 Fengzhen Hou
Fengzhen Hou Zhinan Yu1
Zhinan Yu1 Chung-Kang Peng
Chung-Kang Peng Albert Yang
Albert Yang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Neurosci. , 13 November 2018
Sec. Brain Imaging Methods
Volume 12 - 2018 | https://doi.org/10.3389/fnins.2018.00809
This article is part of the Research Topic Advances in Multi-Scale Analysis of Brain Complexity View all 14 articles
Sleep electroencephalography (EEG) provides an opportunity to study sleep scientifically, whose chaotic, dynamic, complex, and dissipative nature implies that non-linear approaches could uncover some mechanism of sleep. Based on well-established complexity theories, one hypothesis in sleep medicine is that lower complexity of brain waves at pre-sleep state can facilitate sleep initiation and further improve sleep quality. However, this has never been studied with solid data. In this study, EEG collected from healthy subjects was used to investigate the association between pre-sleep EEG complexity and sleep quality. Multiscale entropy analysis (MSE) was applied to pre-sleep EEG signals recorded immediately after light-off (while subjects were awake) for measuring the complexities of brain dynamics by a proposed index, CI1−30. Slow wave activity (SWA) in sleep, which is commonly used as an indicator of sleep depth or sleep intensity, was quantified based on two methods, traditional Fast Fourier transform (FFT) and ensemble empirical mode decomposition (EEMD). The associations between wake EEG complexity, sleep latency, and SWA in sleep were evaluated. Our results demonstrated that lower complexity before sleep onset is associated with decreased sleep latency, indicating a potential facilitating role of reduced pre-sleep complexity in the wake-sleep transition. In addition, the proposed EEMD-based method revealed an association between wake complexity and quantified SWA in the beginning of sleep (90 min after sleep onset). Complexity metric could thus be considered as a potential indicator for sleep interventions, and further studies are encouraged to examine the application of EEG complexity before sleep onset in populations with difficulty in sleep initiation. Further studies may also examine the mechanisms of the causal relationships between pre-sleep brain complexity and SWA, or conduct comparisons between normal and pathological conditions.
Sleep medicine has been increasingly recognized as an important discipline in recent decades; however, the current limitations of electroencephalography (EEG)-based sleep analysis and quantification may have led to ongoing controversy. Sleep is a complex physiological process that involves functions of every organ system at different levels. Alternative metrics have been proposed, aiming to provide insights into the dynamics of sleep. In sleep medicine, EEG is one of the most frequently recorded biological signals, but it is mainly used as the basis for scoring sleep stages in sleep laboratories and clinics, as sleep is classified as either rapid-eye-movement (REM) sleep or non-REM (NREM) sleep (including sleep stages N1, N2, and N3). Owing to the non-linear and dynamic features of EEG, non-linear approaches may lead to better understanding of the profound complexity of sleep.
Non-linear dynamics theory provides new opportunities for understanding the behavior of EEG (Acharya et al., 2005). Previously, EEG has been used to mark features of sleep (He et al., 2005; Janjarasjitt et al., 2008; Yeh et al., 2013; Abeysuriya et al., 2014), and it has been reported that nonlinearity depends on sleep stage (Shen et al., 2003). Recently, studies have increasingly used non-linear methods to investigate the nature of brain activities during sleep (Ma et al., 2018), but there are still limitations in the existing literature. The full advantages of non-linear approaches have yet to be determined (Ma et al., 2018).
According to the complexity theories, somewhat higher complexity is associated with relatively improved health conditions and greater chances of survival (Costa et al., 2002a, 2005), while a reduction in or loss of complexity is often associated with imbalance or disturbed physiological conditions, usually implying disease or aging (Goldberger et al., 2002). In many studies, complexity-based metrics of the dynamics of a physiological system have demonstrated better prognostic power (Mejaddam et al., 2013; Lin et al., 2014; Vandendriessche et al., 2014; Moshirvaziri et al., 2016; Chiu et al., 2017; Ma et al., 2017). The complexity theories also suggest that different levels of complexity can indicate whether a system is under stress or relatively relaxed (Costa et al., 2002a, 2005; Goldberger et al., 2002). In a novel application of complexity theory, Casali et al. found that measuring complexity can provide a reliable way to discriminate the level of consciousness in single individuals during wakefulness, sleep, and anesthesia, as well as in patients who had emerged from coma and recovered a minimal level of consciousness (Casali et al., 2013). When complexity is used in the evaluation of sleep, studies have shown that complexity indices (e.g., entropy) decrease as sleep gets deeper, and reach their lowest level when slow wave sleep (SWS) occurs (Ma et al., 2018). Moreover, Abásolo et al. found that activated brain states—wakefulness and REM sleep—are characterized by higher complexity compared with NREM sleep (Abásolo et al., 2015). Based on these well-established theories and previous studies, investigating the sleep-related temporal structure of brain activity based on measures such as multi-scale entropy (MSE) (Costa et al., 2002a, 2005), the perturbational complexity index (Casali et al., 2013), or Lempel-Ziv complexity (Abásolo et al., 2015) should provide insights that go beyond those obtained with conventional techniques for signal analysis.
In sleep medicine, one question that remains unsolved is whether the complexity of brain waves in the pre-sleep state or during sleep latency can determine or predict sleep quality. In fact, most people with insomnia complain of being unable to fall asleep because they cannot switch off their “racing” mind (Lichstein and Rosenthal, 1980; Espie et al., 1989; Harvey, 2000). Under high mental load, the sense of urgency about falling asleep adversely affects sleep onset latency (Ansfield et al., 1996). Since stressful brain activities always show higher complexity, we can reasonably hypothesize that lower complexity of brain waves at pre-sleep status can facilitate sleep initiation, reduce sleep latency, and further lead to a high quality of sleep characterized by sufficient deep sleep or SWS.
Slow wave sleep SWS is defined as the state in which large-amplitude, low-frequency waves are dominant and it occurs when delta rhythm is dominant in the EEG signal. Slow wave activity (SWA), which is equivalent to delta activity and encompasses components of the EEG signal in the frequency range of ~0.5–4.5 Hz, is considered to be one of the most important functional EEG parameters during sleep (Brunner et al., 1990; Peter Achermann and Borbély, 2011). Under physiological conditions, SWA is commonly used as a quantitative measure of NREM sleep dynamics and an indicator of sleep depth or sleep intensity (Borbély and Achermann, 1999); Olivier et al. (2010). Fast Fourier transform (FFT) analysis is the most popular method for quantifying SWA. However, it has intrinsic limitations in capturing the underlying dynamics of brain oscillations (Ma et al., 2018). First, FFT analysis takes complex EEG oscillations, composed of sine waves with different frequencies (Campbell, 2009), and decomposes them into frequency component bands, such as beta, alpha, theta, and delta. However, it has long been known that brain oscillation is not a linear combination of these arbitrary frequency components, a property called “nonlinearity” (Bedard et al., 2006). Second, FFT analysis assumes that none of these frequency components changes in amplitude or shape as time evolves, which is clearly against what has been observed in complex brain oscillations, a property called “nonstationarity” (Campbell, 2009). In recent years, ensemble empirical mode decomposition (EEMD) has been adopted to solve this problem (Wu and Huang, 2005). EEMD is an adaptive and noise-assisted data analysis method that is based on local characteristics of the data, requiring no predefined basis. EEMD decomposes an original non-linear and non-stationary signal into a series of simple intrinsic mode functions (IMFs), and has the advantage that every IMF can be physically meaningful via the quantification of the instantaneous amplitude and frequency (Wang et al., 2012). EEMD has become popular for analyzing EEG signals in recent years (Chen et al., 2010, 2014; Kuo et al., 2011; Bizopoulos et al., 2013; Al-Subari et al., 2015a,b; Kanoga and Mitsukura, 2015; Bai et al., 2016; Zeng et al., 2016; Gotz et al., 2017; Hassan and Bhuiyan, 2017). Employing EEMD to quantify SWA might lead to additional findings.
Therefore, the aim of the present study was to examine whether the complexity of brain waves in the pre-sleep state is associated with sleep quality. We hypothesized that lower complexity of brain waves before sleep may play a potential facilitating role in wake-sleep transition and improve the subsequent sleep depth. To this end, the MSE method was applied to EEG signals recorded during the first 5 min after light-off, when the participants were still awake. Sleep latency was determined by manual scoring, and sleep depth was quantified by SWA using an EEMD-based approach or traditional FFT analysis.
Overnight polysomnography (PSG) data obtained from the Sleep Heart Health Study-1 (SHHS-1) were used in this study. The SHHS was a multi-center cohort study implemented by the National Heart Lung and Blood Institute to determine the cardiovascular and other consequences of sleep-disordered breathing, and its characteristics have been described in detail elsewhere (Quan et al., 1997; Redline et al., 1998). Unattended overnight PSG was performed with a portable PS-2 system (Compumedics, Abottsville, Australia). Sensors were placed and equipment was calibrated during an evening home visit by a certified technician. Data collection included C3/A2 and C4/A1 EEGs, sampled at 125 Hz; right and left electrooculograms; a bipolar submental electromyogram; thoracic and abdominal excursions (inductive plethysmography bands); airflow (detected by a nasal-oral thermocouple (Protec, Woodinville, WA); finger pulse oximetry (Nonin, Minneapolis, MN) sampled at 1 Hz; electrocardiogram sampled at 125 Hz; body position (mercury gauge sensor); and ambient light (on/off, by a light sensor secured to the recording garment). After equipment retrieval, the data were forwarded to a central reading center (Case Western Reserve University, Cleveland, OH) for scoring according to a standard protocol. Finally, every 30 s epoch was scored (Thomas et al., 2007). The polysomnographic methods, scoring protocol, and quality assurance procedures were as previously described (Quan et al., 1997; Redline et al., 1998; Thomas et al., 2007).
Sleep variables derived from visual scoring were calculated for each participant, including: total sleep time (TST) (time spent asleep between sleep onset and light-on); wake time after sleep onset (WASO, total amount of time awake after falling asleep); and the duration of each sleep stage, calculated as a percentage of TST. Sleep latency, defined as the period from light-off to the first three consecutive epochs of stage N1 sleep or an epoch of any other stage, was also computed.
The study included 103 healthy subjects who met the inclusion criteria: (1) no usual daily alcohol intake; (2) no benzodiazepines or non-tricyclic antidepressants intake within 2 weeks of the SHHS-1 visit; (3) no history of diabetes; (4) no history of stroke; (5) no hypertension status based on second and third blood pressure readings or current treatment with anti-hypertensives; (6) no self-reported hypertension; (7) no self-reported sinus trouble; (8) no coronary angioplasty, heart failure, heart attack, pace maker, or stroke; (9) apnea–hypopnea index, representing the number of apnea and hypopnea events with ≥3% oxygen desaturation per hour of sleep, of < 5; (10) the entire recording was scored, and scoring started before light-off and ended after light-on; (11) no more than 30 min of the sleep period had either lost or unscorable EEG, respiratory, or oximetry data; (12) the time spent on sleep was no less than 50% of the total time spent in bed; (13) at least one epoch during each sleep stage, i.e., REM, N1, N2, and N3; (14) sleep latency no less than 5 min. See the Supplementary Material for identifiers of the included subjects, which were created by the National Sleep Research Resource team for easier matching with file downloads.
MSE was introduced by Costa et al. (2002a,b); Costa et al. (2005) to quantify the complexity of biologic systems. Entropy-based methods characterize uncertainty about a source of information and the probability distribution of the samples drawn from it. The entropy increases with the degree of disorder and reaches its maximum in completely random systems. However, an increase in the entropy may not always be associated with an increase in dynamical complexity. For instance, a randomized time series has higher entropy than the original time series, although the process of generating surrogate data destroys correlations and degrades the information content of the original signal. This inconsistency may be related to the fact that widely used entropy measures are based on single-scale analysis and do not take into account complex temporal fluctuations. Therefore, MSE has been proposed as a method for assessing complexity by measuring the entropy inherent in a time series over multiple time scales.
The procedures involved in calculating MSE have been well reviewed (Costa et al., 2002a,b, 2005) and can be summarized in the following three steps (Yang et al., 2013): (1) construction of a coarse-grained time series according to a scale factor; (2) quantification of the sample entropy of each coarse-grained time series; and (3) examination of the sample entropy profile over a range of scales. The length of each coarse-grained time series is equal to the length of the original time series divided by the scale factor. For scale 1, the time series is simply the original time series. Sample entropy is defined by the negative natural logarithm of the conditional probability that a dataset of length N, having repeated itself within a tolerance r (similarity factor) for m points (pattern length), also repeats itself for m + 1 points, without allowing self-matches (Richman and Moorman, 2000).
One requirement in the calculation of sample entropy is to determine the pattern length m and similarity factor r. In this study, we set m to 2 and r to 0.15 × SD, where SD is the standard deviation of the analyzed time series. As 30 scales were considered and the entropy on each scale does not necessarily have a specific physiological meaning, a complexity index, called CI1−30, was additionally employed in the current study. CI1−30 was defined as the mean value of the entropies from scale 1 to scale 30, as in many other published studies (Vieira et al., 2017). CI1−30 provides insight into the integrated complexity of the system over the time scales of interest. See the Supplementary Material for the MATLAB code for MSE analysis.
Empirical mode decomposition (EMD) is an adaptive data analysis method based on local characteristics of the data, requiring no predefined basis. The details of the method can be found in the work of Huang (2000).
In the EMD approach, the targeted data x(t) is decomposed in terms of IMFs, cj, i.e.,
where rn is the residue of data x(t) after n number of IMFs have been extracted. IMFs are simple oscillatory functions with varying amplitude and frequency. In practice, the EMD is implemented through a sifting process that uses only local extremes, and the process stops when the residue, rn, becomes a monotonic function from which no more IMFs can be extracted (Huang, 2000).
However, mode-mixing, defined as any IMF consisting of oscillations of dramatically disparate scale, can be caused by intermittency of the driving mechanisms and obstructs the true physical interpretations (Wu and Huang, 2005). Therefore, EEMD was developed to alleviate this drawback, making use of the facts that adding noise to the data can provide a uniformly distributed reference scale, and the means of the corresponding IMFs of different white noise series are likely to cancel each other out. The detailed steps for EEMD are (Wu and Huang, 2005): (1) add a white noise series to the targeted data; (2) decompose the data with added white noise into IMFs based on EMD; (3) repeat step (1) and step (2), but with different white noise series each time; and (4) obtain the (ensemble) means of corresponding IMFs of the decompositions as the final result. The MATLAB code for EEMD was shared by RCADA (http://rcada.ncu.edu.tw/research1.htm).
A Hilbert transform can then be used to calculate the instantaneous frequency of the IMFs (Feldman and Braun, 1995). For a IMF c(t), one can define its Hilbert transform ĉ(t) and analytic signal z(t) as shown in Equation (2) and (3), respectively.
Here, ∅(t) is the instantaneous phase (IP) of c(t). The instantaneous frequency f(t) is the derivation of IP and t shown in Equation (4):
Both FFT and EEMD were used for the decomposition of the EEG signal into its constituent frequency components. Once the original EEG signal x(t) was decomposed to n number of IMFs (denoted as ci) and a residue based on EEMD, a different way to measure SWA (denoted as EEMD-SWA) could be proposed, as shown in Equation (5).
In Equation (5), function std(ci) refers to the standard derivation of IMF ci, and the parameters k and l are determined by the orders of the IMFs that fall into the frequency range of SWA, according to the instantaneous frequency range of each IMF. Thus, E1EMD-SWA actually reflects the relative power of slow waves in the signal.
In this study, EEG signals from derivation C3/A2 was imported into MATLAB for offline analysis. For each subject, two key time points were identified, light-off and sleep onset. Figure 1 briefly shows the analysis protocol. The first 5 min EEG immediately after light-out was analyzed by MSE.
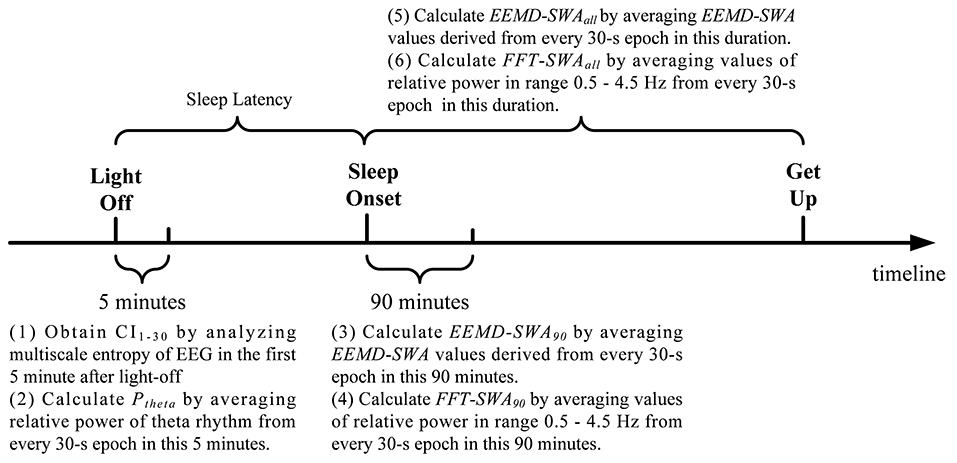
Figure 1. Schematic diagram of the timeline regarding EEG analysis.
On the other hand, SWA was calculated at the beginning of sleep episodes (90 min after sleep onset) and over the whole night's sleep after sleep onset, respectively. EEMD were applied to each 30 s EEG epoch in each time scope. As each 30 s EEG epoch is a time series with 3750 data points, the values of EEMD-SWA for all the epochs in each duration were calculated and averaged, resulting in two metrics, denoted as EEMD-SWA90 and EEMD-SWAall, for each participant. For comparison, the traditional FFT-based evaluation of SWA was applied in a similar way and resulted in another two metrics, denoted as FFT-SWA90 and FFT-SWAall, respectively. In the calculation of FFT-SWA90 and FFT-SWAall, power spectral density was estimated via the period-gram procedure with direct current filtering and Hamming windowing. SWA for each 30 s epoch was calculated by taking the power in the 0.5–4.5 Hz range as a percentage of the total signal power in the frequency range (0.5–62.5 Hz). See the Supplementary Material for the MATLAB code for FFT-SWA.
Then, the correlations between the proposed complexity index CI1−30 and sleep latency, EEMD-SWA90, EEMD-SWAall, FFT-SWA90, and FFT-SWAall, were evaluated to investigate the associations between EEG complexity and proposed measures during the wake-sleep transition and overall sleep. When sleep pressure accumulates, slow brain waves gradually become significant or dominant, which may be in line with a decline in EEG complexity. Inspired by this hypothesis, we investigated the association between brain wave complexity and sleep pressure during pre-sleep wakefulness. Theta activity (4–8 Hz), which is generally considered as a marker for the build-up of sleep pressure (Fattinger et al., 2017), was thus measured by averaging the FFT-based relative power (percentage of the power in the frequency range 0.5–62.5 Hz, denoted as Ptheta in this study) for all the 30 s EEG epochs in the first 5 min after light-off, the same time course as that used for the MSE analysis.
We further analyzed whether early SWA in sleep can be predicted by the pre-sleep EEG complexity using median split subgrouping, where we divided the subjects by ranked SWA into top 50% vs. bottom 50% groups according to EEMD-SWA90 or FFT-SWA90. Such a strategy of median split has an enormous popularity in consumer research, psychology, and numerous other fields (Iacobucci et al., 2015a,b). For instance, Nordström et al. investigated whether age is a suicide risk factor for each sex by median split (Nordström et al., 2010). In addition to that, the subgrouping in the current study provided us an intuitive way to investigate the statistical difference in the values of sample entropy between the two groups over multiple time scales, making a meaningful complementation to CI1−30 used in regressions.
SPSS version 19.0 (IBM SPSS Statistics, NY, United States) and MATLAB (MathWorks R2014a, Inc., Natick, MA, United States) were used for the statistical analyses. The demographics and sleep variables derived from visual scoring were reported as mean with standard deviation if data were normally distributed, and as median with lower and upper quartiles otherwise. Comparisons of the demographics and sleep variables between subgroups from the median split were assessed by chi-square test for categorical variables and unpaired t-test or two-sided Wilcoxon rank sum test for continuous ones. A p < 0.05 was considered statistically significant.
Since sleep may vary with age and sex, and high body mass index (BMI) is a strong risk factor for sleep disorders (Hou et al., 2016), these three variables were included as covariates in the statistical test for correlation between pre-sleep EEG complexity and sleep latency, as well as subsequent sleep quality. General linear models (GLM) were thus employed for the statistical analyses in SPSS 19.0 between CI1−30 and sleep latency, EEMD-SWA90, EEMD-SWAall, FFT-SWA90, FFT-SWAall and Ptheta, controlling age, gender, and BMI.
For each scale in the MSE analysis, the statistical difference in the sample entropy between the EEMD-SWA90 top 50% and bottom 50% groups, as well as between the FFT-SWA90 top 50% and bottom 50% groups, were investigated by covariance analysis, controlling age, gender, and BMI. The false discovery rate (FDR) procedure was included for multiple testing.
In practice, there are three parameters that should be determined for the application of EEMD: the ratio of the standard deviation of the noise to the target data (denoted by ε), the number of prescribed IMFs (denoted by NI), and the number of ensemble members (denoted by NE). In this study, ε was set to 0.1, NI to 7, and NE to 200. Figure 2 shows a typical result of EEMD on a 30 s EEG epoch derived from a subject in the current study.
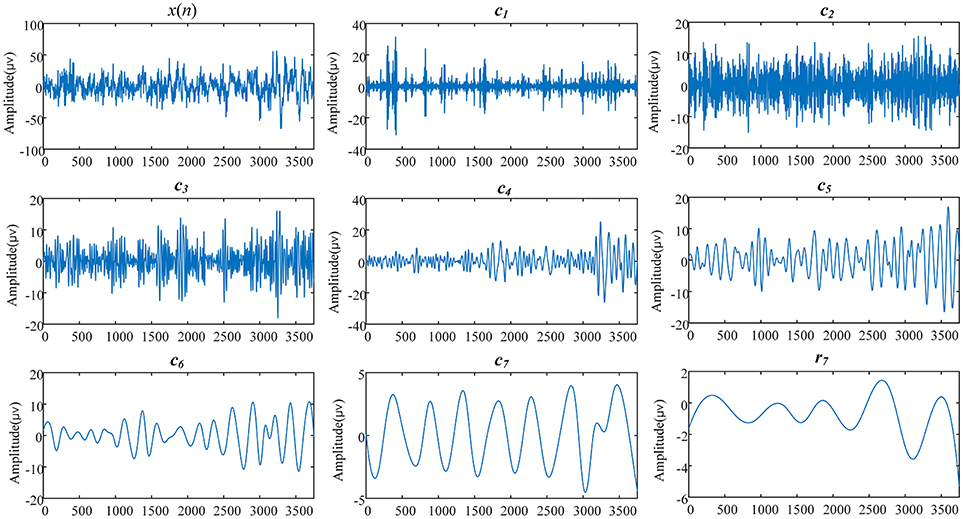
Figure 2. The EEMD results of a 30 s EEG signal derived from a 69-years-old woman (subject ID in SHHS-1 is 200301). x(n) is the original EEG signal, ci(i = 1,2,…,7) are the IMFs, and r7 is the residual part of data x(n) after 7 IMFs were extracted.
Further estimation of the frequency range of IMFs was performed on all epochs in the current study according to Equations (2–4). For each IMF signal, Table 1 lists its frequency range, which uses two quartiles, the 2.5 and 97.5 centiles, and leaves 5% of normal outside the “normal range.” As shown in Table 1 and Figure 2, the IMF1 decomposition was mainly associated with the brain beta rhythm (13–30 Hz), while IMF2 decomposition was associated with the alpha rhythm (8–13 Hz), IMF3 decomposition with the theta rhythm (4–8 Hz), and IMF4, IMF5, IMF6, and IMF7 decompositions with the delta rhythm (< 4 Hz). Therefore, in this study, we set the values of k, l, n in Equation (5) to be 4, 7, and 7, respectively.
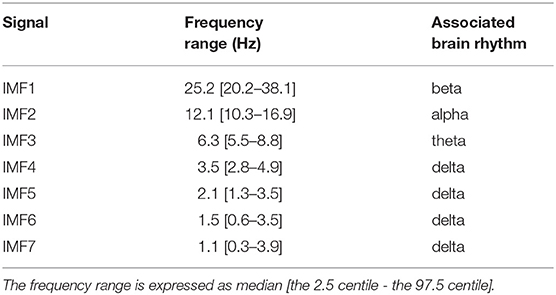
Table 1. Frequency range in seven IMFs of EEMD.
As shown in Table 2, the sample had a median age of 57 years and mean BMI of 25.6 kg/m2. A majority of the participants were identified as being of normal weight (BMI: 18.5–25 kg/m2) or overweight (BMI: 25–30 kg/m2). The standard PSG scoring results revealed that the mean TST was 371 min, while the percentages of scored sleep stages N1, N2, N3, and REM sleep were ~ 4, 55, 20, and 21%, respectively. These results are consistent with the general sleep architecture in adults (Berry, 2012).
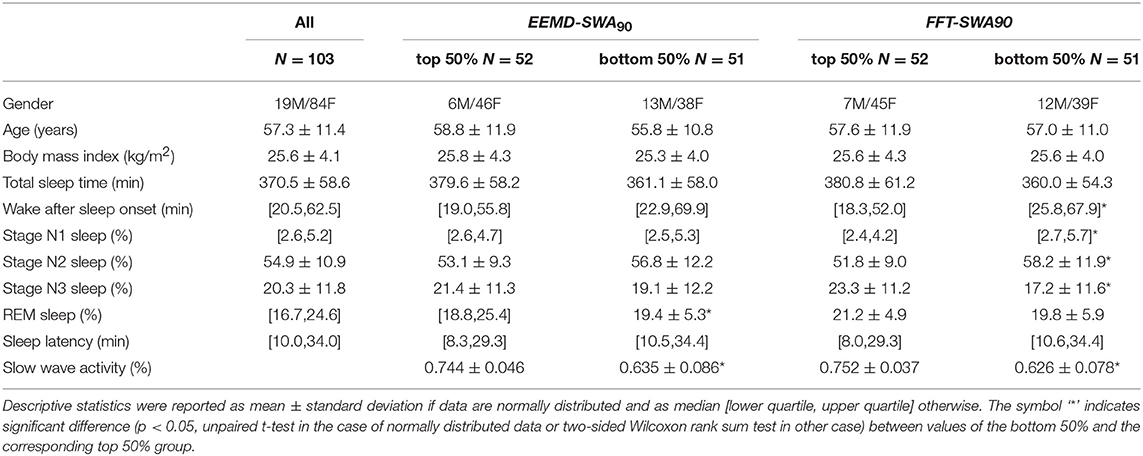
Table 2. Demographics and sleep variables derived from visual scoring.
The demographic characteristics were balanced between the top and bottom 50% groups, with no significant differences regarding age, gender, or BMI. Sleep variables for the EEMD-SWA90 top 50% and bottom 50%, as well as for the FFT-SWA90 top 50% and bottom 50%, are also shown in Table 2. Between the groups divided by EEMD-SWA90, significant differences were found between REM sleep and SWA. However, for the FFT-SWA90 top 50% and bottom 50% groups, in addition to the quantified SWA, WASO, N1, N2, and N3 also showed significant between-group differences. For studied measures (Table 2), no significant differences were found between the two partitions when the same grouping rules (top 50% vs. bottom 50%) were applied.
As shown in Table 3, GLM revealed a significant positive correlation between CI1−30 and sleep latency (r = 0.328, p = 0.001), indicating that higher complexity level was moderately associated with prolonged sleep latency.
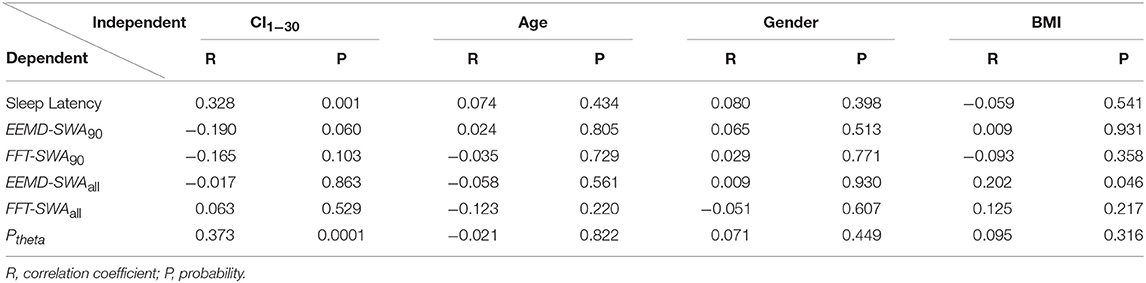
Table 3. Correlations between pre-sleep EEG complexity or demographics and sleep measures in GLM models.
In the regression model, where we adjusted for age, gender, and BMI, EEMD-SWA90 showed a statistical trend for a weak correlation with CI1−30 (r = −0.190, p = 0.06), whereas no association was found between FFT-SWA90 and CI1−30. However, for the whole-night data, no correlation was found between CI1−30 and EEMD-SWAall or FFT-SWAall. Furthermore, the GLM model, controlling age, gender, and BMI, revealed a significant positive correlation (r = 0.373, p = 0.0001) between Ptheta and CI1−30.
When sample entropies on scales 1–30 were compared between the EEMD-SWA90 top 50% and bottom 50% groups, significant differences (controlled by the FDR procedure) were found on all the chosen scales, except that of scale 1. As presented in Figure 3, the group with the higher EEMD-SWA90 in sleep showed a reduced complexity of pre-sleep brain dynamics. When the FFT-SWA90 top 50% and bottom 50% groups were compared, as shown in Figure 4, a similar negative correlation was observed, but significant differences (p < 0.05) between the two groups were found only on the short time scales (scales 2–9). However, after FDR controlling, no significant differences remained for any of the 30 scales, indicating that the EEMD-based method might provide a more robust way to measure SWA.
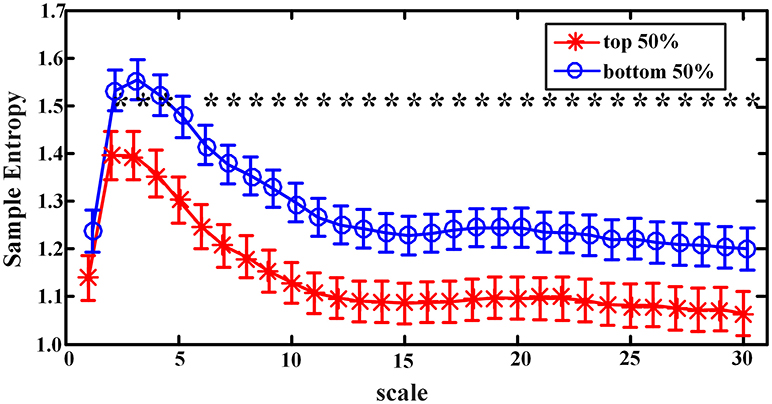
Figure 3. Complexity indices (sample entropy, mean ± SD) on scale 1–30 in groups classified by the rank of EEMD-SWA90 (top 50% vs. bottom 50%). The symbol ‘*' indicates significant difference between groups (p < 0.05, covariance analysis, controlling the age, gender, and BMI). After the procedure of FDR, the significant differences all remained.
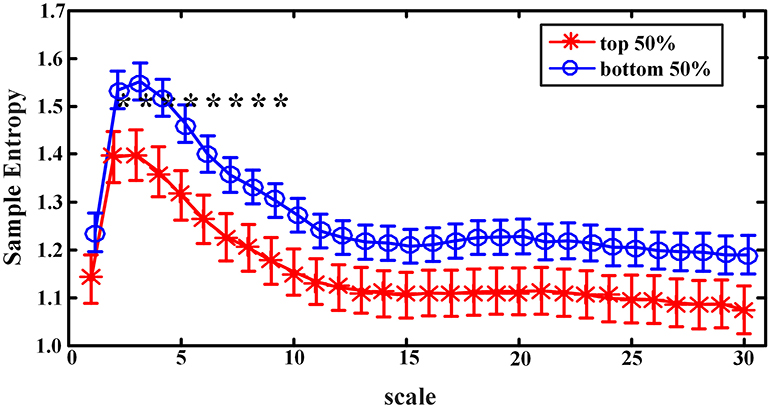
Figure 4. Complexity indices (sample entropy, mean ± standard deviation) on scale 1–30 in groups classified by the rank of FFT-SWA90 (top 50% vs. bottom 50%). The symbol ‘*' indicates significant difference between groups (p < 0.05, covariance analysis, controlling the age, gender and BMI). However, after the procedure of FDR, none of the significant differences remained.
In this study, we examined the associations among wake EEG complexity, sleep latency, and the subsequent SWA quantities during early sleep and over the entire night. Our results revealed a positive correlation between the complexity during the 5 min wakeful EEG and sleep latency. Such complexity also showed a statistical trend for a weak and negative correlation with EEMD-SWA at the beginning of sleep (90 min after sleep onset). Our results suggest that the lower complexity of brain waves in the pre-sleep state may facilitate sleep initiation and reduce sleep latency.
As mentioned in the Introduction, high mental load and urgency to fall asleep increase sleep onset latency (Ansfield et al., 1996) and may lead to non-restorative and unsatisfying sleep, especially in patients with mental disorders or with sleep-onset insomnia, that is, difficulty in sleep initiation. Existing evidence suggests that the manipulation of pre-sleep cognitive activity can lead to changes in sleep onset latency (Ansfield et al., 1996; Nelson and Harvey, 2003; Wuyts et al., 2012). Therefore, future studies are encouraged to examine the complexity before sleep among such populations and to investigate whether interventions that reduce brain wave complexity can assist in promoting restful sleep and improved sleep quality by increasing SWA. In addition, as previously introduced, when a system is under stress or relatively relaxed, levels of complexity can be an appropriate indicator (Costa et al., 2002a, 2005; Goldberger et al., 2002). In recent years, some studies used complexity measures to quantify or evaluate the relaxation states (Aftanas and Golocheikine, 2002; Natarajan et al., 2004). Therefore, we can assume that complexity measures may be useful to examine relaxation interventions such as meditation, slow-paced breathing, music therapy, and cognitive therapy. Effective approaches might be expected to reduce pre-sleep brain complexity and therefore improve sleep quality.
Although in adults, a pronounced increase of theta activity has often been found during the course of sleep deprivation (Aeschbach et al., 1997, 1999; Finelli et al., 2000; Strijkstra et al., 2003; Fattinger et al., 2017), we observed a counterintuitive link between EEG complexity and theta activity during wakefulness in the studied population with normal sleep rather than sleep deprivation. Owing to the counteraction of the circadian process, a linear function with a superimposed 24 h sine wave is generally fitted to theta power and time awake (Åkerstedt and Gillberg, 1990; Finelli et al., 2000). In this way, theta activity may not increase within a session until extreme sleepiness is encountered (Åkerstedt and Gillberg, 1990; Finelli et al., 2000). Aeschbach et al. investigated the influence of the circadian pacemaker and of the duration of time awake on EEG (Aeschbach et al., 1999). They found that the theta rhythm was exhibited a minimum of 1 h after the onset of melatonin secretion (the clock time of melatonin onset in Aeschbach's study ranged from 22:05 to 22:43), indicating evident dissociation, as the circadian maximum of theta activity appeared to be delayed with respect to the maximum of sleep propensity. In our study, 72 out of 103 participants reported their clock time for light-off in the range of 22:05–23:43, however, it is still lack of evidence to make clear the association between sleep pressure and EEG complexity before nocturnal sleep in normal populations. The association between complexity/MSE and sleep-wake regulation is therefore not straightforward and deserves further investigations.
We also analyzed SWA over the entire night for each subject. However, those whole-night results showed significantly weaker correlations with wake complexity compared with SWA90. As it is well-recognized that SWA exhibits a global declining trend over the course of the night and its level in the first non-REM episode increases as a function of prior waking (Achermann et al., 1993), which may indicate the reason why the observed association only present between pre-sleep EEG complexity and SWA in early sleep rather than the whole night. Nonetheless, it would be worth investigating the dynamic changes in pre-sleep complexity and SWA through the entire night, and examining the potential causal relationship in future work. As mentioned above, most of the previous studies of EEG complexity focused on analyses of entropy for each sleep stage, showing a trend of decreasing complexity as sleep becomes deeper (Ma et al., 2018). Non-linear features, including complexity indices, may provide assistance in automatic sleep classification, but, more importantly, studies are encouraged to break the boundaries of limitation to expand the application of non-linear approaches, so that we can better understand the sleep dynamics (Ma et al., 2018). A further step would be to study the complexity differences between normal and pathological conditions, and investigate whether abnormal sleep can be predicted by the first few minutes of pre-sleep EEG recording while a subject is still awake. It would also be of value to determine the potential mechanisms of pre-sleep EEG complexity and sleep-wake transitions and overall sleep quality. Again, we encourage broad applications of such complexity-based approaches in future studies.
Regarding the methodology, the groups defined by FFT-SWA90 showed significant differences regarding manually scored sleep parameters (e.g., WASO, N1, N2, N3), while the differences were not significant when groups were defined by EEMD-SWA90. One possible reason is that both manual scoring and FFT spectral analysis are based on the wave morphology and feature waves. However, for revealing the underlying association between wake complexity and quantified sleep SWA, the proposed EEMD-based method had better performance than traditional FFT-based spectral analysis. GLM revealed a weak correlation between pre-sleep EEG complexity and EEMD-SWA during the first 90 min of sleep. Furthermore, when we split the participants into two groups according to the values of EEMD-SWA90, the sample entropies at almost all time scales exhibited significant differences between the two groups, suggesting that waking EEG complexity can distinguish between lower and higher groups of SWA after sleep onset quantified by the EEMD approach. However, when SWA was measured using a traditional way such as FFT, any association between waking complexity and sleep SWA was insignificant. Owing to the non-linear and non-stationary characteristics of EEG signals during sleep, there would be a drifting of the frequency ranges of the EEG components for each individual. In this situation, as demonstrated in the current study, compared with the frequency band fixed method, a data adaptive approach such as EEMD to measure SWA would be more sensitive to the underlying dynamics of sleep and better able to uncover its mechanisms.
One strength of this study was that we used an existing database and included a reasonably large number of subjects provided by the freely available database (SHHS) to test our hypothesis. Meanwhile, limitations of this study include that associations measured at the population level may not reflect associations at the individual level, and that the study is prone to confounding by other factors. In our study, 82% of the participants were female, which may limit the generalizability of our finding, given that hormonal changes during the menstrual cycle or in peri-menopausal transition may affect sleep (Mallampalli and Carter, 2014; Jehan et al., 2015). However, no such information was provided for the participants in the database. Future studies with prospective designs are strongly encouraged. In addition, the PSG data provided in SHHS-1 were collected from two scalp-placed electrodes: C3 and C4. Since each brain region is associated with different EEG feature waves, future studies are encouraged to perform such complexity and SWA analyses using multiple EEG montages.
In conclusion, lower complexity before sleep onset is associated with a decline of sleep latency and higher SWA after sleep onset, suggesting that reduced complexity of brain waves may improve sleep quality. The application of complexity measures is important to extend our knowledge of sleep. Future studies are encouraged to explore the complexity before sleep among subjects with sleep disorders, to determine its relationship with the efficacy of interventional approaches or for developing screening tools for use with short-term pre-sleep EEG. Although the current study elucidated the association between wakeful brain complexity and nocturnal sleep quality in the healthy population, future investigations should focus on patients with sleep disorders. We would like to encourage interdisciplinary efforts to address this research question.
The study protocol of SHHS was approved by the institutional review board of each participating center, and each participant signed informed consent. All methods were carried out in accordance with relevant guidelines and regulations. The current study only analyzed de-identified data from the SHHS database, and did not involve a research protocol requiring approval by the relevant institutional review board or ethics committee.
FH, C-KP, CW, and YM designed this study. FH, AY and ZY analyzed data. FH and YM wrote the article.
This work was supported by the National Natural Science Foundation of China (61401518, 81673681, and 81473357) and the Delta Environmental and Educational Foundation.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer IK and handling Editor declared their shared affiliation.
We would like to acknowledge the support team of the forum in the Sleep Heart Health Study for their detailed explanations and assistance in our use of the dataset.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnins.2018.00809/full#supplementary-material
Åkerstedt, T., and Gillberg, M. (1990). Subjective and objective sleepiness in the active individual. Int. J. Neurosci. 52, 29–37. doi: 10.3109/00207459008994241
Abásolo, D., Simons, S., Morgado da Silva, R., Tononi, G., and Vyazovskiy, V. V. (2015). Lempel-Ziv complexity of cortical activity during sleep and waking in rats. J. Neurophysiol. 113, 2742–2752. doi: 10.1152/jn.00575.2014
Abeysuriya, R. G., Rennie, C. J., Robinson, P. A., and Kim, J. W. (2014). Experimental observation of a theoretically predicted nonlinear sleep spindle harmonic in human EEG. Clin. Neurophysiol. 125, 2016–2023. doi: 10.1016/j.clinph.2014.01.025
Acharya, U. R., Faust, O., Kannathal, N., Chua, T., and Laxminarayan, S. (2005). Non-linear analysis of EEG signals at various sleep stages. Comput. Methods Prog. Biomed. 80, 37–45. doi: 10.1016/j.cmpb.2005.06.011
Achermann, P., and Borbély, A. A. (2011). “Sleep homeostasis and models of sleep regulation,” in Principles and Practice of Sleep Medicine. (Philadelphia, PA: Elsevier/Saunders, Elsevier Inc.), Chapter 37, 431–444. doi: 10.1016/C2009-0-59875-3
Achermann, P., Dijk, D. J., Brunner, D. P., and Borbély, A. A. (1993). A model of human sleep homeostasis based on EEG slow-wave activity: quantitative comparison of data and simulations. Brain Res. Bull. 31, 97–113. doi: 10.1016/0361-9230(93)90016-5
Aeschbach, D., Matthews, J. R., Postolache, T. T., Jackson, M. A., Giesen, H. A., and Wehr, T. A. (1997). Dynamics of the human EEG during prolonged wakefulness: evidence for frequency-specific circadian and homeostatic influences. Neurosci. Lett. 239, 121–124. doi: 10.1016/S0304-3940(97)00904-X
Aeschbach, D., Matthews, J. R., Postolache, T. T., Jackson, M. A., Giesen, H. A., and Wehr, T. A. (1999). Two circadian rhythms in the human electroencephalogram during wakefulness. Am. J. Physiol. 277, 1771–1779. doi: 10.1152/ajpregu.1999.277.6.R1771
Aftanas, L. I., and Golocheikine, S. A. (2002). Non-linear dynamic complexity of the human EEG during meditation. Neurosci. Lett. 330, 143–146. doi: 10.1016/S0304-3940(02)00745-0
Al-Subari, K., Al-Baddai, S., Tome, A. M., Goldhacker, M., Faltermeier, R., and Lang, E. W. (2015a). EMDLAB: a toolbox for analysis of single-trial EEG dynamics using empirical mode decomposition. J. Neurosci. Methods 253, 193–205. doi: 10.1016/j.jneumeth.2015.06.020
Al-Subari, K., Al-Baddai, S., Tome, A. M., Volberg, G., Hammwohner, R., and Lang, E. W. (2015b). Ensemble empirical mode decomposition analysis of EEG data collected during a contour integration task. PLoS ONE 10:e0119489. doi: 10.1371/journal.pone.0119489
Ansfield, M. E., Wegner, D. M., and Bowser, R. (1996). Ironic effects of sleep urgency. Behav. Res. Ther. 34, 523–531. doi: 10.1016/0005-7967(96)00031-9
Bai, Y., Wan, X., Zeng, K., Ni, Y., Qiu, L., and Li, X. (2016). Reduction hybrid artifacts of EMG-EOG in electroencephalography evoked by prefrontal transcranial magnetic stimulation. J. Neural Eng. 13:066016. doi: 10.1088/1741-2560/13/6/066016
Bedard, C., Kroger, H., and Destexhe, A. (2006). Does the 1/f frequency scaling of brain signals reflect self-organized critical states? Phys. Rev. Lett. 97:118102. doi: 10.1103/PhysRevLett.97.118102
Berry, R. B. (2012). Fundamentals of Sleep Medicine. Philadelphia, PA: Elsevier/Saunders, 1320–1321. doi: 10.1016/B978-1-4377-0326-9.00037-3
Bizopoulos, P. A., Al-Ani, T., Tsalikakis, D. G., Tzallas, A. T., Koutsouris, D. D., and Fotiadis, D. I. (2013). An automatic electroencephalography blinking artefact detection and removal method based on template matching and ensemble empirical mode decomposition. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2013:5853–5856. doi: 10.1109/EMBC.2013.6610883
Borbély, A. A., and Achermann, P. (1999). Sleep homeostasis and models of sleep regulation. J. Biol. Rhythms 14, 557–568.
Brunner, D. P., Dijk, D. J., Tobler, I., and Borbély, A. A. (1990). Effect of partial sleep deprivation on sleep stages and EEG power spectra: evidence for non-REM and REM sleep homeostasis. Electroencephalogr. Clin. Neurophysiol. 75, 492–499. doi: 10.1016/0013-4694(90)90136-8
Campbell, I. G. (2009). EEG recording and analysis for sleep research. Curr. Protoc. Neurosci. 49, 10.2.1-10.2.19. doi: 10.1002/0471142301.ns1002s49
Casali, A. G., Gosseries, O., Rosanova, M., Boly, M., Sarasso, S., Casali, K. R., Tononi, G., et al. (2013). A theoretically based index of consciousness independent of sensory processing and behavior. Sci. Transl. Med. 5:198ra105. doi: 10.1126/scitranslmed.3006294
Chen, D., Li, D., Xiong, M., Bao, H., and Li, X. (2010). GPGPU-aided ensemble empirical-mode decomposition for EEG analysis during anesthesia. IEEE Trans. Inf. Technol. Biomed. 14, 1417–1427. doi: 10.1109/TITB.2010.2072963
Chen, X., Liu, A., McKeown, M. J., Poizner, H., and Wang, Z. J. (2014). An EEMD-IVA framework for concurrent multidimensional EEG and unidimensional kinematic data analysis. IEEE Trans. Biomed. Eng. 61, 2187–2198. doi: 10.1109/TBME.2014.2319294
Chiu, H. C., Ma, H. P., Lin, C., Lo, M. T., Lin, L. Y., Wu, C. K., et al. (2017). Serial heart rhythm complexity changes in patients with anterior wall ST segment elevation myocardial infarction. Sci. Rep. 7:43507. doi: 10.1038/srep43507
Costa, M., Goldberger, A. L., and Peng, C. K. (2002a). Multiscale entropy analysis of complex physiologic time series. Phys. Rev. Lett. 89:068102. doi: 10.1103/PhysRevLett.89.068102
Costa, M., Goldberger, A. L., and Peng, C. K. (2002b). Multiscale entropy to distinguish physiologic and synthetic RR time series. Comput. Cardiol. 29, 137–140. doi: 10.1109/CIC.2002.1166726
Costa, M., Goldberger, A. L., and Peng, C. K. (2005). Multiscale entropy analysis of biological signals. Phys Rev E Stat Nonlin Soft Matter Phys. 71(Pt 2):021906. doi: 10.1103/PhysRevE.71.021906
Espie, C. A., Brooks, D. N., and Lindsay, W. R. (1989). An evaluation of tailored psychological treatment of insomnia. J. Behav. Ther. Exp. Psychiatry 20, 143–153. doi: 10.1016/0005-7916(89)90047-5
Fattinger, S., Kurth, S., Ringli, M., Jenni, O. G., and Huber, R. (2017). Theta waves in children's waking electroencephalogram resemble local aspects of sleep during wakefulness. Sci. Rep. 7:11187. doi: 10.1038/s41598-017-11577-3
Feldman, M., and Braun, S. (1995). Identification of non-linear system parameters via the instantaneous frequency: application of the hilbert transform and Wigner-Ville techniques. Proc. SPIE 2460:637.
Finelli, L. A., Baumann, H., Borbély, A. A., and Achermann, P. (2000). Dual electroencephalogram markers of human sleep homeostasis: correlation between theta activity in waking and slow-wave activity in sleep. Neuroscience 101:523–529. doi: 10.1016/S0306-4522(00)00409-7
Goldberger, A. L., Peng, C. K., and Lipsitz, L. A. (2002). What is physiologic complexity and how does it change with aging and disease? Neurobiol. Aging 23, 23–26. doi: 10.1016/S0197-4580(01)00266-4
Gotz, T., Stadler, L., Fraunhofer, G., Tome, A. M., Hausner, H., and Lang, E. W. (2017). A combined cICA-EEMD analysis of EEG recordings from depressed or schizophrenic patients during olfactory stimulation. J. Neural Eng. 14:016011. doi: 10.1088/1741-2552/14/1/016011
Harvey, A. G. (2000). Pre-sleep cognitive activity: a comparison of sleep-onset insomniacs and good sleepers. Br J Clin Psychol. 39 (Pt 3), 275–286. doi: 10.1348/014466500163284
Hassan, A. R., and Bhuiyan, M. I. H. (2017). Automated identification of sleep states from EEG signals by means of ensemble empirical mode decomposition and random under sampling boosting. Comput. Methods Prog. Biomed. 140, 201–210. doi: 10.1016/j.cmpb.2016.12.015
He, W. X., Yan, X. G., Chen, X. P., and Liu, H. (2005). Nonlinear feature extraction of sleeping EEG signals. Conf. Proc. IEEE Eng. Med. Biol. Soc. 5, 4614–4617. doi: 10.1109/IEMBS.2005.1615498
Hou, F. Z., Li, F. W., Wang, J., and Yan, F. R. (2016). Visibility graph analysis of very short-term heart rate variability during sleep. Phys. A Stat. Mech. Appl. 458, 140–145. doi: 10.1016/j.physa.2016.03.086
Huang, N. E. (2000). New method for nonlinear and nonstationary time series analysis: empirical mode decomposition and Hilbert spectral analysis. Proc. SPIE Int. Soc. Opt. Eng. 4056, 197–209. doi: 10.1117/12.381681
Iacobucci, D., Posavac, S. S., Kardes, F. R., Schneider, M. J., and Popovich, D. L. (2015a). The median split: Robust, refined, and revived ?. J. Consum. Psychol. 25, 690–704. doi: 10.1016/j.jcps.2015.06.014
Iacobucci, D., Posavac, S. S., Kardes, F. R., Schneider, M. J., and Popovich, D. L. (2015b). Toward a more nuanced understanding of the statistical properties of a median split. J. Consum. Psychol. 25, 652–665. doi: 10.1016/j.jcps.2014.12.002
Janjarasjitt, S., Scher, M. S., and Loparo, K. A. (2008). Nonlinear dynamical analysis of the neonatal EEG time series: the relationship between sleep state and complexity. Clin. Neurophysiol. 119, 1812–1823. doi: 10.1016/j.clinph.2008.03.024
Jehan, S., Mastersisarilov, A., Salifu, I., Zizi, F., Jeanlouis, G., Pandiperumal, S. R., et al. (2015). Sleep disorders in postmenopausal women. J. Sleep Disord. Ther. 4:1000212.doi: 10.4172/2167-0277.1000212
Kanoga, S., and Mitsukura, Y. (2015). Eye blink artifact rejection in single-channel electroencephalographic signals by complete ensemble empirical mode decomposition and independent component analysis. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2015, 121–124. doi: 10.1109/EMBC.2015.7318315
Kuo, C. C., Lin, W. S., Dressel, C. A., and Chiu, A. W. (2011). Classification of intended motor movement using surface EEG ensemble empirical mode decomposition. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2011, 6281–6284. doi: 10.1109/IEMBS.2011.6091550
Lichstein, K. L., and Rosenthal, T. L. (1980). Insomniacs' perceptions of cognitive versus somatic determinants of sleep disturbance. J. Abnorm. Psychol. 89, 105–107. doi: 10.1037/0021-843X.89.1.105
Lin, Y. H., Huang, H. C., Chang, Y. C., Lin, C., Lo, M. T., Liu, L. Y., et al. (2014). Multi-scale symbolic entropy analysis provides prognostic prediction in patients receiving extracorporeal life support. Crit Care 18:548. doi: 10.1186/s13054-014-0548-3
Ma, Y., Shi, W., Peng, C. K., and Yang, A. C. (2018). Nonlinear dynamical analysis of sleep electroencephalography using fractal and entropy approaches. Sleep Med. Rev. 37, 85–93. doi: 10.1016/j.smrv.2017.01.003
Ma, Y., Tseng, P. H., Ahn, A., Wu, M. S., Ho, Y. L., Chen, M. F., et al. (2017). Cardiac autonomic alteration and metabolic syndrome: an ambulatory ECG-based study in a general population. Sci. Rep. 7:44363. doi: 10.1038/srep44363
Mallampalli, M. P., and Carter, C. L. (2014). Exploring sex and gender differences in sleep health: a society for women's health research report. J. Womens Health 23:553–562. doi: 10.1089/jwh.2014.4816
Mejaddam, A. Y., Birkhan, O. A., Sideris, A. C., Van der Wilden, G. M., Imam, A. M., Hwabejire, J. O., et al. (2013). Real-time heart rate entropy predicts the need for lifesaving interventions in trauma activation patients. J. Trauma Acute Care Surg. 75, 607–612. doi: 10.1097/TA.0b013e31829bb991
Moshirvaziri, H., Ramezan-Arab, N., and Asgari, S. (2016). Prediction of the outcome in cardiac arrest patients undergoing hypothermia using EEG wavelet entropy. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2016, 3777–3780. doi: 10.1109/EMBC.2016.7591550
Natarajan, K., Acharya, R. U., Alias, F., Tiboleng, T., and Puthusserypady, S. K. (2004). Nonlinear analysis of EEG signals at different mental states. BioMed. Eng. Online 3, 7–7. doi: 10.1186/1475-925X-3-7
Nelson, J., and Harvey, A. G. (2003). An exploration of pre-sleep cognitive activity in insomnia: imagery and verbal thought. Br J Clin Psychol. 42(Pt 3), 271–288. doi: 10.1348/01446650360703384
Nordström, P., Samuelsson, M., and Åsberg, M. (2010). Survival analysis of suicide risk after attempted suicide. Acta Psychiatr. Scand. 91, 336–340. doi: 10.1111/j.1600-0447.1995.tb09791.x
Olivier, J., Mathieu, P., Julie, C., Jacques, M., and Antonio, Z. (2010). Analysis of slow-wave activity and slow-wave oscillations prior to somnambulism. Sleep 33, 1511–1516. doi: 10.1093/sleep/33.11.1511
Quan, S. F., Howard, B. V., Iber, C., Kiley, J. P., Nieto, F. J., O'Connor, G. T., et al. (1997). The Sleep Heart Health Study: design, rationale and methods. Sleep 20, 1077–1085.
Redline, S., Sanders, M. H., Lind, B. K., Quan, S. F., Iber, C., Gottlieb, D. J., et al. (1998). Methods for obtaining and analyzing unattended polysomnography data for a multicenter study.pdf. Sleep 21, 759–767.
Richman, J. S., and Moorman, J. R. (2000). Physiological time-series analysis using approximate entropy and sample entropy. Am. J. Physiol. Heart Circ. Physiol. 278, H2039–H2049. doi: 10.1152/ajpheart.2000.278.6.H2039
Shen, Y., Olbrich, E., Achermann, P., and Meier, P. F. (2003). Dimensional complexity and spectral properties of the human sleep EEG. Electroencephal. Clin. Neurophysiol. 114, 199–209. doi: 10.1016/S1388-2457(02)00338-3
Strijkstra, A. M., Beersma, D. G., Drayer, B., Halbesma, N., and Daan, S. (2003). Subjective sleepiness correlates negatively with global alpha (8-12 Hz) and positively with central frontal theta (4-8 Hz) frequencies in the human resting awake electroencephalogram. Neurosci. Lett. 340, 17–20. doi: 10.1016/S0304-3940(03)00033-8
Thomas, R. J., Mietus, J. E., Peng, C.-K., Gilmartin, G., Daly, R. W., Goldberger, A. L., et al. (2007). Differentiating obstructive from central and complex sleep apnea using an automated electrocardiogram-based method. Sleep 30, 1756–1769. doi: 10.1093/sleep/30.12.1756
Vandendriessche, B., Peperstraete, H., Rogge, E., Cauwels, P., Hoste, E., Stiedl, O., et al. (2014). A multiscale entropy-based tool for scoring severity of systemic inflammation. Crit. Care Med. 42, e560–e569. doi: 10.1097/CCM.0000000000000299
Vieira, M. F., Lemos, T. S., da Costa, P. H. L., and Andrade, A. O. (2017). “On the use of multifractal detrended fluctuation analysis and multiscale entropy to analyze postural sway data,” in Paper Presented at the XVII Congresso Brasileiro de Biomecânica. Porto Alegre, RS.
Wang, T., Zhang, M., Yu, Q., and Zhang, H. (2012). Comparing the applications of EMD and EEMD on time–frequency analysis of seismic signal. J. Appl. Geophys. 83, 29–34. doi: 10.1016/j.jappgeo.2012.05.002
Wu, Z., and Huang, N. E. (2005). Ensemble empirical mode decomposition: a noise-assisted data analysis method. Adv. Adapt. Data Anal. 1, 1–41. doi: 10.1142/S1793536909000047
Wuyts, J., De Valck, E., Vandekerckhove, M., Pattyn, N., Bulckaert, A., Berckmans, D., et al. (2012). The influence of pre-sleep cognitive arousal on sleep onset processes. Int. J. Psychophysiol. 83, 8–15. doi: 10.1016/j.ijpsycho.2011.09.016
Yang, A. C., Wang, S. J., Lai, K. L., Tsai, C. F., Yang, C. H., Hwang, J. P., et al. (2013). Cognitive and neuropsychiatric correlates of EEG dynamic complexity in patients with Alzheimer's disease. Prog. Neuropsychopharmacol. Biol. Psychiatry. 47, 52–61. doi: 10.1016/j.pnpbp.2013.07.022
Yeh, J. R., Peng, C. K., Lo, M. T., Yeh, C. H., Chen, S. C., Wang, C. Y., et al. (2013). Investigating the interaction between heart rate variability and sleep EEG using nonlinear algorithms. J. Neurosci. Methods 219, 233–239. doi: 10.1016/j.jneumeth.2013.08.008
Keywords: EEG, brain activity, non-linear, sleep medicine, sleeps stages, complexity
Citation: Hou F, Yu Z, Peng C-K, Yang A, Wu C and Ma Y (2018) Complexity of Wake Electroencephalography Correlates With Slow Wave Activity After Sleep Onset. Front. Neurosci. 12:809. doi: 10.3389/fnins.2018.00809
Received: 31 May 2018; Accepted: 17 October 2018;
Published: 13 November 2018.
Edited by:
Yaroslav O. Halchenko, Dartmouth College, United StatesReviewed by:
Ilona Kotlewska, Nicolaus Copernicus University in Torun, Poland and Dartmouth College, United StatesCopyright © 2018 Hou, Yu, Peng, Yang, Wu and Ma. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fengzhen Hou, aG91ZnpAY3B1LmVkdS5jbg==
Chunyong Wu, Y3l3dUBjcHUuZWR1LmNu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.