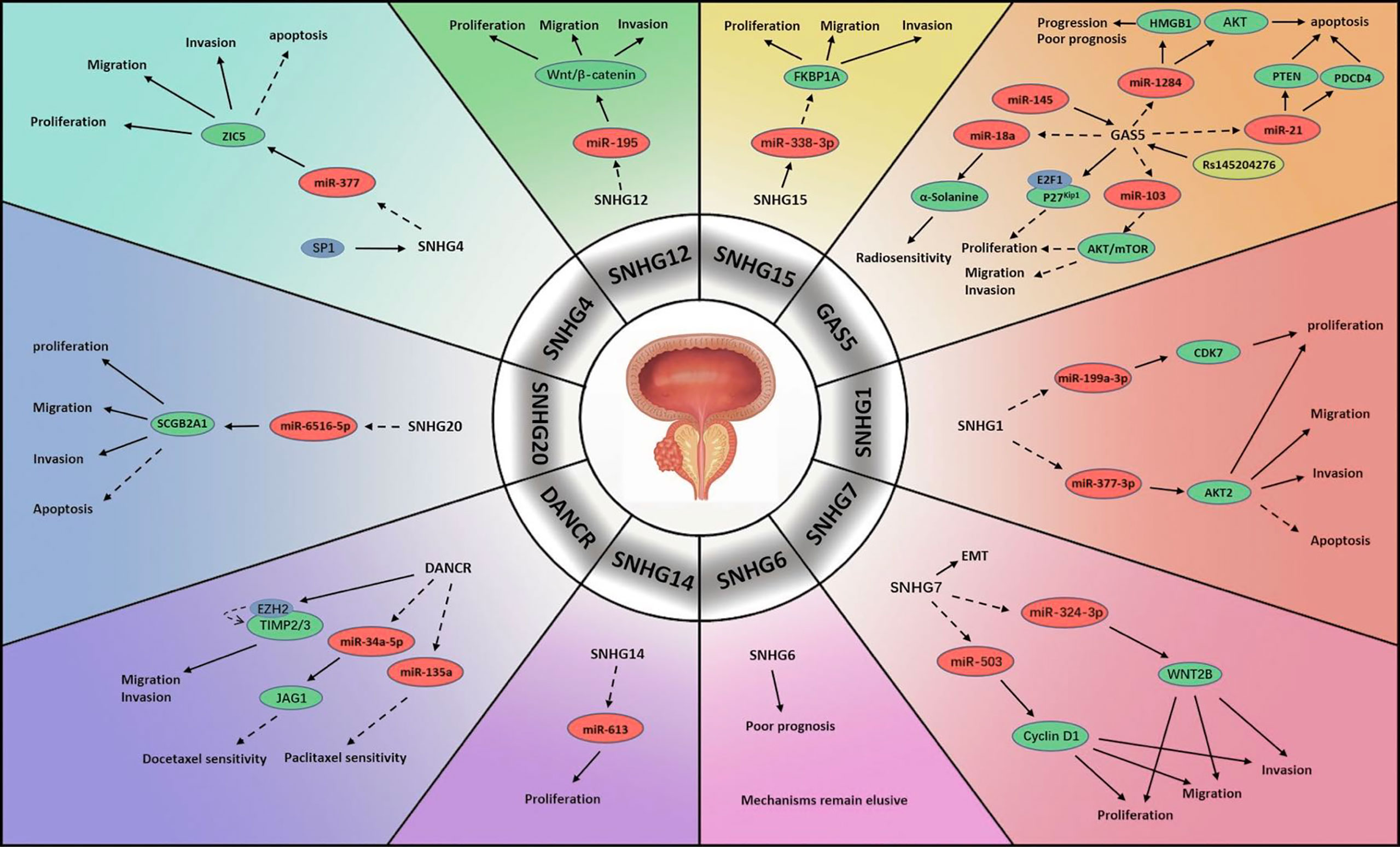
Figure 1 Mechanisms of lncRNA SNHGs in prostate cancer. → Stimulate, ⤏ Inhibit.
SNHG1 is regarded to participate significantly in the proliferation and metastasis of multiple neoplasms (14). PCa is partly attributed to the abnormal expression of SNHG1. In one study, scientists compared PCa specimens and specimens of adjacent non-tumour tissue and found that the expression level of SNHG1 was much higher in the former than in the latter. MiR-199a-3p, which was shown to be negatively associated with SNHG1 expression, was strongly repressed in PCa cells. It was suggested that SNHG1 might enhance cyclin-dependent kinase 7 (CDK7) encoding by mediating miR-199a-3p expression, which boosted cell replication (15). Another possible role of SNHG1 was also observed. MiR-377-3p, which acts as a suppressor of cell proliferation and a promoter of apoptosis, was blocked by the upregulation of SNHG1 in PCa. The downstream target was confirmed to be protein kinase B2 (AKT2), which promotes cell growth and exhibits anti-apoptosis activity. Their results suggested that the SNHG1/miR-377-3p/AKT2 pathway is linked to the progression and prognosis of PCa (16).
SNHG2, also named growth-arrest-specific-5 (GAS5), is a host gene of many snoRNAs and is widely considered a tumour suppressor in numerous human neoplasms (17). Multiple studies have reported that GAS5 is significantly correlated with the tumorigenesis, progression, clinical characteristics, treatment sensitivity and prognosis of PCa. High GAS5 expression might lead to mild clinical features of PCa. It was revealed that GAS5 was capable of repressing tumour cell viability and proliferation by blocking the protein kinase B (AKT)/mammalian target of rapamycin (mTOR) signalling pathway. In this process, miR-103 acted as the intermediate factor, which could be negatively regulated by GAS5 (18). Another pathway was believed to be associated with E2F transcriptional factor 1 (E2F1). Wherein, GAS5 attached E2F1 to the P27Kip1 promoter to initiate effects on the cell cycle, which decelerated PCa development (19). Pickard et al. studied the associations between GAS5 and PCa cell apoptosis and found that GAS5 served as a trigger of apoptosis in various cell lines (20). Yacqub-Usman et al. investigated the interactions between GAS5 and mTOR inhibitors in PCa and demonstrated that GAS5 reduced the tolerance of tumour cells to mTOR inhibitors (21). In a study concerning the modulation of GAS5 in PCa radiotherapy GAS5 was found able to silence miR-18a to promote α-Solanine, a novel anti-tumour molecule able to increase tumour cell sensitivity to radiation (22). A potential upstream regulator of GAS5 was elucidated to be miR-145. Researchers demonstrated that both GAS5 and miR-145 expression was greatly attenuated in PCa specimens. In contrast, enhancing miR-145 level significantly impeded tumour cell proliferation and induced apoptosis by elevating GAS5 expression. However, the intermediate regulator between miR-145 and GAS5 was not identified (23). Interestingly, some studies revealed that a genetic polymorphism in GAS5 known as rs145204276, a 5-bp indel polymorphism (-/AGGCA) in the GAS5 promoter, is involved in PCa (24–26). One demonstrated that the presence of rs145204276 markedly reduced the lymph node and seminal vesicle infiltration of PCa cells, especially among patients older than 65 years old. Furthermore, lymph vascular violation, a potential predictor of PCa biochemical recurrence, was identified to be reduced (25). The mechanism of rs145204276 activity was revealed by another, which showed that rs145204276 greatly upregulated GAS5 and stimulated its transcription. By downregulating miR-1284, activated GAS5 could inhibit high mobility group box 1 (HMGB1), which has been identified as playing a promotive role in carcinogenesis (24). Furthermore, researchers also discovered that miR-21 might be correlated with GAS5. Elevated GAS5 suppressed miR-21, targeting phosphatase and tensin homolog deleted on chromosome ten (PTEN) and programmed cell death 4 (PDCD4) and thereby inhibiting the occurrence of PCa (26). In contrast to the above observations, Zhang et al. reported GAS5 overexpression in PCa cell specimens. They demonstrated that GAS5-007, a transcript of GAS5, accelerated cell proliferation and prevented apoptosis in PCa (27).
Another focus of study is SNHG4, which has been reported to participate in several types of cell activity (28). SNHG4 has also been investigated in PCa. In a study concerning 113 PCa patients and tested their tumour tissues, a positive correlation between SNHG4 level and overall survival (OS) was observed. Further investigation showed that transcription factor SP1 activated SNHG4 upregulation, contributing to tumour cell proliferation and metastasis. Blocking SNHG4 may impair cell growth and migration and inspire apoptosis through elevating miR-377, which is regulated by SNHG1, as mentioned above. In this process, Zinc finger protein 5 (ZIC5) was identified as another downstream target of miR-377 (29).
A meta-analysis concerning SNHG6 summarized its predictive utility regarding the pathological characteristics and clinical outcomes of various neoplasms (30). In PCa, elevated expression of SNHG6 in tumour tissues was speculated to be associated with an unfavourable prognosis. The researchers constructed protein-protein interaction (PPI) networks involving SNHG6 and conducted bioinformatics analysis, which revealed the association of SNHG6 with multiple cell activities such as translation, mRNA modulation and catabolism, rRNA modification and ribosome construction. However, the signalling pathway mediating the SNHG6 functions in PCa was not clarified, a topic that warrants further investigation (31).
Accumulating studies indicate that SNHG7 might be a regulatory molecule in diverse human neoplasms, including urological tumours (32). Researchers have revealed various mechanisms by which SNHG7 regulates tumour cell activities in PCa. Augmented levels of SNHG7 suppressed miR-324-3p and then activated the Wingless-type MMTV integration site family member 2b (WNT2B) in PCa. The epithelial-mesenchymal transition (EMT) was also found to be involved in this pathway, accelerating metastatic process (33). Another approach by which SNHG7 regulates PCa involves the SNHG7/miR-503/Cyclin D1 pathway. SNHG7 depletion inhibited PCa cell replication and growth and froze the cell cycle at the G0/G1 phase. In particular, SNHG7 reduced the miR-324-3p level, which negatively regulates the Cyclin D1 protein (34). Furthermore, SNHG7 might be an independent risk factor that patients with higher SNHG7 levels were more likely to have an unfavourable outcome than those with lower SNHG7 levels (35).
SNHG12 is a dysregulated molecule in the pathogenesis, progression and prognosis of diverse neoplasms (36). In a study, SNHG12 was markedly increased in PCa specimens, energizing tumour cells. SNHG12 activated the miR-195/Wnt/β-catenin pathway, which promoted the pathogenesis of PCa (37).
SNHG13, a lncRNA also known as differentiation antagonizing non-protein coding RNA (DANCR), plays a regulatory role as an oncogene in multiple human cancers. DANCR expression is correlated with pathogenesis and progression and is predictive of the outcome of several neoplasms (38, 39). PCa studies have indicated that DANCR may affect the treatment response to chemotherapy. One literature reported an advanced DANCR level in docetaxel (DTX) nonresponsive cell lines and that DANCR depletion improved the effects of DTX. MiR-34a-5p, extensively sponged by DANCR, was capable of impairing Jagged 1 (JAG1) encoding, which is associated with chemotherapy tolerance (40). The role of DANCR in paclitaxel treatment was also revealed that the upregulation of DANCR inhibited the expression of miR-135a, which could sensitize tumour cells in paclitaxel chemotherapy (41). In another study, the impairment of DANCR in PCa cell lines abrogated cell viability and mobility. In that work, DANCR functioned similarly to SNHG2, downregulating TIMP metallopepitidase inhibitor 2/3 (TIMP2/3) by activating the binding of enhancer of zeste homolog 2 (EZH2) to the promoter of TIMP2/3, which accelerated PCa metastasis. Furthermore, activation of the androgen-androgen receptor (AR) pathway attenuated the DANCR level, decelerating PCa progression (42).
A novel oncogene, SNHG14 exerts regulatory effects in lung cancer, digestive cancer, reproductive cancer, haematological cancer and osteosarcoma (43). SNHG14 is markedly overexpressed in PCa tumour cells. Impeding SNHG14 could greatly suppress cell replication capacity. MiR-613 was found to be blocked by SNHG14 in PCa. Furthermore, silencing SNHG14 led to a reduced PCa tissue volume compared with that in a control group. This research established the mechanism by which SNHG14 promotes PCa through miR-613 (44).
Studies have revealed SNHG15 as a regulator in the progression of several human malignancies (45). Scientists examined its status and modes of function in PCa, finding an aberrant increase in SNHG15 in tumour cells relative to normal tissues. SNHG15 impairment was found to compromise tumour cell activity and reduce tumour aggressiveness. They further revealed miR-338-3p as a target of SNHG15, the downstream molecule of which was found to be FKBP prolyl isomerase 1A (FKBP1A). They revealed the mechanism involving the SNHG15/miR-338-3p/FKBP1A pathway and showed that SNHG15 promoted FKBP1A by negatively regulating miR-338-3p in PCa cells (46).
SNHG20 plays crucial roles in tumorigenesis and progression in several human cancers, including gastrointestinal cancer, liver cancer, breast cancer, cervical cancer, oral cancer and urological cancer (47). Both PCa tissues and cell lines demonstrated greatly elevated SNHG20 levels in one study. Enhancing SNHG20 level activated anti-apoptosis function and accelerated cell propagation. The mechanism was reported to involve the SNHG20/miR-6516-5p/secretoglobin family 2A member 1 (SCGB2A1) signalling pathway: SNHG20 suppressed miR-6516-5p level, which negatively modulated SCGB2A1, a positive regulator of tumour cell activity that is elevated in PCa (48).
lncRNA SNHGs in Bladder Cancer
BCa is another common concern around the globe; although it does not rank as one of the top cancers in incidence or mortality rate, over 400 000 patients are diagnosed every year (49) (Figure 2). Urinary ultrasound combined with cystoscopy is very effective for detection. Transurethral resection and intravesical chemotherapy are the most recommended treatments for non-muscle invasive bladder cancer (NMIBC). However, a considerable number of patients face high risks of recurrence and progression, especially when routine surveillance is absent (50). Once NMIBC advances to muscle invasive bladder cancer (MIBC), the treatments become limited to partial or radical cystectomy and systemic chemotherapy (51). Generally, to prevent recurrence, patients diagnosed with BCa are likely to receive a long period of medical follow up involving procedures such as cystoscopy, which is mentally and physically unpleasant for patients. Therefore, it is crucial to clarify the genetic mechanisms of BCa carcinogenesis, progression and recurrence to identify biomarkers for disease management. The discovery of lncRNA SNHGs may provide clinical options.
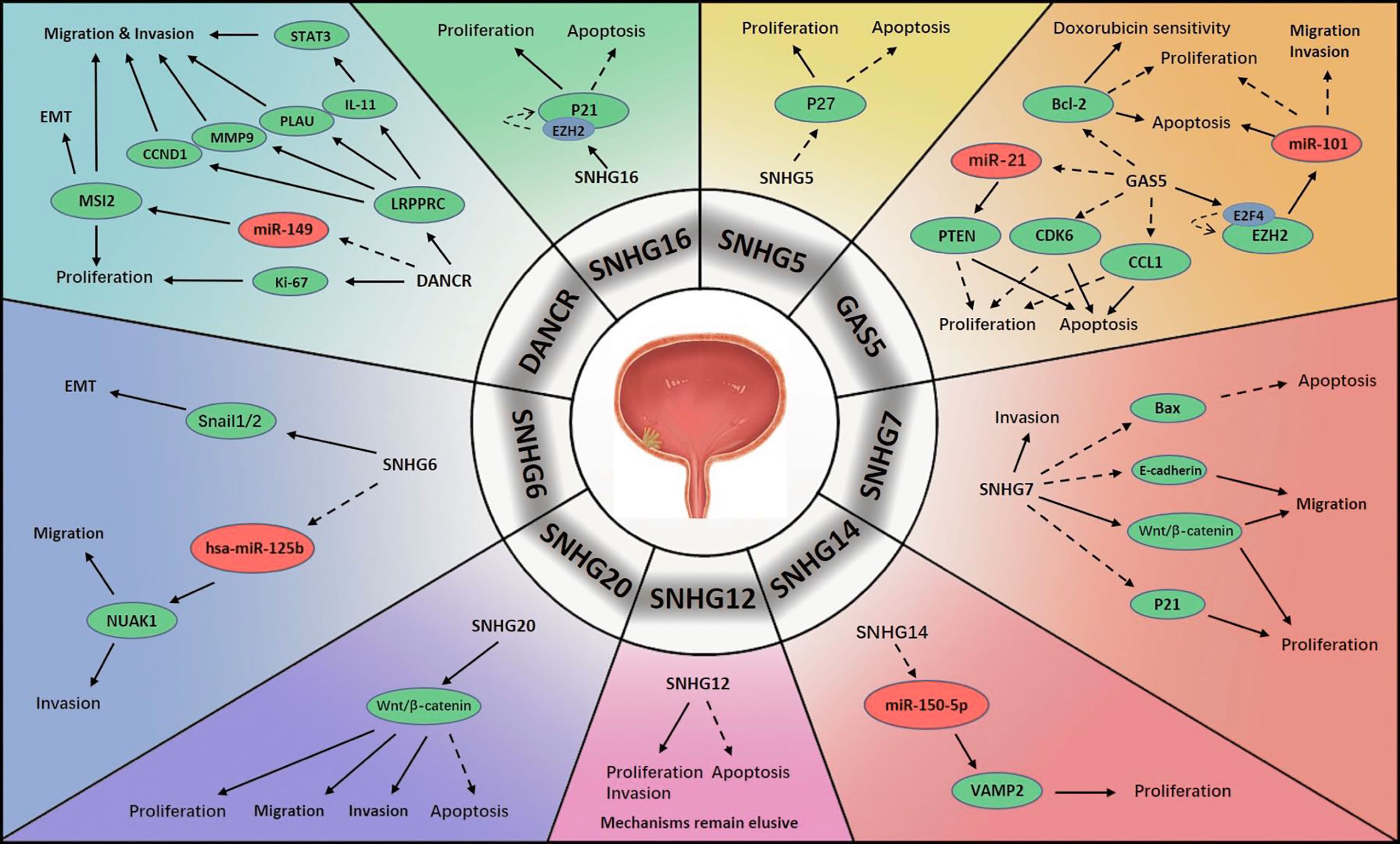
Figure 2 Mechanisms of lncRNA SNHGs in bladder cancer →. Stimulate ⤏, Inhibit.
An extensive decrease in GAS5 level in BCa cells has been found in multiple studies. Similar to PCa, BCa has been shown to be associated with activation of the GAS5/miR-21/PTEN signalling pathway (52). GAS5 was confirmed to inhibit tumour cell proliferation and promote cell apoptosis, and these processes were modified by cyclin-dependent kinase 6 (CDK6), which had a negative correlation with GAS5 in BCa progression (53). Similar correlations between GAS5 and BCa were also discovered that GAS5 impaired the encoding of B-cell lymphoma-2 (Bcl-2), which is an anti-apoptosis factor in carcinogenesis. Apart from this, GAS5 enhanced the response of BCa cell lines to doxorubicin chemotherapy (54). Another literature demonstrated that GAS5 initiated the binding of E2F transcriptional factor 4 (E2F4) with the EZH2 promoter, thereby exerting its proapoptotic function. The prognostic value of GAS5 was also evaluated, and BCa with elevated GAS5 expression tended to display lower malignancy than that with lower GAS5 expression (55). Another possible pathway was explored that GAS5 repressed BCa cell proliferation and activated apoptosis by sponging chemokine ligand 1 (CCL1), a regulatory factor in BCa, which might increase cell viability (56). Additionally, GAS5 was found to be of great value in the diagnosis and prognosis of BCa (57). Avgeris et al. identified GAS5 as an independent predictor of BCa prognosis, especially for NMIBC. In BCa, the absence of GAS5 was associated with more rapid recurrence. Furthermore, GAS5 was suggested to potentially be predictive of disease aggressiveness, and lower levels were found to be associated with higher malignancy (58). Intriguingly, scientists measured lncRNA levels in urine samples and constructed a lncRNA panel consisting of two lncRNAs: GAS5 and uc004cox.4. Analysis revealed that GAS5 had superior diagnostic performance to conventional urine cytology (57).
SNHG5 plays a pivotal role in diverse human neoplasms (59). Unsurprisingly, it also plays an important role in urological malignancies. SNHG5 has been found to be greatly overexpressed in BCa specimens relative to normal urothelial specimens. In addition, increased SNHG5 has been shown to be associated with a more malignant tendency in several aspects, such as tumour burden, aggressiveness and pathological features. Experiments have shown that knocking down SNHG5 has antiproliferative and proapoptotic effects. Additionally, SNHG5 has been shown to exert its tumour-promoting function through the sponging mRNA p27, which is reduced in BCa cells (60).
The regulatory mechanisms of SNHG6 in BCa were revealed. Scientists found that SNHG6 was aberrantly upregulated in BCa tissues, which stimulated EMT by negatively regulating E-cadherin encoding and positively regulating vimentin encoding. Snail1/2 was suggested to modulate this activity. As expected, SNHG6 greatly enhanced the aggressiveness of BCa cells, and silencing SNHG6 reduced BCa cell viability. MiR-125b was subsequently identified as an intermediate molecule of SNHG6, and a negative feedback circle between SNHG6 and hsa-miR-125b in BCa was revealed as SNHG6/hsa-miR-125b/NUAK Family Kinase 1(NUAK1) signalling in BCa promotion (61).
The elevated expression of SNHG7 in BCa was also observed, but the mechanisms varied. Chen et al. repressed SNHG7, which resulted in reduced survival, growth and migration of tumour cells and cell cycle arrest; these findings confirmed that SNHG7 accelerates BCa progression. Moreover, it was verified that Wnt/β-catenin pathway was stimulated by SNHG7 to activate carcinogenesis (62). Others found that SNHG7 depletion greatly reduced cell viability, migration and invasion and triggered cell apoptosis. Possible targets of SNHG7 were revealed to be Bax, p21 and E-cadherin, which were previously demonstrated to initiate apoptosis, modulate the cell cycle and impede cell proliferation, and block cell migration, respectively (63).
One study demonstrated that SNHG12 was abnormally regulated in BCa. Although the mechanism was not identified, they showed that silencing SNHG12 may hamper cell proliferation and invasion and exert proapoptotic effects in BCa. Additionally, they reported that the overexpression of SNHG10 and SNHG12 might lead to worse relapse-free survival (RFS) in BCa (64).
DANCR expression is aberrantly upregulated in BCa pathogenesis and progression. DANCR was considered capable of stimulating lymph node infiltration and metastasis by elevating the levels of interleukin 11 (IL-11), plasminogen activator urokinase (PLAU), matrix metalloproteinase 9 (MMP9), and CCND1(or cyclin D1). In addition, leucine-rich pentatricopeptide repeat containing (LRPPRC), which is differentially expressed between BCa cells and normal cells, was shown to maintain the mRNA structures of these four molecules (65). When DANCR was silenced in BCa cells, cell viability and mobility were markedly compromised. The mechanism involved the upregulation of musashi RNA binding protein 2 (MSI2) by DANCR via the repression of miR-149. High levels of MSI2 were found to be positively associated with the malignant degree of BCa (66).
Researchers discovered that SNHG14 expression was elevated in BCa cells compared with non-tumour cells, which suggested a poorer prognosis. The signalling pathway involved in BCa was the SNHG14/miR-150-5p/vesicle-associated membrane protein 2 (VAMP2) axis. MiR-150-5p acted as a downstream target of SNHG14, and its upregulation antagonized the promoting effects of SNHG14. Moreover, miR-150-5p was found to reduce VAMP2 level, thereby enhancing cell activity and facilitated BCa progression (67).
SNHG16 is a novel factor active in various human cancers and predictive of poor prognosis (68). Scientists detected an abnormally overexpressed SNHG16 level in serum exosomes of BCa patients. SNHG16 was expressed to a greater extent in MIBC than in NMIBC, suggesting that it stimulates tumour progression (69). Similarly, others found that SNHG16 level was greatly elevated in the nuclei of BCa cells. The upregulation of SNHG16 was confirmed to be associated with tumour severity and detrimental to clinical outcomes. When silencing SNHG16, cell growth was delayed and apoptosis was accelerated. The possible target of SNHG16 was p21, which was previously demonstrated to interfere with the cell cycle, maintaining it at G1 stage (70).
SNHG20 is another important oncogene in BCa. Researchers observed a pronounced increase of SNHG20 in BCa specimens and that patients with higher SNHG20 levels tend to have less favourable outcomes. They found that SNHG20 deficiency reduced the viability and aggressiveness of tumour cells and stimulated apoptosis. Furthermore, the impairment of SNHG20 impeded activation of the Wnt/β-catenin pathway and thereby inhibited tumour progression (71).
lncRNA SNHGs in Kidney Cancer
The incidence of kidney cancer is slightly lower than that of bladder cancer, accounting for approximately 2% of human cancers (72) (Figure 3). Renal cell carcinoma (RCC) accounts for most kidney cancers, the predominant and most malignant subtype of which is clear cell renal cell carcinoma (ccRCC) (73). As RCC does not respond well to chemotherapy and radiotherapy, surgical resection has been the most common treatment strategy (74). However, the prognosis remains unsatisfactory and lower than that of BCa and PCa. Thus, research to identify promising biomarkers to improve the diagnosis and treatment of RCC is urgently needed. Several lncRNA SNHGs have been shown to be associated with RCC.
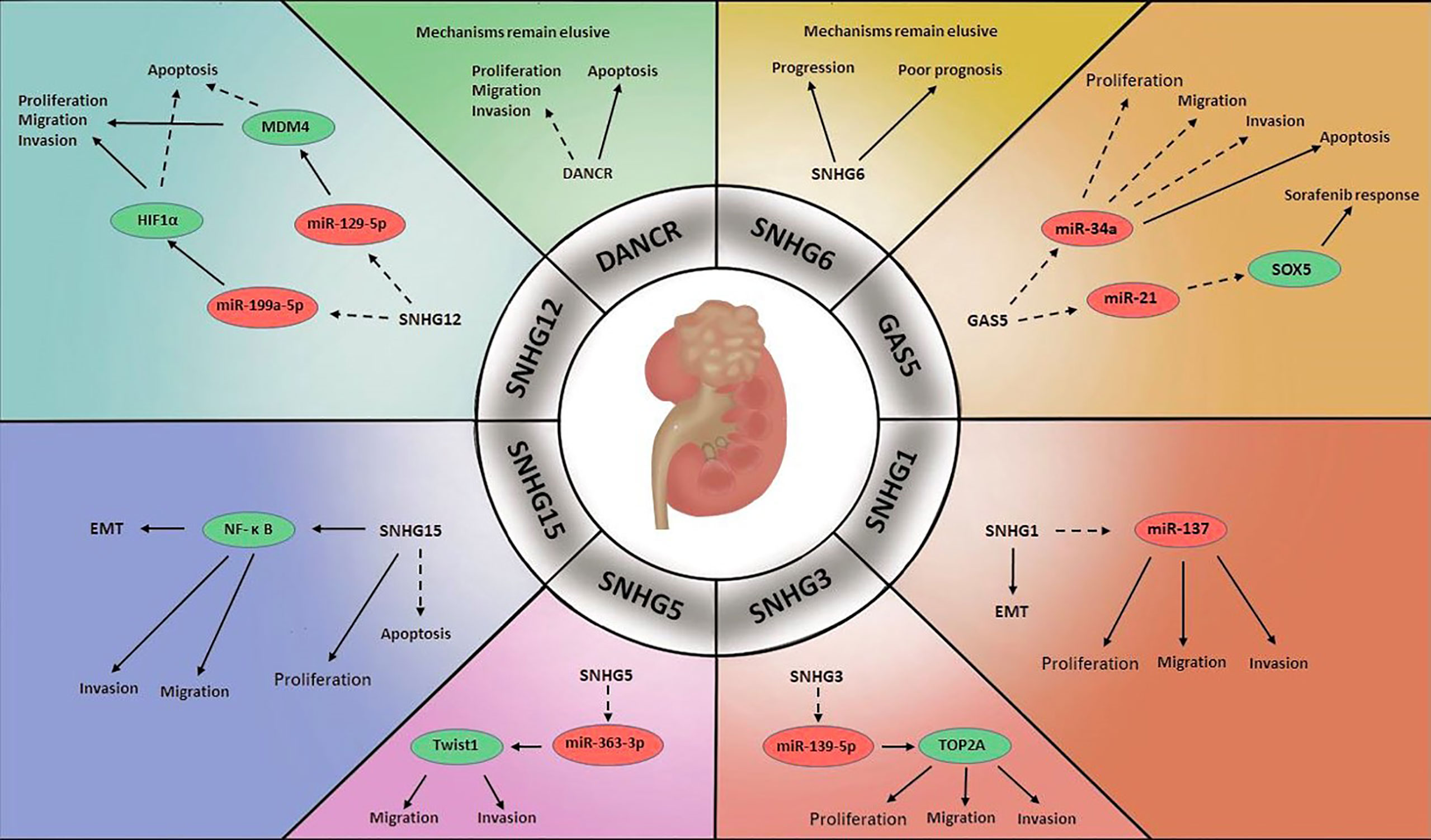
Figure 3 Mechanisms of lncRNA SNHGs in renal cell carcinoma →. Stimulate ⤏, Inhibit.
In one study, SNHG1 was found to be greatly upregulated in RCC cells relative to surrounding non-tumour cells, and SNHG1 inhibition was shown to block the EMT, thereby reducing tumour cell growth and invasion. It was speculated that miR-137 was a target of SNHG1, as SNHG1 knockdown promoted miR-137 expression in RCC cells (75).
Research on RCC has identified GAS5 as a tumour suppressor. Toraih et al. measured GAS5 and miR-34a levels in RCC specimens, and the results suggested that miR-34a was upregulated in the former and downregulated in the latter. Although the mechanism was not clarified, the authors speculated that there might be reciprocal regulation between GAS5 and miR-34a in oncogenesis (76). Others observed a large decrease in GAS5 level in RCC cell lines relative to normal cell lines. They found that enhanced GAS5 expression inhibited cell proliferation, migration and invasive ability and triggered cell apoptosis (77). Liu et al. demonstrated that GAS5 could improve the treatment response to sorafenib in RCC. GAS5 level was markedly increased in sorafenib-sensitive patients compared with sorafenib-resistant patients. They reported that GAS5 and miR-21 interacted with each other and that the target of miR-21 was sex determining region Y-box protein 5 (SOX5), which was negatively modulated by miR-21 (78). GAS5 may be of outstanding prognostic value for RCC clinical features, especially in the prediction of advanced stage and capsular invasion (79).
The dysregulation of SNHG3 has been another focus (80). SNHG3 is overexpressed in tumour specimens of ccRCC, which was significantly correlated with unfavourable clinical and pathological features. SNHG3 depletion has been shown to inhibit cell viability, proliferation and aggressiveness by releasing the negative regulatory effects of miR-139-5p on Topoisomerase II Alpha (TOP2A), identified as the downstream factor of miR-139-5p, and stimulating tumour progression (81).
The role that SNHG5 plays in ccRCC was considered as an oncogene. Scientists revealed that SNHG5 was greatly overexpressed in ccRCC specimens compared with adjacent specimens. Silencing SNHG5 expression activated apoptosis and compromised the invasive capability of ccRCC cells by upregulating miR-363-3p. Further study revealed that the target molecule of miR-363-3p was Twist1, which greatly enhanced cell aggressiveness (82).
SNHG6 has been confirmed to be associated with the clinical features and prognosis of RCC. Elevated SNHG6 expression in RCC specimens were markedly associated with RCC tumour burden and lymph node infiltration. Moreover, SNHG6 was identified as a risk factor in RCC prognosis that patients with higher SNHG6 levels were more likely to have lower OS (83).
An abnormally elevated SNHG12 level in ccRCC was observed. Upregulation of SNHG12 predicted an advanced stage of tumour while knockdown of SNHG12 hampered tumour progression and stimulated apoptosis. SNHG12 was found to exert its regulatory effects through downregulating miR-129-5p, the target of which is murine double minute 4 (MDM4), which has been reported to be overexpressed in several cancers, including ccRCC (84). Another signalling pathway involved this molecular was that via sponging miR-199a-5p, SNHG12 promoted transcription factor HIF1α to accelerate RCC carcinogenesis (85).
In contrast to observations in PCa and BCa, DANCR level was found to be reduced in RCC specimens. Enhanced DANCR expression sharply impeded cell growth and aggressiveness and promoted apoptosis in RCC cells. Although the mechanism was not revealed, these findings suggested that DANCR might act as a tumour inhibitor in RCC (86).
In RCC, increased SNHG15 expression was found to facilitate RCC progression, while SNHG15 depletion resulted in proapoptotic cells and cell cycle arrest. The detailed mechanism was that SNHG15 stimulated the EMT through regulating NF-κB, ultimately enhancing RCC proliferation, migration and invasion (87).
Yang et al. measured lncRNA SNHGs levels in ccRCC and found that SNHG1, GAS5, SNHG3-8, SNHG11, SNHG12, SNHG15-17, SNHG20, SNHG22 and SNHG25 were overexpressed in RCC cells compared with healthy renal cells. In contrast, SNHG9, SNHG10, DANCR and SNHG14 levels were decreased in RCC cells. Among these SNHGs, SNHG3 and SNHG15 exhibited strong prognostic performance. Both were negatively correlated with OS in RCC, and SNHG15 alone was associated with RFS. Additionally, compared with localized RCC, advanced RCC tended to be associated with higher levels SNHG3 and SNHG15 (88).
Wilms’ tumour (WT), or nephroblastoma, is the most common renal malignancy in children, accounting for nearly 7% of all cancers (89, 90). A literature reported the discovery of abnormally increased SNHG6 levels in WT tissues. Impairment of SNHG6 reduced tumour cell viability and mobility and stimulated apoptosis. MiR-15a was confirmed to negatively interact with SNHG6, with the downregulation of SNHG6 leading to enhanced miR-15a expression, which repressed the TGF-β-activated kinase 1 (TAK1)/c-Jun-N-terminal kinase (JNK) and Wnt/β-catenin signalling pathways. Both of these pathways were considered to stimulate tumour progression (91).
Conclusion
Generally, lncRNA SNHGs engage in the modulation of cancer biology in two ways. In one way, lncRNA SNHGs act as competing endogenous RNAs (ceRNAs) that occupy the binding sites between microRNA (miRNA) and its target genes, preventing the inhibitive effects of miRNA on transcription level (92, 93). In the other way, lncRNA SNHGs regulate certain signalling pathways involved in protein encoding and influence tumour pathogenesis and progression on protein expression level. LncRNA SNHGs that significantly involved in urological cancers include SNHG1, GAS5, SNHG3-7, SNHG12, DANCR, SNHG14-16 and SNHG20 (Table 1).
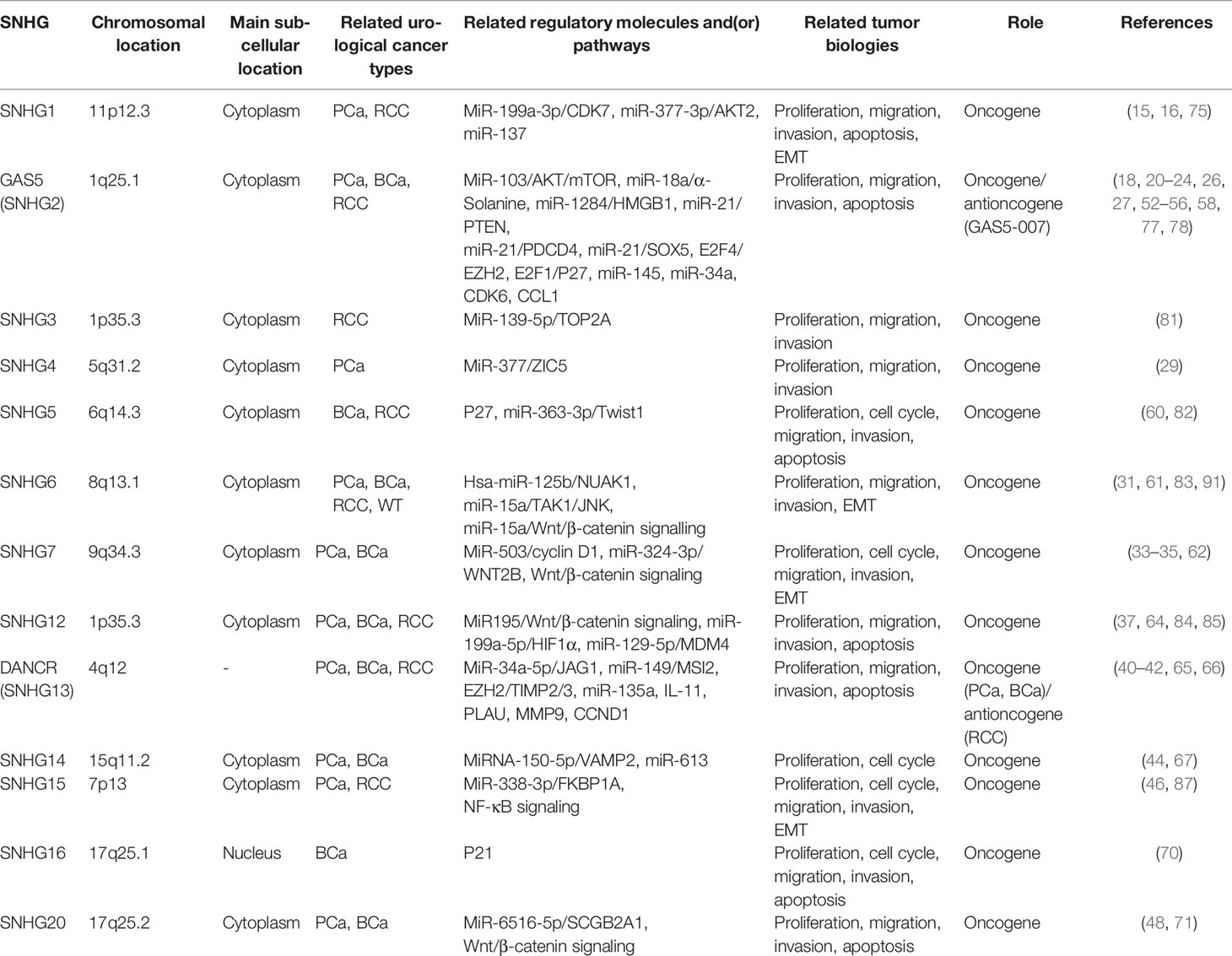
Table 1 LncRNA SNHGs in urological cancers.
The differential expression of lncRNA SNHGs in tumour and non-tumour cells, as well as the discovery of underlying mechanisms by which they function have proved its non-negligible role in tumorigenesis, progression and prognosis of multiple human cancers. In urological cancers, most of lncRNA SNHGs act as an oncogene. While interestingly, DANCER (SNHG13) serves as an oncogene in PCa and BCa but an antioncogene in RCC. Similar phenomenon also exists in GAS5 (SNHG2). The reason why these two molecules play opposite roles in different urological cancers needs further clarification. For sure, the comprehensive and in-depth understanding of the biological features of lncRNA SNHGs may provide us more clues in the exploration of novel biomarkers and potential therapeutic targets in the managements of urological cancers.
Author Contributions
ZY and XZ conceived the structure of manuscript. XZ, LX, ZY and QL critically revised the manuscript. ZY and QL drafted initial manuscript and made the figures and table. All authors contributed to the article and approved the submitted version.
Funding
This study was supported by grants from the National Natural Science Foundation of China (81972374 and 81772744).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
All the authors had full access to all the data and the corresponding author had the final responsibility for the decision to submit for publication. Thanks for the help and support of the Zhejiang University Cancer Institute.
References
1. Siegel RL, Miller KD, Jemal A. Cancer Statistics, 2016. CA Cancer J Clin (2016) 66(1):7–30. doi: 10.3322/caac.21332
2. Tran L, Xiao JF, Agarwal N, Duex JE, Theodorescu D. Advances in Bladder Cancer Biology and Therapy. Nat Rev Cancer (2021) 21(2):104–21. doi: 10.1038/s41568-020-00313-1
3. Lindenberg ML, Turkbey B, Mena E, Choyke PL. Imaging Locally Advanced, Recurrent, and Metastatic Prostate Cancer: A Review. JAMA Oncol (2017) 3(10):1415–22. doi: 10.1001/jamaoncol.2016.5840
4. Porta C, Cosmai L, Leibovich BC, Powles T, Gallieni M, Bex A. The Adjuvant Treatment of Kidney Cancer: A Multidisciplinary Outlook. Nat Rev Nephrol (2019) 15(7):423–33. doi: 10.1038/s41581-019-0131-x
5. Li J, Shen H, Xie H, Ying Y, Jin K, Yan H, et al. Dysregulation of Ncrnas Located At the DLK1DIO3 Imprinted Domain: Involvement in Urological Cancers. Cancer Manag Res (2019) 11:777–87. doi: 10.2147/CMAR.S190764
6. Zeuschner P, Linxweiler J, Junker K. Non-Coding Rnas as Biomarkers in Liquid Biopsies With a Special Emphasis on Extracellular Vesicles in Urological Malignancies. Expert Rev Mol Diagn (2020) 20(2):151–67. doi: 10.1080/14737159.2019.1665998
7. Stepanov GA, Filippova JA, Komissarov AB, Kuligina EV, Richter VA, Semenov DV. Regulatory Role of Small Nucleolar Rnas in Human Diseases. BioMed Res Int (2015) 2015:206849. doi: 10.1155/2015/206849
8. Batista PJ, Chang HY. Long Noncoding Rnas: Cellular Address Codes in Development and Disease. Cell (2013) 152(6):1298–307. doi: 10.1016/j.cell.2013.02.012
9. Krishnan P, Ghosh S, Wang B, Heyns M, Graham K, Mackey JR, et al. Profiling of Small Nucleolar Rnas by Next Generation Sequencing: Potential New Players for Breast Cancer Prognosis. PloS One (2016) 11(9):e0162622. doi: 10.1371/journal.pone.0162622
10. Gong J, Li Y, Liu CJ, Xiang Y, Li C, Ye Y, et al. A Pan-Cancer Analysis of the Expression and Clinical Relevance of Small Nucleolar Rnas in Human Cancer. Cell Rep (2017) 21(7):1968–81. doi: 10.1016/j.celrep.2017.10.070
11. Williams GT, Farzaneh F. Are Snornas and Snorna Host Genes New Players in Cancer? Nat Rev Cancer (2012) 12(2):84–8. doi: 10.1038/nrc3195
12. Zhai Z, Zheng Y, Li N, Deng Y, Zhou L, Tian T, et al. Incidence and Disease Burden of Prostate Cancer From 1990 to 2017: Results From the Global Burden of Disease Study 2017. Cancer (2020) 126(9):1969–78. doi: 10.1002/cncr.32733
13. Halabi S, Lin CY, Kelly WK, Fizazi KS, Moul JW, Kaplan EB, et al. Updated Prognostic Model for Predicting Overall Survival in First-Line Chemotherapy for Patients With Metastatic Castration-Resistant Prostate Cancer. J Clin Oncol (2014) 32(7):671–7. doi: 10.1200/JCO.2013.52.3696
14. Huang L, Jiang X, Wang Z, Zhong X, Tai S, Cui Y. Small Nucleolar RNA Host Gene 1: A New Biomarker and Therapeutic Target for Cancers. Pathol Res Pract (2018) 214(9):1247–52. doi: 10.1016/j.prp.2018.07.033
15. Li J, Zhang Z, Xiong L, Guo C, Jiang T, Zeng L, et al. SNHG1 Lncrna Negatively Regulates Mir-199a-3p to Enhance CDK7 Expression and Promote Cell Proliferation in Prostate Cancer. Biochem Biophys Res Commun (2017) 487(1):146–52. doi: 10.1016/j.bbrc.2017.03.169
16. Xie M, Zhang Z, Cui Y. Long Noncoding RNA SNHG1 Contributes to the Promotion of Prostate Cancer Cells Through Regulating Mir-377-3p/AKT2 Axis. Cancer Biother Radiopharm (2020) 35(2):109–19. doi: 10.1089/cbr.2019.3177
17. Goustin AS, Thepsuwan P, Kosir MA, Lipovich L. The Growth-Arrest-Specific (GAS)-5 Long Non-Coding RNA: A Fascinating Lncrna Widely Expressed in Cancers. Noncoding RNA (2019) 5(3):46. doi: 10.3390/ncrna5030046
18. Xue D, Zhou C, Lu H, Xu R, Xu X, He X. Lncrna GAS5 Inhibits Proliferation and Progression of Prostate Cancer by Targeting Mir-103 Through AKT/Mtor Signaling Pathway. Tumour Biol (2016). doi: 10.1007/s13277-016-5429-8
19. Luo G, Liu D, Huang C, Wang M, Xiao X, Zeng F, et al. Lncrna GAS5 Inhibits Cellular Proliferation by Targeting P27(Kip1). Mol Cancer Res (2017) 15(7):789–99. doi: 10.1158/1541-7786.MCR-16-0331
20. Pickard MR, Mourtada-Maarabouni M, Williams GT. Long Non-Coding RNA GAS5 Regulates Apoptosis in Prostate Cancer Cell Lines. Biochim Biophys Acta (2013) 1832(10):1613–23. doi: 10.1016/j.bbadis.2013.05.005
21. Yacqub-Usman K, Pickard MR, Williams GT. Reciprocal Regulation of GAS5 Lncrna Levels and Mtor Inhibitor Action in Prostate Cancer Cells. Prostate (2015) 75(7):693–705. doi: 10.1002/pros.22952
22. Yang J, Hao T, Sun J, Wei P, Zhang H. Long Noncoding RNA GAS5 Modulates Alpha-Solanine-Induced Radiosensitivity by Negatively Regulating Mir-18a in Human Prostate Cancer Cells. BioMed Pharmacother (2019) 112:108656. doi: 10.1016/j.biopha.2019.108656
23. Xie X, Dai J, Huang X, Fang C, He W. Microrna-145 Inhibits Proliferation and Induces Apoptosis in Human Prostate Carcinoma by Upregulating Long Non-Coding RNA GAS5. Oncol Lett (2019) 18(2):1043–8. doi: 10.3892/ol.2019.10419
24. Deng ZH, Yu GS, Pan B, Feng ZH, Huang Q, Deng JZ, et al. Rs145204276 and Rs4759314 Affect the Prognosis of Prostate Cancer by Modulating the GAS5/Mir-1284/HMGB1 and HOTAIR/Mir-22/HMGB1 Signalling Pathways. Artif Cells Nanomed Biotechnol (2020) 48(1):435–42. doi: 10.1080/21691401.2019.1709859
25. Lin CY, Wang SS, Yang CK, Li JR, Chen CS, Hung SC, et al. Impact of GAS5 Genetic Polymorphism on Prostate Cancer Susceptibility and Clinicopathologic Characteristics. Int J Med Sci (2019) 16(11):1424–9. doi: 10.7150/ijms.38080
26. Zhu L, Zhu Q, Wen H, Huang X, Zheng G. Mutations in GAS5 Affect the Transformation From Benign Prostate Proliferation to Aggressive Prostate Cancer by Affecting the Transcription Efficiency of GAS5. J Cell Physiol (2019) 234(6):8928–40. doi: 10.1002/jcp.27561
27. Zhang Y, Su X, Kong Z, Fu F, Zhang P, Wang D, et al. An Androgen Reduced Transcript of Lncrna GAS5 Promoted Prostate Cancer Proliferation. PloS One (2017) 12(8):e0182305. doi: 10.1371/journal.pone.0182305
28. Li H, Hong J, Wijayakulathilaka W. Long Non-Coding RNA SNHG4 Promotes Cervical Cancer Progression Through Regulating C-Met Via Targeting Mir-148a-3p. Cell Cycle (2019) 18(23):3313–24. doi: 10.1080/15384101.2019.1674071
29. Wang ZY, Duan Y, Wang P. SP1-Mediated Upregulation of Lncrna SNHG4 Functions as a Cerna for Mir-377 to Facilitate Prostate Cancer Progression Through Regulation of ZIC5. J Cell Physiol (2020) 235(4):3916–27. doi: 10.1002/jcp.29285
30. Zhao S, Zhu H, Jiao R, Wu X, Ji G, Zhang X. Prognostic and Clinicopathological Significance of SNHG6 in Human Cancers: A Meta-Analysis. BMC Cancer (2020) 20(1):77. doi: 10.1186/s12885-020-6530-3
31. Yan Y, Chen Z, Xiao Y, Wang X, Qian K. Long Non-Coding RNA SNHG6 is Upregulated in Prostate Cancer and Predicts Poor Prognosis. Mol Biol Rep (2019) 46(3):2771–8. doi: 10.1007/s11033-019-04723-9
32. Zhou Y, Tian B, Tang J, Wu J, Wang H, Wu Z, et al. SNHG7: A Novel Vital Oncogenic Lncrna in Human Cancers. BioMed Pharmacother (2020) 124:109921. doi: 10.1016/j.biopha.2020.109921
33. Han Y, Hu H, Zhou J. Knockdown of Lncrna SNHG7 Inhibited Epithelial-Mesenchymal Transition in Prostate Cancer Though Mir-324-3p/WNT2B Axis in Vitro. Pathol Res Pract (2019) 215(10):152537. doi: 10.1016/j.prp.2019.152537
34. Qi H, Wen B, Wu Q, Cheng W, Lou J, Wei J, et al. Long Noncoding RNA SNHG7 Accelerates Prostate Cancer Proliferation and Cycle Progression Through Cyclin D1 by Sponging Mir-503. BioMed Pharmacother (2018) 102:326–32. doi: 10.1016/j.biopha.2018.03.011
35. Xia Q, Li J, Yang Z, Zhang D, Tian J, Gu B. Long Non-Coding RNA Small Nucleolar RNA Host Gene 7 Expression Level in Prostate Cancer Tissues Predicts the Prognosis of Patients With Prostate Cancer. Med (Baltimore) (2020) 99(7):e18993. doi: 10.1097/MD.0000000000018993
36. Tamang S, Acharya V, Roy D, Sharma R, Aryaa A, Sharma U, et al. SNHG12: An Lncrna as a Potential Therapeutic Target and Biomarker for Human Cancer. Front Oncol (2019) 9:901. doi: 10.3389/fonc.2019.00901
37. Song J, Wu X, Ma R, Miao L, Xiong L, Zhao W. Long Noncoding RNA SNHG12 Promotes Cell Proliferation and Activates Wnt/Beta-Catenin Signaling in Prostate Cancer Through Sponging Microrna-195. J Cell Biochem (2019) 120(8):13066–75. doi: 10.1002/jcb.28578
38. Jin SJ, Jin MZ, Xia BR, Jin WL. Long Non-Coding RNA DANCR as an Emerging Therapeutic Target in Human Cancers. Front Oncol (2019) 9:1225. doi: 10.3389/fonc.2019.01225
39. Liu W, Wang QP, Guo J. Prognostic Significance of Long Non-Coding RNA DANCR Expression in Human Cancers: A Systematic Review and Meta-Analysis. Biosci Rep (2019). doi: 10.1042/BSR20181627
40. Ma Y, Fan B, Ren Z, Liu B, Wang Y. Long Noncoding RNA DANCR Contributes to Docetaxel Resistance in Prostate Cancer Through Targeting the Mir-34a-5p/JAG1 Pathway. Onco Targets Ther (2019) 12:5485–97. doi: 10.2147/OTT.S197009
41. Zhao HF, Zhang ZC, Shi BK, Jiang XZ. DANCR Sponges Mir-135a to Regulate Paclitaxel Sensitivity in Prostate Cancer. Eur Rev Med Pharmacol Sci (2019) 23(16):6849–57. doi: 10.26355/eurrev_201908_18724
42. Jia J, Li F, Tang XS, Xu S, Gao Y, Shi Q, et al. Long Noncoding RNA DANCR Promotes Invasion of Prostate Cancer Through Epigenetically Silencing Expression of TIMP2/3. Oncotarget (2016) 7(25):37868–81. doi: 10.18632/oncotarget.9350
43. Hou XK, Mao JS. Long Noncoding RNA SNHG14 Promotes Osteosarcoma Progression Via Mir-433-3p/FBXO22 Axis. Biochem Biophys Res Commun (2020) 523(3):766–72. doi: 10.1016/j.bbrc.2020.01.016
44. Sun B, Ke KB, Liu DF, Wang Q, Li YN, Chen JH, et al. Long Noncoding RNA SNHG14 Acts as an Oncogene in Prostate Cancer Via Targeting Mir-613. Eur Rev Med Pharmacol Sci (2020) 24(21):10919. doi: 10.26355/eurrev_202011_23556
45. Tong J, Ma X, Yu H, Yang J. SNHG15: A Promising Cancer-Related Long Noncoding RNA. Cancer Manag Res (2019) 11:5961–9. doi: 10.2147/CMAR.S208054
46. Zhang Y, Zhang D, Lv J, Wang S, Zhang Q. Lncrna SNHG15 Acts as an Oncogene in Prostate Cancer by Regulating Mir-338-3p/FKBP1A Axis. Gene (2019) 705:44–50. doi: 10.1016/j.gene.2019.04.033
47. Zhao W, Ma X, Liu L, Chen Q, Liu Z, Zhang Z, et al. SNHG20: A Vital Lncrna in Multiple Human Cancers. J Cell Physiol (2019). doi: 10.1002/jcp.28143
48. Wu X, Xiao Y, Zhou Y, Zhou Z, Yan W. Lncrna SNHG20 Promotes Prostate Cancer Migration and Invasion Via Targeting the Mir-6516-5p/SCGB2A1 Axis. Am J Transl Res (2019) 11(8):5162–9.
49. Mahdavifar N, Ghoncheh M, Pakzad R, Momenimovahed Z, Salehiniya H. Epidemiology, Incidence and Mortality of Bladder Cancer and Their Relationship With the Development Index in the World. Asian Pac J Cancer Prev (2016) 17(1):381–6. doi: 10.7314/apjcp.2016.17.1.381
50. Brausi M, Witjes JA, Lamm D, Persad R, Palou J, Colombel M, et al. A Review of Current Guidelines and Best Practice Recommendations for the Management of Nonmuscle Invasive Bladder Cancer by the International Bladder Cancer Group. J Urol (2011) 186(6):2158–67. doi: 10.1016/j.juro.2011.07.076
51. Matsumoto H, Shiraishi K, Azuma H, Inoue K, Uemura H, Eto M, et al. Clinical Practice Guidelines for Bladder Cancer 2019 Edition by the Japanese Urological Association: Revision Working Position Paper. Int J Urol (2020) 27(5):362–8. doi: 10.1111/iju.14210
52. Chen D, Guo Y, Chen Y, Guo Q, Chen J, Li Y, et al. Lncrna Growth Arrest-Specific Transcript 5 Targets Mir-21 Gene and Regulates Bladder Cancer Cell Proliferation and Apoptosis Through PTEN. Cancer Med (2020)9(8):2846–58. doi: 10.1002/cam4.2664
53. Liu Z, Wang W, Jiang J, Bao E, Xu D, Zeng Y, et al. Downregulation of GAS5 Promotes Bladder Cancer Cell Proliferation, Partly by Regulating CDK6. PloS One (2013) 8(9):e73991. doi: 10.1371/journal.pone.0073991
54. Zhang H, Guo Y, Song Y, Shang C. Long Noncoding RNA GAS5 Inhibits Malignant Proliferation and Chemotherapy Resistance to Doxorubicin in Bladder Transitional Cell Carcinoma. Cancer Chemother Pharmacol (2017) 79(1):49–55. doi: 10.1007/s00280-016-3194-4
55. Wang M, Guo C, Wang L, Luo G, Huang C, Li Y, et al. Long Noncoding RNA GAS5 Promotes Bladder Cancer Cells Apoptosis Through Inhibiting EZH2 Transcription. Cell Death Dis (2018) 9(2):238. doi: 10.1038/s41419-018-0264-z
56. Cao Q, Wang N, Qi J, Gu Z, Shen H. Long Noncoding RNAGAS5 Acts as a Tumor Suppressor in Bladder Transitional Cell Carcinoma Via Regulation of Chemokine (CC Motif) Ligand 1 Expression. Mol Med Rep (2016) 13(1):27–34. doi: 10.3892/mmr.2015.4503
57. Du L, Duan W, Jiang X, Zhao L, Li J, Wang R, et al. Cell-Free Lncrna Expression Signatures in Urine Serve as Novel Non-Invasive Biomarkers for Diagnosis and Recurrence Prediction of Bladder Cancer. J Cell Mol Med (2018) 22(5):2838–45. doi: 10.1111/jcmm.13578
58. Avgeris M, Tsilimantou A, Levis PK, Tokas T, Sideris DC, Stravodimos K, et al. Loss of GAS5 Tumour Suppressor Lncrna: An Independent Molecular Cancer Biomarker for Short-Term Relapse and Progression in Bladder Cancer Patients. Br J Cancer (2018) 119(12):1477–86. doi: 10.1038/s41416-018-0320-6
59. Li YH, Hu YQ, Wang SC, Li Y, Chen DM. Lncrna SNHG5: A New Budding Star in Human Cancers. Gene (2020) p:144724. doi: 10.1016/j.gene.2020.144724
60. Ma Z, Xue S, Zeng B, Qiu D. Lncrna SNHG5 is Associated With Poor Prognosis of Bladder Cancer and Promotes Bladder Cancer Cell Proliferation Through Targeting P27. Oncol Lett (2018) 15(2):1924–30. doi: 10.3892/ol.2017.7527
61. Wang C, Tao W, Ni S, Chen Q. Upregulation of Lncrna Snorna Host Gene 6 Regulates NUAK Family Snf1-Like Kinase-1 Expression by Competitively Binding Microrna-125b and Interacting With Snail1/2 in Bladder Cancer. J Cell Biochem (2019) 120(1):357–67. doi: 10.1002/jcb.27387
62. Chen Y, Peng Y, Xu Z, Ge B, Xiang X, Zhang T, et al. Knockdown of Lncrna SNHG7 Inhibited Cell Proliferation and Migration in Bladder Cancer Through Activating Wnt/Beta-Catenin Pathway. Pathol Res Pract (2019) 215(2):302–7. doi: 10.1016/j.prp.2018.11.015
63. Xu C, Zhou J, Wang Y, Wang A, Su L, Liu S, et al. Inhibition of Malignant Human Bladder Cancer Phenotypes Through the Down-Regulation of the Long Non-Coding RNA SNHG7. J Cancer (2019) 10(2):539–46. doi: 10.7150/jca.25507
64. Jiang B, Hailong S, Yuan J, Zhao H, Xia W, Zha Z, et al. Identification of Oncogenic Long Noncoding RNA SNHG12 and DUXAP8 in Human Bladder Cancer Through a Comprehensive Profiling Analysis. BioMed Pharmacother (2018) 108:500–7. doi: 10.1016/j.biopha.2018.09.025
65. Chen Z, Chen X, Xie R, Huang M, Dong W, Han J, et al. DANCR Promotes Metastasis and Proliferation in Bladder Cancer Cells by Enhancing IL-11-STAT3 Signaling and CCND1 Expression. Mol Ther (2019) 27(2):326–41. doi: 10.1016/j.ymthe.2018.12.015
66. Zhan Y, Chen Z, Li Y, He A, He S, Gong Y, et al. Long Non-Coding RNA DANCR Promotes Malignant Phenotypes of Bladder Cancer Cells by Modulating the Mir-149/MSI2 Axis as a Cerna. J Exp Clin Cancer Res (2018) 37(1):273. doi: 10.1186/s13046-018-0921-1
67. Li J, Wang AS, Wang S, Wang CY, Xue S, Guan H, et al. Lncsnhg14 Promotes the Development and Progression of Bladder Cancer by Targeting Mirna-150-5p. Eur Rev Med Pharmacol Sci (2019) 23(3):1022–9. doi: 10.26355/eurrev_201902_16989
68. Zhang C, Ren X, He J, Wang W, Tu C, Li Z. The Prognostic Value of Long Noncoding RNA SNHG16 on Clinical Outcomes in Human Cancers: A Systematic Review and Meta-Analysis. Cancer Cell Int (2019) 19:261. doi: 10.1186/s12935-019-0971-2
69. Zhang S, Du L, Wang L, Jiang X, Zhan Y, Li J, et al. Evaluation of Serum Exosomal Lncrna-Based Biomarker Panel for Diagnosis and Recurrence Prediction of Bladder Cancer. J Cell Mol Med (2019) 23(2):1396–405. doi: 10.1111/jcmm.14042
70. Cao X, Xu J, Yue D. Lncrna-SNHG16 Predicts Poor Prognosis and Promotes Tumor Proliferation Through Epigenetically Silencing P21 in Bladder Cancer. Cancer Gene Ther (2018) 25(1-2):10–7. doi: 10.1038/s41417-017-0006-x
71. Zhao Q, Gao S, Du Q, Liu Y. Long Non-Coding RNA SNHG20 Promotes Bladder Cancer Via Activating the Wnt/Beta-Catenin Signalling Pathway. Int J Mol Med (2018) 42(5):2839–48. doi: 10.3892/ijmm.2018.3819
72. Siegel RL, Miller KD, Jemal A. Cancer Statistics, 2019. CA Cancer J Clin (2019) 69(1):7–34. doi: 10.3322/caac.21551
73. Zhao Y, Yan Y, Ma R, Lv X, Zhang L, Wang J, et al. Expression Signature of Six-Snorna Serves as Novel Non-Invasive Biomarker for Diagnosis and Prognosis Prediction of Renal Clear Cell Carcinoma. J Cell Mol Med (2020) 24(3):2215–28. doi: 10.1111/jcmm.14886
74. Yang Y. Partial Versus Radical Nephrectomy in Patients With Renal Cell Carcinoma: A Systematic Review and Meta-Analysis. Urol J (2020) 17(2):109–17. doi: 10.22037/uj.v0i0.5358
75. Zhao S, Wang Y, Luo M, Cui W, Zhou X, Miao L. Long Noncoding RNA Small Nucleolar RNA Host Gene 1 (SNHG1) Promotes Renal Cell Carcinoma Progression and Metastasis by Negatively Regulating Mir-137. Med Sci Monit (2018) 24:3824–31. doi: 10.12659/MSM.910866
76. Toraih EA, Alghamdi SA, El-Wazir A, Hosny MM, Hussein MH, Khashana MS, et al. Dual Biomarkers Long Non-Coding RNA GAS5 and Microrna-34a Co-Expression Signature in Common Solid Tumors. PloS One (2018) 13(10):e0198231. doi: 10.1371/journal.pone.0198231
77. Qiao HP, Gao WS, Huo JX, Yang ZS. Long Non-Coding RNA GAS5 Functions as a Tumor Suppressor in Renal Cell Carcinoma. Asian Pac J Cancer Prev (2013) 14(2):1077–82. doi: 10.7314/apjcp.2013.14.2.1077
78. Liu L, Pang X, Shang W, Xie H, Feng Y, Feng G. Long Non-Coding RNA GAS5 Sensitizes Renal Cell Carcinoma to Sorafenib Via Mir-21/SOX5 Pathway. Cell Cycle (2019) 18(3):257–63. doi: 10.1080/15384101.2018.1475826
79. Fawzy MS, Toraih EA, Ageeli EA, Al-Qahtanie SA, Hussein MH, Kandil E. Noncoding Rnas Orchestrate Cell Growth, Death and Drug Resistance in Renal Cell Carcinoma. Epigenomics (2020) 12(3):199–219. doi: 10.2217/epi-2019-0120
80. Shi J, Li J, Yang S, Hu X, Chen J, Feng J, et al. Lncrna SNHG3 is Activated by E2F1 and Promotes Proliferation and Migration of Non-Small-Cell Lung Cancer Cells Through Activating TGF-Beta Pathway and IL-6/JAK2/STAT3 Pathway. J Cell Physiol (2020) 235(3):2891–900. doi: 10.1002/jcp.29194
81. Zhang C, Qu Y, Xiao H, Xiao W, Liu J, Gao Y, et al. Lncrna SNHG3 Promotes Clear Cell Renal Cell Carcinoma Proliferation and Migration by Upregulating TOP2A. Exp Cell Res (2019) 384(1):111595. doi: 10.1016/j.yexcr.2019.111595
82. Li WZ, Zou Y, Song ZY, Wei ZW, Chen G, Cai QL, et al. Long Non-Coding RNA SNHG5 Affects the Invasion and Apoptosis of Renal Cell Carcinoma by Regulating the Mir-363-3p-Twist1 Interaction. Am J Transl Res (2020) 12(2):697–707.
83. An HX, Xu B, Wang Q, Li YS, Shen LF, Li SG. Up-Regulation of Long Non-Coding RNA SNHG6 Predicts Poor Prognosis in Renal Cell Carcinoma. Eur Rev Med Pharmacol Sci (2018) 22(24):8624–9. doi: 10.26355/eurrev_201812_16626
84. Wu Z, Chen D, Wang K, Cao C, Xu X. Long Non-Coding RNA SNHG12 Functions as a Competing Endogenous RNA to Regulate MDM4 Expression by Sponging Mir-129-5p in Clear Cell Renal Cell Carcinoma. Front Oncol (2019) 9:1260. doi: 10.3389/fonc.2019.01260
85. Chen Q, Zhou W, Du SQ, Gong DX, Li J, Bi JB, et al. Overexpression of SNHG12 Regulates the Viability and Invasion of Renal Cell Carcinoma Cells Through Modulation of HIF1alpha. Cancer Cell Int (2019) 19:128. doi: 10.1186/s12935-019-0782-5
86. Jin L, Fu H, Quan J, Pan X, He T, Hu J, et al. Overexpression of Long Non-Coding RNA Differentiation Antagonizing Non-Protein Coding RNA Inhibits the Proliferation, Migration and Invasion and Promotes Apoptosis of Renal Cell Carcinoma. Mol Med Rep (2017) 16(4):4463–8. doi: 10.3892/mmr.2017.7135
87. Du Y, Kong C, Zhu Y, Yu M, Li Z, Bi J, et al. Knockdown of SNHG15 Suppresses Renal Cell Carcinoma Proliferation and EMT by Regulating the NF-Kappab Signaling Pathway. Int J Oncol (2018) 53(1):384–94. doi: 10.3892/ijo.2018.4395
88. Yang W, Zhang K, Li L, Ma K, Hong B, Gong Y, et al. Discovery and Validation of the Prognostic Value of the Lncrnas Encoding Snornas in Patients With Clear Cell Renal Cell Carcinoma. Aging (Albany NY) (2020) 12(5):4424–44. doi: 10.18632/aging.102894
89. Maschietto M, Piccoli FS, Costa CM, Camargo LP, Neves JI, Grundy PE, et al. Gene Expression Analysis of Blastemal Component Reveals Genes Associated With Relapse Mechanism in Wilms Tumour. Eur J Cancer (2011) 47(18):2715–22. doi: 10.1016/j.ejca.2011.05.024
90. Zheng H, Li BH, Liu C, Jia L, Liu FT. Comprehensive Analysis of Lncrna-Mediated Cerna Crosstalk and Identification of Prognostic Biomarkers in Wilms’ Tumor. BioMed Res Int (2020) 2020:4951692. doi: 10.1155/2020/4951692
91. Su L, Wu A, Zhang W, Kong X. Silencing Long Non-Coding RNA SNHG6 Restrains Proliferation, Migration and Invasion of Wilms’ Tumour Cell Lines by Regulating Mir-15a. Artif Cells Nanomed Biotechnol (2019) 47(1):2670–7. doi: 10.1080/21691401.2019.1633338
92. He HT, Xu M, Kuang Y, Han XY, Wang MQ, Yang Q. Biomarker and Competing Endogenous RNA Potential of Tumor-Specific Long Noncoding RNA in Chromophobe Renal Cell Carcinoma. Onco Targets Ther (2016) 9:6399–406. doi: 10.2147/OTT.S116392
Keywords: lncRNA, SNHG, prostate cancer, bladder cancer, kidney cancer
Citation: Yang Z, Li Q, Zheng X and Xie L (2021) Long Noncoding RNA Small Nucleolar Host Gene: A Potential Therapeutic Target in Urological Cancers. Front. Oncol. 11:638721. doi: 10.3389/fonc.2021.638721
Received: 07 December 2020; Accepted: 31 March 2021;
Published: 22 April 2021.
Edited by:
Mantang Qiu, Peking University People’s Hospital, ChinaReviewed by:
Carlo V. Bruschi, University of Salzburg, AustriaHernandes F. Carvalho, State University of Campinas, Brazil
Copyright © 2021 Yang, Li, Zheng and Xie. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Liping Xie, WGllbHBAemp1LmVkdS5jbg==
†These authors have contributed equally to this work