 Amir Goldental
Amir Goldental Pinhas Sabo1†
Pinhas Sabo1† Shira Sardi
Shira Sardi Ido Kanter
Ido Kanter- 1Department of Physics, Bar-Ilan University, Ramat-Gan, Israel
- 2Gonda Interdisciplinary Brain Research Center and The Goodman Faculty of Life Sciences, Bar-Ilan University, Ramat-Gan, Israel
The experimental study of neural networks requires simultaneous measurements of a massive number of neurons, while monitoring properties of the connectivity, synaptic strengths and delays. Current technological barriers make such a mission unachievable. In addition, as a result of the enormous number of required measurements, the estimated network parameters would differ from the original ones. Here we present a versatile experimental technique, which enables the study of recurrent neural networks activity while being capable of dictating the network connectivity and synaptic strengths. This method is based on the observation that the response of neurons depends solely on their recent stimulations, a short-term memory. It allows a long-term scheme of stimulation and recording of a single neuron, to mimic simultaneous activity measurements of neurons in a recurrent network. Utilization of this technique demonstrates the spontaneous emergence of cooperative synchronous oscillations, in particular the coexistence of fast γ and slow δ oscillations, and opens the horizon for the experimental study of other cooperative phenomena within large-scale neural networks.
Introduction
One of the fundamental goals in neuroscience is to understand the mechanisms underlying the emergence of time-dependent collective activities of neural networks (Silva et al., 1991; Gray, 1994; Contreras et al., 1997; Buzsaki and Draguhn, 2004; Buzsaki, 2006; Chialvo, 2010). This understanding will shed light on the way the brain reliably analyzes information and generates behavior (Klimesch, 1999; Basar et al., 2001; Wiest and Nicolelis, 2003; Kahana, 2006; Bollimunta et al., 2008; Fries, 2009; Giraud and Poeppel, 2012). The experimental accomplishment of this goal requires the following two advanced abilities. The first ability is to record from a large number of neurons over a period of seconds and minutes, which reflects the time scale of the collective network phenomena. The second ability is to know all network parameters, e.g., the network connectivity, synaptic delays and synaptic strengths (Figure 1A). Thus, the number of simultaneous measurements has to be in the order of the number of neurons and synapses (Figure 1B). Although the technology of electrophysiological measurements was significantly enhanced during the last decades, there is not yet such a technology which can record from thousands of individual neurons with a single-cell resolution (Marx, 2014) concurrently with real-time gathering of detailed network topology, including synaptic strengths and delays (Pastrana, 2012).
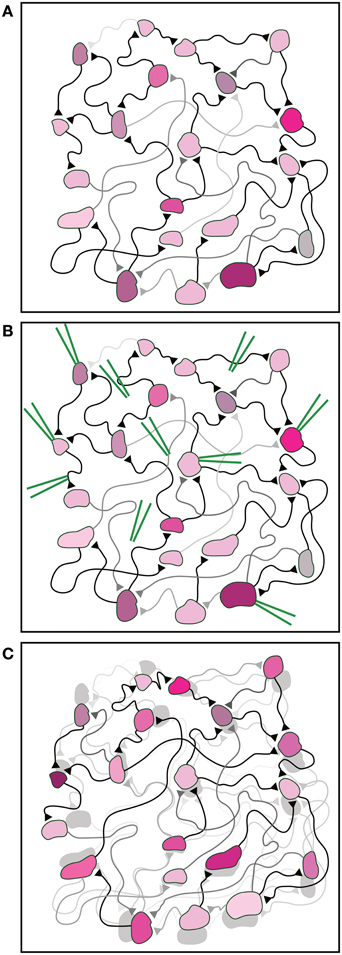
Figure 1. Illustration of the fundamental experimental difficulty. (A) An illustration of a neural network. Synaptic strengths and synaptic delays are indicated by the brightness and length of the connections, respectively. The different properties of each neuron are indicated by different colors and shapes. (B) The knowledge of the current neuronal and synaptic properties requires an enormous number of measurements carried out by many devices (green), e.g., extracellular and intracellular electrodes, inserted in specific targeted spots in the network. (C) The large number of measurements and inserted devices may change the properties of the network. This is schematically exemplified by the difference between the shaded network [identical to the initial network in (A)] and the interfered network as a result of the measurements (front colored network).
It is impartial to assume that the implementation of an enormous number of measurements on the network will influence its activity, and as a byproduct will modify the network parameters. Hence, as a result of many measurements, the estimated network parameters will differ from either the original or from the actual ones (Figure 1C). All in all, this limitation puts in question the ability to experimentally pinpoint the quantitative interplay between the network properties and its functionalities. This limitation reminds the fundamental quantum measurement difficulties (Braginsky et al., 1995), where a measurement affects the state of the system. Although in this case there is no physical principle that prohibits an accurate measurement, the multi-measurements are expected to modify the network and induce unavoidable learning processes, preventing flawless real-time estimations.
We present and utilize a real-time experimental long-term single-neuron stimulation and recording scheme which allows the study of the collective firing activity of a recurrent neural network, given its synaptic strengths and delays. It extends previous attempts to understand network behavior from iterative stimulation and simulations of single cells (Reyes, 2003; Lerchner et al., 2006; Brama et al., 2014; Dummer et al., 2014). Hence, the robustness of the collective firing phenomena can be examined for different sets of synaptic delays and strengths. The presented experimental scheme serves as a mirror image of the reverse engineering methods (Csete and Doyle, 2002; Gregoretti et al., 2010), where the topology of the recurrent network is estimated from its activity and verifies recent simulations and theoretical results, which predicted similar cooperative oscillations in excitatory networks (Goldental et al., 2015).
Materials and Methods
Experimental Procedures
Animals
All procedures were in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals and the University's Guidelines for the Use and Care of Laboratory Animals in Research and were approved and supervised by the Institutional Animal Care and Use Committee.
In vitro Experiments
Culture Preparation
Cortical neurons were obtained from newborn rats (Sprague-Dawley) within 48 h after birth using mechanical and enzymatic procedures. The cortical tissue was digested enzymatically with 0.05% trypsin solution in phosphate-buffered saline (Dulbecco's PBS) free of calcium and magnesium, and supplemented with 20 mM glucose, at 37°C. Enzyme treatment was terminated using heat-inactivated horse serum, and cells were then mechanically dissociated. The neurons were plated directly onto substrate-integrated multi-electrode arrays (MEAs) and allowed to develop functionally and structurally mature networks over a time period of 2–3 weeks in vitro, prior to the experiments. Variability in the number of cultured days in this range had no effect on the observed results. The number of plated neurons in a typical network was in the order of 1,300,000, covering an area of about 380 mm2. The preparations were bathed in minimal essential medium (MEM-Earle, Earle's Salt Base without L-Glutamine) supplemented with heat-inactivated horse serum (5%), glutamine (0.5 mM), glucose (20 mM), and gentamicin (10 g/ml), and maintained in an atmosphere of 37°C, 5% CO2, and 95% air in an incubator as well as during the electrophysiological measurements.
Synaptic Blockers
All experiments were conducted on cultured cortical neurons that were functionally isolated from their network by a pharmacological block of glutamatergic and GABAergic synapses. For each culture 20 μl of a cocktail of synaptic blockers was used, consisting of 10 μM CNQX (6-cyano-7-nitroquinoxaline-2,3-dione), 80 μM APV (amino-5-phosphonovaleric acid) and 5 μM bicuculline. This cocktail did not block the spontaneous network activity completely, but rather made it sparse. At least 1 h was allowed for stabilization of the effect.
Stimulation and Recording
An array of 60 Ti/Au/TiN extracellular electrodes, 30 μm in diameter, and spaced 500 μm from each other (Multi-Channel Systems, Reutlingen, Germany) were used. The insulation layer (silicon nitride) was pre-treated with polyethyleneimine (0.01% in 0.1 M Borate buffer solution). A commercial setup (MEA2100-2x60-headstage, MEA2100-interface board, MCS, Reutlingen, Germany) for recording and analyzing data from two 60-electrode MEAs was used, with integrated data acquisition from 120 MEA electrodes and eight additional analog channels, integrated filter amplifier, and three-channel current or voltage stimulus generator (for each 60 electrode array). Mono-phasic square voltage pulses typically in the range of [−800, −500] mV and [60, 400] μs were applied through extracellular electrodes. Each channel was sampled at a frequency of 50 k samples/s, thus the changes in the neuronal response latency were measured at a resolution of 20 μs.
Cell Selection
A neuron was represented by a stimulation source (source electrode) and a target for the stimulation—the recording electrode (target electrode). These electrodes (source and target) were selected as the ones that evoked well-isolated, well-formed spikes, and reliable response with a high signal-to-noise ratio. This examination was done with a stimulus intensity of −800 mV with a duration of 200 μs using 30 repetitions at a rate of 5 Hz, followed by 1200 repetitions at a rate of 10 Hz.
Data Analysis
Analyses were performed in a Matlab environment (MathWorks, Natwick, MA, USA). The reported results were confirmed based on at least eight experiments each, using different sets of neurons and several tissue cultures. Action potentials were detected on-line by threshold crossing, using a detection window of typically 2–10 ms following the beginning of an electrical stimulation.
Implementation of the Mimicking Scheme
The scheme is based on the neuronal short term memory (Figure 2), and the management of the stimulation history of each of the mimicked neurons, as well as the timings of their evoked spikes, is done in real-time as exemplified in Figure 3. A simplified version of the scheme is presented in Figure 4 in the form of a flowchart as well as in the Supplementary Movie. A detailed description of the mimicking procedure is presented in the Appendices (Supplementary Material).
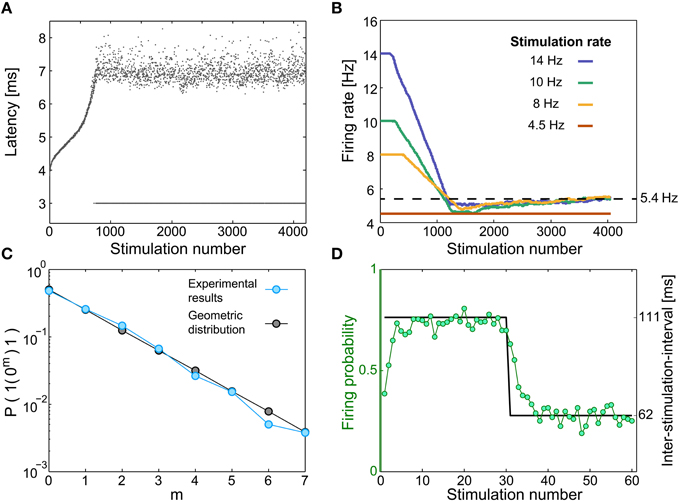
Figure 2. Neuronal short-term memory. (A) The neuronal response latency (NRL) of a cultured neuron, stimulated at 10 Hz. Response failures are denoted at NRL = 3 ms. (B) Firing rates for different stimulation rates (legend), using a sliding window of 1000 stimulations, indicating saturated firing rate (~5.4 Hz, dashed line) independent of the stimulation rate. (C) A semi-log plot of the probability for m successive response failures bounded by evoked spikes, as a function of m, for a stimulation rate of 10 Hz (light blue), and for a geometric distribution (P = 0.5·0.5m, black). (D) The same neuron was given 233 recurrences of 60 stimulations composed of 30 inter-stimulation intervals of 62.5 ms (16 Hz) and 30 inter-stimulation intervals of ~111 ms (9 Hz) (black). The probability for an evoked spike (green circles) indicates a fast adaptation, short memory, of the neuronal response probability.
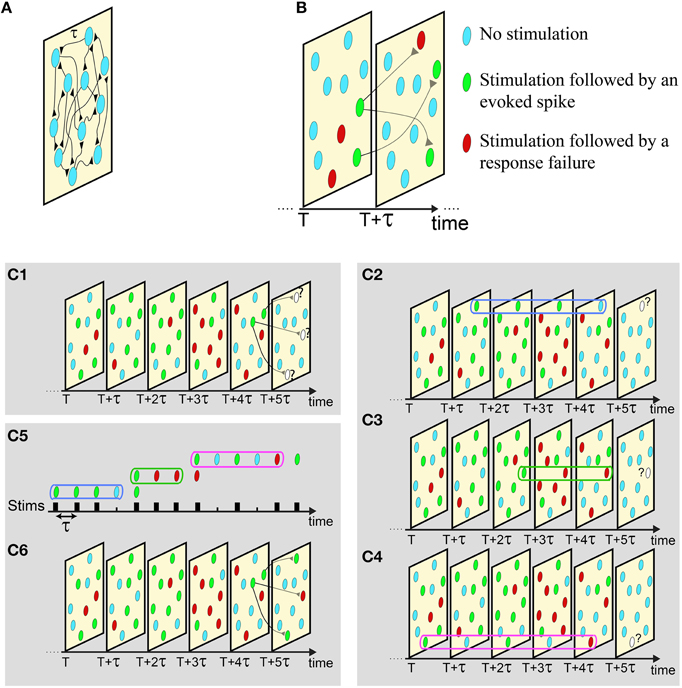
Figure 3. Illustration of the proposed scheme. (A) An excitatory network where all delays are equal to τ. (B) The network dynamics is demonstrated as a set of snapshots for different times, where consecutive snapshots are separated by τ. In each snapshot a neuron can be in one of the following three states: received a stimulation that was followed by an evoked spike (green), received a stimulation that was followed by a response failure (red), or did not receive a stimulation (light blue). (C) C1: The state of the neurons in six consecutive snapshots of the network, where the current state of the three denoted neurons at T + 5τ is unknown. C2−4: The responsiveness of each stimulated neuron in the network is determined by its short-term stimulation memory, three stimulation in the presented example. C5, 6: Mimicking the states of these three neurons using sequential stimulation of a single neuron, C2−4. The responses of the mimicking neuron completely determines the state of the stimulated neurons at snapshot T + 5τ, and the state of the next snapshot can now be revealed by repeating the described procedure.
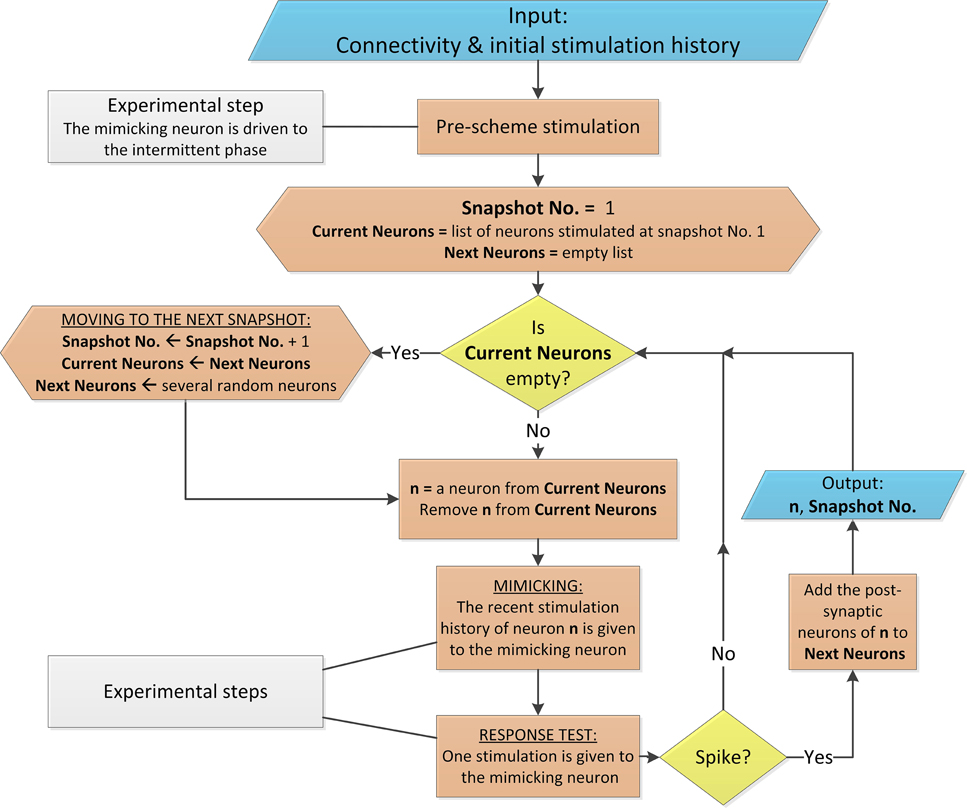
Figure 4. Flowchart of the proposed scheme. The flowchart describes the mimicking process for a networks with above-threshold connections and homogenous delays, similar to Appendix A in Supplementary Material. The flowchart uses conventional shapes. Additionally, the colors light blue, yellow, orange and gray stand for data flow, conditional branching, process and experimental comments, respectively. The process is arbitrarily terminated when Snapshot No. is several hundreds (the mimicked time of the network dynamics is several seconds).
Results
When a neuron is stimulated repeatedly, the time-lag between a stimulation and its corresponding evoked spike, the neuronal response latency (NRL), stretches gradually (Wagenaar et al., 2004; De Col et al., 2008; Vardi et al., 2014, 2015; Figure 2A and Materials and Methods). Above a critical stimulation frequency, fc, which varies much among neurons (Vardi et al., 2015), this stretching terminates at the intermittent phase. This phase is characterized by large fluctuations around a constant NRL and by neuronal response failures, NRFs (Figure 2A). The non-zero fraction of NRFs is such that the average firing frequency is fc, independent of the stimulation frequency; hence, the neuron operates similar to a low pass filter (Vardi et al., 2015; Figure 2B). In addition to the preservation of the neuron's average firing frequency under periodic stimulations, the response failures were found to be statistically independent (Vardi et al., 2015; Figure 2C). Specifically, for inter-stimulation-intervals that are longer than the refractory period, the firing probability is independent of the neuron's firing history. In the general stimulation scenario, aperiodic stimulations, the statistics of the NRFs were experimentally found to depend on the short-term stimulation history of the neuron, which typically consists of several stimulations only (Vardi et al., 2015; Figure 2D). These effects might be an indirect result of some kind of spike-frequency adaptation (Benda and Herz, 2003) or a related mechanism.
The proposed experimental technique allows the mimicking of the activity of a neural network, given the features of the connections and the initial condition of the firing neurons. For the sake of simplicity, we first demonstrate the utilization of the proposed method using a diluted network with above-threshold synapses and with uniform delays between neurons, τ. In such a case the history of a network appears as consecutive “snapshots” of the network separated by τ time-lags between them (Figure 3A). Each “snapshot” of the network defines which are the stimulated neurons and which neurons fire at that time. Specifically, each neuron in each snapshot belongs to one of the following three states: received a stimulation that results in an evoked spike, received a stimulation that results in a response failure, or did not receive stimulation at that time (Figure 3B). The neurons to be stimulated in the consecutive snapshot are determined according to the network connectivity (Figure 3B). For example, assume neuron A is pre-synaptic to neuron B and neuron A fires at time T, consequently neuron B is stimulated at time T + τ. Neurons in the network are stimulated either if their pre-synaptic neurons fired at the previous snapshot, or if they are stimulated by a stochastic noise, e.g., synaptic noise. An example is presented in Figure 3C1, given the network dynamics until the snapshot at time T + 4τ, three neurons will receive a stimulation at the next snapshot, T +5τ (Figure 3C1). The goal now is to determine whether these three neurons will fire, based on their short-term stimulation history. This task is done experimentally using a single mimicking neuron (in vitro or in vivo) (see Supplementary Movie and Materials and Methods) and is based on the following two steps:
The mimicking step: The current responsiveness, response susceptibility to stimulations, of a neuron from the network is mimicked by the enforcement of its short-term stimulation history on the mimicking neuron (see Supplementary Movie), e.g., three last stimulations at Figures 3C2-4. After the completion of this step, the mimicking neuron will have the same responsiveness as the mimicked neuron in the current state of the network (Figure 2D).
The responsive test: τ [ms] after the termination of the first step, the mimicking neuron is stimulated. In case of an evoked spike, we conclude that the mimicked neuron in the network fires and this event is noted in the current snapshot.
For each stimulated neuron in the snapshot these two steps are repeated sequentially in real-time (Figure 3C5), using the same mimicking neuron, until the responsiveness of all neurons in the current snapshot is determined (Figure 3C6). After the snapshot at time T + 5τ was completed, the procedure is repeated to determine the state of the next snapshot, T + 6τ (Figure 3C6), and so on.
The massive management of the stimulation history of each neuron in the network as well as their spike timings is done in real-time (Materials and Methods), demanding faster operations in at least two orders of magnitude than the time scale of τ.
The realization of the proposed real-time method is first demonstrated for an excitatory network consisting of 500 neurons, using a cultured mimicking neuron (Appendix A in Supplementary Material). Each neuron has 2 pre- and 2 post-synaptic connections, all are above-threshold and randomly chosen, τ = 13 ms, with additional external stimulation Poissonian noise with a rate of 1 Hz (Materials and Methods). The network dynamics over ~2 s indicates δ oscillations of ~2.5 Hz which coexist with γ oscillations of ~75 Hz (Figure 5A), which trivially originates from the resolution 1/τ (Figure 3C).
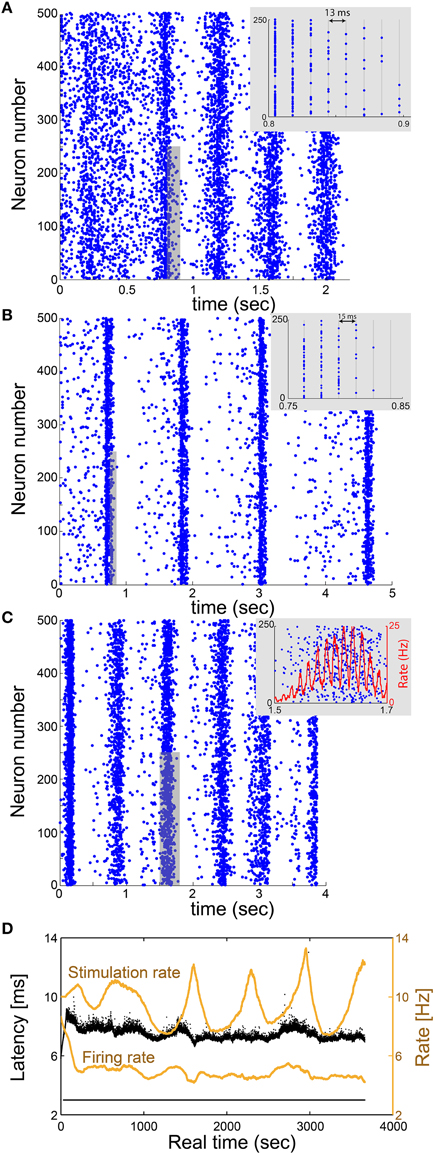
Figure 5. Utilization of the proposed scheme on excitatory large networks reveals δ and γ oscillations. (A) Raster plot of a network, where each blue dot indicates an evoked spike, consisting of 500 mimicked neurons where each neuron in the network has randomly selected 2 pre- and 2 post- above-threshold synaptic connections, and all delays are set to 13 ms. Results, produced using a single-neuron experiment in vitro, indicate fδ~2.5 Hz oscillations which coexist with fγ~75 Hz oscillations (inset). (B) Similar to (A) where each neuron has randomly selected 50 pre- and 50 post- below-threshold synaptic connections, and all delays are set to 15 ms. An above-threshold stimulation requires cooperation of at least four below-threshold stimulations. Results indicate fδ~0.8 Hz oscillations which coexist with fγ~65 Hz oscillations (inset). (C) A raster plot of a network consisting of 500 neurons where each neuron has randomly selected 2 pre- and 2 post- above-threshold synaptic connections, and delays are randomly selected from the uniform distribution U(8,12) ms. Results indicate fδ~1.3 Hz oscillations which coexist with spontaneous fγ~65 Hz oscillations, originated from 1/(average(τ+latency)) (inset). The rate is calculated from the number of spikes in a sliding window of 20 ms with a resolution of 0.1 ms. (D) The NRL of the mimicking neuron in (A) (response failures are denoted at NRL = 3 ms). The stimulation rate (upper orange curve) and firing rate (lower orange curve) are calculated using a sliding average of 2000 stimulations. The average stimulation rate is much higher than fc~5 Hz, indicating that the neuron is in the intermittent phase, which is characterized by large fluctuations of the NRL and response failures which lead to a firing frequency around fc~5 Hz.
This prototypical real-time technique is realized in a more realistic biological network, consisting of sub-threshold synapses as well (Appendix B in Supplementary Material and Figure 5B). The excitatory network consists of N = 500 neurons, where each neuron has 50 pre- and 50 post-synaptic connections with τ = 15 ms, with an additional 1 Hz Poissonian noise (stimulations). An above-threshold stimulation requires the firing of at least four pre-synaptic neurons, or a stimulation originated from the noise. Results indicate δ oscillations of ~0.8 Hz which coexist with γ oscillations of ~65 Hz (Figure 5B), which again originates from the resolution 1/τ.
The generalization of the proposed real-time technique for networks with a continuous distribution of connection delays requires a complicated procedure, since the scheme of discrete time snapshots (Figure 3B) is not valid in this case. The advanced procedure requires the management of the stimulation history and the timings of the evoked spikes of all neurons in a continuous time manner. The mimicking process per neuron is similar, however, technically the complexity of the algorithm is enhanced since the constraint of specific simulations and firing times is released and occur in continuous time. Utilization of the continuous scheme indicates the coexistence of δ and spontaneous γ oscillations (Appendix C in Supplementary Material with Figure 5C and Appendix D in Supplementary Material with Figure 6), where the periods of collective firing are slightly smeared as a result of continuous connection delays.
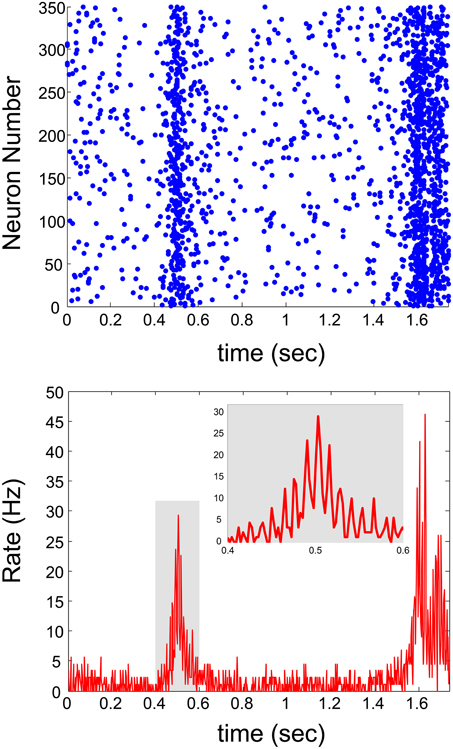
Figure 6. Sub-threshold connections with continuous delays. Real-time scheme with continuous delay-times and sub-threshold connections. Top panel: Raster plot of the network activity. Bottom panel: The average firing rate of the neurons comprising the network, as a function of time.
In the case where all connection delays are equal to τ, the GCD of loops of such random networks is expected to be equal to τ (Kanter et al., 2011; Vardi et al., 2012a,b). In such a case, neurons will fire in synchrony every τ [ms], therefore forming γ oscillations with frequency of 1/τ (Figures 5A,B). On the other hand, in case of random continuous connection delays, the GCD vanishes and no synchrony is expected beyond the δ oscillations. Our results clearly indicate that the non-trivial high frequency synchrony is dominated by the average delay, i.e., the spontaneously originated γ oscillations have the frequency of 1/(average delay) as also observed in simulations (Goldental et al., 2015). The distribution of the connection delays affects only the quality of the synchrony (Figure 5C).
Mimicking the dynamical behavior of a network consisting, for instance, of thousands of neurons over several seconds requires real-time stimulations and recordings of the mimicking neurons over several hours. Specifically, the real-time duration of the experiment is equal to the number of stimulations occurred dynamically in the network, multiplied by the time it takes to mimic a neuron. For illustration, in Figure 5A, a network of N = 500 neurons is mimicked for 2 s. Since, fc = 5 Hz each one of the neurons in the network was mimicked approximately (2 s) · (2fc) = 20 times. The mimicking of one stimulated neuron in the network, requires approximately 0.4 s. Hence, the total real-time of the experiment with a single mimicking neuron is expected to be 20 · N · 0.4 s = 4000 s, which is indeed close to ~3700 s (Figure 5D). During this period, the mimicking neuron remains in the intermittent phase as indicated by the large fluctuations of the NRL and the response failures which resulted from the high stimulation rate (Figure 5D).
Discussion
The presented experimental results verify recent simulations and theoretical work which predicted such oscillations (Goldental et al., 2015). The experimental scheme presents more reliable evidence since it takes into account biological time dependent fluctuations in the responsiveness of neurons and variations in the neuronal critical frequency, as opposed to the simulations and theory.
Currently, there are some limitations to the proposed mimicking method, which is based on short-term neuronal dynamics. Long-term effects and synaptic plasticity are ignored, however they are not expected to dominate the dynamics of the network within several seconds (Figures 5, 6). It might be possible to introduce synaptic dynamics, excitatory and inhibitory, to the mimicking process by stimulating and recording from coupled neurons through synaptic connections, using patch clamp technique (Debanne et al., 2008). Currently this kind of dynamics is simplified to excitatory electrical pulses.
Experimental difficulties arise when the mimicked network is composed of thousands of neurons. Primarily, the experimental time scales linearly with the size of the network, hence it is expected to exceed several hours. Preliminary results (not shown) indicate that it is possible to mimic a network of thousands of neurons, however sailing toward much larger systems is in question. A possible bypass to this obstacle, and a way to mimic more heterogeneous networks, with several types of neurons, is to implement several mimicking neurons in parallel, however it will require the realization of a much more complicated experimental scheme.
The idea of mimicking network dynamics using a single neuron was previously demonstrated for feed-forward networks (Reyes, 2003), where the parameters of the network are adjusted to control the activity, and the mimicking process does not take into consideration short-term neuronal plasticity. This work, on the other hand, examines recurrent random networks, where the parameters are independent of the stability of the firing rates. In addition, the average delay between successive layers in a feed-forward network is irrelevant for the dynamics, since it only shifts the time of the activity by a constant. In contrast, in recurrent networks, the exact delay times are important since each neuron is affected by many delay loops. Hence, the implementation of the mimicking process of a recurrent network consisting of short delays of several milliseconds is a challenge. Additionally, as a result of the many loops, each neuron is revisited many times through the dynamics. Hence, the mimicking process is done many times per neuron, and keeping the network parameters fixed is essential to describe the dynamical properties of the recurrent network.
The proposed real-time experimental method can also be used to mimic the firing patterns of large recurrent neural networks in vivo, based on long-term scheme of stimulation and recording of a single neuron in vivo (Brama et al., 2014). Nevertheless, the real-time management of the in vivo mimicking process, where delays are several milliseconds only, is still an experimental challenge.
The presented experimental technique to use a long-term experiment on a single node in order to mimic the parallel activity of a large scale network is applicable to a variety of networks with propagation delays, where the nodes exhibit a finite or no memory of the preceding conditions. Thus, this technique is expected to be relevant to a wide range of networks that play a key role in other fields such as physics, biology and economics.
Author Contributions
AG developed the theoretical framework and compared experimental results to simulations. PS developed and designed the interface for the real-time experiments. RV and SS prepared the tissue cultures and the experimental materials. All authors performed the experiments and analyzed the data. IK supervised all aspects of the work. All authors discussed the results and commented on the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This research was supported by the Ministry of Science and Technology, Israel.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fnins.2015.00508
References
Basar, E., Basar-Eroglu, C., Karakas, S., and Schurmann, M. (2001). Gamma, alpha, delta, and theta oscillations govern cognitive processes. Int. J. Psychophysiol. 39, 241–248. doi: 10.1016/S0167-8760(00)00145-8
Benda, J., and Herz, A. V. (2003). A universal model for spike-frequency adaptation. Neural Comput. 15, 2523–2564. doi: 10.1162/089976603322385063
Bollimunta, A., Chen, Y., Schroeder, C. E., and Ding, M. (2008). Neuronal mechanisms of cortical alpha oscillations in awake-behaving macaques. J. Neurosci. 28, 9976–9988. doi: 10.1523/JNEUROSCI.2699-08.2008
Braginsky, V. B., Braginskii, V. B., Khalili, F. Y., and Thorne, K. S. (1995). Quantum Measurement. Cambridge: Cambridge University Press.
Brama, H., Guberman, S., Abeles, M., Stern, E., and Kanter, I. (2014). Synchronization among neuronal pools without common inputs: in vivo study. Brain Struct. Funct. 220, 3721–3731. doi: 10.1007/s00429-014-0886-6
Buzsaki, G., and Draguhn, A. (2004). Neuronal oscillations in cortical networks. Science 304, 1926–1929. doi: 10.1126/science.1099745
Chialvo, D. R. (2010). Emergent complex neural dynamics. Nat. Phys. 6, 744–750. doi: 10.1038/nphys1803
Contreras, D., Destexhe, A., Sejnowski, T. J., and Steriade, M. (1997). Spatiotemporal patterns of spindle oscillations in cortex and thalamus. J. Neurosci. 17, 1179–1196.
Csete, M. E., and Doyle, J. C. (2002). Reverse engineering of biological complexity. Science 295, 1664–1669. doi: 10.1126/science.1069981
Debanne, D., Boudkkazi, S., Campanac, E., Cudmore, R. H., Giraud, P., Fronzaroli-Molinieres, L., et al. (2008). Paired-recordings from synaptically coupled cortical and hippocampal neurons in acute and cultured brain slices. Nat. Protoc. 3, 1559–1568. doi: 10.1038/nprot.2008.147
De Col, R., Messlinger, K., and Carr, R. W. (2008). Conduction velocity is regulated by sodium channel inactivation in unmyelinated axons innervating the rat cranial meninges. J. Physiol. 586, 1089–1103. doi: 10.1113/jphysiol.2007.145383
Dummer, B., Wieland, S., and Lindner, B. (2014). Self-consistent determination of the spike-train power spectrum in a neural network with sparse connectivity. Front. Comput. Neurosci. 8:104. doi: 10.3389/fncom.2014.00104
Fries, P. (2009). Neuronal gamma-band synchronization as a fundamental process in cortical computation. Annu. Rev. Neurosci. 32, 209–224. doi: 10.1146/annurev.neuro.051508.135603
Giraud, A.-L., and Poeppel, D. (2012). Cortical oscillations and speech processing: emerging computational principles and operations. Nat. Neurosci. 15, 511–517. doi: 10.1038/nn.3063
Goldental, A., Vardi, R., Sardi, S., Sabo, P., and Kanter, I. (2015). Broadband Macroscopic cortical oscillations emerge from intrinsic neuronal response failures. Front. Neural Circuits 9:65. doi: 10.3389/fncir.2015.00065
Gray, C. M. (1994). Synchronous oscillations in neuronal systems: mechanisms and functions. J. Comput. Neurosci. 1, 11–38. doi: 10.1007/BF00962716
Gregoretti, F., Belcastro, V., Di Bernardo, D., and Oliva, G. (2010). A parallel implementation of the network identification by multiple regression (NIR) algorithm to reverse-engineer regulatory gene networks. PLoS ONE 5:e10179. doi: 10.1371/journal.pone.0010179
Kahana, M. J. (2006). The cognitive correlates of human brain oscillations. J. Neurosci. 26, 1669–1672. doi: 10.1523/JNEUROSCI.3737-05c.2006
Kanter, I., Kopelowitz, E., Vardi, R., Zigzag, M., Kinzel, W., Abeles, M., et al. (2011). Nonlocal mechanism for cluster synchronization in neural circuits. EPL 93:66001. doi: 10.1209/0295-5075/93/66001
Klimesch, W. (1999). EEG alpha and theta oscillations reflect cognitive and memory performance: a review and analysis. Brain Res. Rev. 29, 169–195. doi: 10.1016/S0165-0173(98)00056-3
Lerchner, A., Ursta, C., Hertz, J., Ahmadi, M., Ruffiot, P., and Enemark, S. (2006). Response variability in balanced cortical networks. Neural Comput. 18, 634–659. doi: 10.1162/neco.2006.18.3.634
Marx, V. (2014). Neurobiology: rethinking the electrode. Nat. Methods 11, 1099–1103. doi: 10.1038/nmeth.3149
Pastrana, E. (2012). Sensors and probes: a'nano'era for electrophysiology. Nat. Methods 9, 321–321. doi: 10.1038/nmeth.1961
Reyes, A. D. (2003). Synchrony-dependent propagation of firing rate in iteratively constructed networks in vitro. Nat. Neurosci. 6, 593–599. doi: 10.1038/nn1056
Silva, L. R., Amitai, Y., and Connors, B. W. (1991). Intrinsic oscillations of neocortex generated by layer 5 pyramidal neurons. Science 251, 432–435. doi: 10.1126/science.1824881
Vardi, R., Goldental, A., Marmari, H., Brama, H., Stern, E. A., Sardi, S., et al. (2015). Neuronal response impedance mechanism implementing cooperative networks with low firing rates and microsecond precision. Front. Neural Circuits 9:29. doi: 10.3389/fncir.2015.00029
Vardi, R., Marmari, H., and Kanter, I. (2014). Error correction and fast detectors implemented by ultrafast neuronal plasticity. Phys. Rev. E 89:042712. doi: 10.1103/PhysRevE.89.042712
Vardi, R., Timor, R., Marom, S., Abeles, M., and Kanter, I. (2012a). Synchronization with mismatched synaptic delays: a unique role of elastic neuronal latency. EPL 100:48003. doi: 10.1209/0295-5075/100/48003
Vardi, R., Wallach, A., Kopelowitz, E., Abeles, M., Marom, S., and Kanter, I. (2012b). Synthetic reverberating activity patterns embedded in networks of cortical neurons. EPL 97:66002. doi: 10.1209/0295-5075/97/66002
Wagenaar, D. A., Pine, J., and Potter, S. M. (2004). Effective parameters for stimulation of dissociated cultures using multi-electrode arrays. J. Neurosci. Methods 138, 27–37. doi: 10.1016/j.jneumeth.2004.03.005
Keywords: neuronal plasticity, neural networks, in-vitro, neuronal response latency, neuronal response failures
Citation: Goldental A, Sabo P, Sardi S, Vardi R and Kanter I (2016) Mimicking Collective Firing Patterns of Hundreds of Connected Neurons using a Single-Neuron Experiment. Front. Neurosci. 9:508. doi: 10.3389/fnins.2015.00508
Received: 29 October 2015; Accepted: 21 December 2015;
Published: 20 January 2016.
Edited by:
Mikhail Lebedev, Duke University, USAReviewed by:
Benjamin Lindner, Bernstein Center for Computational Neuroscience, GermanyWolfgang Kinzel, University of Würzburg, Germany
Copyright © 2016 Goldental, Sabo, Sardi, Vardi and Kanter. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ido Kanter, aWRvLmthbnRlckBiaXUuYWMuaWw=
†These authors have contributed equally to this work.