
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Neurosci. , 19 March 2021
Sec. Neurodegeneration
Volume 15 - 2021 | https://doi.org/10.3389/fnins.2021.620929
 Hui-Hui Fan1,2Qing Guo1
Hui-Hui Fan1,2Qing Guo1 Jing Zheng1
Jing Zheng1 Yi-Zhi Lian2
Yi-Zhi Lian2 Shi-Shi Huang2Yue Sun2Ming Zou2
Shi-Shi Huang2Yue Sun2Ming Zou2 Jian-Hong Zhu1,2*
Jian-Hong Zhu1,2* Xiong Zhang1,2*
Xiong Zhang1,2*Background: Studies in animal models have suggested that aldehyde dehydrogenase 1 (encoded by ALDH1A1) protects against Parkinson’s disease (PD) by reducing toxic metabolites of dopamine. Herein we aimed to investigate whether ALDH1A1 was genetically associated with PD susceptibility in humans.
Methods: A Han Chinese population of 1,039 subjects was recruited to analyze six tag-single nucleotide polymorphisms (SNPs), followed by haplotype analyses and variants interaction analyses. Real-time PCR was used to analyze mRNA levels of ALDH1A1 in peripheral blood of 42 subjects.
Results: The tag-SNP rs7043217 of ALDH1A1 was significantly associated with PD susceptibility with the T serving as a risk allele (genotype frequency, P = 0.030; allele frequency, P = 0.013, OR = 1.258, 95% CI = 1.050–1.508). Multiple haplotypes were linked to abnormalities of PD risk, topped by a 4-SNP GGTA module in the order of rs4646547, rs1888202, rs7043217, and rs647880 (P = 9.610 × 10–8, OR = 6.420, 95% CI = 2.944–13.998). Interaction analyses showed that a simultaneous presence of the CC genotype of rs7043217 and the TT genotype of ALDH2 variant rs4767944 conferred an elevated protection against PD (P = 4.68 × 10–4, OR = 0.378, 95% CI = 0.219–0.652). The mRNA expression of ALDH1A1 showed a trend of reduction (P = 0.084) in PD patients compared to the controls.
Conclusion: Our results provide novel genetic insights into the role of ALDH1 in PD pathogenesis.
Parkinson’s disease (PD) is mainly of sporadic origin and the second most common neurodegenerative disorder characterized by the progressive loss of dopaminergic neurons in the substantia nigra pars compacta (SNpc). Besides familial PD cases which are caused by monogenic mutations such as in SNCA and LRRK2, the etiology of sporadic PD remains largely unknown but is considered to be influenced by both genetic factors and environmental exposure (Kalia and Lang, 2015).
Aldehyde dehydrogenase (ALDH) is a family of enzymes that catalyze the oxidation of aldehydes to their corresponding carboxylic acids. In dopaminergic neurons, ALDH converts the dopamine metabolite 3,4-dihydroxyphenylacetaldehyde (DOPAL) into 3,4-dihydroxyphenylacetic acid (DOPAC), which is further metabolized to form homovanillic acid (Grunblatt and Riederer, 2016; Masato et al., 2019). DOPAL is neurotoxic by triggering α-synuclein aggregates, thus provoking the loss of dopaminergic neurons (Marchitti et al., 2007; Burke et al., 2008). It has been shown that benomyl, a pesticide, inhibits ALDH activity in rat primary cells of SNpc, leading to increased DOPAL production and cytotoxicity (Fitzmaurice et al., 2013).
ALDH has a number of isoforms. Amidst, ALDH1 is cytosolic and highly expressed in dopaminergic neurons. ALDH2 is mitochondrion-located and moderately expressed in these neurons (Cai et al., 2014). Genetic studies of PD patients indicate that a clade of ALDH2 haplotypes by pesticide exposure exacerbates predisposition to this disease (Fitzmaurice et al., 2014), and the rs4767944 variant of ALDH2 is associated with PD risk in Han Chinese (Zhang et al., 2015). On the other hand, ALDH1-positive dopaminergic neurons are reported to be more resistant than the negative ones to α-synuclein-mediated cytotoxicity in mouse SNpc (Liu et al., 2014). Moreover, ALDH1 mRNA and protein are found to be down-regulated in the substantia nigra of post-mortem PD patients (Galter et al., 2003; Grunblatt et al., 2004; Werner et al., 2008). ALDH1 is highly expressed in dopaminergic neurons (Cai et al., 2014) and appears to be the pivotal isoform of ALDH in protection against PD. Nonetheless, an important question to be answered is whether ALDH1A1 is genetically associated with PD in humans. A Han Chinese cohort was thus recruited to address this hypothesis by analyzing six tag-single nucleotide polymorphisms (SNPs) of ALDH1A1.
A total of 1,039 Han Chinese were included for tag-SNP analysis from eastern China, including 506 sporadic PD patients (259 males and 247 females) and 533 controls (282 males and 251 females). The median age of the patients and controls was 67 (interquartile range, 60–74) and 58 (interquartile range, 50–68), respectively. For quantitation of ALDH1A1 mRNA levels, another 19 sporadic PD patients (10 males and nine females; median age, 64, interquartile range, 58–68) and 23 control subjects (11 males and 12 females; median age, 59, interquartile range, 54–70) were recruited. PD patients were diagnosed by two neurologists according to the United Kingdom Parkinson’s Disease Society Brain Bank Criteria (Hughes et al., 1992). Patients with a family history of PD, or with secondary and atypical Parkinsonism were excluded. Control subjects were free of neurological disorders determined by medical history, physical and auxiliary laboratory examinations of blood routine, blood lipid, blood sugar, liver, and kidney functions. Informed written consents were obtained from all participants. The study was approved by the Ethics Committee of The Second Affiliated Hospital and Yuying Children’s hospital, Wenzhou Medical University (2017-31).
Six variants of ALDH1A1 tag-SNP, including rs4646547 (T > G), rs1888202 (G > C), rs348471 (C > T), rs7043217 (C > T), rs647880 (A > G), and rs8187876 (G > A), were selected using Haploview v.4.2 as described previously (Zhang et al., 2015). The construction criteria were minor allele frequency (MAF) ≥ 0.1 and r2 > 0.8 (Supplementary Figure 1).
Genomic DNA was extracted from peripheral blood samples using TIANamp Genomic DNA Kit (Tiangen, Beijing, China) according to the manufacturer’s instruction. Five of the tag-SNPs (rs4646547, rs348471, rs7043217, rs647880, and rs8187876) were analyzed by the polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP). The rs1888202 variant was analyzed by Sanger sequencing due to a lack of restriction enzyme recognition site. Primers and restriction enzymes (New England Biolabs, Ipswich, MA, United States) were detailed in Supplementary Table 1.
Total RNA was extracted using RNAiso plus (Takara, Shiga, Japan). An amount of 400 ng RNA was used for synthesis of the first-strand cDNA by PrimeScript RT reagent Kit (Takara, Shiga, Japan). PCR amplification was performed using ChamQ Universal SYBR qPCR Master Mix (Vazyme, Nanjing, China) with the following conditions: 95°C for 10 min, 40 cycles of 95°C for 10 s, 60°C for 15 s, and 72°C for 20 s, and a final extension of 72°C for 10 min. Primers were detailed in Supplementary Table 1. The mRNA levels were expressed as 2–ΔCT following normalization to their respective ACTB.
Statistical analyses were performed using the Statistical Package for Social Science Program (SPSS for Windows, version 23.0). Hardy–Weinberg equilibrium in genotype distribution was assessed using χ2 test. Following Kolmogorov-Smirnov test for normality, age difference was evaluated by Mann–Whitney U test, and levels of gene expression were expressed as mean ± standard deviation and analyzed by student’s T-test. The difference in genotype and allele frequencies, and the interaction between genes were analyzed using logistic regression model with gender and age as covariates. The χ2 test was used to assess differences in gender, and haplotype frequencies between the PD cases and controls. The haplotype analysis was not adjusted for gender and age, because individual haplotypes are unable to be deduced from subjects with two or more heterozygous SNP sites, and thus individual haplotype assignment would induce additional assumptions that are less reliable (Zhang et al., 2004; Woo et al., 2005).
The haplotype construction and association analysis were performed using SHEsis Online Version1 (Shi and He, 2005). A backward elimination method was used to identify the highest-risk haplotype for PD (Francis et al., 2007). In brief, all the six tag-SNPs were used to produce the first haplotype model. One SNP was then removed each time to create 5-SNP models. Of these, the one with the lowest associated P value was deemed the best model for next round of elimination. As such, the best 4-, 3-, and 2-SNP models were determined. A two-tailed P value < 0.05 was considered statistically significant.
Genotype distributions of the six tag-SNPs were in accordance with Hardy-Weinberg equilibrium (P > 0.05). The PD cases and controls were comparable in gender (P > 0.05) but different in age (P < 0.05). Results showed that genotype and allele frequencies of rs4646547, rs1888202, rs647880, and rs8187876 were comparable (P > 0.05) between the PD cases and controls (Table 1). The rs348471 significantly differed (P = 0.045) in genotype but not in allele frequency. In contrast, significant differences were found in both genotype (P = 0.030) and allele (P = 0.013) frequencies of rs7043217 between the cases and controls, with the T allele serving as a risk allele for PD (OR = 1.258, 95% CI = 1.050–1.508). Analyzing of rs7043217 using three genetic models (additive, dominant, and recessive) showed that rs7043217 was significantly associated with PD in the dominant model (P = 0.009; Table 2) and additive model (P = 0.015; Table 2), further suggesting that the T allele was a risk allele for PD.
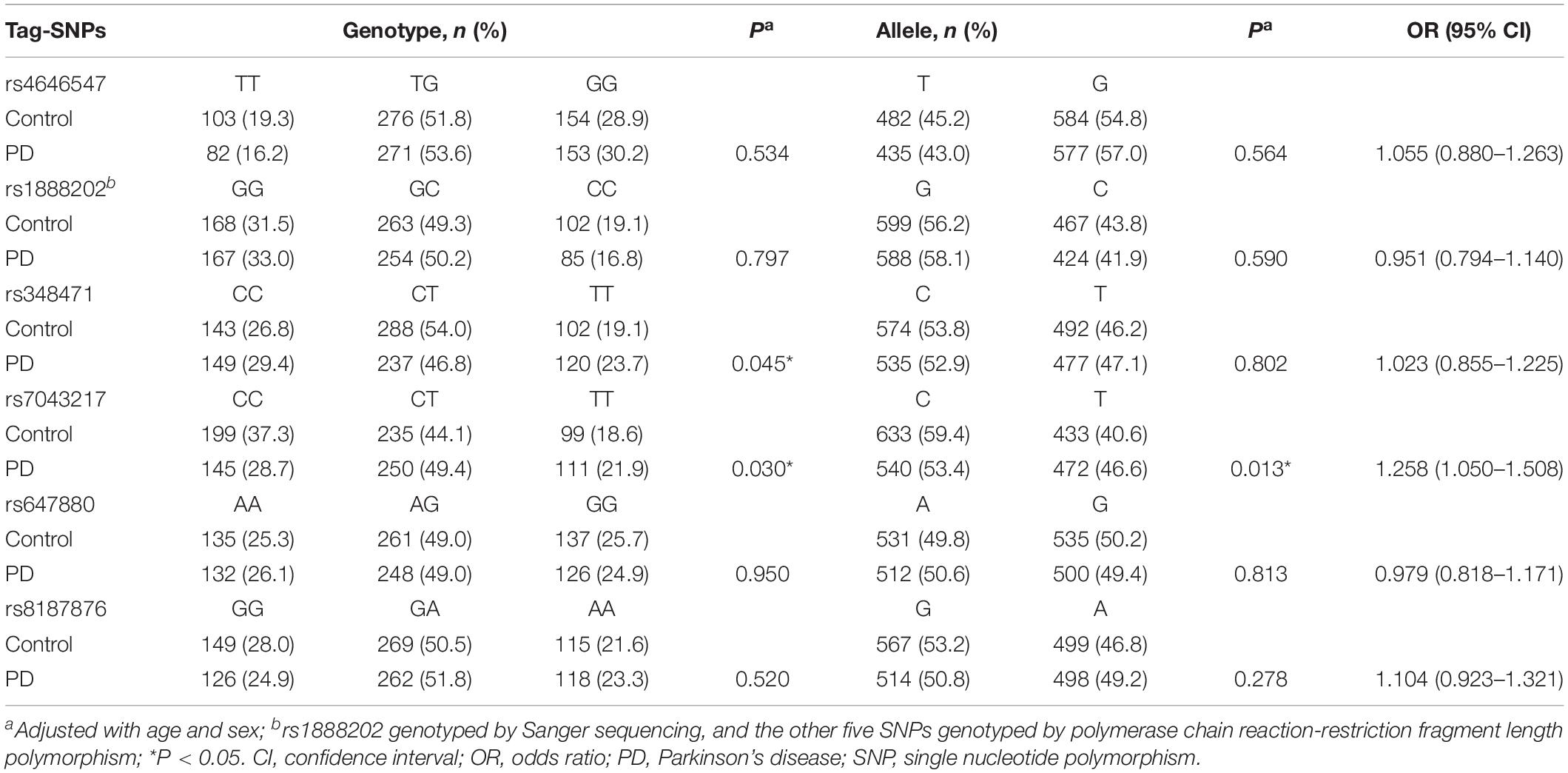
Table 1. Genotype and allele frequencies of the six ALDH1A1 tag-SNPs in PD patients and controls.
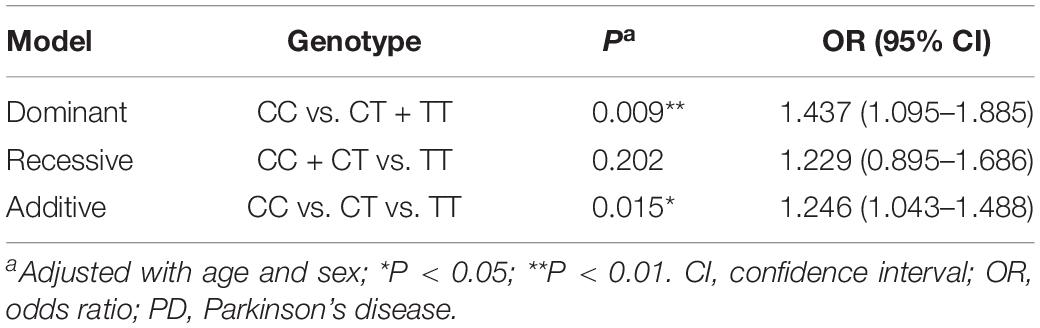
Table 2. Association between rs7043217 and PD using dominant, recessive and additive models.
A total of 415 PD patients with subtype records were classified into three subtypes, that is, tremor dominant, postural instability/gait difficulty, and indeterminate. Association analysis of the SNPs with PD subtypes showed that the associations were present in the genotype frequency of rs7043217 between patients of the tremor-dominant subtype and the controls (P = 0.007), and in the allele frequency of rs7043217 between patients of the postural instability/gait difficulty subtype and the controls (P = 0.032, OR = 1.329, 95% CI, 1.025–1.722; Supplementary Table 2).
We analyzed whether ALDH1A1 haplotypes of the tag-SNPs were associated with the risk of PD. Haplotypes were constructed following the order of rs4646547, rs1888202, rs348471, rs7043217, rs647880, and rs8187876, and those with a frequency < 3.0% in both PD patients and controls were excluded. The remaining 10 haplotypes were listed in Table 3. A significant difference (P = 2.880 × 10–11) in overall haplotype distribution was observed between the PD patients and controls. Four haplotypes, including GGCTAG, GGCCGA, GCCTGG, and GCCCAG, displayed a statistically significant difference between the cases and controls. In particular, the haplotype GGCTAG rendered an increased risk of PD (P = 8.540 × 10–6, OR = 5.619, 95% CI = 2.397–13.172), while the GCCCAG carriers might be resistant to PD (P = 1.400 × 10–8, OR = 0.133, 95% CI = 0.059–0.299).
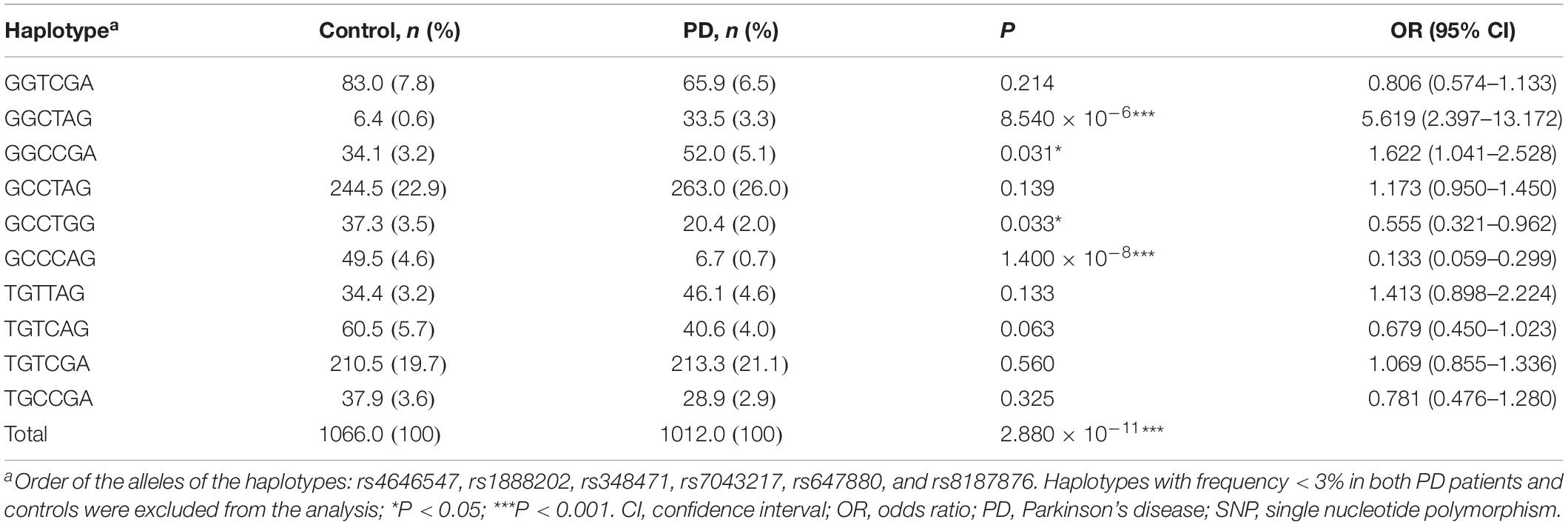
Table 3. Haplotype analysis of ALDH1A1 tag-SNPs in PD patients and controls.
We further analyzed the highest-risk haplotype of ALDH1A1 toward PD using backward elimination models. Results showed that the strongest PD-associated haplotype was the GGTA in order of rs4646547, rs1888202, rs7043217, and rs647880 from the best 4-SNP model (P = 9.610 × 10–8, OR = 6.420, 95% CI = 2.944–13.998; Table 4). The frequencies of the GGTA were 0.7 and 4.4%, respectively, in the controls and PD patients.
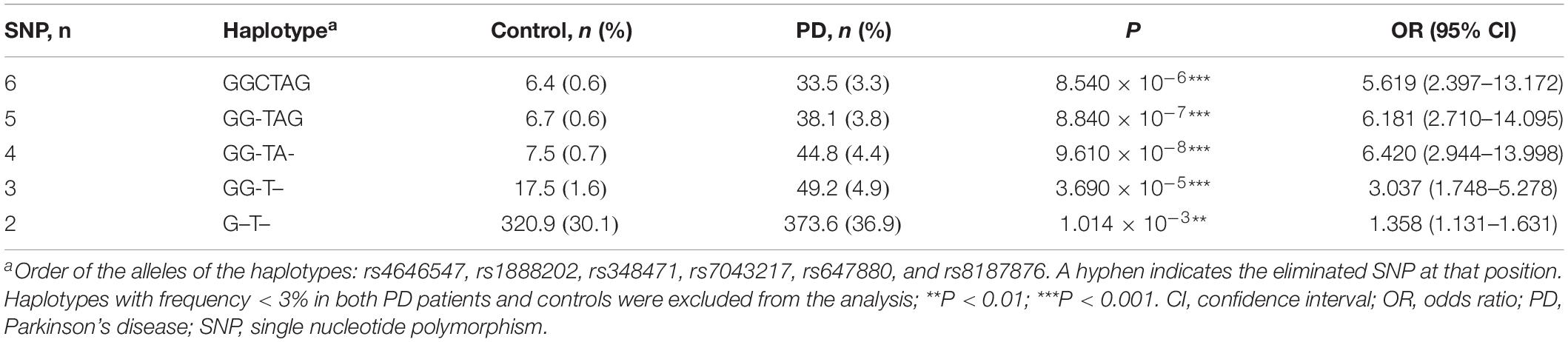
Table 4. The highest-risk haplotype analysis of ALDH1A1 in association with PD.
We have previously discovered that the rs4767944 variant of ALDH2 is associated with PD risk (Zhang et al., 2015), where samples of a total of 766 subjects were also used in the current study, including 402 controls and 364 PD patients. To characterize whether ALDH1A1 and ALDH2 were genetically interplayed, this overlapped subgroup was extracted to analyze the interaction between rs7043217 and rs4767944. Results showed that a simultaneous presence of the CC genotype of rs7043217 and the TT genotype of rs4767944 conferred a strong protection against PD (P = 4.68 × 10–4, OR = 0.378, 95% CI = 0.219–0.652; Table 5).
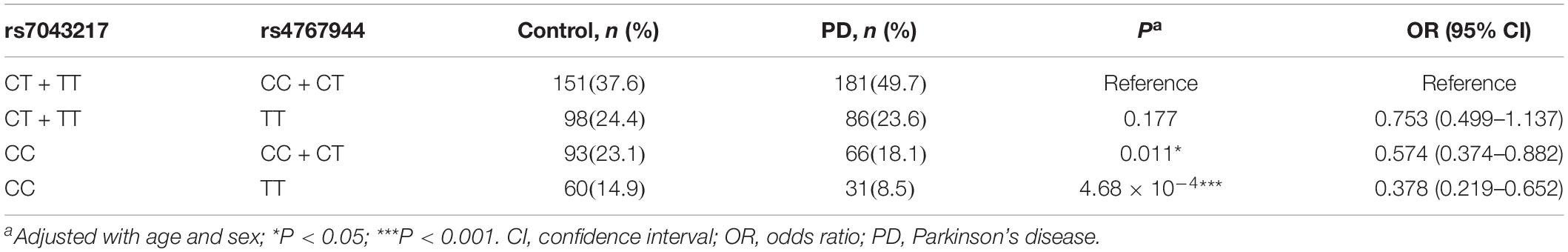
Table 5. Interaction analysis between rs7043217 of ALDH1A1 and rs4767944 of ALDH2 on risk for PD.
We also performed the interaction analysis between the other five tag-SNPs of ALDH1A1 and rs4767944 of ALDH2. Since these five tag-SNPs were not significantly associated with PD, both recessive and dominant models were used. Results showed that the higher interaction was present between rs348471 and rs4767944, where the TT genotype of rs348471 and the CC + TT genotypes of rs4767944 led to an aggravated PD risk (P = 0.005, OR = 1.930, 95% CI = 1.220–3.055; Supplementary Tables 3, 4).
It was previously reported that reduced mRNA expression of ALDH1A1 was a potential blood biomarker of PD (Molochnikov et al., 2012). To replicate this finding, we analyzed ALDH1A1 mRNA levels in peripheral blood of 19 PD patients and 23 controls with sex and age being matched (P > 0.05). Results of real-time PCR showed a trend of decrease in ALDH1A1 mRNA levels in the PD patients (P = 0.084; Supplementary Figure 2).
Animal studies have provided strong evidence linking ALDH1 to PD by dissecting ALDH1-positive neuronal populations in the substantia nigra (Liu et al., 2014). However, it remains elusive whether ALDH1 is genetically associated with PD in humans. By systematically analyzing six tag-SNPs of the ALDH1-encoding gene ALDH1A1 in a Han Chinese population, we demonstrate that the SNP rs7043217 and the haplotypes of GGCTAG and GGTA are genetically associated with PD susceptibility.
The current study represents the first effort in understanding the genetic association between ALDH1A1 and PD. Although ALDH1 can catalyze DOPAL oxidation in the brain, the best-known function of this enzyme is to regulate alcohol metabolism. Indeed, genetic variants of ALDH1A1 have been reported in the context of alcohol drinking. For instance, compared to its T allele, the C allele of rs7043217 is associated with alcohol metabolism capacity in European Americans (Sherva et al., 2009). The G allele of rs610529, which is in linkage disequilibrium with the C allele of rs7043217, is also related to the development of alcohol addiction (Lind et al., 2008). Since a low activity of ALDH protects individuals from alcohol dependence (Luczak et al., 2006), the above results indicate that the C allele of rs7043217 may be associated with high or normal expression or activity of ALDH1. In line with the observation of increased PD risk carrying the rs7043217/T allele, the low ALDH1 activity could lead to DOPAL accumulation in the brain and promote PD pathogenesis (Marchitti et al., 2007).
The association of rs7043217 with overall PD is present in both genotype and allele frequencies. A further PD subtype analysis suggests such an association of rs7043217 is in genotype frequency of the tremor-dominant subtype, and in allele frequency of the postural instability/gait difficulty subtype. Common genetic models include the dominant, recessive, additive, over-dominant, and multiplicative models. All of them are genotype-based, suggesting that genotype frequencies are most commonly used in genetic association studies (Dorak, 2017). However, under multiplicative and additive genetic models, analyzing by allele frequencies provides a more powerful method of testing, suggesting that the allele frequency may be a better indicator in these models (Lewis, 2002). Therefore, both genotype and allele frequencies could serve as an important indicator for certain genetic associations.
It is known that haplotypes are more powerful than individual variants in suggesting genetic association (Gabriel et al., 2002). Indeed, analysis of the six tag-SNPs identifies two haplotypes greatly associated with PD: the GGCTAG serving as a risk factor and the GCCCAG as a protective factor. Further analysis of the effect SNPs suggests that the GGTA of rs4646547, rs1888202, rs7043217, and rs647880 is the highest ranked risk haplotype to discriminate PD, indicating a potential value for genetic diagnosis of PD. Further validation is needed in other populations.
After searching genome-wide association study data, we found a marginal P value, 0.058, of rs7043217 in association with PD (Nalls et al., 2019). Given the ALDH is a type of metabolic enzymes, the discrepancy of significance between the studies may result from confounding factors such as differential environmental exposures and interactions with other genetic susceptibilities. For instance, ALDH is susceptible to the inhibition of pesticides. A clade of ALDH2 haplotypes is shown to exacerbate the risk of PD predisposition by pesticide exposure, but ALDH2 per se is not associated with this disorder (Fitzmaurice et al., 2014). Indeed, there are severe abuses of pesticides in China, especially in the East (Jin and Zhou, 2018; Yu et al., 2020), which thus may contribute to the elevated association between rs7043217 of ALDH1A1 and PD in our cohort. Unfortunately, a lack of pesticide exposure history impedes us from further assessment in this regard. In addition, mice deficient in both Aldh1a1 and Aldh2 exhibit age-dependent loss of dopaminergic neurons in the SNpc and motor dysfunctions (Wey et al., 2012). In contrast, genetic deletion of Aldh1a1 in mice fails to reproduce any overt motor symptoms or loss of dopaminergic neuron in SNpc (Anderson et al., 2011), while no data regarding mice deficient in Aldh2 per se are available. These results indicate that ALDH1 and ALDH2 may collectively aggravate PD pathogenesis. Such an interplay is also suggested by our genetic interaction analysis between the tag-SNPs of ALDH1A1 and rs4767944 of ALDH2, with the highest interplay present in the variant rs7043217. The obtained ALDH1A1 genetic variations thus do not monogenically cause PD.
Besides in the substantia nigra of PD patients (Galter et al., 2003; Grunblatt et al., 2004; Werner et al., 2008), the mRNA expression of ALDH1A1 is also shown to be reduced in the peripheral blood of sporadic PD patients, and the gene is classified as an optimal predictor for PD risk in a panel with four other genes (Molochnikov et al., 2012). Our results appeared to be in support of such a reduction in PD patients. Nonetheless, a further confirmation in large sample size may be needed. Considering its protective effects on dopaminergic neurons (Liu et al., 2014), the reduced expression of ALDH1A1 may be a cause rather than a consequence to PD. The associated mechanism likely involves its detoxicating catalyzation against aldehyde risk factors. We herein disclose a genetic association between ALDH1A1 and PD, represented by the tag-SNP rs7043217. However, all of the 19 SNPs in its linkage disequilibrium block (Supplementary Figure 1) are either located in introns of ALDH1A1 or a synonymous polymorphism in an exon. It remains to be further determined whether certain SNPs in this block and the associated haplotypes may functionally alter the ALDH1A1 expression or ALDH1 activity. Indeed, one of the limitations of genetic association studies is that the identified SNPs may not be causative or functional, but they sometimes represent the only practical approach to begin to address a particular biological hypothesis (Hegele, 2002).
In conclusion, the current study demonstrates that PD susceptibility in Han Chinese is associated with ALDH1A1 genetic variations, notable examples of which include the tag-SNP rs7043217 and the haplotypes GGCTAG and GGTA. These findings provide genetic insights into the role of ALDH1 in PD pathogenesis, and facilitate the development of screening strategies targeting the alleles susceptible to PD for early control of the disease.
The original contributions presented in the study are publicly available. This data can be found here: https://datadryad.org/stash/share/iPw_jVzOJqOBMAtM_fGeH-TpxEwjB1aV93LU3eBugh8.
The studies involving human participants were reviewed and approved by Ethics Committee of The Second Affiliated Hospital and Yuying Children’s hospital, Wenzhou Medical University. The patients/participants provided their written informed consent to participate in this study.
J-HZ and XZ designed the research. H-HF, QG, JZ, and S-SH conducted the experiments. H-HF, Y-ZL, and YS analyzed the data. XZ and MZ contributed to the samples. H-HF and J-HZ wrote the manuscript. All authors have read and approved the final manuscript.
This work was supported by the National Natural Science Foundation of China (31701080, 81771380, and 81771510), Zhejiang Provincial Natural Science Foundation (LD19H090001 and LZ19H090002), and Science and Technology Department of Zhejiang Province (2018C37111).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors are thankful to their colleagues for clinical data collection and management of patients. The authors appreciate all of the subjects participating in this study.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnins.2021.620929/full#supplementary-material
Anderson, D. W., Schray, R. C., Duester, G., and Schneider, J. S. (2011). Functional significance of aldehyde dehydrogenase ALDH1A1 to the nigrostriatal dopamine system. Brain Res. 1408, 81–87. doi: 10.1016/j.brainres.2011.06.051
Burke, W. J., Kumar, V. B., Pandey, N., Panneton, W. M., Gan, Q., Franko, M. W., et al. (2008). Aggregation of alpha-synuclein by DOPAL, the monoamine oxidase metabolite of dopamine. Acta Neuropathol. 115, 193–203. doi: 10.1007/s00401-007-0303-9
Cai, H., Liu, G., Sun, L., and Ding, J. (2014). Aldehyde Dehydrogenase 1 making molecular inroads into the differential vulnerability of nigrostriatal dopaminergic neuron subtypes in Parkinson’s disease. Transl. Neurodegener. 3:27. doi: 10.1186/2047-9158-3-27
Dorak, M. (2017). Genetic association studies: background, conduct, analysis, interpretation. Garland Science. New York, NY: Taylor & Francis Group.
Fitzmaurice, A. G., Rhodes, S. L., Lulla, A., Murphy, N. P., Lam, H. A., O’Donnell, K. C., et al. (2013). Aldehyde dehydrogenase inhibition as a pathogenic mechanism in Parkinson disease. Proc. Natl. Acad. Sci. U. S. A. 110, 636–641. doi: 10.1073/pnas.1220399110
Fitzmaurice, A. G., Rhodes, S. L., Cockburn, M., Ritz, B., and Bronstein, J. M. (2014). Aldehyde dehydrogenase variation enhances effect of pesticides associated with Parkinson disease. Neurology. 82, 419–426. doi: 10.1212/WNL.0000000000000083
Francis, P. J., Schultz, D. W., Hamon, S., Ott, J., Weleber, R. G., and Klein, M. L. (2007). Haplotypes in the complement factor H (CFH) gene: associations with drusen and advanced age-related macular degeneration. PLoS One. 2:e1197. doi: 10.1371/journal.pone.0001197
Gabriel, S. B., Schaffner, S. F., Nguyen, H., Moore, J. M., Roy, J., Blumenstiel, B., et al. (2002). The structure of haplotype blocks in the human genome. Science 296, 2225–2229. doi: 10.1126/science.1069424
Galter, D., Buervenich, S., Carmine, A., Anvret, M., and Olson, L. (2003). ALDH1 mRNA: presence in human dopamine neurons and decreases in substantia nigra in Parkinson’s disease and in the ventral tegmental area in schizophrenia. Neurobiol. Dis. 14, 637–647. doi: 10.1016/j.nbd.2003.09.001
Grunblatt, E., Mandel, S., Jacob-Hirsch, J., Zeligson, S., Amariglo, N., Rechavi, G., et al. (2004). Gene expression profiling of parkinsonian substantia nigra pars compacta; alterations in ubiquitin-proteasome, heat shock protein, iron and oxidative stress regulated proteins, cell adhesion/cellular matrix and vesicle trafficking genes. J. Neural Transm. 111, 1543–1573. doi: 10.1007/s00702-004-0212-1
Grunblatt, E., and Riederer, P. (2016). Aldehyde dehydrogenase (ALDH) in Alzheimer’s and Parkinson’s disease. J. Neural Transm. 123, 83–90. doi: 10.1007/s00702-014-1320-1
Hegele, R. A. (2002). SNP judgments and freedom of association. Arterioscler. Thromb. Vasc. Biol. 22, 1058–1061. doi: 10.1161/01.atv.0000026801.56080.14
Hughes, A. J., Daniel, S. E., Kilford, L., and Lees, A. J. (1992). Accuracy of clinical diagnosis of idiopathic Parkinson’s disease: a clinico-pathological study of 100 cases. J. Neurol. Neurosurg. Psychiatry. 55, 181–184. doi: 10.1136/jnnp.55.3.181
Jin, S., and Zhou, F. (2018). Zero Growth of Chemical Fertilizer and Pesticide Use: China’s Objectives. Progress and Challenges. J. Resources. Ecol. 9, 50–58. doi: 10.5814/j.issn.1674-764x.2018.01.006
Kalia, L. V., and Lang, A. E. (2015). Parkinson’s disease. Lancet 386, 896–912. doi: 10.1016/s0140-6736(14)61393-3
Lewis, C. M. (2002). Genetic association studies: design, analysis and interpretation. Brief Bioinform. 3, 146–153. doi: 10.1093/bib/3.2.146
Lind, P. A., Eriksson, C. J., and Wilhelmsen, K. C. (2008). The role of aldehyde dehydrogenase-1 (ALDH1A1) polymorphisms in harmful alcohol consumption in a Finnish population. Hum. Genomics. 3, 24–35. doi: 10.1186/1479-7364-3-1-24
Liu, G., Yu, J., Ding, J., Xie, C., Sun, L., Rudenko, I., et al. (2014). Aldehyde dehydrogenase 1 defines and protects a nigrostriatal dopaminergic neuron subpopulation. J. Clin. Invest. 124, 3032–3046. doi: 10.1172/jci72176
Luczak, S. E., Glatt, S. J., and Wall, T. L. (2006). Meta-analyses of ALDH2 and ADH1B with alcohol dependence in Asians. Psychol. Bull. 132, 607–621. doi: 10.1037/0033-2909.132.4.607
Marchitti, S. A., Deitrich, R. A., and Vasiliou, V. (2007). Neurotoxicity and metabolism of the catecholamine-derived 3,4-dihydroxyphenylacetaldehyde and 3,4-dihydroxyphenylglycolaldehyde: the role of aldehyde dehydrogenase. Pharmacol. Rev. 59, 125–150. doi: 10.1124/pr.59.2.1
Masato, A., Plotegher, N., Boassa, D., and Bubacco, L. (2019). Impaired dopamine metabolism in Parkinson’s disease pathogenesis. Mol. Neurodegener. 14:35. doi: 10.1186/s13024-019-0332-6
Molochnikov, L., Rabey, J. M., Dobronevsky, E., Bonucelli, U., Ceravolo, R., Frosini, D., et al. (2012). A molecular signature in blood identifies early Parkinson’s disease. Mol. Neurodegener. 7:26. doi: 10.1186/1750-1326-7-26
Nalls, M. A., Blauwendraat, C., Vallerga, C. L., Heilbron, K., Bandres-Ciga, S., Chang, D., et al. (2019). Identification of novel risk loci, causal insights, and heritable risk for Parkinson’s disease: a meta-analysis of genome-wide association studies. Lancet Neurol. 18, 1091–1102. doi: 10.1016/S1474-4422(19)30320-5
Sherva, R., Rice, J. P., Neuman, R. J., Rochberg, N., Saccone, N. L., and Bierut, L. J. (2009). Associations and interactions between SNPs in the alcohol metabolizing genes and alcoholism phenotypes in European Americans. Alcohol. Clin. Exp. Res. 33, 848–857. doi: 10.1111/j.1530-0277.2009.00904.x
Shi, Y. Y., and He, L. (2005). SHEsis, a powerful software platform for analyses of linkage disequilibrium, haplotype construction, and genetic association at polymorphism loci. Cell Res. 15, 97–98. doi: 10.1038/sj.cr.7290272
Werner, C. J., Haussen, R. H. V., Mall, G., and Wolf, S. (2008). Proteome analysis of human substantia nigra in Parkinson’s disease. Proteome Science. 6:8. doi: 10.1186/1477-5956-6-8
Wey, M. C.-Y., Fernandez, E., Martinez, P. A., Sullivan, P., Goldstein, D. S., and Strong, R. (2012). Neurodegeneration and Motor Dysfunction in Mice Lacking Cytosolic and Mitochondrial Aldehyde Dehydrogenases: Implications for Parkinson’s Disease. PLoS One 7:e31522. doi: 10.1371/journal.pone.0031522
Woo, D., Kaushal, R., Chakraborty, R., Woo, J., Haverbusch, M., Sekar, P., et al. (2005). Association of apolipoprotein E4 and haplotypes of the apolipoprotein E gene with lobar intracerebral hemorrhage. Stroke 36, 1874–1879. doi: 10.1161/01.STR.0000177891.15082.b9
Yu, H. Y., Liu, Y. F., Shu, X. Q., Ma, L. M., and Pan, Y. W. (2020). Assessment of the spatial distribution of organochlorine pesticides (OCPs) and polychlorinated biphenyls (PCBs) in urban soil of China. Chemosphere 243:11. doi: 10.1016/j.chemosphere.2019.125392
Zhang, J., Cui, Y., Kuang, G., Li, Y., Wang, N., Wang, R., et al. (2004). Association of the thymidylate synthase polymorphisms with esophageal squamous cell carcinoma and gastric cardiac adenocarcinoma. Carcinogenesis 25, 2479–2485. doi: 10.1093/carcin/bgh247
Keywords: ALDH1A1, Parkinson’s disease, single nucleotide polymorphism, rs7043217, genetic association
Citation: Fan H-H, Guo Q, Zheng J, Lian Y-Z, Huang S-S, Sun Y, Zou M, Zhu J-H and Zhang X (2021) ALDH1A1 Genetic Variations May Modulate Risk of Parkinson’s Disease in Han Chinese Population. Front. Neurosci. 15:620929. doi: 10.3389/fnins.2021.620929
Received: 30 November 2020; Accepted: 10 February 2021;
Published: 19 March 2021.
Edited by:
Luigi Bubacco, University of Padua, ItalyReviewed by:
Qian Li, Washington University School of Medicine in St. Louis, United StatesCopyright © 2021 Fan, Guo, Zheng, Lian, Huang, Sun, Zou, Zhu and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jian-Hong Zhu, amh6aHVAd211LmVkdS5jbg==; Xiong Zhang, emhhbmd4aW9uZzk4QGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.