
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Neurosci., 23 September 2022
Sec. Neural Technology
Volume 16 - 2022 | https://doi.org/10.3389/fnins.2022.966772
 Hanci Zhang1
Hanci Zhang1 Hui Fang1Deqiang Liu1Yiming Zhang1
Hui Fang1Deqiang Liu1Yiming Zhang1 Joseph Adu-Amankwaah2
Joseph Adu-Amankwaah2 Jinxiang Yuan1,3,4*
Jinxiang Yuan1,3,4* Rubin Tan2*Jianping Zhu1*
Rubin Tan2*Jianping Zhu1*Optogenetics is an emerging bioengineering technology that has been rapidly developed in recent years by cross-integrating optics, genetic engineering, electrophysiology, software control, and other disciplines. Since the first demonstration of the millisecond neuromodulation ability of the channelrhodopsin-2 (ChR2), the application of optogenetic technology in basic life science research has been rapidly progressed, especially in neurobiology, which has driven the development of the discipline. As the optogenetic tool protein, microbial rhodopsins have been continuously explored, modified, and optimized, with many variants becoming available, with structural characteristics and functions that are highly diversified. Their applicability has been broadened, encouraging more researchers and clinicians to utilize optogenetics technology in research. In this review, we summarize the species and variant types of the most important class of tool proteins in optogenetic techniques, the microbial rhodopsins, and review the current applications of optogenetics based on rhodopsin qualitative light in biology and other fields. We also review the challenges facing this technology, to ultimately provide an in-depth technical reference to support the application of optogenetics in translational and clinical research.
Optogenetics is a technique that integrates the fields of optics, genetic engineering, electrophysiology, and software control to regulate specific activity in cells with the help of various light-responsive proteins (Kim et al., 2017). Compared with chemical molecules and electrical signals, optical signals used in optogenetics have higher temporal and spatial resolution and cause lower damage to cells. As such, they have been widely used in basic research such as neurobiology, cell biology, and molecular biology (Kolesov et al., 2021).
Light-responsive proteins are a class of proteins commonly found in nature that can perform specific functions in response to stimulation by specific light wavelengths. For instance, they can mediate ion transmembrane transport, gene expression, and cell signal transduction (Govorunova et al., 2017a; Duan et al., 2022). Through genetic engineering optimization of optogenetics, a series of optogenetic tool proteins have been successfully developed. Microbial rhodopsins are one of the most widely used. At the beginning of this century, channelrhodopsins (ChRs) were first evaluated and functionally identified, opening the door to optogenetics research (Nagel et al., 2002, 2003). Boyden et al. used channelrhodopsins-2 (ChR2) to successfully modulate the activity state of neurons in the isolated hippocampus, thereby ushering in the Optogenetics 1.0 era of neurobiological research (Boyden et al., 2005; Nagel et al., 2005; Bi et al., 2006). With the construction of additional subtypes and variants of light-responsive proteins, the scope of application of optogenetics has been greatly expanded, including in a wider range of organisms. For instance, the introduction of GtACR1 into the guard cells of tobacco by Huang et al. was used to show that the continuous stimulation of green light caused depolarization of the plasma membrane, which led to voltage-gated K+ channels opening, guard cell swelling, and stomatal closure; this is a prerequisite technical condition for analyzing the effect of patchy stomatal conductance on water use efficiency. This crosses the limits of plant and animal species and applies optogenetic techniques to the field of plant physiology, enabling artificial control of stomatal closure and broadening the application areas of the technology. Optogenetics technology provides a time-saving and labor-saving technical tool for the study of long-duration plant physiological phenomena (Huang et al., 2021). In a more complex biological reaction process, such as the LicV system described by Liu et al., which can be an effective and tunable optogenetic control of transcriptional and genomic locus markers is achieved (Liu et al., 2022). In subsequent studies, the sources of optogenetic tool proteins have become more diverse, such as UV-B, cryptochrome (CRY), and light-oxygen-voltage (LOV) domains derived from plant photoreceptor proteins. The implicated biological reaction processes span from the regulation of gene expression to cell signal transduction, and from macroscopic neurobiological research to microscopic molecular and cell biology research. These works have facilitated and promoted the technical research of optogenetics in the Optogenetics 2.0 era (Tan et al., 2022). The utility of optogenetics 2.0 has moved beyond ion transport control to the exploration of multiple sources of instrumental proteins, photoreceptor modification, and the use of different types of light to control more complex biochemical reactions in cells. Organisms that have been studied using optogenetics have broadened from animals to plants and microorganisms. Therefore, in addition to neuroscience, optogenetics 2.0 provides a powerful problem-solving approach for disciplines such as plant science, cell biology, and molecular biology.
Attempts have been made to merge optogenetic technologies with clinical medical research, boosting its translation into clinical research applications and even into clinical practice. To date, such research has mainly focused on cardiac pacing and heart rate regulation, visual acuity recovery, optical cochlear implant (oCls), and other fields. For example, optogenetic treatment was used to effect the partial recovery of visual function in a patient with retinitis pigmentosa, providing the first report of its use in a neurodegenerative context (Sahel et al., 2021). In this paper, we review the current variants of microbial rhodopsins, provide a comprehensive description and systematic evaluation of the potential applications of rhodopsin-based optogenetics in the biomedical field, and provide an outlook of the opportunities and challenges facing its future development. We believe that rhodopsin-based optogenetics will not only play an important leading role in basic research but also have a strong potential for biomedical and clinical applications. However, many issues need to be considered such as the safety of viral vectors, trafficking of the protein, targeting light to the protein, and the efficiency of the process. Prevailing over these challenges may allow optogenetics to leap from biomedical to clinical translational applications.
From the invention of optogenetics to the present decade, the family size of microbial rhodopsins (opsin + ovalently-bound retinal = rhodopsin) has been expanding, and its variants have been continuously enriched. Based on its structure and functional characteristics, rhodopsins can be divided into three categories: light-driven ion pump, light-gated ion channel, and light-activated signaling/enzyme rhodopsins. The emergence of new microbal rhodopsin mutants not only opens up the range of optogenetic applications in basic research but also provides a powerful weapon for using optogenetics technology to explore the pathogenic mechanisms, diagnosis, and treatment of various neurological diseases, such as neurodegenerative diseases. The classifications of the microbial rhodopsin family of proteins are shown in Figure 1.
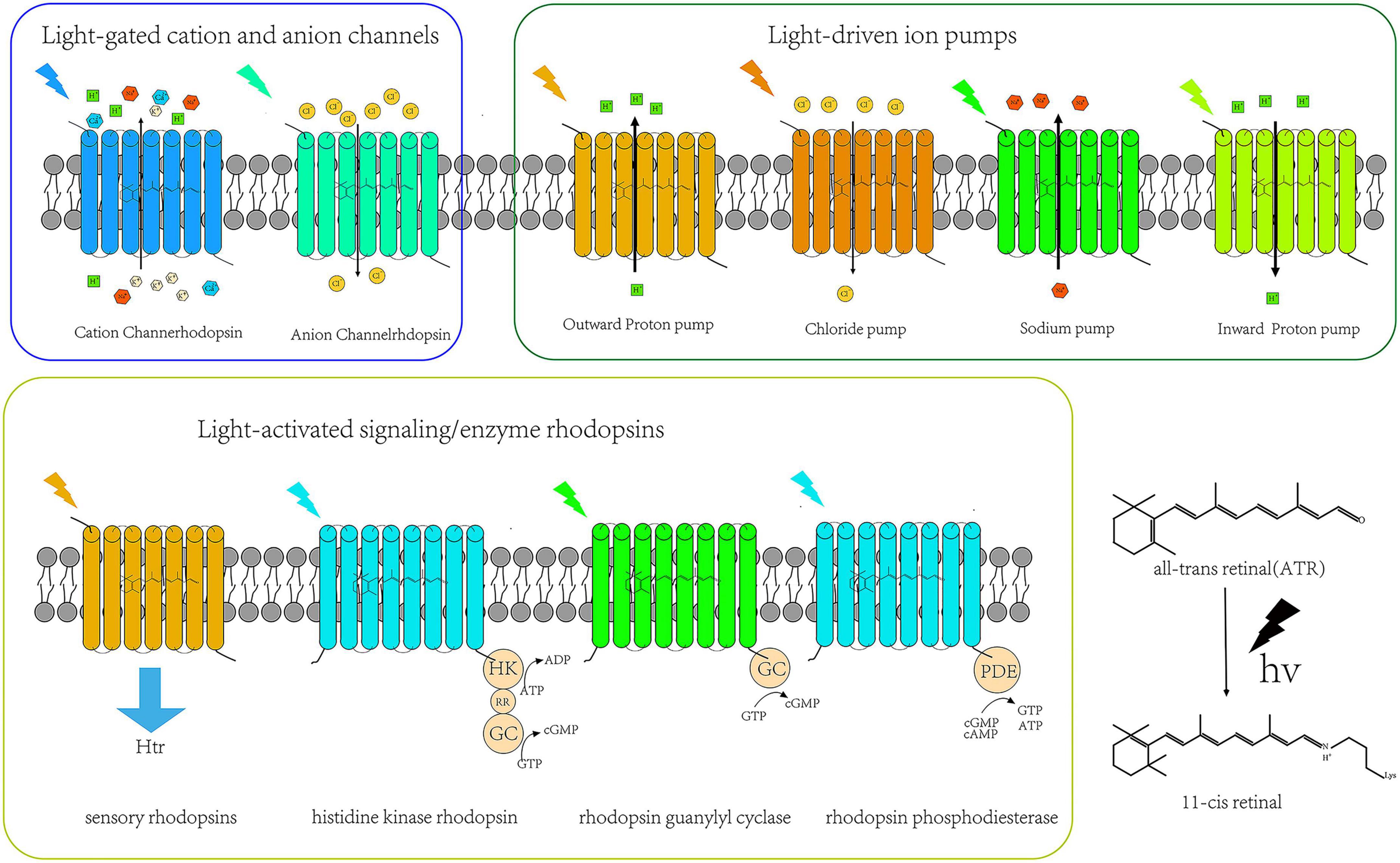
Figure 1. Types of microbial rhodopsins.
As early as 1971, researchers discovered the first microbial Bacteriorhodopsin (BR) in Halobacterium salinarum. This is a light-driven hydrogen ion pump that mediates hydrogen ion transmembrane outflow and participates in photosynthetic phosphorylation (Oesterhelt and Stoeckenius, 1971; Oesterhelt and Stoeckenius, 1973; Danon and Stoeckenius, 1974). BR provided several milestones for structural biology as the first detailed and thoroughly studied microbial rhodopsin, the first seven-transmembrane protein with a resolved molecular configuration, and the first membrane protein with a determined amino acid sequence (Khorana et al., 1979). Unfortunately, at the time, its unique transmembrane ion transport mode did not attract the attention of neurobiologists. It was only when channelrhodopsin-2 (ChR2) was discovered and started to be exploited in neurobiology that BR was gradually adopted by neuroscientists (Boyden et al., 2005). Subsequently, more hydrogen ion pumps homologous to BR were discovered, such as Arch (Chow et al., 2010) and Mac (Husson et al., 2012). Arch, a hydrogen ion pump from Halorubrum sodomense, is usually used to suppress neuronal activity, but it is more noteworthy as a genetically encoded voltage indicators (GEVIs) for the real-time imaging of brain action discharges (Kralj et al., 2011). Mac is a fungal hydrogen ion pump. Husson et al. (2012) found that Mac can control the behavior of caenorhabditis elegans by inhibiting neuronal activity. There are also many other exciting potential photogenetic tool proteins in fungi. For instance, the recently discovered fungal ApOps series of bacteriorhodopsin are also photo-controlled ion pump bacteriorhodopsins. For example, this protein can be localized on the plasma membrane of specific organelles, such as lysosomes, and green light can be used to change their internal and external acid-base environments to study the biological consequences (Panzer et al., 2021).
The second microbial rhodopsin to be discovered was the Cl– ion pump-Halorhodopsin (HR) (Matsuno-Yagi and Mukohata, 1977). In the HR family, the most mature application is NpHR, which was found in Natronomonas pharaonis. NpHR mediates the transmembrane influx of Cl– by ∼590 nm light thereby silencing neurons; this has mainly been used to explore the neural circuit architecture of animals during critical behavioral activities (Lanyi, 1986; Zhang et al., 2007; Chen et al., 2021). However, its disadvantage is that the generated photocurrents are low, which limits its effectiveness in inhibiting neuronal activity. As a result, the behavioral phenotype remains relatively unaffected thus limiting its utility in research. Subsequently, various variants have been developed based on wild-type NpHR, such as eNpHR (Gradinaru et al., 2008), eNpHR 2.0 (Gradinaru et al., 2009) and eNpHR 3.0. The modified and optimized eNpHR 3.0 protein is more highly enriched and localized on the cell membrane, rather than in the cytoplasm, which allows it to produce greater photocurrents while having little effect on the intracellular environment. This variant has been used as a neurosilencing tool in optogenetics (Gradinaru et al., 2010).
From the 1970s and over the following 40 years, no new light-driven pumps for aprotic cations were been reported, except for the chloride and hydrogen ion pump. In 2013, Inoue K et al. reported that they isolated a light-driven Na+ ion pump-KR2 from Krokinobacter eikastu (also known as DeNaR), and that KR2 can pump sodium ions out of cells to achieve neuronal silencing (Inoue et al., 2013; Hososhima et al., 2021). Later, Shevchenko V et al. described an internal flow H+ pump rhodopsin (Inoue et al., 2016; Shevchenko et al., 2017). Studies have shown that the largest advantage of proton pumps over chloride pumps is that they produce higher photocurrent rates and have less interference with γ−aminobutyric acid (GABA) energy interneurons (Chow et al., 2010). With the continuous discovery of the ion pump type of rhodopsin, the types of optogenetics tool proteins are becoming greatly enriched, making it possible for optogenetics to be applied in wider scientific research fields and more disciplines. The summary of light-driven ion pump rhodopsins are shown in Table 1.
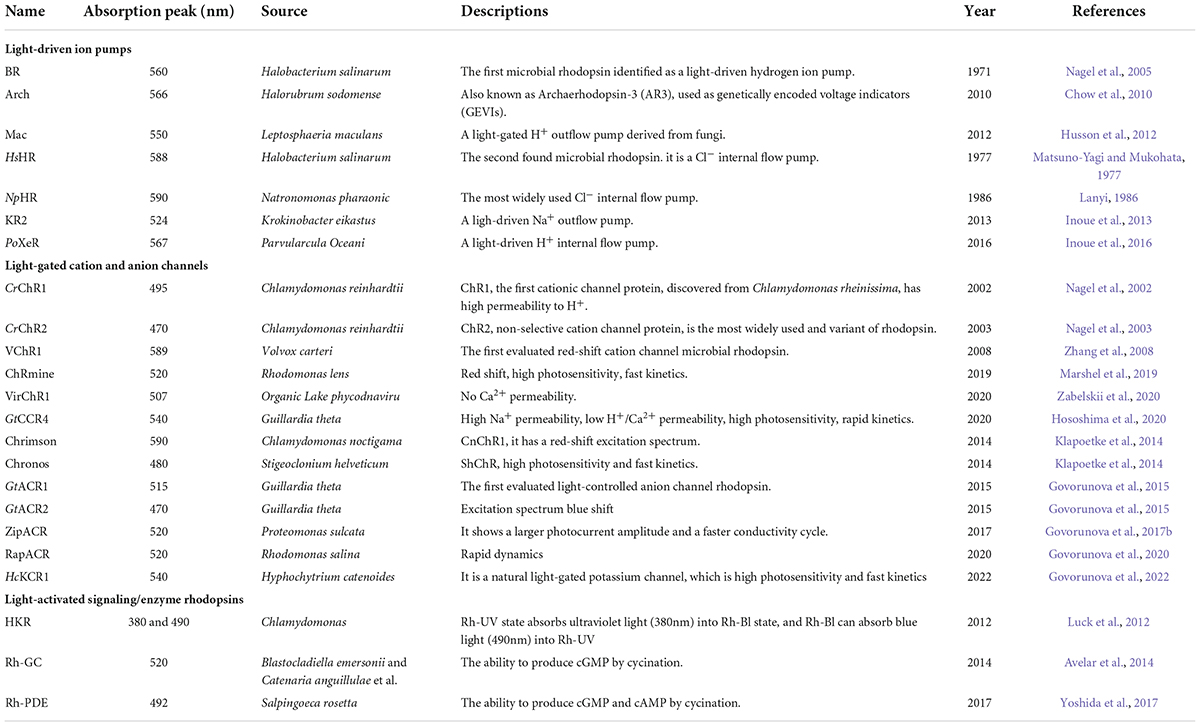
Table 1. Widely used naturally occurring microbial rhodopsins.
Channelrhodopsin-2 (ChR2) is derived from Chlamydomonas reinhardtii; this genus is the earliest and most widely used microbial rhodopsin (Boyden et al., 2005; Kim et al., 2017). ChR2 is a light-controlled non-selective cation channel protein that mediates the inward flow of Na+, K+, H+, Ca2+, and other cations across the membrane under blue light (420 nm) stimulation, resulting in the depolarization and activation of neurons. It is widely used in the analysis and functional characterization of neuronal circuits. In addition, other ion channel rhodopsins are naturally present in nature. The unique redshift properties of VChR1 from Volvox carteri, as reported by Zhang et al., provided an important reference for other genetically engineered rhodopsin variants (Zhang et al., 2008); Marshel et al. obtained ChRmine with high photosensitivity and high electrical conductivity through biogenic analysis and gene mining, which can be activated by light to a depth of 7cm in the cerebral cortex, which suggests that it has broad applicability (Marshel et al., 2019; Chen et al., 2020); VirChR1, a viral rhodopsin discovered by Zabelskii et al. (2020) in Antarctica, blocks Ca2+ penetration, effectively solving the problem of neurotransmitter release from the presynaptic membrane caused by Ca2+ penetration and avoiding the adverse effects on downstream neuronal activities. Other wild-type light-gated ion channel microbial rhodopsins are presented in Table 1.
Most excitingly, A new type of engineed rhodopsin developed by targeted mutations or chimeric modifications of the ChR protein family is particularly well characterized for various scenarios of optogenetic applications. Based on their excellent properties, the variants can be roughly classified into three categories: kinetic opsins, red-shifted opsins, and step-function opsins (SFO) (Kim et al., 2017).
Kinetic opsins are ChR2 variants with high-frequency excitation and rapid inactivation properties, including ChETA (Gunaydin et al., 2010), CHIEF (Lin et al., 2009) and ET/TC (Lin et al., 2013). These variants are excited by high-frequency excitation light and can better simulate the excitation state of neurons under natural conditions, which is of great significance for studying neural encoding. Alternatively, step-function opsins (SFO) include stabilized step-function opsin (SSFO) (Yizhar et al., 2011) and step-waveform inhibitory ChR (SwiChR) (Berndt et al., 2015); this is a class of opsins that can maintain the state of neurons for a long time by brief light excitation and their function can be decreased gradually by a certain order of magnitude, thus explaining the name step-function opsin. For example, they can be used for prolonged, chronic neuromodulation of specific neurons or neuronal circuits.
Due to the low tissue penetration of ChR2 stimulated by blue light, it cannot meet the demand of deep brain projection. Zhang et al. (2008) developed the first red-shifted optic protein C1V1 using an assembly chimera. However, the remote projection performance of C1V1 is inadequate and is not yet able to transmit the signal to the projection end of neural circuits. Subsequently Lin et al. (2013) developed another red-shifted opsin-ReaChR. Although ReaChR can deliver a sufficient amount of opsin to the end of a neural circuit, it is poorly compatible with mammals and can only induce a weak photocurrent with poor fidelity. Later, Rajasethupathy et al. combined the above-mentioned research strategies and developed variants with high photocurrent, redshiftable, and high teleprojection abilities using the system of target mutation and chimera assembly-bReaChES (Rajasethupathy et al., 2015). In addition, the red-shifted property of the excitation light in redshift proteins allows the combined use of redshift and blueshift proteins, and genetically encoded voltage indicators (GECIs), for multiple photorecording and simultaneous acquisition of neural circuits.
The aforementioned ion pump rhodopsins for neuronal silencing can only transport a single ion during the photocycle induced by a single photon, thus showing low ion transport efficiency. Therefore, a more desirable neurosilencing tool would be a natural anion-conducting channelrhodopsin like the cation-conducting ChR2. In 2014, Deisseroth and Hegemann independently designed and successfully obtained the anionic channel rhodopsins: iC1V1 (Berndt et al., 2014) and ChloC (Wietek et al., 2014). Deisseroth obtained iC1V1 via a targeted mutation at 9 positions (T98S/E129S/E140S V156K/E162S H173R/V281K/T285N/N297Q) based on the molecular structure information of the C1V1. Hegemann obtained ChloC by introducing two mutation sites (E90R/T159C) into ChR2. However, the original iC1V1 still transported H+, and the optimized, and successfully obtained iC + + and iChloC have higher selectivity and conductivity to Cl– in two variants while solving the efficiency problem (Berndt et al., 2015). Meanwhile, Spudich isolated naturally occurring ACRs from Guillardia Theta, including GtACR1 and GtACR2. These ACRs overcame the shortcomings of conventional ion pump transport efficiency, such that ACR can transfer 104–105 ions per second and exhibit a 102–104-fold higher photosensitivity than the inhibitory ion pump rhodopsin. In recent years, an increasing number of ACRs have been obtained through genome mining and engineering, such as ZipACR (Govorunova et al., 2017b), RapACR (Govorunova et al., 2018), and RubyACR (Govorunova et al., 2020). Recently, Govorunova et al. reported that the long-sought kalium channelrhodopsins (KCRs) from Hyphochytrium catenoides, HcKCR1 and HcKCR2, show tunable inhibition of mouse cortical neurons with millisecond precision, providing a promising optogenetic tool for the study and treatment of Parkinson’s disease, epilepsy, and other disorders (Govorunova et al., 2022).
Along with engineering modifications of rhodopsins, novel variants with many favorable characteristics have been developed. For instance, rhodopsins that can simultaneously activate and inhibit the same neuron in a single experiment, such as the eNPAC2.0 (NpHR-TS-p2A-hChR2(H134R)-EYFP) designed by Carus-Cadavieco et al. (2017). The experimental requirement of yellow light (586/20 nm) for inhibition and blue light (475/28 nm) for activation helped to elucidate the neural loop mechanisms involved in foraging behavior. Unfortunately, the results were not completely satisfactory. The reliability of blue light neuronal has not yet reached 100% (5 Hz: shows 84.0 ± 11.0% and 20 Hz shows 65.0 ± 12.0) (Carus-Cadavieco et al., 2017; Gao et al., 2021). Vierock et al. (2021) further optimized the blue-light-activated GtACR2 and red-light-activated Chrimson chimerism to obtain BiPOLES, which effectively solved the above problems and realized that the inhibitory anionic and excitatory cationic currents were observed maximum photocurrent density in different excitation light peak. The effect was validated in Drosophila melanogaster, mice, and ferrets, thereby satisfying the need for bidirectional and simultaneous control of neuronal activity in a single experiment (Vierock et al., 2021). The above examples demonstrate that engineered modified bicolor rhodopsins can meet the demand for rapid switching of activating/inhibiting neuronal activity and have great potential for biomedical applications. More variant types and characteristics are detailed in Table 2.
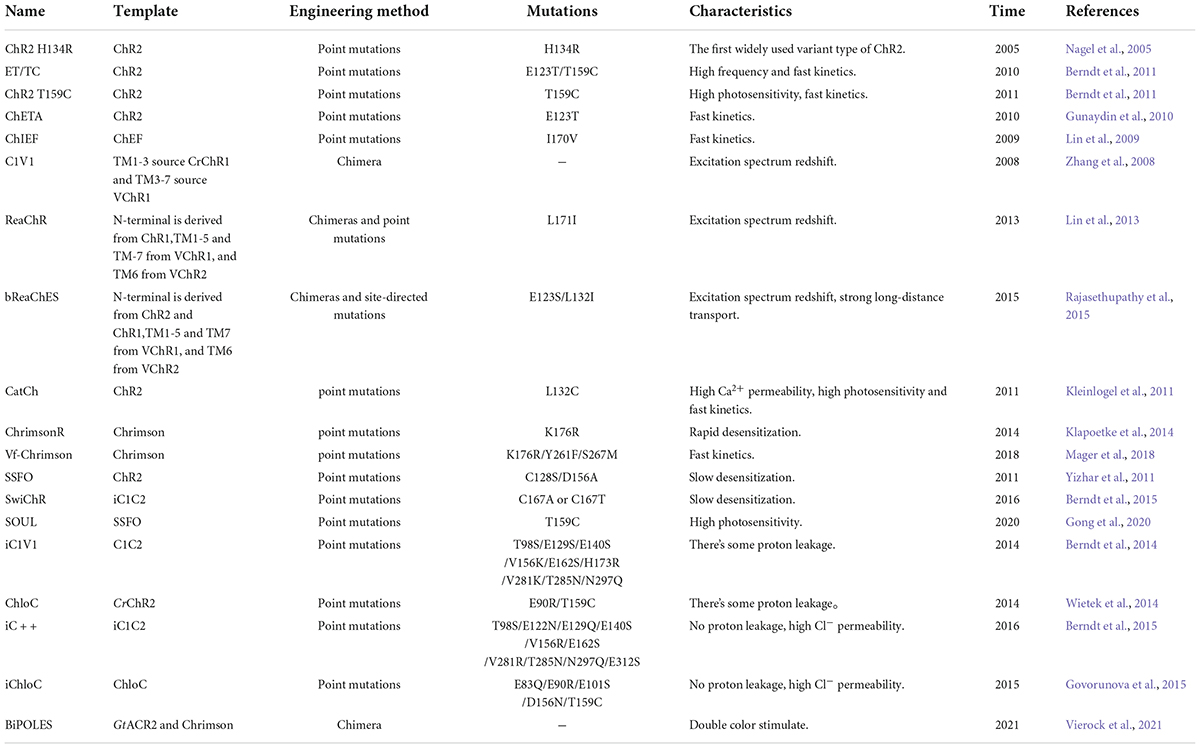
Table 2. Widely used engineered variants.
The first class of signaling/enzyme rhodopsins was discovered in 1982 in Halobacterium. Salinarum; these included sensory rhodopsin I (SRI) (Spudich and Spudich, 1982) and sensory rhodopsin II (SRII, also known as phoborhodopsin, pR) (Bogomolni and Spudich, 1982). Sensory rhodosins (SRs) are embedded in the cell membrane in seven transmembrane alpha helices. When excited, SRs convert the light signal to the cognate transducer halobacterial transducer proteins (Htrs), and form 2:2 complexes with their cognate transducer proteins (halobacterial transducer protein for SRI (HtrI) and halobacterial transducer protein for SRII (HtrII)), which causes a downstream cascade reaction leading to flagellar motility and participation in phototropism. Other signaling/enzyme rhodopsins were subsequently identified. Unlike the previous microbial rhdopsins, the subsequently identified signaling/enzyme rhodopsins are eight-transmembrane (8-TM) proteins, with the C- and N-terminal ends of the protein located on the cytoplasmic side. All the identified signaling/enzyme rhodopsins exist as dimers, including histidine kinase rhodopsin (HKR) (Luck et al., 2012), guanylate cyclase (Rh-GC) (Avelar et al., 2014), and rhodopsin phosphodiesterase (Rh-PDE) (Yoshida et al., 2017). It has been demonstrated that light-controlled signaling/enzyme rhodopsins can be utilized to solve basic science problems. For example, Henss et al. expressed Rh-GC in Cryptobacterium hidrophilum and used this to control the locomotion speed of nematodes via light activation (Henss et al., 2022).
With the advent of optogenetics, there has been an uptake of application in basic biomedical medicine research, such as in the exploration of the mechanism of neurodegenerative diseases. Simultaneously, clinical applications of optogenetics have been explored, including in areas such as vision restoration and optical cochlear implants (oCIs).
Alzheimer’s disease (AD) is a neurodegenerative disease that typically occurs in middle-aged and elderly people. With an increasing aging population, the number of AD patients is rising every year (Scheltens et al., 2021). According to current epidemiological data, the global prevalence of AD will increase three folds by 2050. Current research on the pathogenesis of AD has generated a number of pathophysiological hypotheses that include amyloid-beta (Aβ) toxicity (Selkoe and Hardy, 2016), tau protein hyperphosphorylation (Liang et al., 2022), and synaptic dysfunction (Skaper et al., 2017). Optogenetics provides a novel approach to the study of AD pathogenesis and its clinical treatment. Zhang et al. (2020) used ChR2 to drive GABA neurons and found that this could attenuate neuroinflammation and amyloid β protein levels in the hippocampal CA1 region of APP/PS1 mice and help to restore memory. Guillaume Etter et al. expressed ChETA in MSPV cells and found that activation of parvalbumin interneurons at a 40 Hz frequency restored the normal hippocampal slow-wave amplitude and phase-amplitude coupling in the J20 amyloid β AD mouse model (Etter et al., 2019). Roy et al. reported that the reduction in dendritic ridge density of DG neurons could be reversed by optogenetic techniques in animals with early AD, and that activation of DG engram cells using optogenetic techniques could also restore situational memory (Roy et al., 2016). The aforementioned animal experiments suggest that optogenetic techniques may provide a potential avenue for the treatment of AD. At present, because of the multiplicity of neuronal degenerative lesions in the AD brain, current treatments generally target the entire brain. In addition, most viral vectors need to be embedded in the host cell genome to achieve stable expression, yet there are no effective means to achieve the simultaneous integration of viral sequences within the host genome of the whole brain. As such, current optogenetic techniques are not yet matured enough to be exploited to target the whole brain.
Parkinson’s disease is another clinically common neurodegenerative disorder that begins with the degenerative death of dopaminergic neurons in the substantia nigra of the midbrain, eventually leading to abnormalities in the glutamatergic excitatory pathways in the cerebral cortex, limbic system to the basal ganglia, and motor areas of the brainstem spinal cord (Bloem et al., 2021). Clinical symptoms are mainly motor abnormalities, such as slow and increasing motor dysfunction in individuals, and non-motor clinical symptoms such as cognitive impairment, depression, and pain. The neuropathological markers of PD include the presence of Lewy bodies and Lewy neurites (neuronal inclusions immunopositive for the protein α-synuclein); similar pathological changes have been demonstrated in skin, blood, colon and other tissues (Shahmoradian et al., 2019). Optogenetic stimulation of glutamatergic neurons in the cuneiform nucleus of the midbrain motor area, which in turn activates glutamatergic excitatory pathways extending to the motor areas of the brainstem spinal cord, has been shown to effectively improve dyskinesia in a mouse PD model (Fougère et al., 2021). Deep brain stimulation (DBS) is a common and effective treatment method utilized in clinical practice; combining DBS with optogenetics can be used to explore the neural mechanisms that underlie symptomatic improvement in response to DBS (Gittis and Yttri, 2018). We have reason to believe that, as optogenetic technology improves, it will prove to be critical to revealing the pathogenic mechanisms of Parkinson’s disease and in aiding patient treatment.
Astrocytes are an important component of the tripartite synapse structure; they receive neurotransmitter signals through cell surface receptors and regulate synaptic activity via gliotransmitters. In recent years, research on neurodegenerative diseases has begun to focus on glial cells. Xie et al. (2019) reported that activation of signaling pathways related to synaptic activity in astrocytes by optogenetic techniques affects the release of gliotransmitters, which can modulate the communication network between astrocytes and neurons. Other studies have focused on microglia, which are another important neuroglial cell that plays a major role in phagocytosis and immunomodulation. Microglial activation can be accompanied by the inward flow of chloride ions which further initiates their phagocytic activity. The introduction of chloride ions could be achieved by NpHR, and we speculate that this may offer a potential therapeutic application if microglia are specifically activated in PD lesioned brain regions, which in turn initiates phagocytosis of α-synuclein (Averaimo et al., 2010).
Similar to neurons, cardiomyocytes are also excitable cells. Thus, optogenetics can be used to regulate their physiological activity. Optogenetics have been exploited in the study of the heart for around 12 years (Entcheva and Kay, 2020). In 2014, Bingen et al. transfected CatCh into neonatal rat cardiomyocytes as a strategy to defibrillate the heart through cell activation, thereby suggesting an optogenetic therapeutic strategy for patients with atrial fibrillation (Bingen et al., 2014). Such an approach would address some of the limitations of electric defibrillators, such as reducing the risk of myocardial tissue damage and pain. A recent paper utilizing computational simulation pointed out that GtACR1 may be used to defibrillate human hearts (Ochs et al., 2021). Majumder et al. (2020) developed a theoretical ion channel called the Biologically Integrated Cardiac Defibrillator (BioICD), whose activation can lead to a rapid and repeated restoration of normal rhythm for arrhythmia in the human atrium and ventricle (Majumder et al., 2020). It is believed that this theoretical simulation may soon be developed. Unlike the nervous system however, cardiac optogenetics lacks well-characterized tissue-specific promoters.
Ophthalmic optogenetics is the earliest biomedical application of optogenetics. Bi et al. (2006) pioneered the expression of ChR2-encapsulated AAV vectors in retinal ganglion cells (RGCs), demonstrating the potential of optogenetics in visual recovery (Bi et al., 2006). Several variants of ChR2 introduced earlier have shown promising ophthalmic applications. For example Sengupta et al. (2016) used ReaChR carried by the AAV vector and injected into the rd1 mouse model of blindness and orange light with an intensity lower than the human safety threshold to restore retinal, cortical, and behavioral levels of light-response. Chuong et al. (2014) showed that Jaws-encapsulated AAV vectors achieved light responses from isolated ON- and OFF-retinal ganglion cells in transgenic mice after 600 nm light stimulation. Recently, ChrimsonR was successfully transfected using AAV into a patient’s retinal ganglion cells, resulting in a partial recovery of the degenerated retina. This is the first successful optogenetics case in neurodegenerative disease (Sahel et al., 2021). In addition, four companies have promoted ophthalmic optogenetics in clinical trials: (1) GenSight Biologics, GS030(Paris, France), (2) Allergan, RST-001 (Dublin, Ireland), (3) Bionic Sight, BS01(New York, NY, USA), and (4) Nanoscope Therapeutics, vMCO-010 (Bedford, TX, USA) (Prosseda et al., 2022). Although optogenetics has advanced our ability to effect the recovery of visual acuity, there are marked differences between optogenetic vision and natural vision. One is that photoreceptor cells can detect the stimulation intensity of a single photon, and there is a significant difference in sensitivity. The other is that optogenetic vision is not yet involved in the formation of color perception. If different light wavelength optogenetic tool proteins are expressed in specific cell types, color or color-like perception may emerge. However, it is believed that optogenetics will continue to play a strong role in ophthalmic research and clinical practice.
Electronic cochlear implants (eCIs) are currently the primary clinical alternative to the organ of Corti for restoring hearing function. When hearing loss or hearing impairment is severe, eCIs bypass the sensory hair cells to directly convert sound information into a digital signal, which is reprocessed into a radio frequency (RF) signal that is embedded in the patient’s skin with an internal receiver and stimulator. The RF is decoded and the spiral ganglion neurons (SGNs) are directly stimulated along the electrodes to produce hearing (Zeng et al., 2008). The drawbacks of eCIs include a low frequency resolution due to the wide distribution of electrode contacts and current diffusion, etc., which makes them less effective in noisy environments. Optical cochlear implants (oCIs) modulate the sound signal into an optical signal with the assistance of an acousto-optic modulator, which directly replaces electrical stimulation with optical stimulation. This process improves the frequency resolution of artificial sound encoding due to a lower spatial spread of neural excitation. Currently, Keppeler et al. have established the expression system of CatCh by AAV in rodents and experimentally validated LED-based piggyback oCIs. Usually, LED-based cochlear implants carry 10-15 LED exciters (emitters), while μLED-based oCIs can increase the number of emitters to 144 emitters of 60 × 60 lm, which has been established by utilizing 16 microscale thin-film light-emitting diodes (μLEDs) for oCIs (Klein et al., 2018; Dieter et al., 2020; Keppeler et al., 2020). A recent study showed that the Chrimson variants f-Chrimson and vf-Chrimson also have a potential for optical cochlear implant applications (Bali et al., 2021). At present, the development of oCIs is still in its formative stages, and researchers are working hard to find better matching conditions for rhodopsins and to develop oCIs with higher frequency resolution. It is believed that this work will facilitate the design of next generation oCIs and facilitate their clinical translatability.
Optogenetics has empowered basic research scientists and clinicians to precisely manipulate cellular activity. Its application to basic research and therapeutics has attracted numerous research teams. The ultimate goal is the widespread safe and effective use of optogenetics in the human body. One step toward this goal would involve solving the problem of light delivery, such as through the development of wireless optogenetics. In this section we discuss some of the most pressing and core issues in the therapeutic translatability of optogenetics.
As is the case with other gene therapies, optogenetics requires a vehicle to deliver its proteins to specific cell types; viruses are the ideal transporters. Adeno-associated viruses (AAV) are one of the most widely used viral tools in basic research and clinical trials. As a non-pathogenic member of the parvoviridae family, AAV is composed of a single-stranded DNA genome encapsidated in a 23-28 nm, T = 1, non-invariant capsid (El Andari and Grimm, 2020). Three serotypes of AAV have been licensed for clinical use, including Glybera (AAV1), Luxturna (AAV2), and Zolgensma (AAV9) (Domenger and Grimm, 2019). These viruses are used as opsin transporters for the following reasons: (1) they are non-encapsulated viruses, which are more stable and less likely to break down under laboratory conditions; (2) they are wild-type viruses without virulent potential; although this increases their safety, immunogenicity must be considered. Because AAV is prevalent in humans, most patients carry antibodies to AAV, so they may respond to endogenous AAV vectors through pre-existing immunity (PEI). Therefore, there is a need to develop isolates or serotypes that do not trigger such responses or that only display mild immunogenicity. Another strategy is to modify the gene sequence of the antigenic part of the AAV sequence so that its antigen cannot bind antibodies in the circulatory system to minimize the immune response induced by AAV, thus increasing the efficiency and possibility of targeting cells; (3) they are efficient opsin transporters, including in non-human primates. Their application in non-human primate studies provides sufficient experimental data and theoretical support for the clinical application of optogenetics, as non-human primates have a high similarity with humans in terms of anatomical structure and genetic information; (4) they are free from host DNA, stable by satellite form, and cannot integrate into the host genome. A recent study, in which AAV2-CAG-ChrimsonR:tdTomato was injected into the vitreous humor of macaques demonstrated that optogenetics-mediated RGCs activity persisted for at least 12 months after 12 weeks of transfection (McGregor et al., 2022).
In addition, the precise delivery of the viral vector to the target cells is also problematic. Currently, mainly stereotactic techniques are used, but they carry the risk of infection and may not allow all desirable regions to be accurately targeted. Alternative techniques include ventricular, spinal, and vascular injection modalities, but these methods have their own problems. Because AAVs are hepatophilic, most are injected into the organism and become distributed to the liver, which may produce hepatotoxicity and reduce their targetability. To obtain AAV serotypes that can better target the CNS, PHP.B (Deverman et al., 2016) and PHP.S (Challis et al., 2019) were designed. Recently, scientists have shown organ-specific targeting of adeno-associated virus (AAV) capsids after intravenous delivery, which we achieved by employing a Cre-transgenic-based screening platform and sequential engineering of AAV-PHP.eB between the surface-exposed AA452 and AA460 of VP3. This variant exhibits good CNS targeting and low hepatotropism and has been validated in Callithrix Jacchus (Goertsen et al., 2021).
Light delivery is another major challenge for the clinical utilization of optogenetics. In traditional basic optogenetics research, stereotactic positioning followed by fiber optic implantation is usually adopted. There are certain drawbacks to using this method: the surgery is difficult to precisely perform and leaves the animals at risk of infection. In addition, the tissue damage caused by the fiber optic implants is irreversible and behavioral activities are easily restricted by a combination of the surgery and attendant equipment. Moreover, the heat generated by the light can cause phototoxicity (Owen et al., 2019). Previous studies have shown that activation of the blue light-activated stGtACR2 opsin, expressed in the hippocampus of rats, increased the local peak brain temperature, thereby significantly decreasing the population spike (PS) amplitudes and latencies of Eps, thus confounding the central experimental methods (Acharya et al., 2021). The emergence of wireless optogenetics has solved the problem of wired fiber implants. Researchers have developed an improved optical transmission system that transmits light through a wireless headset and uses small light-emitting diodes (LEDs) to activate the rhodopsin mass, eliminating the need for optical fibers and allowing for wireless charging (Kwon et al., 2015; Montgomery et al., 2015). Recently, a less invasive wireless optogenetic technique has been developed using light with wavelengths shorter than the visible spectrum, such as ultraviolet or X-ray with wavelengths < 450 nm or higher wavelengths, such as infrared light with wavelengths > 650 nm; this inorganic scintillator has been used for the bidirectional modulation of neurons (Miyazaki et al., 2019; Matsubara et al., 2021).Therefore, wireless optogenetics is expected to help advance the uptake of optogenetic-based clinical treatments.
It is worth noting that most of the excitation wavelengths of the current optogenetic tool protein - microbial rhodopsins - involve visible light (400nm-700nm). However, if blue light optic violet matter is used in very deep target brain regions, wireless optogenetics may appear airy and ineffective. Although the use of redshift protein has solved part of the problem, gaps remain in the clinical application of optogenetics (Chen, 2019). Therefore, both traditional and wireless technical means are limited by the microbial rhodopsins itself. Upconversion nanoparticles (UCNPs) can absorb near-infrared light and emit visible light from ultraviolet to near-red light. Compared with traditional optical fiber implantation, UCNP-mediated optogenetics has the advantages of less tissue damage, wireless transmission, and negligible toxicity, More importantly, it reduces the consideration of microbial rhodopsins properties. One interesting report demonstrated a UCNP with a photoreceptor for anchoring retinal photoreceptors and sending signals to the brain upon near-red light stimulation, allowing mammals to see infrared light (Ma et al., 2019); other applications of UCNPs in biomedicine have also been summarized (Mahata et al., 2021).
Rhodopsin-based optogenetics have been widely used in various fields of life science research. Although their development in the field of clinical medicine is still in its early stages, their great potential in this area is well recognized. For the sake of brevity, in this review, we focused on the application of optogenetics in several specific fields of medicine, but we recognize that their utility will become more widespread in the future. In the field of autism, optogenetics has been used to activate the glutamatergic BF-VTA pathway, leading to an immediate shift from sleep to awakening in mice. This showed that excessive excitation of the glutamatergic BF-VTA pathway is associated with clinical disorders characterized by excessive defensive behaviors (Cai et al., 2022). In regenerative medicine, a recent article reported that optogenetic tools improve the differentiation of primary murine myotubes (Chapotte-Baldacci et al., 2022). In addition, optogenetics plays an important role in revealing depression-like neural circuits (Biselli et al., 2019). Of course, many problems need to be overcome. The selection and transmission of viral vectors and optical transmission are some of the major core issues. In addition, immunogenicity, cell specificity, and other challenges stemming from the use of viral vectors need to be urgently solved. To solve these problems, experiments must be conducted on non-human primates. At present, an Open Resource for Non-human Primate Optogenetics has been established (Tremblay et al., 2020), which will provide important technical support for the experimentation of optogenetic technology in non-human primates and promote its clinical translation.
It is clearly that optogenetic techniques have unique applications and prospects for development, but more importantly their combination with other techniques will provide strategies and technical means to solve many problems. One of the promising research directions is closed-loop experiments based on optogenetics and optical imaging techniques, but an ideal algorithm for the accurate identification of active neurons is lacking. Sheng et al. constructed analysis software called Online Real-time activity and offline Cross-session Analysis (ORCA), which performs image alignment, neuron segmentation, and activity extraction at over 100 frames per second; this is fast enough to support real-time neuron detection and readout activity, providing a solution for full optical closed-loop control of neuronal activity (Sheng et al., 2022). This will provide software support for real-time feedback control of neurons based on neuronal activity, with far-reaching promise in addressing neurological disorders such as epilepsy. We believe that optogenetics will play an important role in advancing basic research and that it will ultimately greatly benefit patients.
JZ, RT, and JY: conceptualization. HZ, HF, DL, and YZ: methodology. HZ and JA-A: formal analysis. JZ, HZ, RT, and JY: resources. JZ, HZ, HF, DL, and YZ: writing-original draft. JA-A and RT: revising and editing. All authors: writing-review and editing, investigation.
This work was supported by the National Natural Science Foundation of China (81700055, RT); the Outstanding Talent Research Funding of Xuzhou Medical University (Grant No. D2016021, RT); the Natural Science Foundation of Jiangsu Province (BK20160229, RT); and Research Startup Fund of Jining Medical University (600791001, JY).
We would like to acknowledge the support of the research team at the Core Facility Center for College of Life Sciences, Shandong Normal University. We would also like to express their gratitude to EditSprings (https://www.editsprings.cn) for the expert linguistic services provided.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acharya, A. R., Vandekerckhove, B., Larsen, L. E., Delbeke, J., Wadman, W. J., Vonck, K., et al. (2021). In vivo blue light illumination for optogenetic inhibition: Effect on local temperature and excitability of the rat hippocampus. J. Neural Eng. 18:066038. doi: 10.1088/1741-2552/ac3ef4
Avelar, G. M., Schumacher, R. I., Zaini, P. A., Leonard, G., Richards, T. A., and Gomes, S. L. (2014). A rhodopsin-guanylyl cyclase gene fusion functions in visual perception in a fungus. Curr. Biol. 24, 1234–1240. doi: 10.1016/j.cub.2014.04.009
Averaimo, S., Milton, R. H., Duchen, M. R., and Mazzanti, M. (2010). Chloride intracellular channel 1 (CLIC1): Sensor and effector during oxidative stress. FEBS Lett. 584, 2076–2084. doi: 10.1016/j.febslet.2010.02.073
Bali, B., Lopez de la Morena, D., Mittring, A., Mager, T., Rankovic, V., Huet, A. T., et al. (2021). Utility of red-light ultrafast optogenetic stimulation of the auditory pathway. EMBO Mol. Med. 13:e13391. doi: 10.15252/emmm.202013391
Berndt, A., Lee, S. Y., Ramakrishnan, C., and Deisseroth, K. (2014). Structure-guided transformation of channelrhodopsin into a light-activated chloride channel. Science 344, 420–424. doi: 10.1126/science.1252367
Berndt, A., Lee, S. Y., Wietek, J., Ramakrishnan, C., Steinberg, E. E., Rashid, A. J., et al. (2015). Structural foundations of optogenetics: Determinants of channelrhodopsin ion selectivity. Proc. Natl. Acad. Sci. U.S.A. 113, 822–829. doi: 10.1073/pnas.1523341113
Berndt, A., Schoenenberger, P., Mattis, J., Tye, K. M., Deisseroth, K., Hegemann, P., et al. (2011). High-efficiency channelrhodopsins for fast neuronal stimulation at low light levels. Proc. Natl. Acad. Sci. U.S.A. 108, 7595–7600. doi: 10.1073/pnas.1017210108
Bi, A., Cui, J., Ma, Y.-P., Olshevskaya, E., Pu, M., Dizhoor, A. M., et al. (2006). Ectopic Expression of a microbial-type rhodopsin restores visual responses in mice with photoreceptor degeneration. Neuron 50, 23–33. doi: 10.1016/j.neuron.2006.02.026
Bingen, B. O., Engels, M. C., Schalij, M. J., Jangsangthong, W., Neshati, Z., Feola, I., et al. (2014). Light-induced termination of spiral wave arrhythmias by optogenetic engineering of atrial cardiomyocytes. Cardiovasc. Res. 104, 194–205. doi: 10.1093/cvr/cvu179
Biselli, T., Lange, S. S., Sablottny, L., Steffen, J., and Walther, A. (2019). Optogenetic and chemogenetic insights into the neurocircuitry of depression-like behaviour: A systematic review. Eur. J. Neurosci. 9:38. doi: 10.1111/ejn.14603
Bloem, B. R., Okun, M. S., and Klein, C. (2021). Parkinson’s disease. Lancet 397, 2284–2303. doi: 10.1016/s0140-6736(21)00218-x
Bogomolni, R. A., and Spudich, J. L. (1982). Identification of a third rhodopsin-like pigment in phototactic Halobacterium halobium. Proc. Natl. Acad. Sci. U.S.A. 79, 6250–6254. doi: 10.1073/pnas.79.20.6250
Boyden, E. S., Zhang, F., Bamberg, E., Nagel, G., and Deisseroth, K. (2005). Millisecond-timescale, genetically targeted optical control of neural activity. Nat. Neurosci. 8, 1263–1268. doi: 10.1038/nn1525
Cai, P., Chen, H.-Y., Tang, W.-T., Hu, Y.-D., Chen, S.-Y., Lu, J.-S., et al. (2022). A glutamatergic basal forebrain to midbrain circuit mediates wakefulness and defensive behavior. Neuropharmacology 208:108979. doi: 10.1016/j.neuropharm.2022.108979
Carus-Cadavieco, M., Gorbati, M., Ye, L., Bender, F., van der Veldt, S., Kosse, C., et al. (2017). Gamma oscillations organize top-down signalling to hypothalamus and enable food seeking. Nature 542, 232–236. doi: 10.1038/nature21066
Challis, R. C., Ravindra Kumar, S., Chan, K. Y., Challis, C., Beadle, K., Jang, M. J., et al. (2019). Systemic AAV vectors for widespread and targeted gene delivery in rodents. Nat. Protoc. 14, 379–414. doi: 10.1038/s41596-018-0097-3
Chapotte-Baldacci, C.-A., Cognard, C., Bois, P., Chatelier, A., and Sebille, S. (2022). Handling a mature calcium signature through optogenetics improves the differentiation of primary murine myotubes. Cell Calcium 103:102546. doi: 10.1016/j.ceca.2022.102546
Chen, R., Gore, F., Nguyen, Q.-A., Ramakrishnan, C., Patel, S., Kim, S. H., et al. (2020). Deep brain optogenetics without intracranial surgery. Nat. Biotechnol. 39, 161–164. doi: 10.1038/s41587-020-0679-9
Chen, S. (2019). Optical modulation goes deep in the brain. Science 365, 456–457. doi: 10.1126/science.aay4350
Chen, Y.-H., Wu, J.-L., Hu, N.-Y., Zhuang, J.-P., Li, W.-P., Zhang, S.-R., et al. (2021). Distinct projections from the infralimbic cortex exert opposing effects in modulating anxiety and fear. J. Clin. Investig. 131:e145692. doi: 10.1172/jci145692
Chow, B. Y., Han, X., Dobry, A. S., Qian, X., Chuong, A. S., Li, M., et al. (2010). High-performance genetically targetable optical neural silencing by light-driven proton pumps. Nature 463, 98–102. doi: 10.1038/nature08652
Chuong, A. S., Miri, M. L., Busskamp, V., Matthews, G. A. C., Acker, L. C., Sørensen, A. T., et al. (2014). Noninvasive optical inhibition with a red-shifted microbial rhodopsin. Nat. Neurosci. 17, 1123–1129. doi: 10.1038/nn.3752
Danon, A., and Stoeckenius, W. (1974). Photophosphorylation in Halobacterium halobium. Proc. Natl. Acad. Sci. U.S.A. 71, 1234–1238. doi: 10.1073/pnas.71.4.1234
Deverman, B. E., Pravdo, P. L., Simpson, B. P., Kumar, S. R., Chan, K. Y., Banerjee, A., et al. (2016). Cre-dependent selection yields AAV variants for widespread gene transfer to the adult brain. Nat. Biotechnol. 34, 204–209. doi: 10.1038/nbt.3440
Dieter, A., Klein, E., Keppeler, D., Jablonski, L., Harczos, T., Hoch, G., et al. (2020). μLED-based optical cochlear implants for spectrally selective activation of the auditory nerve. EMBO Mol. Med. 12:e12387. doi: 10.15252/emmm.202012387
Domenger, C., and Grimm, D. (2019). Next-generation AAV vectors—do not judge a virus (only) by its cover. Hum. Mol. Genet. 28:R3–R14. doi: 10.1093/hmg/ddz148
Duan, L., Huang, P., and Zhao, Z. (2022). Optogenetic activation of intracellular signaling based on light-inducible protein-protein homo-interactions. Neural Regen. Res. 17:25. doi: 10.4103/1673-5374.314293
El Andari, J., and Grimm, D. (2020). Production, processing, and characterization of synthetic AAV gene therapy vectors. Biotechnol. J. 16:e2000025. doi: 10.1002/biot.202000025
Entcheva, E., and Kay, M. W. (2020). Cardiac optogenetics: A decade of enlightenment. Nat. Rev. Cardiol. 18, 349–367. doi: 10.1038/s41569-020-00478-0
Etter, G., van der Veldt, S., Manseau, F., Zarrinkoub, I., Trillaud-Doppia, E., and Williams, S. (2019). Optogenetic gamma stimulation rescues memory impairments in an Alzheimer’s disease mouse model. Nat. Commun. 10:5322. doi: 10.1038/s41467-019-13260-9
Fougère, M., van der Zouwen, C. I., Boutin, J., Neszvecsko, K., Sarret, P., and Ryczko, D. (2021). Optogenetic stimulation of glutamatergic neurons in the cuneiform nucleus controls locomotion in a mouse model of Parkinson’s disease. Proc. Natl. Acad. Sci. U.S.A. 118:e2110934118. doi: 10.1073/pnas.2110934118
Gao, X., Bender, F., Soh, H., Chen, C., Altafi, M., Schütze, S., et al. (2021). Place fields of single spikes in hippocampus involve Kcnq3 channel-dependent entrainment of complex spike bursts. Nat. Commun. 12:4801. doi: 10.1038/s41467-021-24805-2
Gittis, A. H., and Yttri, E. A. (2018). Translating insights from optogenetics into therapies for Parkinson’s disease. Curr. Opin. Biomed. Eng. 8, 14–19. doi: 10.1016/j.cobme.2018.08.008
Goertsen, D., Flytzanis, N. C., Goeden, N., Chuapoco, M. R., Cummins, A., Chen, Y., et al. (2021). AAV capsid variants with brain-wide transgene expression and decreased liver targeting after intravenous delivery in mouse and marmoset. Nat. Neurosci. 25, 106–115. doi: 10.1038/s41593-021-00969-4
Gong, X., Mendoza-Halliday, D., Ting, J. T., Kaiser, T., Sun, X., Bastos, A. M., et al. (2020). An ultra-sensitive step-function opsin for minimally invasive optogenetic stimulation in mice and macaques. Neuron 107, 38–51.e8. doi: 10.1016/j.neuron.2020.03.032
Govorunova, E. G., Gou, Y., Sineshchekov, O. A., Li, H., Lu, X., Wang, Y., et al. (2022). Kalium channelrhodopsins are natural light-gated potassium channels that mediate optogenetic inhibition. Nat. Neurosci. 25, 967–974. doi: 10.1038/s41593-022-01094-6
Govorunova, E. G., Sineshchekov, O. A., Hemmati, R., Janz, R., Morelle, O., Melkonian, M., et al. (2018). Extending the time domain of neuronal silencing with cryptophyte anion channelrhodopsins. eNeuro 5:ENEURO.0174–18.2018. doi: 10.1523/eneuro.0174-18.2018
Govorunova, E. G., Sineshchekov, O. A., Janz, R., Liu, X., and Spudich, J. L. (2015). Natural light-gated anion channels: A family of microbial rhodopsins for advanced optogenetics. Science 349, 647–650. doi: 10.1126/science.aaa7484
Govorunova, E. G., Sineshchekov, O. A., Li, H., and Spudich, J. L. (2017a). Microbial rhodopsins: Diversity, mechanisms, and optogenetic applications. Annu. Rev. Biochem. 86, 845–872. doi: 10.1146/annurev-biochem-101910-144233
Govorunova, E. G., Sineshchekov, O. A., Rodarte, E. M., Janz, R., Morelle, O., Melkonian, M., et al. (2017b). The expanding family of natural anion channelrhodopsins reveals large variations in kinetics, conductance, and spectral sensitivity. Sci. Rep. 7:43358. doi: 10.1038/srep43358
Govorunova, E. G., Sineshchekov, O. A., Li, H., Wang, Y., Brown, L. S., and Spudich, J. L. (2020). RubyACRs, nonalgal anion channelrhodopsins with highly red-shifted absorption. Proc. Natl. Acad. Sci. U.S.A. 117, 22833–22840. doi: 10.1073/pnas.2005981117
Gradinaru, V., Mogri, M., Thompson, K. R., Henderson, J. M., and Deisseroth, K. (2009). Optical deconstruction of parkinsonian neural circuitry. Science 324, 354–359. doi: 10.1126/science.1167093
Gradinaru, V., Thompson, K. R., and Deisseroth, K. (2008). eNpHR: A natronomonas halorhodopsin enhanced for optogenetic applications. Brain Cell Biol. 36, 129–139. doi: 10.1007/s11068-008-9027-6
Gradinaru, V., Zhang, F., Ramakrishnan, C., Mattis, J., Prakash, R., Diester, I., et al. (2010). Molecular and cellular approaches for diversifying and extending optogenetics. Cell 141, 154–165. doi: 10.1016/j.cell.2010.02.037
Gunaydin, L. A., Yizhar, O., Berndt, A., Sohal, V. S., Deisseroth, K., and Hegemann, P. (2010). Ultrafast optogenetic control. Nat. Neurosci. 13, 387–392. doi: 10.1038/nn.2495
Henss, T., Schneider, M., Vettkötter, D., Costa, W. S., Liewald, J. F., and Gottschalk, A. (2022). Photoactivated adenylyl cyclases as optogenetic modulators of neuronal activity. Methods Mol. Biol. 2483, 61–76. doi: 10.1007/978-1-0716-2245-2_4
Hososhima, S., Kandori, H., and Tsunoda, S. P. (2021). Ion transport activity and optogenetics capability of light-driven Na+-pump KR2. PLoS One 16:e0256728. doi: 10.1371/journal.pone.0256728
Hososhima, S., Shigemura, S., Kandori, H., and Tsunoda, S. P. (2020). Novel optogenetics tool: Gt_CCR4, a light-gated cation channel with high reactivity to weak light. Biophys. Rev. 12, 453–459. doi: 10.1007/s12551-020-00676-7
Huang, S., Ding, M., Roelfsema, M. R. G., Dreyer, I., Scherzer, S., Al-Rasheid, K. A. S., et al. (2021). Optogenetic control of the guard cell membrane potential and stomatal movement by the light-gated anion channel GtACR1. Sci. Adv. 7:eabg4619. doi: 10.1126/sciadv.abg4619
Husson, S. J., Liewald, J. F., Schultheis, C., Stirman, J. N., Lu, H., and Gottschalk, A. (2012). Microbial light-activatable proton pumps as neuronal inhibitors to functionally dissect neuronal networks in C. elegans. PLoS One 7:e40937. doi: 10.1371/journal.pone.0040937
Inoue, K., Ito, S., Kato, Y., Nomura, Y., Shibata, M., Uchihashi, T., et al. (2016). A natural light-driven inward proton pump. Nat. Commun. 7:13415. doi: 10.1038/ncomms13415
Inoue, K., Ono, H., Abe-Yoshizumi, R., Yoshizawa, S., Ito, H., Kogure, K., et al. (2013). A light-driven sodium ion pump in marine bacteria. Nat. Commun. 4:1678. doi: 10.1038/ncomms2689
Keppeler, D., Schwaerzle, M., Harczos, T., Jablonski, L., Dieter, A., Wolf, B., et al. (2020). Multichannel optogenetic stimulation of the auditory pathway using microfabricated LED cochlear implants in rodents. Sci. Transl. Med. 12:eabb8086. doi: 10.1126/scitranslmed.abb8086
Khorana, H. G., Gerber, G. E., Herlihy, W. C., Gray, C. P., Anderegg, R. J., Nihei, K., et al. (1979). Amino acid sequence of bacteriorhodopsin. Proc. Natl. Acad. Sci. U.S.A. 76, 5046–5050. doi: 10.1073/pnas.76.10.5046
Kim, C. K., Adhikari, A., and Deisseroth, K. (2017). Integration of optogenetics with complementary methodologies in systems neuroscience. Nat. Rev. Neurosci. 18, 222–235. doi: 10.1038/nrn.2017.15
Klapoetke, N. C., Murata, Y., Kim, S. S., Pulver, S. R., Birdsey-Benson, A., Cho, Y. K., et al. (2014). Independent optical excitation of distinct neural populations. Nat. Methods 11, 338–346. doi: 10.1038/nmeth.2836
Klein, E., Gossler, C., Paul, O., and Ruther, P. (2018). High-density μLED-based optical cochlear implant with improved thermomechanical behavior. Front. Neurosci. 12:659. doi: 10.3389/fnins.2018.00659
Kleinlogel, S., Feldbauer, K., Dempski, R. E., Fotis, H., Wood, P. G., Bamann, C., et al. (2011). Ultra light-sensitive and fast neuronal activation with the Ca2+-permeable channelrhodopsin CatCh. Nat. Neurosci. 14, 513–518. doi: 10.1038/nn.2776
Kolesov, D. V., Sokolinskaya, E. L., Lukyanov, K. A., and Bogdanov, A. M. (2021). Molecular tools for targeted control of nerve cell electrical activity. Part I. Acta Nat. 13, 52–64. doi: 10.32607/actanaturae.11414
Kralj, J. M., Douglass, A. D., Hochbaum, D. R., Maclaurin, D., and Cohen, A. E. (2011). Optical recording of action potentials in mammalian neurons using a microbial rhodopsin. Nat. Methods 9, 90–95. doi: 10.1038/nmeth.1782
Kwon, K. Y., Lee, H.-M., Ghovanloo, M., Weber, A., and Li, W. (2015). Design, fabrication, and packaging of an integrated, wirelessly-powered optrode array for optogenetics application. Front. Syst. Neurosci. 9:69. doi: 10.3389/fnsys.2015.00069
Lanyi, J. K. (1986). Halorhodopsin: A light-driven chloride ion pump. Annu. Rev. Biophys. Biophys. Chem. 15, 11–28. doi: 10.1146/annurev.bb.15.060186.000303
Liang, S.-Y., Wang, Z.-T., Tan, L., and Yu, J.-T. (2022). Tau toxicity in neurodegeneration. Mol. Neurobiol. 59, 3617–3634. doi: 10.1007/s12035-022-02809-3
Lin, J. Y., Knutsen, P. M., Muller, A., Kleinfeld, D., and Tsien, R. Y. (2013). ReaChR: A red-shifted variant of channelrhodopsin enables deep transcranial optogenetic excitation. Nat. Neurosci. 16, 1499–1508. doi: 10.1038/nn.3502
Lin, J. Y., Lin, M. Z., Steinbach, P., and Tsien, R. Y. (2009). Characterization of engineered channelrhodopsin variants with improved properties and kinetics. Biophys. J. 96, 1803–1814. doi: 10.1016/j.bpj.2008.11.034
Liu, R., Yang, J., Yao, J., Zhao, Z., He, W., Su, N., et al. (2022). Optogenetic control of RNA function and metabolism using engineered light-switchable RNA-binding proteins. Nat. Biotechnol. 40, 779–786. doi: 10.1038/s41587-021-01112-1
Luck, M., Mathes, T., Bruun, S., Fudim, R., Hagedorn, R., Tran Nguyen, T. M., et al. (2012). A photochromic histidine kinase rhodopsin (HKR1) that is bimodally switched by ultraviolet and blue light. J. Biol. Chem. 287, 40083–40090. doi: 10.1074/jbc.m112.401604
Ma, Y., Bao, J., Zhang, Y., Li, Z., Zhou, X., Wan, C., et al. (2019). Mammalian near-infrared image vision through injectable and self-powered retinal nanoantennae. Cell 177, 243–255.e15. doi: 10.1016/j.cell.2019.01.038
Mager, T., Lopez de la Morena, D., Senn, V., Schlotte, J., D’Errico, A., Feldbauer, K., et al. (2018). High frequency neural spiking and auditory signaling by ultrafast red-shifted optogenetics. Nat. Commun. 9:1750. doi: 10.1038/s41467-018-04146-3
Mahata, M. K., De, R., and Lee, K. T. (2021). Near-infrared-triggered upconverting nanoparticles for biomedicine applications. Biomedicines 9:756. doi: 10.3390/biomedicines9070756
Majumder, R., De Coster, T., Kudryashova, N., Verkerk, A. O., Kazbanov, I. V., Ördög, B., et al. (2020). Self-restoration of cardiac excitation rhythm by anti-arrhythmic ion channel gating. Elife 9:e55921. doi: 10.7554/elife.55921
Marshel, J. H., Kim, Y. S., Machado, T. A., Quirin, S., Benson, B., Kadmon, J., et al. (2019). Cortical layer–specific critical dynamics triggering perception. Science 365:eaaw5202. doi: 10.1126/science.aaw5202
Matsubara, T., Yanagida, T., Kawaguchi, N., Nakano, T., Yoshimoto, J., Sezaki, M., et al. (2021). Remote control of neural function by X-ray-induced scintillation. Nat. Commun. 12:4478. doi: 10.1038/s41467-021-24717-1
Matsuno-Yagi, A., and Mukohata, Y. (1977). Two possible roles of bacteriorhodopsin; A comparative study of strains of Halobacterium halobium differing in pigmentation. Biochem. Biophys. Res. Commun. 78, 237–243. doi: 10.1016/0006-291x(77)91245-1
McGregor, J. E., Kunala, K., Xu, Z., Murphy, P. J., Godat, T., Strazzeri, J. M., et al. (2022). Optogenetic therapy restores retinal activity in primate for at least a year following photoreceptor ablation. Mol. Ther. 30, 1315–1328. doi: 10.1016/j.ymthe.2021.09.014
Miyazaki, T., Chowdhury, S., Yamashita, T., Matsubara, T., Yawo, H., Yuasa, H., et al. (2019). Large timescale interrogation of neuronal function by fiberless optogenetics using lanthanide micro-particles. Cell Rep. 26, 1033–1043.e5. doi: 10.1016/j.celrep.2019.01.001
Montgomery, K. L., Yeh, A. J., Ho, J. S., Tsao, V., Mohan Iyer, S., Grosenick, L., et al. (2015). Wirelessly powered, fully internal optogenetics for brain, spinal and peripheral circuits in mice. Nat Methods 12, 969–974. doi: 10.1038/nmeth.3536
Nagel, G., Brauner, M., Liewald, J. F., Adeishvili, N., Bamberg, E., and Gottschalk, A. (2005). Light activation of channelrhodopsin-2 in excitable cells of caenorhabditis elegans triggers rapid behavioral responses. Curr. Biol. 15, 2279–2284. doi: 10.1016/j.cub.2005.11.032
Nagel, G., Ollig, D., Fuhrmann, M., Kateriya, S., Musti, A. M., Bamberg, E., et al. (2002). Channelrhodopsin-1: A light-gated proton channel in green algae. Science 296, 2395–2398. doi: 10.1126/science.1072068
Nagel, G., Szellas, T., Huhn, W., Kateriya, S., Adeishvili, N., Berthold, P., et al. (2003). Channelrhodopsin-2, a directly light-gated cation-selective membrane channel. Proc. Natl. Acad. Sci.U.S.A. 100, 13940–13945. doi: 10.1073/pnas.1936192100
Ochs, A. R., Karathanos, T. V., Trayanova, N. A., and Boyle, P. M. (2021). Optogenetic stimulation using anion channelrhodopsin (GtACR1) facilitates termination of reentrant arrhythmias with low light energy requirements: A computational study. Front. Physiol. 12:718622. doi: 10.3389/fphys.2021.718622
Oesterhelt, D., and Stoeckenius, W. (1971). Rhodopsin-like protein from the purple membrane of Halobacterium halobium. Nat. New Biol. 233, 149–152. doi: 10.1038/newbio233149a0
Oesterhelt, D., and Stoeckenius, W. (1973). Functions of a New Photoreceptor Membrane. Proc. Natl. Acad. Sci. U.S.A. 70, 2853–2857. doi: 10.1073/pnas.70.10.2853
Owen, S. F., Liu, M. H., and Kreitzer, A. C. (2019). Thermal constraints on in vivo optogenetic manipulations. Nat. Neurosci. 22, 1061–1065. doi: 10.1038/s41593-019-0422-3
Panzer, S., Zhang, C., Konte, T., Bräuer, C., Diemar, A., Yogendran, P., et al. (2021). Modified rhodopsins from Aureobasidium pullulans excel with very high proton-transport rates. Front. Mol. Biosci. 8:750528. doi: 10.3389/fmolb.2021.750528
Prosseda, P. P., Tran, M., Kowal, T., Wang, B., and Sun, Y. (2022). Advances in ophthalmic optogenetics: Approaches and applications. Biomolecules 12:269. doi: 10.3390/biom12020269
Rajasethupathy, P., Sankaran, S., Marshel, J. H., Kim, C. K., Ferenczi, E., Lee, S. Y., et al. (2015). Projections from neocortex mediate top-down control of memory retrieval. Nature 526, 653–659. doi: 10.1038/nature15389
Roy, D. S., Arons, A., Mitchell, T. I., Pignatelli, M., Ryan, T. J., and Tonegawa, S. (2016). Memory retrieval by activating engram cells in mouse models of early Alzheimer’s disease. Nature 531, 508–512. doi: 10.1038/nature17172
Sahel, J.-A., Boulanger-Scemama, E., Pagot, C., Arleo, A., Galluppi, F., Martel, J. N., et al. (2021). Partial recovery of visual function in a blind patient after optogenetic therapy. Nat. Med. 27, 1223–1229. doi: 10.1038/s41591-021-01351-4
Scheltens, P., De Strooper, B., Kivipelto, M., Holstege, H., Chételat, G., Teunissen, C. E., et al. (2021). Alzheimer’s disease. Lancet 397, 1577–1590. doi: 10.1016/s0140-6736(20)32205-4
Selkoe, D. J., and Hardy, J. (2016). The amyloid hypothesis of Alzheimer’s disease at 25 years. EMBO Mol. Med. 8, 595–608. doi: 10.15252/emmm.201606210
Sengupta, A., Chaffiol, A., Macé, E., Caplette, R., Desrosiers, M., Lampič, M., et al. (2016). Red-shifted channelrhodopsin stimulation restores light responses in blind mice, macaque retina, and human retina. EMBO Mol. Med. 8, 1248–1264. doi: 10.15252/emmm.201505699
Shahmoradian, S. H., Lewis, A. J., Genoud, C., Hench, J., Moors, T. E., Navarro, P. P., et al. (2019). Lewy pathology in Parkinson’s disease consists of crowded organelles and lipid membranes. Nat. Neurosci. 22, 1099–1109. doi: 10.1038/s41593-019-0423-2
Sheng, W., Zhao, X., Huang, X., and Yang, Y. (2022). Real-time image processing toolbox for all-optical closed-loop control of neuronal activities. Front. Cell. Neurosci. 16:917713. doi: 10.3389/fncel.2022.917713
Shevchenko, V., Mager, T., Kovalev, K., Polovinkin, V., Alekseev, A., Juettner, J., et al. (2017). Inward H+ pump xenorhodopsin: Mechanism and alternative optogenetic approach. Sci. Adv. 3:e1603187. doi: 10.1126/sciadv.1603187
Skaper, S. D., Facci, L., Zusso, M., and Giusti, P. (2017). Synaptic plasticity, dementia and alzheimer disease. CNS Neurol. Disord. Drug Targets 16, 220–233. doi: 10.2174/1871527316666170113120853
Spudich, E. N., and Spudich, J. L. (1982). Control of transmembrane ion fluxes to select halorhodopsin-deficient and other energy-transduction mutants of Halobacterium halobium. Proc. Natl. Acad. Sci. U.S.A. 79, 4308–4312. doi: 10.1073/pnas.79.14.4308
Tan, P., He, L., Huang, Y., and Zhou, Y. (2022). Optophysiology: Illuminating cell physiology with optogenetics. Physiol. Rev. 102, 1263–1325. doi: 10.1152/physrev.00021.2021
Tremblay, S., Acker, L., Afraz, A., Albaugh, D. L., Amita, H., Andrei, A. R., et al. (2020). An open resource for non-human primate optogenetics. Neuron 108, 1075–1090.e6. doi: 10.1016/j.neuron.2020.09.027
Vierock, J., Rodriguez-Rozada, S., Dieter, A., Pieper, F., Sims, R., Tenedini, F., et al. (2021). BiPOLES is an optogenetic tool developed for bidirectional dual-color control of neurons. Nat. Commun. 12:4527. doi: 10.1038/s41467-021-24759-5
Wietek, J., Wiegert, J. S., Adeishvili, N., Schneider, F., Watanabe, H., Tsunoda, S. P., et al. (2014). Conversion of channelrhodopsin into a light-gated chloride channel. Science 344, 409–412. doi: 10.1126/science.1249375
Xie, Z., Yang, Q., Song, D., Quan, Z., and Qing, H. (2019). Optogenetic manipulation of astrocytes from synapses to neuronal networks: A potential therapeutic strategy for neurodegenerative diseases. Glia 68, 215–226. doi: 10.1002/glia.23693
Yizhar, O., Fenno, L. E., Prigge, M., Schneider, F., Davidson, T. J., O’Shea, D. J., et al. (2011). Neocortical excitation/inhibition balance in information processing and social dysfunction. Nature 477, 171–178. doi: 10.1038/nature10360
Yoshida, K., Tsunoda, S. P., Brown, L. S., and Kandori, H. (2017). A unique choanoflagellate enzyme rhodopsin exhibits light-dependent cyclic nucleotide phosphodiesterase activity. J. Biol. Chem. 292, 7531–7541. doi: 10.1074/jbc.m117.775569
Zabelskii, D., Alekseev, A., Kovalev, K., Rankovic, V., Balandin, T., Soloviov, D., et al. (2020). Viral rhodopsins 1 are an unique family of light-gated cation channels. Nat. Commun. 11:5707. doi: 10.1038/s41467-020-19457-7
Zeng, F.-G., Rebscher, S., Harrison, W., Sun, X., and Feng, H. (2008). Cochlear implants: System design, integration, and evaluation. IEEE Rev. Biomed. Eng. 1, 115–142. doi: 10.1109/rbme.2008.2008250
Zhang, F., Prigge, M., Beyrière, F., Tsunoda, S. P., Mattis, J., Yizhar, O., et al. (2008). Red-shifted optogenetic excitation: A tool for fast neural control derived from Volvox carteri. Nat. Neurosci. 11, 631–633. doi: 10.1038/nn.2120
Zhang, F., Wang, L.-P., Brauner, M., Liewald, J. F., Kay, K., Watzke, N., et al. (2007). Multimodal fast optical interrogation of neural circuitry. Nature 446, 633–639. doi: 10.1038/nature05744
Keywords: optogenetics, microbial rhodopsin, Alzheimer’s disease, Parkinson’s disease, cardiac optogenetics, optical cochlear implant
Citation: Zhang H, Fang H, Liu D, Zhang Y, Adu-Amankwaah J, Yuan J, Tan R and Zhu J (2022) Applications and challenges of rhodopsin-based optogenetics in biomedicine. Front. Neurosci. 16:966772. doi: 10.3389/fnins.2022.966772
Received: 11 July 2022; Accepted: 05 September 2022;
Published: 23 September 2022.
Edited by:
Marco Canepari, UMR 5588 Laboratoire Interdisciplinaire de Physique (LIPhy), FranceReviewed by:
John Spudich, University of Texas Health Science Center at Houston, United StatesCopyright © 2022 Zhang, Fang, Liu, Zhang, Adu-Amankwaah, Yuan, Tan and Zhu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jinxiang Yuan, eXVhbmppbnhpYW5nMThAMTYzLmNvbQ==; Rubin Tan, dGFucnViaW4xMUAxMjYuY29t; Jianping Zhu, d2x6anAyMDAyQDE2My5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.