 Kefei Gu
Kefei Gu Tan Wang
Tan Wang Liying Peng4
Liying Peng4 Yueliang Zhao
Yueliang Zhao- 1Institute for Agri-Food Standards and Testing Technology, Shanghai Academy of Agricultural Sciences, Shanghai, China
- 2College of Food Science and Technology, Shanghai Ocean University, Shanghai, China
- 3Laboratory of Quality and Safety Risk Assessment for Aquatic Products on Storage and Preservation (Shanghai), Ministry of Agriculture, Shanghai, China
- 4Institute of Animal Husbandry and Veterinary Science, Shanghai Academy of Agricultural Science, Shanghai, China
FIP-fve, a fungal fruiting body protein from Flammulina velutipes, has potential immunomodulatory properties. Here, we investigated the immunomodulation mechanism of FIP-fve in Jurkat E6-1 cells by conducting a cell viability assay and IL-2 release assay. Kinase inhibitors experiment and proteomics analysis were also involved in the mechanism study. It was found that FIP-fve stimulated cell proliferation and enhanced IL-2 secretion in a dose-dependent manner in Jurkat E6-1 cells. Unbiased high-throughput proteomics analysis showed that 4 T cell immune activation markers, including ZAP-70, CD69, CD82, and KIF23, were upregulated in response to FIP-fve treatment. Further pathway analysis indicated that MAP2K3/p38 pathway-related proteins, including MAP2K, p38, ELK, AATF, FOS, and JUN-B, were unregulated. In addition, losmapimod (p38 inhibitor) and gossypetin (MAP2K3 inhibitor) inhibited FIP-fve enhanced cell proliferation and IL-2 release in Jurkat E6-1 cells. Our results demonstrate that FIP-fve stimulates cell proliferation and enhances IL-2 secretion through MAP2K3/p38α activation.
Introduction
FIP-fve is a fungal fruiting body protein from golden needle mushroom (Flammulina velutipes) which is widely consumed in the orient. It has been reported to possess immunomodulatory, antiviral, and antiallergic activities (1–4). The current research showed that FIP-fve stimulates mitogenesis of human peripheral blood mononuclear cells (PBMCs), promotes cell proliferation by inducing G0/G1 to S phase progression, and enhances the transcription of interleukin-2 (IL-2) and interferon-γ (IFN-γ) (5). It was a new mitogen and immunomodulator with therapeutic potential, which may have clinical utility in the adjuvant treatment of diseases, whose mechanisms of action were not fully understood. Therefore it is necessary to further study.
Mitogen-activated protein kinase (MAPK) pathways consisting of the extracellular signal-related kinase (ERK), c-jun amino-terminal kinase (JNK), and p38 mitogen-activated protein kinase signal pathway (p38MAPK/p38) has been reported to play a critical role in the activation of T cells during the immune response (5, 6). p38 is also considered to be involved in positive selection in early thymocyte development (5–12). The upstream signal factors of the p38 signaling pathway include MAP2K3 (MMK3) and MAP2K6 (MKK6). Previous studies demonstrated that T cells activated by FIP-fve could secrete IFN-γ and IL-2 and increase the expression of intercellular adhesion molecule-1 (ICAM-1) in a process mediated by the p38 α mitogen-activated protein kinase (p38α) signaling pathway (5). However, it is not clear whether MAP2K3 (MMK3) and MAP2K6 (MKK6) are involved in the immune regulation of T cells by FIP-fve.
Janus kinase signal transducer and activator of transcription (JAK/STAT) pathways play a major role in transferring signals from cell-membrane receptors to the nucleus (8). Studies have shown that JAK/STAT pathways are conventionally involved in the MAPK pathway in the regulation of T cells (9, 13–19). However, it is not clear whether JAK/STAT pathway is involved in the immune regulation of T cells induced by FIP-fve.
In this study, the Jurkat E6-1 cells, which are activated by exogenous lectin (such as PHA) and can produce a large amount of IL-2, were used to investigate the immunomodulation activity of FIP-fve. Cell viability assay, IL-2 release assay, kinase inhibitors, and proteomics analysis were employed to investigate the potential intracellular signal transduction pathways involved in FIP-fve-induced Jurkat E6-1 proliferation.
Materials and Methods
Chemicals and Reagents
Flammulina velutipes was provided by Shanghai Senyuan biology, China. Losmapimod (purity ≥ 98%) was obtained from Shanghai Beyotime Biotechnology (China). Gossypetin (purity ≥ 99%) was purchased from Shanghai ZhenZhun Biotechnology (China). Acetic acid, ammonium sulfate, NaCl, acetonitrile, Tris, and HCl were provided by Sinopharm Chemical Reagent (China). Coomassie Brilliant Blue R250 was obtained from Shanghai yuanye Bio-Technology (China). Cell lysis buffer (N8031) was obtained from Solarbio (China). DMSO, phytohemagglutinin (PHA), protease inhibitor cocktail (P8340), phosphatase inhibitor cocktail 2 (P5726), and phosphatase inhibitor cocktail 3 (Sigma-Aldrich) were obtained from Sigma (USA).
Isolation and Characterization of FIP-fve
FIP-fve extraction and purification were performed according to a reported method (1). Briefly, 500 g of the fruiting bodies of Flammulina velutipes were homogenized with 500 ml ice-cold 5% acetic acid solution with an ultrasonic processor (FS-N, Shanghai Ultrasonic Instrument, China), followed by sonication for 90 min (600 W, 3 S, 6 S; 90 min in total) in an ice bath. The soluble proteins in the supernatant were precipitated by the addition of ammonium sulfate to 95% saturated. After centrifugation at 2,000 rpm for 5 min, the precipitate was re-suspended in 10 m Mof Tris-HCl buffer (pH 8.0) and dialyzed against 1 L of Tris-HCl buffer at 4°C for 72 h with five times change of dialysis solution (total of 5 L of buffer). After that, the dialysate was applied to a HiTrapQ-HP column (GE, USA), which was previously equilibrated with 10 mM Tris/HCl at pH 8.0. The column was eluted and purified with a gradient of 0–40% 10 mM Tris/HCl at pH 8.0 and 1 M NaCl (on the chromatograph). The active fractions were collected according to the ultraviolet value and further purified on a Superdex75 column (GE, USA). The Superdex 75 column was previously equilibrated with 20 mM Tris/HCl at pH 8.0 containing 150 mM NaCl. Elution was performed with 20 mM Tris-HCl at pH 8.0 containing 150 mM NaCl. The collected fractions were checked by sodium dodecyl sulfate polyacrylamide Gel Electrophoresis (SDS-PAGE) and a BCA assay. Similar fractions were combined. Then a total of 19.3 mg of purified FIP-fve was obtained (2.14 mg/ml). The molecular weight of purified FIP-fve was measured by matrix-assisted laser desorption ionization time-of-flight mass spectrometry (5800 MALDI-TOF/TOF, AB Sciex, USA) in a positive ion mode, with the calibration ranges of the linear medium molecular weight calibration materials being 5737.609 + 50, 12,362 + 50, and 16,952 + 50 (Sigma, USA).
SDS/PAGE
The purification of FIP-fve was performed according to a previous study (20). Briefly, cellular proteins (15 μg) from each sample were separated by 12% SDS-PAGE (Mini-Protean TGX™ Gels, 10-wellcomb/Bio-Rad). The separation gel was then stained with Coomassie Brilliant Blue R250 for 12 h. After staining, the gel was washed with a destaining solution(10% acetic acid and 25% anhydrous ethanol)until the bands were visualized. Finally, the stained gel was scanned by Image Scanner (GE Healthcare, USA) at a resolution of 300 dpi (full-color mode).
Cell Culture and Proliferation Assay
Jurkat E6-1 cells were obtained from the American Type Culture Collection (ATCC). The cells were cultured in RPMI 1640 medium (Gibco, Grand Island, USA) supplemented with 10% fetal bovine serum (FBS, Gibco, Grand Island, USA) at 37°C under a 5% CO2. Cell viability was examined using the Cell Counting Kit (CCK-8) (Dojindo, Japan) according to previous studies (21–23). Briefly, cells with a density of 1.0 × 104 cells per well were seeded in 96-well microtiter plates overnight. After incubated with vehicle (PBS) or FIP-fve at concentrations (25, 50, 100, or 200 μg/ml), the culture medium was discarded to remove the FIP-fve. After that, the cells were incubated with 100 μl of CCK-8 solution (10%) in a culture medium for about 1 h. PHA (5 μg/ml) was used as a positive control. The absorbance of the mixture culture medium was measured at 450 nm using a microplate reader (Biotek, USA).
IL-2 Release Assay
Jurkat E6-1 cells with a density of 1.0 × 104 cells per well were seeded in 96-well microtiter plates overnight. The cells were then treated with vehicle (PBS), FIP-fve at various concentrations (25, 50, 100, or 200 μg/ml), or PHA (1.5 or 5.0 μg/ml, positive control) for 6 h. After that, the supernatant was collected, and the amount of IL-2 in the supernatant was measured using an ELISA kit for human IL-2 (USCN Life Science Inc, China). Absorbance was measured at 450 nm with a Bio-Rad 3550 Microplate Reader (Hercules, CA, USA).
Proteomic Analysis
After being treated with FIP-fve for 24 h, Jurkat E6-1 cells were lysed in cell lysis buffer (Solarbio, China) supplemented with protease inhibitor cocktail (Sigma, USA), phosphatase inhibitor cocktail 2 (Sigma, USA), and phosphatase inhibitor cocktail 3 (Sigma, USA) by ultrasonication on an ice bath for 3 min. The protein concentration was measured using the BCA Protein Assay kit (Shanghai EpiZyme Biotechnology, China). The protein was enzymolyzed and labeled with a TMT reagent according to FASP (24) before being sent to Shanghai Luming Biotechnology Co., Ltd. China for Tandem Mass Tag™ (TMT™) proteomic analysis.
Reverse-phase (RP) separation was performed on an 1100 HPLC System (Agilent, USA) equipped with an Agilent Zorbax Extend RP column (5 μm, 150 × 2.1 mm). The mobile phase was composed of 2% acetonitrile in water (solvent A) and 98% acetonitrile in water (solvent B). The flow rate was 0.3 mL/min in the following gradients: 8 min, 98%A; 8.01 min, 95%A; 28 min, 75%A; 50 min, 60%A; 50.01 min, 10%A; 60 min, 10%A; 60.01 min, 98%A; and 65 min, 98%A. The injection volume was 1 μl. PDA was set at 210 and 280 nm. The samples were harvested from 8 to 50 min, and elution buffer was collected in 10 centrifuge tubes every minute and was numbered. The separated peptides were lyophilized for MS detection.
All analyses were performed by a Q Exactive mass spectrometer (Thermo, USA) equipped with a Nanospray Flex source (Thermo, USA). The samples were loaded and separated by a C-18 column (75 μm × 150 nm, 2 μm, 100 A, Dionex, USA) on an EASYnLC™1200 system (Thermo Fisher Scientific, United States). The flow rate was 300 nl/min, and the linear gradient lasted 90 min (0–55 min, 8%B; 55–79 min, 30%B; 79–80 min, 50% B; 80–90 min, 100% B; mobile phase A = 0.1% FA in water; mobile phase B = 80% ACN/0.1% FA in water). Full MS scans were acquired in the mass range of 300–1,600 m/z with a mass resolution of 70,000, and the AGC target value was set at 1e6. The ten most intense MS peaks were fragmented with higher-energy collisional dissociation (HCD) with an NCE of 32. MS/MS spectra were obtained with a resolution of 17,500 with an AGC target of 2e5 and a max infection time of 8 ms. The Q-E dynamic exclusion was set at 30.0 s and run under positive mode.
Database Search Proteome Discoverer (v 2.2) was used to thoroughly search all of the Q Exactive raw data against the sample protein database. Database searches were performed with trypsin digestion specificity. Cysteine alkylation was considered a fixed modification in the database search. For the protein quantification method, TMT6-plex was selected. The global false discovery rate (FDR) was set to 0.01, and the protein groups considered for quantification required at least 2 peptides.
Xcalibur 2.1 data processing software was used for FIP-fve characteristic mass spectrometry identification. Proteome Discoverer TM 2.2 software was used for cell proteomics experimental data analysis, and the UniProt human database was used. The false-positive rate of peptide identification was less than 1%.
Statistical Analysis
All experimental data were the average of at least three independent experiments. Differences between experimental groups were calculated using the Student’s paired t-test. A value of p < 0.05 was considered statistically significant.
Results and Discussion
Preparation of FIP-fve
The crude extract yielded two distinct protein bands with apparent molecular masses of 13 and 30 kDa in SDS/PAGE (Supplementary Figure 1A). Then, the crude extract was purified with a HiTrap Q-HP column, and the active FIP-fve (13 kDa) (1) obtained from the crude extract was further purified with a Superdex 75 column. The purified FIP-fve identified by an LC, yielded a single band with a molecular mass of 13 kDa (Supplementary Figures 1B,C) as determined by the SDS/PAGE assay. A total of 19.3 mg of purified FIP-fve with a concentration of 2.14 mg/ml was obtained as determined by the BCA method. Moreover, the molecular weight of the purified FIP-fve was 12,737.28 Da as determined by the 5800 MALDI-TOF/TOF (Figure 1).
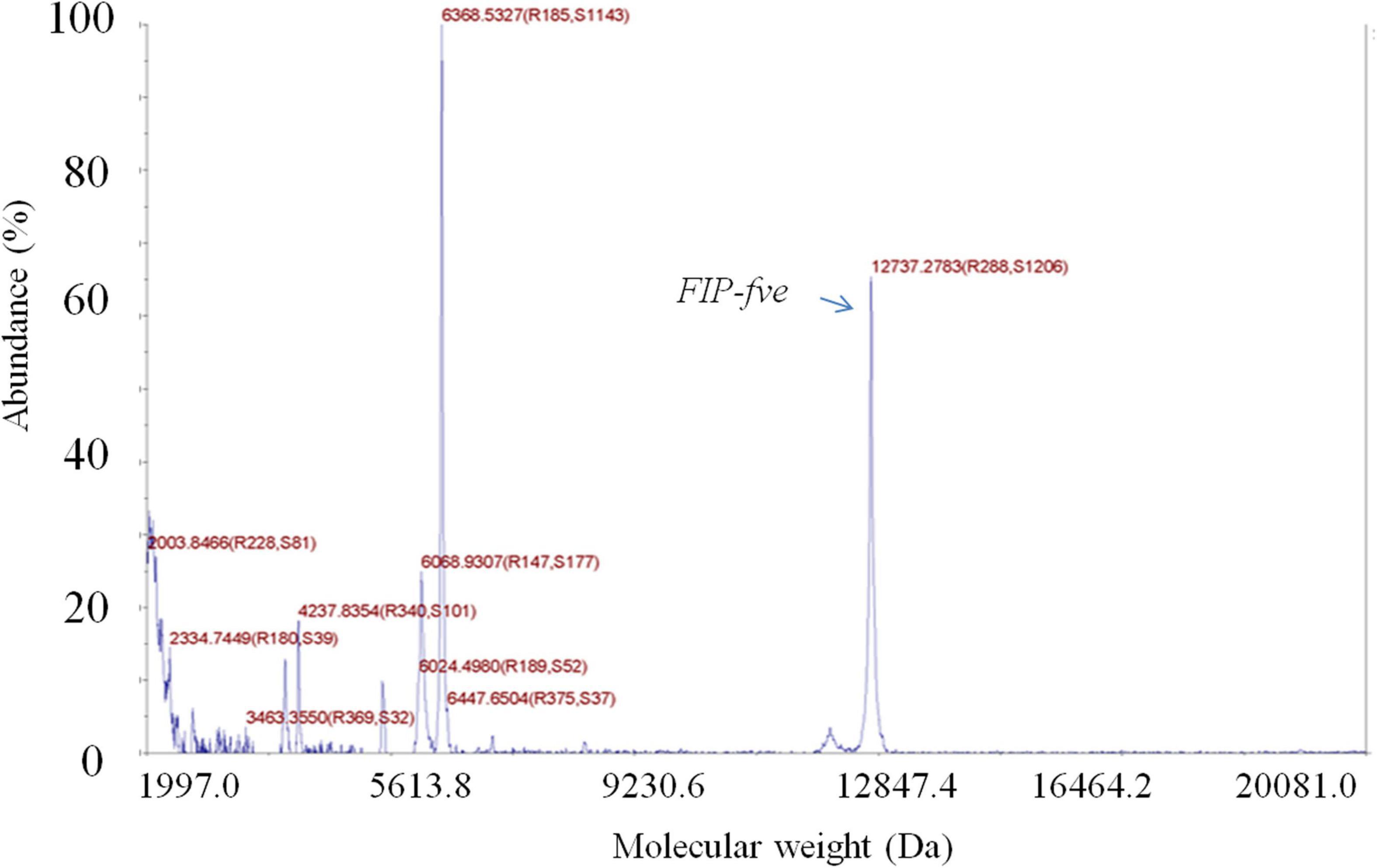
Figure 1. FIP-fve is detected by MALDI-TOF/TOF. The molecular weight of FIP-fve was measured by matrix-assisted laser desorption ionization time-of-flight mass spectrometry in a positive ion mode, with the calibration ranges of the linear medium molecular weight calibration materials being 5,737.609 + 50, 12,362 + 50, and 16,952 + 50.
FIP-fve Stimulates Cell Proliferation and Enhances IL-2 Release
To evaluate the immunomodulation activity of FIP-fve, we next determined the effect of FIP-fve on cell proliferation and IL-2 release in Jurkat E6-1 cells as the binding of IL-2 to its receptor initiates a series of events that result in cell-cycle progression and T cell proliferation (25). Moreover, the Jurkat E6-1 cells activated by exogenous lectin (such as PHA) can produce a large amount of IL-2. And PHA was used as a positive control (5). The proliferation of Jurkat E6-1 cells was evaluated by CCK-8 kit after 12 and 24 h treatment with control, PHA (5 μg/ml), and FIP-fve (100 μg/ml). Although weaker than PHA, FIP-fve was found to stimulate Jurkat E6-1 cells proliferation significantly (Figure 2). The results were consistent with a previous study (5) where FIP-fve was found to be capable of promoting PBMCs proliferation through enhancing the G1 to S phase progression of lymhocytes.
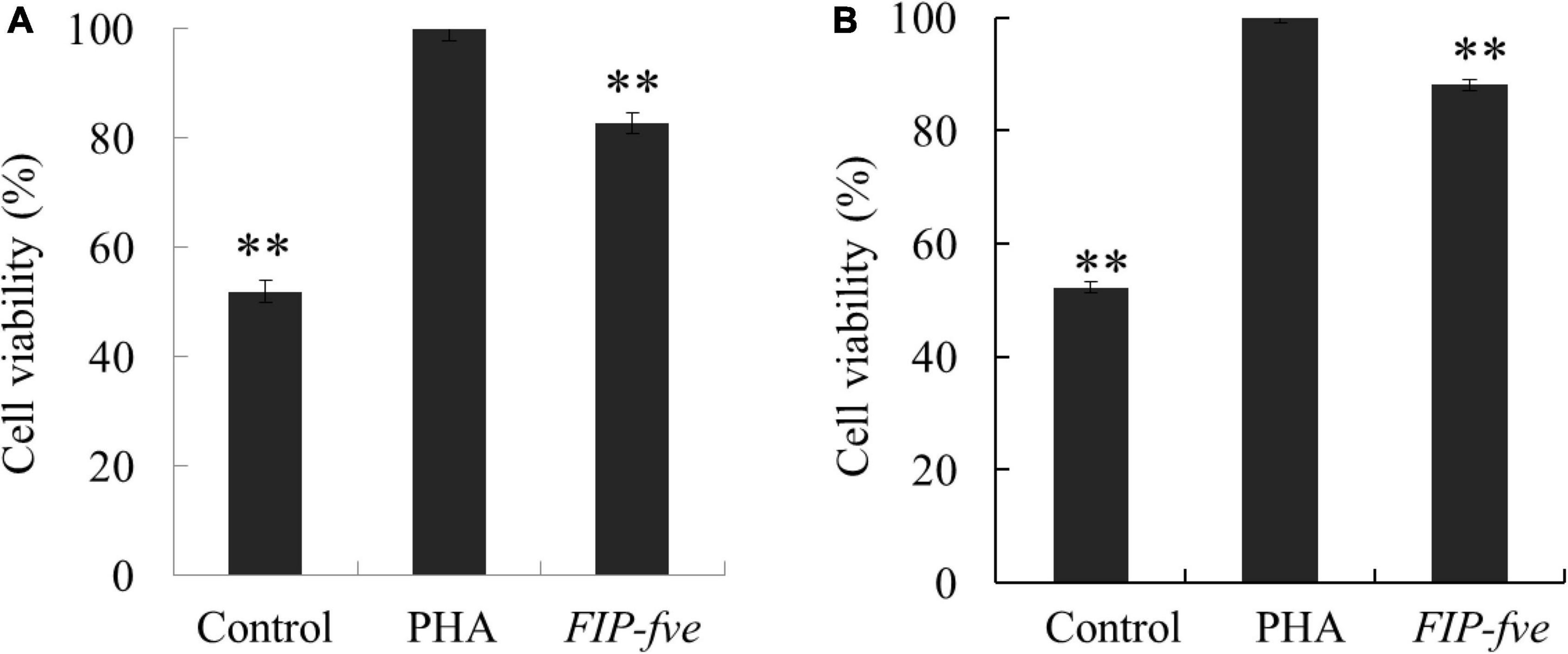
Figure 2. FIP-fve treatment results in Jurkat E6-1 cells Proliferation. Jurkat E6-1 cells were treated with FIP-fve (100 μg/ml) or PHA (5 μg/ml) for 12 h (A) and 24 h (B). Cell viability was assessed by Cell Counting kit-8 assay (CCK-8). PHA was used as a positive control. Values are mean ± SD, n = 3, **p < 0.01.
The in vitro immunomodulatory activity of FIP-fve was investigated by examining IL-2 release in Jurkat E6-1 cells. Jurkat E6-1 cells were cultured with different concentrations of FIP-fve ranging from 35.32 to 766.02 pg/ml for 6 h, and IL-2 production was measured by a human IL-2 ELISA kit. As shown in Figure 3, FIP-fve enhanced IL-2 release in a dose-dependent manner. For example, the levels of IL-2 released in the culture medium of cells treated with FIP-fve at a concentration of 25, 50, 100, and 200 μg/ml were 35.32, 106.88, 321.59, and 766.02 pg/ml, respectively. Moreover, the levels of IL-2 released from the FIP-fve (200 μg/ml) treated cells were almost reachedthat of PHA (5 μg/ml) treated cells (766.02 vs. 798.97 pg/ml). These results indicated that FIP-fve could induce T cell immune activation in Jurkat E6-1 cells.
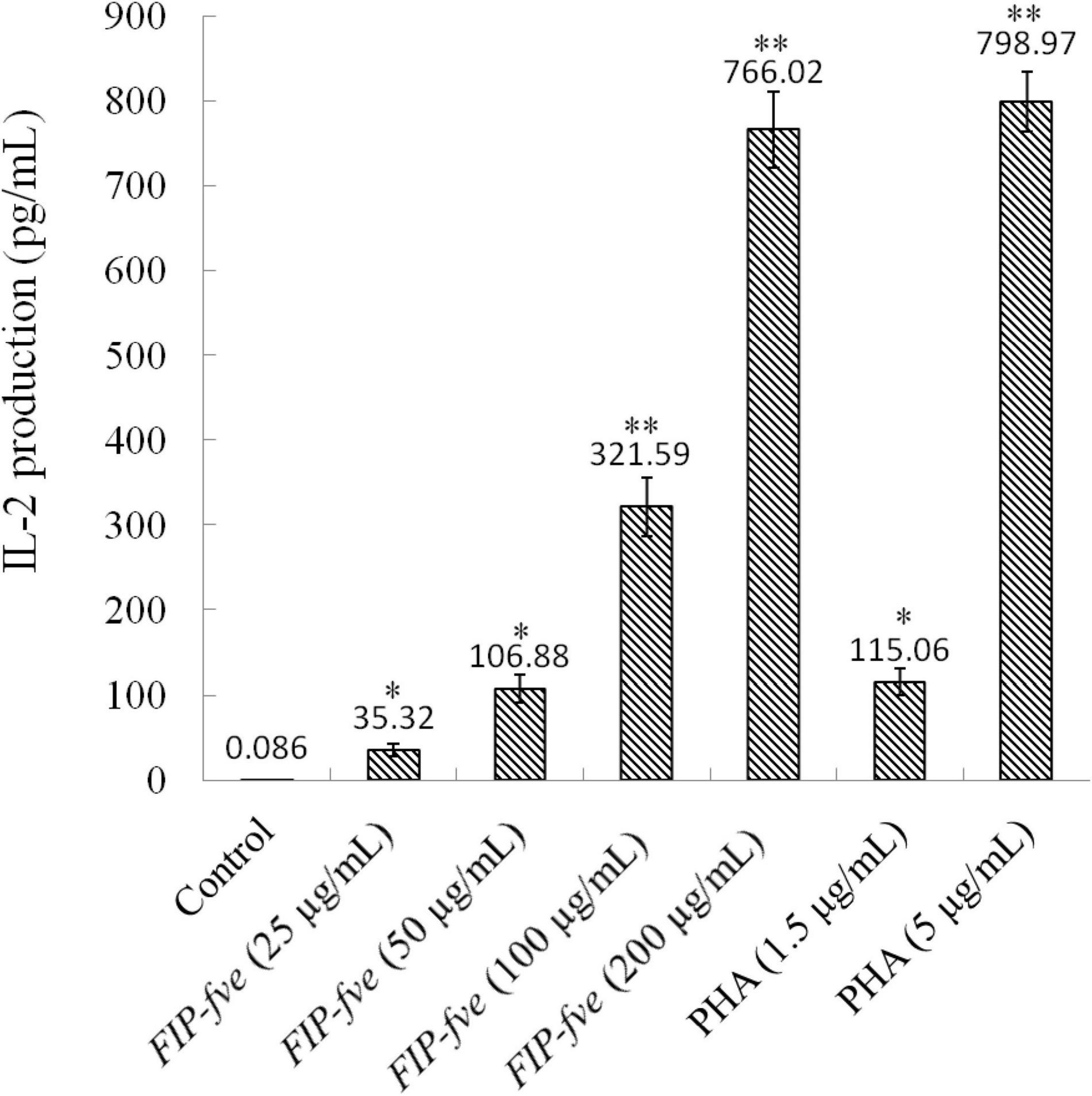
Figure 3. FIP-fve induced IL-2 release in Jurkat E6-1 cells. Jurkat E6-1 cells were treated with or without FIP-fve (25, 50, 100, and 200 μg/ml) for 6 h. PHA (1.5 and 5 μg/ml) was used as a positive control. IL-2 release was detected by ELISA kit for human IL-2 carried out according to the operating manual of the kit (enzyme marker, optical density, 0). Values are mean ± SD, n = 3, *p < 0.05, **p < 0.01.
FIP-fve Activated MAP2K3/p38α (MAPK14) Signaling Pathway in Jurkat E6-1 Cells as Evidenced by Proteomics Analysis
The above observations confirmed that FIP-fve stimulates cell proliferation and enhances IL-2 release in Jurkat E6-1 cells. Furthermore, unbiased high-throughput proteomics was conducted to explore the mechanism of action of FIP-fve in Jurkat E6-1 cells. As shown in Figure 4A, the principal component analysis (PCA) score chart could distinguish the FIP-fve group from the control group, indicating that there were differences between the two groups. The samples labeled by enzymatic hydrolysis were analyzed by liquid chromatography-mass spectrometry, and the database was searched. After the blank value was removed, a total of 5,668 credible proteins were detected according to the criteria of score sequence HT > 0 and unique peptide ≥ 1, in which 290 differentially expressed proteins were identified according to FC > 1.5 or FC < 2/3 (fold change, FC) and p < 0.05 by the t-test (Figure 4B). Based on 290 differentially expressed proteins data, Gene Ontology (GO) molecular function (MF) enrichment and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis were conducted by Proteome Discoverer TM 2.2 software. As shown in Figure 4C, the main upregulated GO MF were related to protein binding, poly (A) RNA binding, and threonine-type peptidase activity, indicating that FIP-fve treatment increased proteins and nucleic acid anabolism in Jurkat E6-1 cells. Moreover, KEGG analysis showed that FIP-fve treatment upregulated signal pathways involved in amino acid biosynthesis, carbon metabolism, proteasome, and glycolysis/glycogenesis (Figure 4D). Glycolysis, providing ATP for cell protein and nucleic acid anabolism, is considered to be the main metabolic pathway required for T cell activation. Amino acid synthesis is necessary for protein synthesis, nucleic acid synthesis, regulation of the mtroc1 pathway, and T cell stress pathway. Collectively, GO and KEGG pathway analysis indicated that FIP-fve treatment activated pathways related to cell proliferation, further confirming that FIP-fve stimulates cell proliferation in Jurkat E6-1 cells.
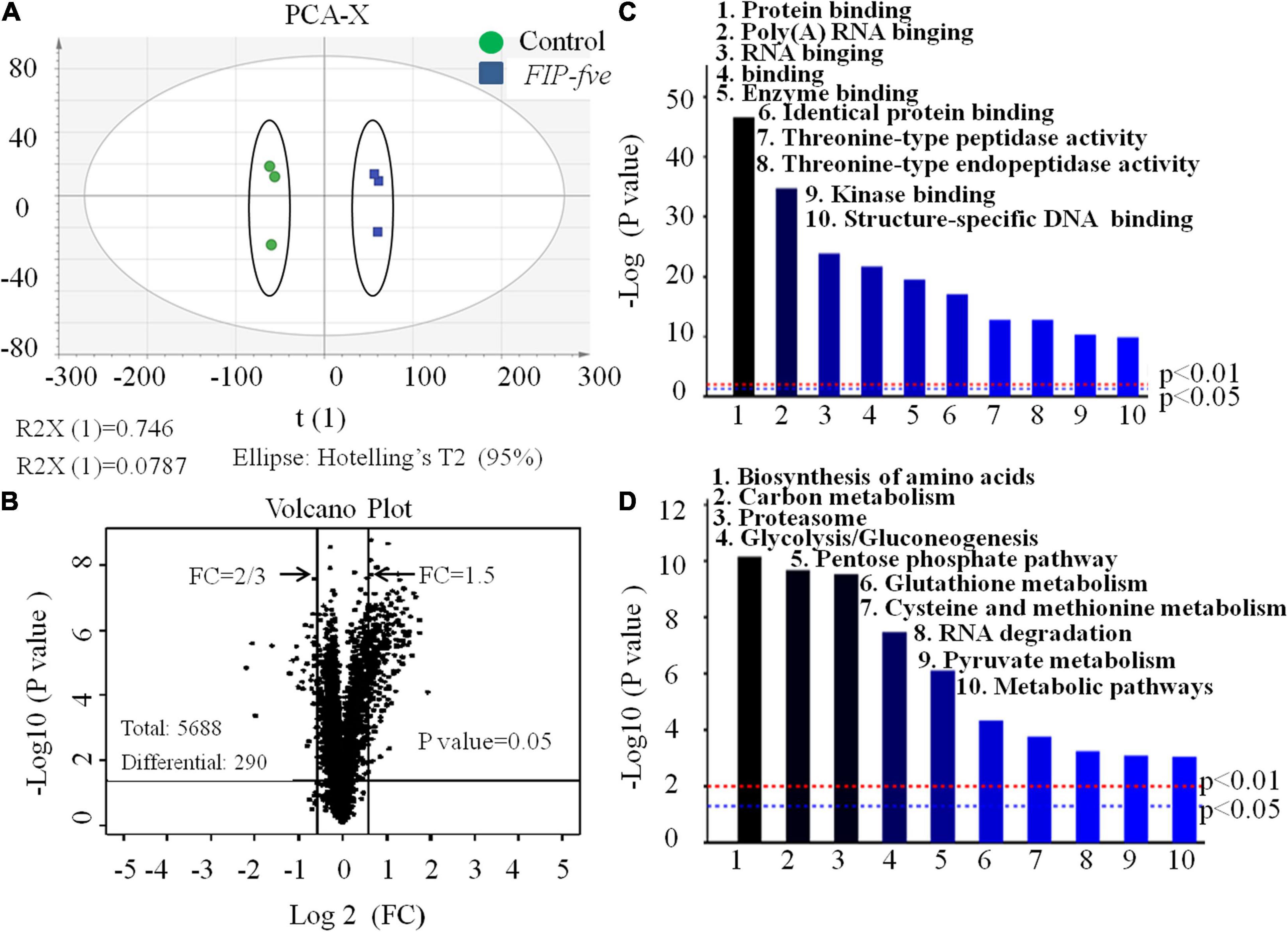
Figure 4. Gene Ontology (GO) molecular function (MF), and Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis of expression of intracellular proteins in FIP-fve-stimulated Jurkat E6-1 cells. (A) PCA analysis. (B) Volcanic map. A black dot in the picture represents a protein. The proteins in the range of FC > 1.5 and FC < 2/3 were differentially expressed proteins. Analysis of GO MF enrichment (C) and KEGG (D) pathway were conducted by Proteome Discoverer TM 2.2 software based on 290 differential expressed proteins.
Among the 290 differentially expressed proteins, 27 proteins were unregulated, including 4 T cell immune activation markers (ZAP-70, CD69, CD82, and KIF23) (10–12, 26–30) (Table 1). In combination with the enhanced IL-2 release, FIP-fve was further confirmed to induce T cell immune activation. Moreover, the unregulated 27 proteins include 13 MAPK-pathway-related signaling mediators (Table 1). As shown in Table 1, the expression of MAP2K3 in Jurkat E6-1 cells treated with FIP-fve was 1.5 times than that of control. The expression of p38α/MAPK14 was significantly unregulated. The expression of ELK1, AATF, FOS, and JUN-B, which were involved in the nuclear transcription process of the p38 kinase pathway, was also unregulated. Since the MAPK pathway has been reported to be one of the important pathways for T cell activation in the immune response and p38 kinase, the downstream effector of MAPK is considered to be an important kinase for the activation and differentiation of T cells (31), FIP-fve might induce immune activation through MAP2K3/p38α (MAPK14) signaling pathway in Jurkat E6-1 cells. In addition, it was also found that the expression of 5 JAK-STAT-pathway related signaling mediators (JAK3, STAT1, STAT3, STAT5B, and STAT6) was unregulated in response to FIP-fve (50 μg/ml) treatment (Table 1). As JAK/STAT pathways are conventionally involved in the MAPK pathway in the regulation of T cells, JAK3/STAT signaling pathway may be also involved in the immune regulation activity of FIP-fve in Jurkat E6-1 cells.
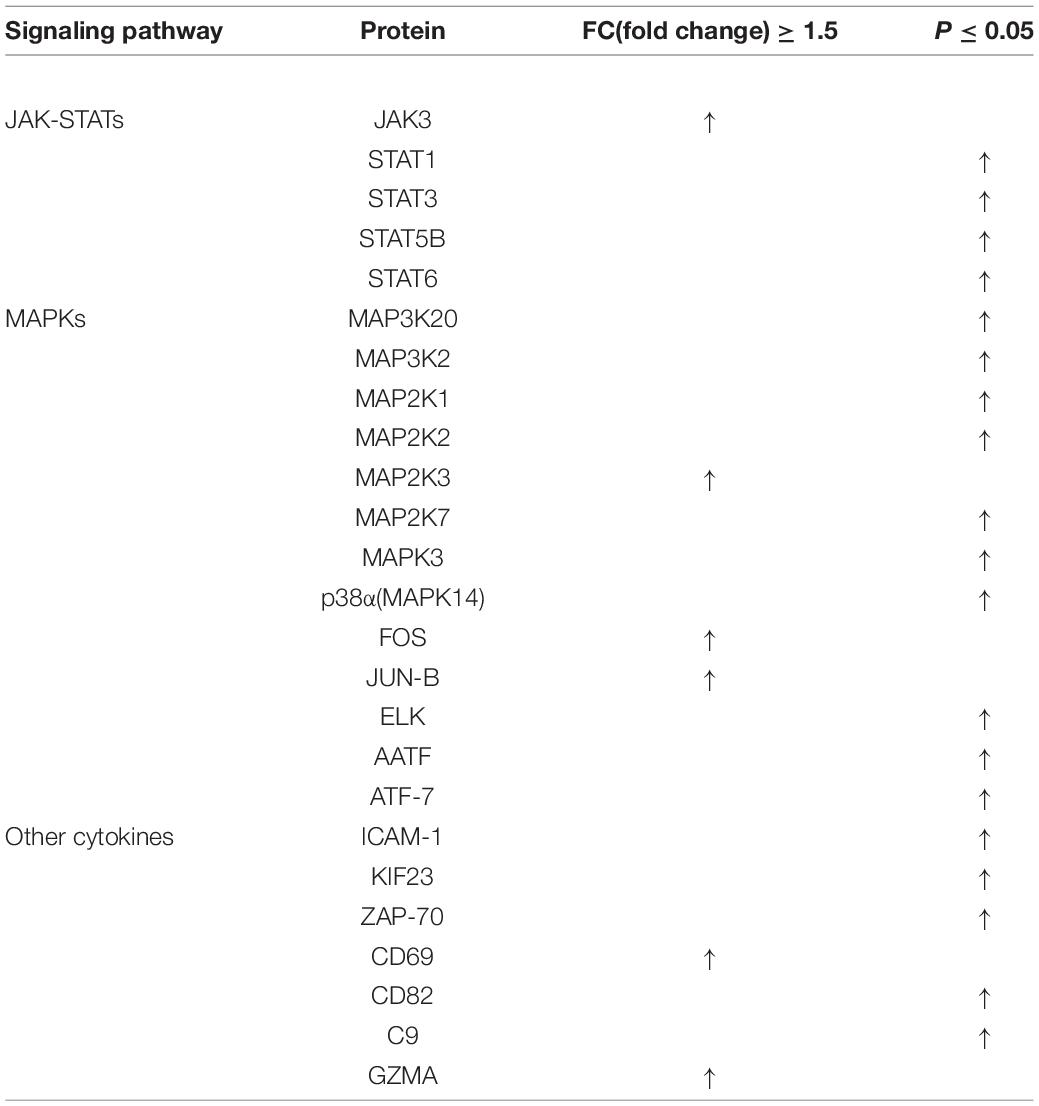
Table 1. Upregulated cell activation related proteins in FIP-fve-induced Jurkat E6-1 cells.
Gossypetin and Losmapimod Attenuated FIP-fve-Induced Immune Activation in Jurkat E6-1 Cells
Gossypetin (a MAP2K3 inhibitor) and losmapimod (a p38α inhibitor) were then used to study the involvement of the MAP2K3/p38α pathway in FIP-fve-induced immune activation in Jurkat E6-1 cells. As shown in Figure 5, 0.1 μM losmapimod and 40 μM gossypetin reduced cells growth stimulated by FIP-fve in Jurkat E6-1 cells by 64 and 71%, respectively (Figure 5A). Moreover, losmapimod and gossypetin attenuated FIP-fve-enhanced IL-2 released in Jurkat E6-1 cells by 83 and 90%, respectively (Figure 5B). These data in combination with results obtained from proteomics analysis indicate that FIP-fve may promote cell proliferation and IL-2 release by activating MAP2K3/p38α signaling pathway in Jurkat E6-1cells.
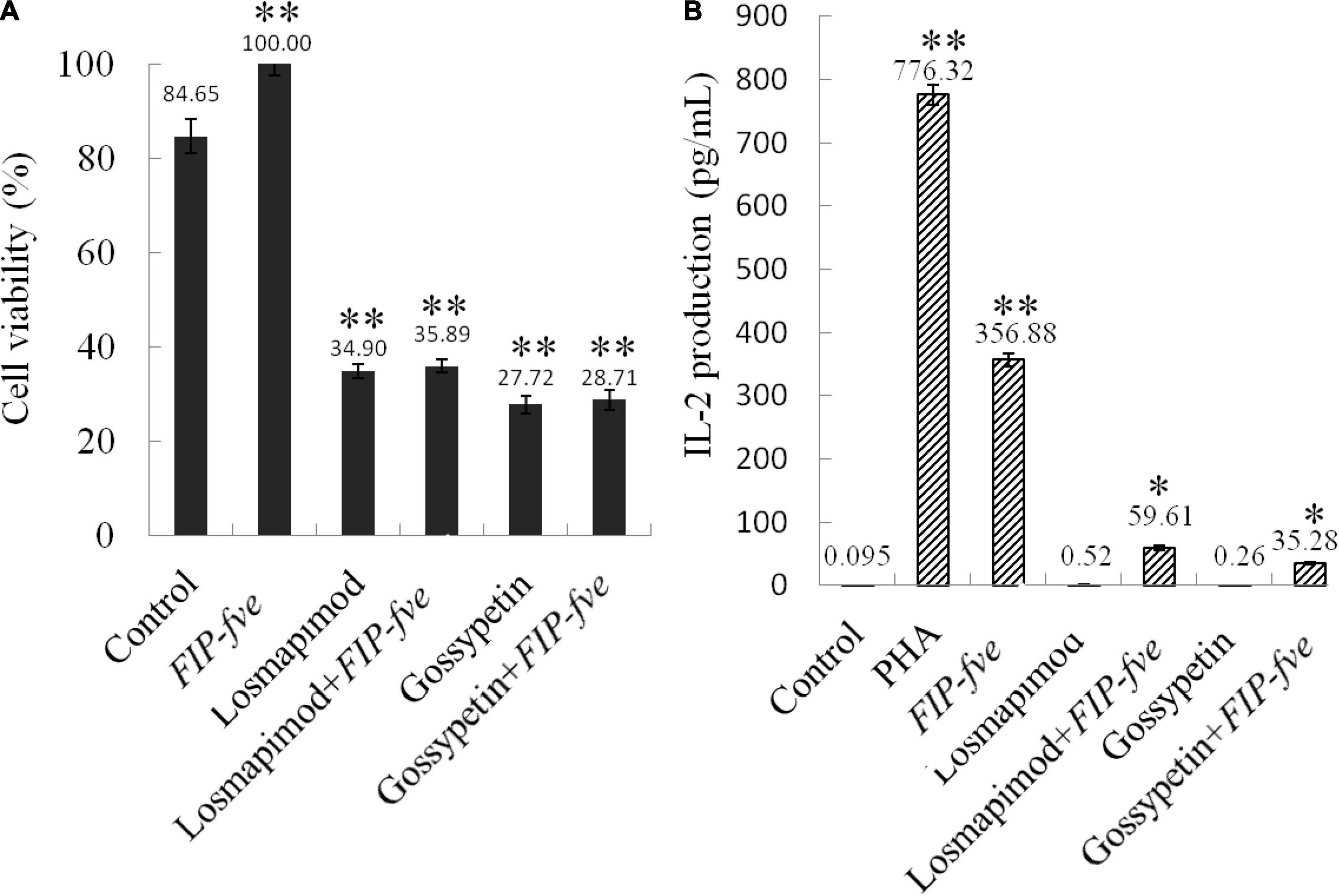
Figure 5. Gossypetin and losmapimod inhibit cell proliferation and IL-2 release in FIP-fve-induced Jurkat E6-1 cells. (A) Gossypetin and losmapimod attenuated proliferation of FIP-fve-induced Jurkat E6-1 cells. Cell viability was determined after treatment within RPMI 1640 medium supplemented with 10% fetal bovine serum FIP-fve (100 μg/ml) and/or Losmapimod (0.1 μM/ml) and/or Gossypetin (40 μM/ml) for 12 h. Values are mean ± standard deviation, n = 3. PHA (5 μg/ml) was used as a positive control. (B) Gossypetin and losmapimod inhibit FIP-fve-induced IL-2 release in Jurkat E6-1 cells. IL-2 release was detected by ELISA kit for human IL-2 after treatment with as in (A) for 6 h. Values are mean ± SD, n = 3, *p < 0.05, **p < 0.01.
Conclusion
FIP-fve, a fungal fruiting body protein from Flammulina velutipes, stimulated cell proliferation and enhanced IL-2 secretion in Jurkat E6-1 cells. Unbiased high-throughput proteomics analysis indicated that FIP-fve may induce immune activation through MAP2K3/p38α (MAPK14) signaling pathway in Jurkat E6-1 cells. Gossypetin (a MAP2K3 inhibitor) and losmapimod (a p38α inhibitor) attenuated cells growth stimulated by FIP-fve and IL-2 release enhanced by FIP-fve, further confirming that FIP-fve stimulates cell proliferation and enhances IL-2 secretion through MAP2K3/p38α activation in Jurkat E6-1 cells.
Data Availability Statement
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE [1] partner repository with the dataset identifier PXD032302.
Author Contributions
KG and YZ conceived the study. KG, TW, and YZ contributed to the design and drafted the work. KG, TW, and LP collected the data. KG and TW analyzed the data. All authors interpreted the data, read the manuscript, and approved the final version.
Funding
This work was supported by the National Ministry of Agriculture for Agricultural Products Risk Evaluation Project (No. GJFP201700604) and the Youth Program of the National Natural Science Foundation of China (No. 31601391).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Dong Shijuan from the Institute of Animal Husbandry and Veterinary Science, Shanghai Key Laboratory of Agricultural Genetics and Breeding for acknowledging the skillful assistance of Jurkat E 6-1 cells culture and analysis.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnut.2022.881924/full#supplementary-material
References
1. Ko JL, Hsu CI, Lin RH, Kao CL, Lin JY. A new fungal immunomodulatory protein, FIP-fve isolated from the edible mushroom, Flammulina velutipes and its complete amino acid sequence. Eur J Biochem. (1995) 228:244–9. doi: 10.1111/j.1432-1033.1995.0244n.x
2. Wasser SP, Weis AL. Therapeutic effects of substances occurring inhigher Basidiomycetes mushrooms: a m odern perspective. Crit Rev Immunol. (1999) 19:65–96.
3. Kong XH, Sun YF, Ren Y, Xi XW, Yang ZX. Application and research on imunomodulatory proteins of Flammulina velutipes. Biotechnology. (2006) 16:84–8.
4. Li S, Zhao Z, Li Y, Tang X. Advances of fungal immunomodulatory proteins in macromycete. Biotechnol Bull. (2012) 5:49–53.
5. Wang PH, Hsu CI, Tang SC, Huang YL, Lin JY, Ko JL. Fungal immunomodulat-ory protein from Flammulina velutipes induces interferon-gamma production through p38 mitogen-activated protein kinase signaling pathway. J Agric Food Chem. (2004) 52:2721–5. doi: 10.1021/jf034556s
6. Chan ED, Riches DW. IFN-gamma + LPS induction of iNOS is modulated by ERK, JNK/SAPK, and p38(mapk) in a mouse macrophage cell line. Am J Physiol Cell Physiol. (2001) 280:C441–50. doi: 10.1152/ajpcell.2001.280.3.C441
7. Chen C, Chen YH, Lin WW. Involvement of p38 mitogen-activated protein kinase in lipopolysaccharide-induced iNOS and COX-2 expression in J774 macrophages. Immunology. (1999) 97:124–9. doi: 10.1046/j.1365-2567.1999.00747.x
8. Renauld JC. Class II cytokine receptors and their ligands: key antiviral and inflammatory modulators. Nat Rev Immunol. (2003) 3:667–76. doi: 10.1038/nri1153
9. Seif F, Khoshmirsafa M, Aazami H, Mohsenzadegan M, Sedighi G, Bahar M. The role of JAK-STAT signaling pathway and its regulators in the fate of T helper cells. Cell Commun Signal. (2017) 15:23. doi: 10.1186/s12964-017-0177-y
10. Marzio R, Mauël J, Betz-Corradin S. CD69 and regulation of the immune function. Immunopharmacol Immunotoxicol. (1999) 21:565–82. doi: 10.3109/08923979909007126
11. Woods M, Guy R, Waldmann H, Glennie M, Alexander DR. A humanised therapeutic CD4 mAb inhibits TCR-induced IL-2, IL-4, and IL-10 secretion and expression of CD25, CD40L, and CD69. Cell Immunol. (1998) 185:101–13. doi: 10.1006/cimm.1998.1287
12. Ashe JM, Wiest DL, Abe R, Singer A. ZAP-70 protein promotes tyrosine phosphorylation of T cell receptor signaling motifs (ITAMs) in immature CD4+8+ thymocytes with limiting p56lck. J Exp Med. (1999) 189:1163–8. doi: 10.1084/jem.189.7.1163
13. Rincón M, Enslen H, Raingeaud J, Recht M, Zapton T, Su MS, et al. Interferon-gamma expression by Th1 effector T cells mediated by the p38 MAP kinase signaling pathway. EMBO J. (1998) 17:2817–29. doi: 10.1093/emboj/17.10.2817
14. Swan KA, Alberola-Ila J, Gross JA, Appleby MW, Forbush KA, Thomas JF, et al. Involvement of p21ras distinguishes positive and negative selection in thymocytes. EMBO J. (1995) 14:276–85. doi: 10.1002/j.1460-2075.1995.tb07001.x
15. Kim HH, Kwack K, Lee ZH. Activation of c-jun N-terminal kinase by 4-1BB (CD137), a T cell co-stimulatory molecule. Mol Cells. (2000) 10:247–52.
16. Hsu SC, Wu CC, Han J, Lai MZ. Involvement of p38 mitogen-activated protein kinase in different stages of thymocyte development. Blood. (2003) 101:970–6. doi: 10.1182/blood-2002-03-0744
17. Fernández E. Thymocyte development past the CD4(+)CD8(+) stage requires an active p38 mitogen-activated protein kinase. Blood. (2000) 95:1356–61. doi: 10.1182/blood.v95.4.1356.004k23_1356_1361
18. Choudhury BK, Wild JS, Alam R, Klinman DM, Boldogh I, Dharajiya N, et al. In vivo role of p38 mitogen-activated protein kinase in mediating the anti-inflammatory effects of CpG oligodeoxynucleotide in murine asthma. J Immunol. (2002) 169:5955–61. doi: 10.4049/jimmunol.169.10.5955
19. Chen CH, Zhang DH, LaPorte JM, Ray A. Cyclic AMP activates p38 mitogen-activated protein kinase in Th2 cells: phosphorylation of GATA-3 and stimulation of Th2 cytokine gene expression. J Immunol. (2000) 165:5597–605. doi: 10.4049/jimmunol.165.10.5597
20. Li F, Wen H, Zhang Y, Aa M, Liu X. Purification and characterization of a novel immunomodulatory protein from the medicinal mushroom Trametes versicolor. Sci China Life Sci. (2011) 54:379–85. doi: 10.1007/s11427-011-4153-2
21. Kellett ME, Greenspan P, Pegg RB. Modification of the cellular antioxidant activity (CAA) assay to study phenolic antioxidants in a Caco-2 cell line. Food Chem. (2018) 244:359–63. doi: 10.1016/j.foodchem.2017.10.035
22. Zhao Y, Zhou Y, Wang M. Brosimone I, an isoprenoid-substituted flavonoid, induces cell cycle G 1 phase arrest and apoptosis through ROS-dependent endoplasmic reticulum stress in HCT116 human colon cancer cells. Food Funct. (2019) 10:2729–38. doi: 10.1039/c8fo02315h
23. Zhao Y, Li ETS, Wang M. Alisol B 23-acetate induces autophagic-dependent apoptosis in human colon cancer cells via ROS generation and JNK activation. Oncotarget. (2017) 8:70239–49. doi: 10.18632/oncotarget.19605
24. Wiśniewski JR, Zougman A, Nagaraj N, Mann M. Universal sample preparation method for proteome analysis. Nat Methods. (2009) 6:359–62. doi: 10.1038/nmeth.1322
25. Zhou X, Xing J, Tang X, Sheng X, Zhan W. Immunological characteristics of Interleukin-2 receptor subunit beta (IL-2Rβ) in flounder (Paralichtlys olivaceus):implication for IL-2R function. Fish Shellfish Immunol. (2019) 93:641–51. doi: 10.1016/j.fsi.2019.07.059
26. Cambiaggi C, Scupoli MT, Cestari T, Gerosa F, Carra G, Tridente G, et al. Constitutive expression of CD69 in interspecies T-cell hybrids and locus assignment to human chromosome 12. Immnogenetics. (1992) 36:117–20. doi: 10.1007/BF00215288
27. Kimura MY, Koyama-Nasu R, Yagi R, Nakayama T. A new therapeutic target: the CD69-Myl9 system in immuneresponses. Semin Immunopathol. (2019) 41:349–58. doi: 10.1007/s00281-019-00734-7
28. Shibagaki N, Hanada K, Yamaguchi S, Yamashita H, Shimada S, Hamada H. Functional analysis of CD82 in the early phase of T cell activation: roles in cell adhesion and signal transduction. Eur J Immunol. (1998) 28:1125–33. doi: 10.1002/(SICI)1521-4141(199804)28:04<1125::AID-IMMU1125>3.0.CO;2-C
29. Korać P, Ajduković R, Kardum Paro MM, Jaksić B, Dominis M. Immunohistochemical analysis of ZAP-70 expression in chronic lymphocytic leukemia. J Mol Histol. (2009) 40:81–6. doi: 10.1007/s10735-009-9213-6
30. Kim G, Jang MS, Son YM, Seo MJ, Ji SY, Han SH, et al. Curcumin inhibits CD4(+) T cell activation, but augments CD69 expression and TGF-β1-mediated generation of regulatory T cells at late phase. PLoS One. (2013) 8:e62300. doi: 10.1371/journal.pone.0062300
Keywords: FIP-fve, immunomodulation mechanism, proteomics analysis, MAP2K3/p38α, IL-2
Citation: Gu K, Wang T, Peng L and Zhao Y (2022) FIP-fve Stimulates Cell Proliferation and Enhances IL-2 Release by Activating MAP2K3/p38α (MAPK14) Signaling Pathway in Jurkat E6-1 Cells. Front. Nutr. 9:881924. doi: 10.3389/fnut.2022.881924
Received: 23 February 2022; Accepted: 28 March 2022;
Published: 09 May 2022.
Edited by:
Ren-You Gan, Chinese Academy of Agricultural Sciences, ChinaCopyright © 2022 Gu, Wang, Peng and Zhao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yueliang Zhao, eXVlbGlhbmcyMTEwQDE2My5jb20=, eWx6aGFvQHNob3UuZWR1LmNu
†These authors have contributed equally to this work