 Marco Greco
Marco Greco Anas Munir
Anas Munir Debora Musarò
Debora Musarò Chiara Coppola
Chiara Coppola Michele Maffia
Michele Maffia- 1Department of Biological and Environmental Science and Technology, University of Salento, Lecce, Italy
- 2Department of Mathematics and Physics “E. De Giorgi”, University of Salento, Lecce, Italy
Parkinson’s disease (PD) is a predominantly idiopathic pathological condition characterized by protein aggregation phenomena, whose main component is alpha-synuclein. Although the main risk factor is ageing, numerous evidence points to the role of type 2 diabetes mellitus (T2DM) as an etiological factor. Systemic alterations classically associated with T2DM like insulin resistance and hyperglycemia modify biological processes such as autophagy and mitochondrial homeostasis. High glucose levels also compromise protein stability through the formation of advanced glycation end products, promoting protein aggregation processes. The ability of antidiabetic drugs to act on pathways impaired in both T2DM and PD suggests that they may represent a useful tool to counteract the neurodegeneration process. Several clinical studies now in advanced stages are looking for confirmation in this regard.
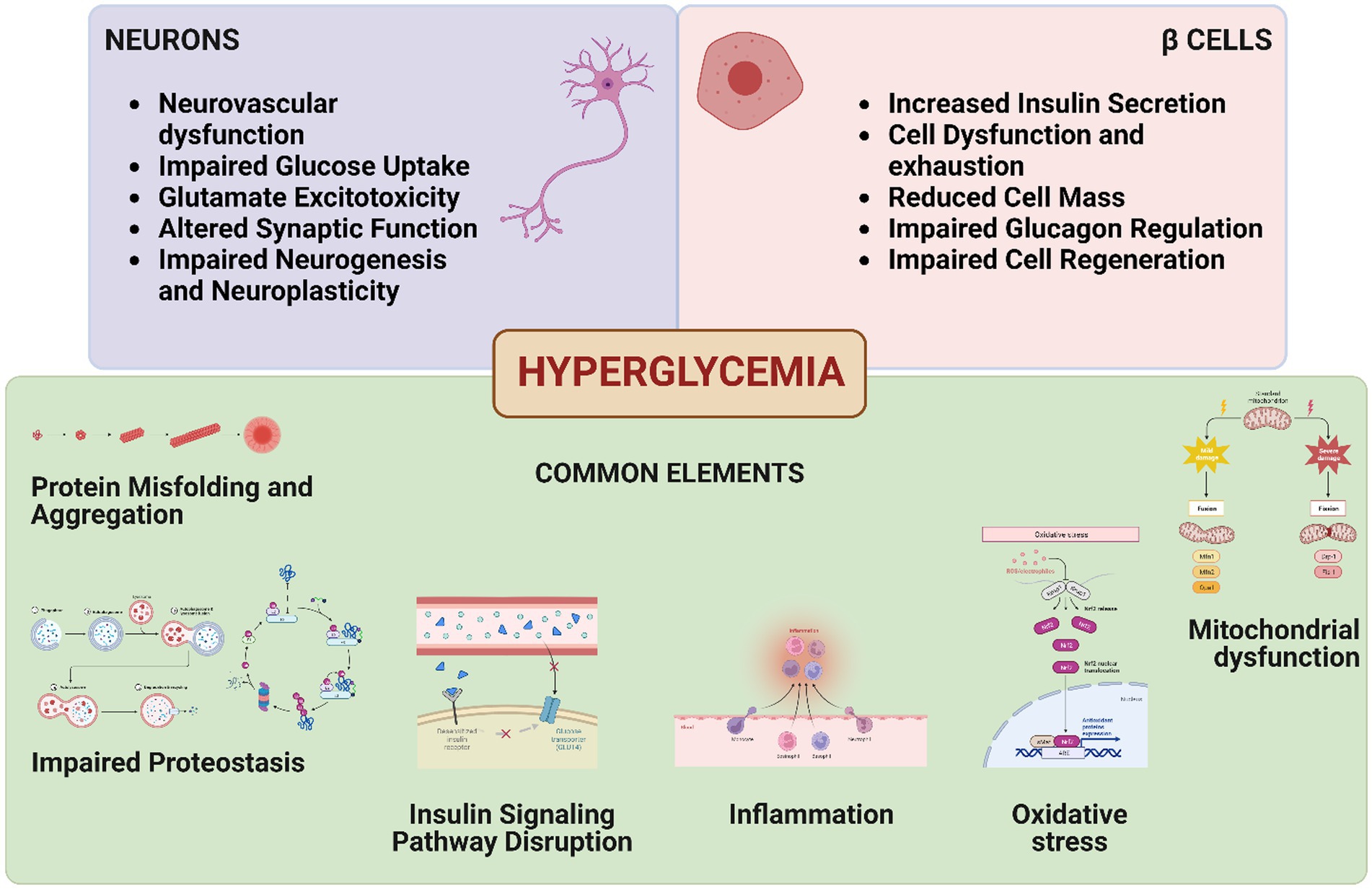
Graphical Abstract. This figure represents the shared elements between neurodegeneration and β-cell dysfunction, linked through the overarching condition of hyperglycemia.
1. Introduction
Parkinson’s disease (PD) is an irreversible and progressive neurological condition that affects 8.5 million people worldwide (World Health Organization, 2022), with an incidence expected to double by 2040 (Dorsey and Bloem, 2018). PD classically involves midbrain structures, causing the depletion of the residing dopaminergic neuronal population and a predominantly motor symptomatology. The disease belongs to the class of proteinopathies and, apart from 5% of genetically linked cases, it is considered idiopathic (Reed et al., 2019).
Proteinopathies are chronic conditions characterized by an imbalance between protein synthesis and degradation (Bayer, 2015). This imbalance, which is caused by ageing, mitochondrial dysfunction, oxidative stress, inflammation and alterations in post-translational processes (PTMs), is responsible for protein aggregation and Lewy bodies formation at the neuronal level in PD. Type 2 diabetes mellitus (T2DM) is another proteinopathy with an alarming prevalence rate of 536 million worldwide, characterized by peripheral insulin resistance, high blood glucose levels and increased insulin secretion, which in time lead to a diminishment of β-cell functionality and number (Sun et al., 2022). Furthermore, these disease symptoms when sustained cause mitochondrial dysfunction, inflammation, and protein misfolding.
An association between type 2 diabetes mellitus (T2DM) and PD has long been advocated (Sandyk, 1993), and subsequently strengthened by a better understanding of the underlying pathologically involved pathways. Numerous population-based studies have failed to unequivocally confirm the hypothesis of a causal relationship, with often contradictory conclusions requiring further investigation (Driver et al., 2008; Cereda et al., 2011, 2012; Schernhammer et al., 2011; Savica et al., 2012; Lu et al., 2014; Pagano et al., 2018). However, because of the brain requirement of insulin and the similar signaling mechanisms in the two body districts, impaired insulin release or uptake can contribute to both T2DM and PD pathogenesis. Both the conditions share pathophysiological elements like chronic inflammation, lysosomal and mitochondrial disfunction, whose molecular bases are associated with a loss in glucose metabolism (Burbulla et al., 2017; Ma et al., 2017). Moreover, insulin signaling has been shown to be involved in several mechanisms regulating apoptosis and oxidative stress prevention, with a profound role as neuroprotective agent (Duarte et al., 2008; Serhan et al., 2020; Gayen et al., 2022).
Furthermore, the dysfunction of glucose metabolism, typical of T2DM has been demonstrated as an early marker of PD (Dunn et al., 2014). Another important process that is impacted in both diseases is the cellular recycling and degradation mechanism of autophagy, which is interestingly both disturbed and contributes to the overwhelming aggregation of proteinaceous Lewy bodies and amyloid plaques in PD and T2DM, respectively (Hou et al., 2020). Moreover, mutations in autophagy-related genes (ARGs) also inherently impair the autophagic flux in both the diseases (Simón-Sánchez et al., 2009; Cui and Li, 2023). In view of this, a limited number of studies have demonstrated that the activation of pathways like the mammalian target of rapamycin (mTOR) can restore autophagy, alleviating symptoms in in vitro and in vivo models of the two diseases (Pupyshev et al., 2021; Cui and Li, 2023). Targeting autophagy, therefore, presents a lucrative drug target common to both disorders.
2. Protein aggregation as a two-way connection
It has long been known, thanks to the pioneering observations of Polymeropoulos and co-workers first, Spillantini and colleagues later, that Lewy bodies in PD, initially localized in the substantia nigra pars compacta, are predominantly composed of alpha-synuclein (aS) protein (Polymeropoulos et al., 1997; Spillantini et al., 1997). It was subsequently discovered by Fujiwara et al. (2002) that aggregates were mainly composed of the phospho-Ser129 form of the protein (Fujiwara et al., 2002). The loss of function of the protein because of its aggregation has several consequences in dopaminergic neurons, it being involved in the modulation of neurotransmitter synthesis and, once incorporated into vesicles, also in their transport and release (Abeliovich et al., 2000; Cabin et al., 2002). Interactors of aS include tyrosine 3-monooxygenase (TH), the rate-limiting dopamine biosynthetic enzyme (Perez et al., 2002), dopamine decarboxylase (Tehranian et al., 2006), protein phosphatase 2A (PP2A) (Qu et al., 2018), vesicle membranes (Jao et al., 2004) and SNARE proteins (Burré et al., 2014). An important but still poorly understood interactor of aS is the Kir6.2 (internal rectifier potassium channel 6.2) subunit of the ATP-sensitive potassium (KATP) channel and its associated sulfonylurea receptor 1 (SUR1). This interaction modulates dopamine release by reducing it (Vidal-Martinez et al., 2018; Choudhury et al., 2022).
A similar function for the protein was observed by Geng and colleagues in pancreatic β-cell, where it has been shown to act as a regulator of the release of insulin-containing vesicles, in response to glucose concentration (Geng et al., 2007). Aggregation is observed in these cells during T2DM; the major component of which is islet amyloid polypeptide (IAPP) (Cooper et al., 1987), although immunoreactivity for phospho-Ser129-aS has also been observed (Martinez-Valbuena et al., 2018).
The fibrillation process of aS passes through the formation of oligomeric structures which are neurotoxic formations able to pierce membranes and spread. At this point, the protein can be internalised by other cells or flow through the cerebrospinal fluid (CSF) to the bloodstream (Menéndez-González et al., 2018; Karpowicz et al., 2019). A role for aS active internalization process seems to be played by surface prion protein (PrP), expressed both at the nervous and pancreatic levels, where it modulates insulin and glucose homeostasis through metal interaction (Ashok and Singh, 2018; Jucker and Walker, 2018; De Riccardis et al., 2019). Recently, Martinez-Valbuena et al. (2021) have found the presence of cytoplasmic aggregates containing IAPP and phospho-Ser129-aS interacting with PrP in human post-mortem pancreatic tissues (Martinez-Valbuena et al., 2021). This offers a new perspective towards a two-way connection between the pathogenesis of T2DM and PD.
3. Insulin resistance and the impairment of autophagic processes
A persistent condition associated with T2DM is insulin resistance, which determines a loss of glycemic control mechanisms in peripheral organs. The brain, however, is capable of managing its own glucose requirements independently of insulin, although new evidence is casting doubt on this assumption (García-Cáceres et al., 2016; Kleinridders, 2016; Pomytkin et al., 2018). Therefore, in the brain, insulin resistance results mainly in an altered signaling pathway.
Encephalic insulin receptors are predominantly expressed in neurons, where they play a role in modulating dendritic and synaptic plasticity, being most highly expressed at these levels (Sportelli et al., 2020). The hormone is associated with a plethora of activities in the brain, such as learning, memory, cognitive functions (Craft et al., 1993), neurotransmitter release (de Bartolomeis et al., 2023), neuroprotection, neuronal proliferation, migration, and differentiation (Roger and Fellows, 1980; Schubert et al., 2003; Xu et al., 2004; Sousa-Nunes et al., 2011). These activities are all modulated by the binding of insulin to its receptors and the activation of their downstream pathways, for instance, Raf-1/MAPK/ERK or PI3K/protein kinase B (AKT) (Arnold et al., 2018). These effectors then modulate mTOR activity, promote protein synthesis activating ribosomal protein S6 kinase (S6K) (Huang et al., 2019), inhibiting the eukaryotic translation initiation factor 4E-binding protein 1 (4E-BP1) (Le Bacquer et al., 2007), while activating transcription factors like forkhead box protein O1 (FoxO1) (Tsai et al., 2003), sterol regulatory element-binding protein (SREBP) and carbohydrate-responsive element-binding protein (ChREBP) (Iizuka et al., 2004; Suzuki et al., 2010).
The mTOR protein, a central element of the mTORC1 and mTORC2 complexes, modulated by the PI3K/AKT pathway, regulates the autophagic process. Alterations in this mechanism are associated with protein aggregation and reduction in ATP/ADP and NAD+/NADH ratios (Heras-Sandoval et al., 2014; Steinberg and Carling, 2019; Katsyuba et al., 2020). A 2010 study found high amounts of autophagosomes with permeabilized lysosomes, indicating a shunted lysosomal-mediated autophagosome clearance contributing to disease onset in post-mortem PD brain (Dehay et al., 2010). This, in addition to the high levels of autophagic markers that colocalize with aS in Lewy bodies, such as microtubule-associated proteins 1A/1B light chain 3B (LC3) in its lipidated form LC3-II and ubiquitin-binding protein p62 (sequestosome-1), provides important evidence for a failure of the autophagic process during the disease (Alvarez-Erviti et al., 2010; Fellner et al., 2021).
4. Hyperglycemia compromises cellular proteostasis
An impaired protein degradation, although sufficient to alter cellular proteostasis, is not the only event observed during T2DM. Increased level of circulating glucose also has detrimental effects on cellular homeostasis and protein function. An engulfment of the glycolytic pathway diverts glucose towards the polyol pathway (Du et al., 2003). Then, the sustained activity of aldose reductase and sorbitol dehydrogenase causes the depletion of NADPH and NAD+, which impairs the capability of the cell to restore reduced glutathione (GSH) levels and causes inhibition of GAPDH (Mathebula, 2015; Lutchmansingh et al., 2018). The accumulation of triose phosphates results in methylglyoxal (MG) formation, an α-keto reactive aldehyde, and then to advanced glycation end products (AGEs) (Strom et al., 2021). Hyperglycemia also impairs the hexosamine pathway, increasing UDP-N-acetylglucosamine levels and altering PTMs (Du et al., 2000; Del Coco et al., 2023). As the MG detoxification is mainly mediated by the highly conserved glutathione-dependent glyoxalase I/II (GloI/II) system, a reduction in intracellular levels of GSH is toxic for the cells (Blair et al., 1979; Yadav et al., 2005). Altered glycosylation and glycation processes involve both aS and IAPP, promoting their aggregation and impairing their degradation (Vicente Miranda et al., 2017; Milordini et al., 2020).
5. Mitochondrial dysfunction as a fatal consequence
Insulin resistance and hyperglycemia cause mitochondria dysfunction, with ROS increase and loss in Ca2+ homeostasis further contributing to AGEs formation (Soejima et al., 1996). Mitochondria are responsible for cellular energy metabolism, regulating also proliferation (Diers et al., 2012), apoptosis (Wang and Youle, 2009), protein degradation (Liao et al., 2020), neurotransmitter transport, uptake and recycle (Varoqui and Erickson, 1996; Gasnier, 2000; Vos, 2010). Their dysfunction is strongly related to inflammation, which in the brain is supported by microglia through the release of pro-inflammatory cytokines.
Hyperglycemia causes alterations in the organelle morphology, triggering fission processes (Wang et al., 2012). Mitophagy, a selective form of autophagy, is responsible for the removal of damaged mitochondria; during this process, organelles are labelled for degradation by PTEN-induced putative kinase 1 (PINK1), E3 ubiquitin-protein ligase Parkin, Ubiquitin and Sequestosome-1 (Lewis and Lewis, 1915; Palikaras et al., 2018). An important role in mitochondria homeostasis is held by DJ-1 protein; it acts as a transcription factor and an antioxidant and modulates chaperones, proteases, and mitochondria activity. DJ-1 functions are modulated by its oxidative state in the cell (Canet-Avilés et al., 2004; Hao et al., 2010; Wilson, 2011; Takahashi-Niki et al., 2017).
Finally, DJ-1 has a key role in repairing MG-related damages, with a recent study demonstrating glyoxalase and weak deglyoxalase activity for the protein (Mazza et al., 2022). Mutations in PINK1, Parkin and DJ-1 are correlated with the onset of monogenic forms of PD (Guadagnolo et al., 2021). This emphasizes how mitophagy and organelle homeostasis are crucial for cell well-being, and how glycemic dyshomeostasis can induce PD.
6. Autophagy and hyperglycemia as new drug targets
The pharmacological approach to PD is merely symptomatic, therefore the identification of a trigger capable of inducing the pathology, and its medication, stand to represent a new complementary route of intervention. Hyperglycemia, hyperinsulinemia, hypercholesterolemia, inflammation, mitochondrial dyshomeostasis, oxidative stress, alteration of protein-degrading pathways, and aggregation are all potential targets to be exploited.
Several widely used antidiabetic and hypoglycemic drugs are currently undergoing pharmaceutical trials in various studies on cohorts of PD patients with a perspective of drug repurposing (Table 1).
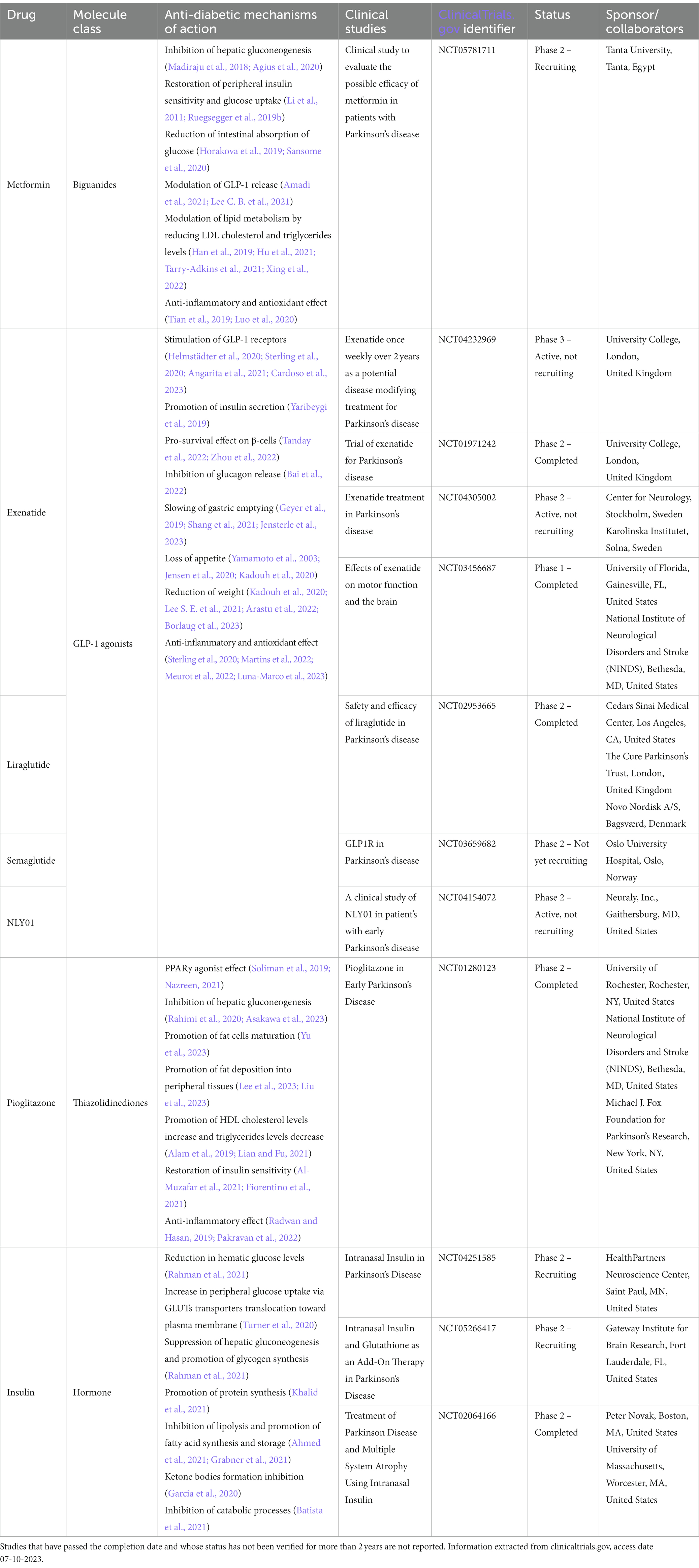
Table 1. Clinical trials on anti-diabetic drugs in PD.
In line with this premise, this review focuses on the compilation of a list of main drugs initially developed for treating hyperglycemia in the context of diabetes, but the activation of whose principal molecular pathways is potentially beneficial in the context of PD. The knowledge of these drugs from a pharmacokinetic and toxicological point of view has great advantages, allowing for drastically shortened trial times.
6.1. Metformin
Metformin is considered the first-line treatment for T2DM since the early 2000s, according to the guidelines of the International Diabetes Foundation (IDF Clinical Guidelines Task Force, 2006), but it is currently also considered a prime candidate for PD therapy. It inhibits hepatic gluconeogenesis, increases peripheral insulin sensitivity and glucagon-like peptide 1 (GLP-1) secretion, promoting peripheral glucose uptake, and reducing its bowel absorption at the same time. The mechanism of action is thought to be mainly through the inhibition of complex I of the mitochondrial transport chain, lowering ATP levels and indirectly activating the cellular metabolic state sensor AMPK (Zhou et al., 2016; Bahne et al., 2018; Baker et al., 2021; Drzewoski and Hanefeld, 2021). This increases fatty acid oxidation, reduces ROS production, and inhibits mTORC1 activating autophagic pathways and lysosomal biogenesis (Amin et al., 2019; Ma et al., 2023). Metformin also activates several downstream interactors including Bcl-1, CREB and PGC1, that promote cell viability and rescue mitochondrial defects, increasing their mass and improving their function (Kang et al., 2017; de Marañón et al., 2022). Finally, by modulation of Nrf2, FoxO3 and NF-κB activity, it exerts a protective effect in astrocytes and microglia (Ryu et al., 2018, 2020; Zhou et al., 2021).
In recent years, metformin has shown neuroprotective effects in both in vitro and in vivo models of PD, in the latter case by decreasing the loss of dopaminergic neurons and reducing motor symptoms, countering phospho-Ser129-aS aggregation in several ways. Pérez-Revuelta and colleagues have observed that metformin promotes the activity of PP2A, an enzyme of primary importance in reducing intracellular levels of phospho-Ser129-aS (Pérez-Revuelta et al., 2014; Greco et al., 2021); PP2A activity has indeed been shown to inhibit mTOR, promoting autophagy. Experiments in C. elegans and murine models of PD have both confirmed the effect of metformin in lowering phospho-Ser129-aS levels through an evolutionarily conserved mechanism (Pérez-Revuelta et al., 2014; Katila et al., 2017; Saewanee et al., 2021). Finally, metformin shows anti-glycating properties, acting as a scavenger of the aldehydic part of MG, preventing the accumulation of AGEs in subjects with T2DM (Kinsky et al., 2016).
6.2. GLP-1 agonists
Secreted by intestinal cells in response to food intake, especially carbohydrates, GLP-1 acts on several organs regulating glucose homeostasis. It stimulates insulin secretion from pancreatic β-cells in a glucose-dependent manner, reduces the secretion of glucagon, slows down gastric emptying, reduces the perception of hunger and thirst, and enhances peripheral glucose uptake (Hira et al., 2021).
Studies have shown that GLP-1 can inhibit mTOR stimulating autophagy in pancreatic β-cells and neurons. The binding of GLP-1 with its ubiquitously expressed membrane receptor GLP-1R triggers activation of the PI3K/AKT pathway. Subsequently, several downstream actors such as GSK3-β, FoxO1, NF-κB and Nrf2 exert cytoprotective effects (Buteau et al., 2006; Dai et al., 2013; Park et al., 2018; Costantino and Paneni, 2019; Li et al., 2020).
Interestingly, GLP-1R presence has been observed in the dopaminergic neuron of the encephalic area postrema, known for its role in modulating autonomic and reward responses, but also addiction mechanisms. GLP-1 seems therefore to be involved, at least in this specific region of the nervous system, in the modulation of dopamine synthesis and release (Yamamoto et al., 2003; Jensen et al., 2020).
Among the most important GLP-1R artificial agonists are exenatide, liraglutide, lixisenatide, semaglutide and NLY01. This class of molecules, found to be neuroprotective in mouse models treated with 6-OHDA or MPTP, is now a subject of interest for human use (Aviles-Olmos et al., 2013; Aslan et al., 2014; Jalewa et al., 2017). Promising results have been observed, with early-PD patients responding better to the treatment and showing motor and cognitive improvements (Athauda et al., 2019).
6.3. Glitazones
Glitazones, which includes pioglitazone, lobeglitazone and rosiglitazone are a class of oral antidiabetic drugs used in patients where first-line therapies fail to achieve glycemic control. Exerting their effect as agonists of the peroxisome proliferator-activated receptor PPARγ, a transcription factor of the nuclear ligand-activated receptor family, they increase tissues’ insulin sensitivity (Nanjan et al., 2018).
PPARγ induces the expression of a plethora of genes related to carbohydrate and lipid metabolism; at an encephalic level, expressed mainly in regions controlling reward and movement mechanisms such as the basal ganglia it has neuroprotective, with antioxidant, anti-inflammatory and anti-apoptotic properties (Warden et al., 2016; Villapol, 2018; Sonne et al., 2023). These effects, also linked to the downstream NF-κB signaling pathway activation, appear to be able to attenuate cognitive decline in various neurological diseases and neurodegenerative conditions (Delerive et al., 1999; Behl et al., 2021).
In several in vivo studies in PD-induced mice models, PPARγ agonists have shown a protective effect on dopaminergic neurons, with an improvement in motor symptoms. This is probably due to an inhibition of MAO-B, COX-1, COX-2 and iNOS activity, which reduce inflammation and preserve mitochondrial function and morphology (Xing et al., 2007; Quinn et al., 2008; Barbiero et al., 2014).
6.4. Insulin
The neuromodulatory, neurotrophic, and neuroprotective properties of insulin on the encephalon are well-known, therefore, its use in neurodegenerative contexts presents several potential beneficial effects (Yang et al., 1981; Shah and Hausman, 1993; Shuaib et al., 1995). However, two problems have hindered this approach until recently: the poor permeability of the blood–brain barrier (BBB) to its passage (Margolis and Altszuler, 1967) and the effect on hematic glycemia when parenterally administered. Therefore, new formulations have been studied to circumvent this problem; the most promising is via inhalation which exploits the permeability of the cribriform plate, shortening drug delivery to the target site and allowing precise modulation of administration. Several in vivo studies on murine PD models have demonstrated the efficacy of the molecule in this form and its neuroprotective activity (Fine et al., 2020; Iravanpour et al., 2021). Rajasekar and colleagues have shown how the use of intranasal insulin in mice treated with streptozocin was able to improve insulin-signaling cascade and attenuate neuroinflammation, neuronal loss, and protein aggregation. This was explained to be due to the modulation of NF-κB and PI3K/Akt pathway (Rajasekar et al., 2017).
Indeed, when intranasal insulin is administered, a canonical activation of PI3K/AKT and MAPK pathways is observed. From a neuroprotective perspective, there is an increased hypoxia-inducible factor-1 (HIF-1) activity, resulting in angiogenesis and endothelial proliferation (Zelzer, 1998). Furthermore, altered insulin signalling is associated with reduced mitochondrial fusion processes at the expense of organelle fission, resulting in increased ROS and reduced ATP levels (Kelley et al., 2002; Bach et al., 2003; Jheng et al., 2012).
Ruegsegger and colleagues demonstrated the ability of intranasal insulin to counteract these processes through the observation of an increase in the MFN1, MFN2, and OPA1 proteins, and a decrease in the DRP1 protein, involved in mitochondrial fusion and fission, respectively, (Ruegsegger et al., 2019a). Moreover, insulin also regulates mitochondrial quality control mechanisms by restoring the compromised activity of PINK1 and DJ-1. This appears particularly beneficial in brain areas of high mitochondrial activity such as the hypothalamus, and hippocampus (Onphachanh et al., 2017; Ruegsegger et al., 2019a; Su et al., 2020). Finally, insulin contributes to the removal of aggregates by promoting PP2A activity, which inhibits mTOR1 and dephosphorylates ULK1, initiating the autophagic process (Axe et al., 2008).
A 2018 clinical interventional study conducted at the University of Massachusetts on 15 patients for 4 weeks, involving intranasal administration of insulin, showed encouraging results. In this study, an improvement in motor performance, visuospatial memory, and verbal fluency was observed in PD subjects compared to placebo, due to a better ability to draw on mnemonic data (Novak et al., 2019).
7. Conclusion
To date, multiple pieces of evidence link T2DM to the onset of PD and other proteinopathies. The absence of cures for such medical conditions and the expected increase in their incidence in the coming years is driving clinical research. Hyperglycemia and insulin resistance, with the resulting protein aggregation, oxidative stress, and mitochondria dysfunction represent important and promising common medical targets, justifying interest in antidiabetic drugs, albeit the apparently different clinical settings. A large body of evidence on the efficacy of such drugs on pathways known to be involved in proteostasis and disrupted in PD exists. The data gathered in in vivo and in vitro models, together with the results of clinical trials leads to the view that the use of antidiabetic drugs, in combination with the current symptomatic medications is extremely encouraging.
Author contributions
MG and AM conceptualized and drafted the original article, then revised it once in its final form. DM wrote a section of the manuscript and prepared the graphical abstract. CC performed data curation. MM funded the publication process, contributed to conception and revised its final form. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by: (i) Italian Ministry of University and Research thanks to the PRIN 2017 project (MIUR) prot. 2017J92TM5 “enhancement of autophagy for the therapy of liver diseases” and to the innovative PhD position made available to AM; (ii) Apulia Region thanks to the post-doc position of the POC PUGLIA FESR ESF 2014/2020 entitled “PaRTiRe - Parkinson’s research through the voice” “Riparti project” made available to MG; (iii) Concorde Project, with grant PNRA18_0 0071_Prot. 20891.21-11-2019, assigned to Michele Samaja, Department of Health Sciences, University of Milan, Milan, Italy. We must also thank the “Alessia Pallara” Foundation for its constant economic support to our Physiology Lab at University of Salento.
Acknowledgments
The authors would like to thank Antonio Danieli for his constant support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abeliovich, A., Schmitz, Y., Fariñas, I., Choi-Lundberg, D., Ho, W.-H., Castillo, P. E., et al. (2000). Mice lacking α-Synuclein display functional deficits in the nigrostriatal dopamine system. Neuron 25, 239–252. doi: 10.1016/S0896-6273(00)80886-7
Agius, L., Ford, B. E., and Chachra, S. S. (2020). The metformin mechanism on gluconeogenesis and AMPK activation: the metabolite perspective. Int. J. Mol. Sci. 21:3240. doi: 10.3390/ijms21093240
Ahmed, B., Sultana, R., and Greene, M. W. (2021). Adipose tissue and insulin resistance in obese. Biomed. Pharmacother. 137:111315. doi: 10.1016/j.biopha.2021.111315
Alam, F., Islam, M. A., Mohamed, M., Ahmad, I., Kamal, M. A., Donnelly, R., et al. (2019). Efficacy and safety of pioglitazone monotherapy in type 2 diabetes mellitus: a systematic review and Meta-analysis of randomised controlled trials. Sci. Rep. 9:5389. doi: 10.1038/s41598-019-41854-2
Al-Muzafar, H. M., Alshehri, F. S., and Amin, K. A. (2021). The role of pioglitazone in antioxidant, anti-inflammatory, and insulin sensitivity in a high fat-carbohydrate diet-induced rat model of insulin resistance. Brazilian J. Med. Biol. Res. 54:e10782. doi: 10.1590/1414-431x2020e10782
Alvarez-Erviti, L., Rodriguez-Oroz, M. C., Cooper, J. M., Caballero, C., Ferrer, I., Obeso, J. A., et al. (2010). Chaperone-mediated autophagy markers in Parkinson disease brains. Arch. Neurol. 67, 1464–1472. doi: 10.1001/archneurol.2010.198
Amadi, J. A., Amadi, P. U., Njoku, U. C., and Osuoha, J. O. (2021). Potentiation of incretin hormones and modulation of metabolic enzymes as possible mechanisms behind the insulin sensitizing effects of cabbage-metformin treatment. Transl. Res. 230, 44–54. doi: 10.1016/j.trsl.2020.10.008
Amin, S., Lux, A., and O’Callaghan, F. (2019). The journey of metformin from glycaemic control to mTOR inhibition and the suppression of tumour growth. Br. J. Clin. Pharmacol. 85, 37–46. doi: 10.1111/bcp.13780
Angarita, G. A., Matuskey, D., Pittman, B., Costeines, J. L., Potenza, M. N., Jastreboff, A. M., et al. (2021). Testing the effects of the GLP-1 receptor agonist exenatide on cocaine self-administration and subjective responses in humans with cocaine use disorder. Drug Alcohol Depend. 221:108614. doi: 10.1016/j.drugalcdep.2021.108614
Arastu, N., Cummins, O., Uribe, W., and Nemec, E. C. (2022). Efficacy of subcutaneous semaglutide compared to placebo for weight loss in obese, non-diabetic adults: a systematic review & meta-analysis. Int. J. Clin. Pharm. 44, 852–859. doi: 10.1007/s11096-022-01428-1
Arnold, S. E., Arvanitakis, Z., Macauley-Rambach, S. L., Koenig, A. M., Wang, H.-Y., Ahima, R. S., et al. (2018). Brain insulin resistance in type 2 diabetes and Alzheimer disease: concepts and conundrums. Nat. Rev. Neurol. 14, 168–181. doi: 10.1038/nrneurol.2017.185
Asakawa, M., Takagi, N., Hamada, D., Yamasaki, Y., and Katsuta, H. (2023). Efficacy of 3 months of additional pioglitazone treatment in type 2 diabetes patients with alcoholic fatty liver disease. Diabetol. Int. 14, 243–251. doi: 10.1007/s13340-023-00619-z
Ashok, A., and Singh, N. (2018). Prion protein modulates glucose homeostasis by altering intracellular iron. Sci. Rep. 8:6556. doi: 10.1038/s41598-018-24786-1
Aslan, I. R., Ranadive, S. A., Valle, I., Kollipara, S., Noble, J. A., and Vaisse, C. (2014). The melanocortin system and insulin resistance in humans: insights from a patient with complete POMC deficiency and type 1 diabetes mellitus. Int. J. Obes. 38, 148–151. doi: 10.1038/ijo.2013.53
Athauda, D., Maclagan, K., Budnik, N., Zampedri, L., Hibbert, S., Aviles-Olmos, I., et al. (2019). Post hoc analysis of the exenatide-PD trial-factors that predict response. Eur. J. Neurosci. 49, 410–421. doi: 10.1111/ejn.14096
Aviles-Olmos, I., Dickson, J., Kefalopoulou, Z., Djamshidian, A., Ell, P., Soderlund, T., et al. (2013). Exenatide and the treatment of patients with Parkinson’s disease. J. Clin. Invest. 123, 2730–2736. doi: 10.1172/JCI68295
Axe, E. L., Walker, S. A., Manifava, M., Chandra, P., Roderick, H. L., Habermann, A., et al. (2008). Autophagosome formation from membrane compartments enriched in phosphatidylinositol 3-phosphate and dynamically connected to the endoplasmic reticulum. J. Cell Biol. 182, 685–701. doi: 10.1083/jcb.200803137
Bach, D., Pich, S., Soriano, F. X., Vega, N., Baumgartner, B., Oriola, J., et al. (2003). Mitofusin-2 determines mitochondrial network architecture and mitochondrial metabolism. J. Biol. Chem. 278, 17190–17197. doi: 10.1074/jbc.M212754200
Bahne, E., Sun, E. W. L., Young, R. L., Hansen, M., Sonne, D. P., Hansen, J. S., et al. (2018). Metformin-induced glucagon-like peptide-1 secretion contributes to the actions of metformin in type 2 diabetes. JCI Insight 3:e93936. doi: 10.1172/jci.insight.93936
Bai, C., Wang, Y., Niu, Z., Guan, Y., Huang, J., Nian, X., et al. (2022). Exenatide improves hepatocyte insulin resistance induced by different regional adipose tissue. Front. Endocrinol. 13:1012904. doi: 10.3389/fendo.2022.1012904
Baker, C., Retzik-Stahr, C., Singh, V., Plomondon, R., Anderson, V., and Rasouli, N. (2021). Should metformin remain the first-line therapy for treatment of type 2 diabetes? Ther. Adv. Endocrinol. Metab. 12:204201882098022. doi: 10.1177/2042018820980225
Barbiero, J. K., Santiago, R. M., Persike, D. S., da Silva Fernandes, M. J., Tonin, F. S., da Cunha, C., et al. (2014). Neuroprotective effects of peroxisome proliferator-activated receptor alpha and gamma agonists in model of parkinsonism induced by intranigral 1-methyl-4-phenyl-1,2,3,6-tetrahyropyridine. Behav. Brain Res. 274, 390–399. doi: 10.1016/j.bbr.2014.08.014
Batista, T. M., Haider, N., and Kahn, C. R. (2021). Defining the underlying defect in insulin action in type 2 diabetes. Diabetologia 64, 994–1006. doi: 10.1007/s00125-021-05415-5
Bayer, T. A. (2015). Proteinopathies, a core concept for understanding and ultimately treating degenerative disorders? Eur. Neuropsychopharmacol. 25, 713–724. doi: 10.1016/j.euroneuro.2013.03.007
Behl, T., Madaan, P., Sehgal, A., Singh, S., Sharma, N., Bhatia, S., et al. (2021). Elucidating the neuroprotective role of PPARs in Parkinson’s disease: a neoteric and prospective target. Int. J. Mol. Sci. 22:10161. doi: 10.3390/ijms221810161
Blair, J. B., James, M. E., and Foster, J. L. (1979). Adrenergic control of glucose output and adenosine 3′:5′-monophosphate levels in hepatocytes from juvenile and adult rats. J. Biol. Chem. 254, 7579–7584. doi: 10.1016/S0021-9258(18)35983-0
Borlaug, B. A., Kitzman, D. W., Davies, M. J., Rasmussen, S., Barros, E., Butler, J., et al. (2023). Semaglutide in HFpEF across obesity class and by body weight reduction: a prespecified analysis of the STEP-HFpEF trial. Nat. Med. 29, 2358–2365. doi: 10.1038/s41591-023-02526-x
Burbulla, L. F., Song, P., Mazzulli, J. R., Zampese, E., Wong, Y. C., Jeon, S., et al. (2017). Dopamine oxidation mediates mitochondrial and lysosomal dysfunction in Parkinson’s disease. Science 357, 1255–1261. doi: 10.1126/science.aam9080
Burré, J., Sharma, M., and Südhof, T. C. (2014). α-Synuclein assembles into higher-order multimers upon membrane binding to promote SNARE complex formation. Proc. Natl. Acad. Sci. 111:E4274-83. doi: 10.1073/pnas.1416598111
Buteau, J., Spatz, M. L., and Accili, D. (2006). Transcription factor FoxO1 mediates glucagon-like Peptide-1 effects on pancreatic β-cell mass. Diabetes 55, 1190–1196. doi: 10.2337/db05-0825
Cabin, D. E., Shimazu, K., Murphy, D., Cole, N. B., Gottschalk, W., McIlwain, K. L., et al. (2002). Synaptic vesicle depletion correlates with attenuated synaptic responses to prolonged repetitive stimulation in mice lacking α-Synuclein. J. Neurosci. 22, 8797–8807. doi: 10.1523/JNEUROSCI.22-20-08797.2002
Canet-Avilés, R. M., Wilson, M. A., Miller, D. W., Ahmad, R., McLendon, C., Bandyopadhyay, S., et al. (2004). The Parkinson’s disease protein DJ-1 is neuroprotective due to cysteine-sulfinic acid-driven mitochondrial localization. Proc. Natl. Acad. Sci. 101, 9103–9108. doi: 10.1073/pnas.0402959101
Cardoso, L. E. M., Marinho, T. S., Martins, F. F., Aguila, M. B., and Mandarim-de-Lacerda, C. A. (2023). Treatment with semaglutide, a GLP-1 receptor agonist, improves extracellular matrix remodeling in the pancreatic islet of diet-induced obese mice. Life Sci. 319:121502. doi: 10.1016/j.lfs.2023.121502
Cereda, E., Barichella, M., Cassani, E., Caccialanza, R., and Pezzoli, G. (2012). Clinical features of Parkinson disease when onset of diabetes came first: a case-control study. Neurology 78, 1507–1511. doi: 10.1212/WNL.0b013e3182553cc9
Cereda, E., Barichella, M., Pedrolli, C., Klersy, C., Cassani, E., Caccialanza, R., et al. (2011). Diabetes and risk of Parkinson’s disease. Diabetes Care 34, 2614–2623. doi: 10.2337/dc11-1584
Choudhury, S. P., Bano, S., Sen, S., Suchal, K., Kumar, S., Nikolajeff, F., et al. (2022). Altered neural cell junctions and ion-channels leading to disrupted neuron communication in Parkinson’s disease. NPJ Park. Dis. 8:66. doi: 10.1038/s41531-022-00324-9
Cooper, G. J., Willis, A. C., Clark, A., Turner, R. C., Sim, R. B., and Reid, K. B. (1987). Purification and characterization of a peptide from amyloid-rich pancreases of type 2 diabetic patients. Proc. Natl. Acad. Sci. 84, 8628–8632. doi: 10.1073/pnas.84.23.8628
Costantino, S., and Paneni, F. (2019). GLP-1-based therapies to boost autophagy in cardiometabolic patients: from experimental evidence to clinical trials. Vasc. Pharmacol. 115, 64–68. doi: 10.1016/j.vph.2019.03.003
Craft, S., Dagogo-Jack, S. E., Wiethop, B. V., Murphy, C., Nevins, R. T., Fleischman, S., et al. (1993). Effects of hyperglycemia on memory and hormone levels in dementia of the Alzheimer type: a longitudinal study. Behav. Neurosci. 107, 926–940. doi: 10.1037/0735-7044.107.6.926
Cui, K., and Li, Z. (2023). Identification and analysis of type 2 diabetes-mellitus-associated autophagy-related genes. Front. Endocrinol. 14:1164112. doi: 10.3389/fendo.2023.1164112
Dai, Y., Mehta, J. L., and Chen, M. (2013). Glucagon-like Peptide-1 receptor agonist Liraglutide inhibits Endothelin-1 in endothelial cell by repressing nuclear factor-kappa B activation. Cardiovasc. Drugs Ther. 27, 371–380. doi: 10.1007/s10557-013-6463-z
de Bartolomeis, A., De Simone, G., De Prisco, M., Barone, A., Napoli, R., Beguinot, F., et al. (2023). Insulin effects on core neurotransmitter pathways involved in schizophrenia neurobiology: A meta-analysis of preclinical studies. Implications for the treatment. Mol. Psychiatry. doi: 10.1038/s41380-023-02065-4 [Epub ahead of print].
de Marañón, A. M., Díaz-Pozo, P., Canet, F., Díaz-Morales, N., Abad-Jiménez, Z., López-Domènech, S., et al. (2022). Metformin modulates mitochondrial function and mitophagy in peripheral blood mononuclear cells from type 2 diabetic patients. Redox Biol. 53:102342. doi: 10.1016/j.redox.2022.102342
De Riccardis, L., Rizzo, F., Urso, E., Garzarelli, V., Intini, V., Greco, M., et al. (2019). Physiological role of prion protein in copper homeostasis and angiogenic mechanisms of endothelial cells. Euro Biotech J. 3, 57–70. doi: 10.2478/ebtj-2019-0007
Dehay, B., Bové, J., Rodríguez-Muela, N., Perier, C., Recasens, A., Boya, P., et al. (2010). Pathogenic lysosomal depletion in Parkinson’s disease. J. Neurosci. 30, 12535–12544. doi: 10.1523/JNEUROSCI.1920-10.2010
Del Coco, L., Greco, M., Inguscio, A., Munir, A., Danieli, A., Cossa, L., et al. (2023). Blood metabolite profiling of Antarctic expedition members: an 1H NMR spectroscopy-based study. Int. J. Mol. Sci. 24:8459. doi: 10.3390/ijms24098459
Delerive, P., De Bosscher, K., Besnard, S., Vanden Berghe, W., Peters, J. M., Gonzalez, F. J., et al. (1999). Peroxisome proliferator-activated receptor α negatively regulates the vascular inflammatory gene response by negative cross-talk with transcription factors NF-κB and AP-1. J. Biol. Chem. 274, 32048–32054. doi: 10.1074/jbc.274.45.32048
Diers, A. R., Broniowska, K. A., Chang, C.-F., and Hogg, N. (2012). Pyruvate fuels mitochondrial respiration and proliferation of breast cancer cells: effect of monocarboxylate transporter inhibition. Biochem. J. 444, 561–571. doi: 10.1042/BJ20120294
Dorsey, E. R., and Bloem, B. R. (2018). The Parkinson pandemic—a call to action. JAMA Neurol. 75:9. doi: 10.1001/jamaneurol.2017.3299
Driver, J. A., Smith, A., Buring, J. E., Gaziano, J. M., Kurth, T., and Logroscino, G. (2008). Prospective cohort study of type 2 diabetes and the risk of Parkinson’s disease. Diabetes Care 31, 2003–2005. doi: 10.2337/dc08-0688
Drzewoski, J., and Hanefeld, M. (2021). The current and potential therapeutic use of metformin—the good old drug. Pharmaceuticals. 14:122. doi: 10.3390/ph14020122
Du, X.-L., Edelstein, D., Rossetti, L., Fantus, I. G., Goldberg, H., Ziyadeh, F., et al. (2000). Hyperglycemia-induced mitochondrial superoxide overproduction activates the hexosamine pathway and induces plasminogen activator inhibitor-1 expression by increasing Sp1 glycosylation. Proc. Natl. Acad. Sci. 97, 12222–12226. doi: 10.1073/pnas.97.22.12222
Du, X., Matsumura, T., Edelstein, D., Rossetti, L., Zsengellér, Z., Szabó, C., et al. (2003). Inhibition of GAPDH activity by poly (ADP-ribose) polymerase activates three major pathways of hyperglycemic damage in endothelial cells. J. Clin. Invest. 112, 1049–1057. doi: 10.1172/JCI18127
Duarte, A. I., Santos, P., Oliveira, C. R., Santos, M. S., and Rego, A. C. (2008). Insulin neuroprotection against oxidative stress is mediated by Akt and GSK-3β signaling pathways and changes in protein expression. Biochim. Biophys. Acta-Mol. Cell Res. 1783, 994–1002. doi: 10.1016/j.bbamcr.2008.02.016
Dunn, L., Allen, G. F., Mamais, A., Ling, H., Li, A., Duberley, K. E., et al. (2014). Dysregulation of glucose metabolism is an early event in sporadic Parkinson’s disease. Neurobiol. Aging 35, 1111–1115. doi: 10.1016/j.neurobiolaging.2013.11.001
Fellner, L., Gabassi, E., Haybaeck, J., and Edenhofer, F. (2021). Autophagy in α-synucleinopathies — an overstrained system. Cells 10:3143. doi: 10.3390/cells10113143
Fine, J. M., Stroebel, B. M., Faltesek, K. A., Terai, K., Haase, L., Knutzen, K. E., et al. (2020). Intranasal delivery of low-dose insulin ameliorates motor dysfunction and dopaminergic cell death in a 6-OHDA rat model of Parkinson’s disease. Neurosci. Lett. 714:134567. doi: 10.1016/j.neulet.2019.134567
Fiorentino, T. V., Monroy, A., Kamath, S., Sotero, R., Cas, M. D., Daniele, G., et al. (2021). Pioglitazone corrects dysregulation of skeletal muscle mitochondrial proteins involved in ATP synthesis in type 2 diabetes. Metabolism 114:154416. doi: 10.1016/j.metabol.2020.154416
Fujiwara, H., Hasegawa, M., Dohmae, N., Kawashima, A., Masliah, E., Goldberg, M. S., et al. (2002). α-Synuclein is phosphorylated in synucleinopathy lesions. Nat. Cell Biol. 4, 160–164. doi: 10.1038/ncb748
Garcia, E., Shalaurova, I., Matyus, S. P., Oskardmay, D. N., Otvos, J. D., Dullaart, R. P. F., et al. (2020). Ketone bodies are mildly elevated in subjects with type 2 diabetes mellitus and are inversely associated with insulin resistance as measured by the lipoprotein insulin resistance index. J. Clin. Med. 9:321. doi: 10.3390/jcm9020321
García-Cáceres, C., Quarta, C., Varela, L., Gao, Y., Gruber, T., Legutko, B., et al. (2016). Astrocytic insulin signaling couples brain glucose uptake with nutrient availability. Cells 166, 867–880. doi: 10.1016/j.cell.2016.07.028
Gasnier, B. (2000). The loading of neurotransmitters into synaptic vesicles. Biochimie 82, 327–337. doi: 10.1016/S0300-9084(00)00221-2
Gayen, M., Benoit, M. R., Fan, Q., Hudobenko, J., and Yan, R. (2022). The CX3CL1 intracellular domain exhibits neuroprotection via insulin receptor/insulin-like growth factor receptor signaling. J. Biol. Chem. 298:102532. doi: 10.1016/j.jbc.2022.102532
Geng, X., Li, L., Bottino, R., Balamurugan, A. N., Bertera, S., Densmore, E., et al. (2007). Antidiabetic sulfonylurea stimulates insulin secretion independently of plasma membrane K ATP channels. Am. J. Physiol. Metab. 293, E293–E301. doi: 10.1152/ajpendo.00016.2007
Geyer, M. C., Sullivan, T., Tai, A., Morton, J. M., Edwards, S., Martin, A. J., et al. (2019). Exenatide corrects postprandial hyperglycaemia in young people with cystic fibrosis and impaired glucose tolerance: a randomized crossover trial. Diabetes. Obes. Metab. 21, 700–704. doi: 10.1111/dom.13544
Grabner, G. F., Xie, H., Schweiger, M., and Zechner, R. (2021). Lipolysis: cellular mechanisms for lipid mobilization from fat stores. Nat. Metab. 3, 1445–1465. doi: 10.1038/s42255-021-00493-6
Greco, M., Spinelli, C. C., De Riccardis, L., Buccolieri, A., Di Giulio, S., Musarò, D., et al. (2021). Copper dependent modulation of α-Synuclein phosphorylation in differentiated SHSY5Y neuroblastoma cells. Int. J. Mol. Sci. 22:2038. doi: 10.3390/ijms22042038
Guadagnolo, D., Piane, M., Torrisi, M. R., Pizzuti, A., and Petrucci, S. (2021). Genotype-phenotype correlations in monogenic Parkinson disease: a review on clinical and molecular findings. Front. Neurol. 12:648588. doi: 10.3389/fneur.2021.648588
Han, Y., Xie, H., Liu, Y., Gao, P., Yang, X., and Shen, Z. (2019). Effect of metformin on all-cause and cardiovascular mortality in patients with coronary artery diseases: a systematic review and an updated meta-analysis. Cardiovasc. Diabetol. 18:96. doi: 10.1186/s12933-019-0900-7
Hao, L.-Y., Giasson, B. I., and Bonini, N. M. (2010). DJ-1 is critical for mitochondrial function and rescues PINK1 loss of function. Proc. Natl. Acad. Sci. 107, 9747–9752. doi: 10.1073/pnas.0911175107
Helmstädter, J., Frenis, K., Filippou, K., Grill, A., Dib, M., Kalinovic, S., et al. (2020). Endothelial GLP-1 (glucagon-like Peptide-1) receptor mediates cardiovascular protection by Liraglutide in mice with experimental arterial hypertension. Arterioscler. Thromb. Vasc. Biol. 40, 145–158. doi: 10.1161/atv.0000615456.97862.30
Heras-Sandoval, D., Pérez-Rojas, J. M., Hernández-Damián, J., and Pedraza-Chaverri, J. (2014). The role of PI3K/AKT/mTOR pathway in the modulation of autophagy and the clearance of protein aggregates in neurodegeneration. Cell. Signal. 26, 2694–2701. doi: 10.1016/j.cellsig.2014.08.019
Hira, T., Trakooncharoenvit, A., Taguchi, H., and Hara, H. (2021). Improvement of glucose tolerance by food factors having glucagon-like Peptide-1 releasing activity. Int. J. Mol. Sci. 22:6623. doi: 10.3390/ijms22126623
Horakova, O., Kroupova, P., Bardova, K., Buresova, J., Janovska, P., Kopecky, J., et al. (2019). Metformin acutely lowers blood glucose levels by inhibition of intestinal glucose transport. Sci. Rep. 9:6156. doi: 10.1038/s41598-019-42531-0
Hou, X., Watzlawik, J. O., Fiesel, F. C., and Springer, W. (2020). Autophagy in Parkinson’s disease. J. Mol. Biol. 432, 2651–2672. doi: 10.1016/j.jmb.2020.01.037
Hu, D., Guo, Y., Wu, R., Shao, T., Long, J., Yu, B., et al. (2021). New insight into metformin-induced cholesterol-lowering effect crosstalk between glucose and cholesterol homeostasis via ChREBP (carbohydrate-responsive element-binding protein)-mediated PCSK9 (Proprotein convertase Subtilisin/Kexin type 9) regulation. Arterioscler. Thromb. Vasc. Biol. 41, e208–e223. doi: 10.1161/ATVBAHA.120.315708
Huang, Y., Wan, Z., Wang, Z., and Zhou, B. (2019). Insulin signaling in Drosophila melanogaster mediates Aβ toxicity. Commun. Biol. 2:13. doi: 10.1038/s42003-018-0253-x
IDF Clinical Guidelines Task Force (2006). Global guideline for type 2 diabetes: recommendations for standard, comprehensive, and minimal care. Diabet. Med. 23, 579–593. doi: 10.1111/j.1464-5491.2006.01918.x
Iizuka, K., Bruick, R. K., Liang, G., Horton, J. D., and Uyeda, K. (2004). Deficiency of carbohydrate response element-binding protein (ChREBP) reduces lipogenesis as well as glycolysis. Proc. Natl. Acad. Sci. 101, 7281–7286. doi: 10.1073/pnas.0401516101
Iravanpour, F., Dargahi, L., Rezaei, M., Haghani, M., Heidari, R., Valian, N., et al. (2021). Intranasal insulin improves mitochondrial function and attenuates motor deficits in a rat 6-OHDA model of Parkinson’s disease. CNS Neurosci. Ther. 27, 308–319. doi: 10.1111/cns.13609
Jalewa, J., Sharma, M. K., Gengler, S., and Hölscher, C. (2017). A novel GLP-1/GIP dual receptor agonist protects from 6-OHDA lesion in a rat model of Parkinson’s disease. Neuropharmacology 117, 238–248. doi: 10.1016/j.neuropharm.2017.02.013
Jao, C. C., Der-Sarkissian, A., Chen, J., and Langen, R. (2004). Structure of membrane-bound α-synuclein studied by site-directed spin labeling. Proc. Natl. Acad. Sci. 101, 8331–8336. doi: 10.1073/pnas.0400553101
Jensen, M. E., Galli, A., Thomsen, M., Jensen, K. L., Thomsen, G. K., Klausen, M. K., et al. (2020). Glucagon-like peptide-1 receptor regulation of basal dopamine transporter activity is species-dependent. Neurochem. Int. 138:104772. doi: 10.1016/j.neuint.2020.104772
Jensterle, M., Ferjan, S., Ležaič, L., Sočan, A., Goričar, K., Zaletel, K., et al. (2023). Semaglutide delays 4-hour gastric emptying in women with polycystic ovary syndrome and obesity. Diabetes Obes. Metab. 25, 975–984. doi: 10.1111/dom.14944
Jheng, H.-F., Tsai, P.-J., Guo, S.-M., Kuo, L.-H., Chang, C.-S., Su, I.-J., et al. (2012). Mitochondrial fission contributes to mitochondrial dysfunction and insulin resistance in skeletal muscle. Mol. Cell. Biol. 32, 309–319. doi: 10.1128/MCB.05603-11
Jucker, M., and Walker, L. C. (2018). Propagation and spread of pathogenic protein assemblies in neurodegenerative diseases. Nat. Neurosci. 21, 1341–1349. doi: 10.1038/s41593-018-0238-6
Kadouh, H., Chedid, V., Halawi, H., Burton, D. D., Clark, M. M., Khemani, D., et al. (2020). GLP-1 analog modulates appetite, taste preference, gut hormones, and regional body fat Stores in Adults with obesity. J. Clin. Endocrinol. Metab. 105, 1552–1563. doi: 10.1210/clinem/dgz140
Kang, H., Khang, R., Ham, S., Jeong, G. R., Kim, H., Jo, M., et al. (2017). Activation of the ATF2/CREB-PGC-1α pathway by metformin leads to dopaminergic neuroprotection. Oncotarget 8, 48603–48618. doi: 10.18632/oncotarget.18122
Karpowicz, R. J., Trojanowski, J. Q., and Lee, V. M.-Y. (2019). Transmission of α-synuclein seeds in neurodegenerative disease: recent developments. Lab. Investig. 99, 971–981. doi: 10.1038/s41374-019-0195-z
Katila, N., Bhurtel, S., Shadfar, S., Srivastav, S., Neupane, S., Ojha, U., et al. (2017). Metformin lowers α-synuclein phosphorylation and upregulates neurotrophic factor in the MPTP mouse model of Parkinson’s disease. Neuropharmacology 125, 396–407. doi: 10.1016/j.neuropharm.2017.08.015
Katsyuba, E., Romani, M., Hofer, D., and Auwerx, J. (2020). NAD+ homeostasis in health and disease. Nat. Metab. 2, 9–31. doi: 10.1038/s42255-019-0161-5
Kelley, D. E., He, J., Menshikova, E. V., and Ritov, V. B. (2002). Dysfunction of mitochondria in human skeletal muscle in type 2 diabetes. Diabetes 51, 2944–2950. doi: 10.2337/diabetes.51.10.2944
Khalid, M., Alkaabi, J., Khan, M. A. B., and Adem, A. (2021). Insulin signal transduction perturbations in insulin resistance. Int. J. Mol. Sci. 22:8590. doi: 10.3390/ijms22168590
Kinsky, O. R., Hargraves, T. L., Anumol, T., Jacobsen, N. E., Dai, J., Snyder, S. A., et al. (2016). Metformin scavenges methylglyoxal to form a novel imidazolinone metabolite in humans. Chem. Res. Toxicol. 29, 227–234. doi: 10.1021/acs.chemrestox.5b00497
Kleinridders, A. (2016). Deciphering brain insulin receptor and insulin-like growth factor 1 receptor Signalling. J. Neuroendocrinol. 28. doi: 10.1111/jne.12433
Le Bacquer, O., Petroulakis, E., Paglialunga, S., Poulin, F., Richard, D., Cianflone, K., et al. (2007). Elevated sensitivity to diet-induced obesity and insulin resistance in mice lacking 4E-BP1 and 4E-BP2. J. Clin. Invest. 117, 387–396. doi: 10.1172/JCI29528
Lee, C. B., Chae, S. U., Jo, S. J., Jerng, U. M., and Bae, S. K. (2021). The relationship between the gut microbiome and metformin as a key for treating type 2 diabetes mellitus. Int. J. Mol. Sci. 22:3566. doi: 10.3390/ijms22073566
Lee, E., Korf, H., and Vidal-Puig, A. (2023). An adipocentric perspective on the development and progression of non-alcoholic fatty liver disease. J. Hepatol. 78, 1048–1062. doi: 10.1016/j.jhep.2023.01.024
Lee, S. E., Lee, N. Y., Kim, S. H., Kim, K.-A., and Kim, Y. S. (2021). Effect of liraglutide 3.0mg treatment on weight reduction in obese antipsychotic-treated patients. Psychiatry Res. 299:113830. doi: 10.1016/j.psychres.2021.113830
Lewis, M. R., and Lewis, W. H. (1915). Mitochondria (and other cytoplasmic structures) in tissue cultures. Am. J. Anat. 17, 339–401. doi: 10.1002/aja.1000170304
Li, Z., Li, S., Wang, N., Xue, P., and Li, Y. (2020). Liraglutide, a glucagon-like peptide-1 receptor agonist, suppresses osteoclastogenesis through the inhibition of NF-κB and MAPK pathways via GLP-1R. Biomed. Pharmacother. 130:110523. doi: 10.1016/j.biopha.2020.110523
Li, X., Zhang, N., Li, Y., Shi, Y., Li, D., Xie, Y., et al. (2011). Effects of metformin and rosiglitazone on peripheral insulin resistance and β-cell function in obesity: a double-blind, randomized, controlled study. J. Int. Med. Res. 39, 358–365. doi: 10.1177/147323001103900203
Lian, J., and Fu, J. (2021). Pioglitazone for NAFLD patients with prediabetes or type 2 diabetes mellitus: a meta-analysis. Front. Endocrinol. 12:615409. doi: 10.3389/fendo.2021.615409
Liao, P.-C., Wolken, D. M. A., Serrano, E., Srivastava, P., and Pon, L. A. (2020). Mitochondria-associated degradation pathway (MAD) function beyond the outer membrane. Cell Rep. 32:107902. doi: 10.1016/j.celrep.2020.107902
Liu, J., Wang, D., Xie, Z., Ding, L., Li, S., Ma, X., et al. (2023). Combination of pioglitazone and metformin actions on liver lipid metabolism in obese mice. Biomol. Ther. 13:1199. doi: 10.3390/biom13081199
Lu, L., Fu, D., Li, H., Liu, A., Li, J., and Zheng, G. (2014). Diabetes and risk of Parkinson’s disease: an updated Meta-analysis of case-control studies. PLoS One 9:e85781. doi: 10.1371/journal.pone.0085781
Luna-Marco, C., de Marañon, A. M., Hermo-Argibay, A., Rodriguez-Hernandez, Y., Hermenejildo, J., Fernandez-Reyes, M., et al. (2023). Effects of GLP-1 receptor agonists on mitochondrial function, inflammatory markers and leukocyte-endothelium interactions in type 2 diabetes. Redox Biol. 66:102849. doi: 10.1016/j.redox.2023.102849
Luo, X., Hu, R., Zheng, Y., Liu, S., and Zhou, Z. (2020). Metformin shows anti-inflammatory effects in murine macrophages through dicer/microribonucleic acid-34a-5p and microribonucleic acid-125b-5p. J. Diabetes Investig. 11, 101–109. doi: 10.1111/jdi.13074
Lutchmansingh, F. K., Hsu, J. W., Bennett, F. I., Badaloo, A. V., McFarlane-Anderson, N., Gordon-Strachan, G. M., et al. (2018). Glutathione metabolism in type 2 diabetes and its relationship with microvascular complications and glycemia. PLoS One 13:e0198626. doi: 10.1371/journal.pone.0198626
Ma, Z., Liu, Z., Li, X., Zhang, H., Han, D., Xiong, W., et al. (2023). Metformin collaborates with PINK1/Mfn2 overexpression to prevent cardiac injury by improving mitochondrial function. Biology 12:582. doi: 10.3390/biology12040582
Ma, L.-Y., Lv, Y.-L., Huo, K., Liu, J., Shang, S.-H., Fei, Y.-L., et al. (2017). Autophagy-lysosome dysfunction is involved in Aβ deposition in STZ-induced diabetic rats. Behav. Brain Res. 320, 484–493. doi: 10.1016/j.bbr.2016.10.031
Madiraju, A. K., Qiu, Y., Perry, R. J., Rahimi, Y., Zhang, X.-M., Zhang, D., et al. (2018). Metformin inhibits gluconeogenesis via a redox-dependent mechanism in vivo. Nat. Med. 24, 1384–1394. doi: 10.1038/s41591-018-0125-4
Margolis, R. U., and Altszuler, N. (1967). Insulin in the cerebrospinal fluid. Nature 215, 1375–1376. doi: 10.1038/2151375a0
Martinez-Valbuena, I., Amat-Villegas, I., Valenti-Azcarate, R., del Carmona-Abellan, M. M., Marcilla, I., Tuñon, M. T., et al. (2018). Interaction of amyloidogenic proteins in pancreatic β cells from subjects with synucleinopathies. Acta Neuropathol. 135, 877–886. doi: 10.1007/s00401-018-1832-0
Martinez-Valbuena, I., Valenti-Azcarate, R., Amat-Villegas, I., Marcilla, I., Marti-Andres, G., Caballero, M.-C., et al. (2021). Mixed pathologies in pancreatic β cells from subjects with neurodegenerative diseases and their interaction with prion protein. Acta Neuropathol. Commun. 9:64. doi: 10.1186/s40478-021-01171-0
Martins, F. F., Marinho, T. S., Cardoso, L. E. M., Barbosa-da-Silva, S., Souza-Mello, V., Aguila, M. B., et al. (2022). Semaglutide (GLP-1 receptor agonist) stimulates browning on subcutaneous fat adipocytes and mitigates inflammation and endoplasmic reticulum stress in visceral fat adipocytes of obese mice. Cell Biochem. Funct. 40, 903–913. doi: 10.1002/cbf.3751
Mathebula, S. D. (2015). Polyol pathway: a possible mechanism of diabetes complications in the eye. African Vis. Eye Heal. 74:5. doi: 10.4102/aveh.v74i1.13
Mazza, M. C., Shuck, S. C., Lin, J., Moxley, M. A., Termini, J., Cookson, M. R., et al. (2022). DJ-1 is not a deglycase and makes a modest contribution to cellular defense against methylglyoxal damage in neurons. J. Neurochem. 162, 245–261. doi: 10.1111/jnc.15656
Menéndez-González, M., Padilla-Zambrano, H., Tomás-Zapico, C., and García, B. (2018). Clearing extracellular alpha-Synuclein from cerebrospinal fluid: a new therapeutic strategy in Parkinson’s disease. Brain Sci. 8:52. doi: 10.3390/brainsci8040052
Meurot, C., Martin, C., Sudre, L., Breton, J., Bougault, C., Rattenbach, R., et al. (2022). Liraglutide, a glucagon-like peptide 1 receptor agonist, exerts analgesic, anti-inflammatory and anti-degradative actions in osteoarthritis. Sci. Rep. 12:1567. doi: 10.1038/s41598-022-05323-7
Milordini, G., Zacco, E., Percival, M., Puglisi, R., Dal Piaz, F., Temussi, P., et al. (2020). The role of glycation on the aggregation properties of IAPP. Front. Mol. Biosci. 7:104. doi: 10.3389/fmolb.2020.00104
Nanjan, M. J., Mohammed, M., Prashantha Kumar, B. R., and Chandrasekar, M. J. N. (2018). Thiazolidinediones as antidiabetic agents: a critical review. Bioorg. Chem. 77, 548–567. doi: 10.1016/j.bioorg.2018.02.009
Nazreen, S. (2021). Design, synthesis, and molecular docking studies of thiazolidinediones as PPAR-γ agonists and thymidylate synthase inhibitors. Arch. Pharm. (Weinheim). 354:e2100021. doi: 10.1002/ardp.202100021
Novak, P., Pimentel Maldonado, D. A., and Novak, V. (2019). Safety and preliminary efficacy of intranasal insulin for cognitive impairment in Parkinson disease and multiple system atrophy: a double-blinded placebo-controlled pilot study. PLoS One 14:e0214364. doi: 10.1371/journal.pone.0214364
Onphachanh, X., Lee, H. J., Lim, J. R., Jung, Y. H., Kim, J. S., Chae, C. W., et al. (2017). Enhancement of high glucose-induced PINK1 expression by melatonin stimulates neuronal cell survival: involvement of MT 2/Akt/NF-κB pathway. J. Pineal Res. 63:e12427. doi: 10.1111/jpi.12427
Pagano, G., Polychronis, S., Wilson, H., Giordano, B., Ferrara, N., Niccolini, F., et al. (2018). Diabetes mellitus and Parkinson disease. Neurology 90, e1654–e1662. doi: 10.1212/WNL.0000000000005475
Pakravan, G., Peymani, M., Abedpoor, N., Safaeinejad, Z., Yadegari, M., Derakhshan, M., et al. (2022). Antiapoptotic and anti-inflammatory effects of Pparγ agonist, pioglitazone, reversed dox-induced cardiotoxicity through mediating of miR-130a downregulation in C57BL/6 mice. J. Biochem. Mol. Toxicol. 36:e23041. doi: 10.1002/jbt.23041
Palikaras, K., Lionaki, E., and Tavernarakis, N. (2018). Mechanisms of mitophagy in cellular homeostasis, physiology and pathology. Nat. Cell Biol. 20, 1013–1022. doi: 10.1038/s41556-018-0176-2
Park, S. W., Mansur, R. B., Lee, Y., Lee, J.-H., Seo, M. K., Choi, A. J., et al. (2018). Liraglutide activates mTORC1 signaling and AMPA receptors in rat hippocampal neurons under toxic conditions. Front. Neurosci. 12:756. doi: 10.3389/fnins.2018.00756
Perez, R. G., Waymire, J. C., Lin, E., Liu, J. J., Guo, F., and Zigmond, M. J. (2002). A role for α-Synuclein in the regulation of dopamine biosynthesis. J. Neurosci. 22, 3090–3099. doi: 10.1523/JNEUROSCI.22-08-03090.2002
Pérez-Revuelta, B. I., Hettich, M. M., Ciociaro, A., Rotermund, C., Kahle, P. J., Krauss, S., et al. (2014). Metformin lowers Ser-129 phosphorylated α-synuclein levels via mTOR-dependent protein phosphatase 2A activation. Cell Death Dis. 5, –e1209. doi: 10.1038/cddis.2014.175
Polymeropoulos, M. H., Lavedan, C., Leroy, E., Ide, S. E., Dehejia, A., Dutra, A., et al. (1997). Mutation in the α-Synuclein gene identified in families with Parkinson’s disease. Science 276, 2045–2047. doi: 10.1126/science.276.5321.2045
Pomytkin, I., Costa-Nunes, J. P., Kasatkin, V., Veniaminova, E., Demchenko, A., Lyundup, A., et al. (2018). Insulin receptor in the brain: mechanisms of activation and the role in the CNS pathology and treatment. CNS Neurosci. Ther. 24:763. doi: 10.1111/cns.12866
Pupyshev, A. B., Tenditnik, M. V., Ovsyukova, M. V., Akopyan, A. A., Dubrovina, N. I., and Tikhonova, M. A. (2021). Restoration of Parkinson’s disease-like deficits by activating autophagy through mTOR-dependent and mTOR-independent mechanisms in pharmacological and transgenic models of Parkinson’s disease in mice. Bull. Exp. Biol. Med. 171, 425–430. doi: 10.1007/s10517-021-05242-z
Qu, J., Yan, H., Zheng, Y., Xue, F., Zheng, Y., Fang, H., et al. (2018). The molecular mechanism of alpha-Synuclein dependent regulation of protein phosphatase 2A activity. Cell. Physiol. Biochem. 47, 2613–2625. doi: 10.1159/000491657
Quinn, L. P., Crook, B., Hows, M. E., Vidgeon-Hart, M., Chapman, H., Upton, N., et al. (2008). The PPARγ agonist pioglitazone is effective in the MPTP mouse model of Parkinson’s disease through inhibition of monoamine oxidase B. Br. J. Pharmacol. 154, 226–233. doi: 10.1038/bjp.2008.78
Radwan, R. R., and Hasan, H. F. (2019). Pioglitazone ameliorates hepatic damage in irradiated rats via regulating anti-inflammatory and antifibrogenic signalling pathways. Free Radic. Res. 53, 748–757. doi: 10.1080/10715762.2019.1624742
Rahimi, M., Sajadimajd, S., Mahdian, Z., Hemmati, M., Malekkhatabi, P., Bahrami, G., et al. (2020). Characterization and anti-diabetic effects of the oligosaccharide fraction isolated from Rosa canina in STZ-induced diabetic rats. Carbohydr. Res. 489:107927. doi: 10.1016/j.carres.2020.107927
Rahman, M. S., Hossain, K. S., Das, S., Kundu, S., Adegoke, E. O., Rahman, M. A., et al. (2021). Role of insulin in health and disease: an update. Int. J. Mol. Sci. 22:6403. doi: 10.3390/ijms22126403
Rajasekar, N., Nath, C., Hanif, K., and Shukla, R. (2017). Intranasal insulin administration ameliorates Streptozotocin (ICV)-induced insulin receptor dysfunction, Neuroinflammation, Amyloidogenesis, and memory impairment in rats. Mol. Neurobiol. 54, 6507–6522. doi: 10.1007/s12035-016-0169-8
Reed, X., Bandrés-Ciga, S., Blauwendraat, C., and Cookson, M. R. (2019). The role of monogenic genes in idiopathic Parkinson’s disease. Neurobiol. Dis. 124, 230–239. doi: 10.1016/j.nbd.2018.11.012
Roger, L. J., and Fellows, R. E. (1980). Stimulation of ornithine decarboxylase activity by insulin in developing rat brain. Endocrinology 106, 619–625. doi: 10.1210/endo-106-2-619
Ruegsegger, G. N., Manjunatha, S., Summer, P., Gopala, S., Zabeilski, P., Dasari, S., et al. (2019a). Insulin deficiency and intranasal insulin alter brain mitochondrial function: a potential factor for dementia in diabetes. FASEB J. 33, 4458–4472. doi: 10.1096/fj.201802043R
Ruegsegger, G. N., Vanderboom, P. M., Dasari, S., Klaus, K. A., Kabiraj, P., McCarthy, C. B., et al. (2019b). Exercise and metformin counteract altered mitochondrial function in the insulin-resistant brain. JCI Insight 4:e130681. doi: 10.1172/jci.insight.130681
Ryu, Y.-K., Go, J., Park, H.-Y., Choi, Y.-K., Seo, Y. J., Choi, J. H., et al. (2020). Metformin regulates astrocyte reactivity in Parkinson’s disease and normal aging. Neuropharmacology 175:108173. doi: 10.1016/j.neuropharm.2020.108173
Ryu, Y.-K., Park, H.-Y., Go, J., Choi, D.-H., Kim, Y.-H., Hwang, J. H., et al. (2018). Metformin inhibits the development of l-DOPA-induced dyskinesia in a murine model of Parkinson’s disease. Mol. Neurobiol. 55, 5715–5726. doi: 10.1007/s12035-017-0752-7
Saewanee, N., Praputpittaya, T., Malaiwong, N., Chalorak, P., and Meemon, K. (2021). Neuroprotective effect of metformin on dopaminergic neurodegeneration and α-synuclein aggregation in C. elegans model of Parkinson’s disease. Neurosci. Res. 162, 13–21. doi: 10.1016/j.neures.2019.12.017
Sandyk, R. (1993). The relationship between diabetes mellitus and Parkinson’s disease. Int. J. Neurosci. 69, 125–130. doi: 10.3109/00207459309003322
Sansome, D. J., Xie, C., Veedfald, S., Horowitz, M., Rayner, C. K., and Wu, T. (2020). Mechanism of glucose-lowering by metformin in type 2 diabetes: role of bile acids. Diabetes. Obes. Metab. 22, 141–148. doi: 10.1111/dom.13869
Savica, R., Grossardt, B. R., Ahlskog, J. E., and Rocca, W. A. (2012). Metabolic markers or conditions preceding Parkinson’s disease: a case-control study. Mov. Disord. 27, 974–979. doi: 10.1002/mds.25016
Schernhammer, E., Hansen, J., Rugbjerg, K., Wermuth, L., and Ritz, B. (2011). Diabetes and the risk of developing Parkinson’s disease in Denmark. Diabetes Care 34, 1102–1108. doi: 10.2337/dc10-1333
Schubert, M., Brazil, D. P., Burks, D. J., Kushner, J. A., Ye, J., Flint, C. L., et al. (2003). Insulin receptor Substrate-2 deficiency impairs brain growth and promotes tau phosphorylation. J. Neurosci. 23, 7084–7092. doi: 10.1523/JNEUROSCI.23-18-07084.2003
Serhan, A., Aerts, J. L., Boddeke, E. W. G. M., and Kooijman, R. (2020). Neuroprotection by insulin-like growth Factor-1 in rats with ischemic stroke is associated with microglial changes and a reduction in Neuroinflammation. Neuroscience 426, 101–114. doi: 10.1016/j.neuroscience.2019.11.035
Shah, B. H., and Hausman, R. E. (1993). Effect of insulin on GABAergic development in the embryonic chick retina. Dev. Brain Res. 72, 151–158. doi: 10.1016/0165-3806(93)90180-I
Shang, P., Baker, M., Banks, S., Hong, S.-I., and Choi, D.-S. (2021). Emerging nondopaminergic medications for Parkinson’s disease: focusing on A2A receptor antagonists and GLP1 receptor agonists. J. Mov. Disord. 14, 193–203. doi: 10.14802/jmd.21035
Shuaib, A., Ijaz, M. S., Waqar, T., Voll, C., Kanthan, R., Miyashita, H., et al. (1995). Insulin elevates hippocampal GABA levels during ischemia. This is independent of its hypoglycemic effect. Neuroscience 67, 809–814. doi: 10.1016/0306-4522(95)00093-X
Simón-Sánchez, J., Schulte, C., Bras, J. M., Sharma, M., Gibbs, J. R., Berg, D., et al. (2009). Genome-wide association study reveals genetic risk underlying Parkinson’s disease. Nat. Genet. 41, 1308–1312. doi: 10.1038/ng.487
Soejima, A., Inoue, K., Takai, D., Kaneko, M., Ishihara, H., Oka, Y., et al. (1996). Mitochondrial DNA is required for regulation of glucose-stimulated insulin secretion in a mouse pancreatic Beta cell line, MIN6. J. Biol. Chem. 271, 26194–26199. doi: 10.1074/jbc.271.42.26194
Soliman, E., Behairy, S. F., El-maraghy, N. N., and Elshazly, S. M. (2019). PPAR-γ agonist, pioglitazone, reduced oxidative and endoplasmic reticulum stress associated with L-NAME-induced hypertension in rats. Life Sci. 239:117047. doi: 10.1016/j.lfs.2019.117047
Sonne, J., Reddy, V., and Beato, M. R. (2023). Neuroanatomy, Substantia Nigra. Treasure Island: StatPearls Publishing.
Sousa-Nunes, R., Yee, L. L., and Gould, A. P. (2011). Fat cells reactivate quiescent neuroblasts via TOR and glial insulin relays in Drosophila. Nature 471, 508–512. doi: 10.1038/nature09867
Spillantini, M. G., Schmidt, M. L., Lee, V. M.-Y., Trojanowski, J. Q., Jakes, R., and Goedert, M. (1997). α-Synuclein in Lewy bodies. Nature 388, 839–840. doi: 10.1038/42166
Sportelli, C., Urso, D., Jenner, P., and Chaudhuri, K. R. (2020). Metformin as a potential neuroprotective agent in prodromal Parkinson’s disease—viewpoint. Front. Neurol. 11, 1–10. doi: 10.3389/fneur.2020.00556
Steinberg, G. R., and Carling, D. (2019). AMP-activated protein kinase: the current landscape for drug development. Nat. Rev. Drug Discov. 18, 527–551. doi: 10.1038/s41573-019-0019-2
Sterling, J. K., Adetunji, M. O., Guttha, S., Bargoud, A. R., Uyhazi, K. E., Ross, A. G., et al. (2020). GLP-1 receptor agonist NLY01 reduces retinal inflammation and neuron death secondary to ocular hypertension. Cell Rep. 33:108271. doi: 10.1016/j.celrep.2020.108271
Strom, A., Strassburger, K., Schmuck, M., Shevalye, H., Davidson, E., Zivehe, F., et al. (2021). Interaction between magnesium and methylglyoxal in diabetic polyneuropathy and neuronal models. Mol. Metab. 43:101114. doi: 10.1016/j.molmet.2020.101114
Su, C.-J., Shen, Z., Cui, R.-X., Huang, Y., Xu, D.-L., Zhao, F.-L., et al. (2020). Thioredoxin-interacting protein (TXNIP) regulates Parkin/PINK1-mediated Mitophagy in dopaminergic neurons under high-glucose conditions: implications for molecular links between Parkinson’s disease and diabetes. Neurosci. Bull. 36, 346–358. doi: 10.1007/s12264-019-00459-5
Sun, H., Saeedi, P., Karuranga, S., Pinkepank, M., Ogurtsova, K., Duncan, B. B., et al. (2022). IDF diabetes atlas: global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pract. 183:109119. doi: 10.1016/j.diabres.2021.109119
Suzuki, R., Lee, K., Jing, E., Biddinger, S. B., McDonald, J. G., Montine, T. J., et al. (2010). Diabetes and insulin in regulation of brain cholesterol metabolism. Cell Metab. 12, 567–579. doi: 10.1016/j.cmet.2010.11.006
Takahashi-Niki, K., Niki, T., Iguchi-Ariga, S. M. M., and Ariga, H. (2017). Transcriptional regulation of DJ-1. Adv. Exp. Med. Biol. 2017, 89–95. doi: 10.1007/978-981-10-6583-5_7
Tanday, N., Lafferty, R. A., Flatt, P. R., and Irwin, N. (2022). Beneficial metabolic effects of recurrent periods of beta-cell rest and stimulation using stable neuropeptide Y1 and glucagon-like peptide-1 receptor agonists. Diabetes. Obes. Metab. 24, 2353–2363. doi: 10.1111/dom.14821
Tarry-Adkins, J. L., Grant, I. D., Ozanne, S. E., Reynolds, R. M., and Aiken, C. E. (2021). Efficacy and side effect profile of different formulations of metformin: a systematic review and Meta-analysis. Diabetes Ther. 12, 1901–1914. doi: 10.1007/s13300-021-01058-2
Tehranian, R., Montoya, S. E., Van Laar, A. D., Hastings, T. G., and Perez, R. G. (2006). Alpha-synuclein inhibits aromatic amino acid decarboxylase activity in dopaminergic cells. J. Neurochem. 99, 1188–1196. doi: 10.1111/j.1471-4159.2006.04146.x
Tian, R., Li, R., Liu, Y., Liu, J., Pan, T., Zhang, R., et al. (2019). Metformin ameliorates endotoxemia-induced endothelial pro-inflammatory responses via AMPK-dependent mediation of HDAC5 and KLF2. Biochim. Biophys. Acta Mol. Basis Dis. 1865, 1701–1712. doi: 10.1016/j.bbadis.2019.04.009
Tsai, W.-C., Bhattacharyya, N., Han, L.-Y., Hanover, J. A., and Rechler, M. M. (2003). Insulin inhibition of transcription stimulated by the Forkhead protein Foxo1 is not solely due to nuclear exclusion. Endocrinology 144, 5615–5622. doi: 10.1210/en.2003-0481
Turner, M. C., Martin, N. R. W., Player, D. J., Ferguson, R. A., Wheeler, P., Green, C. J., et al. (2020). Characterising hyperinsulinemia-induced insulin resistance in human skeletal muscle cells. J. Mol. Endocrinol. 64, 125–132. doi: 10.1530/JME-19-0169
Varoqui, H., and Erickson, J. D. (1996). Active transport of acetylcholine by the human vesicular acetylcholine transporter. J. Biol. Chem. 271, 27229–27232. doi: 10.1074/jbc.271.44.27229
Vicente Miranda, H., Szegő, É. M., Oliveira, L. M. A., Breda, C., Darendelioglu, E., de Oliveira, R. M., et al. (2017). Glycation potentiates α-synuclein-associated neurodegeneration in synucleinopathies. Brain 140, 1399–1419. doi: 10.1093/brain/awx056
Vidal-Martinez, G., Yang, B., Vargas-Medrano, J., and Perez, R. G. (2018). Could α-Synuclein modulation of insulin and dopamine identify a novel link between Parkinson’s disease and diabetes as well as potential therapies? Front. Mol. Neurosci. 11:465. doi: 10.3389/fnmol.2018.00465
Villapol, S. (2018). Roles of peroxisome proliferator-activated receptor gamma on brain and peripheral inflammation. Cell. Mol. Neurobiol. 38, 121–132. doi: 10.1007/s10571-017-0554-5
Vos, M. (2010). Synaptic mitochondria in synaptic transmission and organization of vesicle pools in health and disease. Front. Synaptic Neurosci. 2:139. doi: 10.3389/fnsyn.2010.00139
Wang, W., Wang, Y., Long, J., Wang, J., Haudek, S. B., Overbeek, P., et al. (2012). Mitochondrial fission triggered by hyperglycemia is mediated by ROCK1 activation in podocytes and endothelial cells. Cell Metab. 15, 186–200. doi: 10.1016/j.cmet.2012.01.009
Wang, C., and Youle, R. J. (2009). The role of mitochondria in apoptosis. Annu. Rev. Genet. 43, 95–118. doi: 10.1146/annurev-genet-102108-134850
Warden, A., Truitt, J., Merriman, M., Ponomareva, O., Jameson, K., Ferguson, L. B., et al. (2016). Localization of PPAR isotypes in the adult mouse and human brain. Sci. Rep. 6:27618. doi: 10.1038/srep27618
Wilson, M. A. (2011). The role of cysteine oxidation in DJ-1 function and dysfunction. Antioxid. Redox Signal. 15, 111–122. doi: 10.1089/ars.2010.3481
World Health Organization (2022). Parkinson Disease. Available at: https://www.who.int/news-room/fact-sheets/detail/parkinson-disease (Accessed June 14, 2023).
Xing, H., Liang, C., Wang, C., Xu, X., Hu, Y., and Qiu, B. (2022). Metformin mitigates cholesterol accumulation via the AMPK/SIRT1 pathway to protect osteoarthritis chondrocytes. Biochem. Biophys. Res. Commun. 632, 113–121. doi: 10.1016/j.bbrc.2022.09.074
Xing, B., Liu, M., and Bing, G. (2007). Neuroprotection with pioglitazone against LPS insult on dopaminergic neurons may be associated with its inhibition of NF-κB and JNK activation and suppression of COX-2 activity. J. Neuroimmunol. 192, 89–98. doi: 10.1016/j.jneuroim.2007.09.029
Xu, Q.-G., Li, X.-Q., Kotecha, S. A., Cheng, C., Sun, H. S., and Zochodne, D. W. (2004). Insulin as an in vivo growth factor. Exp. Neurol. 188, 43–51. doi: 10.1016/j.expneurol.2004.03.008
Yadav, S. K., Singla-Pareek, S. L., Ray, M., Reddy, M. K., and Sopory, S. K. (2005). Methylglyoxal levels in plants under salinity stress are dependent on glyoxalase I and glutathione. Biochem. Biophys. Res. Commun. 337, 61–67. doi: 10.1016/j.bbrc.2005.08.263
Yamamoto, H., Kishi, T., Lee, C. E., Choi, B. J., Fang, H., Hollenberg, A. N., et al. (2003). Glucagon-like Peptide-1-responsive catecholamine neurons in the area Postrema link peripheral glucagon-like Peptide-1 with central autonomic control sites. J. Neurosci. 23, 2939–2946. doi: 10.1523/JNEUROSCI.23-07-02939.2003
Yang, J. W., Raizada, M. K., and Fellows, R. E. (1981). Effects of insulin on cultured rat brain cells: stimulation of ornithine decarboxylase activity. J. Neurochem. 36, 1050–1057. doi: 10.1111/j.1471-4159.1981.tb01699.x
Yaribeygi, H., Sathyapalan, T., and Sahebkar, A. (2019). Molecular mechanisms by which GLP-1 RA and DPP-4i induce insulin sensitivity. Life Sci. 234:116776. doi: 10.1016/j.lfs.2019.116776
Yu, P., Wang, W., Guo, W., Cheng, L., Wan, Z., Cheng, Y., et al. (2023). Pioglitazone-enhanced brown fat whitening contributes to weight gain in diet-induced obese mice. Exp. Clin. Endocrinol. Diabetes. doi: 10.1055/a-2178-9113 [Epub ahead of print].
Zelzer, E. (1998). Insulin induces transcription of target genes through the hypoxia-inducible factor HIF-1alpha/ARNT. EMBO J. 17, 5085–5094. doi: 10.1093/emboj/17.17.5085
Zhou, Y., Ma, X.-Y., Han, J.-Y., Yang, M., Lv, C., Shao, Y., et al. (2021). Metformin regulates inflammation and fibrosis in diabetic kidney disease through TNC/TLR4/NF-κB/miR-155-5p inflammatory loop. World J. Diabetes 12, 19–46. doi: 10.4239/wjd.v12.i1.19
Zhou, S., Sun, Y., Xing, Y., Wang, Z., Wan, S., Yao, X., et al. (2022). Exenatide ameliorates hydrogen peroxide-induced pancreatic β-cell apoptosis through regulation of METTL3-mediated m6A methylation. Eur. J. Pharmacol. 924:174960. doi: 10.1016/j.ejphar.2022.174960
Keywords: type 2 diabetes mellitus, Parkinson’s disease, alpha-synuclein, islet amyloid peptide protein, insulin-resistance, autophagy, hyperglycemia
Citation: Greco M, Munir A, Musarò D, Coppola C and Maffia M (2023) Restoring autophagic function: a case for type 2 diabetes mellitus drug repurposing in Parkinson’s disease. Front. Neurosci. 17:1244022. doi: 10.3389/fnins.2023.1244022
Edited by:
Giovanna Valenti, University of Bari Aldo Moro, ItalyReviewed by:
Valerio Magnaghi, University of Milan, ItalyCopyright © 2023 Greco, Munir, Musarò, Coppola and Maffia. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michele Maffia, bWljaGVsZS5tYWZmaWFAdW5pc2FsZW50by5pdA==
†These authors have contributed equally to this work and share first authorship