Commentary: Supplier-dependent differences in intermittent voluntary alcohol intake and response to naltrexone in Wistar rats
 Lova Segerström
Lova Segerström Erika Roman
Erika Roman- Research Group Neuropharmacology, Addiction and Behavior, Department of Pharmaceutical Biosciences, Uppsala University, Uppsala, Sweden
A commentary on
Commentary: Supplier-dependent differences in intermittent voluntary alcohol intake and response to naltrexone in Wistar rats
by Kalueff, A. V. (2016). Front. Neurosci. 10:82. doi: 10.3389/fnins.2016.00082
The authors appreciate Prof. Kalueffs interest in our work (Momeni et al., 2015). The issues highlighted (Kalueff, 2016) are indeed of great importance. Many factors associated with animal husbandry can potentially have an impact on behavior (Castelhano-Carlos and Baumans, 2009), and a more thorough description of experimental conditions is often warranted for replicability and/or comparison across labs. Indeed, attempts to compare results across labs through careful standardization show that the lab environment, including experimenter handling, give rise to variation (Crabbe et al., 1999; Wahlsten et al., 2003; Riedel and Spruijt, 2016); also observable in experiments with automated home-cage based tracking equipment with minimal experimenter handling (Riedel and Spruijt, 2016). An alternative approach of reduced standardization has therefore been suggested (Voelkl and Würbel, 2016).
Momeni et al. (2015) attempted to harness of some of the known differences in Wistar rats from different suppliers (Palm et al., 2011a,b, 2012; Goepfrich et al., 2013) in order to create a more heterogeneous group of animals. Kalueff (2016) suggests that the differences observed by Momeni et al. (2015) are due to environmental factors and/or in combination with acclimatization. These are important aspects, often poorly described in publications, but in our opinion not the full explanation behind the results (Momeni et al., 2015).
Animal handling and husbandry vary, see Supplementary Table and Langer et al. (2011) for additional information, which may impact not only our results. The most striking observation (Supplementary Table) is that Wistar rats and CD-1 mice are kept in the same housing room at Charles River GmbH (Personal communication, Charles River distributor, Scanbur employee), a particular concern to users of CD-1 mice from that supplier. A comparison between Supplementary Table and Kalueff (2016) render discrepancies in information, i.e., number of animals per cage. This may be referred to EU Guidelines, stating that body weight and not age determine the number of animals per cage, applicable to Momeni et al. (2015). Notably, the number of animals allowed per cage in different cage types varies between EU, UK and USA Guidelines, respectively (Tecniplast, 2016).
The impact of transportation procedures (Supplementary Table) should not be underestimated. The distance transported per se (Kalueff, 2016) is likely of less importance with regard to the animals used in Momeni et al. (2015), considering that negligible effects were demonstrated in mice bred locally vs. shipped (Wahlsten et al., 2003). Moreover, the undisturbed acclimatization period of 2 weeks upon arrival is in line with recommendations for male rats (Arts et al., 2012, 2014a,b), also when transferred from normal to reversed light/dark cycle (Arts et al., 2016).
An additional factor of potentially large importance for behavioral studies worth emphasizing is the light/dark cycle. Some publications lack specific information about photoperiod for behavioral testing. Behavioral testing of nocturnal animals should preferably be conducted during the dark (active) phase, unless it is established that the behavioral measures collected are not impacted by photoperiod (Castelhano-Carlos and Baumans, 2009; Hawkins and Golledge, 2016). Moreover, the introduction of individually ventilated cage (IVC) systems may impact on animal physiology (Castelhano-Carlos and Baumans, 2009), and result in behavioral differences when IVC-housed animals are compared to animals housed in conventional open cage systems (Kallnik et al., 2007; Mineur and Crusio, 2009; Logge et al., 2013).
Despite acknowledging mentioned differences in animal handling and husbandry, additional factors may be of importance for the marked supplier-dependent differences between Wistar rats (Langer et al., 2011; Palm et al., 2011a,b, 2012; Goepfrich et al., 2013; Momeni et al., 2015; Theilmann et al., 2016; Wood et al., 2016). Wood et al. (2016) presents a simplified scheme of derivation of commercially available Wistar rats. Recently, a single point mutation in exon 3 of the Grm2 gene was reported, resulting in a premature stop codon of the metabotropic glutamate 2 (mGlu2) receptor, and concomitant loss of functional protein expression. This mutation has a widespread prevalence in Wistar rats, particularly those known as Han Wistar (Wood et al., 2016). The possible prevalence of other mutations in Wistar rats from different suppliers remains to be investigated.
In Momeni et al. (2015) the aim was indeed to apply a behavioral phenomics approach (Gerlai, 2002; Kalueff, 2016) to create a heterogenic group by combining Wistar rats from different suppliers; known to display different characteristics and thus pose as a representative of the heterogeneity in a human population (Stewart and Kalueff, 2015) to explore how subgroups with different behavioral characteristics acquire and maintain voluntary alcohol intake. We were hoping to find a pattern similar to that described by Kalueff (2016). However, analysis of behavioral data (open field and Y-maze) and voluntary alcohol intake revealed differences and concomitant skew subgroup formations, primarily in open field performance and voluntary alcohol intake, that were of supplier-dependent origin rather than equally distributed in the heterogeneous group (Table 1 in Momeni et al., 2015). Kalueff (2016) applied a Spearman correlation analysis to the summary subgroup data obtained from Table 1 in Momeni et al. (2015), i.e., the distribution of the heterogeneous cohort of animals into different subgroups based on behavior and voluntary alcohol intake, and presented correlations that imply possible “strain-specific differences in the higher-order phenotypes,” while also acknowledging that more accurate results may be generated by analyzing raw data (Kalueff, 2016). Due to the skew subgroup formations in Momeni et al. (2015), such correlations were originally not considered relevant to perform. Indeed, Spearman rank order correlations on selected raw data presented in Momeni et al. (2015) reveal no correlations (Table 1) in support of the preliminary results presented by Kalueff (2016).
TABLE 1
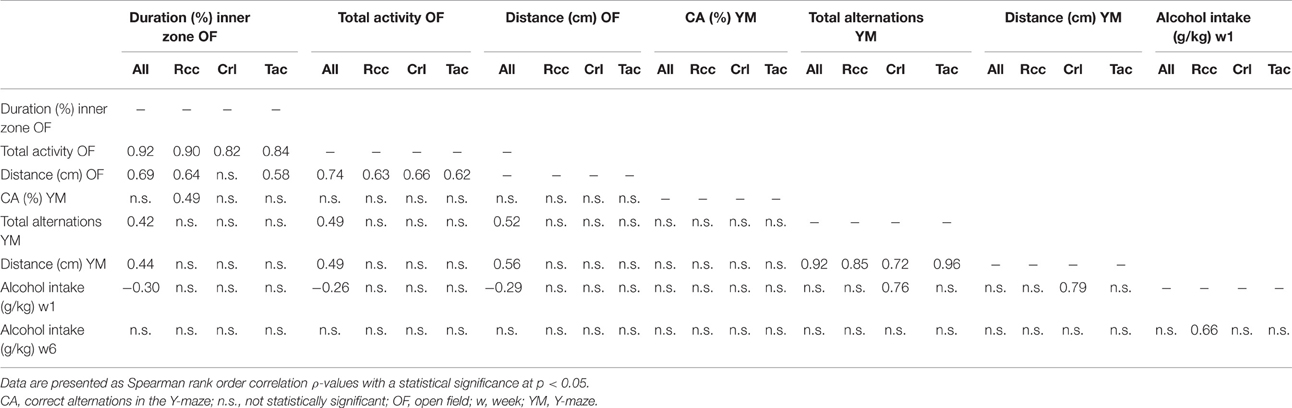
Table 1. Correlation matrix comprising selected parameters from the open field (OF) and Y-maze (YM) tests, and voluntary alcohol intake during weeks one and six of access (see Momeni et al., 2015 for details).
The analysis of open field data in all rats, i.e., the heterogeneous group, revealed a positive correlation between time spent in the inner zone (interpreted as risk-taking behavior) and total activity; an effect also present within the respective group of rats (Table 1). This finding was reported in Momeni et al. (2015) and is in agreement with previous findings (Momeni et al., 2014). Moreover, measures of general activity (distance moved, total activity and total alternations) in the open field and Y-maze tests were correlated in the heterogeneous group of all rats, while only correlations within the respective tests were found in the Rcc, Crl, and Tac groups, respectively (Table 1). A novel finding was that in the heterogeneous group (all rats), total activity, distance moved, and the time spent in the inner zone of the open field was negatively correlated to voluntary alcohol intake during the first week of access, i.e., at acquisition. These correlations were weak, driven predominantly by Crl rats (Supplementary Figure), as they are characterized by longer duration in the inner zone and higher total activity compared to Rcc and Tac rats (Momeni et al., 2015), and absent in the respective group of Rcc, Crl, or Tac rats (Table 1). In agreement with our previous finding (Momeni et al., 2014) and proof of robustness, no association between risk-taking behavior and voluntary alcohol intake was revealed in Rcc rats. To summarize, the complementary correlational analyses performed herein do not support the preliminary findings presented by Kalueff (2016).
In conclusion, the important questions raised by Kalueff (2016), to some extent expanded upon herein, can hopefully raise the attention to the many factors to consider when performing animal experiments in general, and studies in the field of behavioral phenomics and neuroscience in particular. This is all in a strive toward more robust behavioral research with the aim of developing experimental tests and models with high population validity that better parallel findings in humans.
Author Contributions
Study PI: ER. Response outline: LS and ER. Analyzed the data: ER. Drafted the manuscript: ER. Revised and approved the final version: LS and ER.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to acknowledge the first author of the original article Shima Momeni, no longer working in the group. Financial support (ER) from the Alcohol Research Council of the Swedish Alcohol Retailing Monopoly, and the Facias and Åke Wiberg Foundations is gratefully acknowledged. The original work was conducted by support from the Uppsala University Behavioral Facility (UUBF).
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fnins.2016.00442
References
Arts, J. W., Bartholomeus, M. V. D., Meulen, A., Kramer, K., and Ohl, F. (2016). “Effects of reversing light-dark cycle following transfer and re-housing on behavioural and physiological parameters in rats,” in Transportation in Laboratory Rats: Effects of a Black Box, ed J. W. Arts [dissertation thesis] (Utrecht: Brain Center Rudolf Magnus), 135–157.
Arts, J. W., Kramer, K., Arndt, S. S., and Ohl, F. (2012). The impact of transportation on physiological and behavioral parameters in Wistar rats: implications for acclimatization periods. ILAR J. 53, E82–E98. doi: 10.1093/ilar.53.1.82
Arts, J. W., Kramer, K., Arndt, S. S., and Ohl, F. (2014a). Sex differences in physiological acclimatization after transfer in Wistar rats. Animals (Basel) 4, 693–711. doi: 10.3390/ani4040693
Arts, J. W., Oosterhuis, N. R., Kramer, K., and Ohl, F. (2014b). Effects of transfer from breeding to research facility on the welfare of rats. Animals (Basel) 4, 712–728. doi: 10.3390/ani4040712
Castelhano-Carlos, M. J., and Baumans, V. (2009). The impact of light, noise, cage cleaning and in-house transport on welfare and stress of laboratory rats. Lab. Anim. 43, 311–327. doi: 10.1258/la.2009.0080098
Crabbe, J. C., Wahlsten, D., and Dudek, B. C. (1999). Genetics of mouse behavior: interactions with laboratory environment. Science 284, 1670–1672. doi: 10.1126/science.284.5420.1670
Gerlai, R. (2002). Phenomics: fiction or the future? Trends Neurosci. 25, 506–509. doi: 10.1016/S0166-2236(02)02250-6
Goepfrich, A. A., Gluch, C., Friemel, C. M., and Schneider, M. (2013). Behavioral differences in three Wistar Han rat lines for emotional reactivity, cognitive processing and ethanol intake. Physiol. Behav. 110–111, 102–108. doi: 10.1016/j.physbeh.2012.12.019
Hawkins, P., and Golledge, H. (2016). “Overview of key issues associated with using nocturnal rodents during the working day,” in Proceedings of Measuring Behavior 2016, eds A. Spink, G. Riedel, L. Zhou, L. Teekens, R. Albatal, and C. Gurrin (Dublin), 380–383.
Kallnik, M., Elvert, R., Ehrhardt, N., Kissling, D., Mahabir, E., Welzl, G., et al. (2007). Impact of IVC housing on emotionality and fear learning in male C3HeB/FeJ and C57BL/6J mice. Mamm. Genome 18, 173–186. doi: 10.1007/s00335-007-9002-z
Kalueff, A. V. (2016). Commentary: supplier-dependent differences in intermittent voluntary alcohol intake and response to naltrexone in Wistar rats. Front. Neurosci. 10:82. doi: 10.3389/fnins.2016.00082
Langer, M., Brandt, C., and Löscher, W. (2011). Marked strain and substrain differences in induction of status epilepticus and subsequent development of neurodegeneration, epilepsy, and behavioral alterations in rats. [corrected]. Epilepsy Res. 96, 207–224. doi: 10.1016/j.eplepsyres.2011.06.005
Logge, W., Kingham, J., and Karl, T. (2013). Behavioural consequences of IVC cages on male and female C57BL/6J mice. Neuroscience 237, 285–293. doi: 10.1016/j.neuroscience.2013.02.012
Mineur, Y. S., and Crusio, W. E. (2009). Behavioral effects of ventilated micro-environment housing in three inbred mouse strains. Physiol. Behav. 97, 334–340. doi: 10.1016/j.physbeh.2009.02.039
Momeni, S., Segerström, L., and Roman, E. (2015). Supplier-dependent differences in intermittent voluntary alcohol intake and response to naltrexone in Wistar rats. Front. Neurosci. 9:424. doi: 10.3389/fnins.2015.00424
Momeni, S., Sharif, M., Ågren, G., and Roman, E. (2014). Individual differences in risk-related behaviors and voluntary alcohol intake in outbred Wistar rats. Behav. Pharmacol. 25, 206–215. doi: 10.1097/FBP.0000000000000036
Palm, S., Hävermark, Å., Meyerson, B. J., Nylander, I., and Roman, E. (2011a). When is a Wistar a Wistar? Behavioral profiling of outbred Wistar rats from five different suppliers using the MCSF test. Appl. Anim. Behav. Sci. 135, 128–137. doi: 10.1016/j.applanim.2011.08.010
Palm, S., Roman, E., and Nylander, I. (2011b). Differences in voluntary ethanol consumption in Wistar rats from five different suppliers. Alcohol 45, 607–614. doi: 10.1016/j.alcohol.2010.11.005
Palm, S., Roman, E., and Nylander, I. (2012). Differences in basal and ethanol-induced levels of opioid peptides in Wistar rats from five different suppliers. Peptides 36, 1–8. doi: 10.1016/j.peptides.2012.04.016
Riedel, G., and Spruijt, B. (2016). “Alone at home: reliability of circadian activity pattern within and between laboratories,” in Proceedings of Measuring Behavior 2016, eds A. Spink, G. Riedel, L. Zhou, L. Teekens, R. Albatal, and C. Gurrin (Dublin), 393–397.
Stewart, A. M., and Kalueff, A. V. (2015). Developing better and more valid animal models of brain disorders. Behav. Brain Res. 276, 28–31. doi: 10.1016/j.bbr.2013.12.024
Tecniplast (2016). Conventional Cages. Available online at: http://www.tecniplast.it/en/product/conventional-housing-for-small-rodents.html
Theilmann, W., Kleimann, A., Rhein, M., Bleich, S., Frieling, H., Löscher, W., et al. (2016). Behavioral differences of male Wistar rats from different vendors in vulnerability and resilience to chronic mild stress are reflected in epigenetic regulation and expression of p11. Brain Res. 1642, 505–515. doi: 10.1016/j.brainres.2016.04.041
Voelkl, B., and Würbel, H. (2016). Reproducibility crisis: are we ignoring reaction norms? Trends Pharmacol. Sci. 37, 509–510. doi: 10.1016/j.tips.2016.05.003
Wahlsten, D., Metten, P., Phillips, T. J., Boehm Ii, S. L., Burkhart-Kasch, S., Dorow, J., et al. (2003). Different data from different labs: lessons from studies of gene-environment interaction. J. Neurobiol. 54, 283–311. doi: 10.1002/neu.10173
Wood, C. M., Nicolas, C. S., Choi, S. L., Roman, E., Nylander, I., Fernandez-Teruel, A., et al. (2016). Prevalence and influence of cys407* Grm2 mutation in Hannover-derived Wistar rats: mGlu2 receptor loss links to alcohol intake, risk taking and emotional behaviour. Neuropharmacology. doi: 10.1016/j.neuropharm.2016.03.020. [Epub ahead of print].
Keywords: animal model, behavior, phenomics, phenotypes, addiction, alcohol use disorders, Y-maze, open field
Citation: Segerström L and Roman E (2016) Response: Commentary: Supplier-dependent differences in intermittent voluntary alcohol intake and response to naltrexone in Wistar rats. Front. Neurosci. 10:442. doi: 10.3389/fnins.2016.00442
Received: 11 July 2016; Accepted: 12 September 2016;
Published: 30 September 2016.
Edited by:
Juan J. Canales, University of Leicester, UKReviewed by:
Abraham A. Palmer, University of California, San Diego, USA (Amanda Barkley-Levenson contributed to the review of Abraham A. Palmer)Byron C. Jones, University of Tennessee Health Science Center, USA
Copyright © 2016 Segerström and Roman. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Erika Roman, erika.roman@farmbio.uu.se