
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Oncol. , 20 July 2015
Sec. Stem Cell Research
Volume 5 - 2015 | https://doi.org/10.3389/fonc.2015.00155
 Mohit Kumar Jolly1,2
Mohit Kumar Jolly1,2 Marcelo Boareto1,3
Marcelo Boareto1,3 Bin Huang1,4
Bin Huang1,4 Dongya Jia1,5
Dongya Jia1,5 Mingyang Lu1
Mingyang Lu1 Eshel Ben-Jacob1,6,7†José N. Onuchic1,4,7,8*
Eshel Ben-Jacob1,6,7†José N. Onuchic1,4,7,8* Herbert Levine1,2,7,8*
Herbert Levine1,2,7,8*
Transitions between epithelial and mesenchymal phenotypes – the epithelial to mesenchymal transition (EMT) and its reverse the mesenchymal to epithelial transition (MET) – are hallmarks of cancer metastasis. While transitioning between the epithelial and mesenchymal phenotypes, cells can also attain a hybrid epithelial/mesenchymal (E/M) (i.e., partial or intermediate EMT) phenotype. Cells in this phenotype have mixed epithelial (e.g., adhesion) and mesenchymal (e.g., migration) properties, thereby allowing them to move collectively as clusters. If these clusters reach the bloodstream intact, they can give rise to clusters of circulating tumor cells (CTCs), as have often been seen experimentally. Here, we review the operating principles of the core regulatory network for EMT/MET that acts as a “three-way” switch giving rise to three distinct phenotypes – E, M and hybrid E/M – and present a theoretical framework that can elucidate the role of many other players in regulating epithelial plasticity. Furthermore, we highlight recent studies on partial EMT and its association with drug resistance and tumor-initiating potential; and discuss how cell–cell communication between cells in a partial EMT phenotype can enable the formation of clusters of CTCs. These clusters can be more apoptosis-resistant and have more tumor-initiating potential than singly moving CTCs with a wholly mesenchymal (complete EMT) phenotype. Also, more such clusters can be formed under inflammatory conditions that are often generated by various therapies. Finally, we discuss the multiple advantages that the partial EMT or hybrid E/M phenotype have as compared to a complete EMT phenotype and argue that these collectively migrating cells are the primary “bad actors” of metastasis.
Despite remarkable progress in charting the hallmarks of cancer, understanding the cell-fate decisions during tumor initiation, progression, dormancy, and relapse is a major challenge in modern oncology (1). These dynamic decisions enable the tumor cells to tolerate therapeutic assaults such as chemotherapy or radiation; adapt to common micro-environmental stress that they face during cancer progression such as hypoxia, nutrient deprivation, and inflammation; and complete their “metastasis-invasion cascade” to seed tumors in distant organs at early stages; thereby posing unpleasant surprises in the clinical trials.
Recently, there has been rapid progress in characterizing these cell-fate decisions or cellular plasticity by mapping the underlying regulatory networks associated with the tumor–stroma ecosystem such as epithelial–mesenchymal plasticity, dedifferentiation of cancer cells to Cancer Stem Cells (CSCs), drug resistance, cell senescence, metabolic reprograming, response to hypoxia, and tumor angiogenesis (1–3). Cell-fate determination in these examples involve changes in expression of various transcription factors (TFs), miRNA (miRs), and epigenetic regulators that govern the underlying regulatory networks and consequently generate genome-wide distinct expression patterns of genes and proteins corresponding to a particular cell fate (3).
An archetypical example of cell-fate decisions or cellular plasticity during tumor progression is the transition between epithelial and mesenchymal phenotypes – epithelial to mesenchymal transition (EMT) and its reverse MET. EMT marks the first step of “invasion-metastasis cascade” where epithelial cells of the primary tumor lose their cell–cell adhesion and apico-basal polarity, and gain the ability to migrate individually and invade basement membrane and blood vessels. Upon intravasation, these cells stay in the bloodstream as circulating tumor cells (CTCs), until they exit at some distant organs to seed micrometastases. During seeding, they undergo the reverse of EMT – MET – to regain their epithelial characteristics and form secondary tumors or macrometastases, thereby completing their “metastasis-invasion cascade”. Therefore, EMT and MET enable solid tumors, over 90% of which are epithelial in nature (carcinomas) (4), to disseminate and colonize distant organs. However, EMT and MET are not exclusive to cancer, rather they play crucial roles in organogenesis during embryonic development, and wound healing or tissue regeneration where they are regulated tightly, but cancer cells “hijack” this developmental process for metastasis (5, 6) – the cause of 9 out of 10 cancer-related deaths (7).
Importantly, EMT and MET, whether in physiological or pathological contexts, are not binary processes (5, 6). Some cells can attain a hybrid epithelial/mesenchymal (E/M) phenotype, also referred to as partial or intermediate or incomplete EMT phenotype (8–10). In fact, many carcinoma cells may metastasize without completely losing an epithelial morphology and/or completely attaining mesenchymal traits (4, 11). Cells in the hybrid E/M phenotype have both epithelial (cell–cell adhesion) and mesenchymal (migration) traits, hence allowing collective cell migration, as seen during migration of multicellular aggregates in the ECM (4) and clusters of CTCs in bloodstream of breast, lung, and prostate cancer patients (12–14) (Figure 1A). Cells in the CTC clusters co-express epithelial and mesenchymal markers (15), can exit the bloodstream more efficiently (16), are apoptosis-resistant, and can be up to 50 times more metastatic than individually migrating CTCs (17). Therefore, the ability of metastatic cells to attain this hybrid E/M phenotype, rather than a complete EMT phenotype, poses a higher metastatic risk in patients (18).
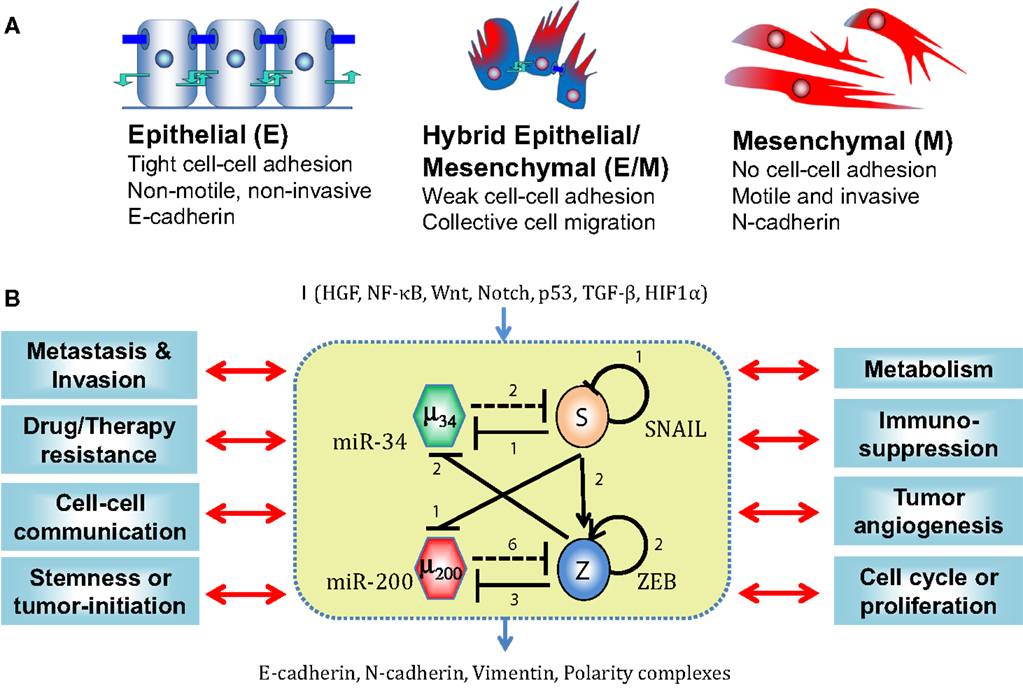
Figure 1. EMT phenotypes and core EMT network. (A) Canonical morphological and functional characteristics of the three phenotypes – epithelial (E), hybrid epithelial/mesenchymal (E/M), and mesenchymal (M). (B) Core EMT regulatory network (shown in yellow box) consists of two interconnected mutually inhibitory feedback loops – (miR-34/SNAIL and miR-200/ZEB). Solid bars represent transcriptional inhibition, solid arrows represent transcriptional activation, and dotted lines denote miRNA-mediated regulation. Numbers mentioned alongside each regulation are the number of binding sites for that particular regulation, as experimentally determined or proposed. This core network receives inputs from a variety of signals (shown by I), modulates many cytoskeletal elements (E-cadherin, N-cadherin, Vimentin, and polarity complexes), and couples with many other cellular traits. [adapted from Refs. (22) and (41)]
Here, we focus our review on elucidating how cells attain this phenotype, characterizing this hybrid E/M phenotype, and discussing why cells in this phenotype are the primary “bad actors” of cancer metastasis.
Epithelial cells can undergo EMT under the influence of many signaling pathways such as TGFβ, EGF, HGF, Notch, FGF, Wnt, and IGF (19), and mechanical factors such as ECM density (20). These signals usually activate one of the EMT-inducing transcription factors (EMT-TFs) – TWIST1, SNAI1, SNAI2 (SLUG), ZEB1, ZEB2 (SIP1), Brachyury, Goosecoid, SIX1, and PRRX1 – that directly or indirectly repress E-cadherin, the hallmark of epithelial phenotype. Conversely, EMT can be inhibited by p53, MET-TFs such as GRHL2 and ELF5, and microRNA (miR) families such as miR-200 and miR-34 (3).
In many carcinomas, these signals converge on a core EMT regulatory network, also referred to as “motor of cellular plasticity” owing to its coupling with many key cellular properties such as apoptosis, cell cycle, metabolism, and immunosuppression (21–25). This regulatory network is composed of two TF families – SNAIL and ZEB and two miR families – miR-200 and miR-34. The epithelial phenotype corresponds to high levels of miR-200 and miR-34, whereas the mesenchymal phenotype corresponds to high levels of ZEB and SNAIL. These components form two interlinked mutually inhibitory feedback loops – miR-34/SNAIL and miR-200/ZEB (26–28), such that EMT-inducing signals such as TGFβ, EGF, HGF, and Notch activate ZEB and SNAIL, and p53 activates miR-200 and miR-34 (3) (Figure 1B).
Mutually inhibitory feedback loops between two fate-determining TFs are one of the simplest gene circuits, and form a central motif in many cell-fate decisions. For instance, CDX2 and OCT4 control the fate of pluripotent embryonic stem cells – CDX2 induces trophoectoderm (TE) fate and OCT4 induces the opposite “sister” fate – inner cell mass (ICM) (29). Similarly, cross-inhibitory TFs PU.1 and GATA.1 are situated at the branch point of erythroid and myeloid lineages in hematopoiesis. The mutual repression between the two TFs guarantees mutual exclusivity of the two identities (for instance, an erythroid cell cannot be a myeloid cell and vice versa), and hence distinct cell identities (29). Therefore, a mutually inhibitory loop between two TFs A and B usually behave as binary or bistable switches allowing two distinct cell-fates – one corresponding to (high A, low B) expression and the other by (low A, high B), or in other words, (1,0) and (0,1) states where “0” denotes relatively low expression, and “1” denotes high expression (30–32) (Figure 2).
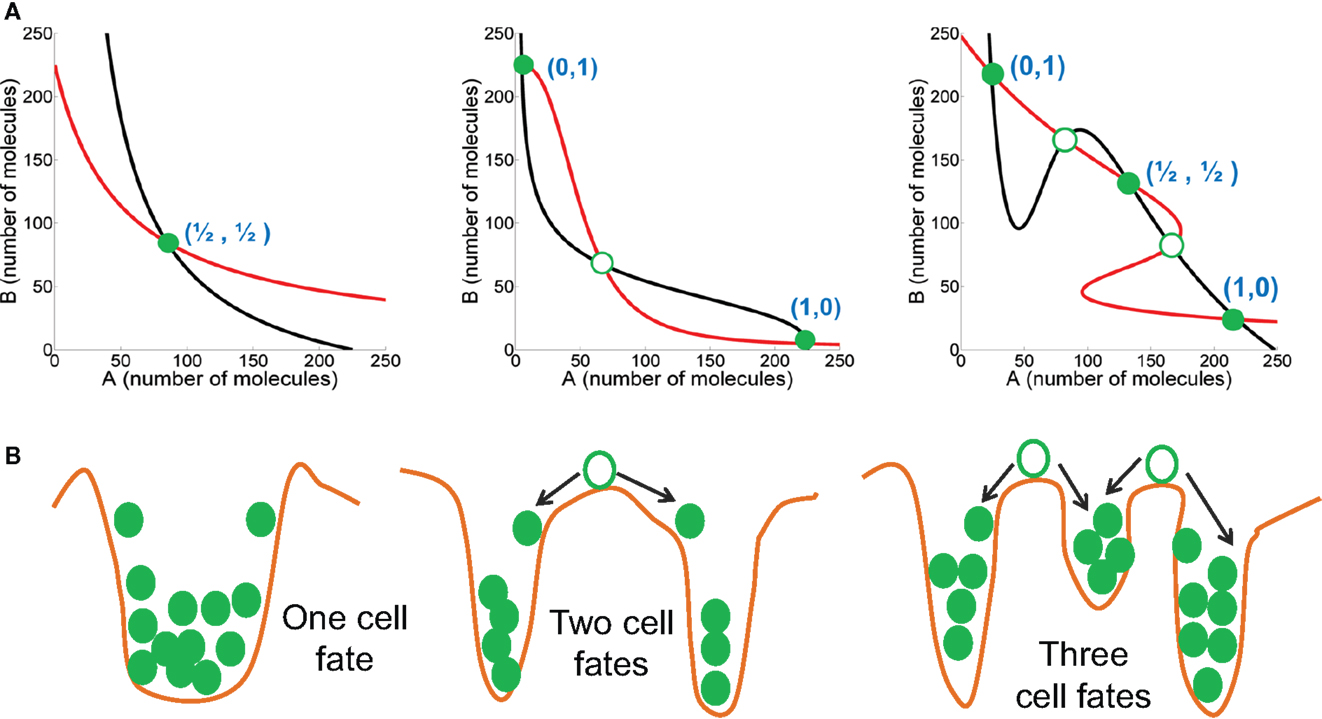
Figure 2. Dynamic characteristics of mutually inhibitory feedback loops. (A) (left) Weak mutual inhibition between A and B allows monostability where the steady state has intermediate levels of both A and B; (middle) Strong mutual inhibition between A and B can drive one species to extremely low levels, and therefore bistability, such that the two steady states are – (high A, low B) or (1,0), and (low A, high B) or (0,1); (right) Strong mutual inhibition between A and B, coupled with strong self-activation of both A and B can enable the system to be tristable, such that the three steady states are – (high A, low B) or (1,0), and (low A, high B) or (0,1), and (medium A, medium B) or (1/2, 1/2). Red and black curves describe nullclines for A and B, and their intersections are the steady states. Green-filled circles represent stable steady states, and green hollow circles show unstable steady states. The thickness of lines representing mutual inhibition between A and B, and self-activation of A and B represent relative strength of those interactions. (B) Cartoons (corresponding to the circuit drawn in the same column) representing the potential energy of the system, where valleys represent stable steady states, and crests denote unstable steady states.
There can be two important variations to this bistable behavior of a mutually inhibitory feedback loop. First, if mutual repression between the two TFs is not strong enough, both A and B are co-expressed at some intermediate level (1/2, 1/2) and the feedback loop does not give rise to two distinct cell fates (33, 34). Second, if one or both TFs auto-activate themselves strongly in addition to strongly repressing the other TF, the circuit can allow for three distinct phenotypes – (1, 0), (0, 1), and (1/2, 1/2) – or (high A, low B), (low A, high B), and (medium A, medium B). The (1/2, 1/2) state can act as the “poised” state of a progenitor cell that can differentiate to attain either of the two lineages – (1, 0) or (0, 1) (31, 35) (Figure 2).
Importantly, distinct cell fates, as discussed above, are different than quantitative trait variation between two cells belonging to the same fate. For instance, CDX2 levels in two cells both belonging to TE are most likely to be slightly different because of cellular stochasticity or non-genetic heterogeneity (36). However, neither of these cells spontaneously, or upon a small perturbation, transdifferentiate to adopt a different fate. Transdifferentiation often requires a large external signal such as overexpression of some cell-fate “master regulator” TFs (37, 38). This robust behavior of cell-fates reflects that they are “stable steady states” of the underlying regulatory network, characterized by a particular range of values of all variables (expression levels) of all the elements in the system (genes, chromatin states, etc.) (29). Therefore, the existence of distinct cell fates in a cell population is often manifested as a multimodal distribution of some of these elements that can be captured in FACS experiments (Figure 3A). Conversely, if a system is monostable but exhibits quantitative trait variation as a function of external drivers, the FACS distribution will be roughly Gaussian (Figure 3B). Consequently, switching cell-fates usually entails a discontinuous jump in the expression levels of many genes, and can therefore be observed only in a multistable system, but not in stochastic variations within a monostable system (same cell fate). For instance, when a bipotent progenitor cell type differentiates to adopt any one of the two daughter lineages, the steady state corresponding to the bipotent progenitor disappears or loses its stability, and two new stable steady states emerge – each corresponding to a daughter lineage and each with a new and distinctive expression pattern (39) (Figure 3A).
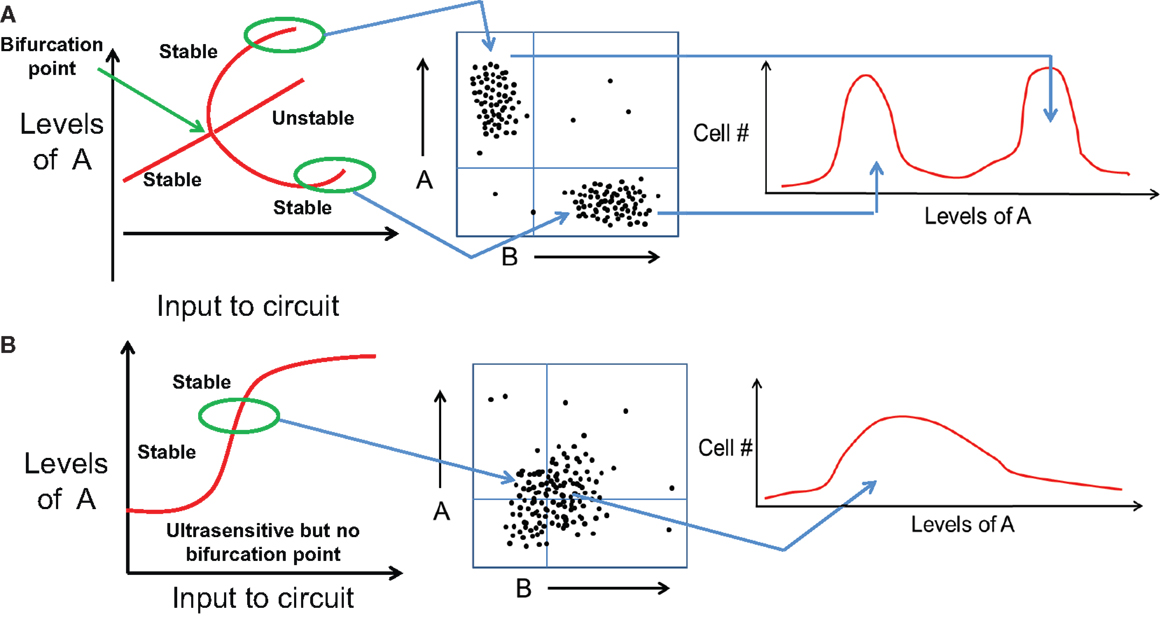
Figure 3. Distinct cell fates vs. quantitative trait variation of same cell fate. (A) (Left) bifurcation diagram representing variation in levels of A as an input is applied to a bistable mutually inhibitory circuit between A and B. At some threshold value of the input signal (marked by bifurcation point), the initial cell fate disappears and gives rise to two new stable steady state or cell fates. (middle) These two cell fates can be observed as different subpopulations in a FACS experiment. (right) Most cells attain one of the two cell fates, and the population distribution is bimodal with different range of values of A. (B) (left) Bifurcation diagram representing variation in levels of A as an input is applied to a monostable mutually inhibitory circuit between A and B. The circuit responds in an ultrasensitive manner but no bifurcation of cell fates observed. (middle) FACS experiments show a population with continuously varying levels of A without any sharp boundaries, hence (right) the population distribution is unimodal and broadly Gaussian.
These major qualitative as well as quantitative differences between distinct cell fates that emerge and then disappear as inputs are varied vs. quantitative trait variation of a single fate have important implications. In a typical experiment, some specific attributes of the population are monitored as a control parameter is varied. Monostable systems exhibit no hysteresis and no multimodality in the population structure; they can however be ultrasensitive and thereby exhibit sharp thresholds in dose–response curve (Figure 4A). A system that exhibits multiple states with individual cells making fate decisions will in general exhibit hysteresis, will often exhibit multimodality, in addition to being able to exhibit sharp thresholds as the systems reaches points of bifurcation (Figure 4B). We shall argue below that the experimental data currently available for the EMT process strongly suggests an interpretation in terms of distinct cell fates, but this needs to be carefully addressed in more quantitative and carefully designed future experiments.
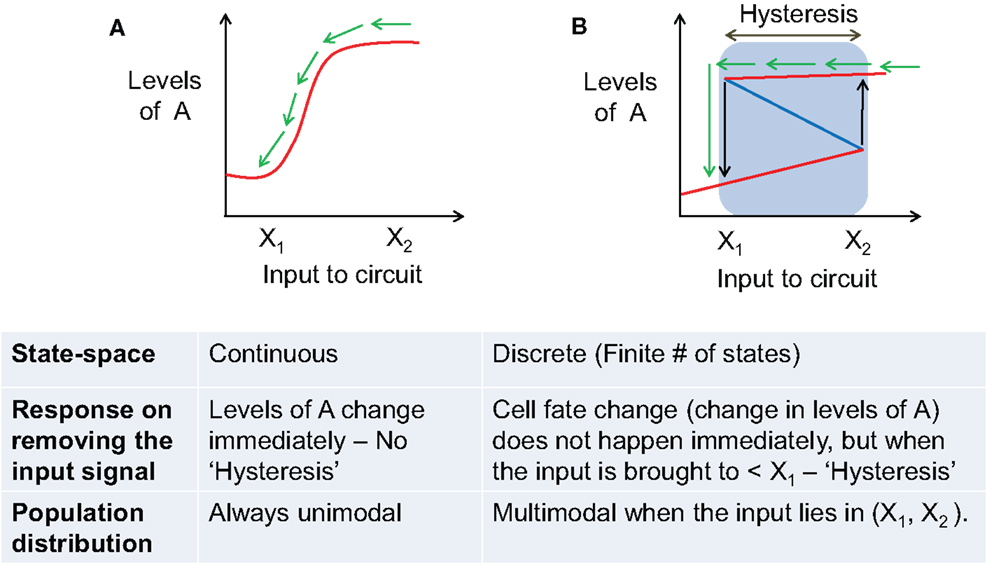
Figure 4. Comparing the behavior of a phenotypic transition that has continuous state-space vs. the one with discrete state-space. (A) Behavior of a system with continuous state-space (infinite stable steady states) during a phenotypic transition as induced by an external input signal. Solid red lines show stable states, and blue dotted lines show unstable steady states. (B) Behavior of a system with discrete state-space [finite number (n = 2 here) of stable steady states (cell fates)]. Blue lines represent stable steady states, and red line denotes unstable steady states. Blue shaded region shows the range of hysteresis and bistability. Black arrows mark the levels of input where the cell switches fate (or transitions from one stable steady state to another) – X1, X2. The table presented below compares the behavior of the two scenarios depicted in (A,B). In both (A,B), green arrows denote the response of the system when the input signal is removed.
As mentioned above, the core EMT network comprises two mutually inhibitory loops – (miR-34/SNAIL) and (miR-200/ZEB). Two computational models of this network have proposed different functions for these two loops. Tian et al. (40) have proposed that both the loops – (miR-34/SNAIL) and (miR-200/ZEB) – function as bistable or binary switches that initiate and complete EMT, respectively. They define the E phenotype as (high miR-200 and miR-34, low ZEB and SNAIL), M phenotype as (low miR-200 and miR-34, high ZEB and SNAIL), and partial EMT as (low miR-34 and ZEB, high SNAIL and miR-200). On the other hand, we propose that miR-34/SNAIL acts as a noise-buffering integrator of various EMT- and MET-inducing signals, preventing aberrant activation of EMT or MET due to transient signals, but not giving rise to any phenotypic transitions by itself. In other words, we argue that this subsystem would be monostable if it could be detached from any feedback from downstream effectors. Conversely, miR-200/ZEB, with input from SNAIL, behaves as a tristable or three-way switch allowing for the existence of three phenotypes – E (high miR-200, low ZEB), M (low miR-200, high ZEB), and E/M or partial EMT (medium miR-200, medium ZEB) (22, 41).
Both existing models provide similar explanations for the E and M phenotypes, and can therefore be compared to experiments that focus on cells that undergo a complete EMT. Experiments showing that SNAIL can initiate repression of E-cadherin but ZEB is required for its complete inhibition (42), and that most genes repressed during EMT are inhibited by ZEB irrespective of the EMT-inducing signal (43), are consistent with either model; both approaches argue for ZEB activation to be necessary for a complete EMT (transition to a completely mesenchymal phenotype). Similarly, experiments showing that upon withdrawing the EMT-inducing signal, only the cells with low ZEB levels, but not high ZEB levels, revert to being epithelial immediately, indicate that ZEB activation marks a commitment point for cells to undergo an EMT (44) – a prediction both models make. Parenthetically, this lack of reversion is direct evidence in favor of the multistability picture (Figure 4B). Further, both the models predict that reverting EMT requires suppressing the EMT-inducing signal as well as ZEB, and that SNAIL knockdown does not suffice. Experiments validating these predictions, again, fail to discriminate between the two models (45–47).
However, experimental studies focusing on partial EMT can distinguish between the two models, and appear in our opinion to be more consistent with (medium miR-200, medium ZEB) definition of partial EMT rather than with (high miR-200, low ZEB). For example, studies in mammary morphogenesis – a canonical case of partial EMT – identified a TF that can maintain the terminal end bud (TEB) cells in a partial EMT phenotype – OVOL, and knockdown of OVOL leads to complete EMT. Thus, OVOL acts as a “critical molecular brake on EMT” (48). OVOL is coupled with EMT core circuit in an intricate manner – it forms a mutually inhibitory switch with ZEB, inhibits miR-200 indirectly, and self-inhibits (49–52). Adding these interactions to our model, we showed that OVOL expands the range of parameters or physiological conditions for the existence of partial EMT or hybrid E/M phenotype (53), thereby explaining its role in maintaining the partial EMT phenotype. Similarly, co-expression of ZEB1 and E-cadherin in cells undergoing gastrulation (another example of partial EMT) (54) and, as discussed later in this review, the association of partial EMT with high tumor-initiating potential (“stemness”), are more likely to correspond to the (medium miR-200, medium ZEB) state structure for partial EMT.
The different results for partial EMT in the two models emerge from different modeling assumptions. The study by Tian et al (40) assumes simple universal forms for the various repressive interactions in the double-switch circuit. This assumption ignores key experimentally identified differences between the architecture of these two loops – two binding sites of miR-34 on SNAIL mRNA vs. six binding sites of miR-200 on ZEB mRNA (55, 56), self-inhibition of SNAIL vs. (indirect) self-activation of ZEB (57, 58), and finally the difference in transcriptional regulation vs. translational regulation by miRs (59–63). Importantly, the number of binding sites of a miR on an mRNA is crucial for determining the fold-change repression in protein expression, as shown by experiments that overexpression of miR-34 reduces SNAIL levels to 50% of the initial levels, but overexpression of miR-200 reduces ZEB levels to 10% (55, 56). Further, the self-inhibition of SNAIL is critical to avoid any aberrant activation of EMT from transient activation of signals, and sets a sensitivity threshold for various EMT-inducing signals (42). Finally, the mechanisms of transcriptional regulation and miR-mediated sequestration and degradation of target mRNAs are distinct from each other and hence typically represented by different functional forms (31, 59–63). Nevertheless, the manifestation of partial EMT state can be cell line-specific (64), because, for instance, not all cell lines might have same number of available miR-200 binding sites on ZEB mRNA, therefore, more quantitative measurement at the single-cell level is required to decipher which characterization of partial EMT holds in a particular context.
Different levels of SNAIL enable different phenotypes and/or combinations thereof; for instance, low levels of SNAIL cannot induce an EMT, and very high levels can induce a complete EMT in almost the entire population (41). However, as observed in both physiological and pathological EMT contexts, the population can be highly heterogeneous, allowing for the emergence of distinct subpopulations of cells with different phenotypes. Cells in these distinct subpopulations may also interconvert their phenotypes due to intracellular stochastic fluctuations. Of course, different cell lines (or biological contexts) would be expected to have different ratios of these subpopulations (18, 65) (Figure 5A). Such cell-to-cell heterogeneity might have crucial functional consequences, especially in adaptive drug resistance, tumor dormancy, and the heterogeneity in CTCs (36, 66–69). These variations ride above purely genomic variations, which themselves can be quite extensive, given the compromised genome integrity in most cancers (1).
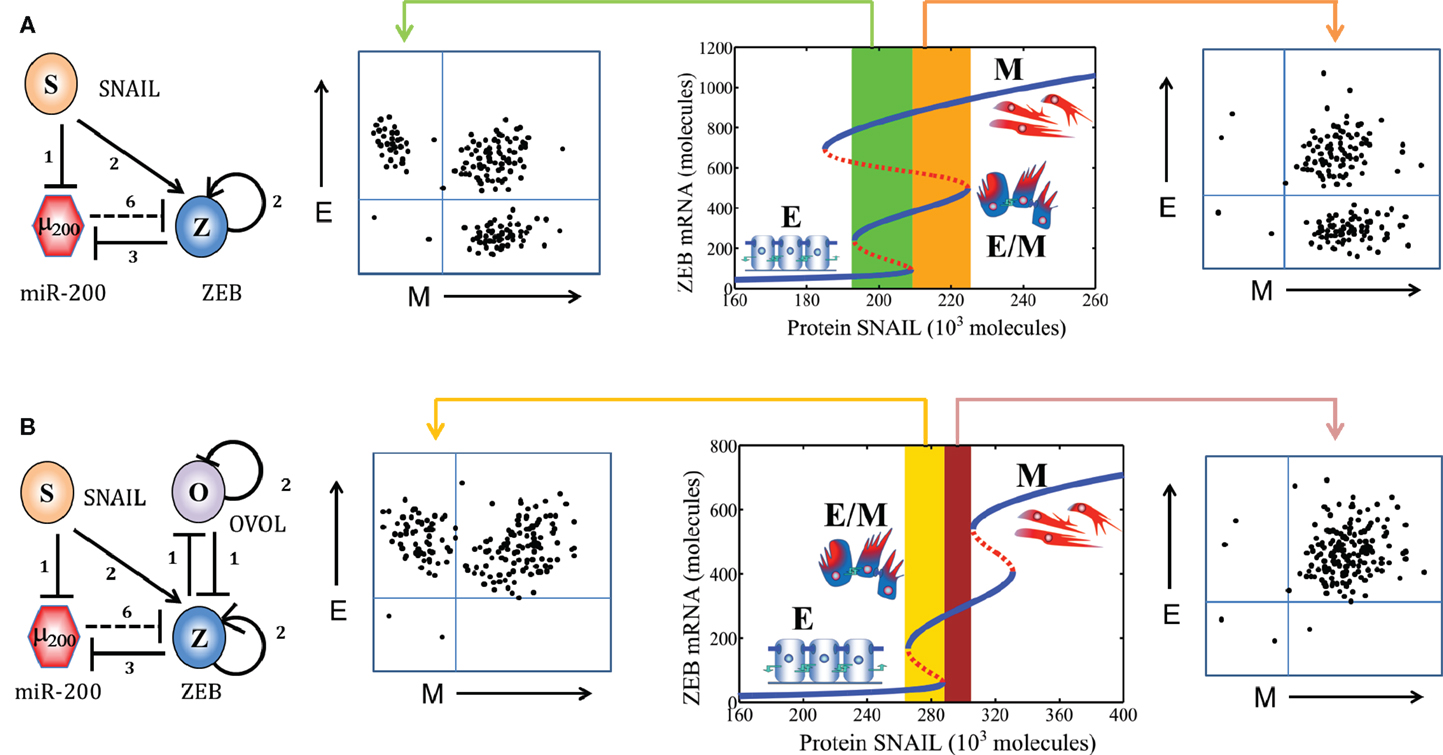
Figure 5. Population distribution or multimodality in EMT response. (A) (Middle) bifurcation of ZEB mRNA levels in response to protein SNAIL (EMT-inducing signal) for the miR-200/ZEB/SNAIL circuit (shown at extreme left). For a certain range of SNAIL values (marked by green rectangle), cells can attain any of the three phenotypes – E, M, and E/M, giving rise to a trimodal population distribution as shown in FACS figure (left). For a different range of SNAIL values (marked by orange rectangle), cells can adopt either E/M or M phenotype, and be distributed in a bimodal manner in FACS figure (right). (B) Bifurcation of ZEB mRNA levels in response to protein SNAIL (EMT-inducing signal) for the miR-200/ZEB/SNAIL/OVOL circuit (shown at extreme left). For a certain range of SNAIL values (marked by yellow rectangle), cells can adopt either E/M or E phenotype, and be distributed in a bimodal manner (FACS figure, left); and for a different range (marked by brown rectangle) all cells are likely to be in E/M phenotype as shown in FACS figure (right). Importantly, as compared to the behavior of miR-200/ZEB/SNAIL circuit, miR-200/ZEB/SNAIL/OVOL circuit allows the existence of new phases (combinations of phenotypes) such as {E/M} and {E, E/M}, and precludes the existence of phases {E, E/M, M}.
Recently, an important quantitative metric – an “EMT score” – has been proposed to represent the overall proclivity of a cell line or primary tumor toward undergoing EMT, however, it largely ignores the cellular heterogeneity and possible clonal heterogeneity inherent to a particular cell line (70). Not surprisingly, these scores vary continuously. Given the evidence described above that the E and M states are truly different cell fates, the continuous variation argues in favor of additional stable intermediate states that occupy different positions on “EMT axis” (64, 70); otherwise, we would in general expect to see a sharper score variation. This is of course what we have already expected based on the circuit models and based on the analogy between pathological EMT and the physiological EMT examples of wound healing and branching morphogenesis. It remains to be investigated precisely how many stable intermediate states are present en route EMT and whether this inference is proven correct by individual cell studies. Also, it must be noted that unlike developmental EMT, pathological EMT might not necessarily involve a real lineage-switching of cells in an epithelial lineage to a mesenchymal one (71).
Another related important question that needs to be answered is that how morphologically stable is (are) the intermediate state(s) of EMT. Partial EMT has been usually labeled as a “metastable” state (10), indicating that it is less stable than pure E or pure M ones. However, recent experimental studies have identified that some epigenetic changes (72) as well as some “phenotypic stability factors” such as OVOL (73) can stabilize the partial EMT phenotype and/or fine-tune the transitions into and from it. Cells expressing endogenous levels of OVOL can maintain their partial EMT phenotype, knockdown of OVOL leads to complete EMT and overexpression of OVOL induces the reversal of EMT – a MET (48, 49). These experimental findings can be unified via our theoretical framework by coupling OVOL to the core EMT network, where we show that OVOL can both act as a “critical molecular brake on EMT” preventing the cells “that have gained partial plasticity” to undergo a complete EMT, and a driver of MET when overexpressed (48, 53) (Figure 5B). Our work on OVOL serves as an example of how our theoretical framework for the core EMT network renders itself to analyzing the role of other regulatory players in epithelial plasticity (53).
Cells that become motile as a result of (complete) EMT appear to come in two distinct shapes and concomitant behaviors, namely mesenchymal and amoeboid (74). Note that there is no guarantee that cells described as M from the genetic network perspective always have mesenchymal shapes. Cells labeled as mesenchymal are spindle-shaped, have lamellopodia and/or filopodia on their leading edge, adhere strongly to the ECM, and act as “path generators” by secreting matrix metallo-proteinases (MMPs). Conversely, amoeboid cells are round-shaped, often have blebby structures, have low adhesion to ECM, and show a higher shape plasticity that helps them squeeze through the gaps in ECM and act as “path finders” (75, 76). Further, cells can adopt a shape representing both amoeboid and mesenchymal traits (hybrid A/M) such as cells with both lamellopodia and blebs (77). In cancer, there is a rich plasticity that allows cells to adopt functional behaviors depending on external signals, phenotypic choices, and of course genetic changes – such as switching between amoeboid and mesenchymal morphologies – a mesenchymal to amoeboid transition (MAT) and its reverse – AMT, and direct bidirectional switching between hybrid E/M and A phenotypes – a collective to amoeboid transition (CAT) and its reverse – ACT (78–82). Presumably, this plasticity enables them to adapt to different environments encountered during metastasis, and is therefore critical for tumor dissemination (79) (Figure 6A).
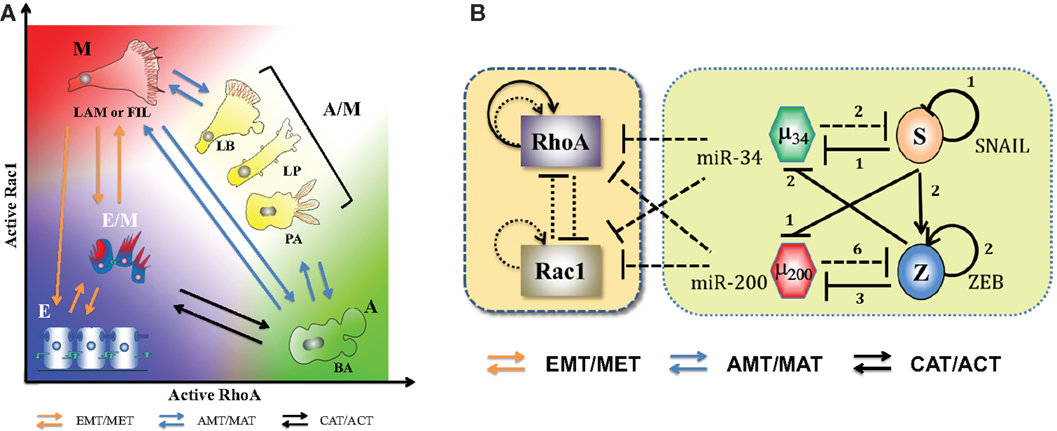
Figure 6. Landscape of cellular shape plasticity during carcinoma metastasis. (A) Cartoon representation of different cell shapes/phenotypes with their respective places on the two-dimensional space of levels of active RhoA (RhoA-GTP) and active Rac1 (Rac1-GTP). As miR-34 and miR-200 inhibit both RhoA and Rac1, both epithelial and hybrid E/M phenotypes have low levels of active forms of RhoA and Rac1. The (high RhoA-GTP, low Rac1-GTP) profile associates with amoeboid (A) morphology with blebs [blebby amoeboid (BA)], whereas (low RhoA-GTP, high Rac1-GTP) associates with mesenchymal (M) shape – cells with lamellopodia or filopodia (LAM or FIL). Cells with (high RhoA-GTP, high Rac1-GTP) adopt a hybrid A/M morphology that can be manifested in multiple ways – lamellipoida with blebs (LB), lobopodia (LP), and pseudopodal amoeboid (PA). Transitions among E, E/M, and M phenotypes (EMT/MET) are represented by orange arrows, those between amoeboid and mesenchymal morphologies – A, A/M, and M – are denoted by blue arrows, and transitions between E/M and A phenotypes – CAT/ACT – are denoted by black arrows. (B) Circuits showing the coupling of core EMT circuit with RhoA and Rac1 – the two GTPases that are critical in regulating cell shape. They inhibit the GTP loading (switching from inactive GDP-bound state to active GTP-bound state) of each other and promote that of themselves (shown by dotted lines). Also, RhoA can activate itself indirectly on a transcription level (solid black lines) (see Ref. (74) and references therein). The microRNAs miR-34 and miR-200 inhibit the translation of RhoA and Rac1. Figure adapted from Ref. (74).
Elucidating the principles of this plasticity requires investigating the coupling between the core EMT circuits and the downstream effectors actually responsible for actualization of motility biophysics. One key piece is the mutually repressing feedback loop between the two GTPases – RhoA and Rac1 – that promote their own GTP loading and inhibit that of the other. Activation of RhoA increases actomyosin contractility resulting in membrane blebbing and facilitating a rounded amoeboid phenotype. Conversely, activation of Rac1 results in focal adhesions and actin polymerization, leading to formation of lamellopodia, enabling a front-back polarized spindle-shaped mesenchymal cell (74, 83). Importantly, these two GTPases play crucial roles during EMT in converting apico-basal polarity typical of cells in epithelial layers to front-back polarity needed for motion. For instance, Rac1 activation at the leading edge stimulates PI3K that leads to indirect self-activation of Rac1, therefore setting up a positive feedback loop for cytoskeletal reorganization necessary for cells to gain directional migration abilities. Similarly, RhoA promotes actin stress fiber formation and prevents formation of the polarity complexes PAR at the rear end of the cell (2). Quite reasonably, the epithelial gatekeepers miR-34 and miR-200 inhibit the translation of RhoA and Rac1 (84–86) (Figure 6B). However, the resultant dynamics of this complex interplay remains a challenge for the future. In particular, a comprehensive understanding of cell shape dynamics and its coupling to EMT will require integrating live-cell imaging with a multi-compartment spatiotemporal model capturing the spatial segregation of the GTPases (87–89).
The partial EMT phenotype – (medium miR-200, medium ZEB) – has been studied extensively in embryonic development and wound healing (8, 90, 91). A canonical process showing the role of partial EMT in development is the branching morphogenesis of the trachea and mammary gland – a mechanism that enables the repeated splitting of a tubular epithelial structure to generate a ductal tree. During branching morphogenesis, the tip cells located at the cap of the TEBs of the growing tubule maintain cell–cell adhesion with neighbors and transiently display mesenchymal features such as loss of apico-basal polarity and increased motility in response to extracellular signals such as FGF (8, 9). These collectively migrating cells express P-cadherin, a proposed marker for partial EMT (11, 92), and form finger-like projections and maintain their partial EMT phenotype (i.e., do not proceed to a complete EMT) possibly due to the action of a “critical molecular brake on EMT” – the TF OVOL – whose knockdown leads to solitary and impaired migration (48). Similar to TEB migration, during sprouting angiogenesis, the “tip” endothelial cells display a partial endothelial to mesenchymal transition (pEndMT) transition and lead the collective migration of a train of “stalk” cells (93). Further, in wound healing, immature basal keratinocytes at the wound edge partially remodel their basement membrane and migrate collectively in a “metastable” partial EMT phenotype, and finally, revert to being epithelial or, in other words, undergo re-epithelialization to close the wound (9, 10, 94, 95). Collective migration in most partial EMT cases is mediated by SLUG (SNAIL2) (93, 96–100). Such collective migration has multiple advantages – it obviates the need for all cells to detect external signals for migration, allows coupling of mechanical forces among the cells, and provides them with maximum plasticity to be able to switch to being epithelial or mesenchymal (complete EMT) phenotypes (8). These advantages can be utilized by carcinomas during invasion and intravasation of multicellular strands (101).
Recent studies have highlighted the crucial significance of partial EMT in cancer metastasis. Cells co-expressing various epithelial and mesenchymal markers are present in primary breast and ovarian cancer (15, 102), in multiple cell lines belonging to ovarian, lung, and renal cell carcinoma (64, 103, 104), as well as in mouse models of pancreatic ductal adenocarcinoma (PDAC) and prostate cancer (105, 106). Importantly, among breast cancer subtypes, the ones with poor clinical outcomes – triple-negative breast cancer (TNBC) and basal-like breast cancer (BLBC) – are most enriched for such biphenotypic cells, indicating a strong association between aggressiveness and E/M phenotype (15, 107, 108). Further, co-expression of mesenchymal marker vimentin and epithelial/luminal markers cytokeratins (CK) 8 and 18, rather than the expression of vimentin alone, correlates with increased invasive and metastatic potential and poor survival and is often observed in many aggressive tumors such as BLBC and melanomas (109–113). Besides, a gene signature consisting of both epithelial and mesenchymal genes predicts poor outcomes independent of breast cancer subtype (18), suggesting that the association of partial EMT phenotype with aggressiveness can be context-independent.
Cells co-expressing E and M markers can also be present in the bloodstream of breast, lung, colon, and prostate cancer patients as clusters of CTCs that contain a median level of three cells per cluster (12–15, 114). These clusters, also referred to as “microemboli”, can be apoptosis-resistant, are more likely to be trapped in narrow blood vessels for extravasation, and often correlate with poor prognosis in patients (13, 16, 17, 115). Although these clusters constitute only 3% of total CTC “events” observed (97% being individually migrating CTCs), they contribute 50% of the total metastases, reflecting their increased metastatic propensity (17). Further, they can be found in the bloodstream of patients with COPD (chronic obstructive pulmonary disease) around 3 years before a lung nodule can be detected, and therefore they might be useful to identify patients at a greater risk of developing lung cancer (116). Importantly, the heightened metastatic potential of such clusters as compared to that of the same numbers of individual cells was recognized (117, 118) even before EMT was characterized as a metastasis mechanism (119).
However, CTC clusters need not necessarily contain only the hybrid E/M cells, and a comprehensive understanding of other cell types that might be present in these clusters is necessary for advancing the clinical application of CTCs analysis as a “liquid biopsy” (16, 120). There may be admixtures of E and M cells in a single cluster. Also, representing the cellular heterogeneity of the primary tumor, some of these clusters may contain leukocytes as well as platelets and megakaryocytes (15, 121), therefore true “seeds” of metastasis (metastasis-initiating cells) within these CTCs must be identified carefully using functional assays such as xenotransplantation in immunodeficient mice. Initial attempts in this direction have elucidated that the cancer cells in a hybrid E/M phenotype (identified by SNAIL+ E-cad+ in colon, and by EPCAM+ CD44+ CD47+ MET+ in breast cancer) can more efficiently act as seeds of metastasis (122, 123), hence establishing a clinical and prognostic relevance of the cells in the hybrid E/M phenotype. Nonetheless, not all cells in a hybrid E/M phenotype might be capable of initiating a tumor in vivo (124).
A subpopulation of cancer cells that seed metastasis or, in other words, have self-renewal as well as clonal tumor initiation ability along with long-term clonal repopulation potential are referred to as Cancer Stem Cells (CSCs). These cells with stem-cell properties (“stemness”) can evade cell death and cancer therapeutics, and may stay dormant for long periods of time (125). However, in the context of cancer, “stemness” is not a fixed inherent trait of a few privileged cells, rather CSCs and non-CSCs can interconvert among themselves, and this plasticity or dynamic equilibrium drives tumor growth as well as invasion (126–129). Functional assays of isolating CSCs include mammosphere-formation in vitro and limiting dilution assays of tumor-initiating potential in vivo in NOD/SCID mice. In other words, CSCs are usually characterized by high evolvability (capacity to give rise to heritable phenotypic variation) (130).
Under some conditions, cells undergoing a full EMT have been shown to be highly likely to gain “stemness” and behave operationally as Cancer Stem Cells (CSCs). This EMT-stemness coupling was first reported for immortalized human mammary epithelial cells (131, 132). Similar findings in many carcinomas such as pancreatic, hepatocellular, and colorectal have strengthened this notion (133). Therefore, aberrant activation of EMT can serve at least two functions – (a) increases the invasion ability to reach distant organs for metastasis and (b) enhances tumor-initiating properties of the cells that reach the metastatic sites (134). However, this notion of a full EMT coupled with stemness has been challenged by studies showing that repression of EMT is required for effective tumor-initiation (135–138) and that reprograming often involves MET (139, 140).
A few recent studies attempt to resolve this contradiction by suggesting that instead of the cells in pure epithelial (E) or pure mesenchymal (M) states, cells in hybrid E/M or partial EMT state are most likely to gain stemness (18, 106, 124, 141). Grosse-Wilde et al. show that co-expression of E and M genes in the very same cell promotes mammosphere formation and stemness, independent of the breast cancer subtype (18). Further, Strauss et al. showed that some cells in hybrid E/M phenotype in primary ovarian cultures and tumors in situ can be multipotent, express markers of other lineages, and drive tumor growth in vivo by giving rise to another E/M subset as well as completely differentiated epithelial cells (124). Ruscetti et al. isolated hybrid E/M cells in vivo in a prostate cancer mouse model and demonstrated their comparable or even higher sphere formation and tumor-initiating potential as compared to completely mesenchymal cells (106). Also, our study that mathematically models the stemness-decision circuit (LIN28/let-7) with inputs from miR-200 and NF-κB suggests that especially at high levels of NF-κB, hybrid E/M state is more likely to gain stemness than complete EMT (141). These studies propose that cells undergoing partial EMT, but not necessarily complete EMT, can gain stemness, or in other words, the “stemness window” lies somewhere close to midway on the “EMT axis” (Figure 7) (134); and are consistent with experiments showing that more than 80% CTCs in men with castration-resistant prostate cancer (CRPC) and over 75% of CTCs in women with metastatic breast cancer co-express epithelial markers CK, mesenchymal markers N-cadherin, and stem cell markers (14).
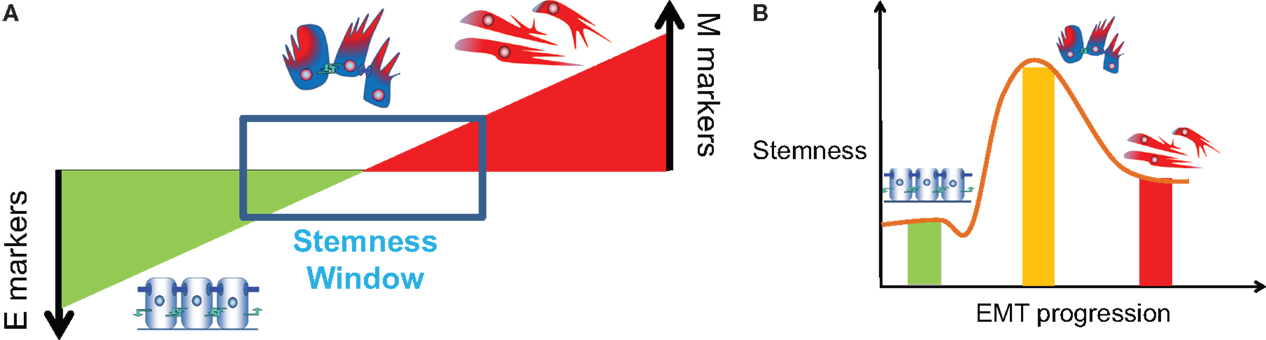
Figure 7. Association of partial EMT with stemness. (A) “EMT gradient” model proposed by Ombrato and Malanchi (134) where stemness is maintained within a window between a fully differentiated epithelial cell and a fully de-differentiated mesenchymal cell [Figure adapted from Ref. (134), cartoons included]. (B) Stemness or tumor-initiating potential of E, E/M, and M phenotype, or variation of stemness during EMT progression (brown line).
This association of hybrid E/M phenotype with stemness is not specific to tumor progression, but has also been reported in physiological EMT examples where adult hepatic stem/progenitor cells (HSCs) co-express epithelial and mesenchymal genes, and give rise to both epithelial and mesenchymal lineages in the liver (142–145). Similar to HSCs, adult renal progenitors are in a “metastable” hybrid E/M state upon tissue injury and mediate renal repair and regeneration (146). Collectively, these studies present strong evidence for the emerging notion that CTCs in a semi-mesenchymal phenotype, rather than those “frozen” or locked in a full EMT phenotype, have the highest plasticity to switch between proliferative and invasive modes, are capable of completing the invasion-metastasis cascade, and should therefore be regarded as CSCs (120, 147–150).
An alternative hypothesis that attempts to resolve the connection between EMT, MET, and stemness proposes that CSCs come in two distinct states – “epithelial-like” and “mesenchymal-like” (151, 152). Importantly, these studies show that most epithelial-like CSCs (identified by ALDH+ by Liu et al. (151) and by CD44high EPCAMhigh by Biddle et al. (152)) can give rise to both epithelial-like and mesenchymal-like populations and hence bilineage colonies in vitro. However, this plasticity was significantly impaired in the mesenchymal-like CSCs (identified by CD44+/CD24− by Liu et al. (151) and by CD44high EPCAMlow by Biddle et al. (152), thereby contributing to the notion that cells locked in a complete EMT phenotype significantly lose their plasticity. Neither of these studies considered the possibility that there might exist hybrid E/M states, but did show that not all CSC’s have to have the same EMT properties. At present, the exact mapping between EMT and stemness appears to be complex and context-dependent (6, 153), but with evidence suggesting that the major target to avoid tumor relapse and metastasis might be the CTCs in a hybrid E/M phenotype which have same degree of stemness (154). It, therefore, becomes essential to find a set of markers that would enable such cells to be identified.
Identifying a robust set of markers to isolate CTCs in a hybrid E/M phenotype remains an open question. CTCs with an epithelial phenotype can be identified via cell surface markers such as EPCAM (epithelial cell adhesion molecule) or cytoskeletal markers such as CK8, CK18, and CK19. However, establishing similar markers for CTCs with at least a partially mesenchymal phenotype has been challenging because vimentin is expressed in most normal blood cells as well (120). A potential signature for identifying the “stem-like” hybrid E/M CTCs may be CD24+CD44+, the expression pattern for pancreatic and gastric CSCs (155, 156), because CD24 is a canonical epithelial marker, and CD44 is a mesenchymal stem cell one (157, 158), and CD24+CD44+ expression pattern overlaps with high levels of P-cadherin, another proposed marker of partial EMT phenotype (92). Recent studies have highlighted that CD24+CD44+ cells can have up to 10 times higher mammosphere-initiating capacity, and can form more aggressive tumors than CD44+/CD24− cells (18, 66) that have been traditionally considered to be CSCs (159). CD24+CD44+ cells are present in multiple cell lines belonging to the luminal and basal-like subtypes, and their population is enriched significantly upon exposure to an acute cytotoxic shock, suggesting that they represent a drug-tolerant subpopulation that can repopulate a tumor (66). Collectively, these studies show that CD44+/CD24− expression does not necessarily correlates with tumorigenicity (160), and consolidate the mounting evidence that cells with a biphenotypic E/M expression tend to have high tumorigenicity in mice (156, 161–163).
While establishing a robust set of markers such as CD24+CD44+ for detecting the E/M cells in CTCs, at least two cautionary steps must be taken. First, a more quantified characterization of the presence of markers is required to identify the intermediate state(s) of EMT. For instance, CD24neg cells must be segregated from CD24lo cells, as they mark different lineages in mouse mammary gland, have dissimilar tumorigenic potential and respond differently to gamma-secretase inhibitors (GSI) due to their distinct gene expression profiles (158, 161). Second, the clusters of CTCs need to be isolated and investigated for different cell types present in them. Owing to their residual cell–cell adhesion, the CTCs in a hybrid E/M phenotype are likely to attach to cancer cells and/or stromal cells to form CTC clusters. Therefore, isolating CTC clusters should have two major advantages – (a) capturing hybrid E/M cells that are not necessarily present on the surface of the cluster and (b) revealing novel insights into the cooperation of cancer cells and/or cancer cells and stromal cells present in the same cluster. Such cooperation is expected to recapitulate the tumor–stroma ecology seen in primary tumors and metastasis, where some stromal cells can be “activated” by cancer through cytokines to provide metabolic synergy and signals for survival and maintaining stemness (164–167). “Activated” stromal cells can also be carried along as the “soil” by the accompanying “seed” metastatic cells to gain early growth advantage during colonization (168).
Cell–cell communication among cancer cells and/or between cancer cells and stromal cells (fibroblasts, immune cells, endothelial cells, etc.) can have a significant influence on phenotypic plasticity (EMT/MET), CSC self-renewal, and a dynamic equilibrium between CSCs and non-CSCs (125, 128). Spatial heterogeneity in the tumor can lead to spatial variations of secreted factors, cell types that are in direct contact, ECM density, etc. each of which can affect the “EMT score” of individual cancer cells in the tissue (20, 169). A key signaling pathway that is involved in multiple aspects of this cross-talk both via cell–cell contact and via soluble factors is Notch signaling. Notch signaling can induce EMT and maintain stemness (170); however, our understanding of the different roles of the two sub-families of ligands (Delta and Jagged) of Notch signaling is only recent (171–173), and incomplete in the context of EMT and/or CSCs.
Notwithstanding this lack of knowledge, Jagged1 is emerging as a potential therapeutic target for its roles in maintaining and increasing CSCs, inhibiting apoptosis, inducing angiogenesis, and affecting the immune cells (171). It can both be secreted by endothelial cells as well as present on the membrane of stromal and cancer cells and can activate Notch signaling in cancer cells to increase the CSC population (174, 175). Further, it is implicated in colonization where it is present on the surface of breast cancer cells and can activate Notch signaling in the bone (176), correlates with poor survival outcome, is overexpressed in CSCs, and has much higher levels in the more aggressive forms of breast cancer such as TNBC and BLBC than in its luminal subtypes (171, 177).
Importantly, Notch-Jagged (N-J) communication might be the preferred mode of tumor–stroma signaling than Notch-Delta (N-D) signaling due to its multiple potential synergistic effects in the tumor ecology. Two cells interacting via N-D signaling usually adopt distinct fates – one cell behaves as Sender [high ligand (Delta), low receptor (Notch)] and the other as Receiver [low ligand (Delta), high receptor (Notch)], therefore allowing only one-directional signaling and “salt-and-pepper” cell-fate patterns (178) (Figure 8B). Conversely, the two cells interacting via N-J signaling can adopt similar fates – hybrid Sender/Receiver [medium ligand (Jagged), medium receptor (Notch)] that enables bidirectional communication between them (172, 173), and allows lateral induction, i.e., a cell induces its neighbor to adopt the same fate as that of its own (179–181) (Figure 8C). Due to this lateral induction mechanism observed in N-J signaling, a cluster of E/M cells interacting via N-J signaling might mutually stabilize their “metastable” phenotype and consequently maintain high “stemness” (172). This notion is supported by the involvement of Notch signaling in wound healing (182). N-J signaling in collectively moving cells can induce or maintain similar fates as that of the neighboring cells, thereby coordinating wound healing, but excessive N-D signaling might impair it. Importantly, if partial EMT is defined as (high miR-200, low ZEB) rather than (medium miR-200, medium ZEB), collectively moving cells with active Notch signaling are likely to have suppressed N-J signaling almost completely because miR-200 inhibits Jagged1 strongly (183) and therefore might diversify their fates via N-D signaling, a phenomenon that would impair wound healing (41) (Figures 8A,D). A tantalizing possibility nevertheless, it remains to be tested both experimentally and via a theoretical model of the coupled core EMT circuit and Notch-Delta-Jagged signaling via interactions such as miR-200 inhibits Jagged (183), miR-34 inhibits Delta and Notch (184, 185), and NICD activates SNAIL (98, 186).
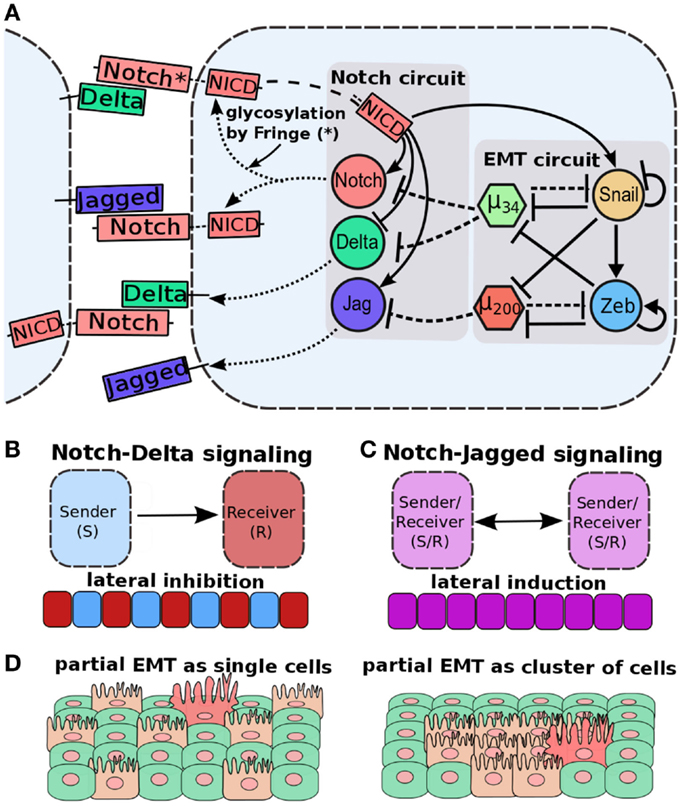
Figure 8. Cell–cell communication and partial EMT. (A) Coupling of EMT circuit with Notch circuit. Notch pathway, when activated by Jagged or Delta, belonging to neighboring cell, can activate Jagged and Notch, but inhibit Delta. EMT circuit couples with Notch circuit in many ways – miR-200 inhibits Jagged1, miR-34 inhibits both Notch and Delta, and NICD can activate SNAIL to drive EMT. (B) Notch-Delta signaling between two cells induces opposite fates in them – one cell behaves as a Sender (high Delta, low Notch) and the other a Receiver (high Notch, low Delta). Due to this lateral inhibition, it can promote “salt-and-pepper” based patterns. (C) Notch-Jagged signaling between two cells induces similar fates in them – lateral induction – and thus leads to patterns with all cells with the same fate. (D) (Left) cells in a partial EMT and interacting via N-D signaling might not be spatially close to each other, because N-D signaling inhibits two neighbors to adopt the same fate. (right) Cells in a partial EMT and interacting via N-J signaling can mutually stabilize the E/M phenotype and stay together as a cluster. Figure adapted from Ref. (173).
Notch-Jagged signaling can also mediate tumor–stroma interaction via regulating the secretion of many cytokines that can enslave or “activate” stromal cells (125). For instance, IL-6 secreted by cancer cells drives the activation of normal fibroblasts toward becoming cancer-associated fibroblasts (CAF) that in turn elicit an EMT response in cancer cells and increases the CSC population (164). Further, IL-6 can also promote the generation of tumor-associated macrophages (TAM) that support tumor metastasis (187). Many inflammatory cytokines such as IFN-γ and IL-6 can also increase production of Jagged and/or decrease that of Delta (177, 188, 189) hence possibly forming a positive feedback loop that rakes up N-J signaling and mediates chronic inflammation, a hallmark of cancer, in the stroma (1, 190).
Therefore, cancer cells can be considered as “ecological engineers” taking advantage of its niche in multiple ways, such as metabolic synergy and gaining CSC and migration traits (165–167). The outcome of such dynamic cross-talk can be best understood using an integrated computational and experimental approach – reconstructing the ecological dynamics of cancer via co-culture experiments, and building a multi-scale model combining intracellular signaling with population level spatial models. Such an understanding might provide valuable insights into therapies targeted at managing the stroma, as well as combinatorial therapies targeting both the cancer and stroma to avoid tumor relapse (165, 191, 192).
EMT has been posited to be involved in drug resistance (193–195), however, characterizing cell lines based on EMT scores indicates that this correlation might not be universally applicable in all carcinomas (70). Importantly, most studies connecting EMT to drug resistance has viewed EMT as an “all-or-none” process (193–195), leaving little scope for assessing the possible drug resistance in partial EMT phenotype, and comparing it with that corresponding to a complete EMT phenotype. Because CSCs have been reported to be primarily responsible for drug resistance (196), the association of hybrid E/M phenotype with stemness (18, 106, 124, 141) proposes that a hybrid E/M or partial EMT phenotype can also be the phenotype maximally correlated with drug resistance.
The partial EMT or “EMT-like” phenotype can associate with both de novo and adaptive drug resistance. Among various breast cancer subtypes, the TNBC contains the maximum number of hybrid E/M cells in the primary tumor (15), and exhibits de novo resistance to current standard therapies such as anthracyclines and taxanes (197). There is also a strong relationship to adaptive therapy. Significantly, a paradigm that emerges from many recent studies is that cancer cells that become resistant to many therapeutic assaults often undergo partial EMT. Development of tamoxifen-insensitivity in MCF7 breast cancer cells and that of trastuzumab resistance in HER2-overexpressing breast cancer cells is usually accompanied by a partial EMT (198, 199). Further, the radiation-resistant colorectal cancer cells generate cellular progeny with an “EMT-like” phenotype (200), and exposure to taxanes induces a phenotypic transition to a chemotherapy-tolerant state (CD44+CD24+) in multiple cell lines belonging to both basal-like and luminal subtypes (66). It must be noted that CD44+CD24+ expression pattern proposed is what we proposed above to be a hallmark of hybrid E/M cells in many cancer subtypes (18).
The underlying signaling pathways and molecular mechanisms of this interplay between partial EMT and drug resistance remain largely elusive. It is not even clear what aspects of drug resistance are phenotypic in character (reduction of growth rate, upregulation of pumps, etc.) and what depend on actual genetic changes and whether these are coupled via regulation of genomic instability. At the signaling level, key intermediary pathways involve N-J signaling (171), that as discussed above, can play a key role in shepherding the epithelial-mesenchymal plasticity by stabilizing a “metastable” partial EMT phenotype.
A key difference in partial EMT during wound healing and that during tumor progression is that during wound healing, cells often re-epithelialize after closing the wound, thereby limiting their plasticity, but during tumor progression, this ubiquitous plasticity spearheads aggressive tumor progression (94, 201). Further, wound healing often elicits an acute inflammatory response that is resolved later (94); however, during cancer, “the wounds that do not heal” (202), inflammatory response is chronic and is a hallmark of cancer (1, 190). Therefore, inflammation can regulate the timespan of heightened epithelial plasticity and more specifically, the timespan over which a hybrid E/M or partial EMT phenotype can be maintained. Such a “stabilizing” effect of inflammation on the “metastable” partial EMT phenotype can be mediated largely by N-J signaling, because many inflammatory factors such as TNF-α, IFN-γ, and IL-6 can increase the production of Jagged and/or decrease that of Delta (177, 188, 189), thereby promoting N-J signaling that can maintain a cluster of cells in a partial EMT phenotype. Consistently, the breast cancer subtype that has maximum number of cells co-expressing E and M genes among all breast cancer subtypes (15) – TNBC – has elevated levels of Jagged1 as well as NF-kB (203).
Inflammatory stress conditions in tissues are also created by both chemotherapy and radiation by activating NF-κB, the central link between inflammation, tumor progression, and radiation resistance (204–207). NF-kB and Jagged can activate each other (188, 208), thereby forming a self-perpetuating loop that maintains both high levels of NF-kB and N-J signaling. NF-kB can promote the likelihood for hybrid E/M cells to gain stemness (141), and as discussed above, N-J signaling can stabilize cells in a hybrid E/M phenotype; therefore post-therapy inflammatory conditions can promote a drug-resistant subpopulation that can be in hybrid E/M phenotype (Figure 9).
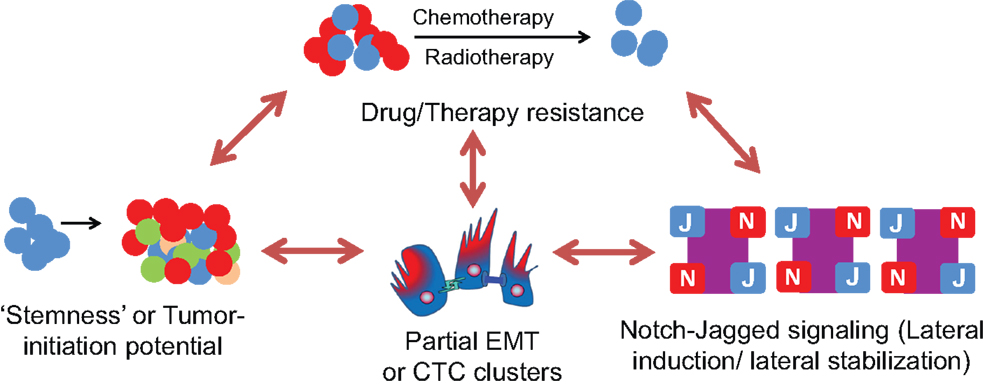
Figure 9. Interplay between Notch-Jagged signaling, Partial EMT or CTC clusters, stemness, and therapy/drug resistance. Cells in a hybrid E/M or partial EMT phenotype (as present in CTC clusters) can possess a much higher tumor-initiation potential (“stemness”) and drug resistance as compared to a completely mesenchymal phenotype. These cells can maintain their “metastable” hybrid E/M phenotype via Notch-Jagged signaling that promotes lateral stabilization (maintenance of same cell fate in neighboring cells) and/or lateral induction (propagation of the same fate as of its own to the neighbor) among a population of cells. This lateral induction can also be utilized to propagate drug resistance among a small subpopulation of cells known as “Cancer Stem Cells” (CSCs).
Epithelial to mesenchymal transition is a fundamental process in embryonic development and tissue repair that is aberrantly activated during the progression of cancer and fibrosis. Multiple cycles of EMT and MET are involved in organogenesis but usually not during adult homeostasis (5, 6). EMT was first described as “epithelial-mesenchymal transformation” in the pioneering work by Elizabeth Hay on primitive streak formation in the chick (209), however, later the term “transformation” was replaced with “transition” with the evidence accumulating that EMT was different from neoplastic transformation and that it was a reversible process. Recently, the term “transition” is giving way to “plasticity” with an increasing appreciation of the notion that EMT is not an “all-or-none” response, rather involves intermediate state(s) with important functional consequences in cancer metastasis as well as drug resistance and subsequent tumor relapse (6, 210–212).
The appreciation of EMT not being an “all-or-none” process is relatively recent in EMT associated with cancer and fibrosis, but has been generally accepted in wound healing and collective cell migration during embryonic development, especially gastrulation, neural crest migration, and branching morphogenesis (9, 10, 96, 213, 214). In cancer-related EMT, the concept of partial or incomplete EMT was initially proposed to reconcile the paradox that despite a presumed role of EMT in cancer progression, most metastatic carcinomas had well-differentiated epithelial characteristics; and it was difficult to identify cells having undergone EMT within the carcinoma tissue in vivo (4). Recent experimental evidence about cancer cells in primary tumor, cell lines, as well as in circulation (CTCs) (15, 64, 103, 104) have bolstered the concept of partial EMT and has moved it to a focal point in the EMT research.
Cells in a partial or intermediate EMT phenotype are likely to score multiple advantages over cells that have completed EMT or crossed the full mesenchymal “tipping point”. First, these cells can garner advantages specific to collective sheet or cluster migration – such a migration obviates the need for all cells to respond to external chemotactic signals, allowing for the passive migration of many carcinoma cells, and underlying the unexpected association of E-cadherin with tumor aggression (8, 215). Second, these cells display sufficient plasticity to switch to enable a switch back to colonization, yet primed for subsequent metastatic rounds (41, 216). Third, these cells are likely to be clustered together in the blood and are therefore anoikis-resistant, an essential trait for efficient metastasis (17). Also, clusters have a greater chance to get trapped in narrow blood vessels, therefore favoring extravasation into distant organs (16). Fourth, these cells can be immune-resistant and chemo-tolerant; and can even be enriched in the population following many therapy-related stresses such as inflammation and radiation (66, 217, 218). Fifth, these cells can have a much higher (~50-times) tumor-initiating and metastatic potential than cells in complete EMT phenotype (17, 18, 117, 124, 134, 141). Sixth, due to their residual cell–cell adhesion, these cells might form clusters of CTCs with other cell types such as leukocytes and fibroblasts and/or maintain the clusters via N-J signaling among themselves, thereby harnessing their “ecological engineering” skills during circulation (165). Collectively, the cells in a partial EMT or hybrid E/M phenotype have a much large repertoire of survival strategies in all stress conditions – be it shear stress in circulation, or stress due to therapeutic assaults; and are therefore better armed to seed metastases at distant organs and coordinate tumor relapse. Not surprisingly, these cells are being increasingly observed in many aggressive malignancies (105, 198, 219–228), strongly suggesting that partial EMT phenomena are more likely to happen in vivo than complete EMT (169, 229). Isolating CTC clusters (230) and testing them for partial EMT characteristics might be the most promising diagnostic approach in the clinic.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We have benefited from useful discussions with Mary C. Farach-Carson, Donald S. Coffey, Samir A. Hanash, Kenneth J. Pienta, Ilan Tsarfaty, and Sendurai A. Mani. This work was supported by National Science Foundation (NSF) Center for Theoretical Biological Physics (NSF PHY-1427654) and NSF grant MCB-1214457. HL and JNO are also supported as CPRIT (Cancer Prevention and Research Institute of Texas) Scholar in Cancer Research of the State of Texas at Rice University. EB-J was also supported by CPRIT and a grant from the Tauber Family Funds and the Maguy-Glass Chair in Physics of Complex Systems. MB was also supported by FAPESP (Grant 2013/14438-8). ML was also supported by a training fellowship from Keck Center for Interdisciplinary Bioscience Training of the Gulf Coast Consortia (CPRIT Grant RP140113).
ACT, amoeboid collective transition; AMT, amoeboid mesenchymal transition; BLBC, basal-like breast cancer; CAT, collective amoeboid transition; CSC, cancer stem cell; CTC, circulating tumor cell; EMT, epithelial mesenchymal transition; FACS, fluorescence-activated cell sorting; MAT, mesenchymal amoeboid transition; MET, mesenchymal epithelial transition; NICD, notch intracellular domain; TNBC, triple negative breast cancer.
1. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell (2011) 144:646–74. doi: 10.1016/j.cell.2011.02.013
2. Lamouille S, Xu J, Derynck R. Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol (2014) 15:178–96. doi:10.1038/nrm3758
3. De Craene B, Berx G. Regulatory networks defining EMT during cancer initiation and progression. Nat Rev Cancer (2013) 13:97–110. doi:10.1038/nrc3447
4. Christiansen JJ, Rajasekaran AK. Reassessing epithelial to mesenchymal transition as a prerequisite for carcinoma invasion and metastasis. Cancer Res (2006) 66:8319–26. doi:10.1158/0008-5472.CAN-06-0410
5. Kalluri R, Weinberg RA. Review series the basics of epithelial-mesenchymal transition. J Clin Invest (2009) 119:1420–8. doi:10.1172/JCI39104
6. Nieto MA. Epithelial plasticity: a common theme in embryonic and cancer cells. Science (2013) 342:1234850. doi:10.1126/science.1234850
7. Gupta GP, Massagué J. Cancer metastasis: building a framework. Cell (2006) 127:679–95. doi:10.1016/j.cell.2006.11.001
8. Revenu C, Gilmour D. EMT 2.0: shaping epithelia through collective migration. Curr Opin Genet Dev (2009) 19:338–42. doi:10.1016/j.gde.2009.04.007
9. Micalizzi DS, Farabaugh SM, Ford HL. Epithelial-mesenchymal transition in cancer: parallels between normal development and tumor progression. J Mammary Gland Biol Neoplasia (2010) 15:117–34. doi:10.1007/s10911-010-9178-9
10. Arnoux V, Côme C, Kusewitt DF, Hudson LG, Savagner P. Cutaneous wound reepithelialization. In: Savagner P, editor. Rise and Fall of Epithelial Phenotype. Berlin: Springer (2005). p. 111–34.
11. Klymkowsky MW, Savagner P. Epithelial-mesenchymal transition: a cancer researcher’s conceptual friend and foe. Am J Pathol (2009) 174:1588–93. doi:10.2353/ajpath.2009.080545
12. Lecharpentier A, Vielh P, Perez-Moreno P, Planchard D, Soria JC, Farace F. Detection of circulating tumour cells with a hybrid (epithelial/mesenchymal) phenotype in patients with metastatic non-small cell lung cancer. Br J Cancer (2011) 105:1338–41. doi:10.1038/bjc.2011.405
13. Hou J-M, Krebs M, Ward T, Sloane R, Priest L, Hughes A, et al. Circulating tumor cells as a window on metastasis biology in lung cancer. Am J Pathol (2011) 178:989–96. doi:10.1016/j.ajpath.2010.12.003
14. Armstrong AJ, Marengo MS, Oltean S, Kemeny G, Bitting RL, Turnbull JD, et al. Circulating tumor cells from patients with advanced prostate and breast cancer display both epithelial and mesenchymal markers. Mol Cancer Res (2011) 9:997–1007. doi:10.1158/1541-7786.MCR-10-0490
15. Yu M, Bardia A, Wittner BS, Stott SL, Smas ME, Ting DT, et al. Circulating breast tumor cells exhibit dynamic changes in epithelial and mesenchymal composition. Science (2013) 339:580–4. doi:10.1126/science.1228522
16. Joosse SA, Gorges TM, Pantel K. Biology, detection, and clinical implications of circulating tumor cells. EMBO Mol Med (2015) 7:1–11. doi:10.15252/emmm.201303698
17. Aceto N, Bardia A, Miyamoto DT, Donaldson MC, Wittner BS, Spencer JA, et al. Circulating tumor cell clusters are oligoclonal precursors of breast cancer metastasis. Cell (2014) 158:1110–22. doi:10.1016/j.cell.2014.07.013
18. Grosse-Wilde A, Fouquier d’Herouei A, McIntosh E, Ertaylan G, Skupin A, Kuestner RE, et al. Stemness of the hybrid epithelial/mesenchymal state in breast cancer and its association with poor survival. PLoS One (2015) 10:e0126522. doi:10.1371/journal.pone.0126522
19. Thiery JP, Sleeman JP. Complex networks orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell Biol (2006) 7:131–42. doi:10.1038/nrm1835
20. Kumar S, Das A, Sen S. Extracellular matrix density promotes EMT by weakening cell-cell adhesions. Mol Biosyst (2014) 10:838–50. doi:10.1039/c3mb70431a
21. Brabletz S, Brabletz T. The ZEB/miR-200 feedback loop – a motor of cellular plasticity in development and cancer? EMBO Rep (2010) 11:670–7. doi:10.1038/embor.2010.117
22. Lu M, Jolly MK, Onuchic J, Ben-Jacob E. Toward decoding the principles of cancer metastasis circuits. Cancer Res (2014) 74:4574–87. doi:10.1158/0008-5472.CAN-13-3367
23. Chen L, Gibbons DL, Goswami S, Cortez MA, Ahn Y, Byers LA, et al. Metastasis is regulated via microRNA-200/ZEB1 axis control of tumour cell PD-L1 expression and intratumoral immunosuppression. Nat Commun (2014) 5:1–12. doi:10.1038/ncomms6241
24. Pecot CV, Rupaimoole R, Yang D, Akbani R, Ivan C, Lu C, et al. Tumour angiogenesis regulation by the miR-200 family. Nat Commun (2013) 4:2427. doi:10.1038/ncomms3427
25. Sun Y, Daemen A, Hatzivassiliou G, Arnott D, Wilson C, Zhuang G, et al. Metabolic and transcriptional profiling reveals pyruvate dehydrogenase kinase 4 as a mediator of epithelial-mesenchymal transition and drug resistance in tumor cells. Cancer Metab (2014) 2:20. doi:10.1186/2049-3002-2-20
26. Park SM, Gaur AB, Lengyel E, Peter ME. The miR-200 family determines the epithelial phenotype of cancer cells by targeting the E-cadherin repressors ZEB1 and ZEB2. Genes Dev (2008) 22:894–907. doi:10.1101/gad.1640608
27. Burk U, Schubert J, Wellner U, Schmalhofer O, Vincan E, Spaderna S, et al. A reciprocal repression between ZEB1 and members of the miR-200 family promotes EMT and invasion in cancer cells. EMBO Rep (2008) 9:582–9. doi:10.1038/embor.2008.74
28. Bracken CP, Gregory PA, Kolesnikoff N, Bert AG, Wang J, Shannon MF, et al. A double-negative feedback loop between ZEB1-SIP1 and the microRNA-200 family regulates epithelial-mesenchymal transition. Cancer Res (2008) 68:7846–54. doi:10.1158/0008-5472.CAN-08-1942
29. Zhou JX, Huang S. Understanding gene circuits at cell-fate branch points for rational cell reprogramming. Trends Genet (2011) 27:55–62. doi:10.1016/j.tig.2010.11.002
30. Macía J, Widder S, Solé R. Why are cellular switches Boolean? General conditions for multistable genetic circuits. J Theor Biol (2009) 261:126–35. doi:10.1016/j.jtbi.2009.07.019
31. Andrecut M, Halley JD, Winkler DA, Huang S. A general model for binary cell fate decision gene circuits with degeneracy: indeterminacy and switch behavior in the absence of cooperativity. PLoS One (2011) 6:e19358. doi:10.1371/journal.pone.0019358
32. Guantes R, Poyatos JF. Multistable decision switches for flexible control of epigenetic differentiation. PLoS Comput Biol (2008) 4:e1000235. doi:10.1371/journal.pcbi.1000235
33. Cherry JL, Adler FR. How to make a biological switch. J Theor Biol (2000) 203:117–33. doi:10.1006/jtbi.2000.1068
34. Gardner TS, Cantor CR, Collins JJ. Construction of a genetic toggle switch in Escherichia coli. Nature (2000) 403:339–42. doi:10.1038/35002131
35. Huang S, Guo YP, May G, Enver T. Bifurcation dynamics in lineage-commitment in bipotent progenitor cells. Dev Biol (2007) 305:695–713. doi:10.1016/j.ydbio.2007.02.036
36. Balázsi G, Van Oudenaarden A, Collins JJ. Cellular decision making and biological noise: from microbes to mammals. Cell (2011) 144:910–25. doi:10.1016/j.cell.2011.01.030
37. Chin MT. Reprogramming cell fate: a changing story. Front Cell Dev Biol (2014) 2:46. doi:10.3389/fcell.2014.00046
38. Graf T, Enver T. Forcing cells to change lineages. Nature (2009) 462:587–94. doi:10.1038/nature08533
39. Pal M, Ghosh S, Bose I. Non-genetic heterogeneity, criticality and cell differentiation. Phys Biol (2015) 12:016001. doi:10.1088/1478-3975/12/1/016001
40. Tian X-J, Zhang H, Xing J. Coupled reversible and irreversible bistable switches underlying TGFβ-induced epithelial to mesenchymal transition. Biophys J (2013) 105:1079–89. doi:10.1016/j.bpj.2013.07.011
41. Lu M, Jolly MK, Levine H, Onuchic JN, Ben-Jacob E. MicroRNA-based regulation of epithelial-hybrid-mesenchymal fate determination. Proc Natl Acad Sci U S A (2013) 110:18174–9. doi:10.1073/pnas.1318192110
42. De Herreros AG, Baulida J. Cooperation, amplification, and feed-back in epithelial-mesenchymal transition. Biochim Biophys Acta (2012) 1825:223–8. doi:10.1016/j.bbcan.2012.01.003
43. Taube JH, Herschkowitz JI, Komurov K, Zhou AY, Gupta S, Yang J, et al. Core epithelial-to-mesenchymal transition interactome gene-expression signature is associated with claudin-low and metaplastic breast cancer subtypes. Proc Natl Acad Sci U S A (2010) 107:15449–54. doi:10.1073/pnas.1004900107
44. Gregory PA, Bracken CP, Smith E, Bert AG, Wright JA, Roslan S, et al. An autocrine TGF-beta/ZEB/miR-200 signaling network regulates establishment and maintenance of epithelial-mesenchymal transition. Mol Biol Cell (2011) 22:1686–98. doi:10.1091/mbc.E11-02-0103
45. Aigner K, Dampier B, Descovich L, Mikula M, Sultan A, Schreiber M, et al. The transcription factor ZEB1 (deltaEF1) promotes tumour cell dedifferentiation by repressing master regulators of epithelial polarity. Oncogene (2007) 26:6979–88. doi:10.1038/sj.onc.1210508
46. Zhang J, Tian X-J, Zhang H, Teng Y, Li R, Bai F, et al. TGF-β-induced epithelial-to-mesenchymal transition proceeds through stepwise activation of multiple feedback loops. Sci Signal (2014) 7:ra91. doi:10.1126/scisignal.2005304
47. Das S, Becker BN, Hoffmann FM, Mertz JE. Complete reversal of epithelial to mesenchymal transition requires inhibition of both ZEB expression and the Rho pathway. BMC Cell Biol (2009) 10:94. doi:10.1186/1471-2121-10-94
48. Watanabe K, Villarreal-Ponce A, Sun P, Salmans ML, Fallahi M, Andersen B, et al. Mammary morphogenesis and regeneration require the inhibition of EMT at terminal end buds by Ovol2 transcriptional repressor. Dev Cell (2014) 29:59–74. doi:10.1016/j.devcel.2014.03.006
49. Roca H, Hernandez J, Weidner S, McEachin RC, Fuller D, Sud S, et al. Transcription factors OVOL1 and OVOL2 induce the mesenchymal to epithelial transition in human cancer. PLoS One (2013) 8:e76773. doi:10.1371/journal.pone.0076773
50. Roca H, Pande M, Huo JS, Hernandez J, Cavalcoli JD, Pienta KJ, et al. A bioinformatics approach reveals novel interactions of the OVOL transcription factors in the regulation of epithelial – mesenchymal cell reprogramming and cancer progression. BMC Syst Biol (2014) 8:29. doi:10.1186/1752-0509-8-29
51. Guo L, Chen C, Shi M, Wang F, Chen X, Diao D, et al. Stat3-coordinated Lin-28-let-7-HMGA2 and miR-200-ZEB1 circuits initiate and maintain oncostatin M-driven epithelial-mesenchymal transition. Oncogene (2013) 32:5272–82. doi:10.1038/onc.2012.573
52. Nair M, Bilanchone V, Ortt K, Sinha S, Dai X. Ovol1 represses its own transcription by competing with transcription activator c-Myb and by recruiting histone deacetylase activity. Nucleic Acids Res (2007) 35:1687–97. doi:10.1093/nar/gkl1141
53. Jia D, Jolly MK, Boareto M, Parsana P, Mooney SM, Pienta KJ, et al. OVOL guides the epithelial-hybrid-mesenchymal transition. Oncotarget (2015) 6:15436–48.
54. Vannier C, Mock K, Brabletz T, Driever W. Zeb1 regulates E-cadherin and Epcam (epithelial cell adhesion molecule) expression to control cell behavior in early zebrafish development. J Biol Chem (2013) 288:18643–59. doi:10.1074/jbc.M113.467787
55. Kim NH, Kim HS, Li XY, Lee I, Choi HS, Kang SE, et al. A p53/miRNA-34 axis regulates Snail1-dependent cancer cell epithelial-mesenchymal transition. J Cell Biol (2011) 195:417–33. doi:10.1083/jcb.201103097
56. Gregory PA, Bert AG, Paterson EL, Barry SC, Tsykin A, Farshid G, et al. The miR-200 family and miR-205 regulate epithelial to mesenchymal transition by targeting ZEB1 and SIP1. Nat Cell Biol (2008) 10:593–601. doi:10.1038/ncb1722
57. Peiró S, Escrivà M, Puig I, Barberà MJ, Dave N, Herranz N, et al. Snail1 transcriptional repressor binds to its own promoter and controls its expression. Nucleic Acids Res (2006) 34:2077–84. doi:10.1093/nar/gkl141
58. Hill L, Browne G, Tulchinsky E. ZEB/miR-200 feedback loop: at the crossroads of signal transduction in cancer. Int J Cancer (2013) 132:745–54. doi:10.1002/ijc.27708
59. Bintu L, Buchler NE, Garcia HG, Gerland U, Hwa T, Kondev J, et al. Transcriptional regulation by the numbers: models. Curr Opin Genet Dev (2005) 15:116–24. doi:10.1016/j.gde.2005.02.007
60. Nyayanit D, Gadgil CJ. Mathematical modeling of combinatorial regulation suggests that apparent positive regulation of targets by miRNA could be an artifact resulting from competition for mRNA. RNA (2015) 21:307–19. doi:10.1261/rna.046862.114
61. Mitarai N, Benjamin JA, Krishna S, Semsey S, Csiszovszki Z, Massé E, et al. Dynamic features of gene expression control by small regulatory RNAs. Proc Natl Acad Sci U S A (2009) 106:10655–9. doi:10.1073/pnas.0901466106
62. Zhou P, Liu Z, Cai S, Wang R. Mechanisms generating bistability and oscillations in microRNA-mediated motifs. Phys Rev E Stat Nonlin Soft Matter Phys (2012) 85(4 Pt 1):041916. doi:10.1103/PhysRevE.85.041916
63. Levine E, Ben Jacob E, Levine H. Target-specific and global effectors in gene regulation by MicroRNA. Biophys J (2007) 93:L52–4. doi:10.1529/biophysj.107.118448
64. Schliekelman MJ, Taguchi A, Zhu J, Dai X, Rodriguez J, Celiktas M, et al. Molecular portraits of epithelial, mesenchymal and hybrid states in lung adenocarcinoma and their relevance to survival. Cancer Res (2015) 75:1789–800. doi:10.1158/0008-5472.CAN-14-2535
65. Keller PJ, Lin AF, Arendt LM, Klebba I, Jones AD, Rudnick JA, et al. Mapping the cellular and molecular heterogeneity of normal and malignant breast tissues and cultured cell lines. Breast Cancer Res (2010) 12:R87. doi:10.1186/bcr2755
66. Goldman A, Majumder B, Dhawan A, Ravi S, Goldman D, Kohandel M, et al. Temporally sequenced anticancer drugs overcome adaptive resistance by targeting a vulnerable chemotherapy-induced phenotypic transition. Nat Commun (2015) 6:6139. doi:10.1038/ncomms7139
67. Chéry L, Lam HM, Coleman I, Lakely B, Coleman R, Larson S, et al. Characterization of single disseminated prostate cancer cells reveals tumor cell heterogeneity and identifies dormancy associated pathways. Oncotarget (2014) 5:9939–51.
68. Polioudaki H, Agelaki S, Chiotaki R, Politaki E, Mavroudis D, Matikas A, et al. Variable expression levels of keratin and vimentin reveal differential EMT status of circulating tumor cells and correlation with clinical characteristics and outcome of patients with metastatic breast cancer. BMC Cancer (2015) 15:399. doi:10.1186/s12885-015-1386-7
69. Frank SA, Rosner MR. Nonheritable cellular variability accelerates the evolutionary processes of cancer. PLoS Biol (2012) 10:e1001296. doi:10.1371/journal.pbio.1001296
70. Tan TZ, Miow QH, Miki Y, Noda T, Mori S, Huang RY, et al. Epithelial-mesenchymal transition spectrum quantification and its efficacy in deciphering survival and drug responses of cancer patients. EMBO Mol Med (2014) 6:1279–93. doi:10.15252/emmm.201404208
71. Bill R, Christofori G. The relevance of EMT in breast cancer metastasis: correlation or causality? FEBS Lett (2015) 589(14):1577–87. doi:10.1016/j.febslet.2015.05.002
72. Jordan NV, Johnson GL, Abell AN. Tracking the intermediate stages of epithelial-mesenchymal transition in epithelial stem cells and cancer. Cell Cycle (2011) 10:2865–73. doi:10.4161/cc.10.17.17188
73. Yaswen P. Reinforcing targeted therapeutics with phenotypic stability factors. Cell Cycle (2015) 13:3818–22. doi:10.4161/15384101.2014.985071
74. Huang B, Lu M, Jolly MK, Tsarfaty I, Onuchic J, Ben-Jacob E. The three-way switch operation of Rac1/RhoA GTPase-based circuit controlling amoeboid-hybrid-mesenchymal transition. Sci Rep (2014) 4:6449. doi:10.1038/srep06449
75. Paňková K, Rösel D, Novotný M, Brábek J. The molecular mechanisms of transition between mesenchymal and amoeboid invasiveness in tumor cells. Cell Mol Life Sci (2010) 67:63–71. doi:10.1007/s00018-009-0132-1
76. Ben-Jacob E, Coffey DS, Levine H. Bacterial survival strategies suggest rethinking cancer cooperativity. Trends Microbiol (2012) 20:403–10. doi:10.1016/j.tim.2012.06.001
77. Bergert M, Chandradoss SD, Desai RA, Paluch E. Cell mechanics control rapid transitions between blebs and lamellipodia during migration. Proc Natl Acad Sci U S A (2012) 109:14434–9. doi:10.1073/pnas.1207968109
78. Chao Y, Wu Q, Acquafondata M, Dhir R, Wells A. Partial mesenchymal to epithelial reverting transition in breast and prostate cancer metastases. Cancer Microenviron (2011) 5:19–28. doi:10.1007/s12307-011-0085-4
79. Friedl P, Wolf K. Plasticity of cell migration: a multiscale tuning model. J Cell Biol (2010) 188:11–9. doi:10.1083/jcb.200909003
80. Wolf K, Wu YI, Liu Y, Geiger J, Tam E, Overall C, et al. Multi-step pericellular proteolysis controls the transition from individual to collective cancer cell invasion. Nat Cell Biol (2007) 9:893–904. doi:10.1038/ncb1616
81. Hegerfeldt Y, Tusch M, Bröcker E-B, Friedl P. Collective cell movement in primary melanoma explants: plasticity of cell-cell interaction, beta1-integrin function, and migration strategies. Cancer Res (2002) 62:2125–30.
82. Hecht I, Bar-El Y, Balmer F, Natan S, Tsarfaty I, Schweitzer F, et al. Tumor invasion optimization by mesenchymal-amoeboid heterogeneity. Sci Rep (2015) 5:10622. doi:10.1038/srep10622
83. Parri M, Chiarugi P. Rac and Rho GTPases in cancer cell motility control. Cell Commun Signal (2010) 8:23. doi:10.1186/1478-811X-8-23
84. Wang Z, Humphries B, Xiao H, Jiang Y, Yang C. MicroRNA-200b suppresses arsenic-transformed cell migration by targeting protein kinase Cα and Wnt5b-protein kinase Cα positive feedback loop and inhibiting Rac1 activation. J Biol Chem (2014) 289:18373–86. doi:10.1074/jbc.M114.554246
85. Kim D, Kim S, Jin E-J, Park HM, Sonn J, Song J, et al. MicroRNA-34a modulates cytoskeletal dynamics through regulating RhoA/Rac1 cross-talk in chondroblasts. J Biol Chem (2012) 287:12501–9. doi:10.1074/jbc.M111.264382
86. Elson-Schwab I, Lorentzen A, Marshall CJ. MicroRNA-200 family members differentially regulate morphological plasticity and mode of melanoma cell invasion. PLoS One (2010) 5:e13176. doi:10.1371/journal.pone.0013176
87. Sailem H, Bousgouni V, Cooper S, Bakal C. Cross-talk between Rho and Rac GTPases drives deterministic exploration of cellular shape space and morphological heterogeneity. Open Biol (2014) 4:130132. doi:10.1098/rsob.130132
88. Jilkine A, Marée AF, Edelstein-Keshet L. Mathematical model for spatial segregation of the Rho-family GTPases based on inhibitory crosstalk. Bull Math Biol (2007) 69:1943–78. doi:10.1007/s11538-007-9200-6
89. Chauhan BK, Lou M, Zheng Y, Lang RA. Balanced Rac1 and RhoA activities regulate cell shape and drive invagination morphogenesis in epithelia. Proc Natl Acad Sci U S A (2011) 108:18289–94. doi:10.1073/pnas.1108993108
90. Futterman MA, García AJ, Zamir EA. Evidence for partial epithelial-to-mesenchymal transition (pEMT) and recruitment of motile blastoderm edge cells during avian epiboly. Dev Dyn (2011) 240:1502–11. doi:10.1002/dvdy.22607
91. Johnen N, Francart M-E, Thelen N, Cloes M, Thiry M. Evidence for a partial epithelial-mesenchymal transition in postnatal stages of rat auditory organ morphogenesis. Histochem Cell Biol (2012) 138:477–88. doi:10.1007/s00418-012-0969-5
92. Ribeiro AS, Paredes J. P-cadherin linking breast cancer stem cells and invasion: a promising marker to identify an “intermediate/metastable” EMT state. Front Oncol (2015) 4:371. doi:10.3389/fonc.2014.00371
93. Welch-Reardon KM, Ehsan SM, Wang K, Wu N, Newman AC, Romero-Lopez M, et al. Angiogenic sprouting is regulated by endothelial cell expression of slug. J Cell Sci (2014) 127:2017–28. doi:10.1242/jcs.143420
94. Leopold PL, Vincent J, Wang H. A comparison of epithelial-to-mesenchymal transition and re-epithelialization. Semin Cancer Biol (2012) 22:471–83. doi:10.1016/j.semcancer.2012.07.003
95. Yan C, Grimm WA, Garner WL, Qin L, Travis T, Tan N, et al. Epithelial to mesenchymal transition in human skin wound healing is induced by tumor necrosis factor-alpha through bone morphogenic protein-2. Am J Pathol (2010) 176:2247–58. doi:10.2353/ajpath.2010.090048
96. Leroy P, Mostov KE. Slug is required for cell survival during partial epithelial-mesenchymal transition of HGF-induced tubulogenesis. Mol Biol Cell (2007) 18:1943–52. doi:10.1091/mbc.E06-09-0823
97. Hudson LG, Newkirk KM, Chandler HL, Choi C, Fossey SL, Parent AE, et al. Cutaneous wound reepithelialization is compromised in mice lacking functional Slug (Snai2). J Dermatol Sci (2009) 56:19–26. doi:10.1016/j.jdermsci.2009.06.009
98. Niessen K, Fu Y, Chang L, Hoodless PA, McFadden D, Karsan A. Slug is a direct Notch target required for initiation of cardiac cushion cellularization. J Cell Biol (2008) 182:315–25. doi:10.1083/jcb.200710067
99. Lee K, Gjorevski N, Boghaert E, Radisky DC, Nelson CM. Snail1, Snail2, and E47 promote mammary epithelial branching morphogenesis. EMBO J (2011) 30:2662–74. doi:10.1038/emboj.2011.159
100. Savagner P, Kusewitt DF, Carver EA, Magnino F, Choi C, Gridley T, et al. Developmental transcription factor slug is required for effective re-epithelialization by adult keratinocytes. J Cell Physiol (2005) 202:858–66. doi:10.1002/jcp.20188
101. Friedl P, Locker J, Sahai E, Segall JE. Classifying collective cancer cell invasion. Nat Cell Biol (2012) 14:777–83. doi:10.1038/ncb2548
102. Strauss R, Sova P, Liu Y, Zong YL, Tuve S, Pritchard D, et al. Epithelial phenotype confers resistance of ovarian cancer cells to oncolytic adenoviruses. Cancer Res (2009) 69:5115–25. doi:10.1158/0008-5472.CAN-09-0645
103. Huang RY, Wong MK, Tan TZ, Kuay KT, Nga HC, Chung VY, et al. An EMT spectrum defines an anoikis-resistant and spheroidogenic intermediate mesenchymal state that is sensitive to e-cadherin restoration by a src-kinase inhibitor, saracatinib (AZD0530). Cell Death Dis (2013) 4:e915. doi:10.1038/cddis.2013.442
104. Sampson VB, David JM, Puig I, Patil PU, de Herreros AG, Thomas GV, et al. Wilms’ tumor protein induces an epithelial-mesenchymal hybrid differentiation state in clear cell renal cell carcinoma. PLoS One (2014) 9:e102041. doi:10.1371/journal.pone.0102041
105. Rhim AD, Mirek ET, Aiello NM, Maitra A, Jennifer M, McAllister F, et al. EMT and dissemination precede pancreatic tumor formation. Cell (2013) 148:349–61. doi:10.1016/j.cell.2011.11.025
106. Ruscetti M, Quach B, Dadashian EL, Mulholland DJ, Hong W. Tracking and functional characterization of epithelial-mesenchymal transition and mesenchymal tumor cells during prostate cancer metastasis. Cancer Res (2015) 75(13):2749–59. doi:10.1158/0008-5472.CAN-14-3476
107. Sarrio D, Rodriguez-Pinilla SM, Hardisson D, Cano A, Moreno-Bueno G, Palacios J. Epithelial-mesenchymal transition in breast cancer relates to the basal-like phenotype. Cancer Res (2008) 68:989–97. doi:10.1158/0008-5472.CAN-07-2017
108. Prat A, Parker JS, Karginova O, Fan C, Livasy C, Herschkowitz JI, et al. Phenotypic and molecular characterization of the claudin-low intrinsic subtype of breast cancer. Breast Cancer Res (2010) 12:R68. doi:10.1186/bcr2635
109. Hendrix MJ, Seftor EA, Seftor RE, Trevor KT. Experimental co-expression of vimentin and keratin intermediate filaments in human breast cancer cells results in phenotypic interconversion and increased invasive behavior. Am J Pathol (1997) 150:483–95.
110. Thomas PA, Kirschmann DA, Cerhan JR, Folberg R, Seftor EA, Sellers TA, et al. Association between keratin and vimentin expression, malignant phenotype, and survival in postmenopausal breast cancer patients. Clin Cancer Res (1999) 5:2698–703.
111. Ramaekers FC, Haag D, Kant A, Moesker O, Jap PH, Vooijs GP. Coexpression of keratin- and vimentin-type intermediate filaments in human metastatic carcinoma cells. Proc Natl Acad Sci U S A (1983) 80:2618–22. doi:10.1073/pnas.80.9.2618
112. Hendrix MJ, Seftor EA, Chu Y, Seftor RE, Nagle RB, McDaniel KM, et al. Coexpression of vimentin and keratins by human melanoma tumor cells: correlation with invasive and metastatic potential. J Natl Cancer Inst (1992) 84:165–74. doi:10.1093/jnci/84.3.165
113. Livasy CA, Karaca G, Nanda R, Tretiakova MS, Olopade OI, Moore DT, et al. Phenotypic evaluation of the basal-like subtype of invasive breast carcinoma. Mod Pathol (2006) 19:264–71. doi:10.1038/modpathol.3800528
114. Wu S, Liu S, Liu Z, Huang J, Pu X, Li J, et al. Classification of circulating tumor cells by epithelial-mesenchymal transition markers. PLoS One (2015) 10:e0123976. doi:10.1371/journal.pone.0123976
115. Hou JM, Krebs MG, Lancashire L, Sloane R, Backen A, Swain RK, et al. Clinical significance and molecular characteristics of circulating tumor cells and circulating tumor microemboli in patients with small-cell lung cancer. J Clin Oncol (2012) 30:525–32. doi:10.1200/JCO.2010.33.3716
116. Ilie M, Hofman V, Long-Mira E, Selva E, Vignaud J-M, Padovani B, et al. “Sentinel” circulating tumor cells allow early diagnosis of lung cancer in patients with chronic obstructive pulmonary disease. PLoS One (2014) 9:e111597. doi:10.1371/journal.pone.0111597
117. Liotta LA, Klelnerman J, Saldel GM. The significance of hematogenous tumor cell clumps in the metastatic process. Cancer Res (1976) 36:889–94.
118. Fidler IJ. The relationship of embolic homogeneity, number, size and viability to the incidence of experimental metastasis. Eur J Cancer (1973) 9:223–7. doi:10.1016/S0014-2964(73)80022-2
119. Hugo H, Ackland LM, Blick T, Lawrence MG, Clementa JA, Williams ED, et al. Epithelial-mesenchymal and mesenchymal-epithelial transitions in carcinoma progression. J Cell Physiol (2007) 213:374–83. doi:10.1002/jcp.21223
120. Alix-Panabières C, Pantel K. Challenges in circulating tumour cell research. Nat Rev Cancer (2014) 14:623. doi:10.1038/nrc3820
121. Ting DT, Wittner BS, Ligorio M, Vincent Jordan N, Shah AM, Miyamoto DT, et al. Single-cell RNA sequencing identifies extracellular matrix gene expression by pancreatic circulating tumor cells. Cell Rep (2014) 8(6):1905–18. doi:10.1016/j.celrep.2014.08.029
122. Baccelli I, Schneeweiss A, Riethdorf S, Stenzinger A, Schillert A, Vogel V, et al. Identification of a population of blood circulating tumor cells from breast cancer patients that initiates metastasis in a xenograft assay. Nat Biotechnol (2013) 31:539–44. doi:10.1038/nbt.2576
123. Cayrefourcq L, Mazard T, Joosse S, Solassol J, Ramos J, Assenat E, et al. Establishment and characterization of a cell line from human circulating colon cancer cells. Cancer Res (2015) 75:892–902. doi:10.1158/0008-5472.CAN-14-2613
124. Strauss R, Li Z-Y, Liu Y, Beyer I, Persson J, Sova P, et al. Analysis of epithelial and mesenchymal markers in ovarian cancer reveals phenotypic heterogeneity and plasticity. PLoS One (2011) 6:e16186. doi:10.1371/journal.pone.0016186
125. Plaks V, Kong N, Werb Z. The cancer stem cell niche: how essential is the niche in regulating stemness of tumor cells? Cell Stem Cell (2015) 16:225–38. doi:10.1016/j.stem.2015.02.015
126. Gupta PB, Fillmore CM, Jiang G, Shapira SD, Tao K, Kuperwasser C, et al. Stochastic state transitions give rise to phenotypic equilibrium in populations of cancer cells. Cell (2011) 146:633–44. doi:10.1016/j.cell.2011.07.026
127. Yang G, Quan Y, Wang W, Fu Q, Wu J, Mei T, et al. Dynamic equilibrium between cancer stem cells and non-stem cancer cells in human SW620 and MCF-7 cancer cell populations. Br J Cancer (2012) 106:1512–9. doi:10.1038/bjc.2012.126
128. Iliopoulos D, Hirsch HA, Wang G, Struhl K. Inducible formation of breast cancer stem cells and their dynamic equilibrium with non-stem cancer cells via IL6 secretion. Proc Natl Acad Sci U S A (2011) 108:1397–402. doi:10.1073/pnas.1018898108
129. Enderling H. Cancer stem cells: small subpopulation or evolving fraction? Integr Biol (Camb) (2015) 7:14–23. doi:10.1039/c4ib00191e
130. Csermely P, Hódsági J, Korcsmáros T, Módos D, Perez-Lopez ÁR, Szalay K, et al. Cancer stem cells display extremely large evolvability: alternating plastic and rigid networks as a potential mechanism network models, novel therapeutic target strategies, and the contributions of hypoxia, inflammation and cellular senescence. Semin Cancer Biol (2015) 30:42–51. doi:10.1016/j.semcancer.2013.12.004
131. Mani SA, Guo W, Liao M-J, Eaton EN, Ayyanan A, Zhou AY, et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell (2008) 133:704–15. doi:10.1016/j.cell.2008.03.027
132. Morel A-P, Lièvre M, Thomas C, Hinkal G, Ansieau S, Puisieux A. Generation of breast cancer stem cells through epithelial-mesenchymal transition. PLoS One (2008) 3:e2888. doi:10.1371/journal.pone.0002888
133. Scheel C, Weinberg RA. Cancer stem cells and epithelial-mesenchymal transition: concepts and molecular links. Semin Cancer Biol (2012) 22:396–403. doi:10.1016/j.semcancer.2012.04.001
134. Ombrato L, Malanchi I. The EMT universe: space between cancer cell dissemination and metastasis initiation. Crit Rev Oncog (2014) 19:349–61. doi:10.1615/CritRevOncog.2014011802
135. Celià-Terrassa T, Meca-Cortés Ó, Mateo F, De Paz AM, Rubio N, Arnal-Estapé A, et al. Epithelial-mesenchymal transition can suppress major attributes of human epithelial tumor-initiating cells. J Clin Invest (2012) 122:1849–68. doi:10.1172/JCI59218
136. Ocaña OH, Córcoles R, Fabra A, Moreno-Bueno G, Acloque H, Vega S, et al. Metastatic colonization requires the repression of the epithelial-mesenchymal transition inducer Prrx1. Cancer Cell (2012) 22:709–24. doi:10.1016/j.ccr.2012.10.012
137. Tsai JH, Donaher JL, Murphy DA, Chau S, Yang J. Spatiotemporal regulation of epithelial-mesenchymal transition is essential for squamous cell carcinoma metastasis. Cancer Cell (2012) 22:725–36. doi:10.1016/j.ccr.2012.09.022
138. Tsuji T, Ibaragi S, Shima K, Hu MG, Katsurano M, Sasaki A, et al. Epithelial-mesenchymal transition induced by growth suppressor p12 CDK2-AP1 promotes tumor cell local invasion but suppresses distant colony growth. Cancer Res (2008) 68:10377–86. doi:10.1158/0008-5472.CAN-08-1444
139. Samavarchi-Tehrani P, Golipour A, David L, Sung HK, Beyer TA, Datti A, et al. Functional genomics reveals a BMP-driven mesenchymal-to-epithelial transition in the initiation of somatic cell reprogramming. Cell Stem Cell (2010) 7:64–77. doi:10.1016/j.stem.2010.04.015
140. Li R, Liang J, Ni S, Zhou T, Qing X, Li H, et al. A mesenchymal-to-epithelial transition initiates and is required for the nuclear reprogramming of mouse fibroblasts. Cell Stem Cell (2010) 7:51–63. doi:10.1016/j.stem.2010.04.014
141. Jolly MK, Huang B, Lu M, Mani SA, Levine H, Ben-Jacob E. Towards elucidating the connection between epithelial − mesenchymal transitions and stemness. J R Soc Interface (2014) 11:20140962. doi:10.1098/rsif.2014.0962
142. Yovchev MI, Grozdanov PN, Zhou H, Racherla H, Guha C, Dabeva MD. Identification of adult hepatic progenitor cells capable of repopulating injured rat liver. Hepatology (2008) 47:636–47. doi:10.1002/hep.22047
143. Conigliaro A, Amicone L, Costa V, De Santis Puzzonia M, Mancone C, Sacchetti B, et al. Evidence for a common progenitor of epithelial and mesenchymal components of the liver. Cell Death Differ (2013) 20:1116–23. doi:10.1038/cdd.2013.49
144. Cicchini C, Amicone L, Alonzi T, Marchetti A, Mancone C, Tripodi M. Molecular mechanisms controlling the phenotype and the EMT/MET dynamics of hepatocyte. Liver Int (2015) 35:302–10. doi:10.1111/liv.12577
145. Choi SS, Diehl AM. Epithelial-to-mesenchymal transitions in the liver. Hepatology (2009) 50:2007–13. doi:10.1002/hep.23196
146. Swetha G, Chandra V, Phadnis S, Bhonde R. Glomerular parietal epithelial cells of adult murine kidney undergo EMT to generate cells with traits of renal progenitors. J Cell Mol Med (2011) 15:396–413. doi:10.1111/j.1582-4934.2009.00937.x
147. Tam WL, Weinberg RA. The epigenetics of epithelial-mesenchymal plasticity in cancer. Nat Med (2013) 19:1438–49. doi:10.1038/nm.3336
148. Das R, Gregory PA, Hollier BG, Tilley WD, Selth LA. Epithelial plasticity in prostate cancer: principles and clinical perspectives. Trends Mol Med (2014) 20:643–51. doi:10.1016/j.molmed.2014.09.004
149. Ansieau S, Collin G, Hill L. EMT or EMT-promoting transcription factors, where to focus the light? Front Oncol (2014) 4:353. doi:10.3389/fonc.2014.00353
150. Strauss R, Hamerlik P, Lieber A, Bartek J. Regulation of stem cell plasticity: mechanisms and relevance to tissue biology and cancer. Mol Ther (2012) 20:887–97. doi:10.1038/mt.2012.2
151. Liu S, Cong Y, Wang D, Sun Y, Deng L, Liu Y, et al. Breast cancer stem cells transition between epithelial and mesenchymal states reflective of their normal counterparts. Stem Cell Reports (2014) 2:78–91. doi:10.1016/j.stemcr.2013.11.009
152. Biddle A, Liang X, Gammon L, Fazil B, Harper LJ, Emich H, et al. Cancer stem cells in squamous cell carcinoma switch between two distinct phenotypes that are preferentially migratory or proliferative. Cancer Res (2011) 71:5317–26. doi:10.1158/0008-5472.CAN-11-1059
153. Brabletz T. To differentiate or not – routes towards metastasis. Nat Rev Cancer (2012) 12:425–36. doi:10.1038/nrc3265
154. Barriere G, Fici P, Gallerani G, Fabbri F, Rigaud M. Epithelial mesenchymal transition: a double-edged sword. Clin Transl Med (2015) 4:4–9. doi:10.1186/s40169-015-0055-4
155. Li C, Heidt DG, Dalerba P, Burant CF, Zhang L, Adsay V, et al. Identification of pancreatic cancer stem cells. Cancer Res (2007) 67:1030–7. doi:10.1158/0008-5472.CAN-06-2030
156. Zhang C, Li C, He F, Cai Y, Yang H. Identification of CD44+CD24+ gastric cancer stem cells. J Cancer Res Clin Oncol (2011) 137:1679–86. doi:10.1007/s00432-011-1038-5
157. Uccelli A, Moretta L, Pistoia V. Mesenchymal stem cells in health and disease. Nat Rev Immunol (2008) 8:726–36. doi:10.1038/nri2395
158. Sleeman KE, Kendrick H, Ashworth A, Isacke CM, Smalley MJ. CD24 staining of mouse mammary gland cells defines luminal epithelial, myoepithelial/basal and non-epithelial cells. Breast Cancer Res (2006) 8:R7. doi:10.1186/bcr1550
159. Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF. Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci U S A (2003) 100:3983–8. doi:10.1073/pnas.0530291100
160. Fillmore CM, Kuperwasser C. Human breast cancer cell lines contain stem-like cells that self-renew, give rise to phenotypically diverse progeny and survive chemotherapy. Breast Cancer Res (2008) 10:R25. doi:10.1186/bcr1982
161. Azzam DJ, Zhao D, Sun J, Minn AJ, Ranganathan P, Drews-Elger K, et al. Triple negative breast cancer initiating cell subsets differ in functional and molecular characteristics and in γ-secretase inhibitor drug responses. EMBO Mol Med (2013) 5:1502–22. doi:10.1002/emmm.201302558
162. Bhat-Nakshatri P, Appaiah H, Ballas C, Pick-Franke P, Goulet R, Badve S, et al. SLUG/SNAI2 and tumor necrosis factor generate breast cells with CD44+/CD24- phenotype. BMC Cancer (2010) 10:411. doi:10.1186/1471-2407-10-411
163. Yang C-H, Wang H-L, Lin Y-S, Kumar KP, Lin H-C, Chang C-J, et al. Identification of CD24 as a cancer stem cell marker in human nasopharyngeal carcinoma. PLoS One (2014) 9:e99412. doi:10.1371/journal.pone.0099412
164. Giannoni E, Bianchini F, Masieri L, Serni S, Torre E, Calorini L, et al. Reciprocal activation of prostate cancer cells and cancer-associated fibroblasts stimulates epithelial-mesenchymal transition and cancer stemness. Cancer Res (2010) 70:6945–56. doi:10.1158/0008-5472.CAN-10-0785
165. Yang KR, Mooney SM, Zarif JC, Coffey DS, Taichman RS, Pienta KJ. Niche inheritance: a cooperative pathway to enhance cancer cell fitness though ecosystem engineering. J Cell Biochem (2014) 115:1478–85. doi:10.1002/jcb.24813
166. Chen W-J, Ho C-C, Chang Y-L, Chen HY, Lin CA, Ling T-Y, et al. Cancer-associated fibroblasts regulate the plasticity of lung cancer stemness via paracrine signalling. Nat Commun (2014) 5:3472. doi:10.1038/ncomms4472
167. Sanità P, Capulli M, Teti A, Galatioto GP, Vicentini C, Chiarugi P, et al. Tumor-stroma metabolic relationship based on lactate shuttle can sustain prostate cancer progression. BMC Cancer (2014) 14:154. doi:10.1186/1471-2407-14-154
168. Duda DG, Duvyerman AMMJ, Kohno M, Snuderi M, Snuderi M, Steller EJ, et al. Malignant cells facilitate lung metastasis by bringing their own soil. Proc Natl Acad Sci U S A (2007) 107:21677–82. doi:10.1073/pnas.1016234107
169. De Herreros AG. Epithelial to mesenchymal transition in tumor cells as consequence of phenotypic instability. Front Cell Dev Biol (2014) 2:71. doi:10.3389/fcell.2014.00071
170. Espinoza I, Miele L. Deadly crosstalk: Notch signaling at the intersection of EMT and cancer stem cells. Cancer Lett (2013) 341:41–5. doi:10.1016/j.canlet.2013.08.027
171. Li D, Masiero M, Banham AH, Harris AL. The Notch ligand JAGGED1 as a target for anti-tumor therapy. Front Oncol (2014) 4:254. doi:10.3389/fonc.2014.00254
172. Boareto M, Jolly MK, Lu M, Onuchic JN, Clementi C, Ben-Jacob E. Jagged-delta asymmetry in Notch signaling can give rise to a sender/receiver hybrid phenotype. Proc Natl Acad Sci U S A (2015) 112:E402–9. doi:10.1073/pnas.1416287112
173. Jolly MK, Boareto M, Lu M, Onuchic JN, Clementi C, Ben-Jacob E. Operating principles of Notch-delta-jagged module of cell-cell communication. New J Phys (2015) 17:055021. doi:10.1088/1367-2630/17/5/055021
174. Lu J, Ye X, Fan F, Xia L, Bhattacharya R, Bellister S, et al. Endothelial cells promote the colorectal cancer stem cell phenotype through a soluble form of jagged-1. Cancer Cell (2013) 23:171–85. doi:10.1016/j.ccr.2012.12.021
175. Yamamoto M, Taguchi Y, Ito-Kureha T, Semba K, Yamaguchi N, Inoue J. NF-κB non-cell-autonomously regulates cancer stem cell populations in the basal-like breast cancer subtype. Nat Commun (2013) 4:2299. doi:10.1038/ncomms3299
176. Sethi N, Dai X, Winter CG, Kang Y. Tumor-derived JAGGED1 promotes osteolytic bone metastasis of breast cancer by engaging Notch signaling in bone cells. Cancer Cell (2011) 19:192–205. doi:10.1016/j.ccr.2010.12.022
177. Sansone P, Storci G, Tavolari S, Guarnieri T, Giovannini C, Taffurelli M, et al. IL-6 triggers malignant features in mammospheres from human ductal breast carcinoma and normal mammary gland. J Clin Invest (2007) 117:3988–4002. doi:10.1172/JCI32533
178. Shaya O, Sprinzak D. From Notch signaling to fine-grained patterning: modeling meets experiments. Curr Opin Genet Dev (2011) 21:732–9. doi:10.1016/j.gde.2011.07.007
179. Petrovic J, Formosa-Jordan P, Luna-Escalante JC, Abelló G, Ibañes M, Neves J, et al. Ligand-dependent Notch signaling strength orchestrates lateral induction and lateral inhibition in the developing inner ear. Development (2014) 141:2313–24. doi:10.1242/dev.108100
180. Owen MR, Sherratt JA, Wearing HJ. Lateral induction by juxtacrine signaling is a new mechanism for pattern formation. Dev Biol (2000) 217:54–61. doi:10.1006/dbio.1999.9536
181. Hartman BH, Reh TA, Bermingham-McDonogh O. Notch signaling specifies prosensory domains via lateral induction in the developing mammalian inner ear. Proc Natl Acad Sci U S A (2010) 107:15792–7. doi:10.1073/pnas.1002827107
182. Chigurupati S, Arumugam TV, Son TG, Lathia JD, Jameel S, Mughal MR, et al. Involvement of Notch signaling in wound healing. PLoS One (2007) 2:e1167. doi:10.1371/journal.pone.0001167
183. Brabletz S, Bajdak K, Meidhof S, Burk U, Niedermann G, Firat E, et al. The ZEB1/miR-200 feedback loop controls Notch signalling in cancer cells. EMBO J (2011) 30:770–82. doi:10.1038/emboj.2010.349
184. De Antonellis P, Medaglia C, Cusanelli E, Andolfo I, Liguori L, De Vita G, et al. MiR-34a targeting of Notch ligand delta-like 1 impairs CD15+/CD133+ tumor-propagating cells and supports neural differentiation in medulloblastoma. PLoS One (2011) 6:e24584. doi:10.1371/journal.pone.0024584
185. Bu P, Chen K-Y, Chen JH, Wang L, Walters J, Shin YJ, et al. A microRNA miR-34a-regulated bimodal switch targets Notch in colon cancer stem cells. Cell Stem Cell (2013) 12:602–15. doi:10.1016/j.stem.2013.03.002
186. Sahlgren C, Gustafsson MV, Jin S, Poellinger L, Lendahl U. Notch signaling mediates hypoxia-induced tumor cell migration and invasion. Proc Natl Acad Sci U S A (2008) 105:6392–7. doi:10.1073/pnas.0802047105
187. Duluc D, Delneste Y, Tan F, Moles M-P, Grimaud L, Lenoir J, et al. Tumor-associated leukemia inhibitory factor and IL-6 skew monocyte differentiation into tumor-associated macrophage-like cells. Blood (2007) 110:4319–30. doi:10.1182/blood-2007-02-072587
188. Johnston DA, Dong B, Hughes CC. TNF induction of jagged-1 in endothelial cells is NFkappaB-dependent. Gene (2009) 435:36–44. doi:10.1016/j.gene.2009.01.003
189. Benedito R, Roca C, Sörensen I, Adams S, Gossler A, Fruttiger M, et al. The Notch ligands Dll4 and jagged1 have opposing effects on angiogenesis. Cell (2009) 137:1124–35. doi:10.1016/j.cell.2009.03.025
190. Colotta F, Allavena P, Sica A, Garlanda C, Mantovani A. Cancer-related inflammation, the seventh hallmark of cancer: links to genetic instability. Carcinogenesis (2009) 30:1073–81. doi:10.1093/carcin/bgp127
191. McGuire MF, Enderling H, Wallace DI, Batra J, Jordan M, Kumar S, et al. Formalizing an integrative, multidisciplinary cancer therapy discovery workflow. Cancer Res (2013) 73:6111–7. doi:10.1158/0008-5472.CAN-13-0310
192. Enderling H, Hahnfeldt P, Hlatky L, Almog N. Systems biology of tumor dormancy: linking biology and mathematics on multiple scales to improve cancer therapy. Cancer Res (2012) 72:2172–5. doi:10.1158/0008-5472.CAN-11-3269
193. Marchini S, Fruscio R, Clivio L, Beltrame L, Porcu L, Nerini IF, et al. Resistance to platinum-based chemotherapy is associated with epithelial to mesenchymal transition in epithelial ovarian cancer. Eur J Cancer (2012) 49:520–30. doi:10.1016/j.ejca.2012.06.026
194. Arumugam T, Ramachandran V, Fournier KF, Wang H, Marquis L, Abbruzzese JL, et al. Epithelial to mesenchymal transition contributes to drug resistance in pancreatic cancer. Cancer Res (2009) 69:5820–8. doi:10.1158/0008-5472.CAN-08-2819
195. Witta SE, Gemmill RM, Hirsch FR, Coldren CD, Hedman K, Ravdel L, et al. Restoring E-cadherin expression increases sensitivity to epidermal growth factor receptor inhibitors in lung cancer cell lines. Cancer Res (2006) 66:944–50. doi:10.1158/0008-5472.CAN-05-1988
196. May CD, Sphyris N, Evans KW, Werden SJ, Guo W, Mani SA. Epithelial-mesenchymal transition and cancer stem cells: a dangerously dynamic duo in breast cancer progression. Breast Cancer Res (2011) 13:202. doi:10.1186/bcr2789
197. André F, Zielinski CC. Optimal strategies for the treatment of metastatic triple-negative breast cancer with currently approved agents. Ann Oncol (2012) 23:vi46–51. doi:10.1093/annonc/mds195
198. Vadgama JV, Wu Y, Ginther C, Kim J, Mosher N, Chung S, et al. Expression of Wnt3 activates Wnt/β-catenin pathway and promotes EMT-like phenotype in trastuzumab resistant HER2-overexpressing breast cancer cells. Mol Cancer Res (2012) 10:1597–606. doi:10.1158/1541-7786.MCR-12-0155-T
199. Hiscox S, Jiang WG, Obermeier K, Taylor K, Morgan L, Burmi R, et al. Tamoxifen resistance in MCF7 cells promotes EMT-like behaviour and involves modulation of β-catenin phosphorylation. Int J Cancer (2006) 118:290–301. doi:10.1002/ijc.21355
200. Bastos LGDR, de Marcondes PG, de-Freitas-Junior JC, Leve F, Mencalha AL, de Souza WF, et al. Progeny from irradiated colorectal cancer cells acquire an EMT-like phenotype and activate Wnt/β-catenin pathway. J Cell Biochem (2014) 115:2175–87. doi:10.1002/jcb.24896
201. Nieto MA, Cano A. The epithelial – mesenchymal transition under control: global programs to regulate epithelial plasticity. Semin Cancer Biol (2012) 22:361–8. doi:10.1016/j.semcancer.2012.05.003
202. Dvorak H. Tumors: wounds that do not heal. N Engl J Med (1986) 315:1650–9. doi:10.1056/NEJM198612253152606
203. Zhu H, Bhaijee F, Ishaq N, Pepper DJ, Backus K, Brown AS, et al. Correlation of Notch1, pAKT and nuclear NF-κB expression in triple negative breast cancer. Am J Cancer Res (2013) 3:230–9.
204. Schaue D, Micewicz ED, Ratikan JA, Xie MW, Cheng G, McBride WH. Radiation and inflammation. Semin Radiat Oncol (2015) 25:4–10. doi:10.1016/j.semradonc.2014.07.007
205. Multhoff G, Radons J. Radiation, inflammation, and immune responses in cancer. Front Oncol (2012) 2:58. doi:10.3389/fonc.2012.00058
206. Torres MA, Pace TW, Liu T, Felger JC, Mister D, Gregory H. Predictors of depression in breast cancer patients treated with radiation: role of prior chemotherapy and nuclear factor kappa B. Cancer (2013) 119:1951–9. doi:10.1002/cncr.28003
207. Harless WW. Cancer treatments transform residual cancer cell phenotype. Cancer Cell Int (2011) 11:1. doi:10.1186/1475-2867-11-1
208. Wang Z, Li Y, Kong D, Banerjee S, Ahmad A, Azmi AS, et al. Acquisition of epithelial-mesenchymal transition phenotype of gemcitabine-resistant pancreatic cancer cells is linked with activation of the Notch signaling pathway. Cancer Res (2009) 69:2400–7. doi:10.1158/0008-5472.CAN-08-4312
209. Hay ED. An overview of epithelio-mesenchymal transformation. Acta Anat (1995) 154:8–20. doi:10.1159/000147748
210. Corallino S, Malabarba MG, Zobel M, Di Fiore PP, Scita G. Epithelial-to-mesenchymal plasticity harnesses endocytic circuitries. Front Oncol (2015) 5:45. doi:10.3389/fonc.2015.00045
211. Pinto CA, Widodo E, Waltham M, Thompson EW. Breast cancer stem cells and epithelial mesenchymal plasticity – implications for chemoresistance. Cancer Lett (2013) 341:56–62. doi:10.1016/j.canlet.2013.06.003
212. McInnes LM, Jacobson N, Redfern A, Dowling A, Thompson EW, Saunders CM. Clinical implications of circulating tumor cells of breast cancer patients: role of epithelial mesenchymal plasticity. Front Oncol (2015) 5:42. doi:10.3389/fonc.2015.00042
213. Theveneau E, Mayor R. Collective cell migration of epithelial and mesenchymal cells. Cell Mol Life Sci (2013) 70:3481–92. doi:10.1007/s00018-012-1251-7
214. Rogers CD, Saxena A, Bronner ME. Sip1 mediates an E-cadherin-to-N-cadherin switch during cranial neural crest EMT. J Cell Biol (2013) 203:835–47. doi:10.1083/jcb.201305050
215. Rodriguez FJ, Lewis-Tuffin LJ, Anastasiadis PZ. E-cadherin’s dark side: possible role in tumor progression. Biochim Biophys Acta (2012) 1826:23–31. doi:10.1016/j.bbcan.2012.03.002
216. Bednarz-Knoll N, Alix-Panabières C, Pantel K. Plasticity of disseminating cancer cells in patients with epithelial malignancies. Cancer Metastasis Rev (2012) 31:673–87. doi:10.1007/s10555-012-9370-z
217. Chouaib S, Janji B, Tittarelli A, Eggermont A, Thiery JP. Tumor plasticity interferes with anti-tumor immunity. Crit Rev Immunol (2014) 34:91–102. doi:10.1615/CritRevImmunol.2014010183
218. Cai X, Dai Z, Reeves RS, Caballero-Benitez A, Duran KL, Delrow JJ, et al. Autonomous stimulation of cancer cell plasticity by the human NKG2D lymphocyte receptor coexpressed with its ligands on cancer cells. PLoS One (2014) 9:e108942. doi:10.1371/journal.pone.0108942
219. Zuo J, Ishikawa T, Boutros S, Xiao Z, Humtsoe JO, Kramer RH. Bcl-2 overexpression induces a partial epithelial to mesenchymal transition and promotes squamous carcinoma cell invasion and metastasis. Mol Cancer Res (2010) 8:170–82. doi:10.1158/1541-7786.MCR-09-0354
220. Mitra A, Menezes ME, Shevde LA, Samant RS. DNAJB6 induces degradation of β-catenin and causes partial reversal of mesenchymal phenotype. J Biol Chem (2010) 285:24686–94. doi:10.1074/jbc.M109.094847
221. Morbini P, Inghilleri S, Campo I, Oggionni T, Zorzetto M, Luisetti M. Incomplete expression of epithelial-mesenchymal transition markers in idiopathic pulmonary fibrosis. Pathol Res Pract (2011) 207:559–67. doi:10.1016/j.prp.2011.06.006
222. Damonte P, Gregg JP, Borowsky AD, Keister BA, Cardiff RD. EMT tumorigenesis in the mouse mammary gland. Lab Invest (2007) 87:1218–26. doi:10.1038/labinvest.3700683
223. Umbreit C, Flanjak J, Weiss C, Erben P, Aderhold C, Faber A, et al. Incomplete epithelial – mesenchymal transition in p16-positive squamous cell carcinoma cells correlates with β-catenin expression. Anticancer Res (2014) 34:7061–69.
224. Zeisberg M, Neilson EG. Mechanisms of tubulointerstitial fibrosis. J Am Soc Nephrol (2010) 21:1819–34. doi:10.1681/ASN.2010080793
225. Gopal SK, Greening DW, Mathias RA, Ji H, Rai A, Chen M, et al. YBX1/YB-1 induces partial EMT and tumourigenicity through secretion of angiogenic factors into the extracellular microenvironment. Oncotarget (2015) 6:13718–30.
226. Lundgren K, Nordenskjöld B, Landberg G. Hypoxia, snail and incomplete epithelial-mesenchymal transition in breast cancer. Br J Cancer (2009) 101:1769–81. doi:10.1038/sj.bjc.6605369
227. Schuetz AN, Rubin BP, Goldblum JR, Shehata B, Weiss SW, Liu W, et al. Intercellular junctions in Ewing sarcoma/primitive neuroectodermal tumor: additional evidence of epithelial differentiation. Mod Pathol (2005) 18:1403–10. doi:10.1038/modpathol.3800435
228. Burns WC, Twigg SM, Forbes JM, Pete J, Tikellis C, Thallas-Bonke V, et al. Connective tissue growth factor plays an important role in advanced glycation end product-induced tubular epithelial-to-mesenchymal transition: implications for diabetic renal disease. J Am Soc Nephrol (2006) 17:2484–94. doi:10.1681/ASN.2006050525
229. Voulgari A, Pintzas A. Epithelial-mesenchymal transition in cancer metastasis: mechanisms, markers and strategies to overcome drug resistance in the clinic. Biochim Biophys Acta (2009) 1796:75–90. doi:10.1016/j.bbcan.2009.03.002
Keywords: partial EMT, intermediate EMT, cancer stem cells, cell-fate decisions, cancer systems biology
Citation: Jolly MK, Boareto M, Huang B, Jia D, Lu M, Ben-Jacob E, Onuchic JN and Levine H (2015) Implications of the hybrid epithelial/mesenchymal phenotype in metastasis. Front. Oncol. 5:155. doi: 10.3389/fonc.2015.00155
Received: 22 May 2015; Accepted: 29 June 2015;
Published: 20 July 2015
Edited by:
Lars Bolund, Aarhus University, DenmarkReviewed by:
Shree Ram Singh, National Cancer Institute, USACopyright: © 2015 Jolly, Boareto, Huang, Jia, Lu, Ben-Jacob, Onuchic and Levine. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José N. Onuchic and Herbert Levine, Bioscience Research Collaborative, Rice University, 6100 Main Street, MS 654, Houston, TX 77005-1827, USA,am9udWNoaWNAcmljZS5lZHU=;aGVyYmVydC5sZXZpbmVAcmljZS5lZHU=
†Deceased on June 5, 2015.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.