
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Neurosci., 30 June 2023
Sec. Neuropharmacology
Volume 17 - 2023 | https://doi.org/10.3389/fnins.2023.1118253
 José Ramón Gutiérrez-Casares1*
José Ramón Gutiérrez-Casares1* Cristina Segú-Vergés2,3
Cristina Segú-Vergés2,3 Juncal Sabate Chueca4
Juncal Sabate Chueca4 Tamara Pozo-Rubio4
Tamara Pozo-Rubio4 Mireia Coma2
Mireia Coma2 Carmen Montoto4
Carmen Montoto4 Javier Quintero5
Javier Quintero5Attention-deficit/hyperactivity disorder (ADHD) is a psychiatric condition well recognized in the pediatric population that can persist into adulthood. The vast majority of patients with ADHD present psychiatric comorbidities that have been suggested to share, to some extent, the pathophysiological mechanism of ADHD. Lisdexamfetamine (LDX) is a stimulant prodrug approved for treating ADHD and, in the US, also for binge eating disorder (BED). Herein, we evaluated, through a systems biology-based in silico method, the efficacy of a virtual model of LDX (vLDX) as ADHD treatment to improve five common ADHD psychiatric comorbidities in adults and children, and we explored the molecular mechanisms behind LDX’s predicted efficacy. After the molecular characterization of vLDX and the comorbidities (anxiety, BED, bipolar disorder, depression, and tics disorder), we created a protein-protein interaction human network to which we applied artificial neural networks (ANN) algorithms. We also generated virtual populations of adults and children-adolescents totaling 2,600 individuals and obtained the predicted protein activity from Therapeutic Performance Mapping System models. The latter showed that ADHD molecular description shared 53% of its protein effectors with at least one studied psychiatric comorbidity. According to the ANN analysis, proteins targeted by vLDX are predicted to have a high probability of being related to BED and depression. In BED, vLDX was modeled to act upon neurotransmission and neuroplasticity regulators, and, in depression, vLDX regulated the hypothalamic-pituitary-adrenal axis, neuroinflammation, oxidative stress, and glutamatergic excitotoxicity. In conclusion, our modeling results, despite their limitations and although requiring in vitro or in vivo validation, could supplement the design of preclinical and potentially clinical studies that investigate treatment for patients with ADHD with psychiatric comorbidities, especially from a molecular point of view.
Attention-deficit/hyperactivity disorder (ADHD) is an impairing psychiatric condition affecting children and adults and is characterized by symptoms of inattention, hyperactivity, and impulsivity (Fayyad et al., 2007; Thomas et al., 2015; Caye et al., 2019). Noteworthy, 80% of adult (Mannuzza et al., 1998; Biederman, 2004; McGough and Barkley, 2004; Wilens et al., 2004; Katzman et al., 2017; Ohnishi et al., 2019; Howard et al., 2020; Kittel-Schneider and Reif, 2020) and 67% of pediatric-adolescent (Larson et al., 2011) patients with ADHD presents psychiatric comorbidities, including depression, anxiety, substance abuse, learning and coordination disorders, and conduct disorders (Mannuzza et al., 1998; Wilens et al., 2004). In ADHD adult patients, the most common psychiatric comorbidities are depression, substance-related disorders, anxiety, and eating disorders (Wilens et al., 2004; Sobanski, 2006; Sobanski et al., 2007; Ohnishi et al., 2019). On the other hand, children and adolescents with ADHD present more often learning and coordination disorders and conduct disorders (including oppositional defiant disorder) but also anxiety and depression (Gillberg et al., 2004; Joelsson et al., 2016; Cuffe et al., 2020). Since ADHD symptoms and its psychiatric comorbidities share similarities, a partial overlap of their pathophysiological mechanisms has been suggested (Franke et al., 2012; Lee et al., 2013; Katzman et al., 2017; Laas et al., 2017).
Currently, a combination of both pharmacological and psychotherapeutic treatments such as cognitive-behavioral therapy (CBT) are treatment options in the clinic for children, adolescents and adults patients with ADHD (Coelho et al., 2017; Pan et al., 2019). CBT has been regarded as one of the most effective psychological treatments for ADHD and, when combined with medication, is associated with greater improvements in adherence to treatment and in core ADHD symptoms, social functions and comorbid symptoms (Coelho et al., 2017; Pan et al., 2019). On the other hand, ADHD medication includes stimulants, among which lisdexamfetamine (LDX), a prodrug with proven efficacy for treating binge eating disorder (BED) and approved for this indication in the adult population of the US (U. S. Food and Drug Administration, 2017). However, the efficacy of LDX on patients with ADHD and psychiatric comorbidities has been poorly studied (Kollins et al., 2011; Roncero and Álvarez, 2014). Also, mechanistic studies on LDX are scarce and could inform clinicians and improve clinical trial designs (Hutson et al., 2014). In this sense, systems biology methods have aided in untangling the molecular effects of drugs in complex clinical settings, such as treatment-resistant pathologies (Akil et al., 2018), data analysis for personalized medicine (Davis et al., 2019), mechanistic studies in drug-refractory patients (Lorén et al., 2019), and in silico head-to-head trials (Caye et al., 2019). We recently performed in silico studies between virtual models of LDX (vLDX) and methylphenidate to treat ADHD in adults and children (Gutiérrez-Casares et al., 2021; Gutierrez-Casares et al., 2023). As an extension of that study, we considered essential to investigate the effects of vLDX over ADHD common psychiatric comorbidities in patients with ADHD.
The main objective of our study was to evaluate, through a systems biology-based in silico method, the efficacy of vLDX as ADHD treatment to improve some common ADHD psychiatric comorbidities. In addition, we explored the molecular mechanisms behind vLDX’s predicted efficacy in virtual adult and pediatric-adolescent comorbid patients with ADHD.
In this study, we applied systems biology-based in silico approaches to evaluate the potential relationship of LDX with psychiatric comorbidities by applying Therapeutic Performance Mapping System (TPMS) technology (Jorba et al., 2020). TPMS is a computational tool that models the protein pathways of a drug or pathology explaining a clinical outcome or phenotype. We applied this technology using two complementary approaches. First, we used artificial neural network (ANN) (Artigas et al., 2020) algorithms, which provide predictive information. Then, we used sampling-based methods combined with other modeling approaches to obtain quantitative systems pharmacology (QSP)-based virtual patient models to explore the mechanisms behind the predicted relationships, as extensively described previously (Gutiérrez-Casares et al., 2021). Briefly, our QSP approach included: the generation of a virtual randomized population following randomized clinical trials’ (Ginsberg et al., 2011; Retz et al., 2012; Coghill et al., 2013) characteristics and European reference population distribution (de Onis et al., 2007; The European Social Survey, 2018), a physiologically based pharmacokinetic (PBPK) modeling approach based on a 14-compartment model (Peters, 2008; Rostami-Hodjegan, 2012; Ciffroy et al., 2016; Brochot and Quindroit, 2018), and TPMS systems biology-based modeling (Jorba et al., 2020), which mimics the human pathophysiology at a protein-network level using machine learning. This method provided virtual patient-specific QSP models of virtual drugs (i.e., defined by the model from available literature) that complied with the accuracy and quality measurements set for each step (Gutiérrez-Casares et al., 2021).
We undertook a bibliographically based structured search to obtain a protein-based molecular characterization of ADHD and of LDX to be used as input for TPMS models (Jorba et al., 2020). Aside from a review of official regulatory documentation and drug-target dedicated databases, a review of the currently available bibliography regarding known targets of the drugs was performed in PubMed on April 27, 2020. The specific searches performed can be found in Gutiérrez-Casares et al. (2021). The list of publications identified in the specific searches was retrieved and assessed at the title and abstract level. If molecular information describing pathophysiological conditions was found, the full texts were thoroughly reviewed to identify the main pathophysiological processes known to be involved in the diseases. We defined ADHD by four motives (or biological processes) that describe the pathophysiology of the disease: neurotransmitter imbalance, neuroinflammation, circadian system imbalance, and altered neural viability. We only included proteins for which a functional role on the disease was reported (Supplementary Table A). Then, we applied the same procedure to characterize all five studied comorbidities, i.e., anxiety, BED, bipolar disorder, depression, and tics disorder (Supplementary Table A).
For the definition and modeling of the virtual drug vLDX, we considered its prodrug nature, and compiled information and modeled the behavior of its active metabolite, d-amphetamine, as previously described (Gutiérrez-Casares et al., 2021). Briefly, we defined vLDX by the protein targets affected by LDX active form, according to a structured search bibliographical analysis, and considered as drug targets those proteins for which the drug had activity either in vitro or in vivo (Table 1). For the QSP modeling, we used d-amphetamine concentration curve associated to LDX treatment (Gutiérrez-Casares et al., 2021), obtained from adjusting a physiologically based pharmacokinetic model (Peters, 2008; Rostami-Hodjegan, 2012; Ciffroy et al., 2016; Brochot and Quindroit, 2018) to reported pharmacokinetic parameters (oral administration; kidney as main clearance organ; bioavailability: 96.4%) and real plasma concentration data (Krishnan and Zhang, 2008; Boellner et al., 2010; Maldonado, 2013; Bundesinstitut für Arzeimittel und Medizinprodukte, 2020). This approach allowed us to obtain a drug concentration curve per virtual patient considering their individual characteristics (weight, height, age, and sex).
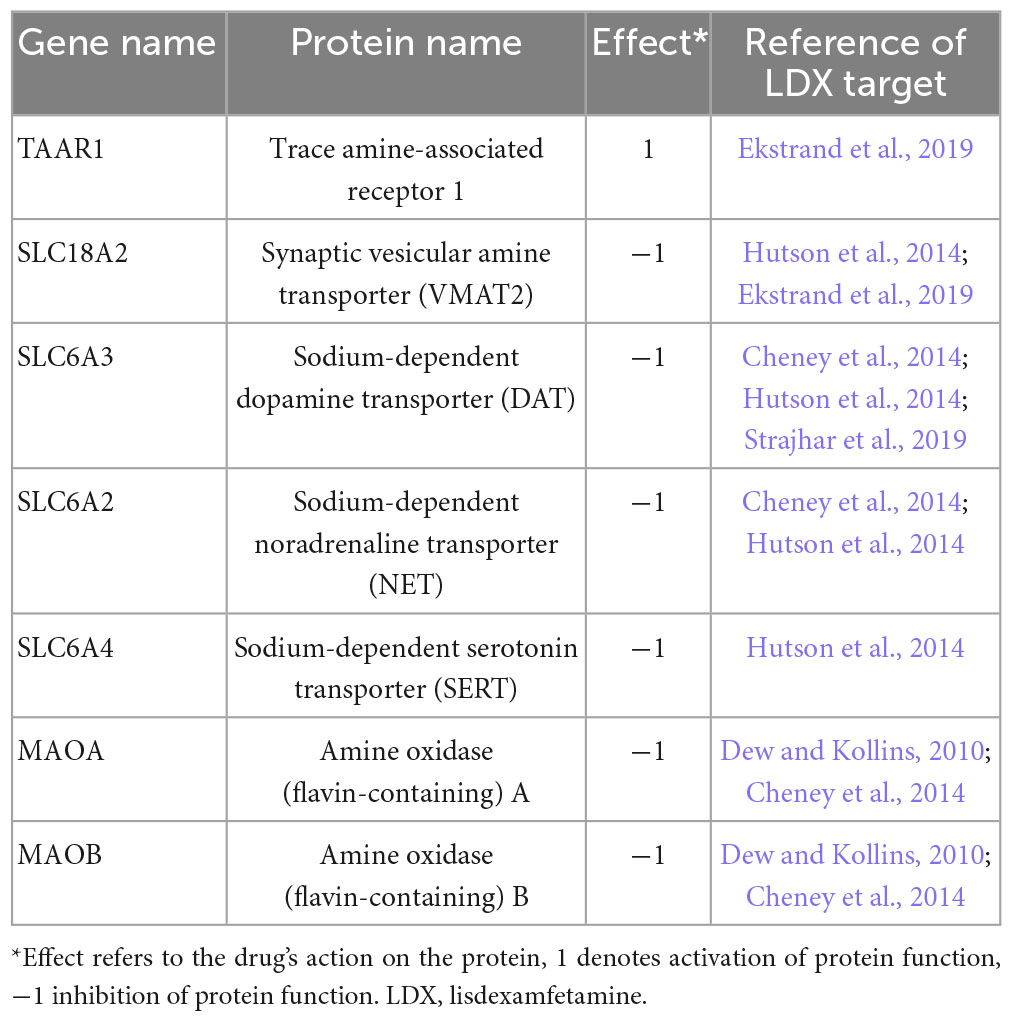
Table 1. Identified protein targets for lisdexamfetamine (Gutiérrez-Casares et al., 2021).
Therapeutic performance mapping system is based on the human protein interaction network and pharmacology and pathophysiology information, which constitutes the training set (Supplementary Table B). To create TPMS models, we exploited an in-house database drawn from public sources—KEGG (Kanehisa et al., 2017), REACTOME (Jassal et al., 2020), INTACT (Orchard et al., 2014), BIOGRID (Oughtred et al., 2019), HPRD (Keshava Prasad et al., 2009), and TRRUST (Han et al., 2018)—as a basis to create the protein-protein interaction human network. We used Cytoscape v. 3.6.0 (Shannon et al., 2003) to visualize this network. We applied ANN (Artigas et al., 2020) algorithms that identify the probability of a specific relationship between two or more protein sets. The ANN algorithm provides a score (ranging from 0 to 100%) associated with a probability (p-value) that the drug (group of protein targets) and the pathology (defined by the molecular description of pathological processes detected in the characterization) being evaluated are functionally connected. Therefore, scores greater than 92% indicate a very strong relationship with a p-value below 0.01; scores between 77 and 92% imply a strong relationship with a p-value between 0.01 and 0.05; scores between 37 and 77% have a medium relationship and a p-value in the range 0.05–0.25; and scores lower than 37% indicate a poor relationship with p-values above 0.25. Finally, we modeled the mechanisms of vLDX for the most relevant comorbidities by using sampling methods after QSP modeling. These models provide mechanisms of action that explain how a stimulus (i.e., proteins activated or inhibited by a drug) produces a response (i.e., proteins active or inhibited in a phenotype) (Jorba et al., 2020). The quality of these models is measured by its accuracy, which is calculated as the percentage of compliance of all drug-pathophysiology relationships included in the training set (Jorba et al., 2020; Gutiérrez-Casares et al., 2021).
The methodology used for developing the in silico clinical trial from which this detailed mechanistic study derives is described in detail elsewhere (Gutiérrez-Casares et al., 2021). Briefly in that study, we used expression data to explore molecular variability and estimate a minimum population size to ensure statistical power; 71 was set as the minimum population size per cohort. Therefore, we generated two virtual populations (adults–18 years old and older–and children-adolescents–from 6 to 18 years old) with ADHD comorbidities, with 100 virtual patients per each age category and comorbidity.
We created a randomized population demographic characteristics using ADHD trials as a reference for mean demographic values (Ginsberg et al., 2011; Retz et al., 2012; Coghill et al., 2013) and standard population information (de Onis et al., 2007; The European Social Survey, 2018) to obtain the randomized patient distribution fitting the desired population values.
Quantitative systems pharmacology models are generated following the TPMS methodology but incorporating drug concentration data at different timepoints in addition to molecular inputs, which add patient-specific quantitative data. To this end, a set of drug concentration timepoints in the target tissue—brain in this study—can be associated with the modulation of the drug’s target proteins. Accordingly, the resulting LDX drug’s target modulation-efficacy relationships were used as extra parameters in the TPMS training set, resulting in the final QSP models. Through the use of clinical efficacy values for various drugs tested in ADHD clinical trials, modeled drug concentration curves were plotted and used to obtain restrictions on target inhibition with ADHD modulation. These restrictions were compatible with systems biology-based TPMS models (Jorba et al., 2020) and conferred them a quantitative dimension. Thus, we obtained a TPMS-derived QSP model per each virtual patient and each virtual drug. We used comorbidity-specific virtual patients for sampling methods-based mechanistic evaluation in each case.
Due to the systems-biology–based nature of the virtual patients’ resulting models, all measures were centered on protein activity. We obtained the predicted protein activity (ranging from −1 to 1) from each QSP model (Jorba et al., 2020; Gutiérrez-Casares et al., 2021). We analyzed these data for each protein individually, calculated the tSignal (Jorba et al., 2020; Gutiérrez-Casares et al., 2021), and defined a reverted protein as one whose activation sign in the comorbidity under study was reverted due to the effect of vLDX. In the current study, we considered reverted proteins as those for which (A) the direction of their activation was opposite to the one to induce the disease and (B) the absolute value of its predicted protein activity was at least half of the maximum value achievable (i.e., higher than 0.5). For mechanisms of action, exploration, we focused on the paths (three-step protein interaction between the stimulus and the response) obtained from TPMS models, as previously described (Jorba et al., 2020; Segú-Vergés et al., 2021), that (A) justified at least a protein effector of the studied comorbidity reverted by vLDX with at least 80% of the maximum value achievable (i.e., | predicted activation| > 0.8) and (B) with a frequency = 100% within the population of comorbidity-specific virtual patients.
According to the bibliography-based molecular characterization of the pathologies under study (Supplementary Table A), the ADHD molecular description shared 53% of its protein effectors with at least one studied psychiatric comorbidity (Figures 1A, B). ADHD shared between 8 and 34% of its effectors with the molecular definition of each individual comorbidity, being anxiety the one with the largest overlap. According to the protein-protein network, 84% of ADHD molecular effectors were shared or directly connected to effectors of at least one comorbidity. The comorbidity with the largest connection was depression, for which 52% of ADHD effectors were shared or directly connected to 54% of depression effectors. All vLDX targeted proteins were found to be effectors of ADHD or at least one of its psychiatric comorbidities (Figure 1C). Interestingly, the LDX target TAAR1, classified as a BED effector, is directly related to DRD2, a protein identified as an effector in ADHD and all five characterized psychiatric comorbidities.
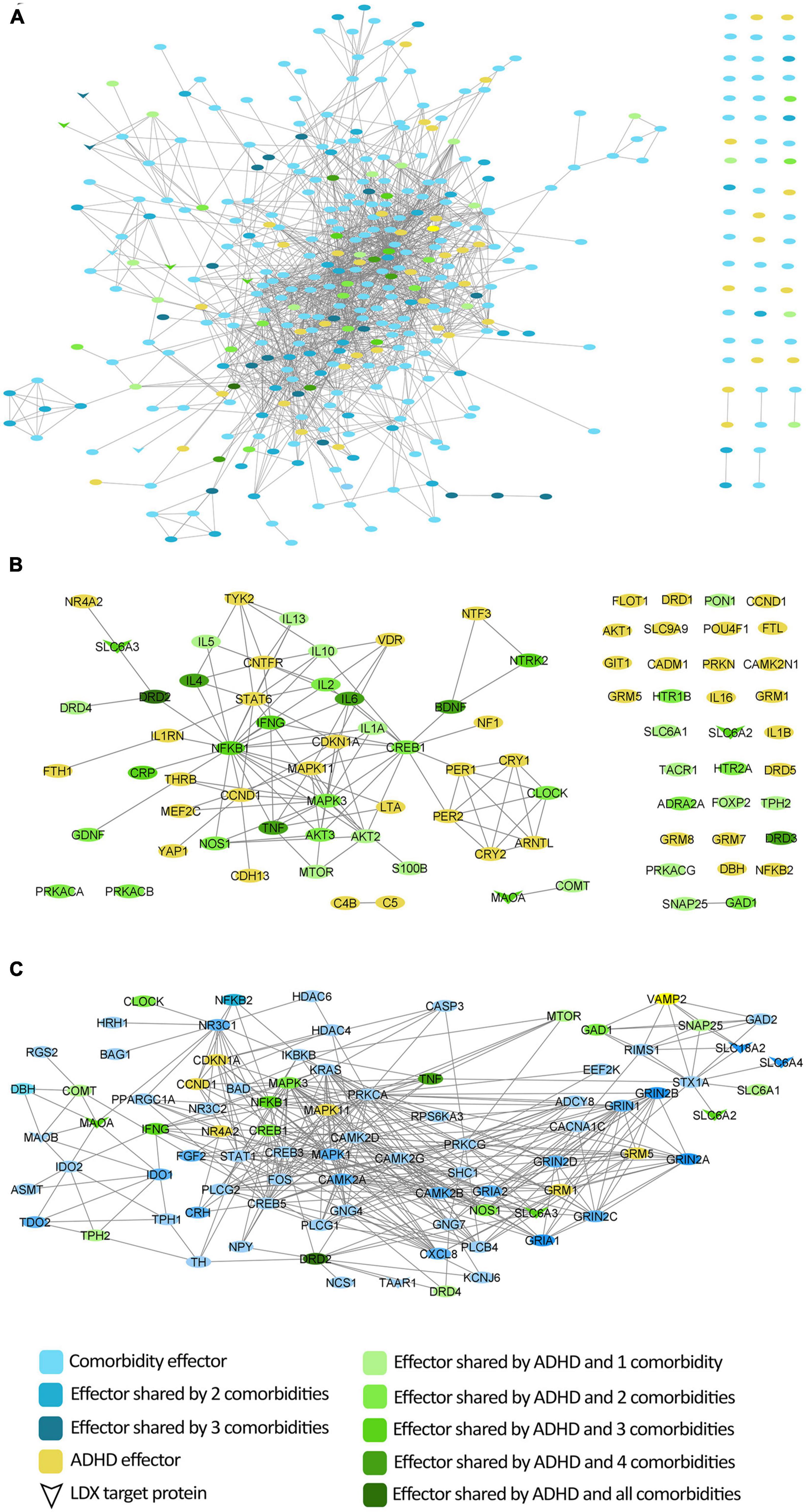
Figure 1. Protein interactome around ADHD molecular characterization considering the protein effectors of its most common comorbidities. (A) Network of direct interactions between the protein effectors of ADHD and those of its comorbidities; (B) Network of ADHD protein effectors representing the overlap with comorbidities; (C) Network of vLDX targets and their interactions with the protein effectors of ADHD and its comorbidities. ADHD, attention-deficit/hyperactivity disorder; vLDX, virtual lisdexamfetamine.
Artificial neural networks showed that the proteins targeted by vLDX had a high probability (p < 0.01) of being related to BED and depression (Table 2). However, this probability was moderate (p < 0.25) for anxiety and tic disorders and very low (p ≥ 0.25) for bipolar disorder. Although the ANN values for anxiety (76%) and tic disorders (71%) were very close to the p-value < 0.05 threshold (77%), only the results for BED and depression were considered statistically significant and subjected to detailed mechanistic evaluation.
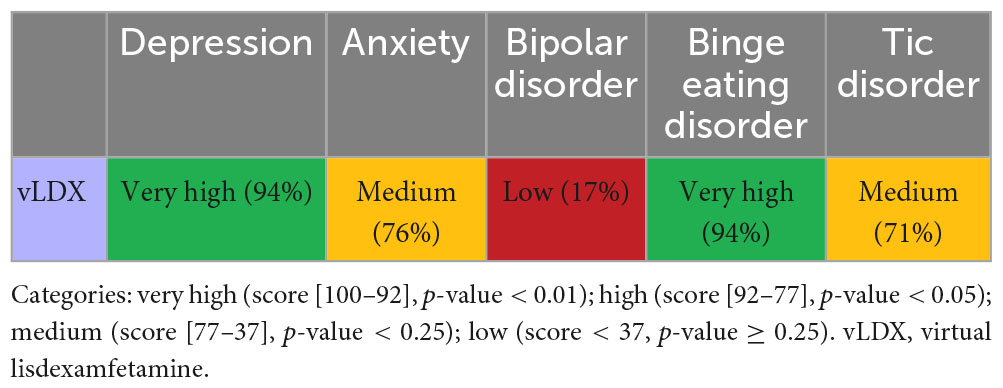
Table 2. Artificial neural network evaluation of the impact of virtual lisdexamfetamine over comorbidities.
Virtual model of Lisdexamfetamine modulated BED by regulating neurotransmission (DAT, MAO, NET1) and neuronal survival and plasticity (BDNF-NTRK2) (Figure 2). In both adult and children-adolescent models, vLDX was able to revert eight BED-related protein effectors with | predicted protein activity| > 0.5, four of which (NET1, DAT1, BDNF, and NTRK2) are also involved in ADHD pathophysiological mechanisms (Table 3). In patients with ADHD presenting BED as a comorbidity, the protein targets DAT1, NET1, SERT, and TAAR1 were highly reverted by vLDX, as well as the neurotrophic factor BDNF (affecting the BDNF-NTRK2 pathway) (Figure 2, Supplementary Figure A and Supplementary Table C). Interestingly, NET1 was predicted to be inhibited by vLDX not only directly but also indirectly through TAAR1 activation, inducing the PKC signaling pathway. In addition, BDNF and its receptor NTRK2 were activated via TAAR1-initiated PKA signaling. vLDX was also found to modulate CRHR1 (Table 3).
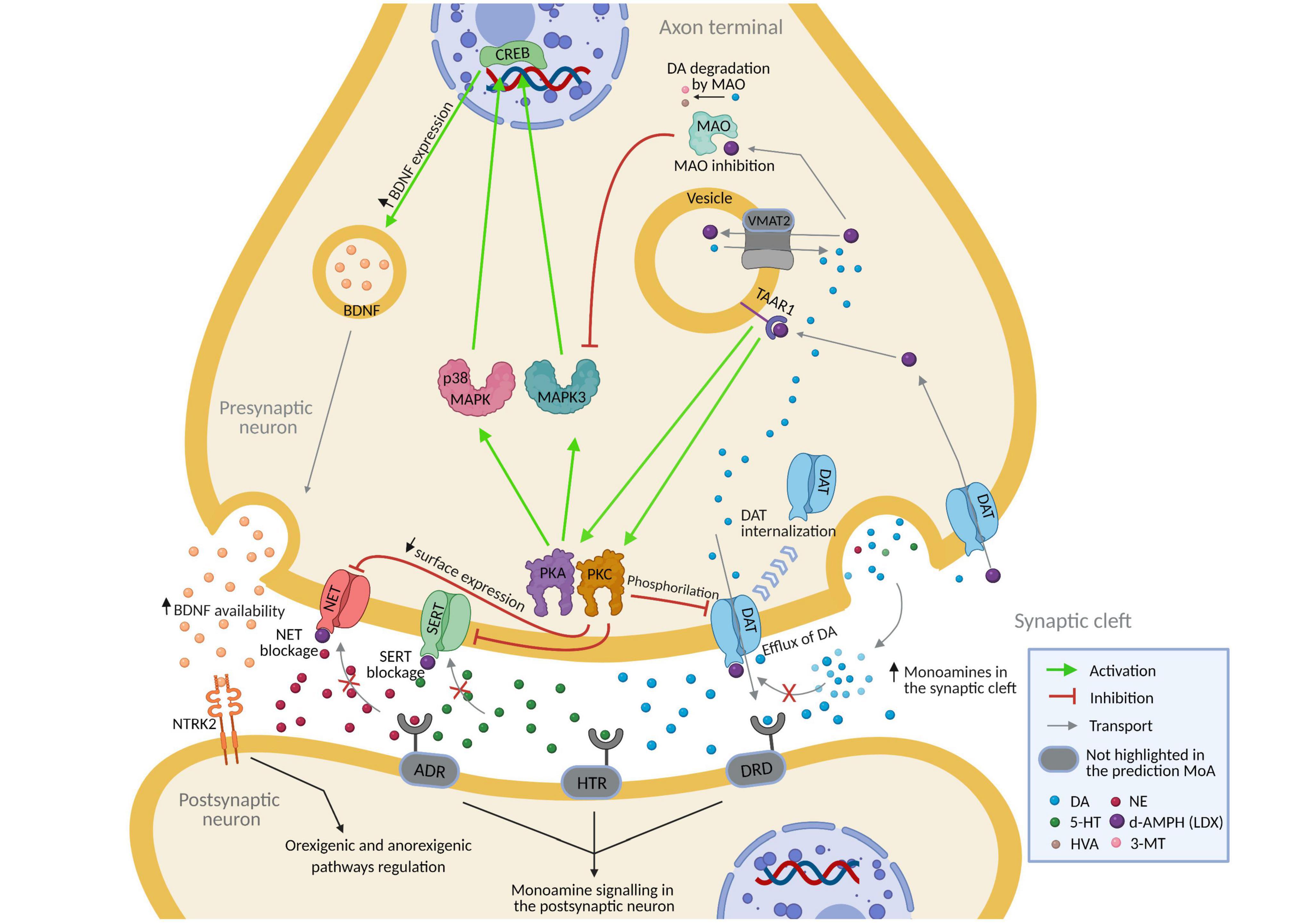
Figure 2. Predicted mechanism of action of lisdexamfetamine (according to vLDX QSP models) over binge eating disorder in patients with ADHD presenting this comorbidity. Supplementary Figure A and Supplementary Table C contain the sources of information found in the scientific literature supporting the predicted mechanism. Created with BioRender.com. 3-MT, 3-Methoxytyramine; 5-HT, Serotonin; ADHD, attention-deficit/hyperactivity disorder; ADR, adrenoreceptors; DA, dopamine; d-AMPH, d-Amphetamine; DAT, sodium-dependent dopamine transporter; DRD, dopamine receptor; HTR, serotonin receptor; HVA, homovanillic acid; NE, norepinephrine; NET, sodium-dependent noradrenaline transporter; NTRK2, neurotrophic receptor tyrosine kinase 2; QSP, quantitative systems pharmacology; SERT, sodium-dependent serotonin transporter; vLDX, virtual lisdexamfetamine.
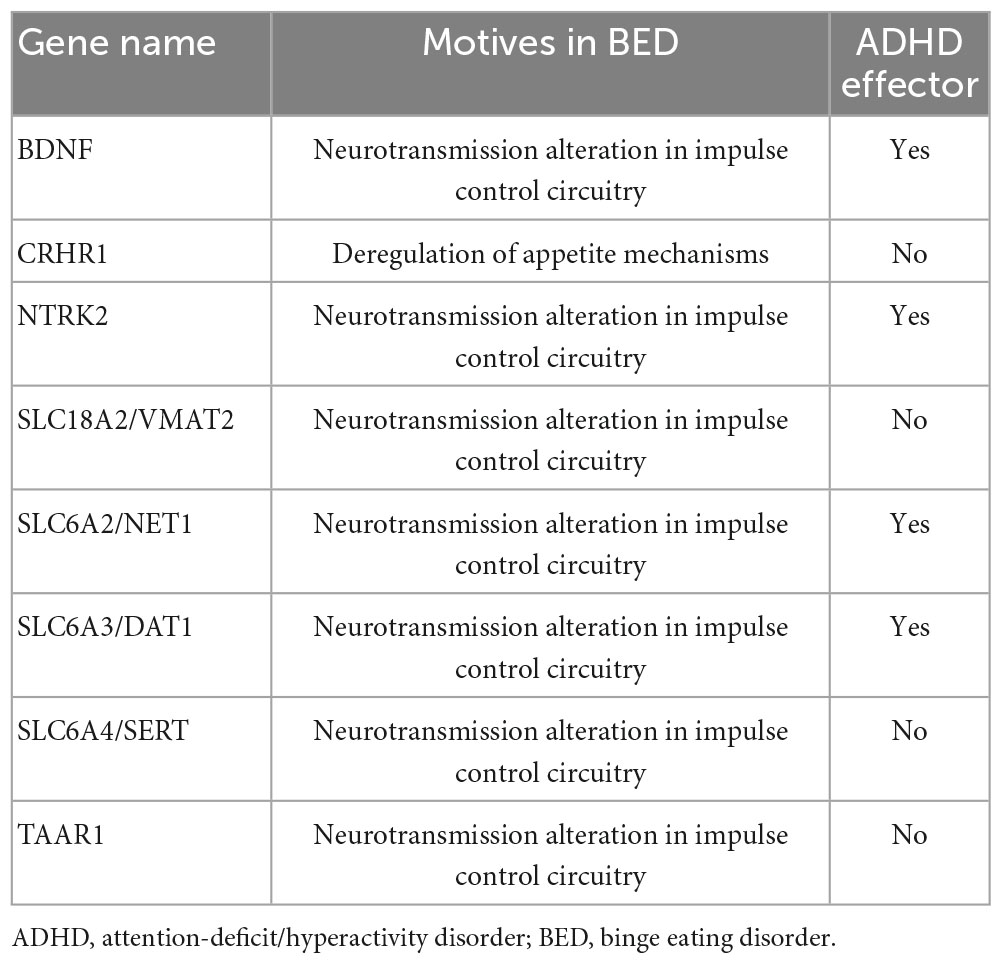
Table 3. Proteins related to binge eating disorder and reverted by virtual lisdexamfetamine with | predicted protein activity| > 0.5 in both adult and pediatric-adolescent populations.
Virtual model of Lisdexamfetamine modulated depression by regulating glutamatergic excitotoxicity, neurotransmission, neuroplasticity, the hypothalamic-pituitary-adrenal (HPA) axis, neuroinflammation, and oxidative stress (Table 4). According to our models, vLDX was able to revert 49 depression-related effectors in adults and 44 in children with | predicted protein activity| > 0.5, 17 of which (BDNF, CREB1, CRP, IL-1A, IL-1B, IL-6, MAO, MTOR, NFKB1, NTRK2, PRKACA, PRKACB, NET1, DAT1, and TNF in both populations and IFNG and IL-2 only in adults) were also involved in ADHD pathophysiological mechanisms. In addition, in both adult and pediatric-adolescent ADHD patient populations, vLDX was found to modulate depression-related processes through glutamatergic excitotoxicity (VAMP2), neurotransmission alteration (SERT, DAT1, MAO, NET1, NOS2, TAAR1, IFN, IFNGR1, IFNGR2), HPA axis hyperactivation (BDNF), neuroinflammation (TNF, IL-1, IL-6, IFNG, NF-κB), and loss of neural plasticity (CREB1, BDNF, PKA) (Figure 3, Supplementary Figure B and Supplementary Table D).
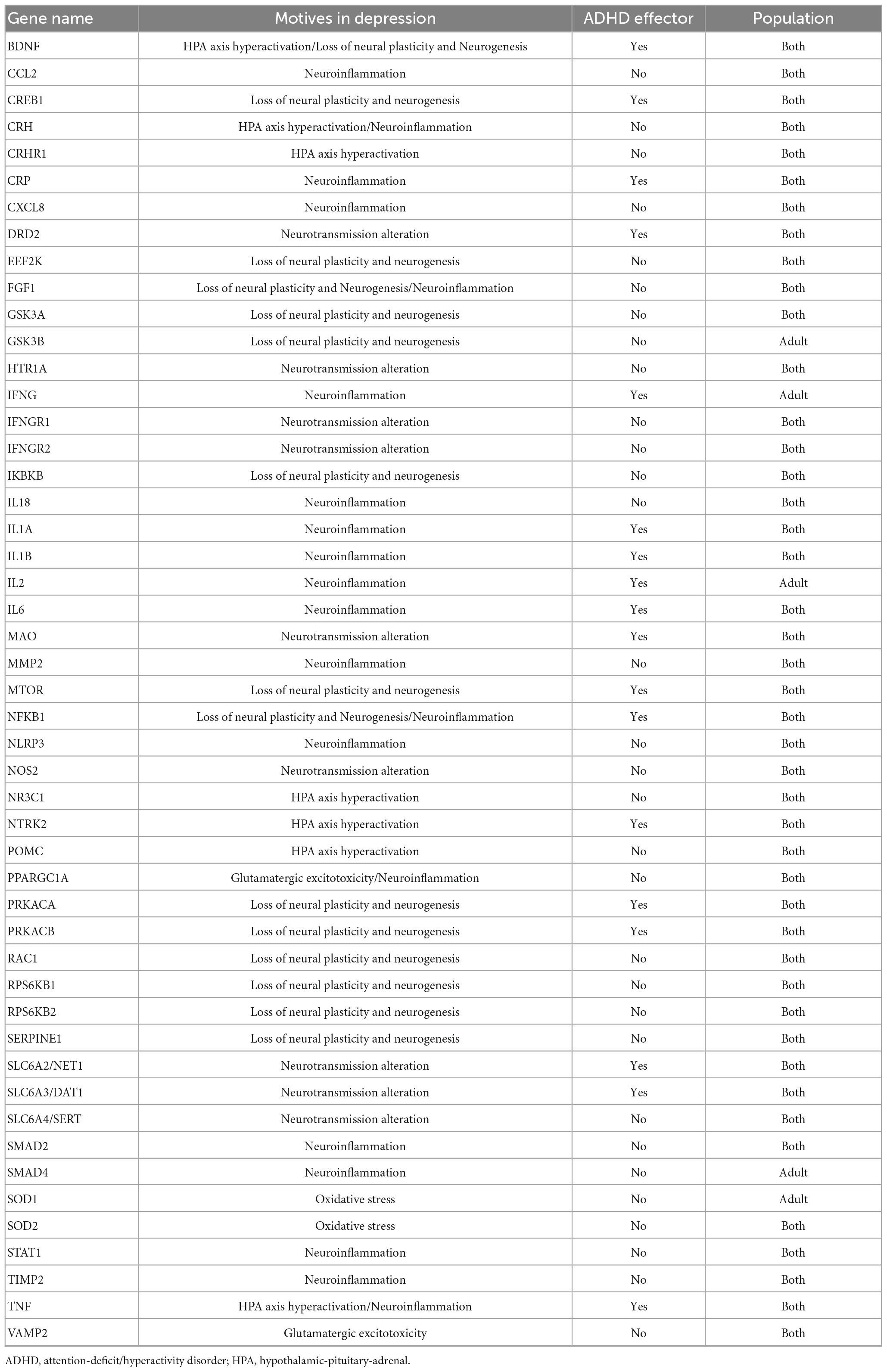
Table 4. Depression-related proteins reverted by virtual lisdexamfetamine with | predicted protein activity| > 0.5 in adult, pediatric-adolescent or both virtual populations.
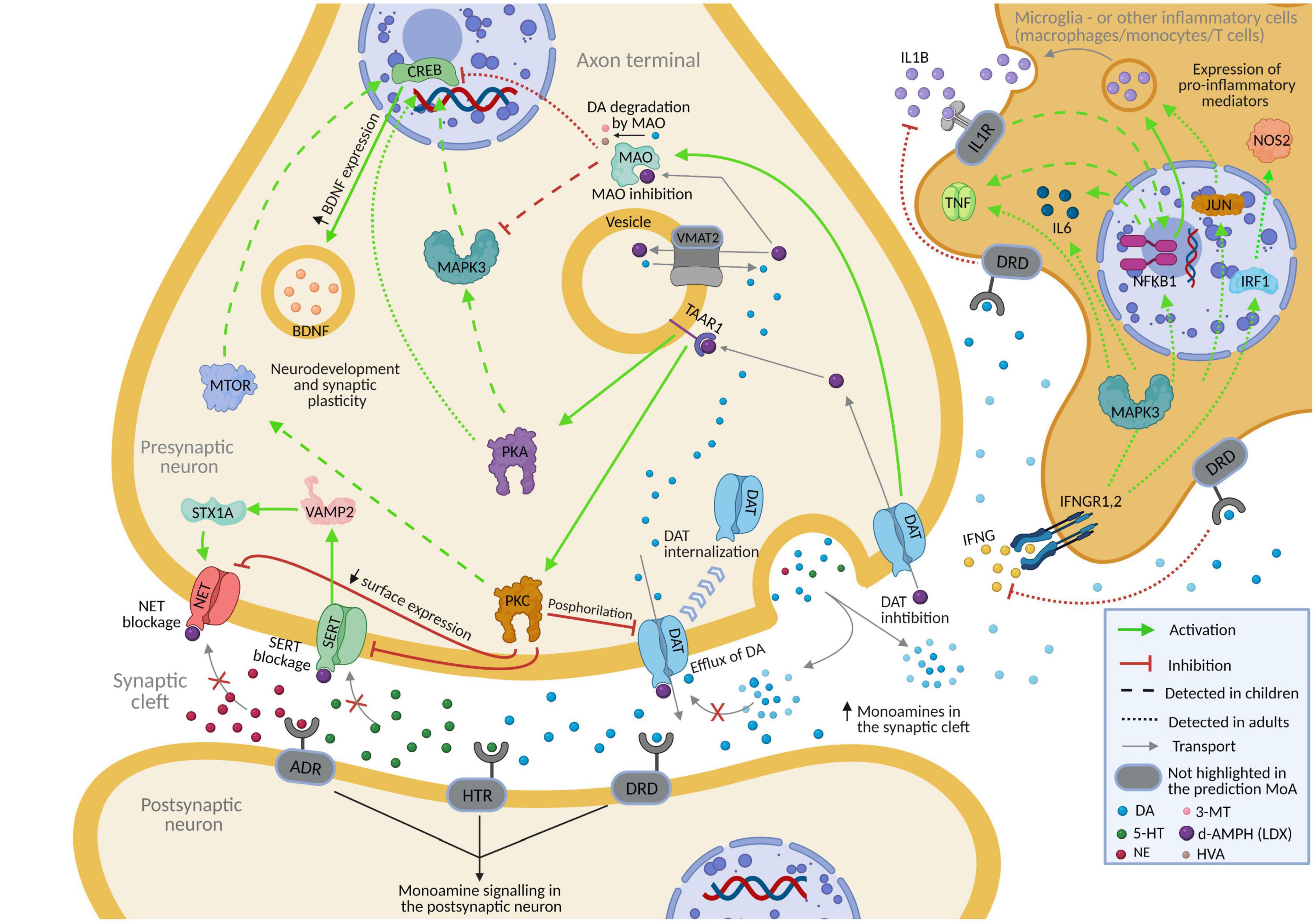
Figure 3. Predicted mechanism of action of lisdexamfetamine (according to vLDX QSP models) over depression in patients with ADHD presenting this comorbidity. Supplementary Figure B and Supplementary Table D contain the sources of information found in the scientific literature supporting the predicted mechanism. Created with BioRender.com. 3-MT, 3-Methoxytyramine; 5-HT, serotonin; ADHD, attention-deficit/hyperactivity disorder; ADR, adrenoreceptor; DA, dopamine; d-AMPH, d-Amphetamine; DAT, sodium-dependent dopamine transporter; DRD, dopamine receptor; HTR, serotonin receptor; HVA, homovanillic acid; IL1R, interleukin-1 receptor; IFNG, interferon-gamma; IFNGR1.2, interferon-gamma receptor; NE, norepinephrine; NET, sodium-dependent noradrenaline transporter; QSP, quantitative systems pharmacology; SERT, sodium-dependent serotonin transporter; vLDX, virtual lisdexamfetamine.
Through a systems biology-based in silico method, we found that vLDX’s target proteins were related to BED and depression. Considering LDX use as treatment in patients with ADHD and taking a previously described virtual population of patients with ADHD and psychiatric comorbidities (Gutiérrez-Casares et al., 2021), we explored the detailed mechanisms behind vLDX activity on BED and depression as comorbidities in patients with ADHD. According to our modeling results, in BED, vLDX was shown to act upon neurotransmission and neuroplasticity regulators, and, in depression, vLDX regulated the HPA axis, neuroinflammation, oxidative stress, and glutamatergic excitotoxicity.
Based on the available literature, the diseases’ molecular characterization showed that more than half of the proteins affected by ADHD were also deregulated in at least one of the evaluated psychiatric comorbidities (i.e., depression, anxiety, BED, bipolar disorder, and tic disorders). In adults, ADHD is highly comorbid with other psychiatric disorders; 65–89% of patients with ADHD present at least one psychiatric comorbidity, mainly anxiety and mood and substance use disorders (Biederman, 2004; Kessler et al., 2006; Sobanski, 2006; Sobanski et al., 2007; Piñeiro-Dieguez et al., 2016). In children and adolescents with ADHD, psychiatric comorbidities are even more prevalent and 60–100% present at least one psychiatric comorbidity, mostly conduct and anxiety disorders and depression (Biederman, 2004; Sobanski, 2006; Sobanski et al., 2007; Joelsson et al., 2016; Cuffe et al., 2020).
The association between ADHD and BED has been established by several clinical studies in adults and children (Cortese et al., 2007; Bleck et al., 2015; Rankin et al., 2016). In our ANN analysis, vLDX pharmacological protein targets were most likely related to BED development, thus supporting LDX’s ability to manage BED. These results are supported by several randomized, double-blind, placebo-controlled multicenter studies that have demonstrated the efficacy of LDX in treating BED (McElroy et al., 2016; Hudson et al., 2017). Of note, in the US, LDX is indicated for BED treatment in adults (U. S. Food and Drug Administration, 2017). Although not approved for use in children and adolescents with this indication, LDX has proven to be effective in adolescents >12 years old (Guerdjikova et al., 2019; Srivastava et al., 2019).
Monoamine transporters NET1, DAT1, and SERT, which are reported LDX targets (Popovic et al., 2009; Guerdjikova et al., 2016), were predicted to be related to vLDX mechanisms in BED and depression improvement in ADHD virtual patients. In our sampling methods-based BED model, NET1 was also inhibited indirectly via the PKC signaling pathway initiated by vLDX-mediated TAAR1 stimulation, which has been reported to improve neurotransmission alteration (Xie and Miller, 2007; Bermingham and Blakely, 2016; Appolinario et al., 2019). Similarly, the BDNF-NTRK2 pathway was activated through TAAR1-initiated PKA signaling, which has been reported to enhance neurotransmission (Xue et al., 2016; Ceccarini et al., 2020). Notably, BDNF synthesis and secretion has been located in presynaptic neuronal sites (Song et al., 2017). vLDX was also found to modulate CRHR1, a protein that has been reported to be involved in appetite regulation (Ghitza et al., 2006; Iemolo et al., 2013; Micioni Di Bonaventura et al., 2017, 2020, 2021). A recent review (Schneider et al., 2021) concluded that LDX’s mechanisms to treat BED encompass a complex combination of processes, including effects on appetite/satiety, reward, and cognitive processes (such as attention and impulsivity/inhibition), mainly through catecholamine and serotonin neuronal pathways in the brain, which are in line with our results. This study also evidenced a lack of specific information on mechanisms that could be differential between both populations. According to our results, no differences in the mechanisms reducing BED were found between our two virtual populations. However, as further discussed in detail later in this section, our models only considered age differences derived from PBPK modeling and brain drug concentration curves, which rendered our models incomplete in this sense.
Our ANN analysis also showed a highly probable relation between proteins targeted by vLDX and depression. In this sense, current available evidence in the literature points toward a slight improvement in depressive symptoms when using LDX as antidepressant augmentation therapy (Corp et al., 2014; Madhoo et al., 2014; Giacobbe et al., 2018). In our study, vLDX was found to act upon DAT1, MAO (Popovic et al., 2009), TAAR1 (Moore et al., 2018), NET1, SERT (Guerdjikova et al., 2016), and NOS2 (Inserra et al., 2019), all of which have been linked with the characteristic impairment in neurotransmission observed in depressive patients (Sung et al., 2003; Sotnikova et al., 2009; Gaweska and Fitzpatrick, 2011; Vaughan and Foster, 2013; Moore et al., 2018; Inserra et al., 2019; Kegg Pathway Database, 2020). Interestingly, DAT1 inhibition has been shown to prevent dopamine reuptake into the cytosol (Vaughan and Foster, 2013) and its degradation by MAO (Gaweska and Fitzpatrick, 2011; Kegg Pathway Database, 2020), hence, improving the impaired neurotransmission (Miller and Raison, 2016). Also, TAAR1 may directly interact with DAT, thereby affecting dopamine neurotransmission via modulation of DAT function (Sotnikova et al., 2009). The inhibition of NET1 by vLDX via TAAR1 stimulation could ameliorate the neurotransmission alteration found in depressive patients (Peterson et al., 2015; Bermingham and Blakely, 2016; Moore et al., 2018). Moreover, this effect on NET1 may be enhanced by the blockade of STX1A by vLDX through SERT since it supports surface trafficking of NET1 and regulates the transporter catalytic function (Sung et al., 2003). On the other hand, the reduction in NOS2 levels exerted by LDX could permit the production of norepinephrine, usually blocked by this enzyme in depression (Inserra et al., 2019). According to our results, vLDX affected processes directly related to the HPA axis, whose hyperactivity has been implicated in the pathophysiology of depression (Pariante and Lightman, 2008; Keller et al., 2017; Menke, 2019; Iob et al., 2020; Pitsillou et al., 2020). Besides, vLDX modulated proteins involved in neuroinflammation and oxidative stress in depressive states (Inserra et al., 2019; Pitsillou et al., 2020), processes that have been repeatedly suggested to play a role in these patients (Yanik et al., 2004; Bakunina et al., 2015; Black et al., 2015; Liu et al., 2015; Lindqvist et al., 2017). According to our model, and as previously described in experimental models (Vidal and Pacheco, 2020), dopamine signaling modulated neuroinflammation by regulating different immune cells, including microglia, cells from the monocytic lineage, and T cells. High dopamine levels have been reported to reduce nitric oxide production in rodent microglial cells (Katrin et al., 2005). In relation to neuroinflammation, by regulating dopamine synaptic availability, vLDX could modulate a wide spectrum of cytokines and other inflammatory factors induced by microglia (or other immune cells) such as IL-6 (Garner et al., 2018; Bobbo et al., 2019), IL-1B (Liu and Quan, 2018), TNF (Welser-Alves and Milner, 2013), and IFNG receptors (Tsuda et al., 2009; Ta et al., 2019), all shown to be involved in the inflammatory process of depression (Inserra et al., 2019; Pitsillou et al., 2020). Particularly, the current corpus of evidence points toward a role for high dopamine levels in attenuating the inflammatory activation of microglia through the stimulation of low-affinity dopamine receptors (including DRD1, DRD2, and DRD4) (Vidal and Pacheco, 2020). Our model also showed that vLDX activated BDNF, an essential factor for neurogenesis and neural plasticity and downregulated in depression (Juhasz et al., 2011; Schmid et al., 2015; Cui et al., 2019; Underhill et al., 2021). BDNF activation by CREB1 (Juhasz et al., 2011) might be possible because of the inhibition of DAT1 and MAO by vLDX or the stimulation of PRKACB by TAAR1 (Schmid et al., 2015; Underhill et al., 2021). In addition, we found that vLDX dampened glutamatergic excitotoxicity through VAMP2 modulation, a phenomenon that has been observed in patients with depression (Mitchell and Baker, 2010; Dantzer and Walker, 2014; Miller and Raison, 2016; Cui et al., 2019; Pitsillou et al., 2020). In summary, our models propose several mechanisms involved in the modulation of depression-related processes by vLDX, including, the regulation of the HPA axis, oxidative stress and neuroinflammation, neural plasticity and glutamatergic neurotoxicity, mainly through the joint effect over neurotransmitter transporters (SERT, DAT1 and NET) but also regulation of intracellular PKA and PKC signaling through TAAR1 agonism. Finally, some minor differences were observed between adult and children depression models, which rise from the differences in drug pharmacokinetics in both populations according to physiologically based pharmacokinetic models (Ciffroy et al., 2016; Brochot and Quindroit, 2018; Gutiérrez-Casares et al., 2021). Further studies considering other factors (such as neurodevelopment with age) might provide more specific insights into the different mechanisms of LDX in adults regarding pediatric patients.
Notwithstanding the abovementioned, our study presented some limitations that should also be considered. Firstly, the main constraint was the available information, at the moment of the study, on the drug and diseases under study upon which our models were built. In the current study, this factor might be aggravated by the difficulty of studying psychiatric diseases molecularly and by the high genetic and signaling overlap between ADHD and its psychiatric comorbidities (Faraone and Mick, 2010; Faraone and Larsson, 2019), which may act as confounding factors at the clinical and molecular levels. Thus, our models could have been susceptible to information availability bias, leading to some aspects being overlooked. This information could certainly be more extensive in the future (e.g., unknown drug targets or yet undescribed pathophysiological processes), circumscribing our analysis to the present time. Additionally, our modeling methodology has the inherent restraints of mathematical modeling, which cannot fit 100% of the training data information (TPMS viable solutions have always at least 85% accuracy, easily achieving around 95% mean accuracy). Secondly, our approach considered only the impact of demographic characteristics, including age, on the PBPK modeling. However, psychiatric conditions can involve different neuronal mechanisms depending on the patient’s neurodevelopmental stage (Sonuga-Barke and Halperin, 2010; Dreyfuss et al., 2017), which our models did not contemplate. This limitation could prevent the detection of relevant results regarding differences in the mechanisms mediating the effects of LDX in adults and pediatric and/or adolescent populations. These and other limitations of the QSP modeling and their impact on the in silico clinical trial approach are described in detail elsewhere (Gutiérrez-Casares et al., 2021). Third, our approach is a simplification of the complex real-world clinical and pharmacological context of ADHD patients and we did not consider the outcomes in patients taking other substances. Individuals with ADHD are known to self-medicate with substances such as nicotine and cannabinoids because they could alleviate ADHD symptoms (and its comorbidities) (Lee et al., 2011). Although patients with ADHD have regularly stated that cannabis has helped to improve their ADHD symptoms, it is still controversial how and whether cannabis impacts on ADHD symptoms or on pharmacological therapies (Francisco et al., 2023). Regarding nicotine, this substance has been reported to improve ADHD symptoms, but health issues associated with smoking seems to indicate that stopping the use of nicotine offers even more benefits for people with ADHD (van Amsterdam et al., 2018). Therefore, further in vivo research on polydrug reactions in ADHD is needed to fully understand these complex interactions and how it impacts on ADHD and its psychiatric comorbidities. Lastly, although this in silico modeling allowed us to get more insight into vLDX activity on BED and depression in patients with ADHD disease, which may be helpful to guide further research, the results obtained in this study must be validated or refuted with in vitro and/or in vivo data.
According to our in silico modeling study, vLDX’s targets were predicted to be related with high probability to proteins involved in BED as a comorbidity in patients with ADHD, for which LDX is currently indicated in the US adult population. In addition, vLDX was modeled to impact the pathophysiology of an ADHD comorbidity highly prevalent in adults and adolescents, namely, depression. Reasoned mechanisms were presented to explain our results other than neurotransmitter regulation, including effects on neuroplasticity, neuroinflammation, oxidative stress, and the HPA axis. These results could inform preclinical and even clinical future investigations treatments for patients with ADHD and psychiatric comorbidities.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
JG-C, JQ, and CM designed the study. CS-V and MC performed the investigation and formal analyses. JG-C, JQ, JS, and TP-R contributed to data interpretation. JG-C and JQ drafted the first version of the manuscript. All authors critically revised all drafts of the manuscript for intellectual content.
This study was funded by the Takeda Pharmaceutical Company Limited. Under the direction of the authors, Takeda Pharmaceutical Company Limited provided funding to Matías Rey-Carrizo (BCN Medical Writing) for support in writing and editing this manuscript.
We would like to thank Helena Bartra (Anaxomics Biotech, Barcelona) for her assistance in results visualization.
JG-C has served as speaker for Takeda and has received research funding from Shire. JQ has served as speaker and/or on scientific advisory boards for Takeda, Janssen, and Rubio. JS, CM, and TP-R are full-time employees at Takeda Farmacéutica España. CS-V and MC are full-time employees at Anaxomics Biotech. Anaxomics Biotech was funded by Takeda Farmacéutica España for contracted research. The authors declare that this study received funding from Takeda. The funder had the following involvement with the study: study design, contribution to data interpretation, and critical revision of the manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnins.2023.1118253/full#supplementary-material
Akil, H., Gordon, J., Hen, R., Javitch, J., Mayberg, H., McEwen, B., et al. (2018). Treatment resistant depression: a multi-scale, systems biology approach. Neurosci. Biobehav. Rev. 84, 272–288. doi: 10.1016/j.neubiorev.2017.08.019
Appolinario, J., Nardi, A., and McElroy, S. (2019). Investigational drugs for the treatment of binge eating disorder (BED): an update. Expert Opin. Investig. Drugs 28, 1081–1094. doi: 10.1080/13543784.2019.1692813
Artigas, L., Coma, M., Matos-Filipe, P., Aguirre-Plans, J., Farrés, J., Valls, R., et al. (2020). In-silico drug repurposing study predicts the combination of pirfenidone and melatonin as a promising candidate therapy to reduce SARS-CoV-2 infection progression and respiratory distress caused by cytokine storm. PLoS One 15:e0240149. doi: 10.1371/journal.pone.0240149
Bakunina, N., Pariante, C., and Zunszain, P. (2015). Immune mechanisms linked to depression via oxidative stress and neuroprogression. Immunology 144, 365–373. doi: 10.1111/imm.12443
Bermingham, D., and Blakely, R. (2016). Kinase-dependent regulation of monoamine neurotransmitter transporters. Pharmacol. Rev. 68, 888–953. doi: 10.1124/pr.115.012260
Biederman, J. (2004). Impact of comorbidity in adults with attention-deficit/hyperactivity disorder. J. Clin. Psychiatry 65, 3–7.
Black, C., Bot, M., Scheffer, P., Cuijpers, P., and Penninx, B. (2015). Is depression associated with increased oxidative stress? A systematic review and meta-analysis. Psychoneuroendocrinology 51, 164–175. doi: 10.1016/j.psyneuen.2014.09.025
Bleck, J., DeBate, R., and Olivardia, R. (2015). The comorbidity of ADHD and eating disorders in a nationally representative sample. J. Behav. Heal. Serv. Res. 42, 437–451. doi: 10.1007/s11414-014-9422-y
Bobbo, V., Jara, C., Mendes, N., Morari, J., Velloso, L., and Araújo, E. (2019). Interleukin-6 expression by hypothalamic microglia in multiple inflammatory contexts: a systematic review. Biomed. Res. Int. 2019, 1365210. doi: 10.1155/2019/1365210
Boellner, S., Stark, J., Krishnan, S., and Zhang, Y. (2010). Pharmacokinetics of lisdexamfetamine dimesylate and its active metabolite, d-amphetamine, with increasing oral doses of lisdexamfetamine dimesylate in children with attention-deficit/hyperactivity disorder: a single-dose, randomized, open-label, crossover. Clin. Ther. 32, 252–264. doi: 10.1016/j.clinthera.2010.02.011
Brochot, C., and Quindroit, P. (2018). “Modelling the fate of chemicals in humans using a lifetime physiologically based pharmacokinetic (PBPK) model in MERLIN-Expo,” in Modelling the fate of chemicals in the environment and the human body. The handbook of environmental chemistry, eds P. Ciffroy, A. Tediosi, and E. Capri (Cham: Springer), 215–257. doi: 10.1007/978-3-319-59502-3_10
Bundesinstitut für Arzeimittel und Medizinprodukte (2020). Decentralised procedure. Public assessment report. Medikinet 5, 10, 20 mg. Medikinet retard 5, 10, 20, 30, 40 mg. Methylphenidate hydrochloride. Available online at: https://www.geneesmiddeleninformatiebank.nl/pars/h34026.pdf (accessed November 21, 2021).
Carcereny, E., Fernández-nistal, A., López, A., Montoto, C., Naves, A., Segú-Vergés, C., et al. (2021). Head to head evaluation of second generation ALK inhibitors brigatinib and alectinib as first-line treatment for ALK + NSCLC using an in silico systems biology-based approach. Oncotarget 12, 316–332. doi: 10.18632/oncotarget.27875
Caye, A., Swanson, J., Coghill, D., and Rohde, L. (2019). Treatment strategies for ADHD: an evidence-based guide to select optimal treatment. Mol. Psychiatry 24, 390–408. doi: 10.1038/s41380-018-0116-3
Ceccarini, M., Tasegian, A., Franzago, M., Patria, F., Albi, E., Codini, M., et al. (2020). 5-HT2AR and BDNF gene variants in eating disorders susceptibility. Am. J. Med. Genet. B Neuropsychiatr. Genet. 183, 155–163. doi: 10.1002/ajmg.b.32771
Cheney, M., Chen, M., Zhang, D., Phillips, J., Loffredo, M., and D’Amico, A. V. (2014). Greatest percentage of involved core length and the risk of death from prostate cancer in men with highest Gleason score = 7. Clin. Genitourin Cancer 12, 234–240. doi: 10.1016/j.clgc.2014.01.006
Ciffroy, P., Alfonso, B., Altenpohl, A., Banjac, Z., Bierkens, J., Brochot, C., et al. (2016). Modelling the exposure to chemicals for risk assessment: a comprehensive library of multimedia and PBPK models for integration, prediction, uncertainty and sensitivity analysis - the MERLIN-Expo tool. Sci. Total Environ. 568, 770–784. doi: 10.1016/j.scitotenv.2016.03.191
Coelho, L., Barbosa, D., Rizzutti, S., Bueno, O., and Miranda, M. (2017). Group cognitive behavioral therapy for children and adolescents with ADHD. Psicol. Reflex Crit. 30:11. doi: 10.1186/s41155-017-0063-y
Coghill, D., Banaschewski, T., Lecendreux, M., Soutullo, C., Johnson, M., Zuddas, A., et al. (2013). European, randomized, phase 3 study of lisdexamfetamine dimesylate in children and adolescents with attention-deficit/hyperactivity disorder. Eur. Neuropsychopharmacol. 23, 1208–1218. doi: 10.1016/j.euroneuro.2012.11.012
Corp, S., Gitlin, M., and Altshuler, L. L. A. (2014). Review of the use of stimulants and stimulant alternatives in treating bipolar depression and major depressive disorder. J. Clin. Psychiatry 75, 1010–1018.
Cortese, S., Dalla Bernardina, B., and Mouren, M. (2007). Attention-Deficit / Hyperactivity Disorder (ADHD) and binge eating. Nutr. Rev. 65, 404–411. doi: 10.1111/j.1753-4887.2007.tb00318.x
Cuffe, S., Visser, S., Holbrook, J., Danielson, M., Geryk, L., Wolraich, M., et al. (2020). ADHD and psychiatric comorbidity: functional outcomes in a school-based sample of children. J. Atten. Disord. 24, 1345–1354. doi: 10.1177/1087054715613437
Cui, W., Ning, Y., Hong, W., Wang, J., Liu, Z., and Li, M. (2019). Crosstalk between inflammation and glutamate system in depression: signaling pathway and molecular biomarkers for Ketamine’s antidepressant effect. Mol. Neurobiol. 56, 3484–3500. doi: 10.1007/s12035-018-1306-3
Dantzer, R., and Walker, A. (2014). Is there a role for glutamate-mediated excitotoxicity in inflammation-induced depression? J. Neural Transm. 121, 925–932. doi: 10.1007/s00702-014-1187-1
Davis, J., Kumbale, C., Zhang, Q., and Voit, E. (2019). Dynamical systems approaches to personalized medicine. Curr. Opin. Biotechnol. 58, 168–174. doi: 10.1016/j.copbio.2019.03.005
de Onis, M., Onyango, A., Borghi, E., Siyam, A., Nishida, C., and Siekmann, J. (2007). Development of a WHO growth reference for school-aged children and adolescents. Bull. World Health Organ. 85, 660–667. doi: 10.2471/blt.07.043497
Dew, R., and Kollins, S. (2010). Lisdexamfetamine dimesylate: a new option in stimulant treatment for ADHD. Expert Opin. Pharmacother. 11, 2907–2913. doi: 10.1517/14656566.2010.531009
Dreyfuss, M., Riegel, M., Pedersen, G., Cohen, A., Silverman, M., Dyke, J., et al. (2017). Patients with bulimia nervosa do not show typical neurodevelopment of cognitive control under emotional influences. Psychiatry Res. Neuroimaging 266, 59–65. doi: 10.1016/j.pscychresns.2017.05.001
Ekstrand, E., Murphy, H., and Wideman, C. (2019). The effects of the prodrug Vyvanse on spatial working memory and adiposity in rats. Pharmacol. Biochem. Behav. 186, 172765. doi: 10.1016/j.pbb.2019.172765
Faraone, S. V., and Larsson, H. (2019). Genetics of attention deficit hyperactivity disorder. Mol. Psychiatry 24, 562–575. doi: 10.1038/s41380-018-0070-0
Faraone, S. V., and Mick, E. (2010). Molecular genetics of attention deficit hyperactivity disorder. Psychiatr. Clin. North Am. 33, 159–180. doi: 10.1016/j.psc.2009.12.004
Fayyad, J., De Graaf, R., Kessler, R., Alonso, J., Angermeyer, M., Demyttenaere, K., et al. (2007). Cross-national prevalence and correlates of adult attention-deficit hyperactivity disorder. Br. J. Psychiatry 190, 402–409. doi: 10.1192/bjp.bp.106.034389
Francisco, A., Lethbridge, G., Patterson, B., Goldman Bergmann, C., and Van Ameringen, M. (2023). Cannabis use in attention – deficit/hyperactivity disorder (ADHD): a scoping review. J. Psychiatr. Res. 157, 239–256. doi: 10.1016/j.jpsychires.2022.11.029
Franke, B., Faraone, S. V., Asherson, P., Buitelaar, J., Bau, C., Ramos-Quiroga, J., et al. (2012). The genetics of attention deficit/hyperactivity disorder in adults, a review. Mol. Psychiatry 17, 960–987. doi: 10.1038/mp.2011.138
Garner, K., Amin, R., Johnson, R., Scarlett, E., and Burton, M. (2018). Microglia priming by interleukin-6 signaling is enhanced in aged mice. J. Neuroimmunol. 324, 90–99. doi: 10.1016/j.jneuroim.2018.09.002
Gaweska, H., and Fitzpatrick, P. (2011). Structures and mechanism of the monoamine oxidase family. Biomol. Concepts 2, 365–377. doi: 10.1515/BMC.2011.030
Ghitza, U., Gray, S., Epstein, D., Rice, K., and Shaham, Y. (2006). The anxiogenic drug yohimbine reinstates palatable food seeking in a rat relapse model: a role of CRF1 receptors. Neuropsychopharmacology 31, 2188–2196. doi: 10.1038/sj.npp.1300964
Giacobbe, P., Rakita, U., Lam, R., Milev, R., Kennedy, S., and McIntyre, R. (2018). Efficacy and tolerability of lisdexamfetamine as an antidepressant augmentation strategy: a meta-analysis of randomized controlled trials. J. Affect. Disord. 226, 294–300. doi: 10.1016/j.jad.2017.09.041
Gillberg, C., Gillberg, I., Rasmussen, P., Kadesjö, B., Söderström, H., Råstam, M., et al. (2004). Co-existing disorders in ADHD - Implications for diagnosis and intervention. Eur. Child Adolesc. Psychiatry 13, i80–i93. doi: 10.1007/s00787-004-1008-4
Ginsberg, L., Katic, A., Adeyi, B., Dirks, B., Babcock, T., Lasser, R., et al. (2011). Long-term treatment outcomes with lisdexamfetamine dimesylate for adults with attention-deficit/hyperactivity disorder stratified by baseline severity. Curr. Med. Res. Opin. 27, 1097–1107. doi: 10.1185/03007995.2011.567256
Guerdjikova, A., Blom, T., Mori, N., Matthews, A., Cummings, T., Casuto, L., et al. (2019). Lisdexamfetamine in pediatric binge eating disorder: a retrospective chart review. Clin. Neuropharmacol. 42, 214–216. doi: 10.1097/WNF.0000000000000367
Guerdjikova, A., Mori, N., Casuto, L., and McElroy, S. (2016). Novel pharmacologic treatment in acute binge eating disorder - role of lisdexamfetamine. Neuropsychiatry Dis. Treat. 12, 833–841. doi: 10.2147/NDT.S80881
Gutiérrez-Casares, J., Quintero, J., Jorba, G., Junet, V., Martínez, V., Pozo-Rubio, T., et al. (2021). Methods to develop an in silico clinical trial: computational head-to-head comparison of lisdexamfetamine and methylphenidate. Front. Psychiatry 12:741170. doi: 10.3389/fpsyt.2021.741170
Gutierrez-Casares, J., Quintero, J., Segú-Vergés, C., Rodríguez, P., Pozo-Rubio, T., Coma, M., et al. (2023). In silico clinical trial evaluating lisdexamfetamine’s and methylphenidate’s mechanism of action computational models in an attention-deficit/hyperactivity disorder virtual patients’ population. Front. Psychiatry 14:939650. doi: 10.3389/fpsyt.2023.939650
Han, H., Cho, J., Lee, S., Yun, A., Kim, H., Bae, D., et al. (2018). TRRUST v2: an expanded reference database of human and mouse transcriptional regulatory interactions. Nucleic Acids Res. 46, D380–D386. doi: 10.1093/nar/gkx1013
Howard, A., Kennedy, T., Mitchell, J., Sibley, M., Hinshaw, S., Arnold, L., et al. (2020). Early substance use in the pathway from childhood attention-deficit/hyperactivity disorder (ADHD) to young adult substance use: evidence of statistical mediation and substance specificity. Psychol. Addict. Behav. 34, 281–292. doi: 10.1037/adb0000542
Hudson, J., McElroy, S., Ferreira-Cornwell, M., Radewonuk, J., and Gasior, M. (2017). Efficacy of lisdexamfetamine in adults with moderate to severe binge-eating disorder: a randomized clinical trial. JAMA Psychiatry 74, 903–910. doi: 10.1001/jamapsychiatry.2017.1889
Hutson, P., Pennick, M., and Secker, R. (2014). Preclinical pharmacokinetics, pharmacology and toxicology of lisdexamfetamine: a novel d-amphetamine pro-drug. Neuropharmacology 87, 41–50. doi: 10.1016/j.neuropharm.2014.02.014
Iemolo, A., Blasio, A., St Cyr, S., Jiang, F., Rice, K., Sabino, V., et al. (2013). CRF-CRF 1 receptor system in the central and basolateral nuclei of the amygdala differentially mediates excessive eating of palatable food. Neuropsychopharmacology 38, 2456–2466. doi: 10.1038/npp.2013.147
Inserra, A., Mastronardi, C., Rogers, G., Licinio, J., and Wong, M. (2019). Neuroimmunomodulation in major depressive disorder: focus on caspase 1, inducible nitric oxide synthase, and interferon-gamma. Mol. Neurobiol. 56, 4288–4305. doi: 10.1007/s12035-018-1359-3
Iob, E., Kirschbaum, C., and Steptoe, A. (2020). Persistent depressive symptoms, HPA-axis hyperactivity, and inflammation: the role of cognitive-affective and somatic symptoms. Mol. Psychiatry 25, 1130–1140. doi: 10.1038/s41380-019-0501-6
Jassal, B., Matthews, L., Viteri, G., Gong, C., Lorente, P., Fabregat, A., et al. (2020). The reactome pathway knowledgebase. Nucleic Acids Res. 48, D498–D503. doi: 10.1093/nar/gkz1031
Joelsson, P., Chudal, R., Gyllenberg, D., Kesti, A., Hinkka-Yli-Salomäki, S., Virtanen, J., et al. (2016). Demographic characteristics and psychiatric comorbidity of children and adolescents diagnosed with ADHD in specialized healthcare. Child. Psychiatry Hum. Dev. 47, 574–582. doi: 10.1007/s10578-015-0591-6
Jorba, G., Aguirre-Plans, J., Junet, V., Segu-Verges, C., Ruiz, J., Pujol, A., et al. (2020). In-silico simulated prototype-patients using TPMS technology to study a potential adverse effect of sacubitril and valsartan. PLoS One 15:e0228926. doi: 10.1371/journal.pone.0228926
Juhasz, G., Dunham, J., McKie, S., Thomas, E., Downey, D., Chase, D., et al. (2011). The CREB1-BDNF-NTRK2 pathway in depression: multiple gene-cognition- environment interactions. Biol. Psychiatry 69, 762–771. doi: 10.1016/j.biopsych.2010.11.019
Kanehisa, M., Furumichi, M., Tanabe, M., Sato, Y., and Morishima, K. (2017). KEGG: new perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 45, D353–D361. doi: 10.1093/nar/gkw1092
Katrin, F., Pannasch, U., and Kettenmann, H. (2005). Dopamine and noradrenaline control distinct functions in rodent microglial cells. Mol. Cell Neurosci. 29, 128–138. doi: 10.1016/j.mcn.2005.01.003
Katzman, M., Bilkey, T., Chokka, P., Fallu, A., and Klassen, L. (2017). Adult ADHD and comorbid disorders: clinical implications of a dimensional approach. BMC Psychiatry 17:302. doi: 10.1186/s12888-017-1463-3
Kegg Pathway Database (2020). Dopaminergic synapse - Homo sapiens (human). Available online at: https://www.genome.jp/kegg-bin/show_pathway?hsa04728 (accessed November 21, 2021).
Keller, J., Gomez, R., Williams, G., Lembke, A., Lazzeroni, L., Murphy, G., et al. (2017). HPA axis in major depression: cortisol, clinical symptomatology and genetic variation predict cognition. Mol. Psychiatry 22, 527–536. doi: 10.1038/mp.2016.120
Keshava Prasad, T., Goel, R., Kandasamy, K., Keerthikumar, S., Kumar, S., Mathivanan, S., et al. (2009). Human protein reference database - 2009 update. Nucleic Acids Res. 37, D767–D772. doi: 10.1093/nar/gkn892
Kessler, R., Adler, L., Barkley, R., Biederman, J., Conners, C., Demler, O., et al. (2006). The prevalence and correlates of adult ADHD in the United States: results from the national comorbidity survey replication. Am. J. Psychiatry 163, 716–723. doi: 10.1176/ajp.2006.163.4.716
Kittel-Schneider, S., and Reif, A. (2020). Adulte aufmerksamkeitsdefizit-/hyperaktivitätsstörung und komorbidität: neue befunde zu epidemiologischen und genetischen faktoren [Adult attention deficit hyperactivity disorder and comorbidity: new findings on epidemiological and genetic factors]. Nervenarzt 91, 575–582. doi: 10.1007/s00115-020-00900-5
Kollins, S., Youcha, S., Lasser, R., and Thase, M. (2011). Lisdexamfetamine dimesylate for the treatment of attention deficit hyperactivity disorder in adults with a history of depression or history of substance use disorder. Innov. Clin. Neurosci. 8, 28–32.
Krishnan, S., and Zhang, Y. (2008). Relative bioavailability of lisdexamfetamine 70-mg capsules in fasted and fed healthy adult volunteers and in solution: a single-dose, crossover pharmacokinetic study. J. Clin. Pharmacol. 48, 293–302. doi: 10.1177/0091270007310381
Laas, K., Kiive, E., Mäestu, J., Vaht, M., Veidebaum, T., and Harro, J. (2017). Nice guys: homozygocity for the TPH2 -703G/T (rs4570625) minor allele promotes low aggressiveness and low anxiety. J. Affect. Disord. 215, 230–236. doi: 10.1016/j.jad.2017.03.045
Larson, K., Russ, S., Kahn, R., and Halfon, N. (2011). Patterns of comorbidity, functioning, and service use for US children with ADHD, 2007. Pediatrics 127, 462–470. doi: 10.1542/peds.2010-0165
Lee, S., Humphreys, K., Flory, K., Liu, R., and Glass, K. (2011). Prospective association of childhood attention-deficit/hyperactivity disorder (ADHD) and substance use and abuse/dependence: a meta-analytic review. Clin. Psychol. Rev. 31, 328–341. doi: 10.1016/j.cpr.2011.01.006
Lee, S., Ripke, S., Neale, B., Faraone, S. V., Purcell, S., Perlis, R., et al. (2013). Genetic relationship between five psychiatric disorders estimated from genome-wide SNPs. Nat. Genet. 45, 984–994. doi: 10.1038/ng.2711
Lindqvist, D., Dhabhar, F., James, S., Hough, C., Jain, F., Bersani, F., et al. (2017). Oxidative stress, inflammation and treatment response in major depression. Psychoneuroendocrinology 76, 197–205. doi: 10.1016/j.psyneuen.2016.11.031
Liu, T., Zhong, S., Liao, X., Chen, J., He, T., Lai, S., et al. (2015). A meta-analysis of oxidative stress markers in depression. PLoS One 10:e0138904. doi: 10.1371/journal.pone.0138904
Liu, X., and Quan, N. (2018). Microglia and CNS interleukin-1: beyond immunological concepts. Front. Neurol. 9:8. doi: 10.3389/fneur.2018.00008
Lorén, V., Garcia-Jaraquemada, A., Naves, J., Carmona, X., Mañosa, M., Aransay, A., et al. (2019). ANP32E, a protein involved in steroid-refractoriness in ulcerative colitis, identified by a systems biology approach. J. Crohns Colitis 13, 351–361. doi: 10.1093/ecco-jcc/jjy171
Madhoo, M., Keefe, R., Roth, R., Sambunaris, A., Wu, J., Trivedi, M., et al. (2014). Lisdexamfetamine dimesylate augmentation in adults with persistent executive dysfunction after partial or full remission of major depressive disorder. Neuropsychopharmacology 39, 1388–1398. doi: 10.1038/npp.2013.334
Maldonado, R. (2013). Comparison of the pharmacokinetics and clinical efficacy of new extended-release formulations of methylphenidate. Expert Opin. Drug Metab. Toxicol. 9, 1001–1014. doi: 10.1517/17425255.2013.786041
Mannuzza, S., Klein, R., Bessler, A., Malloy, P., and LaPadula, M. (1998). Adult psychiatric status of hyperactive boys grown up. Am. J. Psychiatry 155, 493–498. doi: 10.1176/ajp.155.4.493
McElroy, S., Hudson, J., Ferreira-Cornwell, M., Radewonuk, J., Whitaker, T., and Gasior, M. (2016). Lisdexamfetamine dimesylate for adults with moderate to severe binge eating disorder: results of two pivotal phase 3 randomized controlled trials. Neuropsychopharmacology 41, 1251–1260. doi: 10.1038/npp.2015.275
McGough, J., and Barkley, R. (2004). Diagnostic controversies in adult attention deficit hyperactivity disorder. Am. J. Psychiatry 161, 1948–1956. doi: 10.1176/appi.ajp.161.11.1948
Menke, A. (2019). Is the HPA axis as target for depression outdated, or is there a new hope? Front. Psychiatry 10:101. doi: 10.3389/fpsyt.2019.00101
Micioni Di Bonaventura, M., Micioni Di Bonaventura, E., Botticelli, L., and Cifani, C. (2021). “Impact of a history of caloric restriction and a frustration stress manipulation on binge-like eating behavior in female rats: preclinical results,” in Animal models of eating disorders, ed. N. M. Avena (New York, NY: Humana Press), 239–260. doi: 10.1007/978-1-0716-0924-8_13
Micioni Di Bonaventura, M., Micioni Di Bonaventura, E., Polidori, C., and Cifani, C. (2020). “Preclinical models of stress and environmental influences on binge eating,” in Binge eating, eds G. Frank and L. Berner (Cham: Springer), 85–101. doi: 10.1007/978-3-030-43562-2_7
Micioni Di Bonaventura, M., Ubaldi, M., Giusepponi, M., Rice, K., Massi, M., Ciccocioppo, R., et al. (2017). Hypothalamic CRF1 receptor mechanisms are not sufficient to account for binge-like palatable food consumption in female rats. Int. J. Eat. Disord. 50, 1194–1204. doi: 10.1002/eat.22767
Miller, A., and Raison, C. (2016). The role of inflammation in depression: from evolutionary imperative to modern treatment target. Nat. Rev. Immunol. 16, 22–34. doi: 10.1038/nri.2015.5
Mitchell, N., and Baker, G. (2010). An update on the role of glutamate in the pathophysiology of depression. Acta Psychiatr. Scand. 122, 192–210. doi: 10.1111/j.1600-0447.2009.01529.x
Moore, C., Sabino, V., and Cottone, P. (2018). Trace amine associated receptor 1 (TAAR1) modulation of food reward. Front. Pharmacol. 9:129. doi: 10.3389/fphar.2018.00129
Ohnishi, T., Kobayashi, H., Yajima, T., Koyama, T., and Noguchi, K. (2019). Psychiatric comorbidities in adult attention-deficit/hyperactivity disorder: prevalence and patterns in the routine clinical setting. Innov. Clin. Neurosci. 16, 11–16.
Orchard, S., Ammari, M., Aranda, B., Breuza, L., Briganti, L., Broackes-Carter, F., et al. (2014). The MIntAct project–IntAct as a common curation platform for 11 molecular interaction databases. Nucleic Acids Res. 42, D358–D363. doi: 10.1093/nar/gkt1115
Oughtred, R., Stark, C., Breitkreutz, B., Rust, J., Boucher, L., Chang, C., et al. (2019). The BioGRID interaction database: 2019 update. Nucleic Acids Res. 47, D529–D541. doi: 10.1093/nar/gky1079
Pan, M., Huang, F., Zhao, M., Wang, Y., Wang, Y., and Qian, Q. J. (2019). A comparison of efficacy between cognitive behavioral therapy (CBT) and CBT combined with medication in adults with attention-deficit/hyperactivity disorder (ADHD). Psychiatry Res. 279, 23–33. doi: 10.1016/j.psychres.2019.06.040
Pariante, C., and Lightman, S. (2008). The HPA axis in major depression: classical theories and new developments. Trends Neurosci. 31, 464–468. doi: 10.1016/j.tins.2008.06.006
Peters, S. (2008). Evaluation of a generic physiologically based pharmacokinetic model for lineshape analysis. Clin. Pharmacokinet. 47, 261–275. doi: 10.2165/00003088-200847040-00004
Peterson, S., Pack, T., Wilkins, A., Urs, N., Urban, D., Bass, C., et al. (2015). Elucidation of G-protein and beta-arrestin functional selectivity at the dopamine D2 receptor. Proc. Natl. Acad. Sci. U.S.A. 112, 7097–7102. doi: 10.1073/pnas.1502742112
Piñeiro-Dieguez, B., Balanzá-Martínez, V., García-García, P., Soler-López, B., Domingo, M., Labarra, J., et al. (2016). Psychiatric comorbidity at the time of diagnosis in adults with ADHD: the CAT study. J. Atten. Disord. 20, 1066–1075. doi: 10.1177/1087054713518240
Pitsillou, E., Bresnehan, S., Kagarakis, E., Wijoyo, S., Liang, J., Hung, A., et al. (2020). The cellular and molecular basis of major depressive disorder: towards a unified model for understanding clinical depression. Mol. Biol. Rep. 47, 753–770. doi: 10.1007/s11033-019-05129-3
Popovic, B., Bhattacharya, P., and Sivaswamy, L. (2009). Lisdexamfetamine: a prodrug for the treatment of attention-deficit/hyperactivity disorder. Am. J. Heal. Pharm. 66, 2005–2012. doi: 10.2146/ajhp080353
Rankin, J., Matthews, L., Cobley, S., Han, A., Sanders, R., Wiltshire, H., et al. (2016). Psychological consequences of childhood obesity: psychiatric comorbidity and prevention. Adolesc. Health Med. Ther. 7, 125–146. doi: 10.2147/AHMT.S101631
Retz, W., Rösler, M., Ose, C., Scherag, A., Alm, B., Philipsen, A., et al. (2012). Multiscale assessment of treatment efficacy in adults with ADHD: a randomized placebo-controlled, multi-centre study with extended-release methylphenidate. World J. Biol. Psychyatry 13, 48–59. doi: 10.3109/15622975.2010.540257
Roncero, C., and Álvarez, F. (2014). The use of lisdexamfetamine dimesylate for the treatment of ADHD and other psychiatric disorders. Expert Rev. Neurother. 14, 849–865. doi: 10.1586/14737175.2014.932691
Rostami-Hodjegan, A. (2012). Physiologically based pharmacokinetics joined with in vitro-in vivo extrapolation of ADME: a marriage under the arch of systems pharmacology. Clin. Pharmacol. Ther. 92, 50–61. doi: 10.1038/clpt.2012.65
Schmid, T., Snoek, L., Frohli, E., van der Bent, M., Kammenga, J., and Hajnal, A. (2015). Systemic regulation of RAS/MAPK signaling by the serotonin metabolite 5-HIAA. PLoS Genet. 11:e1005236. doi: 10.1371/journal.pgen.1005236
Schneider, E., Higgs, S., Dourish, C., and Park, H. (2021). Lisdexamfetamine and binge-eating disorder: a systematic review and meta-analysis of the preclinical and clinical data with a focus on mechanism of drug action in treating the disorder. Eur. Neuropsychopharmacol. 53, 49–78. doi: 10.1016/j.euroneuro.2021.08.001
Segú-Vergés, C., Coma, M., Kessel, C., Smeets, S., Foell, D., and Aldea, A. (2021). Application of systems biology-based in silico tools to optimize treatment strategy identification in Still’s disease. Arthritis Res. Ther. 23:126. doi: 10.1186/s13075-021-02507-w
Shannon, P., Markiel, A., Ozier, O., Baliga, N., Wang, J., Ramage, D., et al. (2003). Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504. doi: 10.1101/gr.1239303
Sobanski, E. (2006). Psychiatric comorbidity in adults with attention-deficit/hyperactivity disorder (ADHD). Eur. Arch. Psychiatry Clin. Neurosci. 256, i26–i31. doi: 10.1007/s00406-006-1004-4
Sobanski, E., Brüggemann, D., Alm, B., Kern, S., Deschner, M., Schubert, T., et al. (2007). Psychiatric comorbidity and functional impairment in a clinically referred sample of adults with attention-deficit/hyperactivity disorder (ADHD). Eur. Arch. Psychiatry Clin. Neurosci. 257, 371–377. doi: 10.1007/s00406-007-0712-8
Song, M., Martinowich, K., and Lee, F. S. (2017). BDNF at the synapse: why location matters. Mol. Psychiatry 22, 1370–1375. doi: 10.1038/mp.2017.144
Sonuga-Barke, E., and Halperin, J. (2010). Developmental phenotypes and causal pathways in attention deficithyperactivity disorder: potential targets for early intervention? J. Child. Psychol. Psychiatry Allied Discip. 51, 368–389. doi: 10.1111/j.1469-7610.2009.02195.x
Sotnikova, T., Caron, M., and Gainetdinov, R. (2009). Trace amine-associated receptors as emerging therapeutic targets. Mol. Pharmacol. 76, 229–235. doi: 10.1124/mol.109.055970
Srivastava, G., O’hara, V., and Browne, N. (2019). Use of lisdexamfetamine to treat obesity in an adolescent with severe obesity and binge eating. Children 6:22. doi: 10.3390/children6020022
Strajhar, P., Vizeli, P., Patt, M., Dolder, P., Kratschmar, D. V., Liechti, M., et al. (2019). Effects of lisdexamfetamine on plasma steroid concentrations compared with d-amphetamine in healthy subjects: a randomized, double-blind, placebo-controlled study. J. Steroid. Biochem. Mol. Biol. 186, 212–225. doi: 10.1016/j.jsbmb.2018.10.016
Sung, U., Apparsundaram, S., Galli, A., Kahlig, K., Savchenko, V., Schroeter, S., et al. (2003). A regulated interaction of syntaxin 1A with the antidepressant-sensitive norepinephrine transporter establishes catecholamine clearance capacity. J. Neurosci. 23, 1697–1709. doi: 10.1523/JNEUROSCI.23-05-01697.2003
Ta, T., Dikmen, H., Schilling, S., Chausse, B., Lewen, A., Hollnagel, J., et al. (2019). Priming of microglia with IFN-γ slows neuronal gamma oscillations in situ. Proc. Natl. Acad. Sci. U.S.A. 116, 4637–4642. doi: 10.1073/pnas.1813562116
The European Social Survey (2018). ESS Round 7: European Social Survey round 7 data (2014). Data file edition 2.2. NSD - Norwegian Centre for Research Data, Norway – Data Archive and distributor of ESS data for ESS ERIC. Available online at: https://www.europeansocialsurvey.org/data/download.html?r=7 (accessed November 21, 2021).
Thomas, R., Sanders, S., Doust, J., Beller, E., and Glasziou, P. (2015). Prevalence of attention-deficit/hyperactivity disorder: a systematic review and meta-analysis. Pediatrics 135, e994–e1001. doi: 10.1542/peds.2014-3482
Tsuda, M., Masuda, T., Kitano, J., Shimoyama, H., Tozaki-Saitoh, H., and Inoue, K. (2009). IFN-γ receptor signaling mediates spinal microglia activation driving neuropathic pain. Proc. Natl. Acad. Sci. U.S.A. 106, 8032–8037. doi: 10.1073/pnas.0810420106
U. S. Food and Drug Administration. (2017). VYVANSE prescribing information. Available online at: https://www.accessdata.fda.gov/drugsatfda_docs/label/2017/208510lbl.pdf (accessed November 21, 2021).
Underhill, S., Hullihen, P., Chen, J., Fenollar-Ferrer, C., Rizzo, M., Ingram, S., et al. (2021). Amphetamines signal through intracellular TAAR1 receptors coupled to Gα13 and GαS in discrete subcellular domains. Mol. Psychiatry 26, 1208–1223. doi: 10.1038/s41380-019-0469-2
van Amsterdam, J., van der Velde, B., Schulte, M., and van den Brink, W. (2018). Causal factors of increased smoking in ADHD: a systematic review. Subst. Use Misuse 53, 432–445. doi: 10.1080/10826084.2017.1334066
Vaughan, R., and Foster, J. (2013). Mechanisms of dopamine transporter regulation in normal and disease states. Trends Pharmacol. Sci. 34, 489–496. doi: 10.1016/j.tips.2013.07.005
Vidal, P., and Pacheco, R. (2020). The cross-talk between the dopaminergic and the immune system involved in schizophrenia. Front. Pharmacol. 11:394. doi: 10.3389/fphar.2020.00394
Welser-Alves, J. V., and Milner, R. (2013). Microglia are the major source of TNF-α and TGF-β in postnatal glial cultures; regulation by cytokines, lipopolysaccharide, and vitronectin. Neurochem. Int. 63, 47–53. doi: 10.1016/j.neuint.2013.04.007
Wilens, T., Faraone, S. V., and Biederman, J. (2004). Attention-deficit/hyperactivity disorder in adults. JAMA 292, 619–623. doi: 10.1001/jama.292.5.619
Xie, Z., and Miller, G. (2007). Trace amine-associated receptor 1 is a modulator of the dopamine transporter. J. Pharmacol. Exp. Ther. 321, 128–136. doi: 10.1124/jpet.106.117382
Xue, W., Wang, W., Gong, T., Zhang, H., Tao, W., Xue, L., et al. (2016). PKA-CREB-BDNF signaling regulated long lasting antidepressant activities of Yueju but not ketamine. Sci. Rep. 6:26331. doi: 10.1038/srep26331
Keywords: attention-deficit/hyperactivity disorder, lisdexamfetamine, mathematical modeling, binge eating disorder, depression
Citation: Gutiérrez-Casares JR, Segú-Vergés C, Sabate Chueca J, Pozo-Rubio T, Coma M, Montoto C and Quintero J (2023) In silico evaluation of the role of lisdexamfetamine on attention-deficit/hyperactivity disorder common psychiatric comorbidities: mechanistic insights on binge eating disorder and depression. Front. Neurosci. 17:1118253. doi: 10.3389/fnins.2023.1118253
Received: 07 December 2022; Accepted: 12 June 2023;
Published: 30 June 2023.
Edited by:
Jacob Raber, Oregon Health and Science University, United StatesReviewed by:
Simon Weissenberger, First Medical Faculty Charles University, CzechiaCopyright © 2023 Gutiérrez-Casares, Segú-Vergés, Sabate Chueca, Pozo-Rubio, Coma, Montoto and Quintero. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José Ramón Gutiérrez-Casares, anJndXRpZXJyZXpjYXNhcmVzQGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.