 Marij J. P. Welters
Marij J. P. Welters Saskia J. Santegoets
Saskia J. Santegoets Sjoerd H. van der Burg
Sjoerd H. van der Burg- Medical Oncology, Oncode Institute, Leiden University Medical Center, Leiden, Netherlands
Oropharyngeal squamous cell carcinoma (OPSCC) develops as a consequence of several mutations in the tumor suppressor pathways or after a progressive infection with high risk human papillomavirus (HPV). The dismal side effects of the current standard of care and the clear involvement of the immune system has led to a surge in clinical trials that aim to reinforce the tumor-specific immune response as a new treatment option. In this review, we have focused on the most recent literature to discuss the new findings and insights on the role of different immune cells in the context of OPSCC and its etiology. We then applied this knowledge to describe potential biomarkers and analyzed the rationale and outcomes of earlier and ongoing immunotherapy trials. Finally, we describe new developments that are still at the preclinical phase and provide an outlook on what the near future may bring, now that several new and exciting techniques to study the immune system at the single cell level are being exploited.
Introduction
Head and neck cancer is the sixth most prevalent cancer type and mainly consists of squamous cell carcinoma [90%; HNSCC (1, 2)]. The tumor can develop in the oral cavity, the larynx, pharynx (hypopharynx, nasopharynx, or oropharynx) and in the sinonasal tract. While the incidence of head and neck cancer located at most oral sites is marginally decreasing due to the knowledge of tobacco and alcohol as risk factors for its development, the incidence of oropharyngeal carcinomas is increasing, especially in the developed world (3). Oropharyngeal squamous cell carcinomas (OPSCC) include oropharyngeal, tonsillar and base-of-tongue tumors. A high percentage (60–80%) of the OPSCC are induced by high risk human papillomavirus type 16 (HPV16) (4–6). Patients with HPV+ OPSCC often tend to be younger, approximately 75% is male, a minority smokes and they often have lymph node metastasis when first visiting the clinic (7, 8). HPV is a double stranded DNA virus encoding for early and late (envelop) proteins (9, 10). The early proteins E6 and E7 are oncoproteins and responsible for the malignant transformation of HPV infected epithelial cells and maintenance hereof (11, 12). Patients with HPV16+ OPSCC display a longer overall survival (OS) and a lower recurrence rate after standard of care treatment than patients with HPV-negative OPSCC (13, 14).
Only recently the difference between these two OPSCC entities has been acknowledged and a debate on treatment has been started (15, 16). To understand the differences between the HPV+ and HPV-negative tumors, in-dept analysis have been performed on various levels including genetics (DNA), epigenetics, (micro)RNA and the immune system (7). De-intensified treatment, by lowering the dose of the (chemotherapeutic) drug or radiotherapy or by replacement of the drug, have been suggested in order to decrease side effects particularly in patients with HPV+ OPSCC (15, 16). This led to inferior survival and new treatment designs are required (17). Immunotherapy may form a new effective treatment in OPSCC and is currently under investigation. While it is clear that during HPV infection and subsequent transformation several ways are exploited to escape from the immune system (18–22), HPV16 E6/E7-specific T-cells are often detected in OPSCC tumors and their presence is associated with improved clinical outcome (6, 23, 24). In addition, a whole series of articles exists on the association between intratumoral T-cell infiltration and better clinical outcome after standard of care therapy (25–28). These type of data paved the way for immunotherapeutic strategies to harness the immune response to OPSCC. Indeed, blockade of CTLA-4 and/or PD-1 has been studied and showed a survival benefit in a subset of patients with different types of HNSCC when compared to the standard of care treatment arm (29). The vast majority of patients did not benefit from this treatment, illustrating the requirement for in-depth studies on systemic and local host-tumor interactions. In particular, studies on the tumor immune microenvironment (TME) are a prerequisite to understand what hurdles are at play and need to be overcome in order for the immune system to effectively control tumor growth.
A search in PubMed using the key terms “Oropharyngeal cancer immunity”, “Oropharyngeal cancer myeloid cells”, “Oropharyngeal cancer immunotherapy trial”, and “Head and Neck squamous cell carcinoma immunotherapy trial” was performed for articles published in the last 10 years (until august 2020) describing cohorts of patients with head and neck cancer, of which at least 25% had OPSCC and the results were not typical for one type of HNSCC, or with <25% of OPSCC patients but with results specific for the OPSCC group. Also, studies comparing the results between HPV-negative and HPV-positive HNSCC were included. In addition, a search in Clinicaltrials.gov for registered and ongoing trials in patients with HNSCC, including OPSCC, was performed. From these articles a selection was made to discuss the insights and developments in OPSCC immunity, biomarkers, and immunotherapy within the last decade. Last but not least, these studies were supplemented with general literature to explain concepts.
The Tumor Immune Microenvironment
The TME plays a pivotal role in the clinical behavior and response to different sorts of therapy of various cancer types (30). In-depth studies on the type, balance and interaction of immune cells in the TME and how this may affect clinical behavior could open the door to rationally designed strategies to treat patients with OPSCC (23).
Lymphocytes
The influx of high number of T cells was positively associated with clinical outcome of OPSCC after standard of care therapy (23, 31–33). In particular, strong infiltration by CD8+ and CD4+ T cells in pretreatment OPSCC tissues was associated with lower T stage, improved disease specific survival (DSS) and prolonged overall survival (OS). The clinical benefit of tumor infiltration with T cells was irrespective of HPV status of the tumor, albeit that HPV-induced tumors were more often strongly infiltrated by these T cells (31, 34–38). The functional activity of these tumor-infiltrating T cells is also important. Higher interferon gamma (IFNγ) and lower interleukin-4 (IL-4) and transforming growth factor-beta (TGF-β) cytokine expression levels were observed in HPV+ than HPV-negative OPSCC (39). In line with this, HPV-induced OPSCC contain high numbers of IFNγ or IL-17 producing CD8+ T cells (40) and the presence of IFNγ-producing CD4+ and CD8+ T cells in OPSCC, as inferred by the expression of the transcription factor T-box (Tbet) expressed in T cells, was related to better OS (6). Interestingly, the presence of IL-17-producing non-T cells in HPV+ OPSCC was associated with worse clinical benefit (38).
The analysis of freshly dissociated OPSCC tissues by mass cytometry revealed that HPV16+ OPSCC, comprising HPV16-specific T cells, also contained high numbers of CD161+ classical T cells, CD103+ tissue resident CD8+ T cells, dendritic cells (DCs) and DC-like macrophages (6). CD161+ T cells were highly activated, as shown by the high expression levels of PD-1, CD38 and HLA-DR, and were shown to be superior effector cells as they produced more IFNγ per cell (24). Notably, a higher frequency of CD161+CD4+ T cells was found to be associated with prolonged survival in OPSCC (24). The abundant presence of CD103+CD8+ T cells in HPV16+ OPSCC and its correlation with better prognosis was also demonstrated by others (41, 42). These CD103+CD8+ T cells represent non-circulating memory T cells that play a key role in local immunosurveillance (43) and are enriched for a number of genes associated with tissue resident cells (44). Single cell RNA analysis of 13 OPSCC revealed that these cells also expressed genes associated with cytotoxic potential, exhaustion/co-inhibition programmed cell death protein 1 (PD-1), lymphocyte activation gene 3 (LAG3), and T cell immunoglobulin and mucin domain-containing protein 3 (TIM3)) as well as activation/co-stimulation [CD27, inducible T cell costimulator (ICOS) and tumor necrosis factor receptor superfamily member 14 (TNFRSF14)] (45).
An important aspect in tumor control by T cells is the interaction with human leukocyte antigen (HLA) molecules presenting tumor-derived antigens at the cell surface of tumor cells, yet HLA expression is often lost or decreased in OPSCC (46, 47). Antigen presentation to CD8+ T cells may also be impaired by alterations in antigen processing pathway components such as the endoplasmic reticulum aminopeptidase 1 (ERAP1), an enzyme involved in trimming the N-terminus of peptide until it fits in the HLA class I molecule. Some polymorphisms in ERAP-1 were associated with high or low T cell infiltration of OPSCC. Interestingly, only the ERAP1 allotypes present in highly infiltrated HPV16+ OPSCC were capable of trimming model antigens, including HPV16 E7, to the proper HLA class I binding epitope (48). In contrast to HPV-negative OPSCC, there is no correlation between HLA class I expression and survival for HPV+ OPSCC (49, 50). The expression of HLA class II, which was more often found on tumor cells in HPV+ OPSCC, is associated with longer OS, DFS and disease specific survival (DSS) in OPSCC (46), supporting a role for CD4+ T cells in the control of OPSCC.
The Curious Relationship Between Regulatory T Cells and OPSCC Survival
Regulatory T cells (Tregs) act as gatekeepers of immunological tolerance, and dysfunction in Treg-mediated control plays an important role in autoimmune and allergic disorders and cancer. Tregs play a dismal role in tumor immunity, with numerous studies revealing their role in suppressing anti-tumor immune responses and promoting tumor progression in various types of cancer. In HNSCC, including OPSCC, there is conflicting evidence regarding their role in suppressing tumor immunity and survival (51). In two small scale studies, a negative impact on clinical outcome and disease progression was observed for high frequencies of Tregs in the peripheral blood of patients with OPSCC (52, 53), whereas other studies including OPSCC demonstrated no impact of tumor-infiltrating Tregs (54) or a positive impact of circulating or tumor-infiltrating Tregs (38, 55, 56) on clinical outcome and/or survival. A similar debate has been described for colorectal cancers (57, 58), resulting in the definition of three populations of CD4+Foxp3+ T cells, namely highly suppressive CD45RA-negative Foxp3hi effector Tregs (Foxp3hi eTregs), suppressive CD45RA+Foxp3int naïve Tregs (Foxp3int nTregs), and non-suppressive cytokine-producing CD45RA-negative Foxp3int T cells (59–61). Patients with colorectal tumors that were predominantly infiltrated with non-suppressive and cytokine-producing CD45RA-negative Foxp3int T cells displayed a much better survival than patients with tumors mainly infiltrated with suppressive Foxp3hi eTregs (60). This phenomenon may also play a role in studies that evaluated the clinical significance of intratumoral Tregs in OPSCC by means of immunohistochemistry (IHC), since discrimination between Foxp3int and Foxp3hi is difficult using this technique.
Part of the controversy on the prognostic value of Tregs may also be attributed to differences in the type of samples assessed, i.e. peripheral blood versus tumor tissues, and the absence of knowledge about the HPV status of the assessed tumors (51). The latter may be important as HPV-associated OPSCC is a distinct clinical entity with a different intratumoral immune cell make-up and a much better prognosis after (chemo)radiotherapy than HPV-negative OPSCC, particularly in patients with a concomitant HPV-specific and type 1-oriented intratumoral T cell response (6, 14, 32, 62).
Alternatively, differences in the effects of Tregs on clinical outcome may also come from their functional adaptability. Tregs can mirror effector cells by adopting the transcriptional profile of the cells they aim to suppress. In mice, Foxp3+ Tregs have been described to upregulate signal transducer and activator of transcription 3 (STAT3), GATA binding protein 3 (GATA-3)/interferon regulatory factor 4 (IRF4), or Tbet to control T helper type 17 (Th17), Th2 or Th1 inflammatory responses, respectively, in persistent infection and autoimmunity (63–66). We recently described that Tbet+ Tregs accumulate in tumors of HPV16+ OPSCC patients, and that their presence was associated with prolonged survival following standard of care therapy (67). Albeit detected at lower levels, the number of infiltrating Tbet+ Tregs strongly correlated with the number of tumor-infiltrating Tbet-positive CD4+ and CD8+ effector T cells as well as with the detection of HPV16-specific T cells, both of which we previously demonstrated to be associated with better tumor control on standard of care therapy. These data suggested that Tbet expression within the intratumoral Treg population and the association with prolonged survival is merely the reflection of an ongoing beneficial Th1-oriented T cell response, and that the balance between pro- and anti-tumor T cells eventually determines clinical outcome.
Myeloid Cells
The myeloid cell compartment constitutes another major player in the TME. One of the most abundant types of myeloid cells in the TME are macrophages. High numbers of tumor associated macrophages (TAMs) in the TME is most often associated with poor prognosis (68, 69). Macrophages are highly plastic and display a functional status ranging between classically activated anti-tumor type 1 macrophages (M1) and the alternatively activated tumor promoting type 2 macrophages (M2) (70). Incoming monocytes from the blood into the tumor can differentiate into macrophages dependent on the different cytokines they sense and the interactions they have with other cells present in the TME. While an inflamed TME with IFNγ results in M1 differentiation, high levels of interleukin 4 (IL-4) and IL-13 turn them into M2 (71). M2 can produce cytokines, such as TGF-β, vascular endothelial growth factor (VEGF), IL-6, IL-10, and prostaglandin E2 (PGE2), and chemokines that promote angiogenesis or destruct the extracellular matrix, thereby helping tumor cells to invade and metastasize and they can stimulate Treg development and expansion while suppressing CD8+ T cell function (72, 73). Moreover, in response to inflammation (IFNγ production) or hypoxia TAMs can upregulate inhibitory receptor ligands for T cells like PDL-1 and PDL-2 (69, 72).
CD68+CD163+ M2 macrophages are often detected in OPSCC (46, 74). A higher infiltration with these macrophages was associated with poor clinical outcome in two studies on HNSCC tumors with the majority or all being OPSCC (46, 75). The strong stromal presence of CD68+CD163+ macrophages was associated with shorter DFS, DSS and OS, independent of HPV status in OPSCC (46), but this does not mean that all macrophages are bad as high infiltration with macrophages, the minority being M2, was associated with better PFS in p16+ OPSCC (47). This suggests that other macrophages may benefit tumor control. Indeed, in various tumor types high infiltration with M1 (CD14+CD163-negative) macrophages is associated with prolonged survival (76, 77). It also implies that simply depleting all macrophages in the TME may have detrimental effects and even accelerate tumor growth or increase recurrences (72, 78, 79). Therefore, reprogramming TAMs is currently under investigation in solid tumors, to move the balance from pro-tumorigenic towards tumor fighting macrophages (72, 80) and it is highly likely that also a strong Th1 cell response may aid in this process (73).
Similar to TAMs also myeloid derived suppressor cells (MDSCs) can be recruited from the blood or generated locally by arresting monocytes in their immature state. The phenotype of these MDSC is often described as CD14+ HLA-DR-CD33+CD11b+ (81, 82), hence monocytic MDSC (mMDSC). High circulating numbers of these cells are associated with metastasis and recurrences in OPSCC patients (83). Interestingly, treatment with tadalafil, a phospohodiesterase 5 inhibitor to block nitric oxide and arginase 1 production by myeloid cells, resulted in the activation of a stronger tumor-specific T-cell response with no difference in responsiveness between HPV16+ OPSCC and HPV-negative (non)oropharyngeal cancer patients (84). Several other studies to deplete mMDSCs, prevent their recruitment or block their suppressive function have been undertaken (85). Next to these mMDSC also granulocytic MDSC (gMDSC) and neutrophils may enter the tumor to suppress effector T cells in the TME. However, no studies on the role of these cells in the TME of OPSCC have been reported.
Finally, recruited monocytes can also differentiate into DCs. High numbers of CD14+CD11b+CD11c+ DC-like macrophages and CD14-negative CD11bdimCD11c+ DCs have been detected in the TME of OPSCC patients with a good clinical outcome (6). In another study, high numbers of stromal CD11c+ DC was an independent prognostic factor for better survival in HNSCC, including OPSCC (55). Current studies have analyzed DCs by only a few markers. Recently, two in-depth studies on blood derived DCs have divided these in several phenotypic and functional subtypes (86, 87). We applied high-dimensional flow cytometry and multispectral imaging to reveal a new subset of DCs, called CD163+ cDC2 or DC3, the presence of which was directly related to survival in HPV16+ OPSCC (74). Moreover, these DCs displayed the capacity to specifically trigger type 1 T cell responses by their production of IL-12 and IL-18 (74) and may be involved in conferring a tissue-resident signature to stimulated T cells (88).
Expression of Immune Checkpoints
Intratumoral T cell reactivity is regulated via so-called co-stimulatory and co-inhibitory (or checkpoint) molecules. Well-known is the suppression of T cells expressing PD-1 via PD-1 ligand (PD-L1) and blockade of this axis has resulted in spectacular clinical responses for a number of tumor types. Analyses of checkpoint expression in OPSCC revealed that the expression of PD-1 and/or PD-L1 was related to a stronger immune infiltration and good prognosis after standard therapy (89–92), most likely as it reflects an ongoing immune response in which type I and II interferons are produced. The presence of intratumoral PD-L1 expressing CD68+ macrophages and CD8+ T cells was found to be associated with improved OS (93). In addition, rich immune infiltration, comprising PD1+CD8+ T cells and CD68+ macrophages, was found to be associated with a better clinical response to checkpoint therapy (94). While the numbers of infiltrating total CD8+ T cells and CD68+ macrophages were higher in HPV+ OPSCC, the percentage of CD8+PD-1+ T cells was similar, and the percentage of CD68+ PD-L1+ macrophages lower in HPV+ OPSCC compared to HPV-negative OPSCC (95).
Another actionable co-inhibitory molecule is natural killer group 2 member A (NKG2A) (96, 97), which together with its co-receptor CD94 is expressed by many of the tumor-infiltrating CD8+ T cells and only by a minority of the CD4+ T cells in OPSCC (45). Remarkably, NKG2A expression on CD8+ in OPSCC is independent from PD-1 and often found on CD103+ early effector tissue resident CD8+ T cells (45, 97). The frequency of intratumoral NKG2A/CD94+ CD8+ T cells was higher in in HPV16+ OPSCC patients with a demonstrable ongoing HPV16-specific T cell response when compared to HPV16+ OPSCC lacking such an anti-tumor response or to HPV-negative OPSCC patients (6, 97). NKG2A interacts with HLA-E, which is a non-classical highly-conserved HLA class I molecule that is expressed by many cancers (96, 98, 99), including OPSCC (50). The interaction between NKG2A and HLA-E is thought to block the cytotoxic activity of CD8+ T cells and NK cells (100) and a couple of studies have shown that expression of HLA-E by tumor cells restrained the prognostic impact of tumor-infiltrating CD8+ T cells (98, 99), including that of HPV16+ OPSCC (97).
Other inhibitory receptors found to be upregulated on activated T cells in the TME of OPSCC include TIM3, LAG3 and T cell immunoreceptor with Ig and ITIM domains (TIGIT) and others (45). All expressed on higher numbers of T cells in HPV+ when compared to virus-negative head and neck tumors, but only in HPV+ tumors each of these markers was associated with prolonged survival (101).
Overall, the expression of inhibitory receptors are more indicative for an inflamed TME with ongoing antitumor immunity than for an exhausted T cell response in OPSCC. Nevertheless, the interaction between inhibitory receptors and their ligands will inhibit the activation and effector functions of T cells impairing their capacity to control OPSCC growth.
The Blood Compartment for Biomarker Analysis
An important question is whether the TME biomarkers associated with clinical outcome are also detectable and prognostic when analyzed on immune cells present in blood, as this compartment is easily accessible and allows for kinetic studies. The easiest approach is to determine differential leukocyte counts on blood samples, which is used in all hospitals as a normal diagnostic routine. High neutrophil counts in OPSCC, and more specifically high neutrophil-to-lymphocyte ratio (NLR) in the blood sample prior, during and after radiotherapy correlated with poor OS, recurrence free survival (RFS) and/or DSS as well as distant metastasis (102–105). Also, in HPV16+ OPSCC patients, a high NLR in the blood sample obtained prior to concurrent chemoradiation correlated with decreased OS. Neutrophils appear to have an unique phenotype of immature granulocytes CD11cbright/CD62Ldim/CD11bbright/CD16bright, which are absent in blood of healthy donors (106).
High circulating monocyte counts prior to therapy also correlated to reduced OS and RFS in HPV16+ OPSCC patients (102, 107). While monocyte counts in HPV-negative OPSCC patients were even higher, the absolute monocyte counts were not related to clinical outcome in this patient group (102), suggesting that these monocytes might have a negative impact only in highly immunogenic tumors. Potentially, the composition of the monocyte subtypes may be of importance as some of them may constitute CD14+ MDSC, which have shown to exhibit immune suppressive effects (6, 108). Furthermore, the percentage of classical (CD14highCD16-negative) monocytes were shown to be increased in OPSCC patients as well as expressed PD-L1 (107). Also a high monocyte-to-lymphocyte ratio was associated with lower OS in OPSCC (107, 109).
So far, no comparative studies have been performed with respect to neutrophils and MDSC in order to analyze if the blood compartment reflects the TME.
The relation between higher numbers of tumor-infiltrating T cells and clinical outcome prompts the question if this can also be detected in the blood. High pre-treatment numbers of CD8+ T cells in the blood correlated with improved OS in HPV16+ OPSCC patients (14). However, absolute CD4 counts as well as frequencies of CD4+ T cells measured in blood of OPSCC was not correlating to clinical outcome (14). This seems logical as the population of CD4+ T cells comprises naïve T cells, type 1 and type 2 effector T cells, memory T cells and also Tregs which dilutes the real impact of one of these subpopulations. Moreover, the peripheral blood levels of CD4+ and CD8+ T cells were not related to the type and degree of specific T cell subset infiltration in OPSCC (34). Interestingly, it was found that HPV+ OPSCC with elevated Treg frequencies in the blood displayed a better OS than patients with HPV-negative tumors or patients with HPV+ OPSCC patients displaying relatively low numbers of circulating Tregs (56).
Last but not least, HPV16-specific T cells have been detected in the circulation of patients with OPSCC using functional assays in which PBMC were stimulated with the HPV-encoded antigens (110–112). The detection of such an HPV16-specific T cell response in the peripheral blood did not always coincide with the presence of these HPV16-specific T cells in the TME, this (110). HPV16-specific T cells could also be detected in a large percentage of healthy individuals (113–115) indicating that their presence is not enough to function as a biomarker.
All together there are a couple of opportunities (neutrophils, monocytes, Tregs) that may be exploited as blood biomarkers reflecting the TME, but more sophisticated measurements of subpopulations and validation of the used techniques are warranted.
The Role of Tumor-Specific T Cells in the TME of OPSCC
There is ample evidence that tumors highly infiltrated with CD4+ and/or CD8+ T cells (also referred to as immunologically “hot” tumors) respond better to (immuno-)therapy (27, 116). In OPSCC, HPV infection may play a very important role in this phenomenon. Superior prognosis of HPV+ OPSCC over HPV-negative OPSCC following chemoradiotherapy has been reported (13, 117). Interestingly, for the majority of HPV+ OPSCC this was related with a more dense and activated T cell infiltrate, suggesting a role for HPV-specific immunity in tumor control (14, 62, 118).
HPV16+ tumors express the virally derived oncoproteins E6 and E7, and it has been suggested that these “non-self” HPV antigens can evoke immune responses. Indeed, cellular and humoral immunity against these antigens can be detected in the peripheral blood and TME of HPV16+ OPSCC patients (110, 119–123). Notably, there is a direct link between the presence of an intratumoral HPV-specific T-cell response and the good prognosis of HPV16-driven OPSCC (6). Patients with an HPV16-driven tumor and a concomitant intratumoral HPV16-specific T cell response have a much better survival after (chemo)radiotherapy than patients with an HPV16-driven tumor without such an HPV16-specific T cell response or patients with HPV-negative OPSCC. Moreover, the presence of these HPV-specific T cells was associated with a type 1 oriented TME and a much higher level of activated CD161-expressing effector memory CD4+ T cells,CD103+ tissue-resident CD8+ T cells, and tissue-resident memory T cell-stimulating CD163+ cDC2 (6, 74).
Although a clear relation could be found between the dense, type 1 oriented HPV-specific immune infiltrate and disease outcome in HPV16-driven OPSCC, there is also evidence for improved survival of HPV-negative OPSCC patients with strongly T-cell infiltrated tumors (37, 124). Unfortunately, knowledge on the tumor-specificity is lacking in these studies. A promising and emerging field in studies of the anti-tumor response is the recognition of tumor neoantigens. Tumors with high numbers of nonsynonymous mutations have a greater likelihood of presenting mutation-derived neoepitopes and consequently mounting a T-cell response against these epitopes thereby improving the clinical response (125). Tumor mutational burden (TMB) in HNSCC is relatively high and comparable to other smoking related tumor types. Viral HNSCC display only half of the mutations rate observed in non-viral HNSCC (125, 126). CD8+ T cells responding to such mutations have been detected in a few patients with non-OPSCC either spontaneously induced (127) or following a complete response after pembrolizumab treatment (128). Notably, such neoantigen specific T cells can also be detected in low TMB tumors (129), suggesting that they may also be present in OPSCC.
Immunotherapy in OPSCC
During the last two decades, several strategies aiming to boost the immune response to cancer have been developed and tested in patients with cancer, ranging from immune modulators to checkpoint therapy, adoptive cell transfer, and vaccines. Table 1 summarizes active/recruiting immunotherapy trials in patients with OPSCC and HNSCC, including OPSCC.
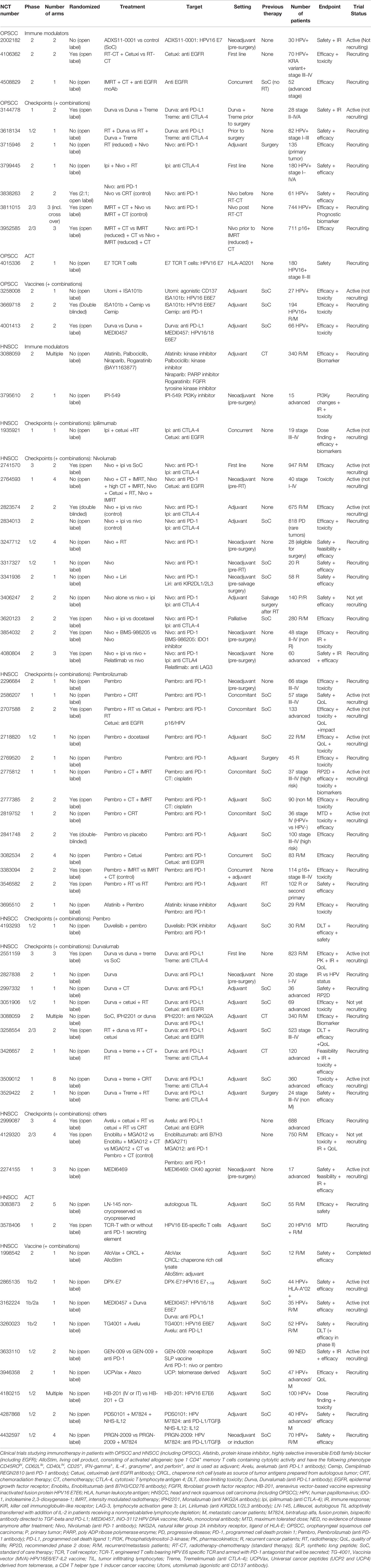
Table 1 Immunotherapy in patients with OPSCC.
Immune Modulators
One of the earliest immune stimulators tested is recombinant IL-2 (rIL-2), used to directly stimulate the activity of tumor-specific T cells. In a phase III trial with 200 patients rIL-2 was injected close to the ipsilateral lymph node, each day for 10 days before surgery and radiotherapy and then each month at the contralateral LN site for 1 year. The treatment had no significant side effects but improved the DFS (from 51 to 64%) and OS (from 55 to 73%) at 5 years (130). At a later stage, IRX-2 was developed to stimulate T cell activity. IRX-2 is a mix of purified cytokines obtained after stimulating peripheral blood mononuclear cells (PBMC) with phytohemagglutinin (PHA). In a phase IIa trial treatment naïve patients with HNSCC received subcutaneous IRX-2 injections near the TDLN for 10 days prior to surgical resection, which with the knowledge of today is an interesting choice as our studies showed that the great majority of TDLN comprise tumor-specific T cells even when they are hardly detectable in the tumor itself (24, 131). Injection of IRX-2 was associated with small radiological reductions in tumor size, including patients with OPSCC (132). In a first analysis, pre-and-post IRX-2 samples were analyzed and showed marked increases in CD3+ T cells and CD68+ macrophages (133). In a subgroup of 7 patients pre-and post-samples could be compared, showing potential increases in CD4+ T cells (6/7) and CD68 macrophages (4/7) as well as decreases in CD8+ T cells (5/7) patients after IRX-2 treatment. Unfortunately, HPV status was not determined but one could envision that most of the responders would have a HPV-positive tumor. Nanostring analyses using the PanCancer IO360 immune profiling panel confirmed the increases in CD4+ T cells and most markedly in DC as well as in the immune modulatory cytokines IL-6 and IL-10 (134). Another compound that indirectly could stimulate the tumor-specific T cell response is the toll-like receptor 8 (TLR8) agonist motolomid, which is known to activate monocytes, DCs and NK cells (135). Treatment of recurrent or metastatic HNSCC patients after the first 6 cycles of chemotherapy with weekly infusions of cetuximab with or without motolomid did not result in improved OS in a trial with n=195 patients. However, in a prespecified subgroup of n=83 OPSCC patients the motolomid arm displayed longer PFS and OS when the tumor was induced by HPV (136), suggesting that motolomid may have boosted the existing but probably weak HPV-specific T cell response in these patients.
Other compounds target immune suppressive cells that are active in the TME. For instance, metformin, which is widely used as a drug to manage diabetes type 2, has been described to have anti-cancer effects (137). In mice, metformin increased the number of tumor infiltrating CD8+ T cells, and the combination of metformin and a tumor vaccine improved the multifunctionality of the vaccine-induced CD8+ tumor infiltrating lymphocytes (TIL) (138). Another potential mechanism is the effect of metformin on Tregs, as it also inhibits mammalian target of rapamycin (mTOR). Metformin treatment for a minimum of 9 days before surgery of HNSCC, more than half being OPSCC, resulted in a strong decrease of intratumoral Foxp3+ Tregs and increase of stromal CD8+ T cells between pre-and post-treatment samples (n=16 HPV+ and n=20 HPV-negative), independent of HPV status (139). Potentially, metformin treatment may be interesting for HPV+ OPSCC patients with ongoing tumor immunity. As high numbers of Tregs have infiltrated those tumors (67), the use of metformin may tip the balance in favor of spontaneous anti-tumor responses in these patients, resulting in tumor shrinkage. Similarly, it may provide benefit in combination with other types of immunotherapy. Several new immune modulators are currently tested in the clinic (Table 1).
Checkpoint Blockade Therapy
The use of antibodies that block the interaction between PD-1 and PD-L1 are successfully used in several types of immunogenic cancers with a high mutation rate (e.g. melanoma, lung cancer) or induced by the Merkel cell virus (140–142). Several studies have been reported on the efficacy of such antibodies for the treatment of HNSCC, including patients with OPSCC (143–147).
In CheckMate 141, a phase III trial, platinum-resistant patients with recurrent or metastatic tumors were treated with the anti-PD-1 antibody nivolumab. The response rate was 13.3% in the nivolumab group versus 5.8% in patients treated with standard of care. The objective response rate (ORR) to nivolumab was 15.9% in HPV+ patients while it was 8.0% in HPV-negative patients (143). The 2-year OS in this study was 16.9% in patients with PD-L1 expression, regardless of the HPV status (148). Nivolumab also induced clinical benefit in patients lacking PD-L1 expression, albeit in a lower proportion of patients (148). Interestingly, continued treatment of patients with nivolumab with slow progressing disease still resulted in clinical benefit, irrespective of the HPV status of the tumor. This was associated with a decrease in the percentage of PD-1+ Tregs in PBMC (149), suggesting that an ongoing immune response was present in these patients but which was simply too weak to be effective. Importantly, nivolumab treatment improved OS irrespective of prior treatment with cetuximab, an antibody known for its immune modulating side effects (150–153), albeit that the reduction in risk of death was lower in pre-treated patients (154). A similar response rate with PD-1 blockade using pembrolizumab was found in the KEYNOTE-012 phase Ib study of 60 patients with recurrent or metastatic tumors with PD-L1 expression. In total 18% of the patients displayed a clinical response, divided as 25% (4/16) in the HPV+ HNSCC patient group and 14% (4/29) in HPV-negative group (144).
The PD-L1 antibody durvalumab was tested in the expansion phase of a phase I/II study for treatment of patients with recurrent or metastatic tumors, 40% of which were HPV+. The ORR was 6.5% (4/62) in the total group with 15% in the PD-L1 expressing subgroup and 2.6% in the group with low PD-L1 expression. None of the HPV+ patients showed a response (145).
Another important checkpoint molecule is NKG2A. Preclinical experiments revealed that anti-NKG2A therapy promoted tumor immunity and synergized with PD-1 blockade. In addition, it was shown that NKG2A blockade improved the efficacy of tumor vaccines and adoptive cell therapy (ACT) (96, 97). The humanized NKG2A antibody monalizumab was tested together with cetuximab in previously treated recurrent or metastatic tumors, including 32% OPSCC. A preliminary report on the outcome of the trial revealed a confirmed clinical partial response (PR) in 8 out of 26 patients (31%) and stable disease (SD) in 14 out of 26 patients, which was regarded as promising when compared to historical cetuximab data (96).
In comparison to the high ORR in virus-driven Merkel cell carcinoma after checkpoint therapy (56% (142)), the response in HPV+ patients to checkpoint therapy is disappointingly low. However, in view of the fact that patients with both an HPV+ tumor and a strong immune response to HPV display an excellent response to standard of care therapy (6, 24), this result was to be expected. The majority of HPV+ patients with recurrent or metastatic tumors most likely are those in whom there was no or a very weak intratumoral T-cell response to HPV during first line treatment and it is known that checkpoint blockade works best in patients with a pre-existing tumor-specific immune response (155). Hence, in OPSCC these therapies should be combined with modalities that enhance the tumor-reactive T cell response in patients.
Adoptive Cell Therapy
The infusion of large numbers of tumor-reactive T cells, being either ex-vivo expanded TILs, T-cell receptor (TCR) transduced T cells or chimeric antigen receptor (CAR)-T cells is one way to enhance the number of tumor-reactive T cells and has proven to be clinically effective in patients with advanced cancer (156–158). The presence of two strong tumor-specific antigens E6 and E7 in HPV-driven cancers provides the opportunity to specifically stimulate and expand tumor-specific T cells for ACT. Stimulation of peripheral blood lymphocytes with overlapping peptide pools resulted in the expansion of HPV16-specific T cells in 33 of 52 HPV16+ OPSCC patients. Most of the cell cultures comprised HPV16-specific CD4+ T cells and infrequently CD8+ T cells with the capacity to kill target tumor cells (122). A highly reproducible method to stimulate these cells under good manufacturing practice (GMP)-conditions resulted in the expansion of IFNγ-producing HPV-specific CD4+ T cells (11 of 11 cases) and CD8+ T cells (3 of 11) cases (131). In a phase II ACT trial, one out of 5 OPSCC patients resolved most of the metastatic lesions (complete response, CR) except for one brain metastasis, which was resected. Responding patients received higher numbers of IFNγ-producing HPV-specific TILs and the frequency of HPV-specific T cells in the blood one month after ACT correlated with clinical outcome. The question is how HPV-specific ACT will be used as most patients of whom their primary tumors display HPV-specific TIL reactivity respond well to chemoradiation while the tumors of most patients with recurrent disease are expected to contain no or few HPV-specific TILs (6). If present in the tumor, ex-vivo expansion and reinfusion of HPV-specific TIL may help to prevent or to reduce chemoradiation as primary treatment.
The absence of HPV-specific T cells mandates alternative approaches such as the infusion of ex-vivo TCR-transduced autologous T cells. Draper et al. reported the isolation of an HLA-A*0201 restricted HPV16 E629-38-specific TCR from a patient with HPV16-induced anal cancer. The use of this TCR is expected to contribute to tumor immunity as the tumor site was highly enriched for this TCR clonotype when compared to blood and T cells transduced with this TCR were shown to recognize several HPV16+ cell lines of different origins (159). Another study reported the isolation of an HLA-DRB1*04 restricted TCR reactive to the HPV16 E770-89 epitope. TCR-transduced cells were shown to specifically produce IFNγ when stimulated with HPV16 E6 and E7 containing tumor lysate (160). The isolation of TCR responsive to HPV epitopes restricted by a series of HLA class I and II molecules will lead to a warehouse of HPV-specific TCR required for personalized treatment of patients with recurrent or metastatic HPV-induced OPSCC. Currently, the use of T cells transgenic for an E7-specific TCR is tested in the clinic (Table 1). The fact that most mutations (neoantigens) are patient specific makes a similar approach for HPV-negative OPSCC complicated but not impossible (161).
Vaccines
Therapeutic vaccination is another option to increase the number of tumor-reactive T cells in OPSCC, preferentially against tumor-specific epitopes created by DNA mutations or oncogenic viruses. Based on a whole series of different trials it was concluded that the induction of tumor reactivity correlated with clinical outcome after vaccination in patients with pre-cancers or in settings of low tumor burden, and with tumor regressions if broad type 1 T cell reactivity was established (162). The treatment of most established cancers will require the combination of therapeutic vaccination with other modalities to overcome immune suppression and escape (163).
Two types of vaccines, one aiming to induce responses to MAGE-A3 and HPV and the other aiming to induce T-cell reactivity against p53 (for HPV-negative tumors) were tested in groups of HNSCC patients, including 30–40% OPSCC patients. These vaccines displayed a weak immunogenicity and did not alter clinical outcome (164, 165).
However, in a single arm phase II trial, designed based on the observation that HPV16+ OPSCC patients lacking tumor infiltration with HPV16-specific T cells are most likely the ones that are diagnosed with incurable HPV16+ cancer (6, 24) and that PD-1 checkpoint therapy has most impact in patients with pre-existing tumor-specific T cell responses (155), HPV16+ OPSCC patients received a combined treatment with ISA101 and nivolumab (146). ISA101 is an HPV16 -E6 and -E7-specific synthetic long peptide vaccine, which previously was shown to induce CR and PR in patients with HPV16-induced high-grade but premalignant disease (166, 167). The ORR of 33% and median OS of 17.5 months was regarded promising (146) when compared to PD-1 inhibition only in a similar patient group (143) or when compared to the study with durvalumab in which none of the HPV+ patients responded (145). A randomized trial comparing the efficacy of PD-1 blockade alone versus vaccination + PD-1 blockade is underway (Table 1). The outcome of the combination ISA101 and nivolumab trial argue for tumor-specific vaccination strategies. Approaches to stimulate the neoantigen repertoire are also warranted for the treatment of in particular HPV-negative OPSCC, similar to what has been done in other types of cancers (168–170).
Another option to stimulate tumor-reactive T cells might be through irradiation, as this has been shown to result in the release of antigens from tumor cells as well as in the induction of proinflammatory signals that trigger the innate immune system (171). Upon radiation, the tumor may serve as an in situ vaccine with the capacity to activate tumor-specific T cells. Spurred by the results with checkpoint blockade after radiation therapy in lung cancer (172), new trials are initiated in patients with HNSCC, including OPSCC (Table 1), using radiotherapy and checkpoint inhibition together or in combination with chemotherapy (173–175). The feasibility of using PD-L1 antibody avelumab with conventional cetuximab-radiotherapy as an alternative for advanced stage OPSCC patients unfit for cisplatin treatment was tested in a phase I study with 8 patients with OPSCC, 4 of which were HPV+. Immune toxicity was transient and manageable, and objective responses were observed in 4 of 6 evaluable OPSCC patients (3 complete responses and one partial response), including 3 HPV+ patients (176).
Preclinical Developments
There are several mouse models for HPV+ HNSCC. Study of cell surface proteins expressed by MTEC tumors, made from mouse tonsillar epithelial cells that are transformed by HPV16 E6, E7, and hRAS, showed the expression of CD47 (177). By binding to signal-regulatory protein alpha (SIRP-alpha) expressed on antigen processing cells, this transmembrane molecule blocks the phagocytosis and clearance of cells expressing CD47, resulting in the suppression of innate and adaptive anti-tumor responses. Targeting of CD47 using antibodies in rituximab-refractory non-Hodgkin’s lymphoma re-sensitized about 50% of these patients (178). The CD47-SIRP-alpha axis can also be targeted pharmacologically (179). The efficacy of radiation, which is a key component of the standard of care treatment for OPSCC, was shown to depend on intact T cell responses and was improved after CD47 knock-down in MTEC tumors (177), suggesting that CD47 may also play a key role in regulating immunity and as such the efficacy of standard of care treatment in OPSCC. Another study with mEER tumor cells, derived from the metastases of an HPV+ oropharyngeal murine cancer, injected into the flank of mice showed that the response to standard cisplatin-radiation therapy could be improved by adding cyclophosphamide and an inducible nitric oxide synthase (iNOS) inhibitor. Standard therapy did not alter the tumor microenvironment, which remained cold as indicated by high numbers of immune suppressive immune cells (MDSC, Tregs) and low numbers of T cells, M1 macrophages, and DCs (180). The addition of cyclophosphamide to the standard treatment converted treated tumors to hot but the combination with the iNOS inhibitor resulted in a strong influx of CD8+ and CD4+ Th1 cells as well as lead to durable control of established tumors (180). The use of cisplatin chemotherapy has been shown to increase tumor immune infiltration (181) and tumor cell killing by tumor necrosis factor alpha (TNFα)-producing T cells (182). The use of a low dose cisplatin was also shown to augment the effects of an adenovirus-based oncolytic virus therapy of subcutaneously injected mEER tumors. The effects of the oncolytic virus were clearly CD8+ T cell dependent and cisplatin was shown to augment the infiltration of the mEER tumors with HPV16 E7-specific CD8+ T cells (183). Interestingly, the anatomical location of the mEER tumors had a profound impact on the composition of the tumor infiltrating immune cells (184), quite reminiscent of what was observed in patients with HPV16-induced OPSCC and cervical cancer (24). Orthotopic injection of mEER tumor cells resulted in a much more inflamed tumor immune microenvironment, reflected by higher numbers of tumor-infiltrating CD8+ T cells and a stronger type 1 and 2 gene signature, than when these tumor cells were injected in the flank. Moreover, whereas the orthotopic growing mEER tumors were directly responsive to treatment with anti-PD-1 (> 50% survival) and even better to the combination of anti-PD-1 and anti-CTLA-4 (> 90% survival), flank injected tumors did not show any response to PD-1 blockade alone whereas only 40% responded to the combination treatment. Intratumoral injections of STING agonist, to increase IFN signaling, resulted in a strong decrease of Tregs and MDSC and an almost complete eradication of flank tumors (184). The decrease of immune suppressive immune cells in flank-injected mEER tumors seems essential for a better outcome after both standard chemoradiation (180) and immunotherapy (184).
Perspectives
The use of 15 parameter flow cytometry, the development of over 30 marker panels by mass cytometry and the introduction of unbiased bioinformatical approaches to cluster cells based on the expression of all proteins assessed have led to a much better definition and quantification of the immune cell phenotypes that are present in OPSCC, solved debates on the prognostic role of certain subtypes and led to the identification of subtypes strongly associated with clinical outcome. The use of single cell RNA sequencing to study the transcriptome of intratumoral cells has provided new means to identify subtypes of immune cells and understand their functional properties as well as defined gene expression signatures with clinical outcome. In a first study of 26 patients with all types of HNSCC, including 7 OPSCC, the transcriptional signatures in helper CD4+ T cells and B cells were quite divergent between HPV-negative and HPV+ tumors, whereas that of CD8+ T cells and Tregs was quite similar (185). These results may not necessarily reflect a difference caused by HPV but can be caused by the different types of tumor and their location (6, 24, 184). In a similar, but purely OPSCC and T cell focused transcriptome study, highly active tumor resident and tumor-reactive populations of CD4+ and CD8+ T cells that displayed actionable checkpoints were found in OPSCC (45). Whilst these techniques give an unprecedented insight in the complexity of the immune infiltrate of tumors, they lack the spatial information on each cell. The fact that a fairly simple classification based on the distribution of T cells has a major impact on prognosis and response to immunotherapy stresses the importance of such analyses (27). Multispectral imaging using the Vectra not only allows for a more in-depth analyses of different types of immune cells within the same tumor section but also to study their interaction. The use of mass cytometry based imaging allows for many more markers to be studied in a spatial context (186) and one can foresee that this will bring a much deeper understanding on the types of cells in the TME as well as their interaction and how this affect clinical outcome.
The much better outcome of HPV+ OPSCC compared to their HPV-negative counterparts after standard of care therapy prompted discussions on de-intensified treatments, specifically with respect to the dose of cisplatin chemotherapy (15), but this led to inferior survival (17). It should be realized that the reason why patients with HPV+ tumors do better on this standard therapy is because of their extensive immune infiltration and this is enhanced by cisplatin (67, 180). Thus, rather than downscaling the use of chemotherapy one should consider to make optimal use of it. For instance, by taking advantage of the fact that chemotherapy may remove some of the immunosuppressive mechanisms that are at play in HPV-induced cancers (187, 188). Rational combination of the immunomodulatory properties of chemotherapy and radiation that turns up the heat in tumors are highly warranted. Based on a recent publication on therapeutic vaccination (189), it is unlikely that checkpoint therapy or ACT approaches will manage to do this. Potentially, oncolytic virus therapy may aid in this (NCT02626000).
Finally, checkpoint therapy has only limited effect in patients with head and neck cancer. One of the problems may be that these patients failed to or mounted only a weak response to tumor antigens. Whereas this can be achieved with therapeutic vaccines in patients with HPV+ tumors, this is more difficult when the tumor antigens are unknown, as is the case in HPV-negative OPSCC. To increase the number of tumor-reactive T cells, approaches are undertaken to either transfer TCR transgenic T cells into patients or by isolation and expansion of T cells which are most likely to be enriched for tumor-reactive T cells using the expression of the activation markers CD137, PD-1, or CD39 to select the T cells (190–193). These approaches have mostly targeted tumor-reactive CD8+ T cells, but in view of the important role (194–196) and efficacy of tumor-reactive CD4+ T cells (197–199) strategies to rapidly isolate that specific T cell fraction are needed. This may be achieved using the expression of CD39 on tumor-infiltrating CD4+ T cells (45).
Author Contributions
The authors contributed equally to the review. All authors contributed to the article and approved the submitted version.
Funding
This study was financially supported by grants from the Dutch Cancer Society 2016–10726 to SB, MW, and SS.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Shield KD, Ferlay J, Jemal A, Sankaranarayanan R, Chaturvedi AK, Bray F, et al. The global incidence of lip, oral cavity, and pharyngeal cancers by subsite in 2012. CA Cancer J Clin (2017) 67(1):51–64. doi: 10.3322/caac.21384
2. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin (2015) 65(2):87–108. doi: 10.3322/caac.21262
3. Mehanna H, Jones TM, Gregoire V, Ang KK. Oropharyngeal carcinoma related to human papillomavirus. BMJ (2010) 340:c1439. doi: 10.1136/bmj.c1439
4. Brown LM, Check DP, Devesa SS. Oropharyngeal cancer incidence trends: diminishing racial disparities. Cancer Causes Control (2011) 22(5):753–63. doi: 10.1007/s10552-011-9748-1
5. van Monsjou HS, van Velthuysen ML, van den Brekel MW, Jordanova ES, Melief CJ, Balm AJ. Human papillomavirus status in young patients with head and neck squamous cell carcinoma. Int J Cancer (2012) 130(8):1806–12. doi: 10.1002/ijc.26195
6. Welters MJP, Ma W, Santegoets S, Goedemans R, Ehsan I, Jordanova ES, et al. Intratumoral HPV16-Specific T Cells Constitute a Type I-Oriented Tumor Microenvironment to Improve Survival in HPV16-Driven Oropharyngeal Cancer. Clin Cancer Res (2018) 24(3):634–47. doi: 10.1158/1078-0432.CCR-17-2140
7. Wang HF, Wang SS, Tang YJ, Chen Y, Zheng M, Tang YL, et al. The Double-Edged Sword-How Human Papillomaviruses Interact With Immunity in Head and Neck Cancer. Front Immunol (2019) 10:653. doi: 10.3389/fimmu.2019.00653
8. Syrjanen S, Syrjanen K. HPV in Head and Neck Carcinomas: Different HPV Profiles in Oropharyngeal Carcinomas - Why? Acta Cytol (2019) 63(2):124–42. doi: 10.1159/000495727
9. Doorbar J, Quint W, Banks L, Bravo IG, Stoler M, Broker TR, et al. The biology and life-cycle of human papillomaviruses. Vaccine (2012) 30 Suppl 5:F55–70. doi: 10.1016/j.vaccine.2012.06.083
10. Stanley M. Immunobiology of HPV and HPV vaccines. Gynecol Oncol (2008) 109(2 Suppl):S15–21. doi: 10.1016/j.ygyno.2008.02.003
11. Gupta S, Kumar P, Das BC. HPV: Molecular pathways and targets. Curr Probl Cancer (2018) 42(2):161–74. doi: 10.1016/j.currproblcancer.2018.03.003
12. zur Hausen H. Papillomaviruses and cancer: from basic studies to clinical application. Nat Rev Cancer (2002) 2(5):342–50. doi: 10.1038/nrc798
13. Ang KK, Harris J, Wheeler R, Weber R, Rosenthal DI, Nguyen-Tan PF, et al. Human papillomavirus and survival of patients with oropharyngeal cancer. N Engl J Med (2010) 363(1):24–35. doi: 10.1056/NEJMoa0912217
14. Wansom D, Light E, Worden F, Prince M, Urba S, Chepeha DB, et al. Correlation of cellular immunity with human papillomavirus 16 status and outcome in patients with advanced oropharyngeal cancer. Arch Otolaryngol Head Neck Surg (2010) 136(12):1267–73. doi: 10.1001/archoto.2010.211
15. Gillison ML, Trotti AM, Harris J, Eisbruch A, Harari PM, Adelstein DJ, et al. Radiotherapy plus cetuximab or cisplatin in human papillomavirus-positive oropharyngeal cancer (NRG Oncology RTOG 1016): a randomised, multicentre, non-inferiority trial. Lancet (2019) 393(10166):40–50. doi: 10.1016/S0140-6736(18)32779-X
16. Patel RR, Ludmir EB, Augustyn A, Zaorsky NG, Lehrer EJ, Ryali R, et al. De-intensification of therapy in human papillomavirus associated oropharyngeal cancer: A systematic review of prospective trials. Oral Oncol (2020) 103:104608. doi: 10.1016/j.oraloncology.2020.104608
17. Ventz S, Trippa L, Schoenfeld JD. Lessons Learned from Deescalation Trials in Favorable Risk HPV-Associated Squamous Cell Head and Neck Cancer-A Perspective on Future Trial Designs. Clin Cancer Res (2019) 25(24):7281–6. doi: 10.1158/1078-0432.CCR-19-0945
18. Karim R, Tummers B, Meyers C, Biryukov JL, Alam S, Backendorf C, et al. Human papillomavirus (HPV) upregulates the cellular deubiquitinase UCHL1 to suppress the keratinocyte’s innate immune response. PloS Pathog (2013) 9(5):e1003384. doi: 10.1371/journal.ppat.1003384
19. Ma W, Melief CJ, van der Burg SH. Control of immune escaped human papilloma virus is regained after therapeutic vaccination. Curr Opin Virol (2017) 23:16–22. doi: 10.1016/j.coviro.2017.02.005
20. Tummers B, et Goedemans R, Pelascini LP, Jordanova ES, van Esch EM, Meyers C, et al. The interferon-related developmental regulator 1 is used by human papillomavirus to suppress NFkappaB activation. Nat Commun (2015) 6:6537. doi: 10.1038/ncomms7537
21. Steinbach A, Riemer AB. Immune evasion mechanisms of human papillomavirus: An update. Int J Cancer (2018) 142(2):224–9. doi: 10.1002/ijc.31027
22. Zhou C, Tuong ZK, Frazer IH. Papillomavirus Immune Evasion Strategies Target the Infected Cell and the Local Immune System. Front Oncol (2019) 9:682. doi: 10.3389/fonc.2019.00682
23. Matlung SE, Wilhelmina van Kempen PM, Bovenschen N, van Baarle D, Willems SM. Differences in T-cell infiltrates and survival between HPV+ and HPV- oropharyngeal squamous cell carcinoma. Future Sci OA (2016) 2(1):FSO88. doi: 10.4155/fso.15.88
24. Santegoets SJ, van Ham VJ, Ehsan I, Charoentong P, Duurland CL, van Unen V, et al. The Anatomical Location Shapes the Immune Infiltrate in Tumors of Same Etiology and Affects Survival. Clin Cancer Res (2019) 25(1):240–52. doi: 10.1158/1078-0432.CCR-18-1749
25. Fridman WH, Galon J, Dieu-Nosjean MC, Cremer I, Fisson S, Damotte D, et al. Immune infiltration in human cancer: prognostic significance and disease control. Curr Top Microbiol Immunol (2011) 344:1–24. doi: 10.1007/82_2010_46
26. Wallis SP, Stafford ND, Greenman J. Clinical relevance of immune parameters in the tumor microenvironment of head and neck cancers. Head Neck (2015) 37(3):449–59. doi: 10.1002/hed.23736
27. Galon J, Bruni D. Approaches to treat immune hot, altered and cold tumours with combination immunotherapies. Nat Rev Drug Discovery (2019) 18(3):197–218. doi: 10.1038/s41573-018-0007-y
28. Van Abel KM, Routman DM, Moore EJ, Ma DJ, Yin LX, Fields PA, et al. T cell fraction impacts oncologic outcomes in human papillomavirus associated oropharyngeal squamous cell carcinoma. Oral Oncol (2020) 111:104894. doi: 10.1016/j.oraloncology.2020.104894
29. Gillison ML, Blumenschein G Jr., Fayette J, Guigay J, Colevas AD, Licitra L, et al. CheckMate 141: 1-Year Update and Subgroup Analysis of Nivolumab as First-Line Therapy in Patients with Recurrent/Metastatic Head and Neck Cancer. Oncologist (2018) 23(9):1079–82. doi: 10.1634/theoncologist.2017-0674
30. Giraldo NA, Sanchez-Salas R, Peske JD, Vano Y, Becht E, Petitprez F, et al. The clinical role of the TME in solid cancer. Br J Cancer (2019) 120(1):45–53. doi: 10.1038/s41416-018-0327-z
31. Jung AC, Guihard S, Krugell S, Ledrappier S, Brochot A, Dalstein V, et al. CD8-alpha T-cell infiltration in human papillomavirus-related oropharyngeal carcinoma correlates with improved patient prognosis. Int J Cancer (2013) 132(2):E26–36. doi: 10.1002/ijc.27776
32. Ward MJ, Thirdborough SM, Mellows T, Riley C, Harris S, Suchak K, et al. Tumour-infiltrating lymphocytes predict for outcome in HPV-positive oropharyngeal cancer. Br J Cancer (2014) 110(2):489–500. doi: 10.1038/bjc.2013.639
33. Gurin D, Slavik M, Hermanova M, Selingerova I, Kazda T, Hendrych M, et al. The tumor immune microenvironment and its implications for clinical outcome in patients with oropharyngeal squamous cell carcinoma. J Oral Pathol Med (2020) 49(9):886–96. doi: 10.1111/jop.13055
34. Wansom D, Light E, Thomas D, Worden F, Prince M, Urba S, et al. Infiltrating lymphocytes and human papillomavirus-16–associated oropharyngeal cancer. Laryngoscope (2012) 122(1):121–7. doi: 10.1002/lary.22133
35. Lee YS, Park JY, Cho KJ, Kim SB, Lee SW, Choi SH, et al. Composition of inflammatory cells regulating the response to concurrent chemoradiation therapy for HPV (+) tonsil cancer. Oral Oncol (2015) 51(12):1113–9. doi: 10.1016/j.oraloncology.2015.10.001
36. Poropatich K, Hernandez D, Fontanarosa J, Brown K, Woloschak G, Paintal A, et al. Peritumoral cuffing by T-cell tumor-infiltrating lymphocytes distinguishes HPV-related oropharyngeal squamous cell carcinoma from oral cavity squamous cell carcinoma. J Oral Pathol Med (2017) 46(10):972–8. doi: 10.1111/jop.12605
37. Nordfors C, Grun N, Tertipis N, Ahrlund-Richter A, Haeggblom L, Sivars L, et al. CD8+ and CD4+ tumour infiltrating lymphocytes in relation to human papillomavirus status and clinical outcome in tonsillar and base of tongue squamous cell carcinoma. Eur J Cancer (2013) 49(11):2522–30. doi: 10.1016/j.ejca.2013.03.019
38. Punt S, Dronkers EA, Welters MJ, Goedemans R, Koljenovic S, Bloemena E, et al. A beneficial tumor microenvironment in oropharyngeal squamous cell carcinoma is characterized by a high T cell and low IL-17(+) cell frequency. Cancer Immunol Immunother (2016) 65(4):393–403. doi: 10.1007/s00262-016-1805-x
39. Lin CM, Lin LW, Chen YW, Ye YL. The expression and prognostic impact of proinflammatory cytokines and their associations with carcinogens in oropharyngeal squamous cell carcinoma. Cancer Immunol Immunother (2020) 69(4):549–58. doi: 10.1007/s00262-020-02488-w
40. Partlova S, Boucek J, Kloudova K, Lukesova E, Zabrodsky M, Grega M, et al. Distinct patterns of intratumoral immune cell infiltrates in patients with HPV-associated compared to non-virally induced head and neck squamous cell carcinoma. Oncoimmunology (2015) 4(1):e965570. doi: 10.4161/21624011.2014.965570
41. Solomon B, Young RJ, Bressel M, Cernelc J, Savas P, Liu H, et al. Identification of an excellent prognosis subset of human papillomavirus-associated oropharyngeal cancer patients by quantification of intratumoral CD103(+) immune cell abundance. Ann Oncol (2019) 30(10):1638–46. doi: 10.1093/annonc/mdz271
42. Hewavisenti R, Ferguson A, Wang K, Jones D, Gebhardt T, Edwards J, et al. CD103+ tumor-resident CD8+ T cell numbers underlie improved patient survival in oropharyngeal squamous cell carcinoma. J Immunother Cancer (2020) 8(1):e000452. doi: 10.1136/jitc-2019-000452
43. Mueller SN, Mackay LK. Tissue-resident memory T cells: local specialists in immune defence. Nat Rev Immunol (2016) 16(2):79–89. doi: 10.1038/nri.2015.3
44. Savas P, Virassamy B, Ye C, Salim A, Mintoff CP, Caramia F, et al. Single-cell profiling of breast cancer T cells reveals a tissue-resident memory subset associated with improved prognosis. Nat Med (2018) 24(7):986–93. doi: 10.1038/s41591-018-0078-7
45. Kortekaas KE, Santegoets SJ, Sturm G, Ehsan I, van Egmond SL, Finotello F, et al. CD39 identifies the CD4+ tumor-specific T cell population in human cancer. Cancer Immunol Res (2020) 8(10):1311–21. doi: 10.1158/2326-6066.CIR-20-0270
46. Cioni B, Jordanova ES, Hooijberg E, van der Linden R, de Menezes RX, Tan K, et al. HLA class II expression on tumor cells and low numbers of tumor-associated macrophages predict clinical outcome in oropharyngeal cancer. Head Neck (2019) 41(2):463–78. doi: 10.1002/hed.25442
47. Ou D, Adam J, Garberis I, Blanchard P, Nguyen F, Levy A, et al. Influence of tumor-associated macrophages and HLA class I expression according to HPV status in head and neck cancer patients receiving chemo/bioradiotherapy. Radiother Oncol (2019) 130:89–96. doi: 10.1016/j.radonc.2018.08.013
48. Reeves E, Wood O, Ottensmeier CH, King EV, Thomas GJ, Elliott T, et al. HPV Epitope Processing Differences Correlate with ERAP1 Allotype and Extent of CD8(+) T-cell Tumor Infiltration in OPSCC. Cancer Immunol Res (2019) 7(7):1202–13. doi: 10.1158/2326-6066.CIR-18-0498
49. Näsman A, Andersson E, Marklund L, Tertipis N, Hammarstedt-Nordenvall L, Attner P, et al. HLA class I and II expression in oropharyngeal squamous cell carcinoma in relation to tumor HPV status and clinical outcome. PloS One (2013) 8(10):e77025. doi: 10.1371/journal.pone.0077025
50. Näsman A, Andersson E, Nordfors C, Grun N, Johansson H, Munck-Wikland E, et al. MHC class I expression in HPV positive and negative tonsillar squamous cell carcinoma in correlation to clinical outcome. Int J Cancer (2013) 132(1):72–81. doi: 10.1002/ijc.27635
51. O’Higgins C, Ward FJ, Abu Eid R. Deciphering the Role of Regulatory CD4 T Cells in Oral and Oropharyngeal Cancer: A Systematic Review. Front Oncol (2018) 8:442. doi: 10.3389/fonc.2018.00442
52. Drennan S, Stafford ND, Greenman J, Green VL. Increased frequency and suppressive activity of CD127(low/-) regulatory T cells in the peripheral circulation of patients with head and neck squamous cell carcinoma are associated with advanced stage and nodal involvement. Immunology (2013) 140(3):335–43. doi: 10.1111/imm.12144
53. Sun W, Li WJ, Wu CY, Zhong H, Wen WP. CD45RA-Foxp3high but not CD45RA+Foxp3low suppressive T regulatory cells increased in the peripheral circulation of patients with head and neck squamous cell carcinoma and correlated with tumor progression. J Exp Clin Cancer Res (2014) 33:35. doi: 10.1186/1756-9966-33-35
54. Distel LV, Fickenscher R, Dietel K, Hung A, Iro H, Zenk J, et al. Tumour infiltrating lymphocytes in squamous cell carcinoma of the oro- and hypopharynx: prognostic impact may depend on type of treatment and stage of disease. Oral Oncol (2009) 45(10):e167–74. doi: 10.1016/j.oraloncology.2009.05.640
55. Bron L, Jandus C, Andrejevic-Blant S, Speiser DE, Monnier P, Romero P, et al. Prognostic value of arginase-II expression and regulatory T-cell infiltration in head and neck squamous cell carcinoma. Int J Cancer (2013) 132(3):E85–93. doi: 10.1002/ijc.27728
56. Lukesova E, Boucek J, Rotnaglova E, Salakova M, Koslabova E, Grega M, et al. High level of Tregs is a positive prognostic marker in patients with HPV-positive oral and oropharyngeal squamous cell carcinomas. BioMed Res Int (2014) 2014:303929. doi: 10.1155/2014/303929
57. deLeeuw RJ, Kost SE, Kakal JA, Nelson BH. The prognostic value of FoxP3+ tumor-infiltrating lymphocytes in cancer: a critical review of the literature. Clin Cancer (2012) 18(11):3022–9. doi: 10.1158/1078-0432.CCR-11-3216
58. Salama P, Phillips M, Grieu F, Morris M, Zeps N, Joseph D, et al. Tumor-infiltrating FOXP3+ T regulatory cells show strong prognostic significance in colorectal cancer. J Clin Oncol (2009) 27(2):186–92. doi: 10.1200/JCO.2008.18.7229
59. Miyara M, Yoshioka Y, Kitoh A, Shima T, Wing K, Niwa A, et al. Functional delineation and differentiation dynamics of human CD4+ T cells expressing the FoxP3 transcription factor. Immunity (2009) 30(6):899–911. doi: 10.1016/j.immuni.2009.03.019
60. Saito T, Nishikawa H, Wada H, Nagano Y, Sugiyama D, Atarashi K, et al. Two FOXP3(+)CD4(+) T cell subpopulations distinctly control the prognosis of colorectal cancers. Nat Med (2016) 22(6):679–84. doi: 10.1038/nm.4086
61. Sugiyama D, Nishikawa H, Maeda Y, Nishioka M, Tanemura A, Katayama I, et al. Anti-CCR4 mAb selectively depletes effector-type FoxP3+CD4+ regulatory T cells, evoking antitumor immune responses in humans. Proc Natl Acad Sci USA (2013) 110(44):17945–50. doi: 10.1073/pnas.1316796110
62. Badoual C, Hans S, Merillon N, Van Ryswick C, Ravel P, Benhamouda N, et al. PD-1-expressing tumor-infiltrating T cells are a favorable prognostic biomarker in HPV-associated head and neck cancer. Cancer Res (2013) 73(1):128–38. doi: 10.1158/0008-5472.CAN-12-2606
63. Chaudhry A, Rudra D, Treuting P, Samstein RM, Liang Y, Kas A, et al. CD4+ regulatory T cells control TH17 responses in a Stat3-dependent manner. Science (2009) 326(5955):986–91. doi: 10.1126/science.1172702
64. Koch MA, Tucker-Heard G, Perdue NR, Killebrew JR, Urdahl KB, Campbell DJ. The transcription factor T-bet controls regulatory T cell homeostasis and function during type 1 inflammation. Nat Immunol (2009) 10(6):595–602. doi: 10.1038/ni.1731
65. Wang Y, Su MA, Wan YY. An essential role of the transcription factor GATA-3 for the function of regulatory T cells. Immunity (2011) 35(3):337–48. doi: 10.1016/j.immuni.2011.08.012
66. Zheng Y, Chaudhry A, Kas A, deRoos P, Kim JM, Chu TT, et al. Regulatory T-cell suppressor program co-opts transcription factor IRF4 to control T(H)2 responses. Nature (2009) 458(7236):351–6. doi: 10.1038/nature07674
67. Santegoets SJ, Duurland CL, Jordanova ES, van Ham JJ, Ehsan I, van Egmond SL, et al. Tbet-positive regulatory T cells accumulate in oropharyngeal cancers with ongoing tumor-specific type 1 T cell responses. J Immunother Cancer (2019) 7(1):14. doi: 10.1186/s40425-019-0497-0
68. Mantovani A, Marchesi F, Malesci A, Laghi L, Allavena P. Tumour-associated macrophages as treatment targets in oncology. Nat Rev Clin Oncol (2017) 14(7):399–416. doi: 10.1038/nrclinonc.2016.217
69. Nakamura K, Smyth MJ. Myeloid immunosuppression and immune checkpoints in the tumor microenvironment. Cell Mol Immunol (2020) 17(1):1–12. doi: 10.1038/s41423-019-0306-1
70. Locati M, Curtale G, Mantovani A. Diversity, Mechanisms, and Significance of Macrophage Plasticity. Annu Rev Pathol (2020) 15:123–47. doi: 10.1146/annurev-pathmechdis-012418-012718
71. Biswas SK, Mantovani A. Macrophage plasticity and interaction with lymphocyte subsets: cancer as a paradigm. Nat Immunol (2010) 11(10):889–96. doi: 10.1038/ni.1937
72. Anfray C, Ummarino A, Andon FT, Allavena P. Current Strategies to Target Tumor-Associated-Macrophages to Improve Anti-Tumor Immune Responses. Cells (2019) 9(1):46. doi: 10.3390/cells9010046
73. Heusinkveld M, de Vos van Steenwijk PJ, Goedemans R, Ramwadhdoebe TH, Gorter A, Welters MJ, et al. M2 macrophages induced by prostaglandin E2 and IL-6 from cervical carcinoma are switched to activated M1 macrophages by CD4+ Th1 cells. J Immunol (2011) 187(3):1157–65. doi: 10.4049/jimmunol.1100889
74. Santegoets SJ, Duurland CL, Jordanova EJ, van Ham VJ, Ehsan I, Loof NM, et al. CD163(+) cytokine-producing cDC2 stimulate intratumoral type 1 T cell responses in HPV16-induced oropharyngeal cancer. J Immunother Cancer (2020) 8(2):e001053. doi: 10.1136/jitc-2020-001053
75. Balermpas P, Rodel F, Liberz R, Oppermann J, Wagenblast J, Ghanaati S, et al. Head and neck cancer relapse after chemoradiotherapy correlates with CD163+ macrophages in primary tumour and CD11b+ myeloid cells in recurrences. Br J Cancer (2014) 111(8):1509–18. doi: 10.1038/bjc.2014.446
76. de Vos van Steenwijk PJ, Ramwadhdoebe TH, Goedemans R, Doorduijn EM, van Ham JJ, Gorter A, et al. Tumor-infiltrating CD14-positive myeloid cells and CD8-positive T-cells prolong survival in patients with cervical carcinoma. Int J Cancer (2013) 133(12):2884–94. doi: 10.1002/ijc.28309
77. Heusinkveld M, van der Burg SH. Identification and manipulation of tumor associated macrophages in human cancers. J Transl Med (2011) 9:216. doi: 10.1186/1479-5876-9-216
78. Etzerodt A, Tsalkitzi K, Maniecki M, Damsky W, Delfini M, Baudoin E, et al. Specific targeting of CD163(+) TAMs mobilizes inflammatory monocytes and promotes T cell-mediated tumor regression. J Exp Med (2019) 216(10):2394–411. doi: 10.1084/jem.20182124
79. van der Sluis TC, Sluijter M, van Duikeren S, West BL, Melief CJ, Arens R, et al. Therapeutic Peptide Vaccine-Induced CD8 T Cells Strongly Modulate Intratumoral Macrophages Required for Tumor Regression. Cancer Immunol Res (2015) 3(9):1042–51. doi: 10.1158/2326-6066.CIR-15-0052
80. Jahchan NS, Mujal AM, Pollack JL, Binnewies M, Sriram V, Reyno L, et al. Tuning the Tumor Myeloid Microenvironment to Fight Cancer. Front Immunol (2019) 10:1611. doi: 10.3389/fimmu.2019.01611
81. Mandruzzato S, Brandau S, Britten CM, Bronte V, Damuzzo V, Gouttefangeas C, et al. Toward harmonized phenotyping of human myeloid-derived suppressor cells by flow cytometry: results from an interim study. Cancer Immunol Immunother (2016) 65(2):161–9. doi: 10.1007/s00262-015-1782-5
82. Cassetta L, Baekkevold ES, Brandau S, Bujko A, Cassatella MA, Dorhoi A, et al. Deciphering myeloid-derived suppressor cells: isolation and markers in humans, mice and non-human primates. Cancer Immunol Immunother (2019) 68(4):687–97. doi: 10.1007/s00262-019-02302-2
83. Lang S, Bruderek K, Kaspar C, Hoing B, Kanaan O, Dominas N, et al. Clinical Relevance and Suppressive Capacity of Human Myeloid-Derived Suppressor Cell Subsets. Clin Cancer Res (2018) 24(19):4834–44. doi: 10.1158/1078-0432.CCR-17-3726
84. Califano JA, Khan Z, Noonan KA, Rudraraju L, Zhang Z, Wang H, et al. Tadalafil augments tumor specific immunity in patients with head and neck squamous cell carcinoma. Clin Cancer Res (2015) 21(1):30–8. doi: 10.1158/1078-0432.CCR-14-1716
85. Davis RJ, Van Waes C, Allen CT. Overcoming barriers to effective immunotherapy: MDSCs, TAMs, and Tregs as mediators of the immunosuppressive microenvironment in head and neck cancer. Oral Oncol (2016) 58:59–70. doi: 10.1016/j.oraloncology.2016.05.002
86. Dutertre CA, Becht E, Irac SE, Khalilnezhad A, Narang V, Khalilnezhad S, et al. Single-Cell Analysis of Human Mononuclear Phagocytes Reveals Subset-Defining Markers and Identifies Circulating Inflammatory Dendritic Cells. Immunity (2019) 51(3):573–89.e578. doi: 10.1016/j.immuni.2019.08.008
87. Villani AC, Satija R, Reynolds G, Sarkizova S, Shekhar K, Fletcher J, et al. Single-cell RNA-seq reveals new types of human blood dendritic cells, monocytes, and progenitors. Science (2017) 356(6335):eaah4573. doi: 10.1126/science.aah4573
88. Bourdely P, Anselmi G, Vaivode K, Ramos RN, Missolo-Koussou Y, Hidalgo S, et al. Transcriptional and Functional Analysis of CD1c(+) Human Dendritic Cells Identifies a CD163(+) Subset Priming CD8(+)CD103(+) T Cells. Immunity (2020) 53(2):335–52.e8. doi: 10.1016/j.immuni.2020.06.002
89. De Meulenaere A, Vermassen T, Aspeslagh S, Deron P, Duprez F, Laukens D, et al. Tumor PD-L1 status and CD8(+) tumor-infiltrating T cells: markers of improved prognosis in oropharyngeal cancer. Oncotarget (2017) 8(46):80443–52. doi: 10.18632/oncotarget.19045
90. Fukushima Y, Someya M, Nakata K, Hori M, Kitagawa M, Hasegawa T, et al. Influence of PD-L1 expression in immune cells on the response to radiation therapy in patients with oropharyngeal squamous cell carcinoma. Radiother Oncol (2018) 129(2):409–14. doi: 10.1016/j.radonc.2018.08.023
91. Lecerf C, Kamal M, Vacher S, Chemlali W, Schnitzler A, Morel C, et al. Immune gene expression in head and neck squamous cell carcinoma patients. Eur J Cancer (2019) 121:210–23. doi: 10.1016/j.ejca.2019.08.028
92. Sato F, Ono T, Kawahara A, Kawaguchi T, Tanaka H, Shimamatsu K, et al. Prognostic impact of p16 and PD-L1 expression in patients with oropharyngeal squamous cell carcinoma receiving a definitive treatment. J Clin Pathol (2019) 72(8):542–9. doi: 10.1136/jclinpath-2019-205818
93. Young RJ, Bressel M, Porceddu S, Cernelc J, Savas P, Liu H, et al. Validation and characterisation of prognostically significant PD-L1(+) immune cells in HPV+ oropharyngeal squamous cell carcinoma. Oral Oncol (2020) 101:104516. doi: 10.1016/j.oraloncology.2019.104516
94. Kim MH, Kim JH, Lee JM, Choi JW, Jung D, Cho H, et al. Molecular subtypes of oropharyngeal cancer show distinct immune microenvironment related with immune checkpoint blockade response. Br J Cancer (2020) 122(11):1649–60. doi: 10.1038/s41416-020-0796-8
95. Oguejiofor K, Galletta-Williams H, Dovedi SJ, Roberts DL, Stern PL, West CM. Distinct patterns of infiltrating CD8+ T cells in HPV+ and CD68 macrophages in HPV- oropharyngeal squamous cell carcinomas are associated with better clinical outcome but PD-L1 expression is not prognostic. Oncotarget (2017) 8(9):14416–27. doi: 10.18632/oncotarget.14796
96. Andre P, Denis C, Soulas C, Bourbon-Caillet C, Lopez J, Arnoux T, et al. Anti-NKG2A mAb Is a Checkpoint Inhibitor that Promotes Anti-tumor Immunity by Unleashing Both T and NK Cells. Cell (2018) 175(7):1731–43.e1713. doi: 10.1016/j.cell.2018.10.014
97. van Montfoort N, Borst L, Korrer MJ, Sluijter M, Marijt KA, Santegoets SJ, et al. NKG2A Blockade Potentiates CD8 T Cell Immunity Induced by Cancer Vaccines. Cell (2018) 175(7):1744–55.e1715. doi: 10.1016/j.cell.2018.10.028
98. Gooden M, Lampen M, Jordanova ES, Leffers N, Trimbos JB, van der Burg SH, et al. HLA-E expression by gynecological cancers restrains tumor-infiltrating CD8(+) T lymphocytes. Proc Natl Acad Sci USA (2011) 108(26):10656–61. doi: 10.1073/pnas.1100354108
99. Talebian Yazdi M, van Riet S, van Schadewijk A, Fiocco M, van Hall T, Taube C, et al. The positive prognostic effect of stromal CD8+ tumor-infiltrating T cells is restrained by the expression of HLA-E in non-small cell lung carcinoma. Oncotarget (2016) 7(3):3477–88. doi: 10.18632/oncotarget.6506
100. van Hall T, Andre P, Horowitz A, Ruan DF, Borst L, Zerbib R, et al. Monalizumab: inhibiting the novel immune checkpoint NKG2A. J Immunother Cancer (2019) 7(1):263. doi: 10.1186/s40425-019-0761-3
101. Gameiro SF, Ghasemi F, Barrett JW, Koropatnick J, Nichols AC, Mymryk JS, et al. Treatment-naive HPV+ head and neck cancers display a T-cell-inflamed phenotype distinct from their HPV- counterparts that has implications for immunotherapy. Oncoimmunology (2018) 7(10):e1498439. doi: 10.1080/2162402X.2018.1498439
102. Huang SH, Waldron JN, Milosevic M, Shen X, Ringash J, Su J, et al. Prognostic value of pretreatment circulating neutrophils, monocytes, and lymphocytes in oropharyngeal cancer stratified by human papillomavirus status. Cancer (2015) 121(4):545–55. doi: 10.1002/cncr.29100
103. Panje C, Riesterer O, Glanzmann C, Studer G. Neutrophil-lymphocyte ratio complements volumetric staging as prognostic factor in patients treated with definitive radiotherapy for oropharyngeal cancer. BMC Cancer (2017) 17(1):643. doi: 10.1186/s12885-017-3590-0
104. So YK, Lee G, Oh D, Byeon S, Park W, Chung MK. Prognostic Role of Neutrophil-to-Lymphocyte Ratio in Patients with Human Papillomavirus-Positive Oropharyngeal Cancer. Otolaryngol Head Neck Surg (2018) 159(2):303–9. doi: 10.1177/0194599818764651
105. Sumner WA, Stokes WA, Oweida A, Berggren KL, McDermott JD, Raben D, et al. Survival impact of pre-treatment neutrophils on oropharyngeal and laryngeal cancer patients undergoing definitive radiotherapy. J Transl Med (2017) 15(1):168. doi: 10.1186/s12967-017-1268-7
106. Trellakis S, Farjah H, Bruderek K, Dumitru CA, Hoffmann TK, Lang S, et al. Peripheral blood neutrophil granulocytes from patients with head and neck squamous cell carcinoma functionally differ from their counterparts in healthy donors. Int J Immunopathol Pharmacol (2011) 24(3):683–93. doi: 10.1177/039463201102400314
107. Takahashi H, Sakakura K, Tada H, Kaira K, Oyama T, Chikamatsu K. Prognostic significance and population dynamics of peripheral monocytes in patients with oropharyngeal squamous cell carcinoma. Head Neck (2019) 41(6):1880–8. doi: 10.1002/hed.25625
108. Santegoets S, de Groot AF, Dijkgraaf EM, Simoes AMC, van der Noord VE, van Ham JJ, et al. The blood mMDSC to DC ratio is a sensitive and easy to assess independent predictive factor for epithelial ovarian cancer survival. Oncoimmunology (2018) 7(8):e1465166. doi: 10.1080/2162402X.2018.1465166
109. Kano S, Homma A, Hatakeyama H, Mizumachi T, Sakashita T, Kakizaki T, et al. Pretreatment lymphocyte-to-monocyte ratio as an independent prognostic factor for head and neck cancer. Head Neck (2017) 39(2):247–53. doi: 10.1002/hed.24576
110. Heusinkveld M, Goedemans R, Briet RJ, Gelderblom H, Nortier JW, Gorter A, et al. Systemic and local human papillomavirus 16-specific T-cell immunity in patients with head and neck cancer. Int J Cancer (2012) 131(2):E74–85. doi: 10.1002/ijc.26497
111. Masterson L, Lechner M, Loewenbein S, Mohammed H, Davies-Husband C, Fenton T, et al. CD8(+) T cell response to human papillomavirus 16 E7 is able to predict survival outcome in oropharyngeal cancer. Eur J Cancer (2016) 67:141–51. doi: 10.1016/j.ejca.2016.08.012
112. Bhatt KH, Neller MA, Srihari S, Crooks P, Lekieffre L, Aftab BT, et al. Profiling HPV-16-specific T cell responses reveals broad antigen reactivities in oropharyngeal cancer patients. J Exp Med (2020) 217(10):e20200389. doi: 10.1084/jem.20200389
113. de Jong A, van der Burg SH, Kwappenberg KM, van der Hulst JM, Franken KL, Geluk A, et al. Frequent detection of human papillomavirus 16 E2-specific T-helper immunity in healthy subjects. Cancer Res (2002) 62(2):472–9.
114. de Jong A, van Poelgeest MI, van der Hulst JM, Drijfhout JW, Fleuren GJ, Melief CJ, et al. Human papillomavirus type 16-positive cervical cancer is associated with impaired CD4+ T-cell immunity against early antigens E2 and E6. Cancer Res (2004) 64(15):5449–55. doi: 10.1158/0008-5472.CAN-04-0831
115. Welters MJ, de Jong A, van den Eeden SJ, van der Hulst JM, Kwappenberg KM, Hassane S, et al. Frequent display of human papillomavirus type 16 E6-specific memory t-Helper cells in the healthy population as witness of previous viral encounter. Cancer Res (2003) 63(3):636–41.
116. Angell H, Galon J. From the immune contexture to the Immunoscore: the role of prognostic and predictive immune markers in cancer. Curr Opin Immunol (2013) 25(2):261–7. doi: 10.1016/j.coi.2013.03.004
117. Licitra L, Perrone F, Bossi P, Suardi S, Mariani L, Artusi R, et al. High-risk human papillomavirus affects prognosis in patients with surgically treated oropharyngeal squamous cell carcinoma. J Clin Oncol (2006) 24(36):5630–6. doi: 10.1200/JCO.2005.04.6136
118. Thurlow JK, Pena Murillo CL, Hunter KD, Buffa FM, Patiar S, Betts G, et al. Spectral clustering of microarray data elucidates the roles of microenvironment remodeling and immune responses in survival of head and neck squamous cell carcinoma. J Clin Oncol (2010) 28(17):2881–8. doi: 10.1200/JCO.2009.24.8724
119. Al-Taei S, Banner R, Powell N, Evans M, Palaniappan N, Tabi Z, et al. Decreased HPV-specific T cell responses and accumulation of immunosuppressive influences in oropharyngeal cancer patients following radical therapy. Cancer Immunol Immunother (2013) 62(12):1821–30. doi: 10.1007/s00262-013-1488-5
120. Hladikova K, Partlova S, Koucky V, Boucek J, Fonteneau JF, Zabrodsky M, et al. Dysfunction of HPV16-specific CD8+ T cells derived from oropharyngeal tumors is related to the expression of Tim-3 but not PD-1. Oral Oncol (2018) 82:75–82. doi: 10.1016/j.oraloncology.2018.05.010
121. Parikh F, Duluc D, Imai N, Clark A, Misiukiewicz K, Bonomi M, et al. Chemoradiotherapy-induced upregulation of PD-1 antagonizes immunity to HPV-related oropharyngeal cancer. Cancer Res (2014) 74(24):7205–16. doi: 10.1158/0008-5472.CAN-14-1913
122. Ramos CA, Narala N, Vyas GM, Leen AM, Gerdemann U, Sturgis EM, et al. Human papillomavirus type 16 E6/E7-specific cytotoxic T lymphocytes for adoptive immunotherapy of HPV-associated malignancies. J Immunother (2013) 36(1):66–76. doi: 10.1097/CJI.0b013e318279652e
123. Smith EM, Pawlita M, Rubenstein LM, Haugen TH, Hamsikova E, Turek LP. Risk factors and survival by HPV-16 E6 and E7 antibody status in human papillomavirus positive head and neck cancer. Int J Cancer (2010) 127(1):111–7. doi: 10.1002/ijc.25015
124. Nasman A, Romanitan M, Nordfors C, Grun N, Johansson H, Hammarstedt L, et al. Tumor infiltrating CD8+ and Foxp3+ lymphocytes correlate to clinical outcome and human papillomavirus (HPV) status in tonsillar cancer. PloS One (2012) 7(6):e38711. doi: 10.1371/journal.pone.0038711
125. Brown SD, Warren RL, Gibb EA, Martin SD, Spinelli JJ, Nelson BH, et al. Neo-antigens predicted by tumor genome meta-analysis correlate with increased patient survival. Genome Res (2014) 24(5):743–50. doi: 10.1101/gr.165985.113
126. Stransky N, Egloff AM, Tward AD, Kostic AD, Cibulskis K, Sivachenko A, et al. The mutational landscape of head and neck squamous cell carcinoma. Science (2011) 333(6046):1157–60. doi: 10.1126/science.1208130
127. Ren L, Leisegang M, Deng B, Matsuda T, Kiyotani K, Kato T, et al. Identification of neoantigen-specific T cells and their targets: implications for immunotherapy of head and neck squamous cell carcinoma. Oncoimmunology (2019) 8(4):e1568813. doi: 10.1080/2162402X.2019.1568813
128. Yang W, Lee KW, Srivastava RM, Kuo F, Krishna C, Chowell D, et al. Immunogenic neoantigens derived from gene fusions stimulate T cell responses. Nat Med (2019) 25(5):767–75. doi: 10.1038/s41591-019-0434-2
129. van den Bulk J, Verdegaal EME, Ruano D, Ijsselsteijn ME, Visser M, van der Breggen R, et al. Neoantigen-specific immunity in low mutation burden colorectal cancers of the consensus molecular subtype 4. Genome Med (2019) 11(1):87. doi: 10.1186/s13073-019-0697-8
130. De Stefani A, Forni G, Ragona R, Cavallo G, Bussi M, Usai A, et al. Improved survival with perilymphatic interleukin 2 in patients with resectable squamous cell carcinoma of the oral cavity and oropharynx. Cancer (2002) 95(1):90–7. doi: 10.1002/cncr.10654
131. van Poelgeest MI, Visconti VV, Aghai Z, van Ham VJ, Heusinkveld M, Zandvliet ML, et al. Potential use of lymph node-derived HPV-specific T cells for adoptive cell therapy of cervical cancer. Cancer Immunol Immunother (2016) 65(12):1451–63. doi: 10.1007/s00262-016-1892-8
132. Wolf GT, Ee WE Jr., Dolan RW, Moyer JS, Kaplan MJ, Spring PM, et al. Novel neoadjuvant immunotherapy regimen safety and survival in head and neck squamous cell cancer. Head Neck (2011) 33(12):1666–74. doi: 10.1002/hed.21660
133. Berinstein NL, Wolf GT, Naylor PH, Baltzer L, Egan JE, Brandwein HJ, et al. Increased lymphocyte infiltration in patients with head and neck cancer treated with the IRX-2 immunotherapy regimen. Cancer Immunol Immunother (2012) 61(6):771–82. doi: 10.1007/s00262-011-1134-z
134. Berinstein NL, McNamara M, Nguyen A, Egan J, Wolf GT. Increased immune infiltration and chemokine receptor expression in head and neck epithelial tumors after neoadjuvant immunotherapy with the IRX-2 regimen. Oncoimmunology (2018) 7(5):e1423173. doi: 10.1080/2162402X.2017.1423173
135. Lu H, Dietsch GN, Matthews MA, Yang Y, Ghanekar S, Inokuma M, et al. VTX-2337 is a novel TLR8 agonist that activates NK cells and augments ADCC. Clin Cancer Res (2012) 18(2):499–509. doi: 10.1158/1078-0432.CCR-11-1625
136. Ferris RL, Saba NF, Gitlitz BJ, Haddad R, Sukari A, Neupane P, et al. Effect of Adding Motolimod to Standard Combination Chemotherapy and Cetuximab Treatment of Patients With Squamous Cell Carcinoma of the Head and Neck: The Active8 Randomized Clinical Trial. JAMA Oncol (2018) 4(11):1583–8. doi: 10.1001/jamaoncol.2018.1888
137. Noto H, Goto A, Tsujimoto T, Noda M. Cancer risk in diabetic patients treated with metformin: a systematic review and meta-analysis. PloS One (2012) 7(3):e33411. doi: 10.1371/journal.pone.0033411
138. Eikawa S, Nishida M, Mizukami S, Yamazaki C, Nakayama E, Udono H. Immune-mediated antitumor effect by type 2 diabetes drug, metformin. Proc Natl Acad Sci U.S.A. (2015) 112(6):1809–14. doi: 10.1073/pnas.1417636112
139. Amin D, Richa T, Mollaee M, Zhan T, Tassone P, Johnson J, et al. Metformin effects on FOXP3(+) and CD8(+) T cell infiltrates of head and neck squamous cell carcinoma. Laryngoscope (2019) 130(9):E490–8. doi: 10.1002/lary.28336
140. Brahmer J, Reckamp KL, Baas P, Crino L, Eberhardt WE, Poddubskaya E, et al. Nivolumab versus Docetaxel in Advanced Squamous-Cell Non-Small-Cell Lung Cancer. N Engl J Med (2015) 373(2):123–35. doi: 10.1056/NEJMoa1504627
141. Larkin J, Chiarion-Sileni V, Gonzalez R, Grob JJ, Cowey CL, Lao CD, et al. Combined Nivolumab and Ipilimumab or Monotherapy in Untreated Melanoma. N Engl J Med (2015) 373(1):23–34. doi: 10.1056/NEJMoa1504030
142. Nghiem PT, Bhatia S, Lipson EJ, Kudchadkar RR, Miller NJ, Annamalai L, et al. PD-1 Blockade with Pembrolizumab in Advanced Merkel-Cell Carcinoma. N Engl J Med (2016) 374(26):2542–52. doi: 10.1056/NEJMoa1603702
143. Ferris RL, Blumenschein G Jr., Fayette J, Guigay J, Colevas AD, Licitra L, et al. Nivolumab for Recurrent Squamous-Cell Carcinoma of the Head and Neck. N Engl J Med (2016) 375(19):1856–67. doi: 10.1056/NEJMoa1602252
144. Seiwert TY, Burtness B, Mehra R, Weiss J, Berger R, Eder JP, et al. Safety and clinical activity of pembrolizumab for treatment of recurrent or metastatic squamous cell carcinoma of the head and neck (KEYNOTE-012): an open-label, multicentre, phase 1b trial. Lancet Oncol (2016) 17(7):956–65. doi: 10.1016/S1470-2045(16)30066-3
145. Segal NH, Ou SI, Balmanoukian A, Fury MG, Massarelli E, Brahmer JR, et al. Safety and efficacy of durvalumab in patients with head and neck squamous cell carcinoma: results from a phase I/II expansion cohort. Eur J Cancer (2019) 109:154–61. doi: 10.1016/j.ejca.2018.12.029
146. Massarelli E, William W, Johnson F, Kies M, Ferrarotto R, Guo M, et al. Combining Immune Checkpoint Blockade and Tumor-Specific Vaccine for Patients With Incurable Human Papillomavirus 16-Related Cancer: A Phase 2 Clinical Trial. JAMA Oncol (2019) 5(1):67–73. doi: 10.1001/jamaoncol.2018.4051
147. Powell SF, Gold KA, Gitau MM, Sumey CJ, Lohr MM, McGraw SC, et al. Safety and Efficacy of Pembrolizumab With Chemoradiotherapy in Locally Advanced Head and Neck Squamous Cell Carcinoma: A Phase IB Study. J Clin Oncol (2020) 38(21):2427–37. doi: 10.1200/JCO.19.03156
148. Ferris RL, Blumenschein G Jr., Fayette J, Guigay J, Colevas AD, Licitra L, et al. Nivolumab vs investigator’s choice in recurrent or metastatic squamous cell carcinoma of the head and neck: 2-year long-term survival update of CheckMate 141 with analyses by tumor PD-L1 expression. Oral Oncol (2018) 81:45–51. doi: 10.1016/j.oraloncology.2018.04.008
149. Haddad R, Concha-Benavente F, Blumenschein G Jr., Fayette J, Guigay J, Colevas AD, et al. Nivolumab treatment beyond RECIST-defined progression in recurrent or metastatic squamous cell carcinoma of the head and neck in CheckMate 141: A subgroup analysis of a randomized phase 3 clinical trial. Cancer (2019) 125(18):3208–18. doi: 10.1002/cncr.32190
150. Concha-Benavente F, Srivastava RM, Trivedi S, Lei Y, Chandran U, Seethala RR, et al. Identification of the Cell-Intrinsic and -Extrinsic Pathways Downstream of EGFR and IFNgamma That Induce PD-L1 Expression in Head and Neck Cancer. Cancer Res (2016) 76(5):1031–43. doi: 10.1158/0008-5472.CAN-15-2001
151. Jie HB, Srivastava RM, Argiris A, Bauman JE, Kane LP, Ferris RL. Increased PD-1(+) and TIM-3(+) TILs during Cetuximab Therapy Inversely Correlate with Response in Head and Neck Cancer Patients. Cancer Immunol Res (2017) 5(5):408–16. doi: 10.1158/2326-6066.CIR-16-0333
152. Ma W, Concha-Benavente F, Santegoets S, Welters MJP, Ehsan I, Ferris RL, et al. EGFR signaling suppresses type 1 cytokine-induced T-cell attracting chemokine secretion in head and neck cancer. PloS One (2018) 13(9):e0203402. doi: 10.1371/journal.pone.0203402
153. Trivedi S, Srivastava RM, Concha-Benavente F, Ferrone S, Garcia-Bates TM, Li J, et al. Anti-EGFR Targeted Monoclonal Antibody Isotype Influences Antitumor Cellular Immunity in Head and Neck Cancer Patients. Clin Cancer Res (2016) 22(21):5229–37. doi: 10.1158/1078-0432.CCR-15-2971
154. Ferris RL, Licitra L, Fayette J, Even C, Blumenschein G Jr., Harrington KJ, et al. Nivolumab in Patients with Recurrent or Metastatic Squamous Cell Carcinoma of the Head and Neck: Efficacy and Safety in CheckMate 141 by Prior Cetuximab Use. Clin Cancer Res (2019) 25(17):5221–30. doi: 10.1158/1078-0432.CCR-18-3944
155. Tumeh PC, Harview CL, Yearley JH, Shintaku IP, Taylor EJ, Robert L, et al. PD-1 blockade induces responses by inhibiting adaptive immune resistance. Nature (2014) 515(7528):568–71. doi: 10.1038/nature13954
156. June CH, O’Connor RS, Kawalekar OU, Ghassemi S, Milone MC. CAR T cell immunotherapy for human cancer. Science (2018) 359(6382):1361–5. doi: 10.1126/science.aar6711
157. Wang X, Riviere I. Manufacture of tumor- and virus-specific T lymphocytes for adoptive cell therapies. Cancer Gene Ther (2015) 22(2):85–94. doi: 10.1038/cgt.2014.81
158. Yang JC, Rosenberg SA. Adoptive T-Cell Therapy for Cancer. Adv Immunol (2016) 130:279–94. doi: 10.1016/bs.ai.2015.12.006
159. Draper LM, Kwong ML, Gros A, Stevanovic S, Tran E, Kerkar S, et al. Targeting of HPV-16+ Epithelial Cancer Cells by TCR Gene Engineered T Cells Directed against E6. Clin Cancer Res (2015) 21(19):4431–9. doi: 10.1158/1078-0432.CCR-14-3341
160. Mercier-Letondal P, Marton C, Deschamps M, Ferrand C, Vauchy C, Chenut C, et al. Isolation and Characterization of an HLA-DRB1*04-Restricted HPV16-E7 T Cell Receptor for Cancer Immunotherapy. Hum Gene Ther (2018) 29(10):1202–12. doi: 10.1089/hum.2018.091
161. Ali M, Foldvari Z, Giannakopoulou E, Boschen ML, Stronen E, Yang W, et al. Induction of neoantigen-reactive T cells from healthy donors. Nat Protoc (2019) 14(6):1926–43. doi: 10.1038/s41596-019-0170-6
162. van der Burg SH. Correlates of immune and clinical activity of novel cancer vaccines. Semin Immunol (2018) 39:119–36. doi: 10.1016/j.smim.2018.04.001
163. van der Burg SH, Arens R, Ossendorp F, van Hall T, Melief CJ. Vaccines for established cancer: overcoming the challenges posed by immune evasion. Nat Rev Cancer (2016) 16(4):219–33. doi: 10.1038/nrc.2016.16
164. Voskens CJ, Sewell D, Hertzano R, DeSanto J, Rollins S, Lee M, et al. Induction of MAGE-A3 and HPV-16 immunity by Trojan vaccines in patients with head and neck carcinoma. Head Neck (2012) 34(12):1734–46. doi: 10.1002/hed.22004
165. Schuler PJ, Harasymczuk M, Visus C, Deleo A, Trivedi S, Lei Y, et al. Phase I dendritic cell p53 peptide vaccine for head and neck cancer. Clin Cancer Res (2014) 20(9):2433–44. doi: 10.1158/1078-0432.CCR-13-2617
166. Kenter GG, Welters MJ, Valentijn AR, Lowik MJ, Berends-van der Meer DM, Vloon AP, et al. Vaccination against HPV-16 oncoproteins for vulvar intraepithelial neoplasia. N Engl J Med (2009) 361(19):1838–47. doi: 10.1056/NEJMoa0810097
167. van Poelgeest MI, Welters MJ, Vermeij R, Stynenbosch LF, Loof NM, Berends-van der Meer DM, et al. Vaccination against Oncoproteins of HPV16 for Noninvasive Vulvar/Vaginal Lesions: Lesion Clearance Is Related to the Strength of the T-Cell Response. Clin Cancer Res (2016) 22(10):2342–50. doi: 10.1158/1078-0432.CCR-15-2594
168. Hilf N, Kuttruff-Coqui S, Frenzel K, Bukur V, Stevanovic S, Gouttefangeas C, et al. Actively personalized vaccination trial for newly diagnosed glioblastoma. Nature (2019) 565(7738):240–5. doi: 10.1038/s41586-018-0810-y
169. Hu Z, Ott PA, Wu CJ. Towards personalized, tumour-specific, therapeutic vaccines for cancer. Nat Rev Immunol (2018) 18(3):168–82. doi: 10.1038/nri.2017.131
170. Sahin U, Derhovanessian E, Miller M, Kloke BP, Simon P, Lower M, et al. Personalized RNA mutanome vaccines mobilize poly-specific therapeutic immunity against cancer. Nature (2017) 547(7662):222–6. doi: 10.1038/nature23003
171. Demaria S, Golden EB, Formenti SC. Role of Local Radiation Therapy in Cancer Immunotherapy. JAMA Oncol (2015) 1(9):1325–32. doi: 10.1001/jamaoncol.2015.2756
172. Shaverdian N, Lisberg AE, Bornazyan K, Veruttipong D, Goldman JW, Formenti SC, et al. Previous radiotherapy and the clinical activity and toxicity of pembrolizumab in the treatment of non-small-cell lung cancer: a secondary analysis of the KEYNOTE-001 phase 1 trial. Lancet Oncol (2017) 18(7):895–903. doi: 10.1016/S1470-2045(17)30380-7
173. Bonomo P, Desideri I, Loi M, Mangoni M, Sottili M, Marrazzo L, et al. Anti PD-L1 DUrvalumab combined with Cetuximab and RadiOtherapy in locally advanced squamous cell carcinoma of the head and neck: A phase I/II study (DUCRO). Clin Transl Radiat Oncol (2018) 9:42–7. doi: 10.1016/j.ctro.2018.01.005
174. Bahig H, Aubin F, Stagg J, Gologan O, Ballivy O, Bissada E, et al. Phase I/II trial of Durvalumab plus Tremelimumab and stereotactic body radiotherapy for metastatic head and neck carcinoma. BMC Cancer (2019) 19(1):68. doi: 10.1186/s12885-019-5266-4
175. Yu Y, Lee NY. JAVELIN Head and Neck 100: a Phase III trial of avelumab and chemoradiation for locally advanced head and neck cancer. Future Oncol (2019) 15(7):687–94. doi: 10.2217/fon-2018-0405
176. Elbers JBW, Al-Mamgani A, Tesseslaar MET, van den Brekel MWM, Lange CAH, van der Wal JE, et al. Immuno-radiotherapy with cetuximab and avelumab for advanced stage head and neck squamous cell carcinoma: Results from a phase-I trial. Radiother Oncol (2020) 142:79–84. doi: 10.1016/j.radonc.2019.08.007
177. Vermeer DW, Spanos WC, Vermeer PD, Bruns AM, Lee KM, Lee JH. Radiation-induced loss of cell surface CD47 enhances immune-mediated clearance of human papillomavirus-positive cancer. Int J Cancer (2013) 133(1):120–9. doi: 10.1002/ijc.28015
178. Advani R, Flinn I, Popplewell L, Forero A, Bartlett NL, Ghosh N, et al. CD47 Blockade by Hu5F9-G4 and Rituximab in Non-Hodgkin’s Lymphoma. N Engl J Med (2018) 379(18):1711–21. doi: 10.1056/NEJMoa1807315
179. Logtenberg MEW, Jansen JHM, Raaben M, Toebes M, Franke K, Brandsma AM, et al. Glutaminyl cyclase is an enzymatic modifier of the CD47- SIRPalpha axis and a target for cancer immunotherapy. Nat Med (2019) 25(4):612–9. doi: 10.1038/s41591-019-0356-z
180. Hanoteau A, Newton JM, Krupar R, Huang C, Liu HC, Gaspero A, et al. Tumor microenvironment modulation enhances immunologic benefit of chemoradiotherapy. J Immunother Cancer (2019) 7(1):10. doi: 10.1186/s40425-018-0485-9
181. Beyranvand Nejad E, van der Sluis TC, van Duikeren S, Yagita H, Janssen GM, van Veelen PA, et al. Tumor Eradication by Cisplatin Is Sustained by CD80/86-Mediated Costimulation of CD8+ T Cells. Cancer Res (2016) 76(20):6017–29. doi: 10.1158/0008-5472.CAN-16-0881
182. van der Sluis TC, van Duikeren S, Huppelschoten S, Jordanova ES, Beyranvand Nejad E, Sloots A, et al. Vaccine-induced tumor necrosis factor-producing T cells synergize with cisplatin to promote tumor cell death. Clin Cancer Res (2015) 21(4):781–94. doi: 10.1158/1078-0432.CCR-14-2142
183. Goh AR, Shin SP, Jung NR, Ryu CH, Eom HS, Lee JH, et al. Low-dose cisplatin converts the tumor microenvironment into a permissive state for HSVtk-induced antitumor immunity in HPV16-related tonsillar carcinoma. Cancer Lett (2015) 356(2 Pt B):743–50. doi: 10.1016/j.canlet.2014.10.022
184. Dorta-Estremera S, Hegde VL, Slay RB, Sun R, Yanamandra AV, Nicholas C, et al. Targeting interferon signaling and CTLA-4 enhance the therapeutic efficacy of anti-PD-1 immunotherapy in preclinical model of HPV(+) oral cancer. J Immunother Cancer (2019) 7(1):252. doi: 10.1186/s40425-019-0728-4
185. Cillo AR, Kurten CHL, Tabib T, Qi Z, Onkar S, Wang T, et al. Immune Landscape of Viral- and Carcinogen-Driven Head and Neck Cancer. Immunity (2020) 52(1):183–99.e189. doi: 10.1016/j.immuni.2019.11.014
186. Ijsselsteijn ME, van der Breggen R, Farina Sarasqueta A, Koning F, de Miranda N. A 40-Marker Panel for High Dimensional Characterization of Cancer Immune Microenvironments by Imaging Mass Cytometry. Front Immunol (2019) 10:2534. doi: 10.3389/fimmu.2019.02534
187. Welters MJ, van der Sluis TC, van Meir H, Loof NM, van Ham VJ, van Duikeren S, et al. Vaccination during myeloid cell depletion by cancer chemotherapy fosters robust T cell responses. Sci Transl Med (2016) 8(334):334ra352. doi: 10.1126/scitranslmed.aad8307
188. Melief CJM, Welters MJP, Vergote I, Kroep JR, Kenter GG, Ottevanger PB, et al. Strong vaccine responses during chemotherapy are associated with prolonged cancer survival. Sci Transl Med (2020) 12(535):eaaz8235. doi: 10.1126/scitranslmed.aaz8235
189. Abdulrahman Z, De Miranda NF, Van Esch EM, De Vos van Steenwijk PJ, Nijman HW, Welters MJP, et al. A pre-existing inflammatory immune microenvironment predicts the clinical response of vulvar high-grade squamous intraepithelial lesions to therapeutic HPV16 vaccination. J Immunother Cancer (2020) 8(1):e000563. doi: 10.1136/jitc-2020-000563
190. Duhen T, Duhen R, Montler R, Moses J, Moudgil T, de Miranda NF, et al. Co-expression of CD39 and CD103 identifies tumor-reactive CD8 T cells in human solid tumors. Nat Commun (2018) 9(1):2724. doi: 10.1038/s41467-018-05072-0
191. Gros A, Robbins PF, Yao X, Li YF, Turcotte S, Tran E, et al. PD-1 identifies the patient-specific CD8(+) tumor-reactive repertoire infiltrating human tumors. J Clin Invest (2014) 124(5):2246–59. doi: 10.1172/JCI73639
192. Parkhurst M, Gros A, Pasetto A, Prickett T, Crystal JS, Robbins P, et al. Isolation of T-Cell Receptors Specifically Reactive with Mutated Tumor-Associated Antigens from Tumor-Infiltrating Lymphocytes Based on CD137 Expression. Clin Cancer Res (2017) 23(10):2491–505. doi: 10.1158/1078-0432.CCR-16-2680
193. Ye Q, Song DG, Poussin M, Yamamoto T, Best A, Li C, et al. CD137 accurately identifies and enriches for naturally occurring tumor-reactive T cells in tumor. Clin Cancer Res (2014) 20(1):44–55. doi: 10.1158/1078-0432.CCR-13-0945
194. Hung K, Hayashi R, Lafond-Walker A, Lowenstein C, Pardoll D, Levitsky H. The central role of CD4(+) T cells in the antitumor immune response. J Exp Med (1998) 188(12):2357–68. doi: 10.1084/jem.188.12.2357
195. Levitsky HI, Lazenby A, Hayashi RJ, Pardoll DM. In vivo priming of two distinct antitumor effector populations: the role of MHC class I expression. J Exp Med (1994) 179(4):1215–24. doi: 10.1084/jem.179.4.1215
196. Shankaran V, Ikeda H, Bruce AT, White JM, Swanson PE, Old LJ, et al. IFNgamma and lymphocytes prevent primary tumour development and shape tumour immunogenicity. Nature (2001) 410(6832):1107–11. doi: 10.1038/35074122
197. Hunder NN, Wallen H, Cao J, Hendricks DW, Reilly JZ, Rodmyre R, et al. Treatment of metastatic melanoma with autologous CD4+ T cells against NY-ESO-1. N Engl J Med (2008) 358(25):2698–703. doi: 10.1056/NEJMoa0800251
198. Quezada SA, Simpson TR, Peggs KS, Merghoub T, Vider J, Fan X, et al. Tumor-reactive CD4(+) T cells develop cytotoxic activity and eradicate large established melanoma after transfer into lymphopenic hosts. J Exp Med (2010) 207(3):637–50. doi: 10.1084/jem.20091918
Keywords: tumor microenvironment, immunotherapy, oropharyngeal cancer, T cells, myeloid cells, clinical outcome, survival
Citation: Welters MJP, Santegoets SJ and van der Burg SH (2020) The Tumor Microenvironment and Immunotherapy of Oropharyngeal Squamous Cell Carcinoma. Front. Oncol. 10:545385. doi: 10.3389/fonc.2020.545385
Received: 24 March 2020; Accepted: 16 November 2020;
Published: 21 December 2020.
Edited by:
Sven Saussez, University of Mons, BelgiumReviewed by:
Tina Dalianis, Karolinska Institutet (KI), SwedenJerome R. Lechien, University of Mons, Belgium
Copyright © 2020 Welters, Santegoets and van der Burg. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sjoerd H. van der Burg, c2h2ZGJ1cmdAbHVtYy5ubA==