 Emanuela Mhillaj
Emanuela Mhillaj Maria G. Morgese
Maria G. Morgese Paolo Tucci
Paolo Tucci Maria Bove
Maria Bove Stefania Schiavone
Stefania Schiavone Luigia Trabace
Luigia Trabace- 1Department of Physiology and Pharmacology, Sapienza University of Rome, Rome, Italy
- 2Department of Clinical and Experimental Medicine, University of Foggia, Foggia, Italy
Androgens are mainly prescribed to treat several diseases caused by testosterone deficiency. However, athletes try to promote muscle growth by manipulating testosterone levels or assuming androgen anabolic steroids (AAS). These substances were originally synthesized to obtain anabolic effects greater than testosterone. Although AAS are rarely prescribed compared to testosterone, their off-label utilization is very wide. Furthermore, combinations of different steroids and doses generally higher than those used in therapy are common. Symptoms of the chronic use of supra-therapeutic doses of AAS include anxiety, depression, aggression, paranoia, distractibility, confusion, amnesia. Interestingly, some studies have shown that AAS elicited electroencephalographic changes similar to those observed with amphetamine abuse. The frequency of side effects is higher among AAS abusers, with psychiatric complications such as labile mood, lack of impulse control and high violence. On the other hand, AAS addiction studies are complex because data collection is very difficult due to the subjects' reticence and can be biased by many variables, including physical exercise, that alter the reward system. Moreover, it has been reported that AAS may imbalance neurotransmitter systems involved in the reward process, leading to increased sensitivity toward opioid narcotics and central stimulants. The goal of this article is to review the literature on steroid abuse and changes to the reward system in preclinical and clinical studies.
Introduction
Anabolic-androgenic steroids (AAS) are synthetic compounds derived from testosterone, which is the main male hormone. The binding of testosterone to androgen receptors has anabolic and androgenic effects. During puberty, the increase in testosterone levels contributes to linear growth augmentation, as well as muscle mass accumulation (Bhasin et al., 1996, 2001; Brower, 2002; Kuhn, 2002) by inducing hypertrophy without changes in the absolute number of both Type 1 and 2 muscle fibers (Sinha-Hikim et al., 2002). Testosterone also acts by increasing the number of muscle progenitor cells (Sinha-Hikim et al., 2003) and promoting their myogenic differentiation (Singh et al., 2003, 2006). Testosterone promotes mitochondrial biogenesis, improves net oxygen delivery to the tissue by increasing red cell mass and tissue capillarity, and facilitates oxygen unloading from oxyhemoglobin (Coviello et al., 2008; Gupta et al., 2008). The idea of designing and developing steroids with anabolic properties arose during the 1930s soon after the identification and isolation of the hormone androsterone by the German investigator Butenandt, who collected this compound from thousands of liters of pooled human urine derived from a number of military service volunteers. Most of the AAS used before the 1990s were pharmacological agents approved for medicinal or veterinary use. By the 1990s, various androgen precursors became nutritional supplements. Androgen precursors are either inactive or weak androgens that the body converts into potent androgens. These include naturally occurring precursors to testosterone, such as 4-androstenediol, 5-androstenediol, 4-androstenedione, and dehydroepiandrosterone, as well as precursors to synthetic AAS including 4-norandrostenedione, 4-norandrostenediol, and 5-norandrostenediol, which the body converts to nandrolone (Pope et al., 2014). Other synthetic AAS, such as 17-desmethylstanozolol, methylclostebol, and methyltrienolone have been recently introduced into the market as dietary supplements. These “designed” steroids have not undergone toxicological or safety testing in humans or animals. Thus, they potentially represent an even more serious health risk than the more traditionally used AAS.
Medical Use
From a clinical standpoint, AAS are commonly prescribed to treat several disorders, such as the androgen deficiency syndromes (Conway et al., 2000), hereditary angioedema, hematological disorders (Shahidi, 2001), catabolic conditions, such as some types of cancer-related cachexia (Langer et al., 2001), metabolic dysfunctions induced by severe burn (Hart et al., 2001), inflammatory pulmonary diseases (Ferreira et al., 2001), radiation therapy, and AIDS-associated malnutrition (Basaria et al., 2001; Polsky et al., 2001). Less common medical uses of AAS deal with heart and renal failure (Basaria et al., 2001). Contrasting data exists in the literature regarding the use of AAS in the treatment of androgen deficiency in aging males, infertility, sexual dysfunctions or impotence, as well as post-menopausal syndrome in women. Thus, while a review of Morley (2001) points toward therapeutic effects on libido and menopause-induced sarcopenia, Conway et al. (2000) consider their therapeutic application in these pathological conditions as ≪misuse of androgens≫. Hence, according to the state of the art presented in their review, they reported no indication for androgen therapy in male infertility because of its suppressing effect on spermatogenesis. Importantly, there is no evidence in available literature that AAS abuse or dependence might develop from the legitimate medical use of AAS.
Non-medical Use
The use of AAS for non-medical intentions can easily determine abuse and lead to dependence. When used by athletes, AAS can improve performance to levels obtainable by virtually any other combination of non-chemical solutions provided by modern sport techniques (Noakes, 2004). Generally, supra-pharmacological doses of AAS act either by a direct mechanism, promoting an increase in mass, force, speed of muscular contraction, and recovery after intense physical exercise (Tremblay et al., 2004) or by an indirect pathway through erythropoietic stimulation, leading to increased synthesis of 2,3-diphospholglycerate and tissutal oxygen transfer facilitation (Shahidi, 2001). Consumption of high doses of AAS typically consists in 6–12 week cycles, followed by a 6–12 week period of wash-out. These patterns of AAS use may easily precipitate in periods of continuous consumption without any AAS-free intervals due to the fact that abusers try to assure their muscle gains while avoiding withdrawal symptoms (Brower, 2002; Kuhn, 2002). Several other drugs are frequently associated with the use of supra-pharmacological doses of AAS by abusers that are designed to increase their effects, diminish side effects or avoid detection by urine testing (Wichstrom and Pedersen, 2001). The abuse of other illicit drugs, such as amphetamines and opioids, has also been shown to be strengthened by AAS use (Arvary and Pope, 2000). Moreover, such abuse might reinforce the occurrence of adverse substance interactions. In particular, in the case of AAS and amphetamine association, the overdose potential appears to be increased, due to cardiotoxicity (Thiblin et al., 2000). The contemporary consumption of AAS and bromocriptine, used to rapidly reduce body fat and total weight, has been described as the cause of a syndrome characterized by syncopal episodes and atrial fibrillation (Manoharan et al., 2002).
Populations of adolescents and young adults have been the subject of several clinical studies that explore the prevalence of AAS misuse and abuse. Irving et al. (2002) conducted a study on a population of 4746 middle and high school students from public schools of Minneapolis completing surveys and anthropometric measurements as part of a population-based study of eating patterns and weight concerns among teenagers (Project EAT: Eating Among Teens). They observed that steroid use was more common in non-Caucasian males and in middle school students as compared to high school. In males, steroid use was associated with poor self-esteem, higher rates of depressed mood and attempted suicide, poor knowledge and attitudes about health, greater participation in sports emphasizing weight and shape, greater parental concern about weight, and higher rates of eating disorders and substance abuse. In a study by Wichstrom and Pedersen (2001), a representative sample of 8877 Norwegian youths (15–22 years of age) was surveyed. Results showed that AAS use did not vary according to sport involvement or demographics. Moreover, AAS use was associated mainly with the abuse of marijuana, aggressive-type conduct problems and eating disorders.
Adverse Effects
The severity and impact of side effects induced by AAS abuse depend on a wide range of factors, such as dose, duration of administration, possible consumption of a combination of AAS, as well as gender and age of the abusers. Data on the impact of sustained administration, failed to show any documented adverse events associated to a single episode of acute consumption of supra-pharmacological doses of AAS. Their abuse has been shown to be associated to greater effects on physical performance in younger individuals and women, together with increased incidence and risk of developing serious side events (Kindlundh et al., 1999). Few data exist on the risk of side effects linked to long-term use of high-dose of AAS for non-therapeutic purposes (Parssinen and Seppala, 2002). Cardiovascular complications have been widely described in AAS abusers, including the occurrence of arrhythmic events (Furlanello et al., 2003). In a recent post-mortem study that compared 87 deceased men positive for AAS with 173 control subjects (Far et al., 2012), AAS users showed significantly greater cardiac mass.
In another clinical investigation, ventricular hypertrophy, associated with fibrosis and myocytolysis, was detected after cardiac death in four AAS users (Montisci et al., 2012). Also, controlled studies realized by echocardiography (Krieg et al., 2007; Hassan et al., 2009; Baggish et al., 2010) or by cardiac magnetic resonance imaging (Luijkx et al., 2013) have demonstrated lower ventricular ejection fractions and reduced diastolic tissue velocities in AAS users.
Pathological effects on urogenital and reproductive systems have been reported. In particular, hypogonadotropic hypogonadism with consequent testicular atrophy in men and development of inhibitory mechanisms for FSH and LH production in women have been described in selected populations of AAS abusers (Anderson and Wu, 1996; Dohle et al., 2003). Increased virility and lowering of voice tone, irregular menstruation with infertility, decreased breast size, hypertrophic clitoris, and increased sexual desire have also been described in a population of female AAS abusers (Franke and Berendonk, 1997; Kutscher et al., 2002). Other complications include liver damage and hepatitis (Tanaka et al., 2000), insulin-resistance secondary to glucose intolerance with alterations of thyroid function (Yesalis et al., 2000), increased risk of infectious diseases caused by inappropriate use of syringes and non-protected sexual relations among AAS users (Aitken et al., 2002). Although several studies point toward a reversibility of undesirable AAS-induced effects following suspension, they can become irreversible complications with prolonged AAS abuse (Kutscher et al., 2002).
Psychiatric Effects
AAS are universally recognized to have psychoactive effects (Yates, 2000). Although some spared studies have reported their therapeutic use in depression to improve mood and anergia (Rabkin et al., 2000), most evidence points toward the association of AAS with depression, mania, psychosis, suicide and increased aggression leading to violence and, in extreme cases, to homicide (Pope and Brower, 2000; Pope et al., 2000; Thiblin et al., 2000). Indeed, suicide and homicide have been shown to be the main cause of premature deaths among steroid users and, in particular, in the teen population (Thiblin et al., 1999b). Although this does not imply that all steroid users will suffer crippling depression or homicidal rage, steroids appear to strongly contribute to psychiatric dysfunctions in susceptible individuals.
Globally, the prevalence of AAS-induced psychiatric disorders has been hard to evaluate and determine, because of sampling biases in clinical case reports. In a review of Pope et al., (Pope et al., 2000), summarizing four prospective, placebo-controlled trials, it has been reported that at least 5% of AAS users will experience AAS dose-dependent maniac or hypomaniac episodes (Pope and Brower, 2000). However, this estimated percentage appears to be influenced by the fact that in most controlled trials, it is not possible to completely mimic the extreme doses and combinations of AAS taken by abusers for ethical reasons. Thus, estimated rates of AAS-induced psychiatric alterations are probably even higher. This is also due to the fact that other factors can increase the likelihood of psychiatric consequences of AAS abuse, such as the presence of a positive psychiatric anamnesis, alcohol, or other drug use (Dean, 2000) as well as other medical comorbidities. For example, in a case-report of Morton et al. (2000), the authors described the case of a man suffering from Axis II psychopathology, who developed severe psychosis after receiving therapeutic doses of an anabolic steroid for burn injuries in combination with lorazepam and opioids.
Psychological motivations contributing to anabolic steroid use and abuse have received little attention in psychiatric literature. Clinical studies demonstrate that steroids are used in part to deal with an earlier trauma, such as childhood physical or sexual abuse (Porcerelli and Sandler, 1995).
Effects on the Brain Reward Function: Dependence and Addiction Potential
The data in the literature show no documented cases of dependence induced by AAS use at therapeutic doses. This suggests that dependence is likely associated to the use of higher doses of AAS (Long et al., 2000; Thiblin et al., 2000; Haupt, 2001; Brower, 2002; Kutscher et al., 2002); Figure 1 graphically represents this hypothesis. However, molecular mechanisms leading to AAS-induced dependence are still unclear.
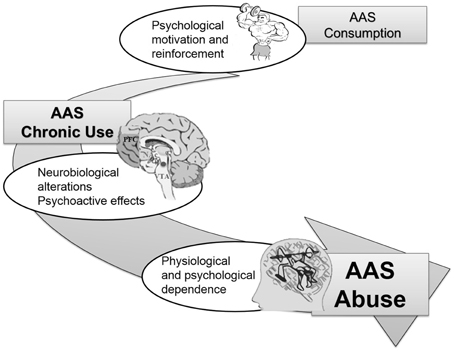
Figure 1. Main clinical observations linking AAS consumption to AAS addiction.
In a review of the scientific literature published between 1988 and 1998 (Brower, 2000), AAS dependence was defined as a diagnosable mental disorder. Between 1999 and 2000, two more diagnostic studies of AAS dependence were published (Midgley et al., 1999; Brower, 2000).
A “withdrawal syndrome” induced by AAS abuse has been clearly described, consisting mainly of depressed mood, fatigue, AAS craving, restlessness, anorexia, insomnia, and decreased libido lasting for several weeks or months (Brower, 1997, 2000). In the 1980s, Tennant et al. (1988) described a case report on which a model of a biphasic course of withdrawal was proposed. The initial phase of the AAS-induced withdrawal (lasting for about 1 week) seemed to be comparable to opioid-induced withdrawal, while the second phase was mostly characterized by clear depressive symptoms and craving (Tennant et al., 1988).
Considerable evidence suggests that AAS dependence might share crucial mechanisms of opioid dependence in humans. In 1989, Kashkin and Kleber (1989) posited that AAS dependence might partly arise via an opioidergic mechanism, through which AAS might enhance the activity of central endogenous opioids, and AAS withdrawal would lead to a decrease in this activity and a subsequent acute hyperadrenergic syndrome (Kashkin and Kleber, 1989). This posited link between AAS and opioids was later confirmed by a large number of observations indicating that AAS users seem to be particularly at risk for developing opioid abuse or dependence (McBride et al., 1996; Wines et al., 1999). Additional clinical studies provided evidence that AAS might decrease the analgesic action of both metamizol and morphine (Philipova et al., 2003).
In 2009, a study by Kanayama et al. (2009) added further evidence for a relationship between AAS and opioids. In the population included in that study, opioid abuse or dependence began either before or after the onset of AAS use, suggesting the possibility that these forms of substance abuse might arise from a common molecular pathway (Kanayama et al., 2009). However, in a study of Negus et al. (2001), authors could not detect any withdrawal phenomena following administration of high doses of AAS (Negus et al., 2001).
AAS seem to act through a more modest reinforcement mechanism compared to cocaine or heroin and resembles the reinforcement mechanism described for caffeine, nicotine, and benzodiazepines. In 2002, Brower (2002) proposed a 2-stage model of steroid dependence. In Stage 1, anabolic effects of AAS provide the initial input and motivation for AAS consumption. Stage 2 deals with consequent chronic use, following which physiological and psychological dependence may develop, thereby making it increasingly difficult for users to quit. Psychoactive effects, such as mood changes and increases in aggressive behavior, characterize this stage of dependence. Diagnostic and Statistical Manual of Mental Disorders criteria for AAS dependence are met and users are not able to stop or discontinue AAS consumption. In Stage 2, addiction treatment may be required, especially when AAS abuse is associated with other substance dependence, such as alcohol, opioids, or amphetamine abuse (Brower, 2002). Arvary and Pope (2000) investigated this phenomenon in a clinical study, including 227 patients admitted to a private facility for dependence on heroin or other opioids. Results of this study strongly suggested that these patients were introduced to opioids through AAS use and bodybuilding physical activity. In particular, 81% of them first purchased opioids from the same drug dealer who had sold them AAS; 67% were introduced to opioids by a fellow body-builder; 86% first used opioids to reduce insomnia and irritability induced by AAS, and 67% used opioids to diminish depression feelings induced by withdrawal from AAS (Arvary and Pope, 2000).
A second model, explaining mechanisms leading to AAS dependence, has also been proposed (Bahrke and Yesalis, 1994). This model holds that AAS-dependence development occurs specifically in socio-cultural contexts that are likely to motivate certain individuals, particularly men, to attain large and strong muscles by frequent and intensive training sessions. These training sessions also improve mood and self-esteem and are generally associated with very strict and controlled dietary regimens. Thus, AAS-induced muscle-active effects might underlie the reinforcing actions of these compounds (Midgley et al., 1999) and the compulsive features of AAS use seem to strengthen the likely compulsive patterns of training and diet. Studies to elucidate mechanisms leading to AAS dependence have also included surveys of current and former AAS users, recruited from gyms, websites, and physicians. Brower et al. (1990) reported numerous criteria for psychoactive substance dependence in a survey of eight AAS abusers, including continued use despite adverse side effects, and withdrawal symptoms (Brower et al., 1990).
Specific dysfunctions of the various components of the brain reward system have been described in clinical studies. For example, alterations in levels of monoamine metabolites, neurohormones, and neuropeptides, which play a crucial role in the reward mechanism, have been investigated in the cerebrospinal fluid of subjects who received methyltestosterone (MT) with respect to placebo-treatment (Daly et al., 2001). Results showed that levels of 5-hydroxyindolacetic acid (5-HIAA) increased while 3-metoxy-4-hydroxyphenylglycol (MHPG) levels decreased in cerebrospinal fluid, following MT administration. In particular, changes in cerebrospinal fluid 5-HIAA significantly correlated with the activation of specific psychiatric symptom cluster scores. In addition, according to this study, a decrease in cerebrospinal fluid MHPG may derive from reduced norepinephrine clearance, even though authors did not detect any significant correlations between changes in MHPG levels and the development of clear psychiatric symptoms, suggesting a less crucial role for noradrenergic changes in this process. An increase in substance P levels and vasopressin (Hallberg et al., 2000; Harrison et al., 2000), as well as dysfunctions of the central opioid system (Schlussman et al., 2000), have been proposed as playing a potential role in the development of aggressive behavior after AAS abuse.
Multiple factors have been associated with the induction of dependence in AAS users, such as low endogenous levels of testosterone. Indeed, it has been demonstrated that women, adolescents and elderly subjects have a lower probability of developing AAS dependence (Wood et al., 2004). Among possible risk factors for dependence development, the most relevant appears to be participation in competitive sports with intense and repetitive physical exercise (Kanayama et al., 2003b). Some investigators have also suggested that personality psychopathology may be a risk factor for AAS abuse. Yates et al. (1990) reported that AAS users and weight lifters had a higher prevalence of histrionic, antisocial, and borderline personality traits than community controls. Although a growing number of reports, current knowledge of molecular mechanisms leading to AAS dependence in humans remains limited. In this regard, the reinforcing effects of AAS may also be biased by intensive physical exercise and by increased narcissistic self-esteem arising from the fulfillment of the desired body appearance. On the other hand, many users practice “stacking” consumption, consisting in the contemporary mixed use of multiple steroids.
Since it has been reported that around 96% of users combine AAS with other drugs in order to relieve non-medical steroid side effects (Parkinson and Evans, 2006), pharmacodynamics, and pharmacokinetic interaction studies are surely warranted, although hardly feasible, in order to exclude further bias.
Behavioral and Neurochemical Responses to AAS Administration in Animal Models
Preclinical studies have contributed in evaluating the impact of AAS exposure on neurochemical mechanisms underlying AAS-induced behavioral outcomes. Animal studies offer a direct measure of behavioral parameters under conditions where age and sex of the subjects, along with AAS administration, are established by the investigator. In this section, we will focus our attention on the data in the literature from animal models employing different AAS exposure paradigms, frequently used to model human abuse patterns. In particular, we will review laboratory animal research findings to assess AAS-induced behavioral effects, such as aggression and reward. Moreover, we will highlight studies that have reported neuronal pathways and signaling molecules involved in these behaviors.
Aggression
Behavioral human studies linking AAS abuse and aggression have confounding factors, such as regimen (multiple steroids over a cycle of use), co-administration with other drugs of abuse and inaccurate measures of behavior simulated by subjective reports (McGinnis, 2004). Conversely, experimental designs in animals that correlate AAS exposure and aggression are less equivocal.
The resident-intruder test is a common paradigm for assessing aggression. Initial studies on animal models have reported that long-term exposure to high doses of testosterone raised levels of aggression in gonadally intact rats and re-established aggression in castrated rats (Lumia et al., 1994). However, indices of aggressive responses depend on environmental context, social cues, sex and hormonal status of the intruder, age of exposure, physical provocation, and type of AAS administered (Clark and Henderson, 2003; Lumia and McGinnis, 2010). Hence, studies in rats showed that AAS-treated males demonstrated a different predisposition for aggression when tested in three different environments (home cage, opponent cage, or neutral cage) (Christie and Barfield, 1979; Lumia et al., 1994; Breuer et al., 2001; Farrell and McGinnis, 2003). Adult male rats receiving high doses of AAS are more aggressive toward the intruder in their home cage and displayed lower levels of aggression in either opponents or neutral cages (Breuer et al., 2001; Farrell and McGinnis, 2003). Investigators extended their interest to other experimental factors demonstrating that AAS-treated rats are typically more aggressive toward intact rather than castrated rats, as well as toward ovariectomized rather than sexually receptive females (Breuer et al., 2001; Farrell and McGinnis, 2003; Cunningham and McGinnis, 2006, 2007). McGinnis et al. (2002a), showed that 12 weeks of testosterone propionate exposure enhanced inter-male aggression in adult rats after physical provocation in the form of a mild tail pinch. Moreover, the environmental and social discriminating cues described above failed to alter testosterone-induced aggressive responses to physical provocation (McGinnis et al., 2002a,b). While testosterone clearly increases aggression, conflicting results have been reported in the literature concerning other commonly abused AAS (stanozolol, nandrolone decanoate, boldenone undecylenate) tested either in combination or individually. Salas-Ramirez et al. (2010) tested whether a 2-week administration of an AAS cocktail containing testosterone cypionate, nandrolone decanoate, and boldenone undecylenate had dissimilar behavioral consequences when drug exposure occurred during adolescence or adulthood. Higher aggression levels were observed in male Syrian hamsters exposed to an AAS cocktail compared to controls, regardless of age treatment (Salas-Ramirez et al., 2010). On the other hand, stanozolol failed to induce aggressive behavior in gonadectomized and intact rats and mice (Clark and Barber, 1994; Martinez-Sanchis et al., 1996; McGinnis et al., 2002a). More conflicting results have been reported by using nandrolone decanoate. Long et al. (1996) showed increased levels of aggression in Sprague–Dawley rats receiving chronic nandrolone decanoate, while no effect has been evidenced in Wistar rats (Zotti et al., 2014). Accordingly, adult rats exposed to mild physical provocation demonstrated decreased inter-male aggression when treated with stanozolol, while no effects of nandrolone have been reported (Breuer et al., 2001; Farrell and McGinnis, 2003). Regardless of the experimental methodologies employed to assess aggression, these findings suggest that strain, AAS chemical composition and regimen reflect the diversity of supra-therapeutic AAS exposure on behavioral responses in animals.
Several studies in preclinical models of aggression have investigated the AAS effects on the neurochemical changes in specific brain areas related to this behavior.
High aggression is often associated to decreased serotonin (5-HT) neurotransmission. Although this may account for high aggression as an individual feature, it has been suggested that serotonergic activity is probably higher during performance of aggressive behavior (van der Vegt et al., 2003).
In particular, testosterone propionate exposure decreased both 5-HT and 5-HT metabolite, 5-HIAA, in the hippocampus but not in the striatum or in the frontal cortex of adult rats (Bonson et al., 1994). Moreover, the aggressive behavior of dominant rats was decreased by treatment with selective agonists of 5-HT1A, 5-HT1B, and 5-HT2A∕2C receptors (Bonson et al., 1994). A significant decrease in 5-HT1A and 5-HT1B receptors immunoreactive staining has been shown in the latero-anterior hypothalamus and amygdala of hamsters treated with a mixture of AAS (Grimes and Melloni, 2005; Ricci et al., 2007). However, no decrease in the number of 5-HT1A receptor-expressing neurons and an increase in 5-HT2A receptor immunoreactivity have been reported in the hypothalamus (Ricci et al., 2006; Schwartzer et al., 2009). Ambar and Chiavegatto (2009) have reported reduced 5-HT1B mRNA levels in the hippocampus, hypothalamus, amygdala, and prefrontal cortex of nandrolone-treated mice suggesting that the serotonergic tone in these brain areas has a pivotal role for AAS-induced aggression in rodents (Ambar and Chiavegatto, 2009).
Reward
The data in literature highlight the potential for AAS addiction in humans (Kashkin and Kleber, 1989; Brower et al., 1990, 1991; Brower, 2002; Wood, 2004). Nevertheless, it is difficult to separate the direct rewarding effects of AAS from the psychological dependence of users on their physical appearance, muscular strength, and athletic performance. Hence, studies in animal models are a useful tool when examining androgen-reinforcing properties in conditions where anabolic effects and athletic performance are not relevant. Conditioned place preference (CPP) and self-administration are relevant experimental paradigms used to study reward in an experimental condition (Wood, 2004; Koob, 2006). Several studies in adult rodents have reported that systemic testosterone injections induced CPP in male rats and mice (de Beun et al., 1992; Alexander et al., 1994; Arnedo et al., 2000, 2002; Frye et al., 2001). In another animal model, it has been demonstrated that 15 days of administration of an AAS cocktail consisting of testosterone cypionate, nandrolone decanoate, and boldenone undecylenate, increased the rate of self-administration and enhanced the sensitivity to amphetamine challenge (Clark et al., 1996). However, in the same study, a 2 week treatment with MT had no effect on reward or performance of intracranial self-stimulation. In this light, Ballard and Wood (2005) have reported that in animals drostanolone and nandrolone tend to be self-administered (Ballard and Wood, 2005) and can cause CPP (Frye et al., 2002). Moreover, such effects can be prevented by dopaminergic antagonists (Schroeder and Packard, 2000) indicating that dopaminergic pathways are necessary for these behavioral outcomes. Indeed, the mesocorticolimbic circuitry, such as nucleus accumbens (NAc) and ventral tegmental area (VTA) are crucial for the reward system.
Parrilla-Carrero et al. (2009) investigated the rewarding effects of three different types of synthetic androgens differing in chemical structure and metabolism by using the CPP test in adult mice. They found that systemic injection of testosterone propionate and nandrolone decanoate, but not 17α-methyltestosterone, produced a dose-dependent shift in CPP suggesting that the rewarding properties of AAS might depend on their interaction with different pathways (Parrilla-Carrero et al., 2009). Very recently, the same research group has demonstrated nandrolone's failure to reward in adolescent mice (Martinez-Rivera et al., 2015). Although the literature reports that the adolescent brain is more sensitive to the reinforcing effects of drugs of abuse, this study suggests that such sensitivity may be drug dependent (Ernst et al., 2009; Galvan, 2010; Martinez-Rivera et al., 2015).
Packard et al. (1997) reported that testosterone induced CPP when directly injected into NAc (Packard et al., 1997). Similarly, Frye et al. (2002) showed that direct implants of testosterone or its metabolites (dihydrotestosterone, 3α-androstanediol) in the NAc shell induced a preference for the androgen-associated compartment, while no effect was observed with androgenic stimulation of the NAc core, suggesting a sub-region-specific functional role in reinforcement and reward pathway.
A growing body of evidence has shown the reinforcing effects of AAS using the experimental self-administration (oral, intravenous iv, intracerebroventricular icv) paradigm, which is considered as a model of addiction with the greatest face validity (Johnson and Wood, 2001; Wood, 2004; Frye, 2007; Frye et al., 2007). Wood (2002) demonstrated that gonadally intact adult male hamsters preferentially self-administer testosterone orally by using a food-induced drinking model (Wood, 2002). Although oral self-administration resembles oral AAS intake in humans, potential effects of taste solution or gut fill might present an inherent limitation on AAS oral consumption. Thus, Wood et al. (2004) used an operant chamber to train animals with chronic jugular cannulae and demonstrated an increase in testosterone iv self-administration compared to controls. Moreover, Syrian hamsters voluntarily consume testosterone through icv self-administration, suggesting that testosterone-reinforcing effects are centrally mediated (DiMeo and Wood, 2004; Wood, 2004). Ballard and Wood (2005) extended their research study on androgens and compared icv self-administration of four commonly abused AAS (nandrolone, drostanolone, oxymetholone, stanozolol) that differ in their method of administration, duration of action and metabolism. Results from this study showed that male hamsters preferentially self-administered nandrolone or drostanolone, which are two of the mostly used injectable androgens in humans. Conversely, animals failed to self-administer the orally active androgens oxymetholone or stanozolol, suggesting that injectable androgens may be more reinforcing than orally active steroids (Ballard and Wood, 2005).
To better understand the behavioral outcomes described above, various neurochemical studies have examined AAS effects on the monoaminergic system by measuring neurotransmitter and metabolite levels or by detecting receptors and enzyme alterations in key brain areas linked to the reward pathway. It has been reported that CPP induced by testosterone was blocked when adult male rats were directly injected into NAc with a D1-like or D2-like dopamine receptor antagonist (SCH23390 or sulpiride, respectively) (Schroeder and Packard, 2000). Sub-chronic administration of high AAS doses reduced dopamine D1-like receptor protein and mRNA levels in the NAc core and shell and increased D4-receptor mRNA expression in NAc, while D2-like receptors were up-regulated in the NAc core but down-regulated in the shell (Kindlundh et al., 2001b, 2003; Birgner et al., 2008a; Martinez-Rivera et al., 2015). An up-regulation of the dopamine transporter (DAT) protein was observed in vivo by a binding study using positron emission tomography (PET), in the striatum of male rat brain after chronic treatment with nandrolone (Kindlundh et al., 2002). Interestingly, Martinez-Rivera et al. (2015), observed no difference of D1-receptor protein expression in adolescent mice suggesting that the mesolimbic dopaminergic system during adolescence is immature or not sensitive to the rewarding response induced by nandrolone. Studies in Syrian hamsters suggested that testosterone reduced dopamine (DA) release in NAc (Triemstra et al., 2008). Likewise, our research group showed a reduction in DA content in NAc of rats treated for 4 weeks with nandrolone, changes which were accompanied by reduced hedonic-related behavior (Zotti et al., 2014). Furthermore, Birgner et al. (2007), in a microdialysis study, demonstrated that sub-chronic nandrolone decreased extracellular levels of DA metabolites (DOPAC and HVA) in rat NAc shell without affecting the release of DA. In line with these results, nandrolone was shown to reduce type A and B activity of monoamine oxidase (MAO) (Birgner et al., 2008b), although a previous study reported no effects of the drug on these enzymes activity in rats (Thiblin et al., 1999a). Further confirming the role of dopaminergic system in AAS effects on reward pathway, subchronic nandrolone has been shown to significantly down-regulate D1 receptors in the NAc and caudate putamen of rats, and to up-regulate D2-like receptors in the NAc core and VTA (Kindlundh et al., 2001b). In this regard, D1 and D2 receptors have been implicated in the reinforcing effects of drugs, as D1 is necessary for the acquisition of the effect and D2 crucial in mediating positive reinforcement (Missale et al., 1998). On the other hand, we have previously reported that stanozolol had no effect on DA content in NAc (Tucci et al., 2012). Findings regarding the impact of different AAS on brain reward function are summarized in Table 1.
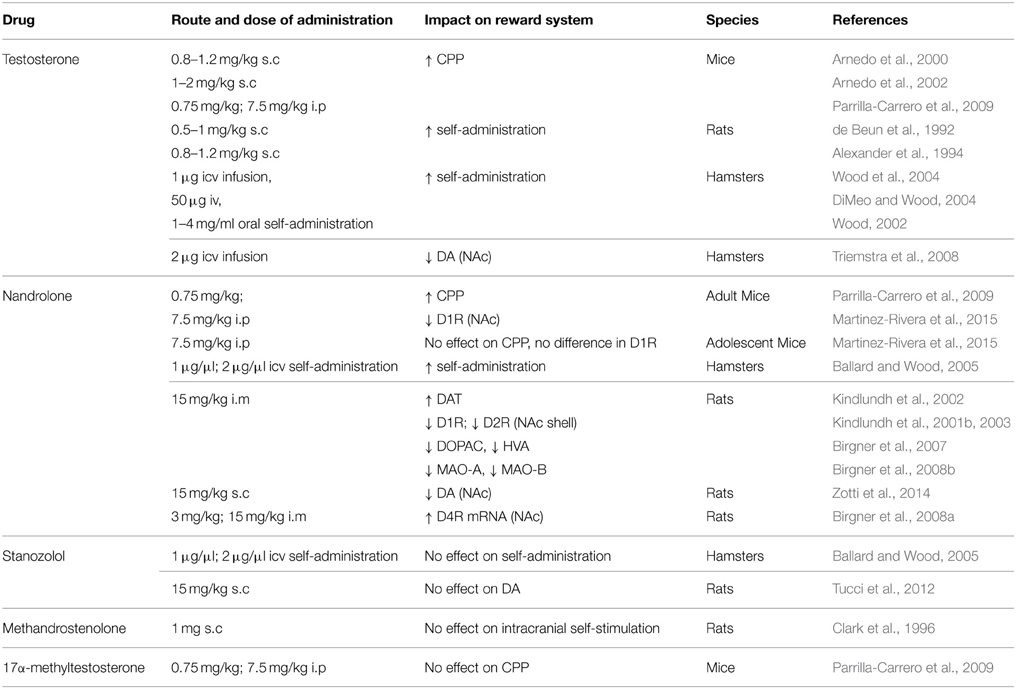
Table 1. Preclinical overview of the impact of different AAS on reward system.
Contradictory neurochemical results have been reported regarding AAS effects on the serotonergic system. In particular, intranasal administration of testosterone has been shown to increase dopaminergic and serotonergic systems in rat neostriatum and NAc (de Souza Silva et al., 2009). Accordingly, nandrolone decanoate and oxymethenolone treatment enhanced 5-HT and 5-HIAA concentrations in rat cerebral cortex and hypothalamus, while decreased levels of 5-HT and 5-HIAA were observed in the striatum of nandrolone-treated rats (Thiblin et al., 1999a; Lindqvist et al., 2002; Tamaki et al., 2003). Moreover, it has been shown that AAS affects 5-HT receptor expression. In particular, sub-chronic nandrolone administration down-regulates 5-HT1B and up-regulates 5-HT2 receptor density in rat brain (Kindlundh et al., 2003). In addition, McQueen et al. (1999) have demonstrated that serotonin transporter (SERT) mRNA-expressing cells in the dorsal raphe nucleus, as well as the density of SERT sites increase after sub-chronic treatment with testosterone (McQueen et al., 1999).
On the other hand, several studies have associated the endogenous opioid system to behaviors linked to reward and reinforcement (Gianoulakis, 2009). Thus, a number of experimental investigations have been carried out to ascertain whether AAS treatment modifies the levels of opioid peptides and their receptors in brain areas mediating reward. In particular, β-endorphin levels have been reported to significantly increase in the paraventricular thalamic nucleus and VTA of rats treated with AAS cocktails or nandrolone decanoate, respectively (Johansson et al., 1997; Harlan et al., 2000). In line with previous reports, chronic exposure to nandrolone decanoate has been linked to enhanced μ-, δ-, and κ-receptor binding in the hypothalamus, striatum, and midbrain periaqueductal gray (Johansson et al., 2000a). However, in the NAc shell and central amygdala of rats treated with the higher dose of nandrolone regimen, a down-regulation of κ-receptor binding, as measured by autoradiography has been demonstrated (Magnusson et al., 2009). Moreover, an increase in dynorphin converting enzyme-like activity was found only in the NAc of rats exposed to chronic nandrolone, suggesting an increased biosynthesis of dynorphin peptides, which, in turn, might affect basal DA levels in the NAc (Spanagel et al., 1992; Steiner and Gerfen, 1998; Magnusson et al., 2007).
Other AAS Pathways and Reward
It is worth noting that AAS effects are commonly described after chronic or sub-chronic drug exposure. Indeed, acute subcutaneous testosterone administration failed to influence accumbal DA release (Triemstra et al., 2008). In this regard, it has been proposed that AAS effects on the reinforcement system may be DA-independent, as happens with other abuse substances such as ethanol and benzodiazepines. Moreover, AAS effects on mesolimbic dopamine might be indirect or rely on non-classic androgen-sensitive pathways. Thus, based on accumulated evidence, AAS have an addictive potential, especially in susceptible subjects.
As reported, many discrepancies need to be better clarified. First, it is important to clarify whether classic nuclear receptors are involved in these effects or if other mechanisms are also involved. Moreover, scientific evidence exists for fast actions of steroids acting on calcium channels, membrane receptors, second messengers and membrane fluidity (for a review see Foradori et al., 2008). In this regard, a recent in vitro study has shown that testosterone, by acting on membrane receptors, was able to increase hippocampal plasticity within 2 h, leading to increased spine density (Li et al., 2015). Sato et al. (2010) investigated the possible involvement of these types of receptors on reinforcement effect of AAS. In particular, their experiments demonstrated that animals, intact or carrying the testicular feminization mutation, preferentially self-administer dihydrotestosterone (DHT) and DHT conjugated to bovine serum albumin, DHT-BSA, which acts only on cell surface. These observations prompted the authors to conclude that androgen self-administration may be mediated by plasma membrane receptors (Sato et al., 2010). Accordingly, it has been postulated that classical genomic action of androgen may be not fast enough to assure reinforcement. In agreement to such hypothesis, the distribution of androgen receptors in NAc and VTA is resulted quite sparse (Kritzer and Creutz, 2008; Sato et al., 2008).
Nonetheless, it is worth to note that further signaling systems, other than dopaminergic or opioidergic, can be implicated in reward. Indeed, chronic nandrolone decanoate was found to down-regulate the NR1 subunit of NMDA receptors in NAc of treated rats (Le Greves et al., 1997). This finding led the Authors to hypothesize that AAS may thus sensitize reward mechanisms.
However, the number of studies investigating the effects of AAS on glutamatergic system in reward is still limited; hence, future investigations should be focused accordingly to clarify whether AAS reinforcement relies on non-classic pathways or on other signaling systems.
AAS Effects on Other Drugs of Abuse
Clinical and epidemiological data have reported that the abuse of AAS in humans is often associated with the abuse of psychotropic drugs, such as cocaine, opiates, alcohol, cannabis, amphetamine, and 3,4-methylenedioxy-methamphetamine (MDMA). These surveys have suggested a role of AAS as a gateway to other dependency-inducing drugs (DuRant et al., 1995; Arvary and Pope, 2000; Kindlundh et al., 2001a; Kanayama et al., 2003a; Thevis et al., 2008). Based on these findings, different animal paradigms have been used to investigate AAS pre-exposure effects on neurochemical and behavioral response to other addictive substances. Consistent with reported higher alcohol intake in AAS abusers, increased voluntary alcohol consumption after cessation of AAS administration has also been observed in male adult rats (Johansson et al., 2000b). In line with these findings, corticotropin releasing factor modulation of GABAergic transmission in the amygdala seems to play a pivotal role in ethanol effects, suggesting that AAS might alter the sensitivity of these circuits and predispose to alcohol abuse (Roberto et al., 2004, 2010; Oberlander and Henderson, 2012). Chronic nandrolone decanoate administration has been found to significantly impair CPP induced by Δ9-tetrahydrocannabinol (THC) without affecting CB1 receptor binding. Interestingly, nandrolone administration increased THC abstinence precipitated by the CB1 cannabinoid antagonist rimonabant (Celerier et al., 2006).
Administration of supra-pharmacological doses of nandrolone decanoate has been shown to decrease the hyper-locomotion and stereotyped behavior induced by amphetamine and MDMA, in a dose-dependent manner (Kurling et al., 2008). Such behavioral outcomes have been corroborated by microdialysis results. In particular, nandrolone decanoate attenuated the effect of amphetamine and MDMA on DA baseline and DA metabolites levels in the NAc. However, the higher dose of nandrolone decanoate has enhanced the acute effects of MDMA-induced release of 5-HT, followed by exhaustion of neuronal 5-HT stores. Thus, high-dose nandrolone decanoate treatment might enhance neuron vulnerability to MDMA, leading to effects resembling MDMA neurotoxicity (Kurling et al., 2008). In addition, it has been demonstrated that the effects of amphetamine on the hippocampal and hypothalamic DOPAC/DA ratio were prevented by nandrolone decanoate, with no changes to DA baseline levels (Birgner et al., 2007). Likewise, it has been shown that pretreatment with nandrolone decanoate attenuates accumbal DA and 5-HT outflow, as well as the consequent stereotyped behavior induced by cocaine (Kurling-Kailanto et al., 2010; Kailanto et al., 2011). Nandrolone might decrease neurochemical and behavioral effects induced by cocaine via up-regulation of DAT and SERT binding sites. In these studies, the authors showed that changes in DA and 5-HT systems endure, even after a long recovery period from the last dose of nandrolone. This confirms the hypothesis that drug abuse causes long lasting changes in brain dopaminergic and serotonergic pathways (Kurling et al., 2008; Kailanto et al., 2011). These data are in line with earlier findings demonstrating that chronic cocaine and methamphetamine decreased D2-receptor and DAT expression during withdrawal and lasted up to 11 months after the last drug administration (Volkow et al., 1990, 2001a,b). Collectively, these results demonstrate that pre-treatment with nandrolone decanoate dose-dependently attenuates neurochemical and behavioral effects relating to the reward system induced by psychostimulant drugs. These findings indicate that such reduced dopaminergic and serotonergic activity in brain regions strictly involved in the reward system might represent the neurochemical substrate that could underlie a higher prevalence of illicit drug use among AAS abusers. Indeed, to achieve the desired effect of psychostimulant drugs, AAS users may require increased doses of these substances.
On the other hand, testosterone has been hypothesized to act as a partial agonist on the opiod system considering that, depending on type of receptors involved, steroid effects are brain region specific (Wood, 2008). As the reinforcing effects of opioids are thought to be mediated principally by μ- and δ-receptors (Peters and Wood, 2005), many data in the literature are available with regard to altered opioid receptor binding after AAS. In particular, nandrolone has been reported to increase binding of μ-, δ-, and κ-receptors in the hypothalamus, striatum, and midbrain periaqueductal gray (Johansson et al., 2000a), while reduced κ-receptors have been found in NAc (Johansson et al., 2000b). In addition, increased β-endorphin levels in the VTA (Johansson et al., 1997) and paraventricular thalamus (PVT) (Harlan et al., 2000) along with and higher β-endorphin fiber staining in bed nucleus of the stria terminalis and PVT (Menard et al., 1995) have been described. However, the total β-endorphin immunoreactivity is lower in arcuate nucleus (Menard et al., 1995).
On the other hand, nandrolone has been found to enhance morphine-induced hypothermia while testosterone increases the antinociceptive effect of a κ-agonist. However, contrasting data exist since no effects of AAS on morphine antinociception have been reported in other animal models (Negus et al., 2001; Celerier et al., 2003; Philipova et al., 2003). In fact, nandrolone pre-exposure has been shown to inhibit tolerance to antinociceptive properties of morphine and CPP induced by morphine in mice (Celerier et al., 2003) and rats (Philipova et al., 2003). Accordingly, pre-exposure to AAS has been shown to prevent morphine-induced striatal Fos expression (Harlan et al., 2000). High variability is present in findings linking AAS to opiate withdrawal. In monkeys no effect of AAS has been described for naloxone-precipitated morphine withdrawal paradigm, although Celerier et al. (2003) found that nandrolone increased withdrawal symptoms induced by naloxone in morphine-dependent mice. Moreover, the dysphoric effect mediated by nandrolone pre-treatment has been correlated to elevated striatal levels of dynorphin B, which in turn may account for the inhibition of dopaminergic activity in this brain region (Steiner and Gerfen, 1998; Johansson et al., 2000a). Finally, testosterone has been shown not to increase motivation for morphine (Cooper and Wood, 2014). Such discrepancies may rely on different AAS and schedule of treatment used, as well as different species or strain of animal used.
Although literature has been populated by many clinical or preclinical reports, many knots in the unraveling of deleterious addictive effects of AAS still need to be untied. Furthermore, taking into account that the use of these substances is becoming popular, especially among adolescents, a deeper knowledge of CNS effects of AAS is nowadays mandatory.
Funding
This review was supported by PRIN 2011 (PT) from MIUR.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Aitken, C., Delalande, C., and Stanton, K. (2002). Pumping iron, risking infection? Exposure to hepatitis C, hepatitis B and HIV among anabolic-androgenic steroid injectors in Victoria, Australia. Drug Alcohol Depend. 65, 303–308. doi: 10.1016/S0376-8716(01)00174-0
Alexander, G. M., Packard, M. G., and Hines, M. (1994). Testosterone has rewarding affective properties in male rats: implications for the biological basis of sexual motivation. Behav. Neurosci. 108, 424–428. doi: 10.1037/0735-7044.108.2.424
Ambar, G., and Chiavegatto, S. (2009). Anabolic-androgenic steroid treatment induces behavioral disinhibition and downregulation of serotonin receptor messenger RNA in the prefrontal cortex and amygdala of male mice. Genes Brain Behav. 8, 161–173. doi: 10.1111/j.1601-183X.2008.00458.x
Anderson, R. A., and Wu, F. C. (1996). Comparison between testosterone enanthate-induced azoospermia and oligozoospermia in a male contraceptive study. II. Pharmacokinetics and pharmacodynamics of once weekly administration of testosterone enanthate. J. Clin. Endocrinol. Metab. 81, 896–901.
Arnedo, M. T., Salvador, A., Martinez-Sanchis, S., and Gonzalez-Bono, E. (2000). Rewarding properties of testosterone in intact male mice: a pilot study. Pharmacol. Biochem. Behav. 65, 327–332. doi: 10.1016/S0091-3057(99)00189-6
Arnedo, M. T., Salvador, A., Martínez-Sanchís, S., and Pellicer, O. (2002). Similar rewarding effects of testosterone in mice rated as short and long attack latency individuals. Addict. Biol. 7, 373–379. doi: 10.1080/1355621021000005955
Arvary, D., and Pope, H. G. Jr. (2000). Anabolic-androgenic steroids as a gateway to opioid dependence. N. Engl. J. Med. 342, 1532. doi: 10.1056/NEJM200005183422018
Baggish, A. L., Weiner, R. B., Kanayama, G., Hudson, J. I., Picard, M. H., Hutter, A. M. Jr., et al. (2010). Long-term anabolic-androgenic steroid use is associated with left ventricular dysfunction. Circ. Heart Fail. 3, 472–476. doi: 10.1161/CIRCHEARTFAILURE.109.931063
Bahrke, M. S., and Yesalis, C. E. III. (1994). Weight training. A potential confounding factor in examining the psychological and behavioural effects of anabolic-androgenic steroids. Sports Med. 18, 309–318. doi: 10.2165/00007256-199418050-00003
Ballard, C. L., and Wood, R. I. (2005). Intracerebroventricular self-administration of commonly abused anabolic-androgenic steroids in male hamsters (Mesocricetus auratus): nandrolone, drostanolone, oxymetholone, and stanozolol. Behav. Neurosci. 119, 752–758. doi: 10.1037/0735-7044.119.3.752
Basaria, S., Wahlstrom, J. T., and Dobs, A. S. (2001). Clinical review 138: anabolic-androgenic steroid therapy in the treatment of chronic diseases. J. Clin. Endocrinol. Metab. 86, 5108–5117. doi: 10.1210/jcem.86.11.7983
Bhasin, S., Storer, T. W., Berman, N., Callegari, C., Clevenger, B., Phillips, J., et al. (1996). The effects of supraphysiologic doses of testosterone on muscle size and strength in normal men. N. Engl. J. Med. 335, 1–7. doi: 10.1056/NEJM199607043350101
Bhasin, S., Woodhouse, L., Casaburi, R., Singh, A. B., Bhasin, D., Berman, N., et al. (2001). Testosterone dose-response relationships in healthy young men. Am. J. Physiol. Endocrinol. Metab. 281, E1172–E1181.
Birgner, C., Kindlundh-Hogberg, A. M., Alsio, J., Lindblom, J., Schioth, H. B., and Bergstrom, L. (2008a). The anabolic androgenic steroid nandrolone decanoate affects mRNA expression of dopaminergic but not serotonergic receptors. Brain Res. 1240, 221–228. doi: 10.1016/j.brainres.2008.09.003
Birgner, C., Kindlundh-Högberg, A. M., Nyberg, F., and Bergström, L. (2007). Altered extracellular levels of DOPAC and HVA in the rat nucleus accumbens shell in response to sub-chronic nandrolone administration and a subsequent amphetamine challenge. Neurosci. Lett. 412, 168–172. doi: 10.1016/j.neulet.2006.11.001
Birgner, C., Kindlundh-Högberg, A. M., Oreland, L., Alsiö, J., Lindblom, J., Schiöth, H. B., et al. (2008b). Reduced activity of monoamine oxidase in the rat brain following repeated nandrolone decanoate administration. Brain Res. 1219, 103–110. doi: 10.1016/j.brainres.2008.05.014
Bonson, K. R., Johnson, R. G., Fiorella, D., Rabin, R. A., and Winter, J. C. (1994). Serotonergic control of androgen-induced dominance. Pharmacol. Biochem. Behav. 49, 313–322. doi: 10.1016/0091-3057(94)90427-8
Breuer, M. E., McGinnis, M. Y., Lumia, A. R., and Possidente, B. P. (2001). Aggression in male rats receiving anabolic androgenic steroids: effects of social and environmental provocation. Horm. Behav. 40, 409–418. doi: 10.1006/hbeh.2001.1706
Brower, K. J. (2000). “Anabolic steroids: potential for physical and psychological dependence,” in Anabolic Steroids in Sport and Exercise, 2nd Edn., ed C. E. Yesalis (Champaign, IL: Human Kinetics), 280–304.
Brower, K. J. (2002). Anabolic steroid abuse and dependence. Curr. Psychiatry Rep. 4, 377–387. doi: 10.1007/s11920-002-0086-6
Brower, K. J., Blow, F. C., Young, J. P., and Hill, E. M. (1991). Symptoms and correlates of anabolic-androgenic steroid dependence. Br. J. Addict. 86, 759–768. doi: 10.1111/j.1360-0443.1991.tb03101.x
Brower, K. J., Eliopulos, G. A., Blow, F. C., Catlin, D. H., and Beresford, T. P. (1990). Evidence for physical and psychological dependence on anabolic androgenic steroids in eight weight lifters. Am. J. Psychiatry 147, 510–512. doi: 10.1176/ajp.147.4.510
Célérier, E., Ahdepil, T., Wikander, H., Berrendero, F., Nyberg, F., and Maldonado, R. (2006). Influence of the anabolic-androgenic steroid nandrolone on cannabinoid dependence. Neuropharmacology 50, 788–806. doi: 10.1016/j.neuropharm.2005.11.017
Célérier, E., Yazdi, M. T., Castañé, A., Ghozland, S., Nyberg, F., and Maldonado, R. (2003). Effects of nandrolone on acute morphine responses, tolerance and dependence in mice. Eur. J. Pharmacol. 465, 69–81. doi: 10.1016/S0014-2999(03)01462-6
Christie, M. H., and Barfield, R. J. (1979). Effects of aromatizable androgens on aggressive behaviour among rats (rattus norvegicus). J. Endocrinol. 83, 17–26. doi: 10.1677/joe.0.0830017
Clark, A. S., and Barber, D. M. (1994). Anabolic-androgenic steroids and aggression in castrated male rats. Physiol. Behav. 56, 1107–1113. doi: 10.1016/0031-9384(94)90351-4
Clark, A. S., and Henderson, L. P. (2003). Behavioral and physiological responses to anabolic-androgenic steroids. Neurosci. Biobehav. Rev. 27, 413–436. doi: 10.1016/S0149-7634(03)00064-2
Clark, A. S., Lindenfeld, R. C., and Gibbons, C. H. (1996). Anabolic-androgenic steroids and brain reward. Pharmacol. Biochem. Behav. 53, 741–745. doi: 10.1016/0091-3057(95)02082-9
Conway, A. J., Handelsman, D. J., Lording, D. W., Stuckey, B., and Zajac, J. D. (2000). Use, misuse and abuse of androgens. The Endocrine Society of Australia consensus guidelines for androgen prescribing. Med. J. Aust. 172, 220–224.
Cooper, S. E., and Wood, R. I. (2014). Androgens and opiates: testosterone interaction with morphine self-administration in male rats. Neuroreport 25, 521–526. doi: 10.1097/wnr.0000000000000125
Coviello, A. D., Kaplan, B., Lakshman, K. M., Chen, T., Singh, A. B., and Bhasin, S. (2008). Effects of graded doses of testosterone on erythropoiesis in healthy young and older men. J. Clin. Endocrinol. Metab. 93, 914–919. doi: 10.1210/jc.2007-1692
Cunningham, R. L., and McGinnis, M. Y. (2006). Physical provocation of pubertal anabolic androgenic steroid exposed male rats elicits aggression towards females. Horm. Behav. 50, 410–416. doi: 10.1016/j.yhbeh.2006.05.002
Cunningham, R. L., and McGinnis, M. Y. (2007). Factors influencing aggression toward females by male rats exposed to anabolic androgenic steroids during puberty. Horm. Behav. 51, 135–141. doi: 10.1016/j.yhbeh.2006.09.003
Daly, R. C., Su, T. P., Schmidt, P. J., Pickar, D., Murphy, D. L., and Rubinow, D. R. (2001). Cerebrospinal fluid and behavioral changes after methyltestosterone administration: preliminary findings. Arch. Gen. Psychiatry 58, 172–177. doi: 10.1001/archpsyc.58.2.172
Dean, C. E. (2000). Prasterone (DHEA) and mania. Ann. Pharmacother. 34, 1419–1422. doi: 10.1345/aph.10115
de Beun, R., Jansen, E., Slangen, J. L., and Van de Poll, N. E. (1992). Testosterone as appetitive and discriminative stimulus in rats: sex- and dose-dependent effects. Physiol. Behav. 52, 629–634. doi: 10.1016/0031-9384(92)90389-J
de Souza Silva, M. A., Mattern, C., Topic, B., Buddenberg, T. E., and Huston, J. P. (2009). Dopaminergic and serotonergic activity in neostriatum and nucleus accumbens enhanced by intranasal administration of testosterone. Eur. Neuropsychopharmacol. 19, 53–63. doi: 10.1016/j.euroneuro.2008.08.003
DiMeo, A. N., and Wood, R. I. (2004). Circulating androgens enhance sensitivity to testosterone self-administration in male hamsters. Pharmacol. Biochem. Behav. 79, 383–389. doi: 10.1016/j.pbb.2004.08.015
Dohle, G. R., Smit, M., and Weber, R. F. (2003). Androgens and male fertility. World J. Urol. 21, 341–345. doi: 10.1007/s00345-003-0365-9
DuRant, R. H., Escobedo, L. G., and Heath, G. W. (1995). Anabolic-steroid use, strength training, and multiple drug use among adolescents in the United States. Pediatrics 96, 23–28.
Ernst, M., Romeo, R. D., and Andersen, S. L. (2009). Neurobiology of the development of motivated behaviors in adolescence: a window into a neural systems model. Pharmacol. Biochem. Behav. 93, 199–211. doi: 10.1016/j.pbb.2008.12.013
Far, H. R., Ågren, G., and Thiblin, I. (2012). Cardiac hypertrophy in deceased users of anabolic androgenic steroids: an investigation of autopsy findings. Cardiovasc. Pathol. 21, 312–316. doi: 10.1016/j.carpath.2011.10.002
Farrell, S. F., and McGinnis, M. Y. (2003). Effects of pubertal anabolic-androgenic steroid (AAS) administration on reproductive and aggressive behaviors in male rats. Behav. Neurosci. 117, 904–911. doi: 10.1037/0735-7044.117.5.904
Ferreira, I., Brooks, D., Lacasse, Y., and Goldstein, R. (2001). Nutritional intervention in COPD: a systematic overview. Chest 119, 353–363. doi: 10.1378/chest.119.2.353
Foradori, C. D., Weiser, M. J., and Handa, R. J. (2008). Non-genomic actions of androgens. Front. Neuroendocrinol. 29, 169–181. doi: 10.1016/j.yfrne.2007.10.005
Franke, W. W., and Berendonk, B. (1997). Hormonal doping and androgenization of athletes: a secret program of the German Democratic Republic government. Clin. Chem. 43, 1262–1279.
Frye, C. A. (2007). Some rewarding effects of androgens may be mediated by actions of its 5alpha-reduced metabolite 3alpha-androstanediol. Pharmacol. Biochem. Behav. 86, 354–367. doi: 10.1016/j.pbb.2006.10.003
Frye, C. A., Babson, A., and Walf, A. A. (2007). Self-administration of 3alpha-androstanediol increases locomotion and analgesia and decreases aggressive behavior of male hamsters. Pharmacol. Biochem. Behav. 86, 415–421. doi: 10.1016/j.pbb.2006.05.025
Frye, C. A., Park, D., Tanaka, M., Rosellini, R., and Svare, B. (2001). The testosterone metabolite and neurosteroid 3alpha-androstanediol may mediate the effects of testosterone on conditioned place preference. Psychoneuroendocrinology 26, 731–750. doi: 10.1016/S0306-4530(01)00027-0
Frye, C. A., Rhodes, M. E., Rosellini, R., and Svare, B. (2002). The nucleus accumbens as a site of action for rewarding properties of testosterone and its 5alpha-reduced metabolites. Pharmacol. Biochem. Behav. 74, 119–127. doi: 10.1016/S0091-3057(02)00968-1
Furlanello, F., Bentivegna, S., Cappato, R., and De Ambroggi, L. (2003). Arrhythmogenic effects of illicit drugs in athletes. Ital. Heart J. 4, 829–837.
Galvan, A. (2010). Adolescent development of the reward system. Front. Hum. Neurosci. 4:6. doi: 10.3389/neuro.09.006.2010
Gianoulakis, C. (2009). Endogenous opioids and addiction to alcohol and other drugs of abuse. Curr. Top. Med. Chem. 9, 999–1015. doi: 10.2174/156802609789630956
Grimes, J. M., and Melloni, R. H. Jr. (2005). Serotonin-1B receptor activity and expression modulate the aggression-stimulating effects of adolescent anabolic steroid exposure in hamsters. Behav. Neurosci. 119, 1184–1194. doi: 10.1037/0735-7044.119.5.1184
Gupta, V., Bhasin, S., Guo, W., Singh, R., Miki, R., Chauhan, P., et al. (2008). Effects of dihydrotestosterone on differentiation and proliferation of human mesenchymal stem cells and preadipocytes. Mol. Cell. Endocrinol. 296, 32–40. doi: 10.1016/j.mce.2008.08.019
Hallberg, M., Johansson, P., Kindlundh, A. M., and Nyberg, F. (2000). Anabolic-androgenic steroids affect the content of substance P and substance P(1-7) in the rat brain. Peptides 21, 845–852. doi: 10.1016/S0196-9781(00)00218-7
Harlan, R. E., Brown, H. E., Lynch, C. S., D'Souza, D., and Garcia, M. M. (2000). Androgenic-anabolic steroids blunt morphine-induced c-fos expression in the rat striatum: possible role of beta-endorphin. Brain Res. 853, 99–104. doi: 10.1016/S0006-8993(99)02257-X
Harrison, R. J., Connor, D. F., Nowak, C., Nash, K., and Melloni, R. H. Jr. (2000). Chronic anabolic-androgenic steroid treatment during adolescence increases anterior hypothalamic vasopressin and aggression in intact hamsters. Psychoneuroendocrinology 25, 317–338. doi: 10.1016/S0306-4530(99)00057-8
Hart, D. W., Wolf, S. E., Ramzy, P. I., Chinkes, D. L., Beauford, R. B., Ferrando, A. A., et al. (2001). Anabolic effects of oxandrolone after severe burn. Ann. Surg. 233, 556–564. doi: 10.1097/00000658-200104000-00012
Hassan, N. A., Salem, M. F., and Sayed, M. A. (2009). Doping and effects of anabolic androgenic steroids on the heart: histological, ultrastructural, and echocardiographic assessment in strength athletes. Hum. Exp. Toxicol. 28, 273–283. doi: 10.1177/0960327109104821
Haupt, H. A. (2001). Upper extremity injuries associated with strength training. Clin. Sports Med. 20, 481–490. doi: 10.1016/S0278-5919(05)70264-7
Irving, L. M., Wall, M., Neumark-Sztainer, D., and Story, M. (2002). Steroid use among adolescents: findings from Project EAT. J. Adolesc. Health 30, 243–252. doi: 10.1016/S1054-139X(01)00414-1
Johansson, P., Hallberg, M., Kindlundh, A., and Nyberg, F. (2000a). The effect on opioid peptides in the rat brain, after chronic treatment with the anabolic androgenic steroid, nandrolone decanoate. Brain Res. Bull. 51, 413–418. doi: 10.1016/S0361-9230(99)00263-4
Johansson, P., Lindqvist, A., Nyberg, F., and Fahlke, C. (2000b). Anabolic androgenic steroids affects alcohol intake, defensive behaviors and brain opioid peptides in the rat. Pharmacol. Biochem. Behav. 67, 271–279. doi: 10.1016/S0091-3057(00)00365-8
Johansson, P., Ray, A., Zhou, Q., Huang, W., Karlsson, K., and Nyberg, F. (1997). Anabolic androgenic steroids increase beta-endorphin levels in the ventral tegmental area in the male rat brain. Neurosci. Res. 27, 185–189. doi: 10.1016/S0168-0102(96)01141-8
Johnson, L. R., and Wood, R. I. (2001). Oral testosterone self-administration in male hamsters. Neuroendocrinology 73, 285–292. doi: 10.1159/000054645
Kailanto, S., Kankaanpää, A., and Seppälä, T. (2011). Subchronic steroid administration induces long lasting changes in neurochemical and behavioral response to cocaine in rats. Steroids 76, 1310–1316. doi: 10.1016/j.steroids.2011.06.011
Kanayama, G., Cohane, G. H., Weiss, R. D., and Pope, H. G. (2003a). Past anabolic-androgenic steroid use among men admitted for substance abuse treatment: an underrecognized problem? J. Clin. Psychiatry 64, 156–160. doi: 10.4088/JCP.v64n0208
Kanayama, G., Hudson, J. I., and Pope, H. G. Jr. (2009). Features of men with anabolic-androgenic steroid dependence: a comparison with nondependent AAS users and with AAS nonusers. Drug Alcohol Depend. 102, 130–137. doi: 10.1016/j.drugalcdep.2009.02.008
Kanayama, G., Pope, H. G., Cohane, G., and Hudson, J. I. (2003b). Risk factors for anabolic-androgenic steroid use among weightlifters: a case-control study. Drug Alcohol Depend. 71, 77–86. doi: 10.1016/S0376-8716(03)00069-3
Kashkin, K. B., and Kleber, H. D. (1989). Hooked on hormones? An anabolic steroid addiction hypothesis. JAMA 262, 3166–3170. doi: 10.1001/jama.1989.03430220089036
Kindlundh, A. M., Bergström, M., Monazzam, A., Hallberg, M., Blomqvist, G., Långström, B., et al. (2002). Dopaminergic effects after chronic treatment with nandrolone visualized in rat brain by positron emission tomography. Prog. Neuropsychopharmacol. Biol. Psychiatry 26, 1303–1308. doi: 10.1016/S0278-5846(02)00293-2
Kindlundh, A. M., Hagekull, B., Isacson, D. G., and Nyberg, F. (2001a). Adolescent use of anabolic-androgenic steroids and relations to self-reports of social, personality and health aspects. Eur. J. Public Health 11, 322–328. doi: 10.1093/eurpub/11.3.322
Kindlundh, A. M., Isacson, D. G., Berglund, L., and Nyberg, F. (1999). Factors associated with adolescent use of doping agents: anabolic-androgenic steroids. Addiction 94, 543–553. doi: 10.1046/j.1360-0443.1999.9445439.x
Kindlundh, A. M., Lindblom, J., Bergström, L., Wikberg, J. E., and Nyberg, F. (2001b). The anabolic-androgenic steroid nandrolone decanoate affects the density of dopamine receptors in the male rat brain. Eur. J. Neurosci. 13, 291–296. doi: 10.1046/j.0953-816X.2000.01402.x
Kindlundh, A. M., Lindblom, J., and Nyberg, F. (2003). Chronic administration with nandrolone decanoate induces alterations in the gene-transcript content of dopamine D(1)- and D(2)-receptors in the rat brain. Brain Res. 979, 37–42. doi: 10.1016/S0006-8993(03)02843-9
Koob, G. F. (2006). The neurobiology of addiction: a neuroadaptational view relevant for diagnosis. Addiction 101(Suppl. 1), 23–30. doi: 10.1111/j.1360-0443.2006.01586.x
Krieg, A., Scharhag, J., Albers, T., Kindermann, W., and Urhausen, A. (2007). Cardiac tissue Doppler in steroid users. Int. J. Sports Med. 28, 638–643. doi: 10.1055/s-2007-964848
Kritzer, M. F., and Creutz, L. M. (2008). Region and sex differences in constituent dopamine neurons and immunoreactivity for intracellular estrogen and androgen receptors in mesocortical projections in rats. J. Neurosci. 28, 9525–9535. doi: 10.1523/JNEUROSCI.2637-08.2008
Kuhn, C. M. (2002). Anabolic steroids. Recent Prog. Horm. Res. 57, 411–434. doi: 10.1210/rp.57.1.411
Kurling, S., Kankaanpää, A., and Seppälä, T. (2008). Sub-chronic nandrolone treatment modifies neurochemical and behavioral effects of amphetamine and 3,4-methylenedioxymethamphetamine (MDMA) in rats. Behav. Brain Res. 189, 191–201. doi: 10.1016/j.bbr.2007.12.021
Kurling-Kailanto, S., Kankaanpää, A., and Seppälä, T. (2010). Subchronic nandrolone administration reduces cocaine-induced dopamine and 5-hydroxytryptamine outflow in the rat nucleus accumbens. Psychopharmacology (Berl.) 209, 271–281. doi: 10.1007/s00213-010-1796-9
Kutscher, E. C., Lund, B. C., and Perry, P. J. (2002). Anabolic steroids: a review for the clinician. Sports Med. 32, 285–296. doi: 10.2165/00007256-200232050-00001
Langer, C. J., Hoffman, J. P., and Ottery, F. D. (2001). Clinical significance of weight loss in cancer patients: rationale for the use of anabolic agents in the treatment of cancer-related cachexia. Nutrition 17, S1–S20. doi: 10.1016/s0899-9007(01)80001-0
Le Grevès, P., Huang, W., Johansson, P., Thörnwall, M., Zhou, Q., and Nyberg, F. (1997). Effects of an anabolic-androgenic steroid on the regulation of the NMDA receptor NR1, NR2A and NR2B subunit mRNAs in brain regions of the male rat. Neurosci. Lett. 226, 61–64. doi: 10.1016/S0304-3940(97)00244-9
Li, S., Kang, L., Zhang, Y., Feng, B., Du, J., and Cui, H. (2015). Detecting the presence of hippocampus membrane androgen receptors in male SAMP8 mice and their induced synaptic plasticity. Mol. Cell. Endocrinol. 414, 82–90. doi: 10.1016/j.mce.2015.07.005
Lindqvist, A. S., Johansson-Steensland, P., Nyberg, F., and Fahlke, C. (2002). Anabolic androgenic steroid affects competitive behaviour, behavioural response to ethanol and brain serotonin levels. Behav. Brain Res. 133, 21–29. doi: 10.1016/S0166-4328(01)00408-9
Long, S. F., Wilson, M. C., and Davis, W. M. (2000). The effects of nandrolone decanoate on cocaine-induced kindling in male rats. Neuropharmacology 39, 2442–2447. doi: 10.1016/S0028-3908(00)00061-7
Long, S. F., Wilson, M. C., Sufka, K. J., and Davis, W. M. (1996). The effects of cocaine and nandrolone co-administration on aggression in male rats. Prog. Neuropsychopharmacol. Biol. Psychiatry 20, 839–856. doi: 10.1016/0278-5846(96)00063-2
Luijkx, T., Velthuis, B. K., Backx, F. J., Buckens, C. F., Prakken, N. H., Rienks, R., et al. (2013). Anabolic androgenic steroid use is associated with ventricular dysfunction on cardiac MRI in strength trained athletes. Int. J. Cardiol. 167, 664–668. doi: 10.1016/j.ijcard.2012.03.072
Lumia, A. R., and McGinnis, M. Y. (2010). Impact of anabolic androgenic steroids on adolescent males. Physiol. Behav. 100, 199–204. doi: 10.1016/j.physbeh.2010.01.007
Lumia, A. R., Thorner, K. M., and McGinnis, M. Y. (1994). Effects of chronically high doses of the anabolic androgenic steroid, testosterone, on intermale aggression and sexual behavior in male rats. Physiol. Behav. 55, 331–335. doi: 10.1016/0031-9384(94)90142-2
Magnusson, K., Birgner, C., Bergström, L., Nyberg, F., and Hallberg, M. (2009). Nandrolone decanoate administration dose-dependently affects the density of kappa opioid peptide receptors in the rat brain determined by autoradiography. Neuropeptides 43, 105–111. doi: 10.1016/j.npep.2008.12.005
Magnusson, K., Hallberg, M., Bergquist, J., and Nyberg, F. (2007). Enzymatic conversion of dynorphin A in the rat brain is affected by administration of nandrolone decanoate. Peptides 28, 851–858. doi: 10.1016/j.peptides.2006.12.011
Manoharan, G., Campbell, N. P., and O'Brien, C. J. (2002). Syncopal episodes in a young amateur body builder. Br. J. Sports Med. 36, 67–68. doi: 10.1136/bjsm.36.1.67
Martinez-Rivera, F. J., Natal-Albelo, E. J., Martínez, N. A., Orozco-Vega, R. A., Muñiz-Seda, O. A., and Barreto-Estrada, J. L. (2015). The effect of the anabolic steroid, nandrolone, in conditioned place preference and D1 dopamine receptor expression in adolescent and adult mice. Behav. Processes 113, 81–85. doi: 10.1016/j.beproc.2015.01.008
Martínez-Sanchis, S., Brain, P. F., Salvador, A., and Simón, V. M. (1996). Long-term chronic treatment with stanozolol lacks significant effects on aggression and activity in young and adult male laboratory mice. Gen. Pharmacol. 27, 293–298. doi: 10.1016/0306-3623(95)02019-5
McBride, A. J., Williamson, K., and Petersen, T. (1996). Three cases of nalbuphine hydrochloride dependence associated with anabolic steroid use. Br. J. Sports Med. 30, 69–70. doi: 10.1136/bjsm.30.1.69
McGinnis, M. Y. (2004). Anabolic androgenic steroids and aggression: studies using animal models. Ann. N.Y. Acad. Sci. 1036, 399–415. doi: 10.1196/annals.1330.024
McGinnis, M. Y., Lumia, A. R., Breuer, M. E., and Possidente, B. (2002a). Physical provocation potentiates aggression in male rats receiving anabolic androgenic steroids. Horm. Behav. 41, 101–110. doi: 10.1006/hbeh.2001.1742
McGinnis, M. Y., Lumia, A. R., and Possidente, B. P. (2002b). Effects of withdrawal from anabolic androgenic steroids on aggression in adult male rats. Physiol. Behav. 75, 541–549. doi: 10.1016/S0031-9384(02)00657-1
McQueen, J. K., Wilson, H., Sumner, B. E., and Fink, G. (1999). Serotonin transporter (SERT) mRNA and binding site densities in male rat brain affected by sex steroids. Brain Res. Mol. Brain Res. 63, 241–247. doi: 10.1016/S0169-328X(98)00281-2
Menard, C. S., Hebert, T. J., Dohanich, G. P., and Harlan, R. E. (1995). Androgenic-anabolic steroids modify beta-endorphin immunoreactivity in the rat brain. Brain Res. 669, 255–262. doi: 10.1016/0006-8993(94)01266-K
Midgley, S. J., Heather, N., and Davies, J. B. (1999). Dependence-producing potential of anabolic-androgenic steroids. Addict. Res. Theory 7, 539–550. doi: 10.3109/16066359909004404
Missale, C., Nash, S. R., Robinson, S. W., Jaber, M., and Caron, M. G. (1998). Dopamine receptors: from structure to function. Physiol. Rev. 78, 189–225.
Montisci, M., El Mazloum, R., Cecchetto, G., Terranova, C., Ferrara, S. D., Thiene, G., et al. (2012). Anabolic androgenic steroids abuse and cardiac death in athletes: morphological and toxicological findings in four fatal cases. Forensic Sci. Int. 217, e13–e18. doi: 10.1016/j.forsciint.2011.10.032
Morley, J. E. (2001). Testosterone replacement in older men and women. J. Gend. Specif. Med. 4, 49–53.
Morton, R., Gleason, O., and Yates, W. (2000). Psychiatric effects of anabolic steroids after burn injuries. Psychosomatics 41, 66–68. doi: 10.1016/S0033-3182(00)71176-5
Negus, S. S., Pope, H. G. Jr., Kanayama, G., Wines, J. D. Jr., and Fischer, B. D. (2001). Lack of evidence for opioid tolerance or dependence in rhesus monkeys following high-dose anabolic-androgenic steroid administration. Psychoneuroendocrinology 26, 789–796. doi: 10.1016/S0306-4530(01)00028-2
Noakes, T. D. (2004). Tainted glory–doping and athletic performance. N. Engl. J. Med. 351, 847–849. doi: 10.1056/NEJMp048208
Oberlander, J. G., and Henderson, L. P. (2012). Corticotropin-releasing factor modulation of forebrain GABAergic transmission has a pivotal role in the expression of anabolic steroid-induced anxiety in the female mouse. Neuropsychopharmacology 37, 1483–1499. doi: 10.1038/npp.2011.334
Packard, M. G., Cornell, A. H., and Alexander, G. M. (1997). Rewarding affective properties of intra-nucleus accumbens injections of testosterone. Behav. Neurosci. 111, 219–224. doi: 10.1037/0735-7044.111.1.219
Parkinson, A. B., and Evans, N. A. (2006). Anabolic androgenic steroids: a survey of 500 users. Med. Sci. Sports Exerc. 38, 644–651. doi: 10.1249/01.mss.0000210194.56834.5d
Parrilla-Carrero, J., Figueroa, O., Lugo, A., Garcia-Sosa, R., Brito-Vargas, P., Cruz, B., et al. (2009). The anabolic steroids testosterone propionate and nandrolone, but not 17alpha-methyltestosterone, induce conditioned place preference in adult mice. Drug Alcohol Depend. 100, 122–127. doi: 10.1016/j.drugalcdep.2008.09.014
Pärssinen, M., and Seppälä, T. (2002). Steroid use and long-term health risks in former athletes. Sports Med. 32, 83–94. doi: 10.2165/00007256-200232020-00001
Peters, K. D., and Wood, R. I. (2005). Androgen dependence in hamsters: overdose, tolerance, and potential opioidergic mechanisms. Neuroscience 130, 971–981. doi: 10.1016/j.neuroscience.2004.09.063
Philipova, T., Ivanova, T., Pavlova, E., Kasakov, L., and Vlaskovska, M. (2003). Nandrolone modulates the non-opioid and opioid analgesia and tolerance/dependence: role of sexual dimorphism. Arch. Physiol. Biochem. 111, 429–436. doi: 10.3109/13813450312331342283
Polsky, B., Kotler, D., and Steinhart, C. (2001). HIV-associated wasting in the HAART era: guidelines for assessment, diagnosis, and treatment. AIDS Patient Care STDS 15, 411–423. doi: 10.1089/108729101316914412
Pope, H. G. J., and Brower, K. J. (2000). “Anabolic-androgenic steroid abuse,” in Comprehensive Textbook of Psychiatry, eds B. J. Sadock, V. A. Sadock, and P. Ruiz, (Philadelphia, PA: Lippincott Williams & Wilkins), 1085–1095.
Pope, H. G. Jr., Kouri, E. M., and Hudson, J. I. (2000). Effects of supraphysiologic doses of testosterone on mood and aggression in normal men: a randomized controlled trial. Arch. Gen. Psychiatry 57, 133–140. discussion 155–136. doi: 10.1001/archpsyc.57.2.133
Pope, H. G. Jr., Wood, R. I., Rogol, A., Nyberg, F., Bowers, L., and Bhasin, S. (2014). Adverse health consequences of performance-enhancing drugs: an Endocrine Society scientific statement. Endocr. Rev. 35, 341–375. doi: 10.1210/er.2013-1058
Porcerelli, J. H., and Sandler, B. A. (1995). Narcissism and empathy in steroid users. Am. J. Psychiatry 152, 1672–1674. doi: 10.1176/ajp.152.11.1672
Rabkin, J. G., Wagner, G. J., and Rabkin, R. (2000). A double-blind, placebo-controlled trial of testosterone therapy for HIV-positive men with hypogonadal symptoms. Arch. Gen. Psychiatry 57, 141–147. discussion 155–146. doi: 10.1001/archpsyc.57.2.141
Ricci, L. A., Grimes, J. M., and Melloni, R. H. Jr. (2007). Lasting changes in neuronal activation patterns in select forebrain regions of aggressive, adolescent anabolic/androgenic steroid-treated hamsters. Behav. Brain Res. 176, 344–352. doi: 10.1016/j.bbr.2006.10.025
Ricci, L. A., Rasakham, K., Grimes, J. M., and Melloni, R. H. Jr. (2006). Serotonin-1A receptor activity and expression modulate adolescent anabolic/androgenic steroid-induced aggression in hamsters. Pharmacol. Biochem. Behav. 85, 1–11. doi: 10.1016/j.pbb.2006.06.022
Roberto, M., Cruz, M. T., Gilpin, N. W., Sabino, V., Schweitzer, P., Bajo, M., et al. (2010). Corticotropin releasing factor-induced amygdala gamma-aminobutyric Acid release plays a key role in alcohol dependence. Biol. Psychiatry 67, 831–839. doi: 10.1016/j.biopsych.2009.11.007
Roberto, M., Madamba, S. G., Stouffer, D. G., Parsons, L. H., and Siggins, G. R. (2004). Increased GABA release in the central amygdala of ethanol-dependent rats. J. Neurosci. 24, 10159–10166. doi: 10.1523/JNEUROSCI.3004-04.2004
Salas-Ramirez, K. Y., Montalto, P. R., and Sisk, C. L. (2010). Anabolic steroids have long-lasting effects on male social behaviors. Behav. Brain Res. 208, 328–335. doi: 10.1016/j.bbr.2009.11.026
Sato, S. M., Johansen, J. A., Jordan, C. L., and Wood, R. I. (2010). Membrane androgen receptors may mediate androgen reinforcement. Psychoneuroendocrinology 35, 1063–1073. doi: 10.1016/j.psyneuen.2010.01.007
Sato, S. M., Schulz, K. M., Sisk, C. L., and Wood, R. I. (2008). Adolescents and androgens, receptors and rewards. Horm. Behav. 53, 647–658. doi: 10.1016/j.yhbeh.2008.01.010
Schlussman, S. D., Zhou, Y., Johansson, P., Kiuru, A., Ho, A., Nyberg, F., et al. (2000). Effects of the androgenic anabolic steroid, nandrolone decanoate, on adrenocorticotropin hormone, corticosterone and proopiomelanocortin, corticotropin releasing factor (CRF) and CRF receptor1 mRNA levels in the hypothalamus, pituitary and amygdala of the rat. Neurosci. Lett. 284, 190–194. doi: 10.1016/S0304-3940(00)01016-8
Schroeder, J. P., and Packard, M. G. (2000). Role of dopamine receptor subtypes in the acquisition of a testosterone conditioned place preference in rats. Neurosci. Lett. 282, 17–20. doi: 10.1016/S0304-3940(00)00839-9
Schwartzer, J. J., Ricci, L. A., and Melloni, R. H. Jr. (2009). Adolescent anabolic-androgenic steroid exposure alters lateral anterior hypothalamic serotonin-2A receptors in aggressive male hamsters. Behav. Brain Res. 199, 257–262. doi: 10.1016/j.bbr.2008.11.048
Shahidi, N. T. (2001). A review of the chemistry, biological action, and clinical applications of anabolic-androgenic steroids. Clin. Ther. 23, 1355–1390. doi: 10.1016/S0149-2918(01)80114-4
Singh, R., Artaza, J. N., Taylor, W. E., Braga, M., Yuan, X., Gonzalez-Cadavid, N. F., et al. (2006). Testosterone inhibits adipogenic differentiation in 3T3-L1 cells: nuclear translocation of androgen receptor complex with beta-catenin and T-cell factor 4 may bypass canonical Wnt signaling to down-regulate adipogenic transcription factors. Endocrinology 147, 141–154. doi: 10.1210/en.2004-1649
Singh, R., Artaza, J. N., Taylor, W. E., Gonzalez-Cadavid, N. F., and Bhasin, S. (2003). Androgens stimulate myogenic differentiation and inhibit adipogenesis in C3H 10T1/2 pluripotent cells through an androgen receptor-mediated pathway. Endocrinology 144, 5081–5088. doi: 10.1210/en.2003-0741
Sinha-Hikim, I., Artaza, J., Woodhouse, L., Gonzalez-Cadavid, N., Singh, A. B., Lee, M. I., et al. (2002). Testosterone-induced increase in muscle size in healthy young men is associated with muscle fiber hypertrophy. Am. J. Physiol. Endocrinol. Metab. 283, E154–E164. doi: 10.1152/ajpendo.00502.2001
Sinha-Hikim, I., Roth, S. M., Lee, M. I., and Bhasin, S. (2003). Testosterone-induced muscle hypertrophy is associated with an increase in satellite cell number in healthy, young men. Am. J. Physiol. Endocrinol. Metab. 285, E197–E205. doi: 10.1152/ajpendo.00370.2002
Spanagel, R., Herz, A., and Shippenberg, T. S. (1992). Opposing tonically active endogenous opioid systems modulate the mesolimbic dopaminergic pathway. Proc. Natl. Acad. Sci. U.S.A. 89, 2046–2050. doi: 10.1073/pnas.89.6.2046
Steiner, H., and Gerfen, C. R. (1998). Role of dynorphin and enkephalin in the regulation of striatal output pathways and behavior. Exp. Brain Res. 123, 60–76. doi: 10.1007/s002210050545
Tamaki, T., Shiraishi, T., Takeda, H., Matsumiya, T., Roy, R. R., and Edgerton, V. R. (2003). Nandrolone decanoate enhances hypothalamic biogenic amines in rats. Med. Sci. Sports Exerc. 35, 32–38. doi: 10.1097/00005768-200301000-00006
Tanaka, K., Sakai, H., Hashizume, M., and Hirohata, T. (2000). Serum testosterone:estradiol ratio and the development of hepatocellular carcinoma among male cirrhotic patients. Cancer Res. 60, 5106–5110.
Tennant, F., Black, D. L., and Voy, R. O. (1988). Anabolic steroid dependence with opioid-type features. N. Engl. J. Med. 319, 578. doi: 10.1056/NEJM198809013190910
Thevis, M., Sauer, M., Geyer, H., Sigmund, G., Mareck, U., and Schänzer, W. (2008). Determination of the prevalence of anabolic steroids, stimulants, and selected drugs subject to doping controls among elite sport students using analytical chemistry. J. Sports Sci. 26, 1059–1065. doi: 10.1080/02640410801910293
Thiblin, I., Finn, A., Ross, S. B., and Stenfors, C. (1999a). Increased dopaminergic and 5-hydroxytryptaminergic activities in male rat brain following long-term treatment with anabolic androgenic steroids. Br. J. Pharmacol. 126, 1301–1306. doi: 10.1038/sj.bjp.0702412
Thiblin, I., Lindquist, O., and Rajs, J. (2000). Cause and manner of death among users of anabolic androgenic steroids. J. Forensic Sci. 45, 16–23.
Thiblin, I., Runeson, B., and Rajs, J. (1999b). Anabolic androgenic steroids and suicide. Ann. Clin. Psychiatry 11, 223–231. doi: 10.3109/10401239909147074
Tremblay, M. S., Copeland, J. L., and Van Helder, W. (2004). Effect of training status and exercise mode on endogenous steroid hormones in men. J. Appl. Physiol. (1985) 96, 531–539. doi: 10.1152/japplphysiol.00656.2003
Triemstra, J. L., Sato, S. M., and Wood, R. I. (2008). Testosterone and nucleus accumbens dopamine in the male Syrian hamster. Psychoneuroendocrinology 33, 386–394. doi: 10.1016/j.psyneuen.2007.12.006
Tucci, P., Morgese, M. G., Colaianna, M., Zotti, M., Schiavone, S., Cuomo, V., et al. (2012). Neurochemical consequence of steroid abuse: stanozolol-induced monoaminergic changes. Steroids 77, 269–275. doi: 10.1016/j.steroids.2011.12.014
van der Vegt, B. J., Lieuwes, N., van de Wall, E. H., Kato, K., Moya-Albiol, L., Martínez-Sanchis, S., et al. (2003). Activation of serotonergic neurotransmission during the performance of aggressive behavior in rats. Behav. Neurosci. 117, 667–674. doi: 10.1037/0735-7044.117.4.667
Volkow, N. D., Chang, L., Wang, G. J., Fowler, J. S., Ding, Y. S., Sedler, M., et al. (2001a). Low level of brain dopamine D2 receptors in methamphetamine abusers: association with metabolism in the orbitofrontal cortex. Am. J. Psychiatry 158, 2015–2021. doi: 10.1176/appi.ajp.158.12.2015
Volkow, N. D., Chang, L., Wang, G. J., Fowler, J. S., Franceschi, D., Sedler, M., et al. (2001b). Loss of dopamine transporters in methamphetamine abusers recovers with protracted abstinence. J. Neurosci. 21, 9414–9418.
Volkow, N. D., Fowler, J. S., Wolf, A. P., Schlyer, D., Shiue, C. Y., Alpert, R., et al. (1990). Effects of chronic cocaine abuse on postsynaptic dopamine receptors. Am. J. Psychiatry 147, 719–724. doi: 10.1176/ajp.147.6.719
Wichstrom, L., and Pedersen, W. (2001). Use of anabolic-androgenic steroids in adolescence: winning, looking good or being bad? J. Stud. Alcohol 62, 5–13. doi: 10.15288/jsa.2001.62.5
Wines, J. D. Jr., Gruber, A. J., Pope, H. G. Jr., and Lukas, S. E. (1999). Nalbuphine hydrochloride dependence in anabolic steroid users. Am. J. Addict. 8, 161–164. doi: 10.1080/105504999305965
Wood, R. I. (2002). Oral testosterone self-administration in male hamsters: dose-response, voluntary exercise, and individual differences. Horm. Behav. 41, 247–258. doi: 10.1006/hbeh.2002.1769
Wood, R. I. (2004). Reinforcing aspects of androgens. Physiol. Behav. 83, 279–289. doi: 10.1016/j.physbeh.2004.08.012
Wood, R. I. (2008). Anabolic-androgenic steroid dependence? Insights from animals and humans. Front. Neuroendocrinol. 29, 490–506. doi: 10.1016/j.yfrne.2007.12.002
Wood, R. I., Johnson, L. R., Chu, L., Schad, C., and Self, D. W. (2004). Testosterone reinforcement: intravenous and intracerebroventricular self-administration in male rats and hamsters. Psychopharmacology (Berl) 171, 298–305. doi: 10.1007/s00213-003-1587-7
Yates, W. (2000). Testosterone in psychiatry. Arch. Gen. Psychiatry 57, 155–156. doi: 10.1001/archpsyc.57.2.155
Yates, W. R., Perry, P. J., and Andersen, K. H. (1990). Illicit anabolic steroid use: a controlled personality study. Acta Psychiatry Scand. 81, 548–550. doi: 10.1111/j.1600-0447.1990.tb05496.x
Yesalis, C. E., Bahrke, M. S., Kopstein, A. N., and Barsukiewicz, C. K. (2000). “Incidence of anabolic steroids use: a discussion of methodological issues,” in Anabolic Steroids in Sport and Exercise, 2nd Edn., ed C. E. Yesalis (Champaign, IL: Human Kinetics), 73–115.
Keywords: anabolic androgenic steroid, reward, dopamine, serotonin, psychosis spectrum disorders, depression
Citation: Mhillaj E, Morgese MG, Tucci P, Bove M, Schiavone S and Trabace L (2015) Effects of anabolic-androgens on brain reward function. Front. Neurosci. 9:295. doi: 10.3389/fnins.2015.00295
Received: 28 May 2015; Accepted: 06 August 2015;
Published: 26 August 2015.
Edited by:
George Panagis, University of Crete, GreeceReviewed by:
Giovanni Laviola, Istituto Superiore di Sanità, ItalyLynn G. Kirby, University of Pennsylvania, USA
S. Stevens Negus, Virginia Commonwealth University, USA
Copyright © 2015 Mhillaj, Morgese, Tucci, Bove, Schiavone and Trabace. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luigia Trabace, Department of Clinical and Experimental Medicine, Faculty of Medicine, University of Foggia, Viale L. Pinto, 71121 Foggia, Italy,bHVpZ2lhLnRyYWJhY2VAdW5pZmcuaXQ=