 Mengyao Song
Mengyao Song Mingxia Zhang
Mingxia Zhang Sixuan He
Sixuan He Le Li
Le Li Huijing Hu
Huijing Hu
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
PERSPECTIVE article
Front. Neurosci., 31 July 2023
Sec. Translational Neuroscience
Volume 17 - 2023 | https://doi.org/10.3389/fnins.2023.1232308
This article is part of the Research TopicNew Insights into Brain Imaging Methods for Rehabilitation of Brain DiseasesView all 19 articles
Ultrasound neuromodulation technology is a promising neuromodulation approach, with the advantages of noninvasiveness, high-resolution, deep penetration and good targeting, which aid in circumventing the side effects of drugs and invasive therapeutic interventions. Ultrasound can cause mechanical effects, activate mechanosensitive ion channels and alter neuronal excitability, producing biological effects. The structural determination of mechanosensitive ion channels will greatly contribute to our understanding of the molecular mechanisms underlying mechanosensory transduction. However, the underlying biological mechanism of ultrasonic neuromodulation remains poorly understood. Hence, this review aims to provide an outline of the properties of ultrasound, the structures of specific mechanosensitive ion channels, and their role in ultrasound neuromodulation.
Ultrasound refers to sound waves with frequencies above 20,000 Hertz (Hz), which exceeds the human hearing range of 20 Hz–20 kHz. With its high frequency and short wavelength, ultrasound has linear propagation capabilities within a finite distance, high beam focusing and excellent directionality features (Dell'Italia et al., 2022). Moreover, ultrasound is a noninvasive technique for neural regulation. It has high safety and can be employed in combination with magnetic resonance imaging. Exosomes produced by human astrocyte (HA) cells stimulated by ultrasound were comparable in terms of size distribution and morphology with those of untreated HA cells (Deng et al., 2021). Moreover, HE staining conducted after 4 weeks of ultrasound treatment showed no major organ damage, which further affirmed the safety of ultrasound therapy in human. Neuroregulatory technology, such as transcranial ultrasound stimulation, transcranial magnetic stimulation, transcranial electrical stimulation, deep brain stimulation, and optogenetics, is a kind of therapeutic approach that implements either implantable or nonimplantable devices to alleviate patients’ symptoms. A comparison of these different neural regulation modes is shown in Table 1 below. It can effectively modulate the activity of the central, peripheral or autonomic nervous systems by either physical (e.g., light, sound, electricity, magnetism, etc.) or chemical (e.g., drugs) means (Jiang et al., 2019). Currently, ultrasound is frequently involved in treatment procedures as a physical factor. High-intensity ultrasound is typically recommended for tumor ablation, additionally, it has been applied to treat several neurological diseases including Parkinson’s disease, essential tremor and obsessive–compulsive disorder (Sperling et al., 2018; Germann et al., 2021; Iorio-Morin et al., 2021). Low-intensity ultrasound is often used for neural regulation.
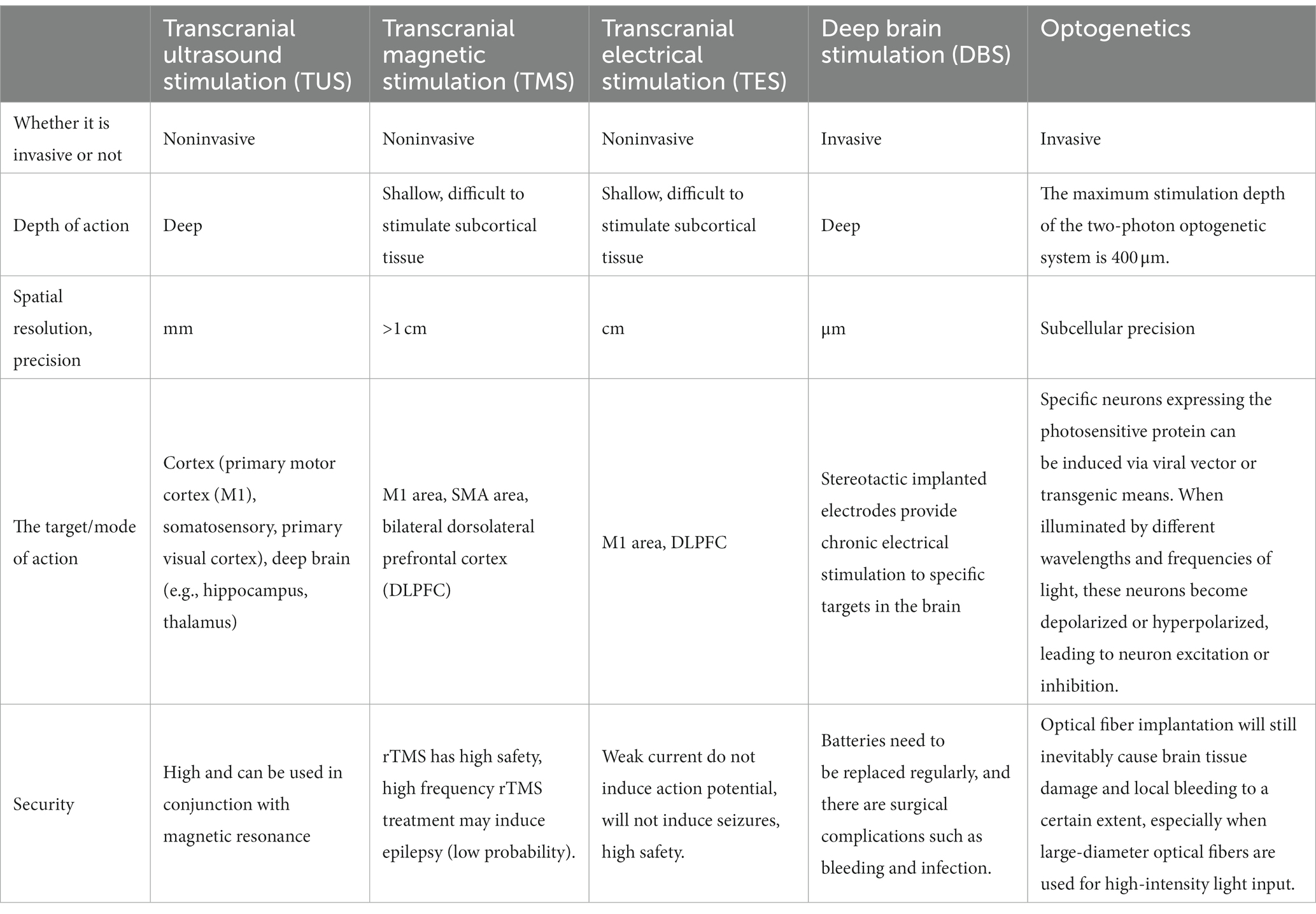
Table 1. Comparison of several neural regulation modes.
The effects of ultrasound mainly result from mechanical, cavitation and thermal effects (Darmani et al., 2022). Mechanical effects stem from ultrasound functioning as a mechanical wave, exerting radiation forces on biological tissues. Acoustic radiation force (ARF) can cause mechanical vibration and cell membrane deformation, and activate mechanosensitive ion channels in neurons, and discharge cells (Peng et al., 2020). On the other hand, thermal effects refer to biological tissue cells absorbing the energy of ultrasound, causing a subsequent temperature increase and mediating cell excitability. However, too high a temperature denatures enzymes and proteins, leading to decreased biological activity. Moreover, cavitation effects arise from the formation of small gas bubbles in tissue under positive and negative pressure phases of ultrasound oscillation stretching (Baek et al., 2017; Blackmore et al., 2019; Feng et al., 2019). Currently, it is commonly surmised that the biological impacts of ultrasound are primarily attributed to its mechanical influence rather than its thermal attributes.
Mechanosensitive (MS) ion channels refer to a group of transmembrane channel proteins that can convert mechanical stimuli signals into electrical or chemical signals. Mechanically-sensitive channels exist widely in bacterial, archaea, and eukaryotic organisms (Martinac, 2004). In bacteria and archaea, mechanosensitive channels serve as protection and survival mechanisms. Their primary function is to release intracellular substances as an emergency valve to lower osmotic pressure when the extracellular environment becomes hypotonic such as during heavy rain (Ajouz et al., 1998). The discovery of new families of mechanically activated ion channels, such as PIEZO, which have important in vivo physiological roles in mammals, opens new avenues for studying the role of mechanotransduction in human health and disease (Coste et al., 2010; Murthy et al., 2017; Kefauver et al., 2020). In 2016, Kubanek reported that ultrasound triggered a current via the heterologous expression of two pore domain potassium channels (K2P channels) including TREK-1, TREK-2, TRAAK and voltage-gated sodium channels (Nav1.5) in the Xenopus oocyte system (Kubanek et al., 2016). Heterologous expression of TRP-4 channels in Caenorhabditis elegans neurons revealed that their motion behavior was provoked by ultrasound. These discoveries suggest that the mechanical force generated by ultrasound could open mechanosensitive ion channels in the cell membrane and induce ion flow, which can alter neuronal excitability and eventually result in biological effects.
Despite substantial recent progress in the identification and characterization of mechanically activated ion channels, a variety of biological processes that depend on mechanotransduction remain poorly understood at the molecular level, and the identities of many mechanosensors remain elusive. Here, this article will first review the structures of several mechanosensitive ion channels and then outline the progress in their research related to ultrasound to help bolster our understanding of mechanotransduction at the molecular level.
Sukharev et al. (1994) discovered two types of mechanosensitive ion channels in E. coli, which were named based on their pore size: the mechanosensitive channel of large conductance (MscL) and the mechanosensitive channel of small conductance (MscS). Perozo et al. (2002) demonstrated that the open state of MscL is highly dynamic, supporting a water-filled pore of at least 25 Å, lined mostly by the first transmembrane helix (TM1), allowing passage of large organic ions and small proteins. The patch-clamp electrical recording technique showed that when the patches were subjected to suction or solution bathing the patch was diluted, and voltage-controlled (−clamped) patches of E. coli membranes produced giant steps in unitary current (Martinac et al., 1987; Sukharev et al., 1993; Levina et al., 1999). MscS showed a single channel conductance of ~1 nS and demonstrated both pressure and voltage dependence, and selectivity for anions. While MscL with ~3 nS conductance when subjected to even stronger suction.
MscL is widely expressed in prokaryotic cells but is not present in eukaryotes. MscL has been extensively studied by scientists in recent years. Chang and colleagues indicated that the gating of MscL is primarily regulated by lipoprotein interactions (Chang et al., 1998). The composition of MscL comprises five identical subunits forming an ion channel, which is in turn regulated by membrane tension. The three-dimensional structure of the Mycobacterium tuberculosis MscL homolog was determined through X-ray crystallography, to a resolution of 3.5 angstroms. Each subunit consists of 136 amino acid residues, alongside two transmembrane α-helical regions (TM1 and TM2), and a loop region that links them on the extracellular side. The termini (N-terminal and C-terminal) are found on the cytoplasmic surface. The region responsible for pore formation by MscL results from TM1 of the five constituent subunits, and TM2 primarily associates with the cytoplasmic membrane lipid bilayer. MscL is a mechanically gated ion channel, featuring considerable conductance and no ion selectivity; physiologically, the protein structure resembles a cylinder. Additionally, MscL can be expressed stably in the cell membranes of eukaryotic cells.
The MscL channel is expressed in prokaryotes and its function in these cells has been extensively researched. However, the application and study of MscL in eukaryotes is just emerging. MscL channels can be expressed in eukaryotic cell lines including CHO and HEK293 cells (Doerner et al., 2012). Through electrophysiological experiments and dye release experiments, the authors demonstrated the mechanosensitive function of the MscL channels. Ye et al. (2018) also confirmed the channel activity of MscL in HEK293T cells and neurons through electrophysiological experiments. However, MscL channels expressed in eukaryotic systems have a lower opening threshold and smaller conductance than those in prokaryotes. Therefore, the authors expressed the functional mutant I92L MscL in neurons. Fluorescent localization and electrophysiological experiments proved that MscL I92L can be expressed and inserted into the neuronal membrane and has channel activity. Furthermore, MscL I92L demonstrated greater sensitivity to mechanical stimulation than the wild-type MscL channel. This is exemplified by only 30 mmHg of force leading to significant channel opening.
Piezo ion channels refer to a group of channels discovered and named by Coste et al. (2010). Piezo1 (Fam38A) was identified as the ion channel essential for generating mechanosensitive potentials in the Neuro2A cell line through expression profile and RNA interference knockdown of candidate genes techniques. Only two types of Piezo families have been discovered so far, Piezo1 (Fam38A) and Piezo2 (Fam38B). Piezo1 is a nonselective cationic channel that can be inhibited by GsMTx4 (tarantula venom), gadolinium (Gd), and ruthenium red (RR) (Copp et al., 2016). Piezo2, a mammalian cognate of Piezo1, records mechanically sensitive electrical currents in separated dorsal root ganglion (DRG)neurons. Piezo1 channels mainly exist in nonsensory tissues, including the skin, lungs, kidneys, and bladder. On the other hand, Piezo2 channels occur mainly in sensory tissues such as the trigeminal ganglion (TG), DRG sensory neurons, and Merkel cells.
Each subunit of Piezo proteins is made up of over 2000 amino acid residues, and their molecules are relatively large, having a molecular mass of 1.2 × 10^6. Ge et al. determined that the full-length of the cryo-electron microscopy structure is 2,547 amino acids, with a resolution of 4.8 Å in mouse Piezo1 (Ge et al., 2015). The findings reveal that Piezo1 consists of a triple helix structure, with extracellular domains comprising three distal blades and a central cap. There are 14 apparently resolved segments per subunit in the transmembrane region that form three peripheral wings and a central pore module that encloses a potential ion-conducting pore. The carboxyl terminal is responsible for the pore section of the ion channel, while the amino terminal receives mechanical stimulation that opens the carboxyl terminal pore.
Piezo1 activation plays an essential role in mechanical transmission via ultrasonic stimulation. Qiu et al. (2019) conducted a study that revealed that the activation of heterologously expressed HEK293T cells and endogenous Piezo1 channels can be achieved through low-intensity and low-frequency ultrasound stimulation. This resulted in Ca2+ influx as well as increased nuclear c-Fos expression levels in primary neurons, although when pre-treated with a Piezo1 inhibitor the effect was inhibited in cells. Furthermore, the study demonstrated that ultrasonic stimulation significantly affected downstream Ca2+ signaling protein levels and induced the expression of important proteins such as phospho-CaMKII, phospho-CREB, and c-Fos in a neuronal cell line. These proteins are known to play significant roles in complex neuronal functions like learning, memory, and neuronal plasticity. Notably, the impact of ultrasonic stimulation on Ca2+ signaling protein levels was found to decrease with the loss of Piezo1 channel functions. In 2023, the authors also reported that Piezo1 knockout (P1KO) in the right motor cortex of mice significantly decreased ultrasound-induced neuronal calcium responses, limb movement, muscle electromyogram (EMG) signaling and C-Fos expression compared to the control. Central amygdala (CEA) neurons, having higher Piezo1 expression levels, displayed greater sensitivity to ultrasound than cortical neurons. Piezo1 is expressed in both neurons and astrocytes. The authors demonstrated that Piezo1 is expressed in different brain regions and that neuronal Piezo1 plays an important role in mediating ultrasound effects directly (Shen et al., 2021). The researchers also developed a Piezo1-targeted microbubble (PTMB) which can bind to the extracellular domains of the Piezo1 channel (Zhu et al., 2023).
The discovery of the transient receptor potential (TRP) channel initially occurred within the visual system of Drosophila melanogaster, and was based upon the peculiar behavior of mutant Drosophila in response to sustained light exposure. This resulted in the production of transient potentials as opposed to sustained peak potentials. TRP channels have been found to be highly conserved genes across a broad spectrum of species, ranging from Caenorhabditis elegans to humans. They exhibit prominent distribution within sensory neurons and play crucial roles in the modulation of external mechanical stimuli, including pressure and sound waves. Additionally, these channels are implicated in the generation of senses associated with touch, pain, hearing, taste, and vision (Voolstra et al., 2010).
TRP channels are nonselective cationic channels that are highly permeable to Ca2+ and Na+, with TRPM6 and TRPM7 being highly permeable to Mg2+. Nilius and Owsianik (2011) classified TRP ion channels into seven subtypes and two categories based on amino acid sequences and three-dimensional structures. The first category includes TRPC (TRP-canonical), TRPV (TRP-vanilloid), TRPM (TRP-melastatin), TRPA (TRP-ankyrin) and TRPN (TRPNompC), whereas the second category includes TRPP (TRP-polycystin) and TRPML (TRP-mucolipin). The TRP channel family consists of nonselective cationic channels that are made up of four tetramer monomers. These monomers contain a hexaxial transmembrane (TM) domain with a pore ring structure located between TM5 and TM6.
TRP-4 channels are primarily expressed in four CEPs including CEPDL, CEPDR, CEPVL and CEPVR dopaminergic neurons and in two ADEs consisting of ADEL and ADER dopaminergic neurons, and DVA and DVC interneurons in a few C. elegans neurons. Ibsen et al. (2015) conducted a study in which they observed the effect of defective TRP-4 mutation in nematodes with respect to their response to ultrasound-combined microbubbles. The mutant nematodes exhibited a reduction in large reversal responses compared to the wild type. Subsequently, researchers transferred the TRP-4 gene to amphid wing ‘C’ (AWC) neurons leading to its induction. This activation of TRP-4 gene expression in AWC neurons was found to result in the accumulation of calcium ions under specific peak negative pressures (0.41 and 0.47 MPa) of ultrasonic stimulation. This response was not observed in the wild-type AWC cells. Additionally, the decrease in contrarian motor behavior in C. elegans mutants with TRP-4 indicated that the induction of TRP-4 by ultrasonic stimulation (peak negative pressure < 0.5 MPa) might regulate the activity of neurons involved in reverse motor behavior, thereby leading to a reduction in reverse movement.
The identification of the K2P channel first occurred in the human kidney, as its characteristic two pore regions prompted its labeling as a two-pore-domain potassium (K2P) channel. This particular form of channel is capable of activation along the entire range of physiological voltages, elucidating both the background potassium current and the baseline potassium current, and is not vulnerable to typical potassium channel blockers (Lesage et al., 1996).
Potassium channels are a diverse group of proteins that play a crucial role in regulating cellular activity and maintaining cellular homeostasis. These channels can be classified into three main types based on their structure and function: calcium-activated (KCa) potassium channels, two-pore-domain potassium channels (K2P) and inward-rectified potassium channels (Kir). Among these, K2P channels have gained particular attention in recent years due to their unique structure and function. Currently, K2P channels are categorized into six subgroups or “clades” each with distinct structural and functional characteristics. These six clades are: TWIK, TASK, TREK, TALK, THIK, and TRESK. Notably, only the TREK subgroup of K2P channels is known to be mechanically sensitive. Thus, understanding the distinct properties of each K2P channel subtype is critical for gaining insight into their role in physiological and pathological processes (Feliciangeli et al., 2015). Kv, KCa, and Kir are tetrameric channels, with each monomer having a single pore domain. However, K2P is a dimeric channel, and each monomer is composed of two pore regions. These channels consist of two pore domains (P1 and P2), two extracellular cap helices (C1 and C2), and four transmembrane domains (M1–M4) with both the amino and carboxyl terminus situated on the cytoplasmic side (Renigunta et al., 2015). K2P channels which are equipped with the capacity to perceive mechanical stimulation at the cellular membrane are classified as mechanically sensitive channels, namely TREK-1, TREK-2, and TRAAK. The high-resolution crystal structures of TWIK-1, TRAAK, and TREK-2 channels have been made available. These structures divulge the existence of multiple helices within the extracellular ring between TM1 and the P1, resulting in the formation of a physical obstruction that urges ions to exit via the side pore.
Sorum et al. (2021) proposed that ultrasound has the potential to trigger the opening of mechanosensitive TRAAK channels through an increase in membrane tension. This study provides insight into the vital role of mechanosensitive channels in physiological responses to ultrasound and presents a promising avenue for gene targeting in the regulation of cellular auditory nerves. The authors observed that the application of short-pulsed, low-intensity ultrasound (10 ms, 5 MHz, 1.2 W/cm2) led to a swift and robust activation of TRAAK channels in plaques of Xenopus oocytes expressing TRAAK as well as in cortical neurons of mice also expressing TRAAK. The K+ selective ultrasonic stimulation current featured a reversal potential that was proximal to the Nernst equilibrium potential for K+ (EK+ = −59 mV), comparable to the TRAAK currents for both base and pressure stimulation. In contrast, the study revealed that the non-mechanosensitive K2P ion channel TASK2 was not activated by ultrasound.
In addition to its role in neuromodulation, ultrasound also has various applications, such as reducing fracture healing time (especially in delayed healing and bone nonunion), preventing inflammatory loosening of prosthetics, and promoting tendon, ligament, and cartilage recovery. Furthermore, it can inhibit lipopolysaccharide-induced inflammation and reduce proinflammatory factors (Chan et al., 2010; Jeremias Júnior et al., 2011; Loyola-Sánchez et al., 2012; Ren et al., 2013; Zhao X et al., 2017; Jiang et al., 2019). Table 2 summarizes the research progress on the above mechanically sensitive ion channels related to ultrasonic neural regulation.
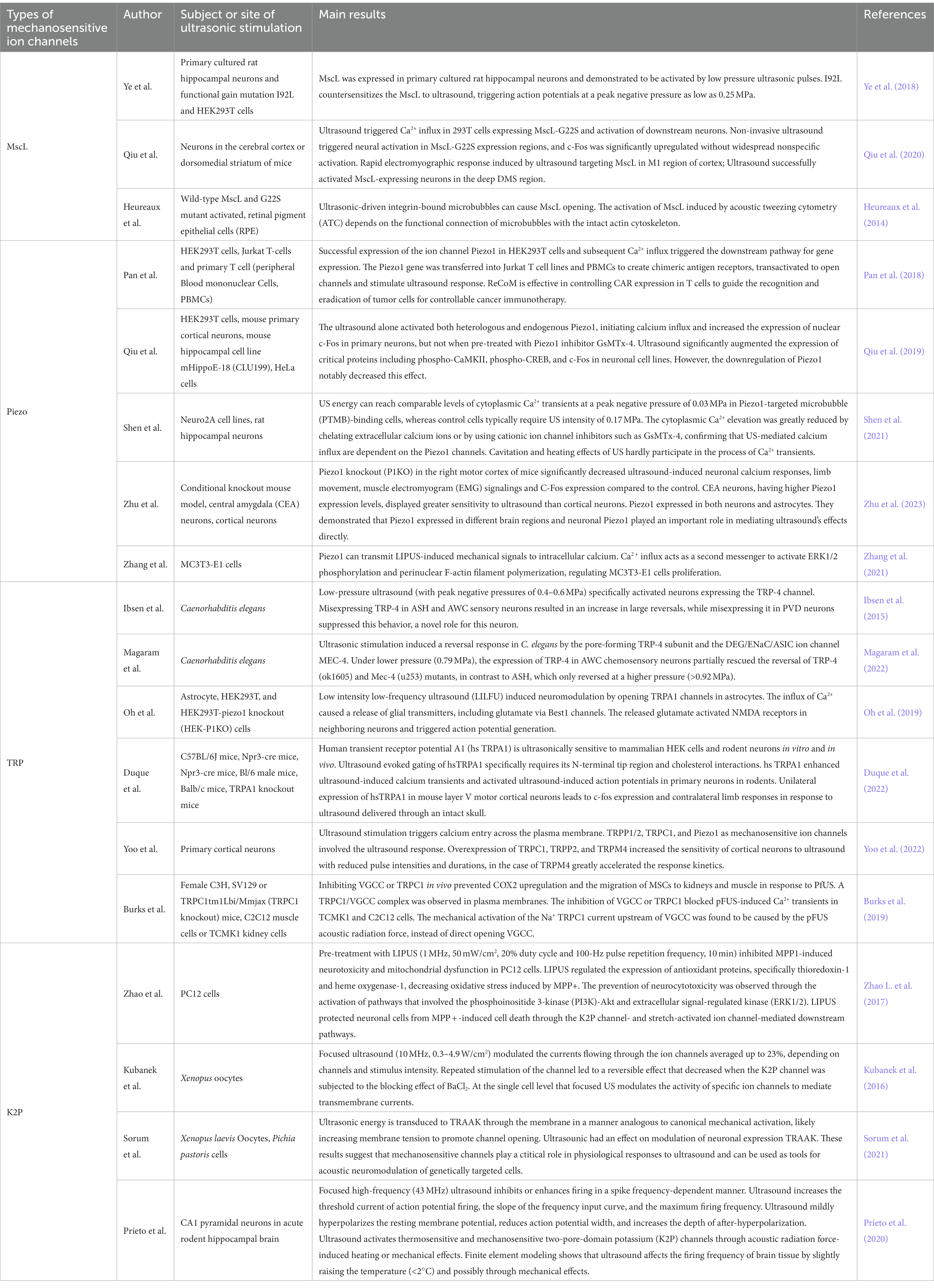
Table 2. Studies on several mechanosensitive ion channels in ultrasonic neuromodulation.
Mechanosensitive ion channels are mechanical force molecular sensors that are activated by mechanical stimuli and located on the cell membrane. These channels can quickly and efficiently convert mechanical stimuli into electrical and chemical signals. Stretching of mechanosensitive proteins can damage molecular binding sites, expose regulatory sites, and change the association or dissociation rates for protein binding (Ingham et al., 1997; Krammer et al., 2002; Cain et al., 2021). MS channel opening can result in ion influx. Additionally, numerous diseases such as muscular dystrophy, and cardiac arrhythmias and et al. have been related to defects in activating MS ion channels (Sukharev and Sachs, 2012). However, the gating mechanism and physiological effects of these channels differ. We also do not know the connection between the structure and the function of these MS ion channels. Ultrasound usually activates more than one MS ion channel, but it is unknown which one is working or how they work together. Our understanding of the mechanisms and functions of ultrasound activated mechanosensitive ion channels remains limited.
We need to develop innovative research methods and conduct more thorough research on the mechanism of these channels and explore screening of mechanically sensitive ion channels or their mutants that can be accurately controlled in the future. Addtionally, these MS ion channels should be expressed in various nerve cells. In future studies, researchers should clarify the expression profile of each ion channel in neuronal cells, and then study their sensitivity to ultrasound modulation, so as to summarize and compare the differences in the role of various ion channels in ultrasound modulation.
To date, previous studies have demonstrated the structure of some specific MS ion channels and their investigated their roles in ultrasonic neuromodulation. Ultrasound offers several advantages, including non-invasiveness, convenient in vitro regulation, intracranial multipoint focusing, spatiotemporal controllability and accuracy. Basic clinical trials have demonstrated that ultrasound can improve specific behaviors, such as increased responsiveness in patients with chronic disorders of consciousness (Cain et al., 2021) and improved mood (Reznik et al., 2020; Sanguinetti et al., 2020). However, further studies are needed to investigate how ultrasound acts on organisms, which mechanically sensitive channels it influences, and how different ultrasonic stimulation parameters produce varying effects. To gain a better understanding of ultrasonic neuromodulation efficacy, it is necessary to conduct these studies in various species and different disease models, and set different ultrasonic stimulation parameters to observe the applicability, persistence and timeliness of ultrasonic stimulation. It is also crucial to address the thermal and cavitation effects of ultrasonic stimulation and improve ultrasonic focusing resolution to increase the precision of ultrasonic regulation. Finally, innovative ultrasonic equipment should be developed to enhance the efficacy and applicability of wearable ultrasonic equipment.
HH: conceptualization. MS and MZ: writing—original draft preparation. MS, MZ, SH, LL, and HH: writing—review and editing. All authors contributed to the article and approved the submitted version.
This work was supported by the Natural Science Foundation of Shaanxi province (2022-JM482), Shenzhen Science and Technology Program (GJHZ20210705143401005), the Education and Teaching Reform Funds for the Central Universities (No. 23GZ230102), and the National Natural Science Foundation of China (No. 31771016).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Ajouz, B., Berrier, C., Garrigues, A., Besnard, M., and Ghazi, A. (1998). Release of thioredoxin via the mechanosensitive channel MscL during osmotic downshock of Escherichia coli cells. J. Biol. Chem. 273, 26670–26674. doi: 10.1074/jbc.273.41.26670
Baek, H., Pahk, K. J., and Kim, H. (2017). A review of low-intensity focused ultrasound for neuromodulation. Biomed. Eng. Lett. 7, 135–142. doi: 10.1007/s13534-016-0007-y
Blackmore, J., Shrivastava, S., Sallet, J., Butler, C. R., and Cleveland, R. O. (2019). Ultrasound neuromodulation: a review of results, mechanisms and safety. Ultrasound. Med. Biol. 45, 1509–1536. doi: 10.1016/j.ultrasmedbio.2018.12.015
Burks, S. R., Lorsung, R. M., Nagle, M. E., Tu, T. W., and Frank, J. A. (2019). Focused ultrasound activates voltage-gated calcium channels through depolarizing TRPC1 sodium currents in kidney and skeletal muscle. Theranostics 9, 5517–5531. doi: 10.7150/thno.33876
Cain, J. A., Spivak, N. M., Coetzee, J. P., Crone, J. S., Johnson, M. A., Lutkenhoff, E. S., et al. (2021). Ultrasonic thalamic stimulation in chronic disorders of consciousness. Brain Stimul. 14, 301–303. doi: 10.1016/j.brs.2021.01.008
Chan, Y. S., Hsu, K. Y., Kuo, C. H., Lee, S. D., Chen, S. C., Chen, W. J., et al. (2010). Using low-intensity pulsed ultrasound to improve muscle healing after laceration injury: an in vitro and in vivo study. Ultrasound Med. Biol. 36, 743–751. doi: 10.1016/j.ultrasmedbio.2010.02.010
Chang, G., Spencer, R. H., Lee, A. T., Barclay, M. T., and Rees, D. C. (1998). Structure of the MscL homolog from Mycobacterium tuberculosis: a gated mechanosensitive ion channel. Science 282, 2220–2226. doi: 10.1126/science.282.5397.2220
Copp, S. W., Kim, J. S., Ruiz-Velasco, V., and Kaufman, M. P. (2016). The mechano-gated channel inhibitor GsMTx4 reduces the exercise pressor reflex in decerebrate rats. J. Physiol. 594, 641–655. doi: 10.1113/jp271714
Coste, B., Mathur, J., Schmidt, M., Earley, T. J., Ranade, S., Petrus, M. J., et al. (2010). Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 330, 55–60. doi: 10.1126/science.1193270
Darmani, G., Bergmann, T. O., Butts Pauly, K., Caskey, C. F., de Lecea, L., Fomenko, A., et al. (2022). Non-invasive transcranial ultrasound stimulation for neuromodulation. Clin. Neurophysiol. 135, 51–73. doi: 10.1016/j.clinph.2021.12.010
Dell'Italia, J., Sanguinetti, J. L., Monti, M. M., Bystritsky, A., and Reggente, N. (2022). Current state of potential mechanisms supporting low intensity focused ultrasound for neuromodulation. Front. Hum. Neurosci. 16:872639. doi: 10.3389/fnhum.2022.872639
Deng, Z., Wang, J., Xiao, Y., Li, F., Niu, L., Liu, X., et al. (2021). Ultrasound-mediated augmented exosome release from astrocytes alleviates amyloid-β-induced neurotoxicity. Theranostics 11, 4351–4362. doi: 10.7150/thno.52436
Doerner, J. F., Febvay, S., and Clapham, D. E. (2012). Controlled delivery of bioactive molecules into live cells using the bacterial mechanosensitive channel MscL. Nat. Commun. 3:990. doi: 10.1038/ncomms1999
Duque, M., Lee-Kubli, C. A., Tufail, Y., Magaram, U., Patel, J., Chakraborty, A., et al. (2022). Sonogenetic control of mammalian cells using exogenous transient receptor potential A1 channels. Nat. Commun. 13:600. doi: 10.1038/s41467-022-28205-y
Feliciangeli, S., Chatelain, F. C., Bichet, D., and Lesage, F. (2015). The family of K2P channels: salient structural and functional properties. J. Physiol. 593, 2587–2603. doi: 10.1113/jphysiol.2014.287268
Feng, B., Chen, L., and Ilham, S. J. (2019). A review on ultrasonic neuromodulation of the peripheral nervous system: enhanced or suppressed activities? Appl. Sci. 9:1637. doi: 10.3390/app9081637
Ge, J., Li, W., Zhao, Q., Li, N., Chen, M., Zhi, P., et al. (2015). Architecture of the mammalian mechanosensitive Piezo1 channel. Nature 527, 64–69. doi: 10.1038/nature15247
Germann, J., Elias, G. J. B., Neudorfer, C., Boutet, A., Chow, C. T., Wong, E. H. Y., et al. (2021). Potential optimization of focused ultrasound capsulotomy for obsessive compulsive disorder. Brain 144, 3529–3540. doi: 10.1093/brain/awab232
Heureaux, J., Chen, D., Murray, V. L., Deng, C. X., and Liu, A. P. (2014). Activation of a bacterial mechanosensitive channel in mammalian cells by cytoskeletal stress. Cell. Mol. Bioeng. 7, 307–319. doi: 10.1007/s12195-014-0337-8
Ibsen, S., Tong, A., Schutt, C., Esener, S., and Chalasani, S. H. (2015). Sonogenetics is a non-invasive approach to activating neurons in Caenorhabditis elegans. Nat. Commun. 6:8264. doi: 10.1038/ncomms9264
Ingham, K. C., Brew, S. A., Huff, S., and Litvinovich, S. V. (1997). Cryptic self-association sites in type III modules of fibronectin. J. Biol. Chem. 272, 1718–1724. doi: 10.1074/jbc.272.3.1718
Iorio-Morin, C., Yamamoto, K., Sarica, C., Zemmar, A., Levesque, M., Brisebois, S., et al. (2021). Bilateral focused ultrasound thalamotomy for essential tremor (BEST-FUS phase 2 trial). Mov. Disord. 36, 2653–2662. doi: 10.1002/mds.28716
Jeremias Júnior, S. L., Camanho, G. L., Bassit, A. C., Forgas, A., Ingham, S. J., and Abdalla, R. J. (2011). Low-intensity pulsed ultrasound accelerates healing in rat calcaneus tendon injuries. J. Orthop. Sports Phys. Ther. 41, 526–531. doi: 10.2519/jospt.2011.3468
Jiang, X., Savchenko, O., Li, Y., Qi, S., Yang, T., Zhang, W., et al. (2019). A review of low-intensity pulsed ultrasound for therapeutic applications. I.E.E.E. Trans. Biomed. Eng. 66, 2704–2718. doi: 10.1109/tbme.2018.2889669
Kefauver, J. M., Ward, A. B., and Patapoutian, A. (2020). Discoveries in structure and physiology of mechanically activated ion channels. Nature 587, 567–576. doi: 10.1038/s41586-020-2933-1
Krammer, A., Craig, D., Thomas, W. E., Schulten, K., and Vogel, V. (2002). A structural model for force regulated integrin binding to fibronectin's RGD-synergy site. Matrix Biol. 21, 139–147. doi: 10.1016/s0945-053x(01)00197-4
Kubanek, J., Shi, J., Marsh, J., Chen, D., Deng, C., and Cui, J. (2016). Ultrasound modulates ion channel currents. Sci. Rep. 6:24170. doi: 10.1038/srep24170
Lesage, F., Guillemare, E., Fink, M., Duprat, F., Lazdunski, M., Romey, G., et al. (1996). TWIK-1, a ubiquitous human weakly inward rectifying K+ channel with a novel structure. EMBO J. 15, 1004–1011. doi: 10.1002/j.1460-2075.1996.tb00437.x
Levina, N., Tötemeyer, S., Stokes, N. R., Louis, P., Jones, M. A., and Booth, I. R. (1999). Protection of Escherichia coli cells against extreme turgor by activation of MscS and MscL mechanosensitive channels: identification of genes required for MscS activity. EMBO J. 18, 1730–1737. doi: 10.1093/emboj/18.7.1730
Loyola-Sánchez, A., Richardson, J., Beattie, K. A., Otero-Fuentes, C., Adachi, J. D., and MacIntyre, N. J. (2012). Effect of low-intensity pulsed ultrasound on the cartilage repair in people with mild to moderate knee osteoarthritis: a double-blinded, randomized, placebo-controlled pilot study. Arch. Phys. Med. Rehabil. 93, 35–42. doi: 10.1016/j.apmr.2011.07.196
Magaram, U., Weiss, C., Vasan, A., Reddy, K. C., Friend, J., and Chalasani, S. H. (2022). Two pathways are required for ultrasound-evoked behavioral changes in Caenorhabditis elegans. PLoS One 17:e0267698. doi: 10.1371/journal.pone.0267698
Martinac, B. (2004). Mechanosensitive ion channels: molecules of mechanotransduction. J. Cell Sci. 117, 2449–2460. doi: 10.1242/jcs.01232
Martinac, B., Buechner, M., Delcour, A. H., Adler, J., and Kung, C. (1987). Pressure-sensitive ion channel in Escherichia coli. Proc. Natl. Acad. Sci. U. S. A. 84, 2297–2301. doi: 10.1073/pnas.84.8.2297
Murthy, S. E., Dubin, A. E., and Patapoutian, A. (2017). Piezos thrive under pressure: mechanically activated ion channels in health and disease. Nat. Rev. Mol. Cell Biol. 18, 771–783. doi: 10.1038/nrm.2017.92
Nilius, B., and Owsianik, G. (2011). The transient receptor potential family of ion channels. Genome Biol. 12:218. doi: 10.1186/gb-2011-12-3-218
Oh, S. J., Lee, J. M., Kim, H. B., Lee, J., Han, S., Bae, J. Y., et al. (2019). Ultrasonic neuromodulation via astrocytic TRPA1. Curr. Biol. 29, 3386–3401.e8. doi: 10.1016/j.cub.2019.08.021
Pan, Y., Yoon, S., Sun, J., Huang, Z., Lee, C., Allen, M., et al. (2018). Mechanogenetics for the remote and noninvasive control of cancer immunotherapy. Proc. Natl. Acad. Sci. U. S. A. 115, 992–997. doi: 10.1073/pnas.1714900115
Peng, X., He, W., Xin, F., Genin, G. M., and Lu, T. J. (2020). The acoustic radiation force of a focused ultrasound beam on a suspended eukaryotic cell. Ultrasonics 108:106205. doi: 10.1016/j.ultras.2020.106205
Perozo, E., Cortes, D. M., Sompornpisut, P., Kloda, A., and Martinac, B. (2002). Open channel structure of MscL and the gating mechanism of mechanosensitive channels. Nature 418, 942–948. doi: 10.1038/nature00992
Prieto, M. L., Firouzi, K., Khuri-Yakub, B. T., Madison, D. V., and Maduke, M. (2020). Spike frequency-dependent inhibition and excitation of neural activity by high-frequency ultrasound. J. Gen. Physiol. 152:e202012672. doi: 10.1085/jgp.202012672
Qiu, Z., Guo, J., Kala, S., Zhu, J., Xian, Q., Qiu, W., et al. (2019). The mechanosensitive ion channel Piezo1 significantly mediates in vitro ultrasonic stimulation of neurons. iScience 21, 448–457. doi: 10.1016/j.isci.2019.10.037
Qiu, Z., Kala, S., Guo, J., Xian, Q., Zhu, J., Zhu, T., et al. (2020). Targeted neurostimulation in mouse brains with non-invasive ultrasound. Cell Rep. 32:108033. doi: 10.1016/j.celrep.2020.108033
Ren, L., Yang, Z., Song, J., Wang, Z., Deng, F., and Li, W. (2013). Involvement of p38 MAPK pathway in low intensity pulsed ultrasound induced osteogenic differentiation of human periodontal ligament cells. Ultrasonics 53, 686–690. doi: 10.1016/j.ultras.2012.10.008
Renigunta, V., Schlichthörl, G., and Daut, J. (2015). Much more than a leak: structure and function of K₂p-channels. Pflugers Arch. 467, 867–894. doi: 10.1007/s00424-015-1703-7
Reznik, S. J., Sanguinetti, J. L., Tyler, W. J., Daft, C., and Allen, J. J. B. (2020). A double-blind pilot study of transcranial ultrasound (TUS) as a five-day intervention: TUS mitigates worry among depressed participants. Neurol. Psychiatry Brain Res. 37, 60–66. doi: 10.1016/j.npbr.2020.06.004
Sanguinetti, J. L., Hameroff, S., Smith, E. E., Sato, T., Daft, C. M. W., Tyler, W. J., et al. (2020). Transcranial focused ultrasound to the right prefrontal cortex improves mood and alters functional connectivity in humans. Front. Hum. Neurosci. 14:52. doi: 10.3389/fnhum.2020.00052
Shen, X., Song, Z., Xu, E., Zhou, J., and Yan, F. (2021). Sensitization of nerve cells to ultrasound stimulation through Piezo1-targeted microbubbles. Ultrason. Sonochem. 73:105494. doi: 10.1016/j.ultsonch.2021.105494
Sorum, B., Rietmeijer, R. A., Gopakumar, K., Adesnik, H., and Brohawn, S. G. (2021). Ultrasound activates mechanosensitive TRAAK K(+) channels through the lipid membrane. Proc. Natl. Acad. Sci. U. S. A. 118:e2006980118. doi: 10.1073/pnas.2006980118
Sperling, S. A., Shah, B. B., Barrett, M. J., Bond, A. E., Huss, D. S., Gonzalez Mejia, J. A., et al. (2018). Focused ultrasound thalamotomy in Parkinson disease: nonmotor outcomes and quality of life. Neurology 91, e1275–e1284. doi: 10.1212/wnl.0000000000006279
Sukharev, S. I., Blount, P., Martinac, B., Blattner, F. R., and Kung, C. (1994). A large-conductance mechanosensitive channel in E. coli encoded by mscL alone. Nature 368, 265–268. doi: 10.1038/368265a0
Sukharev, S. I., Martinac, B., Arshavsky, V. Y., and Kung, C. (1993). Two types of mechanosensitive channels in the Escherichia coli cell envelope: solubilization and functional reconstitution. Biophys. J. 65, 177–183. doi: 10.1016/s0006-3495(93)81044-0
Sukharev, S., and Sachs, F. (2012). Molecular force transduction by ion channels: diversity and unifying principles. J. Cell Sci. 125, 3075–3083. doi: 10.1242/jcs.092353
Voolstra, O., Beck, K., Oberegelsbacher, C., Pfannstiel, J., and Huber, A. (2010). Light-dependent phosphorylation of the drosophila transient receptor potential ion channel. J. Biol. Chem. 285, 14275–14284. doi: 10.1074/jbc.M110.102053
Ye, J., Tang, S., Meng, L., Li, X., Wen, X., Chen, S., et al. (2018). Ultrasonic control of neural activity through activation of the mechanosensitive Channel MscL. Nano Lett. 18, 4148–4155. doi: 10.1021/acs.nanolett.8b00935
Yoo, S., Mittelstein, D. R., Hurt, R. C., Lacroix, J., and Shapiro, M. G. (2022). Focused ultrasound excites cortical neurons via mechanosensitive calcium accumulation and ion channel amplification. Nat. Commun. 13:493. doi: 10.1038/s41467-022-28040-1
Zhang, G., Li, X., Wu, L., and Qin, Y. X. (2021). Piezo1 channel activation in response to mechanobiological acoustic radiation force in osteoblastic cells. Bone Res. 9:16. doi: 10.1038/s41413-020-00124-y
Zhao, L., Feng, Y., Shi, A., Zhang, L., Guo, S., and Wan, M. (2017). Neuroprotective effect of low-intensity pulsed ultrasound against MPP(+)-induced neurotoxicity in PC12 cells: involvement of K2P channels and stretch-activated ion channels. Ultrasound Med. Biol. 43, 1986–1999. doi: 10.1016/j.ultrasmedbio.2017.04.020
Zhao, X., Zhao, G., Shi, Z., Zhou, C., Chen, Y., Hu, B., et al. (2017). Low-intensity pulsed ultrasound (LIPUS) prevents periprosthetic inflammatory loosening through FBXL2-TRAF6 ubiquitination pathway. Sci. Rep. 7:45779. doi: 10.1038/srep45779
Keywords: ultrasound, neuromodulation, Piezo ion channels, transient receptor potential channels, mechanosensitive ion channels
Citation: Song M, Zhang M, He S, Li L and Hu H (2023) Ultrasonic neuromodulation mediated by mechanosensitive ion channels: current and future. Front. Neurosci. 17:1232308. doi: 10.3389/fnins.2023.1232308
Edited by:
Ying Shen, The First Affiliated Hospital of Nanjing Medical University, ChinaReviewed by:
Wen Wu, Southern Medical University, ChinaCopyright © 2023 Song, Zhang, He, Li and Hu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Huijing Hu, aHVodWlqaW5nQG53cHUuZWR1LmNu
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.