 Ning Xu
Ning Xu Yue He1,2
Yue He1,2
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Neurosci. , 13 February 2025
Sec. Neurogenesis
Volume 19 - 2025 | https://doi.org/10.3389/fnins.2025.1512073
Major depressive disorder (MDD) is a highly prevalent and severely disabling psychiatric disorder that decreases quality of life and imposes substantial economic burden. Acupuncture has emerged as an effective adjunctive treatment for depression, it regulates neurotransmitters involved in mood regulation and modulates the activity of specific brain regions associated with emotional processing, as evidenced by neuroimaging and biochemical studies. Despite these insights, the precise neuroplastic mechanisms through which acupuncture exerts its antidepressant effects remain not fully elucidated. This review aims to summarize the current knowledge on acupuncture’s modulation of neuroplasticity in depression, with a focus on the neuroplasticity-based targets associated with acupuncture’s antidepressant effects. We encapsulate two decades of research into the neurobiological mechanisms underpinning the efficacy of acupuncture in treating depression. Additionally, we detail the acupoints and electroacupuncture parameters used in the treatment of depression to better serve clinical application.
Major depressive disorder (MDD) is a highly prevalent and severely disabling psychiatric disorder (Kupfer et al., 2012). It significantly decreases quality of life and imposes substantial economic burden (Fox and Lobo, 2019; Monroe and Harkness, 2022). Although the pathogenesis of MDD has advanced considerably, it remains incompletely understood. Studies have indicated a reduction in neuroplasticity among patients with depression and in animals subjected to stress or various depression models. Brain imaging studies have shown that patients with MDD exhibit volume reductions and decreased connectivity in the prefrontal cortex (PFC) and hippocampus (Evans et al., 2018; MacQueen and Frodl, 2011; Savitz and Drevets, 2009), with some studies also noting changes in the anterior cingulate cortex, striatum, and amygdala (Gerhard et al., 2016; Gujral et al., 2017). Postmortem analyses have shown decreased neuronal soma size and reduced glial numbers in the PFC of MDD patients (Rajkowska and Stockmeier, 2013; Sanacora et al., 2008). Similarly, preclinical studies in rodents and nonhuman primates have demonstrated neuron atrophy in the hippocampus and PFC under chronic stress conditions (Duman et al., 2016; McEwen et al., 2015). Collectively, these findings indicate that depression induces structural and functional alterations in specific brain regions. Two primary hypotheses, i.e., the neuroplasticity and neurogenesis hypotheses, have been proposed at the molecular and cellular level to explain these findings (Boku et al., 2018; Price and Duman, 2020). They proposed that the pathophysiology of depression might be linked to impairments in neuroplasticity, such as neural atrophy, neural apoptosis, deficits in functional neurocircuitry, reduced generation of new neurons, and dysregulation of synaptic plasticity in cortical and limbic regions.
Neuroplasticity can be broadly defined as the capacity of the nervous system to reorganize its structure, function, and connections in response to both intrinsic and extrinsic stimuli (Cramer et al., 2011). It involves the growth and adaptability of neural pathways and synapses at the structural level (Magee and Grienberger, 2020). Functionally, this adaptability is evidenced by increased long-term potentiation (LTP), a key form of synaptic plasticity that underpins the brain’s capacity to adapt through learning and memory to the ever-changing environment. Neurogenesis, a specific form of neuroplasticity, involves the generation of new neurons in the adult brain from pluripotent stem cells (Otte et al., 2016). There is evidence that neurogenesis occurs in the dentate gyrus (DG) of adult humans (Eriksson et al., 1998; Spalding et al., 2013), suggesting that the human hippocampus retains the ability to generate neurons during adulthood, which is likely to contribute to learning and memory (Malhi and Mann, 2018). The neuroplasticity process is influenced by a variety of signaling mechanisms and molecules, including but not limited to neurotrophic factors, growth factors, cytokines, and neurotransmitters (Gonçalves et al., 2016).
Neuroplasticity has been identified as a promising target for managing depression and other neuropsychiatric disorders. The exploration of specific signaling pathways, neurotransmitters (Kraus et al., 2017), and neurotrophic factors (Castrén and Monteggia, 2021) that influence neuroplasticity in specific brain regions, such as the cortical and limbic systems, has garnered significant attention. Encouraging findings from both animal and human studies indicate that novel rapid-acting antidepressants (i.e., ketamine) (Aleksandrova and Phillips, 2021), physical exercise (Vivar et al., 2012), or learning (Sagi et al., 2012) can enhance neuroplasticity, prompting further exploration and clinical application of promising neuroplasticity-based therapies. There is compelling evidence supporting the clinical efficacy of both manual acupuncture and electroacupuncture (EA) as complementary and integrative therapies for depression (Yang et al., 2022). Extensive preclinical and clinical research has been conducted in recent years on the molecular and cellular mechanisms of acupuncture for treating depression. However, much of this research remains descriptive. To date, detailed explanations of the potential antidepressant mechanisms linked to neuroplasticity have yet to be articulated and summarized.
To elucidate the neurobiological mechanisms associated with neuroplasticity that underpin the antidepressant effects of acupuncture on depression and to establish a solid foundation for future research, this review compiles recent data from basic research closely related to the effects of acupuncture on neuroplasticity in the treatment of depression. These studies demonstrated findings through methods such as Nissl staining, Golgi staining, transmission electron microscopy, immunohistochemical analysis, and electrophysiological recording, showing an increase in neuron numbers, the reversal of synaptic ultrastructural pathology, or the successful induction of long-term potentiation (LTP) in specific brain regions. Tables 1–2 summarizes these studies, highlighting the neuroplastic effects and related mechanisms of acupuncture treatment of depression (Figure 1), and provides a framework at the molecular and cellular levels to enhance understanding of the neuroplastic mechanisms involved in acupuncture treatment of depression.
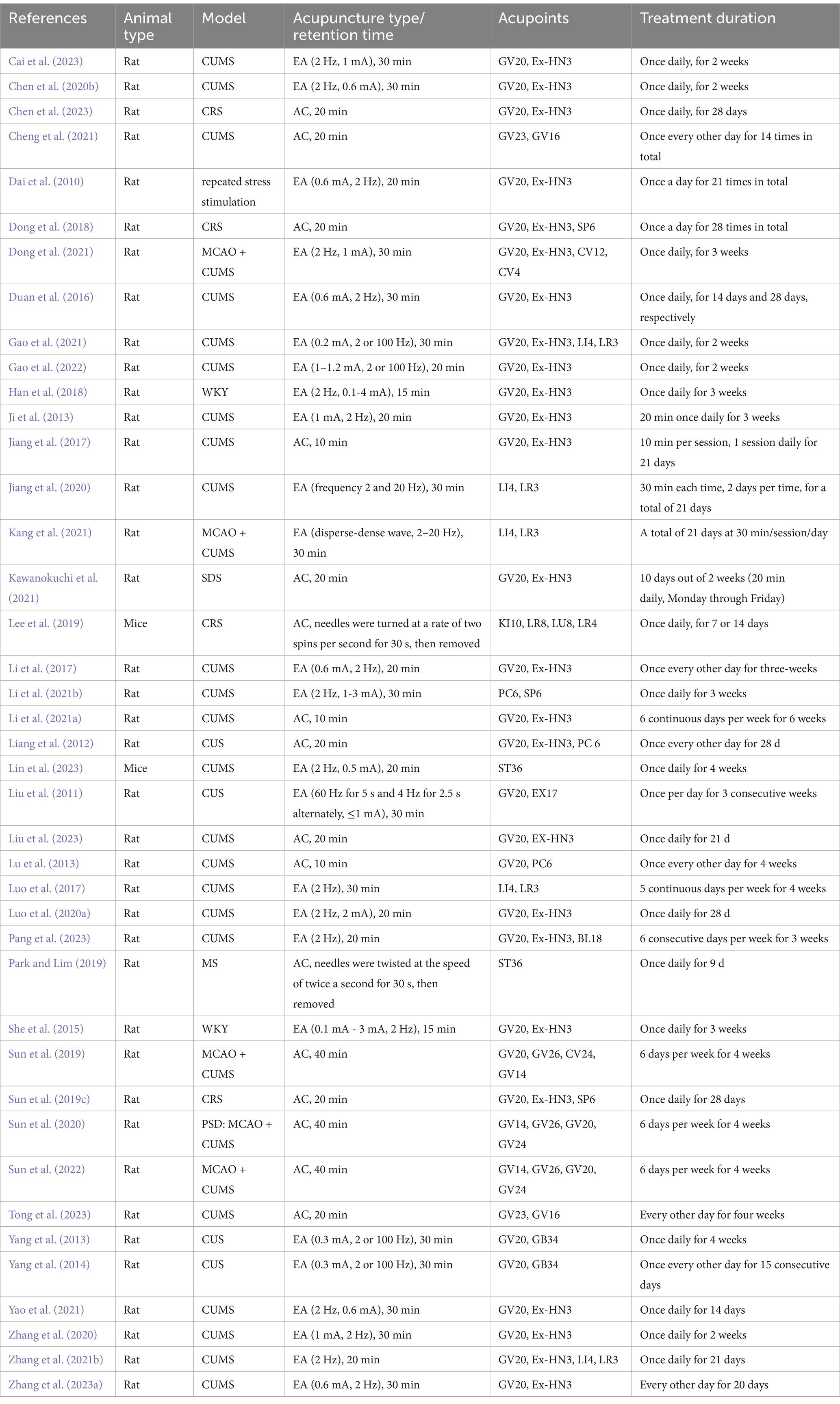
Table 1. Summary for animal intervention strategy of acupuncture in treating depression.
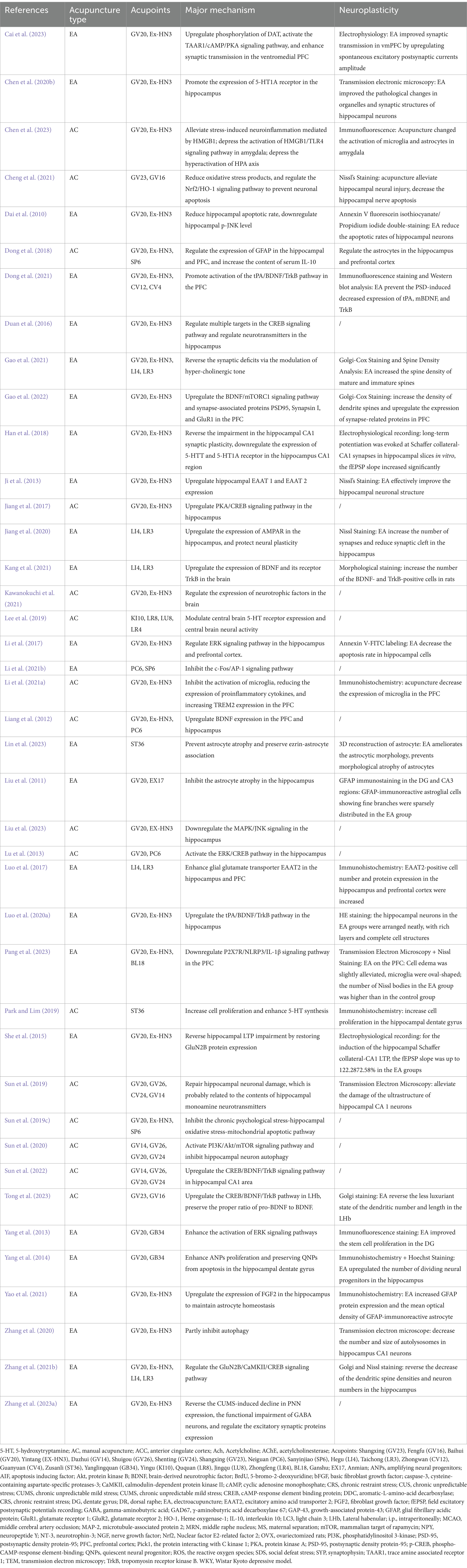
Table 2. Summary for the major mechanism of acupuncture.
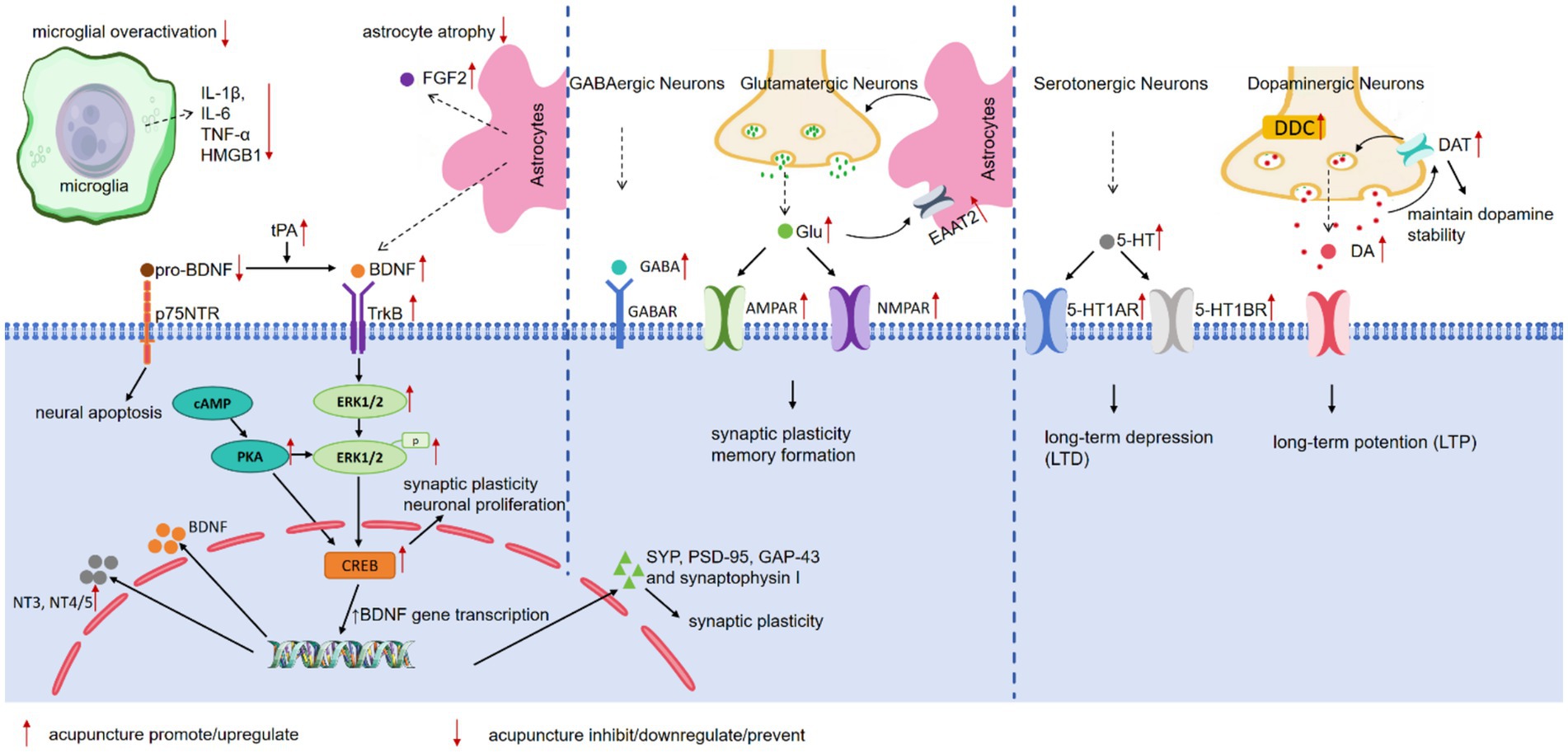
Figure 1. The neuroplasticity related mechanisms of acupuncture in treating depression.
BDNF is a neuropeptide synthesized primarily in the cell bodies of neurons and glia, with the highest expression in the hippocampus, cerebellum, and cerebral cortex (Colucci-D’Amato et al., 2020). BDNF signaling critically modulating synaptic plasticity, neuronal survival, and differentiation, plays a pivotal role in the central nervous system (CNS). BDNF binds with high affinity to the tyrosine kinase B (TrkB) receptor, triggering the activation of TrkB and its downstream signaling cascades, including the mitogen-activated protein kinase (MAPK–ERK), phosphoinositide 3-kinase (PI3K-Akt), and phospholipase-C gamma (PLC-γ) pathways (Wang et al., 2022). These pathways are essential for supporting neuronal survival, function, and synaptic plasticity.
Preclinical research conducted by Liang et al. (2012) in animal models of depression highlighted the beneficial effects of acupuncture in upregulating BDNF mRNA and protein expression levels in the hippocampus and PFC. Additionally, studies by Sun et al. (2022a) reported that manual acupuncture stimulation increases blood BDNF concentrations, as well as BDNF protein and mRNA expression in the hippocampal CA1 area, in a poststroke depression (PSD) rat model. Kawanokuchi et al. (2021a) demonstrated that the neuroplasticity mechanism underlying acupuncture stimulation could be related to its regulatory effects on the expression of neurotrophic factors. In a social defeat stress rat model of depression, acupuncture was found to restore BDNF, neurotrophin (NT)-3, and NT-4/5 production while simultaneously suppressing nerve growth factor expression in the brain.
Cyclic AMP response-binding protein (CREB) is a transcription factor (Zhang et al., 2005) that activates the transcription of various target genes and serves as a critical regulator of BDNF-related gene expression (Finkbeiner et al., 1997). The intracellular TrkB signaling cascade facilitates CREB activation (Zarneshan et al., 2022), which in turn enhances BDNF gene transcription and promotes BDNF expression (Esvald et al., 2020), playing a pivotal role in LTP and synaptic plasticity (Tartt et al., 2022). Tong et al. (2023a) demonstrated that acupuncture may exert antidepressant-like effects by activating the BDNF/TrkB/CREB signaling pathway and decreasing the expression of pro-BDNF in the lateral habenula (LHb). Additionally, acupuncture increases the number of BDNF- and TrkB-positive cells in the brains of PSD rats (Kang et al., 2021a). Tissue plasminogen activator (tPA) regulates the balance between BDNF and pro-BDNF through activating the extracellular protease plasmin (Lee et al., 2001; Liang et al., 2018; Nagappan et al., 2009), which is part of the cleavage pathway that converts pro-BDNF to BDNF. Studies suggest that the tPA/BDNF/TrkB signaling pathway may contribute to the neuroplastic effects of EA on rats by increasing BDNF levels in the hippocampus and PFC (Dong et al., 2021; Luo et al., 2020b) and in the serum. The HE staining results corroborated these findings, showing changes in the morphology and quantity of neurons in the hippocampal CA3 area. However, the study (Luo et al., 2020) also noted lower BDNF content in the raphe nuclei, suggesting differential targeting across brain regions, which warrants further exploration and reflection.
The MAPK cascade, which includes the extracellular signal-regulated kinase (ERK) pathway, is a critical signal transduction pathway activated downstream of the BDNF-activated TrkB receptor (Sweatt, 2004). ERK plays a key role in regulating cellular fates such as growth, proliferation, differentiation, and survival (Ryu et al., 2015; Wang and Mao, 2019). The antidepressant effects of EA may be related to alterations in the extracellular microenvironment of hippocampal neural stem cells (NSCs) and the activation of ERK signaling pathways, which also contribute to the beneficial effects on promoting NSC proliferation (Yang et al., 2013). ERK1/2 is one of several ERK isoforms that has been thoroughly investigated. Research indicates that acupuncture upregulates the expression of p-ERK1/2 and BDNF in the PFC in a depression rat model (Li et al., 2018). The transcription factor CREB serves as a downstream target of ERK. Chronic stress reduces ERK and CREB phosphorylation (activation) in the rat hippocampus and PFC (Qi et al., 2008). Lu et al. (2013) demonstrated that acupuncture increases the level of p-ERK1/2 in the hippocampus and PFC and enhances p-CREB levels in the hippocampus in a depression rat model, suggesting that the ERK-CREB pathway is upregulated in the hippocampus. Additionally, the protein kinase A (PKA)/CREB signaling pathway, which regulates synaptic plasticity and learning memory, involves PKA as the upstream activator of CREB (Hu et al., 2012; Shaywitz and Greenberg, 1999). Jiang et al. (2017) reported that acupuncture upregulates PKA-α and p-CREB expression in the hippocampus, indicating that the mechanisms underlying the antidepressant effect of acupuncture may be related to the regulation of the ERK-CREB and PKA/CREB signaling pathway in the hippocampus.
The c-Jun N-terminal kinases (JNKs), a critical member of the MAPK family (Zeke et al., 2016), mediate cellular responses to a range of abiotic and biotic stressors (de Los Reyes Corrales et al., 2021). The JNK pathway is activated in response to various stress events, such as infection, inflammation, or oxidative stress. Research indicates that JNK signaling activation occurs in animal models of depression (Adzic et al., 2009; Egeland et al., 2015; Rogatsky et al., 1998) and that inhibition of the JNK pathway leads to increased neurogenesis and alleviation of depressive and anxiety-like behaviors (Mohammad et al., 2018). In support of this model, Liu et al. (2023a) reported that acupuncture downregulates the protein expression levels of c-JUN and p-JNK in the hippocampal CA1, CA3, and DG regions, suggesting that, at least in part, acupuncture exerts an antidepressant effect through regulating MAPK/JNK signaling.
Serotonin (5-hydroxytryptamine, 5-HT) is a monoamine neurotransmitter that has diverse functions (Sirek and Sirek, 1970). The differential physiological effects of 5-HT are mediated by the activation of any of the 15 different serotoninergic receptors, which are categorized into 7 different classes (5-HT1 to 5-HT7) (Hoyer et al., 2002). In the CNS, serotonin is primarily synthesized locally in the raphe nucleus (David and Gardier, 2016), especially the dorsal raphe (DR) (Mahar et al., 2014). Although only a minute proportion of the body’s 5-HT (approximately 5%) is found in the mature mammalian brain (Fouquet et al., 2019), it significantly influences neuronal networks during development and modulates various critical neuronal functions (Lesch and Waider, 2012), such as perception, cognitive activities and emotional responses, particularly mood regulation in the mature brain. Serotonin remains a critical neurotransmitter in the CNS with notable neuroplastic capabilities (Kraus et al., 2017), which mainly attributed to interactions between serotonergic receptors and neurotrophic proteins (Mattson et al., 2004), intracellular signaling cascades (Rantamäki and Castrén, 2008) involved in cytoskeletal rearrangement, and the modulation of cell adhesion molecules and glutamatergic transmission (Dalva et al., 2007; Sanacora et al., 2012). The dopamine (DA) system is integral to many aspects of brain function, including locomotion, affect, and cognition (Grace, 2016). Both DA and 5-HT are critical neuromodulators of synaptic plasticity but often play antagonistic roles during reward-driven learning (Wert-Carvajal et al., 2022). For example, DA enhances the induction of LTP in the hippocampus (Broussard et al., 2016; Brzosko et al., 2015). In contrast, 5-HT has been shown to induce long-term depression (LTD) in certain receptor-specific regions of the hippocampus (Kemp and Manahan-Vaughan, 2004; Lecouflet et al., 2021).
Serotonergic imbalance plays a critical role in the pathogenesis of MDD. Aversive external stimuli, such as stress, cause excessive activation of the endocrine immune system (Beurel et al., 2020). The resulting inflammatory response may trigger depression in susceptible individuals by reducing plasma tryptophan levels and diminishing brain serotonin activity (Cowen and Browning, 2015; Wichers et al., 2005). Chronic stress significantly attenuates 5-HT neurotransmission and 5-HT1A autoreceptor sensitivity (Mahar et al., 2014). Additionally, dysfunction in the DA system has also been implicated in the pathophysiology of depression. Preclinical studies using chronic unpredictable mild stress (CUMS) (Chang and Grace, 2014) or learned helplessness models (Belujon and Grace, 2014) of depression have revealed a reduction in the number of spontaneously firing DAergic neurons in the ventral tegmental area, indicating that chronic stress induces plastic changes that diminish the activity of the DAergic neuron population.
Acupuncture has been demonstrated to increase the levels of 5-HT, norepinephrine (NE), and dopamine (DA) in the hippocampus (Sun et al., 2019a), which helps to repair hippocampal neuronal damage and restore neuronal plasticity in PSD rats. One study highlighted that the antidepressant mechanism of EA may be linked to the promotion of 5-HT1A receptor mRNA and protein expression in the hippocampus (Chen et al., 2020b), thereby reversing pathological changes in hippocampal neurons. Additionally, acupuncture has been shown to enhance the expression of the 5-HT1A receptor in the cortex, hippocampus, thalamus, and hypothalamus and the 5-HT1B receptor in the cortex and thalamus (Lee et al., 2019). This provides further evidence that acupuncture can ameliorate alterations in the 5-HT system associated with depression. Acupuncture also promotes cell proliferation in the dentate gyrus of the hippocampus and increases 5-HT levels in the dorsal raphe (Park and Lim, 2019). However, one study presents different view point, it suggested that EA could alleviate depressive-like behaviors by reversing impairment in synaptic plasticity within the hippocampal CA1 region in Wistar Kyoto (WKY) depressive model rats, predominantly through the downregulation of serotonin transporter (5-HTT) and 5-HT1A receptor levels (Han et al., 2018a). Emerging evidence indicates abnormalities in the function of monoamine receptors and transporters in the brain of WKY rat (El Mansari et al., 2023). Consequently, this study holds limited significance regarding changes in 5-HT-related indicators.
Furthermore, research on dopamine has shown that EA upregulates dopaminergic signaling in the PFC, as evidenced by increased expression of a critical enzyme in this pathway, aromatic L-amino acid decarboxylase (DDC) (Zhang et al., 2021a). Studies have also shown that EA promotes the activation of the dopamine transporter (DAT), which plays a crucial role in maintaining dopamine stability in the synaptic cleft, and improves synaptic transmission in the ventromedial PFC by upregulating spontaneous excitatory postsynaptic current amplitude (Cai et al., 2023). The potential mechanism for these effects may be related to the activation of the trace amine-associated receptor 1 (TAAR1)/cyclic adenosine monophosphate (cAMP)/protein kinase A (PKA) signaling pathway, as there is evidence that TAAR1 activation has antidepressant potential and can modulate DAT function or quantity through TAAR1 signaling (Espinoza et al., 2018; Rutigliano and Zucchi, 2020).
Glutamate (Glu) serves as the primary excitatory neurotransmitter in the CNS (Duman et al., 2019) and is indispensable for a broad range of behaviors, including learning and memory, emotional responses, sensory input and integration, and motor system activity (Mattson et al., 2018). Through its specific receptor subtypes, namely, N-methyl-d-aspartate receptors (NMDARs) and α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid receptors (AMPARs), glutamate plays a crucial role in regulating synaptic and neuronal plasticity, which in turn influences fundamental human processes such as mood, cognition, learning, and reward (Murrough et al., 2017). Under normal conditions, moderate levels of NMDA receptor activation are beneficial, promoting neuroprotective signaling pathways. This includes the activation of the RAS-mitogen-activated protein kinase (RAS-MAPK) pathway and CREB-mediated induction of survival gene (Krishnan and Nestler, 2008). Under pathological conditions, however, glutamate-mediated excitotoxicity via the action of extrasynaptic NMDARs is known to be a potent neuronal excitotoxin, inducing either rapid or delayed neurotoxicity (Sanacora et al., 2008).
Clinical studies employing in vivo proton magnetic resonance spectroscopy (MRS) have consistently revealed a reduction in glutamate metabolite concentrations in the medial prefrontal cortex (mPFC) (Moriguchi et al., 2019) of patients with depression. This finding is supported by postmortem studies that reported alterations in glutamate receptor subtypes within PFC subregions of depressed subjects compared to controls (Feyissa et al., 2009; Gray et al., 2015), highlighting abnormalities in glutamatergic transmission in major depression patients. Additionally, morphological studies have revealed that chronic stress reduces the structure and function of glutamate pyramidal neurons in the mPFC and hippocampus, providing further evidence of glutamate neuronal atrophy in these regions.
Preclinical research has indicated that CUMS for 28 days leads to significant decreases in the levels of 5-HT, Glu, and gamma-aminobutyric acid (GABA) in the hippocampus. This finding is consistent with findings from 13C-MRS studies (Banasr et al., 2010) demonstrating that chronic stress exposure reduces the cycling and metabolism of glutamate and glutamine, as well as GABA, in the PFC of rats. EA for 14 days has been shown to reverse this trend by increasing the levels of these neurotransmitters in the hippocampus (Duan et al., 2016). In the context of CUMS, EA reportedly enhances the expression of the glial glutamate transporter EAAT2 in both the hippocampus and PFC, alleviating depressive-like behaviors (Luo et al., 2017a). Moreover, EA has been validated to increase the expression of AMPA glutamate receptor (AMPAR) and AMPAR-related proteins, such as glutamate receptor 1 (GluR1), glutamate receptor 2 (GluR2), and Stargazin, and protein interacting with C kinase 1 (Pick1) in the hippocampus. These modifications in synaptic strength and synaptic responses significantly impact synaptic plasticity (Lu et al., 2009; Tomita et al., 2005). EA also promotes neural plasticity by upregulating synapse-related proteins, including synaptophysin (SYN), postsynaptic density protein-95 (PSD-95), and growth-associated protein-43 (GAP-43), in the hippocampus (Jiang et al., 2020). The neuroplasticity effects of EA were confirmed by Nissl staining, which revealed an increase in synapses and a reduction in synaptic clefts in the hippocampus. The functional properties of NMDARs are determined by their subunit composition (Cull-Candy et al., 2001). At synaptic sites, NMDARs typically contain GluN2A subunits, which mediate long-term synaptic plasticity. In contrast, extra-synaptic NMDARs are generally enriched in GluN2B subunits, and are thought to constitute a major signaling pathway that triggers neuronal death (Tian et al., 2021). Studies have suggested (Zhang et al., 2021b) that EA ameliorates depression-like behaviors, potentially through its influence on synaptic plasticity by reducing GluN2B levels. This reduction may inhibit the overactivation of GluN2B-containing NMDARs and increase the expression of synaptic plasticity-related proteins including microtubule-associated protein 2 (MAP-2), PSD-95, and SYN in the hippocampus. Additionally, there is evidence that EA increased NR2A expression level as well as decreased NR2B expression level in the hippocampus (Guo et al., 2021), leading to an affirmative conclusion that EA promotes synaptic plasticity in the hippocampus of depression-modeled rats. Research examining the effect of acupuncture on glutamate levels in the brain has yielded mixed results. In contrast to the previous finding, one study suggest that EA decreases hippocampal glutamate levels, leading to varying conclusions that necessitate further verification (Guo et al., 2021). We carefully reviewed the protocol in these two articles (Duan et al., 2016; Guo et al., 2021) and found that both studies used male Sprague–Dawley rats for a four-week CUMS modeling process and utilized high-performance liquid chromatography (HPLC) to detect hippocampal neurotransmitters, with no notable methodological differences. However, the modeling outcomes of these two studies vary significantly: one study revealed a decrease in glutamate levels due to CUMS, whereas the other reported an increase. We found through literature review that there is considerable diversity in the early findings reported alterations of glutamate in blood, CSF, and brain tissue. Acute stress increases extracellular glutamate in the mPFC and hippocampus, while studies demonstrate that chronic stress exposure decreases the cycling and metabolism of glutamate and glutaminevin rat PFC (Duman et al., 2019). The discrepancies in the results observed between these two studies may be due to variations in the intensity of the CUMS modeling process. Future research endeavors should meticulously consider this factor, ensuring uniformity and comparability in the modeling intensity across studies to enable the derivation of more precise and dependable conclusions. Furthermore, it is notable that, emerging evidence shows deficits in glutamatergic signaling in the brain of WKY rat (Millard et al., 2020). Future experiments should carefully consider the suitability of using this variety of mice for the relevant research.
Astrocytes are abundant (Freeman, 2010) and intricately structured glial cells in the central nervous system (CNS) (Oberheim et al., 2009; Stogsdill et al., 2017). They form extensive contacts with other brain cells and perform diverse functions, including ion and neurotransmitter homeostasis, synapse formation, modulation, function and elimination (Endo et al., 2022). Astrocytes are closely associated with synapses, as they infiltrate the neuropil via numerous cellular processes to interact with thousands of synapses and actively participate in synapse formation and plasticity through various secreted and contact-mediated signals (Clarke and Barres, 2013; Stogsdill et al., 2017). They also play a crucial role in regulating glutamate neurotransmission (Sanacora and Banasr, 2013) by actively regulating the uptake, metabolism, and recycling of glutamate (Rajkowska and Stockmeier, 2013), thus preventing synaptic spillover and excitotoxicity (Lener et al., 2017).
Emerging evidence suggests that the neuropathology of MDD is characterized by notable reductions in astrocyte density and the expression of astrocyte markers without evident neuronal loss in both patients with MDD and depressive model mice (Rajkowska and Stockmeier, 2013). Postmortem studies have reported a decrease in glial cell density and number, specifically in certain frontal-limbic regions, in subjects with MDD but not in senior citizens (Bowley et al., 2002; Cotter et al., 2002; Gittins and Harrison, 2011; Khundakar et al., 2011a; Khundakar et al., 2011b; Ongür et al., 1998; Rajkowska et al., 1999). Reductions in the protein and mRNA expression of astrocyte markers, particularly glial fibrillary acidic protein (GFAP), as indicated by several preclinical studies and postmortem studies (Araya-Callís et al., 2012; Banasr et al., 2010; Bernard et al., 2011; Chandley et al., 2013; Choudary et al., 2005; Ernst et al., 2011; Johnston-Wilson et al., 2000; Klempan et al., 2009; Sequeira et al., 2009; Si et al., 2004; Sun et al., 2012; Webster et al., 2005), suggest a significant reduction in the number of astrocytes. Moreover, stress leads to diminished secretion of neurotrophic factors and increased production of cytokines (Schmidt et al., 2011) by astrocytes, as well as a reduced astrocytic response to neuronal injury (Laping et al., 1994). This collectively contributes to disrupted neuroplasticity and cellular resilience (Manji et al., 2000; Pittenger and Duman, 2008) in depression.
Preclinical studies have shown that EA prevents astrocyte atrophy in the prefrontal cortex and alleviates depressive-like behavior in mice subjected to CUMS (Lin et al., 2023a), providing experimental evidence that EA enhances the presence of astrocytes in the active milieu of the brain. Fibroblast growth factor 2 (FGF2) is a pleiotropic protein involved in regulating a myriad of cellular processes, including the proliferation, differentiation, and survival of various cell types (Belov and Mohammadi, 2013; Bikfalvi et al., 1997; Krzyscik et al., 2022; Xie et al., 2020). EA has been shown to upregulate the expression of FGF2 in the hippocampus and increase both the protein expression of the astrocyte marker GFAP and the mean optical density of GFAP-immunoreactive astrocytes (Yao et al., 2021a). Considering that FGF2 knockdown significantly reduces astrocyte proliferation and induces astrocyte apoptosis (Yao et al., 2021a), it is suggested that EA maintains astrocyte homeostasis by modulating FGF2 expression. Additionally, animal research has demonstrated that EA enhances GFAP mRNA and protein expression in the hippocampus (Liu et al., 2011), indicating its role in preventing hippocampal glial atrophy. Other studies corroborate these findings, reporting that acupuncture therapy increases hippocampal GFAP protein expression while decreasing it in the prefrontal cortex (Dong et al., 2018).
Microglia are the principal innate immune cells that reside in the central nervous system (CNS) (Nayak et al., 2014). These cells act as crucial sentinels, maintaining CNS homeostasis and responding swiftly to damage or infection (Beurel et al., 2020). In their quiescent state, microglia secrete trophic factors that are essential for neuronal development, maintenance, and function throughout life. Their phagocytic activity is also critical, as they participate in the clearance of dead cells, as well as in synaptogenesis and synaptic pruning—processes vital for maintaining normal brain homeostasis. Even under resting conditions, the processes of microglia are highly dynamic; they perpetually scan their surroundings and communicate directly with neurons, astrocytes, and blood vessels. When faced with damage, inflammation, or other pathological changes, microglia undergo transformations, activate inflammatory functions, and initiate genetic programs designed to address and repair CNS insults.
In pathological states, abnormalities in microglia contribute significantly to the pathology of depression, primarily through the promotion of neuroinflammation (Yirmiya et al., 2015). This is often related to stress-induced activation of the sympathetic nervous system and the HPA axis (Ader et al., 1995; Won and Kim, 2016), which increase circulating glucocorticoids (de Kloet et al., 2008; Hunter et al., 2016) and subsequently activate the immune system. This activation triggers inflammatory responses from proinflammatory microglia, resulting in increased release of proinflammatory cytokines. Studies have highlighted the critical role of microglia-related neuroinflammation and neuronal atrophy in depression (Price and Duman, 2020).
Recent research has focused primarily on clarifying the mechanisms by which acupuncture reduces neuroinflammation. Li et al. (2021a) discovered that manual acupuncture therapy reversed the elevated levels of interleukin-1beta (IL-1beta), interleukin-6 (IL-6), and the microglial marker ionized calcium-binding adaptor molecule 1 (Iba-1) and decreased the gene expression of triggering receptor expressed on myeloid cells 2 (TREM2) in the PFC. These findings suggest that acupuncture mitigates neuroinflammation by inhibiting microglial overactivation, reducing the expression of proinflammatory cytokines, and enhancing TREM2 expression in the PFC. High mobility group box-1 (HMGB1) is recognized as an endogenous risk factor and initiating signal for neuroinflammation (Weber et al., 2015; Zandarashvili et al., 2013) and is actively released by microglia and neurons under CUMS conditions (Rana et al., 2021; Wang et al., 2020). Chen et al. (2022) reported that acupuncture alleviates neuroinflammation by downregulating HMGB1 expression and microglial activation in the hippocampus and reducing tumor necrosis factor-α (TNF-α) levels in the serum.
Several animal studies have shown that acupuncture can prevent neuronal apoptosis or autophagy. Acupuncture intervention significantly reduces oxidative stress markers, such as reactive oxygen species (ROS) and H2O2, by upregulating the nuclear factor E2-related factor 2 (Nrf2)/haem oxygenase-1 (HO-1) signaling pathway. This pathway plays a crucial role in the cerebral antioxidant system (Suzuki and Yamamoto, 2015), thus preventing neuronal apoptosis (Cheng et al., 2021). Chronic stress results in elevated levels of ROS, which are intimately linked to oxidative stress and subsequently trigger the activation of the mitochondrial apoptosis pathway. Acupuncture effectively reduces ROS levels and the expression of key factors in the apoptosis pathway, including cytochrome C, cysteine-containing aspartate-specific protease-3 (caspase-3), and apoptosis-inducing factor (AIF) proteins, in the hippocampus (Sun et al., 2019c).
EA partially inhibits autophagy by reducing the number and size of autolysosomes and decreasing the levels of the autophagic biomarker light chain 3 (LC3) and the LC3-II/LC3-I ratio in hippocampal CA1 neurons (Zhang et al., 2020). Mammalian target of rapamycin (mTOR) is an autophagy inhibitory kinase, and its activation leads to the suppression of cellular autophagy. The phosphatidylinositol 3-kinase (PI3K)/protein kinase B (Akt)/mTOR signaling pathway plays a pivotal regulatory role in autophagy. Acupuncture promotes the activation of the PI3K/Akt/mTOR signaling pathway, thereby inhibiting hippocampal neuron autophagy. This is evidenced by decreased expression of Beclin1, LC3B-II/I, and LC3B-II in the CA1 region of the hippocampus (Sun et al., 2020). Beclin1 is a critical factor in the activation of autophagy, LC3 serves as a reliable marker of autophagosomes, and the LC3B-II/I ratio reflects the level of cellular autophagy. Furthermore, EA enhances the proliferation of amplifying neural progenitor cells (ANPs) while suppressing the apoptosis of quiescent neural progenitor cells (QNPs) in the hippocampal dentate gyrus (Yang et al., 2014).
PNNs are structures within the extracellular matrix of the central nervous system that play crucial roles in synaptic plasticity and protection against external oxidative stress (Hirono et al., 2018). Research has shown that EA enhances the expression of PNN, GABA synthetase glutamic acid decarboxylase 67 (GAD67), and excitatory synaptic proteins, including GLuR1 and PSD-95, in the mPFC of rats subjected to CUMS (Zhang et al., 2023). These findings confirm the antidepressant effects of EA on synaptic plasticity.
Further studies have demonstrated that EA increases the spine density on specific dendrites of layer V pyramidal neurons and increases the expression of synaptic proteins such as BDNF, GluR1, GluR2, PSD-95, and synaptophysin I in functional areas of the PFC (Gao et al., 2021). This finding suggested that the mechanism by which EA ameliorates depressive-like behaviors is related, at least in part, to the promotion of synaptic plasticity. Additionally, findings indicate a decrease in acetylcholine expression and an increase in acetylcholinesterase expression in the PFC, suggesting that modulation of the hypercholinergic tone may contribute to the antidepressant and neuroplastic effects of EA. This area presents new research directions and requires further exploration.
Growing clinical and preclinical evidence supports the effectiveness of acupuncture as a complementary and integrative therapy for depression. The modulatory effect of acupuncture may derive from both acupoint specificity and electrical stimuli, highlighting the complexity of this therapeutic method. As a traditional medical technique, acupuncture demonstrates considerable variability in acupoint selection, manipulative techniques, and stimulation intensity. Different acupuncturists may employ different acupuncture strategies, and even the same practitioner may modify the acupuncture protocol based on timing or patient response heterogeneity. This makes it a challenge to quantify the treatment. Furthermore, the current trend in clinical practice is to integrate acupuncture and mainstream medicine. In treating depression, acupuncture serve as a supplementary and alternative approach, improve symptoms of patients, and enhance the efficacy of antidepressant medications or other therapies. The aforementioned emphasizes the complexity of acupuncture and the challenges for quantify this treatment, and the requirement of more rigorous approaches to investigate the underlying mechanisms of acupuncture.
This review has summarized numerous potential targets related to neuroplasticity that contribute to the antidepressant effect of acupuncture. These targets include the modulation of neurotrophic factors and their receptors, neurotransmitters (primarily within the monoaminergic and glutamatergic system), growth factors, and glia (primarily within astrocytes and microglia). It is important to acknowledge that there are many other potential mechanisms involved in the pathophysiology of depression that were not captured in this paper but may be modulated by acupuncture interventions, such as effects on inflammation, the HPA axis, the gut microbiota, the gene expression of neuropeptides, mitochondrial biogenesis, and extracellular ATP levels. Although this review focused primarily on the interplay between acupuncture, neuroplasticity, and depression in the hippocampus and PFC, it did not fully explore the diversity of plasticity across different brain regions that vary according to brain circuits. Thus, the conclusions presented should be further substantiated with clinical research employing more advanced diagnostic tools such as diffusion tensor imaging and proton magnetic resonance spectroscopy.
NX: Conceptualization, Writing – original draft. YH: Writing – original draft. Y-NW: Writing – original draft. LB: Writing – review & editing. LW: Funding acquisition, Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by the National Natural Science Foundation (81303044), the Natural Science Foundation of Heilongjiang Province (LH2022H082), and the Postdoctoral Scientific Research Developmental Fund of Heilongjiang Province (LBH-Q19185).
DNA and microglia cartoon image used in figure were obtained from Scidraw.io. https://scidraw.io/.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Ader, R., Cohen, N., and Felten, D. (1995). Psychoneuroimmunology: interactions between the nervous system and the immune system. Lancet 345, 99–103. doi: 10.1016/S0140-6736(95)90066-7
Adzic, M., Djordjevic, J., Djordjevic, A., Niciforovic, A., Demonacos, C., Radojcic, M., et al. (2009). Acute or chronic stress induce cell compartment-specific phosphorylation of glucocorticoid receptor and alter its transcriptional activity in Wistar rat brain. J. Endocrinol. 202, 87–97. doi: 10.1677/JOE-08-0509
Aleksandrova, L. R., and Phillips, A. G. (2021). Neuroplasticity as a convergent mechanism of ketamine and classical psychedelics. Trends Pharmacol. Sci. 42, 929–942. doi: 10.1016/j.tips.2021.08.003
Araya-Callís, C., Hiemke, C., Abumaria, N., and Flugge, G. (2012). Chronic psychosocial stress and citalopram modulate the expression of the glial proteins GFAP and NDRG2 in the hippocampus. Psychopharmacology 224, 209–222. doi: 10.1007/s00213-012-2741-x
Banasr, M., Chowdhury, G. M., Terwilliger, R., Newton, S. S., Duman, R. S., Behar, K. L., et al. (2010). Glial pathology in an animal model of depression: reversal of stress-induced cellular, metabolic and behavioral deficits by the glutamate-modulating drug riluzole. Mol. Psychiatry 15, 501–511. doi: 10.1038/mp.2008.106
Belov, A. A., and Mohammadi, M. (2013). Molecular mechanisms of fibroblast growth factor signaling in physiology and pathology. Cold Spring Harb. Perspect. Biol. 5, 239–249. doi: 10.1101/cshperspect.a015958
Belujon, P., and Grace, A. A. (2014). Restoring mood balance in depression: ketamine reverses deficit in dopamine-dependent synaptic plasticity. Biol. Psychiatry 76, 927–936. doi: 10.1016/j.biopsych.2014.04.014
Bernard, R., Kerman, I. A., Thompson, R. C., Jones, E. G., Bunney, W. E., Barchas, J. D., et al. (2011). Altered expression of glutamate signaling, growth factor, and glia genes in the locus coeruleus of patients with major depression. Mol. Psychiatry 16, 634–646. doi: 10.1038/mp.2010.44
Beurel, E., Toups, M., and Nemeroff, C. B. (2020). The bidirectional relationship of depression and inflammation: double trouble. Neuron 107, 234–256. doi: 10.1016/j.neuron.2020.06.002
Bikfalvi, A., Klein, S., Pintucci, G., and Rifkin, D. B. (1997). Biological roles of fibroblast growth factor-2. Endocr. Rev. 18, 26–45
Boku, S., Nakagawa, S., Toda, H., and Hishimoto, A. (2018). Neural basis of major depressive disorder: beyond monoamine hypothesis. Psychiatry Clin. Neurosci. 72, 3–12. doi: 10.1111/pcn.12604
Bowley, M. P., Drevets, W. C., Ongür, D., and Price, J. L. (2002). Low glial numbers in the amygdala in major depressive disorder. Biol. Psychiatry 52, 404–412. doi: 10.1016/S0006-3223(02)01404-X
Broussard, J. I., Yang, K., Levine, A. T., Tsetsenis, T., Jenson, D., Cao, F., et al. (2016). Dopamine regulates aversive contextual learning and associated in vivo synaptic plasticity in the Hippocampus. Cell Rep. 14, 1930–1939. doi: 10.1016/j.celrep.2016.01.070
Brzosko, Z., Schultz, W., and Paulsen, O. (2015). Retroactive modulation of spike timing-dependent plasticity by dopamine. eLife 4:e09685. doi: 10.7554/eLife.09685
Cai, X., Wu, M., Zhang, Z., Liu, H., Huang, S., Song, J., et al. (2023). Electroacupuncture alleviated depression-like behaviors in ventromedial prefrontal cortex of chronic unpredictable mild stress-induced rats: increasing synaptic transmission and phosphorylating dopamine transporter. CNS Neurosci. Ther. 29, 2608–2620. doi: 10.1111/cns.14200
Castrén, E., and Monteggia, L. M. (2021). Brain-derived neurotrophic factor signaling in depression and antidepressant action. Biol. Psychiatry 90, 128–136. doi: 10.1016/j.biopsych.2021.05.008
Chandley, M. J., Szebeni, K., Szebeni, A., Crawford, J., Stockmeier, C. A., Turecki, G., et al. (2013). Gene expression deficits in pontine locus coeruleus astrocytes in men with major depressive disorder. J. Psychiatry Neurosci. 38, 276–284. doi: 10.1503/jpn.120110
Chang, C. H., and Grace, A. A. (2014). Amygdala-ventral pallidum pathway decreases dopamine activity after chronic mild stress in rats. Biol. Psychiatry 76, 223–230. doi: 10.1016/j.biopsych.2013.09.020
Chen, W., Chen, Y., Cheng, W., Li, P., Shen, J., Tong, T., et al. (2023). Acupuncture exerts preventive effects in rats of chronic unpredictable mild stress: the involvement of inflammation in amygdala and brain-spleen axis. Biochem. Biophys. Res. Commun. 646, 86–95. doi: 10.1016/j.bbrc.2023.01.046
Chen, L., Jiang, H., Bao, T., Wang, Y., Meng, H., Sun, Y., et al. (2022). Acupuncture ameliorates depressive behaviors by modulating the expression of hippocampal Iba-1 and HMGB1 in rats exposed to chronic restraint stress. Front. Psych. 13:903004. doi: 10.3389/fpsyt.2022.903004
Chen, L., Yao, Z., Qu, S., Zhang, J., Zhang, J., Zhang, Z., et al. (2020b). Electroacupuncture improves synaptic plasticity by regulating the 5-HT1A receptor in hippocampus of rats with chronic unpredictable mild stress. J. Int. Med. Res. 48:300060520918419. doi: 10.1177/0300060520918419
Cheng, W. J., Li, P., Huang, W. Y., Huang, Y., Chen, W. J., Chen, Y. P., et al. (2021). Acupuncture relieves stress-induced depressive behavior by reducing oxidative stress and Neuroapoptosis in rats. Front. Behav. Neurosci. 15:783056. doi: 10.3389/fnbeh.2021.783056
Choudary, P. V., Molnar, M., Evans, S. J., Tomita, H., Li, J. Z., Vawter, M. P., et al. (2005). Altered cortical glutamatergic and GABAergic signal transmission with glial involvement in depression. Proc. Natl. Acad. Sci. USA 102, 15653–15658. doi: 10.1073/pnas.0507901102
Clarke, L. E., and Barres, B. A. (2013). Emerging roles of astrocytes in neural circuit development. Nat. Rev. Neurosci. 14, 311–321. doi: 10.1038/nrn3484
Colucci-D’Amato, L., Speranza, L., and Volpicelli, F. (2020). Neurotrophic factor BDNF, physiological functions and therapeutic potential in depression, neurodegeneration and brain Cancer. Int. J. Mol. Sci. 21:21. doi: 10.3390/ijms21207777
Cotter, D., Mackay, D., Chana, G., Beasley, C., Landau, S., and Everall, I. P. (2002). Reduced neuronal size and glial cell density in area 9 of the dorsolateral prefrontal cortex in subjects with major depressive disorder. Cerebral Cortex 12, 386–394. doi: 10.1093/cercor/12.4.386
Cowen, P. J., and Browning, M. (2015). What has serotonin to do with depression? World Psychiatry 14, 158–160. doi: 10.1002/wps.20229
Cramer, S. C., Sur, M., Dobkin, B. H., O’Brien, C., Sanger, T. D., Trojanowski, J. Q., et al. (2011). Harnessing neuroplasticity for clinical applications. Brain 134, 1591–1609. doi: 10.1093/brain/awr039
Cull-Candy, S., Brickley, S., and Farrant, M. (2001). NMDA receptor subunits: diversity, development and disease. Curr. Opin. Neurobiol. 11, 327–335. doi: 10.1016/S0959-4388(00)00215-4
Dai, W., Li, W. D., and Lu, J. (2010). Effect of electroacupuncture on hippocampal apoptosis and JNK signal pathway in chronic stress depression rats. Zhen ci yan jiu 35, 330–334
Dalva, M. B., McClelland, A. C., and Kayser, M. S. (2007). Cell adhesion molecules: signalling functions at the synapse. Nat. Rev. Neurosci. 8, 206–220. doi: 10.1038/nrn2075
David, D. J., and Gardier, A. M. (2016). The pharmacological basis of the serotonin system: application to antidepressant response. L’Encephale 42, 255–263. doi: 10.1016/j.encep.2016.03.012
de Kloet, E. R., Karst, H., and Joëls, M. (2008). Corticosteroid hormones in the central stress response: quick-and-slow. Front. Neuroendocrinol. 29, 268–272. doi: 10.1016/j.yfrne.2007.10.002
de Los Reyes Corrales, T., Losada-Pérez, M., and Casas-Tintó, S. (2021). JNK pathway in CNS pathologies. Int. J. Mol. Sci. 22:3883. doi: 10.3390/ijms22083883
Dong, S., Jiang, H.-L., Wang, Y., Lu, J., Chang, L., Zhang, P., et al. (2018). Effect of acupuncture on expression of glial fibrillary acidic protein in Hippocampus and prefrontal cortex and serum Interleukin-10 in chronic restraint stress depression rats. Zhen ci yan jiu 43, 209–214. doi: 10.13702/j.1000-0607.170676
Dong, H., Qin, Y.-Q., Sun, Y.-C., Yao, H.-J., Cheng, X.-K., Yu, Y., et al. (2021). Electroacupuncture ameliorates depressive-like behaviors in Poststroke rats via activating the tPA/BDNF/TrkB pathway. Neuropsychiatr. Dis. Treat. 17, 1057–1067. doi: 10.2147/NDT.S298540
Duan, D. M., Tu, Y., Liu, P., and Jiao, S. (2016). Antidepressant effect of electroacupuncture regulates signal targeting in the brain and increases brain-derived neurotrophic factor levels. Neural Regen. Res. 11, 1595–1602. doi: 10.4103/1673-5374.193238
Duman, R. S., Aghajanian, G. K., Sanacora, G., and Krystal, J. H. (2016). Synaptic plasticity and depression: new insights from stress and rapid-acting antidepressants. Nat. Med. 22, 238–249. doi: 10.1038/nm.4050
Duman, R. S., Sanacora, G., and Krystal, J. H. (2019). Altered connectivity in depression: GABA and glutamate neurotransmitter deficits and reversal by novel treatments. Neuron 102, 75–90. doi: 10.1016/j.neuron.2019.03.013
Egeland, M., Zunszain, P. A., and Pariante, C. M. (2015). Molecular mechanisms in the regulation of adult neurogenesis during stress. Nat. Rev. Neurosci. 16, 189–200. doi: 10.1038/nrn3855
El Mansari, M., Hamoudeh, R., Daniels, S., and Blier, P. (2023). Wistar Kyoto rats exhibit decreased serotonin neuronal firing and increased norepinephrine burst activity but dampened hippocampal α(2)-adrenoceptor sensitivity. J. Psychopharmacol. 37, 1105–1115. doi: 10.1177/02698811231209235
Endo, F., Kasai, A., Soto, J. S., Yu, X., Qu, Z., Hashimoto, H., et al. (2022). Molecular basis of astrocyte diversity and morphology across the CNS in health and disease. Science 378:9020. doi: 10.1126/science.adc9020
Eriksson, P. S., Perfilieva, E., Björk-Eriksson, T., Alborn, A. M., Nordborg, C., Peterson, D. A., et al. (1998). Neurogenesis in the adult human hippocampus. Nat. Med. 4, 1313–1317. doi: 10.1038/3305
Ernst, C., Nagy, C., Kim, S., Yang, J. P., Deng, X., Hellstrom, I. C., et al. (2011). Dysfunction of astrocyte connexins 30 and 43 in dorsal lateral prefrontal cortex of suicide completers. Biol. Psychiatry 70, 312–319. doi: 10.1016/j.biopsych.2011.03.038
Espinoza, S., Leo, D., Sotnikova, T. D., Shahid, M., Kääriäinen, T. M., and Gainetdinov, R. R. (2018). Biochemical and functional characterization of the trace amine-associated receptor 1 (TAAR1) agonist RO5263397. Front. Pharmacol. 9:645. doi: 10.3389/fphar.2018.00645
Esvald, E. E., Tuvikene, J., Sirp, A., Patil, S., Bramham, C. R., and Timmusk, T. (2020). CREB family transcription factors are major mediators of BDNF transcriptional autoregulation in cortical neurons. J. Neurosci. 40, 1405–1426. doi: 10.1523/JNEUROSCI.0367-19.2019
Evans, J. W., Szczepanik, J., Brutsché, N., Park, L. T., Nugent, A. C., and Zarate, C. A. Jr. (2018). Default mode connectivity in major depressive disorder measured up to 10 days after ketamine administration. Biol. Psychiatry 84, 582–590. doi: 10.1016/j.biopsych.2018.01.027
Feyissa, A. M., Chandran, A., Stockmeier, C. A., and Karolewicz, B. (2009). Reduced levels of NR2A and NR2B subunits of NMDA receptor and PSD-95 in the prefrontal cortex in major depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 33, 70–75. doi: 10.1016/j.pnpbp.2008.10.005
Finkbeiner, S., Tavazoie, S. F., Maloratsky, A., Jacobs, K. M., Harris, K. M., and Greenberg, M. E. (1997). CREB: a major mediator of neuronal neurotrophin responses. Neuron 19, 1031–1047. doi: 10.1016/S0896-6273(00)80395-5
Fouquet, G., Coman, T., Hermine, O., and Côté, F. (2019). Serotonin, hematopoiesis and stem cells. Pharmacol. Res. 140, 67–74. doi: 10.1016/j.phrs.2018.08.005
Fox, M. E., and Lobo, M. K. (2019). The molecular and cellular mechanisms of depression: a focus on reward circuitry. Mol. Psychiatry 24, 1798–1815. doi: 10.1038/s41380-019-0415-3
Freeman, M. R. (2010). Specification and morphogenesis of astrocytes. Science 330, 774–778. doi: 10.1126/science.1190928
Gao, J., Lai, M., Fu, W., Wang, M., Thi, T. T. M., Ning, B., et al. (2021). Electroacupuncture ameliorates depressive-like state and synaptic deficits induced by hyper-cholinergic tone During chronic stress in rats. Med. Sci. Monit. 27:e933833. doi: 10.12659/MSM.933833
Gao, J., Lai, M. Y., Mai, T. T., Fu, W., Wang, M. Y., Ning, B. L., et al. (2022). Effects of electroacupuncture on BNDF/mTORC1 signaling pathway and synaptic plasticity in prefrontal cortex of rats exposed to chronic unpredictable mild stress. Zhen ci yan jiu 47, 15–20. doi: 10.13702/j.1000-0607.201293
Gerhard, D. M., Wohleb, E. S., and Duman, R. S. (2016). Emerging treatment mechanisms for depression: focus on glutamate and synaptic plasticity. Drug Discov. Today 21, 454–464. doi: 10.1016/j.drudis.2016.01.016
Gittins, R. A., and Harrison, P. J. (2011). A morphometric study of glia and neurons in the anterior cingulate cortex in mood disorder. J. Affect. Disord. 133, 328–332. doi: 10.1016/j.jad.2011.03.042
Gonçalves, J. T., Schafer, S. T., and Gage, F. H. (2016). Adult neurogenesis in the Hippocampus: from stem cells to behavior. Cell 167, 897–914. doi: 10.1016/j.cell.2016.10.021
Grace, A. A. (2016). Dysregulation of the dopamine system in the pathophysiology of schizophrenia and depression. Nat. Rev. Neurosci. 17, 524–532. doi: 10.1038/nrn.2016.57
Gray, A. L., Hyde, T. M., Deep-Soboslay, A., Kleinman, J. E., and Sodhi, M. S. (2015). Sex differences in glutamate receptor gene expression in major depression and suicide. Mol. Psychiatry 20, 1057–1068. doi: 10.1038/mp.2015.91
Gujral, S., Aizenstein, H., Reynolds, C. F. 3rd, Butters, M. A., and Erickson, K. I. (2017). Exercise effects on depression: Possible neural mechanisms. Gen. Hosp. Psychiatry 49, 2–10. doi: 10.1016/j.genhosppsych.2017.04.012
Guo, Q., Lin, X.-M., Di, Z., Zhang, Q.-A., and Jiang, S. (2021). Electroacupuncture ameliorates CUMS-induced depression-like behavior: involvement of the glutamatergic system and apoptosis in rats. Comb. Chem. High Throughput Screen. 24, 996–1004. doi: 10.2174/1386207323666201027121423
Han, X., Wu, H., Yin, P., Chen, Z., Cao, X., Duan, Y., et al. (2018). Electroacupuncture restores hippocampal synaptic plasticity via modulation of 5-HT receptors in a rat model of depression. Brain Res. Bull. 139, 256–262. doi: 10.1016/j.brainresbull.2018.03.004
Hirono, M., Watanabe, S., Karube, F., Fujiyama, F., Kawahara, S., Nagao, S., et al. (2018). Perineuronal nets in the Deep cerebellar nuclei regulate GABAergic transmission and delay Eyeblink conditioning. J. Neurosci. 38, 6130–6144. doi: 10.1523/JNEUROSCI.3238-17.2018
Hoyer, D., Hannon, J. P., and Martin, G. R. (2002). Molecular, pharmacological and functional diversity of 5-HT receptors. Pharmacol. Biochem. Behav. 71, 533–554. doi: 10.1016/S0091-3057(01)00746-8
Hu, Y., Wu, D. L., Luo, C. X., Zhu, L. J., Zhang, J., Wu, H. Y., et al. (2012). Hippocampal nitric oxide contributes to sex difference in affective behaviors. Proc. Natl. Acad. Sci. USA 109, 14224–14229. doi: 10.1073/pnas.1207461109
Hunter, R. G., Seligsohn, M., Rubin, T. G., Griffiths, B. B., Ozdemir, Y., Pfaff, D. W., et al. (2016). Stress and corticosteroids regulate rat hippocampal mitochondrial DNA gene expression via the glucocorticoid receptor. Proc. Natl. Acad. Sci. USA 113, 9099–9104. doi: 10.1073/pnas.1602185113
Ji, Q., Li, Z. G., Tang, Y. S., Mo, Y. P., Yao, H. J., and Saiyin, C. K. (2013). Effect of electroacupuncture intervention on behavioral changes and hippocampal excitatory amino acid transporter mRNA expression in depression rats. Zhen ci yan jiu 38:202-207, 219
Jiang, H., Zhang, X., Wang, Y., Zhang, H., Li, J., Yang, X., et al. (2017). Mechanisms underlying the antidepressant response of acupuncture via PKA/CREB signaling pathway. Neural Plasticity. 2017:4135164. doi: 10.1155/2017/4135164
Jiang, L., Zhang, H., Zhou, J., Tang, X., Luo, L., Huang, P., et al. (2020). Involvement of hippocampal AMPA receptors in Electroacupuncture attenuating depressive-like behaviors and regulating synaptic proteins in rats subjected to chronic unpredictable mild stress. World Neurosurg. 139, E455–E462. doi: 10.1016/j.wneu.2020.04.042
Johnston-Wilson, N. L., Sims, C. D., Hofmann, J. P., Anderson, L., Shore, A. D., Torrey, E. F., et al. (2000). Disease-specific alterations in frontal cortex brain proteins in schizophrenia, bipolar disorder, and major depressive disorder. The Stanley Neuropathology Consortium. Mol. Psychiatr. 5, 142–149. doi: 10.1038/sj.mp.4000696
Kang, Z., Ye, H., Chen, T., and Zhang, P. (2021). Effect of Electroacupuncture at Siguan Acupoints on expression of BDNF and TrkB proteins in the Hippocampus of post-stroke depression rats. J. Mol. Neurosci. 71, 2165–2171. doi: 10.1007/s12031-021-01844-4
Kawanokuchi, J., Takagi, K., Tanahashi, N., Yamamoto, T., Nagaoka, N., Ishida, T., et al. (2021). Acupuncture treatment for social defeat stress. Front. Behav. Neurosci. 15:685433. doi: 10.3389/fnbeh.2021.685433
Kemp, A., and Manahan-Vaughan, D. (2004). Hippocampal long-term depression and long-term potentiation encode different aspects of novelty acquisition. Proc. Natl. Acad. Sci. USA 101, 8192–8197. doi: 10.1073/pnas.0402650101
Khundakar, A., Morris, C., Oakley, A., and Thomas, A. J. (2011a). A morphometric examination of neuronal and glial cell pathology in the orbitofrontal cortex in late-life depression. Int. Psychogeriatr. 23, 132–140. doi: 10.1017/S1041610210000700
Khundakar, A. A., Morris, C. M., Oakley, A. E., and Thomas, A. J. (2011b). Cellular pathology within the anterior cingulate cortex of patients with late-life depression: a morphometric study. Psychiatry Res. 194, 184–189. doi: 10.1016/j.pscychresns.2011.04.008
Klempan, T. A., Sequeira, A., Canetti, L., Lalovic, A., Ernst, C., Ffrench-Mullen, J., et al. (2009). Altered expression of genes involved in ATP biosynthesis and GABAergic neurotransmission in the ventral prefrontal cortex of suicides with and without major depression. Mol. Psychiatry 14, 175–189. doi: 10.1038/sj.mp.4002110
Kraus, C., Castrén, E., Kasper, S., and Lanzenberger, R. (2017). Serotonin and neuroplasticity - links between molecular, functional and structural pathophysiology in depression. Neurosci. Biobehav. Rev. 77, 317–326. doi: 10.1016/j.neubiorev.2017.03.007
Krishnan, V., and Nestler, E. J. (2008). The molecular neurobiology of depression. Nature 455, 894–902. doi: 10.1038/nature07455
Krzyscik, M. A., Opaliński, Ł., Szymczyk, J., and Otlewski, J. (2022). Cyclic and dimeric fibroblast growth factor 2 variants with high biomedical potential. Int. J. Biol. Macromol. 218, 243–258. doi: 10.1016/j.ijbiomac.2022.07.105
Kupfer, D. J., Frank, E., and Phillips, M. L. (2012). Major depressive disorder: new clinical, neurobiological, and treatment perspectives. Lancet 379, 1045–1055. doi: 10.1016/S0140-6736(11)60602-8
Laping, N. J., Teter, B., Nichols, N. R., Rozovsky, I., and Finch, C. E. (1994). Glial fibrillary acidic protein: regulation by hormones, cytokines, and growth factors. Brain Pathol. 4, 259–275. doi: 10.1111/j.1750-3639.1994.tb00841.x
Lecouflet, P., Roux, C. M., Potier, B., Leger, M., Brunet, E., Billard, J. M., et al. (2021). Interplay between 5-HT4 receptors and GABAergic system within CA1 hippocampal synaptic plasticity. Cerebral Cortex 31, 694–701. doi: 10.1093/cercor/bhaa253
Lee, R., Kermani, P., Teng, K. K., and Hempstead, B. L. (2001). Regulation of cell survival by secreted proneurotrophins. Science 294, 1945–1948. doi: 10.1126/science.1065057
Lee, M.-J., Ryu, J.-S., Won, S.-K., Namgung, U., Jung, J., Lee, S.-M., et al. (2019). Effects of acupuncture on chronic stress-induced depression-like behavior and its central neural mechanism. Front. Psychol. 10:10. doi: 10.3389/fpsyg.2019.01353
Lener, M. S., Niciu, M. J., Ballard, E. D., Park, M., Park, L. T., Nugent, A. C., et al. (2017). Glutamate and gamma-aminobutyric acid Systems in the Pathophysiology of major depression and antidepressant response to ketamine. Biol. Psychiatry 81, 886–897. doi: 10.1016/j.biopsych.2016.05.005
Lesch, K. P., and Waider, J. (2012). Serotonin in the modulation of neural plasticity and networks: implications for neurodevelopmental disorders. Neuron 76, 175–191. doi: 10.1016/j.neuron.2012.09.013
Li, X., Qi, S. M., Wang, Y., Jiang, H. W., Li, Y. H., Zhao, B. C., et al. (2021b). Antidepressant effect of electroacupuncture on modulating the expression of c-Fos/AP-1 through the JNK signaling pathway. Anatomical Record 304, 2480–2493. doi: 10.1002/ar.24740
Li, X.-Y., Wang, H.-M., Zhao, Y., Li, C., Lu, J., Wu, J.-H., et al. (2021a). Effect of acupuncture on microglia activation in prefrontal cortex of chronic stress-induced depression rats. Zhen ci yan jiu 46, 52–57. doi: 10.13702/j.1000-0607.200886
Li, X., Xu, M.-M., Jiang, H.-L., Lu, J., Zhang, X.-H., Zhao, B.-C., et al. (2018). Effect of acupuncture intervention on depression behavior and expression of ERK 1/2 and BDNF in prefrontal cortex in chronic unpredictable mild stress induced depression rats. Zhen ci yan jiu 43, 705–710.
Li, W., Zhu, Y., Saud, S. M., Guo, Q., Xi, S., Jia, B., et al. (2017). Electroacupuncture relieves depression-like symptoms in rats exposed to chronic unpredictable mild stress by activating ERK signaling pathway. Neurosci. Lett. 642, 43–50. doi: 10.1016/j.neulet.2017.01.060
Liang, J., Lu, J., Cui, S.-F., Wang, J.-R., and Tu, Y. (2012). Effect of acupuncture on expression of brain-derived neurotrophic factor gene and protein in frontal cortex and hippocampus of depression rats. Zhen ci yan jiu 37, 20–24
Liang, J., Yue, Y., Jiang, H., Geng, D., Wang, J., Lu, J., et al. (2018). Genetic variations in the p11/tPA/BDNF pathway are associated with post stroke depression. J. Affect. Disord. 226, 313–325. doi: 10.1016/j.jad.2017.09.055
Lin, S.-S., Zhou, B., Chen, B.-J., Jiang, R.-T., Li, B., Illes, P., et al. (2023). Electroacupuncture prevents astrocyte atrophy to alleviate depression. Cell Death Dis. 14:343. doi: 10.1038/s41419-023-05839-4
Liu, Q., Li, B., Zhu, H.-Y., Wang, Y.-Q., Yu, J., and Wu, G.-C. (2011). Glia atrophy in the hippocampus of chronic unpredictable stress-induced depression model rats is reversed by electroacupuncture treatment. J. Affect. Disord. 128, 309–313. doi: 10.1016/j.jad.2010.07.007
Liu, R., Zhang, K., Wang, W.-L., Tong, Q.-Y., Ma, W., Cai, W., et al. (2023). Study on mechanisms of acupuncture underlying improvement of CUMS-induced depression in rats based on tandem mass spectrometry proteomics technique. Zhen ci yan jiu 48, 533–540. doi: 10.13702/j.1000-0607.20220454
Lu, J., Liang, J., Wang, J.-R., Hu, L., Tu, Y., and Guo, J.-Y. (2013). Acupuncture activates ERK-CREB pathway in rats exposed to chronic unpredictable mild stress. Evid. Based Complement. Alternat. Med. 2013, 1–7. doi: 10.1155/2013/469765
Lu, W., Shi, Y., Jackson, A. C., Bjorgan, K., During, M. J., Sprengel, R., et al. (2009). Subunit composition of synaptic AMPA receptors revealed by a single-cell genetic approach. Neuron 62, 254–268. doi: 10.1016/j.neuron.2009.02.027
Luo, D., Ma, R., Wu, Y., Zhang, X., Liu, Y., Wang, L., et al. (2017). Mechanism underlying acupuncture-ameliorated depressive behaviors by enhancing glial glutamate transporter in chronic unpredictable mild stress (CUMS) rats. Med. Sci. Monit. 23, 3080–3087. doi: 10.12659/MSM.902549
Luo, T., Tian, H., Song, H., Zhao, J., Liya, A., Fang, Y., et al. (2020). Possible involvement of tissue plasminogen activator/brain-derived neurotrophic factor pathway in anti-depressant effects of Electroacupuncture in chronic unpredictable mild stress-induced depression in rats. Front. Psych. 11:63. doi: 10.3389/fpsyt.2020.00063
MacQueen, G., and Frodl, T. (2011). The hippocampus in major depression: evidence for the convergence of the bench and bedside in psychiatric research? Mol. Psychiatry 16, 252–264. doi: 10.1038/mp.2010.80
Magee, J. C., and Grienberger, C. (2020). Synaptic plasticity forms and functions. Annu. Rev. Neurosci. 43, 95–117. doi: 10.1146/annurev-neuro-090919-022842
Mahar, I., Bambico, F. R., Mechawar, N., and Nobrega, J. N. (2014). Stress, serotonin, and hippocampal neurogenesis in relation to depression and antidepressant effects. Neurosci. Biobehav. Rev. 38, 173–192. doi: 10.1016/j.neubiorev.2013.11.009
Malhi, G. S., and Mann, J. J. (2018). Depression. Lancet 392, 2299–2312. doi: 10.1016/S0140-6736(18)31948-2
Manji, H. K., Moore, G. J., Rajkowska, G., and Chen, G. (2000). Neuroplasticity and cellular resilience in mood disorders. Mol. Psychiatry 5, 578–593. doi: 10.1038/sj.mp.4000811
Mattson, M. P., Maudsley, S., and Martin, B. (2004). BDNF and 5-HT: a dynamic duo in age-related neuronal plasticity and neurodegenerative disorders. Trends Neurosci. 27, 589–594. doi: 10.1016/j.tins.2004.08.001
Mattson, M. P., Moehl, K., Ghena, N., Schmaedick, M., and Cheng, A. (2018). Intermittent metabolic switching, neuroplasticity and brain health. Nat. Rev. Neurosci. 19, 63–80. doi: 10.1038/nrn.2017.156
McEwen, B. S., Bowles, N. P., Gray, J. D., Hill, M. N., Hunter, R. G., Karatsoreos, I. N., et al. (2015). Mechanisms of stress in the brain. Nat. Neurosci. 18, 1353–1363. doi: 10.1038/nn.4086
Millard, S. J., Weston-Green, K., and Newell, K. A. (2020). The Wistar-Kyoto rat model of endogenous depression: a tool for exploring treatment resistance with an urgent need to focus on sex differences. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 101:109908. doi: 10.1016/j.pnpbp.2020.109908
Mohammad, H., Marchisella, F., Ortega-Martinez, S., Hollos, P., Eerola, K., Komulainen, E., et al. (2018). JNK1 controls adult hippocampal neurogenesis and imposes cell-autonomous control of anxiety behaviour from the neurogenic niche. Mol. Psychiatry 23:487. doi: 10.1038/mp.2017.21
Monroe, S. M., and Harkness, K. L. (2022). Major depression and its recurrences: life course matters. Annu. Rev. Clin. Psychol. 18, 329–357. doi: 10.1146/annurev-clinpsy-072220-021440
Moriguchi, S., Takamiya, A., Noda, Y., Horita, N., Wada, M., Tsugawa, S., et al. (2019). Glutamatergic neurometabolite levels in major depressive disorder: a systematic review and meta-analysis of proton magnetic resonance spectroscopy studies. Mol. Psychiatry 24, 952–964. doi: 10.1038/s41380-018-0252-9
Murrough, J. W., Abdallah, C. G., and Mathew, S. J. (2017). Targeting glutamate signalling in depression: progress and prospects. Nat. Rev. Drug Discov. 16, 472–486. doi: 10.1038/nrd.2017.16
Nagappan, G., Zaitsev, E., Senatorov, V. V. Jr., Yang, J., Hempstead, B. L., and Lu, B. (2009). Control of extracellular cleavage of ProBDNF by high frequency neuronal activity. Proc. Natl. Acad. Sci. USA 106, 1267–1272. doi: 10.1073/pnas.0807322106
Nayak, D., Roth, T. L., and McGavern, D. B. (2014). Microglia development and function. Annu. Rev. Immunol. 32, 367–402. doi: 10.1146/annurev-immunol-032713-120240
Oberheim, N. A., Takano, T., Han, X., He, W., Lin, J. H., Wang, F., et al. (2009). Uniquely hominid features of adult human astrocytes. J. Neurosci. 29, 3276–3287. doi: 10.1523/JNEUROSCI.4707-08.2009
Ongür, D., Drevets, W. C., and Price, J. L. (1998). Glial reduction in the subgenual prefrontal cortex in mood disorders. Proc. Natl. Acad. Sci. USA 95, 13290–13295. doi: 10.1073/pnas.95.22.13290
Otte, C., Gold, S. M., Penninx, B. W., Pariante, C. M., Etkin, A., Fava, M., et al. (2016). Major depressive disorder. Nat. Rev. Dis. Prim. 2:16065. doi: 10.1038/nrdp.2016.65
Pang, F., Yang, Y., Huang, S., Yang, Z., Zhu, Z., Liao, D., et al. (2023). Electroacupuncture alleviates depressive-like behavior by modulating the expression of P2X7/NLRP3/IL-1β of prefrontal cortex and liver in rats exposed to chronic unpredictable mild stress. Brain Sci. 13:436. doi: 10.3390/brainsci13030436
Park, J.-S., and Lim, H.-H. (2019). Effect of acupuncture on depression and cell proliferation in hippocampal gyrus Dentatus of maternal-separated rat pups. J. Kor. Med. Rehabil. 29, 91–99. doi: 10.18325/jkmr.2019.29.2.91
Pittenger, C., and Duman, R. S. (2008). Stress, depression, and neuroplasticity: a convergence of mechanisms. Neuropsychopharmacology 33, 88–109. doi: 10.1038/sj.npp.1301574
Price, R. B., and Duman, R. (2020). Neuroplasticity in cognitive and psychological mechanisms of depression: an integrative model. Mol. Psychiatry 25, 530–543. doi: 10.1038/s41380-019-0615-x
Qi, X., Lin, W., Li, J., Li, H., Wang, W., Wang, D., et al. (2008). Fluoxetine increases the activity of the ERK-CREB signal system and alleviates the depressive-like behavior in rats exposed to chronic forced swim stress. Neurobiol. Dis. 31, 278–285. doi: 10.1016/j.nbd.2008.05.003
Rajkowska, G., Miguel-Hidalgo, J. J., Wei, J., Dilley, G., Pittman, S. D., Meltzer, H. Y., et al. (1999). Morphometric evidence for neuronal and glial prefrontal cell pathology in major depression. Biol. Psychiatry 45, 1085–1098. doi: 10.1016/S0006-3223(99)00041-4
Rajkowska, G., and Stockmeier, C. A. (2013). Astrocyte pathology in major depressive disorder: insights from human postmortem brain tissue. Curr. Drug Targets 14, 1225–1236. doi: 10.2174/13894501113149990156
Rana, T., Behl, T., Mehta, V., Uddin, M. S., and Bungau, S. (2021). Molecular insights into the therapeutic promise of targeting HMGB1 in depression. Pharmacol. Reports 73, 31–42. doi: 10.1007/s43440-020-00163-6
Rantamäki, T., and Castrén, E. (2008). Targeting TrkB neurotrophin receptor to treat depression. Expert Opin. Ther. Targets 12, 705–715. doi: 10.1517/14728222.12.6.705
Rogatsky, I., Logan, S. K., and Garabedian, M. J. (1998). Antagonism of glucocorticoid receptor transcriptional activation by the c-Jun N-terminal kinase. Proc. Natl. Acad. Sci. USA 95, 2050–2055. doi: 10.1073/pnas.95.5.2050
Rutigliano, G., and Zucchi, R. (2020). Molecular variants in human trace amine-associated receptors and their implications in mental and metabolic disorders. Cell. Mol. Neurobiol. 40, 239–255. doi: 10.1007/s10571-019-00743-y
Ryu, H., Chung, M., Dobrzyński, M., Fey, D., Blum, Y., Lee, S. S., et al. (2015). Frequency modulation of ERK activation dynamics rewires cell fate. Mol. Syst. Biol. 11:838. doi: 10.15252/msb.20156458
Sagi, Y., Tavor, I., Hofstetter, S., Tzur-Moryosef, S., Blumenfeld-Katzir, T., and Assaf, Y. (2012). Learning in the fast lane: new insights into neuroplasticity. Neuron 73, 1195–1203. doi: 10.1016/j.neuron.2012.01.025
Sanacora, G., and Banasr, M. (2013). From pathophysiology to novel antidepressant drugs: glial contributions to the pathology and treatment of mood disorders. Biol. Psychiatry 73, 1172–1179. doi: 10.1016/j.biopsych.2013.03.032
Sanacora, G., Treccani, G., and Popoli, M. (2012). Towards a glutamate hypothesis of depression: an emerging frontier of neuropsychopharmacology for mood disorders. Neuropharmacology 62, 63–77. doi: 10.1016/j.neuropharm.2011.07.036
Sanacora, G., Zarate, C. A., Krystal, J. H., and Manji, H. K. (2008). Targeting the glutamatergic system to develop novel, improved therapeutics for mood disorders. Nat. Rev. Drug Discov. 7, 426–437. doi: 10.1038/nrd2462
Savitz, J., and Drevets, W. C. (2009). Bipolar and major depressive disorder: neuroimaging the developmental-degenerative divide. Neurosci. Biobehav. Rev. 33, 699–771. doi: 10.1016/j.neubiorev.2009.01.004
Schmidt, H. D., Shelton, R. C., and Duman, R. S. (2011). Functional biomarkers of depression: diagnosis, treatment, and pathophysiology. Neuropsychopharmacology 36, 2375–2394. doi: 10.1038/npp.2011.151
Sequeira, A., Mamdani, F., Ernst, C., Vawter, M. P., Bunney, W. E., Lebel, V., et al. (2009). Global brain gene expression analysis links glutamatergic and GABAergic alterations to suicide and major depression. PLoS One 4:e6585. doi: 10.1371/journal.pone.0006585
Shaywitz, A. J., and Greenberg, M. E. (1999). CREB: a stimulus-induced transcription factor activated by a diverse array of extracellular signals. Annu. Rev. Biochem. 68, 821–861. doi: 10.1146/annurev.biochem.68.1.821
She, Y., Xu, J., Duan, Y., Su, N., Sun, Y., Cao, X., et al. (2015). Possible antidepressant effects and mechanism of electroacupuncture in behaviors and hippocampal synaptic plasticity in a depression rat model. Brain Res. 1629, 291–297. doi: 10.1016/j.brainres.2015.10.033
Si, X., Miguel-Hidalgo, J. J., O’Dwyer, G., Stockmeier, C. A., and Rajkowska, G. (2004). Age-dependent reductions in the level of glial fibrillary acidic protein in the prefrontal cortex in major depression. Neuropsychopharmacology 29, 2088–2096. doi: 10.1038/sj.npp.1300525
Spalding, K. L., Bergmann, O., Alkass, K., Bernard, S., Salehpour, M., Huttner, H. B., et al. (2013). Dynamics of hippocampal neurogenesis in adult humans. Cell 153, 1219–1227. doi: 10.1016/j.cell.2013.05.002
Stogsdill, J. A., Ramirez, J., Liu, D., Kim, Y. H., Baldwin, K. T., Enustun, E., et al. (2017). Astrocytic neuroligins control astrocyte morphogenesis and synaptogenesis. Nature 551, 192–197. doi: 10.1038/nature24638
Sun, P.-Y., Cai, R.-L., Li, P.-F., Zhu, Y., Wang, T., Wu, J., et al. (2019). Protective effects on hippocampal neurons and the influence on hippocampal monoamine neurotransmitters with acupuncture for promoting the circulation of the governor vessel and regulating the mental state in rats with post-stroke depression. Zhongguo zhen jiu 39, 741–747. doi: 10.13703/j.0255-2930.2019.07.017
Sun, P.-Y., Chu, H.-R., Li, N., Liu, H., Liu, S.-Y., Zhang, F., et al. (2022). Effect of Tongdu Tiaoshen acupuncture on CREB/BDNF/TrkB signaling pathway of hippocampus in rats with post-stroke depression. Zhongguo zhen jiu 42, 907–913. doi: 10.13703/j.0255-2930.20220206-k0003
Sun, P.-Y., Li, P.-F., Wang, T., Wu, J., Li, N., Liu, H., et al. (2020). Effect of Tongdu Tiaoshen acupuncture on PI3K/Akt/mTOR signaling pathway and autophagy-related proteins of hippocampus in rats with post-stroke depression. Zhongguo zhen jiu 40, 1205–1210. doi: 10.13703/j.0255-2930.20200522-k0006
Sun, J. D., Liu, Y., Yuan, Y. H., Li, J., and Chen, N. H. (2012). Gap junction dysfunction in the prefrontal cortex induces depressive-like behaviors in rats. Neuropsychopharmacology 37, 1305–1320. doi: 10.1038/npp.2011.319
Sun, Y., Tu, Y., Guo, Y., Jiang, H.-L., Li, Y.-H., Wang, Y., et al. (2019c). Acupuncture improved depressive behavior by regulating expression of hippocampal apoptosis-related factors in psychological stress-induced depression rats. Zhen ci yan jiu 44, 412–418. doi: 10.13702/j.1000-0607.190098
Suzuki, T., and Yamamoto, M. (2015). Molecular basis of the Keap1-Nrf2 system. Free Radic. Biol. Med. 88, 93–100. doi: 10.1016/j.freeradbiomed.2015.06.006
Sweatt, J. D. (2004). Mitogen-activated protein kinases in synaptic plasticity and memory. Curr. Opin. Neurobiol. 14, 311–317. doi: 10.1016/j.conb.2004.04.001
Tartt, A. N., Mariani, M. B., Hen, R., Mann, J. J., and Boldrini, M. (2022). Dysregulation of adult hippocampal neuroplasticity in major depression: pathogenesis and therapeutic implications. Mol. Psychiatry 27, 2689–2699. doi: 10.1038/s41380-022-01520-y
Tian, M., Stroebel, D., Piot, L., David, M., Ye, S., and Paoletti, P. (2021). GluN2A and GluN2B NMDA receptors use distinct allosteric routes. Nat. Commun. 12:4709. doi: 10.1038/s41467-021-25058-9
Tomita, S., Adesnik, H., Sekiguchi, M., Zhang, W., Wada, K., Howe, J. R., et al. (2005). Stargazin modulates AMPA receptor gating and trafficking by distinct domains. Nature 435, 1052–1058. doi: 10.1038/nature03624
Tong, T., Chen, Y., Hao, C., Shen, J., Chen, W., Cheng, W., et al. (2023). The effects of acupuncture on depression by regulating BDNF-related balance via lateral habenular nucleus BDNF/TrkB/CREB signaling pathway in rats. Behav. Brain Res. 451:114509. doi: 10.1016/j.bbr.2023.114509
Vivar, C., Potter, M. C., Choi, J., Lee, J. Y., Stringer, T. P., Callaway, E. M., et al. (2012). Monosynaptic inputs to new neurons in the dentate gyrus. Nat. Commun. 3:1107. doi: 10.1038/ncomms2101
Wang, B., Huang, X., Pan, X., Zhang, T., Hou, C., Su, W. J., et al. (2020). Minocycline prevents the depressive-like behavior through inhibiting the release of HMGB1 from microglia and neurons. Brain Behav. Immun. 88, 132–143. doi: 10.1016/j.bbi.2020.06.019
Wang, C. S., Kavalali, E. T., and Monteggia, L. M. (2022). BDNF signaling in context: from synaptic regulation to psychiatric disorders. Cell 185, 62–76. doi: 10.1016/j.cell.2021.12.003
Wang, J. Q., and Mao, L. (2019). The ERK pathway: molecular mechanisms and treatment of depression. Mol. Neurobiol. 56, 6197–6205. doi: 10.1007/s12035-019-1524-3
Weber, M. D., Frank, M. G., Tracey, K. J., Watkins, L. R., and Maier, S. F. (2015). Stress induces the danger-associated molecular pattern HMGB-1 in the hippocampus of male Sprague Dawley rats: a priming stimulus of microglia and the NLRP3 inflammasome. J. Neurosci. 35, 316–324. doi: 10.1523/JNEUROSCI.3561-14.2015
Webster, M. J., O’Grady, J., Kleinman, J. E., and Weickert, C. S. (2005). Glial fibrillary acidic protein mRNA levels in the cingulate cortex of individuals with depression, bipolar disorder and schizophrenia. Neuroscience 133, 453–461. doi: 10.1016/j.neuroscience.2005.02.037
Wert-Carvajal, C., Reneaux, M., Tchumatchenko, T., and Clopath, C. (2022). Dopamine and serotonin interplay for valence-based spatial learning. Cell Rep. 39:110645. doi: 10.1016/j.celrep.2022.110645
Wichers, M. C., Koek, G. H., Robaeys, G., Verkerk, R., Scharpé, S., and Maes, M. (2005). IDO and interferon-alpha-induced depressive symptoms: a shift in hypothesis from tryptophan depletion to neurotoxicity. Mol. Psychiatry 10, 538–544. doi: 10.1038/sj.mp.4001600
Won, E., and Kim, Y. K. (2016). Stress, the autonomic nervous system, and the immune-kynurenine pathway in the etiology of depression. Curr. Neuropharmacol. 14, 665–673. doi: 10.2174/1570159X14666151208113006
Xie, Y., Su, N., Yang, J., Tan, Q., Huang, S., Jin, M., et al. (2020). FGF/FGFR signaling in health and disease. Signal Transduct. Target. Ther. 5:181. doi: 10.1038/s41392-020-00222-7
Yang, N. N., Lin, L. L., Li, Y. J., Li, H. P., Cao, Y., Tan, C. X., et al. (2022). Potential mechanisms and clinical effectiveness of acupuncture in depression. Curr. Neuropharmacol. 20, 738–750. doi: 10.2174/1570159X19666210609162809
Yang, L., Yue, N., Zhu, X., Han, Q., Li, B., Liu, Q., et al. (2014). Electroacupuncture promotes proliferation of amplifying neural progenitors and preserves quiescent neural progenitors from apoptosis to alleviate depressive‐like and anxiety‐like Behaviours. Evid. Based Complement. Alternat. Med. 2014:2014. doi: 10.1155/2014/872568
Yang, L., Yue, N., Zhu, X., Han, Q., Liu, Q., Yu, J., et al. (2013). Electroacupuncture upregulates ERK signaling pathways and promotes adult hippocampal neural progenitors proliferation in a rat model of depression. BMC Complement. Altern. Med. 13, 1–9. doi: 10.1186/1472-6882-13-288
Yao, Z., Zhang, Z., Zhang, J., Cai, X., Zhong, Z., Huang, Y., et al. (2021). Electroacupuncture alleviated the depression-like behavior by regulating FGF2 and astrocytes in the hippocampus of rats with chronic unpredictable mild stress. Brain Res. Bull. 169, 43–50. doi: 10.1016/j.brainresbull.2021.01.005
Yirmiya, R., Rimmerman, N., and Reshef, R. (2015). Depression as a microglial disease. Trends Neurosci. 38, 637–658. doi: 10.1016/j.tins.2015.08.001
Zandarashvili, L., Sahu, D., Lee, K., Lee, Y. S., Singh, P., Rajarathnam, K., et al. (2013). Real-time kinetics of high-mobility group box 1 (HMGB1) oxidation in extracellular fluids studied by in situ protein NMR spectroscopy. J. Biol. Chem. 288, 11621–11627. doi: 10.1074/jbc.M113.449942
Zarneshan, S. N., Fakhri, S., and Khan, H. (2022). Targeting Akt/CREB/BDNF signaling pathway by ginsenosides in neurodegenerative diseases: a mechanistic approach. Pharmacol. Res. 177:106099. doi: 10.1016/j.phrs.2022.106099
Zeke, A., Misheva, M., Reményi, A., and Bogoyevitch, M. A. (2016). JNK signaling: regulation and functions based on complex protein-protein partnerships. Microbiol. Mol. Biol. Rev. 80, 793–835. doi: 10.1128/MMBR.00043-14
Zhang, Z., Cai, X., Yao, Z., Wen, F., Fu, Z., Zhang, J., et al. (2020). EA ameliorated depressive behaviors in CUMS rats and was related to its suppressing autophagy in the Hippocampus. Neural Plast. 2020, 1–9. doi: 10.1155/2020/8860968
Zhang, Y., Guo, Z., Yang, L., Cheng, C., Gai, C., Gao, Y., et al. (2023). Possible involvement of Perineuronal nets in anti-depressant effects of Electroacupuncture in chronic-stress-induced depression in rats. Neurochem. Res. 48, 3146–3159. doi: 10.1007/s11064-023-03970-4
Zhang, K., Liu, R., Zhang, J., Wei, X., Gao, Y., Ma, W., et al. (2021b). Electroacupuncture ameliorates depression-like behaviour in rats by enhancing synaptic plasticity via the GluN2B/CaMKII/CREB Signalling pathway. Evid. Based Complement. Alternat. Med. 2146001. doi: 10.1155/2021/2146001
Zhang, X., Odom, D. T., Koo, S. H., Conkright, M. D., Canettieri, G., Best, J., et al. (2005). Genome-wide analysis of cAMP-response element binding protein occupancy, phosphorylation, and target gene activation in human tissues. Proc. Natl. Acad. Sci. USA 102, 4459–4464. doi: 10.1073/pnas.0501076102
Zhang, J., Zhang, J., Zhang, Z., Zheng, Y., Zhong, Z., Yao, Z., et al. (2021a). Dopaminergic signaling in prefrontal cortex contributes to the antidepressant effect of electroacupuncture: an iTRAQ-based proteomics analysis in a rat model of CUMS. Anat. Rec. 304, 2454–2469. doi: 10.1002/ar.24732
5-HT - 5-hydroxytryptamine, Serotonin
5-HTT - The serotonin transporter
AIF - Apoptosis-inducing factor
Akt - Protein kinase B
AMPAR - The expression of AMPA glutamate receptor
AMPARs - α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid receptors
ANPs - The amplifying neural progenitor cells
BDNF - Brain-derived neurotrophic factor
cAMP - Cyclic adenosine monophosphate
caspase-3 - Cysteine-containing aspartate-specific proteases-3
CNS - The central nervous system
CNS - Central nervous system
CREB - Cyclic AMP response-binding protein
CUMS - The chronic unpredictable mild stress
DA - Dopamine
DAT - The dopamine transporter
DDC - Aromatic-L-amino-acid decarboxylase
DG - The dentate gyrus
DR - The dorsal raphe
EA - Electroacupuncture
ERK - The extracellular signal-regulated kinase
FGF2 - Fibroblast growth factor 2
GABA - Gamma-aminobutyric acid
GAD67 - GABA synthetase glutamic acid decarboxylase 67
GAP-43 - Growth-associated protein-43
GFAP - Glial fibrillary acidic protein
Glu - Glutamate
GluR1 - Glutamate receptor 1
GluR2 - Glutamate receptor 2
HMGB1 - High mobility group box-1
HO-1 - Heme oxygenase-1
HPA - Hypothalamic–pituitary–adrenal axis
Iba-1 - The microglial marker Ionized calcium-binding adaptor molecule 1
IL-1beta - Interleukin-1beta
IL-6 - Interleukin-6
JNKs - The c-Jun N-terminal kinases
LC3 - The autophagic biomarker light chain 3
LHb - The lateral habenular
LTD - Long-term depression
LTP - Long-term potentiation
MAP-2 - Microtubule-associated protein 2
MAPK–ERK - The mitogen-activated protein kinase
mPFC - The medial prefrontal cortex
MDD - Major depressive disorder
MRS - Magnetic resonance spectroscopy
mTOR - The mammalian target of rapamycin
NE - Norepinephrine
NMDARs - N-methyl-d-aspartate receptors
Nrf2 - Nuclear factor E2-related factor 2
NSCs - Neural stem cells
NT - Neurotrophin
PFC - The prefrontal cortex
PI3K - The phosphatidylinositol 3-kinase
PI3K-Akt - phosphoinositide 3-kinase
Pick1 - Protein interacting with C kinase 1
PKA - The protein kinase A
PKA - Protein kinase A
PLC-γ - Phospholipase-C gamma
PNN - Perineuronal nets
PSD - Poststroke depression
PSD-95 - Postsynaptic density protein-95
QNPs - The quiescent neural progenitor cells
RAS-MAPK - The RAS-mitogen-activated protein kinase pathway
ROS - Reactive oxygen species
SYN - Synaptophysin
SYP - Synaptophysin
TAAR1 - The trace amine-associated receptor 1
TNF-α - Tumor necrosis factor-α
tPA - Tissue plasminogen activator
TrkB - The tyrosine kinase B receptor
WKY - Wistar Kyoto
Keywords: major depressive disorder, acupuncture, mechanism, neuroplasticity, complementary and alternative medicine
Citation: Xu N, He Y, Wei Y-N, Bai L and Wang L (2025) Possible antidepressant mechanism of acupuncture: targeting neuroplasticity. Front. Neurosci. 19:1512073. doi: 10.3389/fnins.2025.1512073
Edited by:
Tie-Qiang Li, Karolinska University Hospital, SwedenReviewed by:
Jing Du, Capital Medical University, ChinaCopyright © 2025 Xu, He, Wei, Bai and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Long Wang, d2xrZXlhbkAxNjMuY29t; d2FuZ2xvbmc3OEBobGp1Y20uZWR1LmNu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.