 Jingqi Zhang
Jingqi Zhang Tao Guo
Tao Guo Yaxin Chen1
Yaxin Chen1 Lijiao Wu
Lijiao Wu Hui Xie
Hui Xie- 1Hospital of Chengdu University of Traditional Chinese Medicine, Chengdu, China
- 2School of Sports Medicine and Health, Chengdu Sport University, Chengdu, China
Background: Hearing loss affects over 10% of the global population. Inflammation is a key factor in hearing loss caused by noise, infection, and aging, damaging various hearing-related tissues (e.g., spiral ligament, stria vascularis). Mendelian randomization (MR) can help identify potential causal relationships and therapeutic targets.
Methods: We conducted MR analyses on 91 inflammatory proteins (n = 14,824) and genome-wide association study results for various hearing loss types in European ancestry populations, including sensorineural hearing loss (SNHL; ncases = 15,952, ncontrols = 196,592), sudden idiopathic hearing loss (SIHL; ncases = 1,491, ncontrols = 196,592), and other hearing loss (OHL; ncases = 4,157, ncontrols = 196,592). Additionally, hearing loss with difficulty in hearing (ncases = 14,654, ncontrols = 474,839) served as a validation set. To predict inflammatory protein-enriched pathways and tissues, we performed enrichment analysis, functional annotation, and tissue analyses using “OmicsNet2.0” and “FUMA” platforms. We also combined “CoreMine” and molecular docking to explore potential drugs targeting inflammatory proteins and investigate binding efficacy.
Results: CCL19 was identified as a common risk factor for SNHL and OHL, which was validated in the hearing loss with difficulty in hearing dataset. Tissue analysis revealed that SIHL-related inflammatory proteins were enriched in the amygdala. Multi-omics research indicated associations between inflammatory proteins and neurodegenerative diseases. Molecular docking studies suggested that Chuanxiong Rhizoma and Uncariae Ramulus Cumuncis are potential drugs for targeting CCL19.
Conclusion: This study identified CCL19 as a common risk factor for various types of hearing loss through MR analysis, highlighting the crucial role of inflammatory proteins in hearing loss. The enrichment of related inflammatory proteins in the amygdala and their association with neurodegenerative diseases provide new insights into the mechanisms of hearing loss.
1 Introduction
According to 2015 statistics, hearing loss affects over 10% of the world’s population, making it the second most common disability after anemia (1). The increasing noise pollution and the use of ototoxic drugs (aminoglycoside antibiotics, loop diuretics, anti-tumor drugs, etc.) (2) have also exacerbated hearing loss, not only affecting health and quality of life but also causing a significant economic burden.
Multiple factors can lead to hearing loss, including genetic factors (3), noise pollution (4), infections (5), aging (6), and ototoxic drugs (7, 8). Inflammation plays a crucial role in these processes. For instance, the expression of inflammatory factor IL-6 has been observed in the spiral ligament, stria vascularis, and spiral ganglion neurons following noise exposure (9). Infection-induced inflammatory states can damage inner ear hair cells and spiral ganglion neurons (10). Age-related hearing loss (ARHL) is associated with low-grade inflammation (11), while ototoxic drugs like cisplatin increase reactive oxygen species (ROS) levels and inflammatory factor expression, leading to hearing loss (12). Existing studies have explored the relationship between certain inflammatory factors and hearing loss (13), but have not clearly established causal links and are limited by factors such as low protein coverage, small cohort sizes, and potential confounding biases.
Recent advances in genomics have provided powerful tools for investigating the genetic basis of complex diseases like hearing loss. Mendelian randomization (MR) uses genetic variants as instrumental variables to infer causal relationships between exposures and outcomes (14), helping to overcome limitations of observational studies by reducing confounding and reverse causation (15). Previous studies have employed MR to explore the causal relationships between white blood cell count and sudden sensorineural hearing loss (16), serum lipids and ARHL loss (17), and mitochondrial proteins and inflammatory diet with sensorineural hearing loss (SNHL) (18, 19).
Our study utilized MR to investigate the relationship between 91 inflammatory proteins and various types of hearing loss. Additionally, multi-omics analysis and tissue analysis were integrated to explore the mechanisms by which inflammatory proteins cause hearing loss and identify enriched tissues. Finally, potential drug discovery and molecular docking were combined to identify novel drugs targeting inflammatory proteins for the treatment of hearing loss.
2 Methods
The study followed the STROBE-MR checklist for standardized MR analysis (20) (Supplementary Table S1). The overall study design is presented in Figure 1. The exposure data was derived from 91 inflammatory proteins (14,824 participants) provided by Zhao et al. (21). The outcome data for three types of hearing loss were obtained from the FinnGen database.1 SNHL (ncase = 15,952, ncontrol = 196,592) was defined as hearing loss due to the inner ear or sensory organ (cochlea and associated structures) or vestibulocochlear nerve, and patients were diagnosed using ICD-10 codes H90.3, H90.4, and H90.5 (22). Sudden idiopathic hearing loss (SIHL) (ncase = 1,491, ncontrol = 196,592) was diagnosed using ICD-10 code H91.2. Other hearing loss (OHL) (ncase = 4,157, ncontrol = 196,592) includes conditions such as deaf mutism, other specified/unspecified hearing loss, ototoxic hearing loss, presbycusis, etc. All data pertain to European populations. The validation data was sourced from the Biobank Japan database for hearing loss, difficulty in hearing (ncase = 14,654, ncontrol = 474,839) (23). Detailed information of the data can be found in Supplementary Table S2.
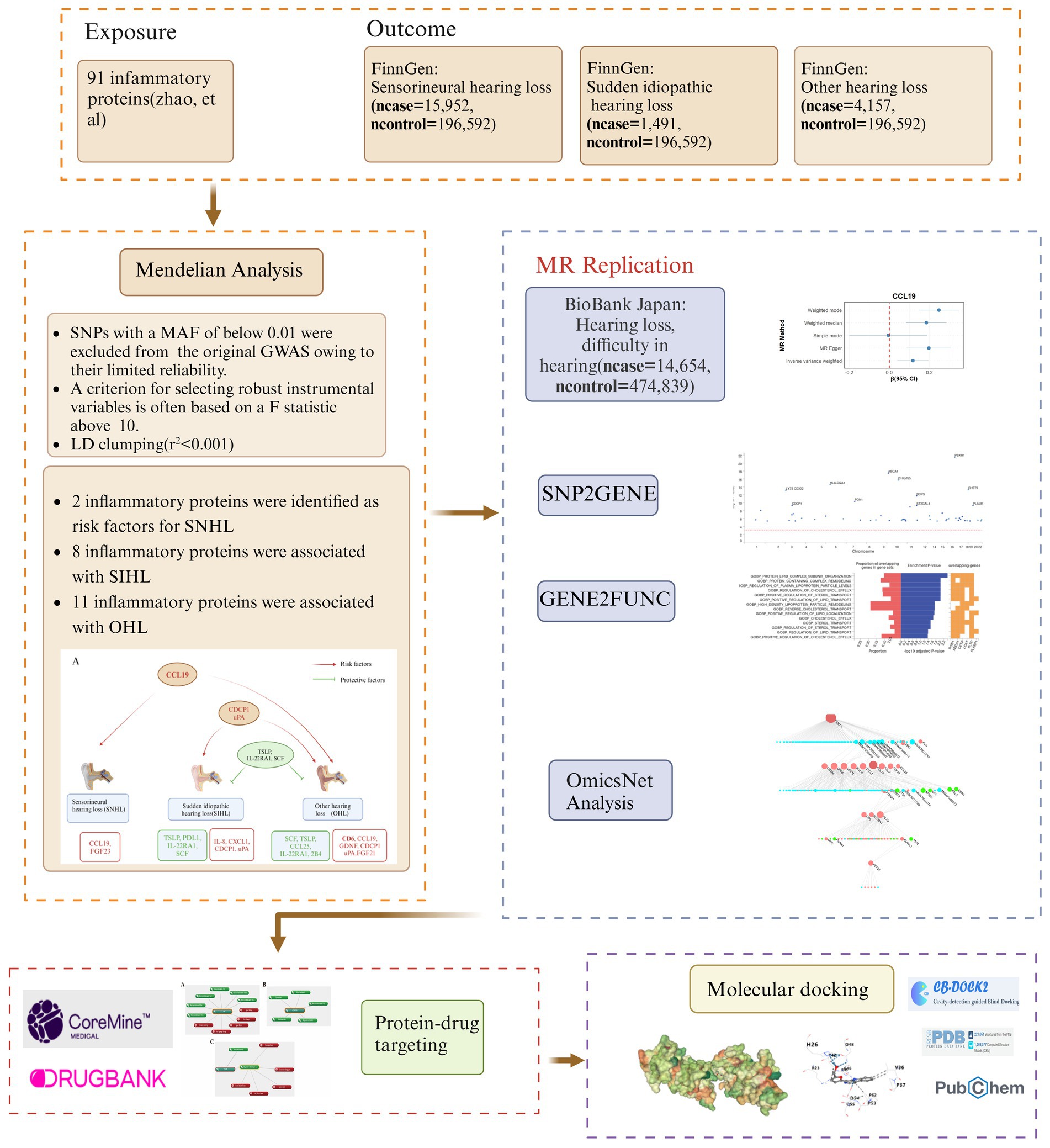
Figure 1. Research flow diagram. MR, Mendelian randomization; SNHL, sensorineural hearing loss; SIHL, sudden idiopathic hearing loss; OHL, other hearing loss.
Subsequently, we utilized the functional mapping and annotation of genome-wide association studies (FUMA) platform, specifically its “SNP2GENE” and “GENE2FUNC” tools, to perform functional annotation, gene-based association, and tissue analyses on SIHL-related proteins. Through multi-omics analysis, we identified miRNAs and transcription factors involved in the crosstalk between inflammatory proteins, as well as explored potential signaling pathways. To discover effective therapeutic agents, we employed “CoreMine” to mine for drugs targeting key proteins. Additionally, we used molecular docking to evaluate the binding capacity of small molecule drugs to these critical proteins.
2.1 Mendelian randomization analysis
This study followed the core assumptions of MR analysis (Supplementary Figure S1). The following criteria were used to select genetic variants associated with inflammatory proteins: (i) we selected genetic variants strongly associated with inflammatory proteins (p < 5 × 10−6) as instrumental variables and excluded any variants that were directly associated with SNHL, SIHL, OHL, or difficulty in hearing in genome-wide association studies; (ii) to ensure independence between single-nucleotide polymorphisms (SNPs), linkage disequilibrium (LD) clustering was performed, with a 10,000 kb window and r2 < 0.001 as the filtering standard (24); (iii) SNPs with a minor allele frequency (MAF) below 0.01 were excluded due to their limited reliability (25); and (iv) a criterion for selecting robust instrumental variables is often based on an F-statistic above 10, calculated as: R2 = 2 × EAF × (1 − EAF) × beta2; F = R2 × (N − 2)/(1 − R2) (26).
The “TwoSampleMR” software package was used to perform MR analysis. Multiple methods were used to evaluate the causal effects of genetic variants on hearing loss, including simple mode, weighted mode, weighted median, and MR-Egger, as well as inverse variance weighted (IVW) method, which is considered the primary method (27). The Q statistic was used to evaluate the heterogeneity of genetic instruments (28). The MR-Egger intercept was used to test for directional pleiotropy, and results with evidence of pleiotropy were excluded. False discovery rate (FDR) correction was used to reduce the likelihood of false positive discoveries, and results with FDR <0.05 were considered significant. All analyses were performed in R software (version 4.2.3).
2.2 Functional annotation, gene-based association and tissue analyses
We utilized FUMA platform to prioritize the most likely causal SNPs and genes, and to provide gene-based, pathway, and tissue enrichment results (29). FUMA integrates information from 18 biological data repositories and tools. We employed FUMA’s “SNP2GENE” tool to further analyze previous results and screen for priority genes. The “GENE2FUN” tool was used for functional enrichment and disease pathway analyses of the identified genes. Additionally, we conducted tissue expression analysis using MAGMA within “SNP2GENE.” MAGMA tests the relationship between tissue-specific gene expression and disease gene associations through gene property analysis.
The “SNP2GENE” analysis parameters were set as follows: the maximum p-value threshold for lead SNPs was 5 × 10−8, with a maximum p-value cutoff of 0.05. To define independent significant SNPs, the r2 threshold was set to 0.6; a second r2 threshold of 0.1 was used to define lead SNPs (29–31). The 1,000 genomes phase 3 European population was selected as the reference panel, matching the study population’s background. We opted to include non-GWAS tagged SNPs from the reference panel for LD analysis, enhancing the precision of LD assessment. The MAF was set to 0, allowing consideration of variants at all frequencies, and the maximum distance between LD blocks to be merged into a locus was set to 250 kb (32). These parameter settings allowed for a comprehensive and stringent functional annotation and mapping of the GWAS results. The maximum p-value threshold ensured focus on the most significant association signals, while the r2 thresholds helped identify independent significant SNPs and lead SNPs. The 250 kb LD block merging distance aided in defining potentially functionally related genomic regions.
2.3 Multi-omics network analysis
Multi-omics analysis was performed using “OmicsNet2.0”2 (33). The input data consisted of 17 proteins associated with SNHL, SIHL, and OHL, as identified through MR analysis. The corresponding Gene IDs for these proteins were obtained from the National Center for Biotechnology Information (NCBI).3
The “protein–protein” interactions were established based on the “InnateDB” database, while the “transcription factor-target gene” relationships were determined using the “TRRUST” database. The “miRNA-mRNA” interactions were derived from the “miRTarBase” database (34). By integrating these diverse data sources, a comprehensive multi-omics network was constructed, with the resulting visualization output in the form of a tree diagram.
This multi-omics approach allowed for the exploration of the complex relationships between proteins, transcription factors, miRNAs, and their target genes, providing a systems-level understanding of the molecular mechanisms underlying hearing loss. The integration of multiple omics data types enabled the identification of key regulatory pathways and potential therapeutic targets, offering valuable insights into the pathogenesis of SNHL, SIHL, and OHL.
2.4 Drug repurposing opportunities
To explore the potential of the core protein identified through multiple screening steps as a drug target and to investigate drug repurposing opportunities, the “CoreMine” database4 (35) was utilized. “CoreMine” was searched for potential drug targets (including both Western and Chinese medicines) related to the core protein, aiming to assess the feasibility of targeting this protein for therapeutic purposes.
The drugs identified through the “CoreMine” search were further validated using the “DrugBank” database5 to ensure their accuracy and to obtain information on their mechanisms of action and disease associations. This validation step helped to confirm the relevance of the identified drugs to the core protein and their potential for repurposing in the context of hearing loss treatment.
By leveraging the information from both “CoreMine” and “DrugBank,” a comprehensive understanding of the drug targeting potential of the core protein was obtained. This approach not only considered the direct targeting of the core protein but also explored the targeting of its associated pathways and regulatory mechanisms, as identified through the multi-omics network analysis.
2.5 Molecular docking analysis
Following the identification of traditional Chinese medicines (TCMs) targeting the inflammatory proteins through “CoreMine,” molecular docking simulations were conducted between the main active components of these TCMs and inflammatory proteins. The effective components of TCMs were retrieved from the TCMSP database, selecting those that met the criteria of oral bioavailability (OB ≥30%) and drug-likeness (DL ≥0.18) (36).
The small molecule structures of the qualifying TCM components were obtained from the PubChem database.6 The UniProt ID for inflammatory proteins was acquired from the UniProt database,7 and its structural data was downloaded from the RCSB Protein Data Bank (PDB, https://www.rcsb.org/).
Prior to molecular docking, energy minimization was performed on the small molecules using Chem3D software (version 22.0) to achieve more stable conformations. The protein structure underwent preprocessing in CB-Dock, which included removing water molecules, adding hydrogen atoms, and repairing missing residues. Molecular docking simulations were carried out using the CB-Dock2 online tool8 (37).
This comprehensive approach allowed for a systematic evaluation of potential interactions between TCM components and inflammatory proteins, providing insights into possible therapeutic mechanisms and drug repurposing opportunities for hearing loss treatment.
3 Results
3.1 Mendelian randomization analysis results
In the MR analysis, the IVW method was used as the standard to identify 17 inflammatory proteins associated with hearing loss. Among them, SNHL and OHL shared a common risk factor (CCL19), while SIHL and OHL had common risk factors (CDCP1, uPA) and common protective factors (TSLP, IL-22RA1, SCF) (Figure 2A). Two inflammatory proteins were identified to be associated with SNHL, both of which were risk factors for SNHL: CCL19 (β = 0.21, 95% CI = 0.05–0.36, p = 0.01), FGF23 (β = 0.20, 95% CI = 0.02–0.39, p = 0.03) (Figure 2B) (Supplementary Table S3A). Eight inflammatory proteins were found to be related to SIHL, with four being protective factors for SIHL: TSLP (β = −0.27, 95% CI = −0.48 to −0.05, p = 0.015), PDL1 (β = −0.22, 95% CI = −0.42 to −0.03, p = 0.024), IL-22RA1 (β = −0.30, 95% CI = −0.56 to −0.03, p = 0.028), SCF (β = −0.13, 95% CI = −0.25 to −0.01, p = 0.035). The other four were risk factors for SIHL: IL-8 (β = 0.28, 95% CI = 0.07–0.48, p = 0.008), CXCL1 (β = 0.18, 95% CI = 0.02–0.35, p = 0.031), CDCP1 (β = 0.14, 95% CI = 0.01–0.27, p = 0.033), uPA (β = 0.18, 95% CI = 0.01–0.35, p = 0.043) (Figure 2C) (Supplementary Table S4A).
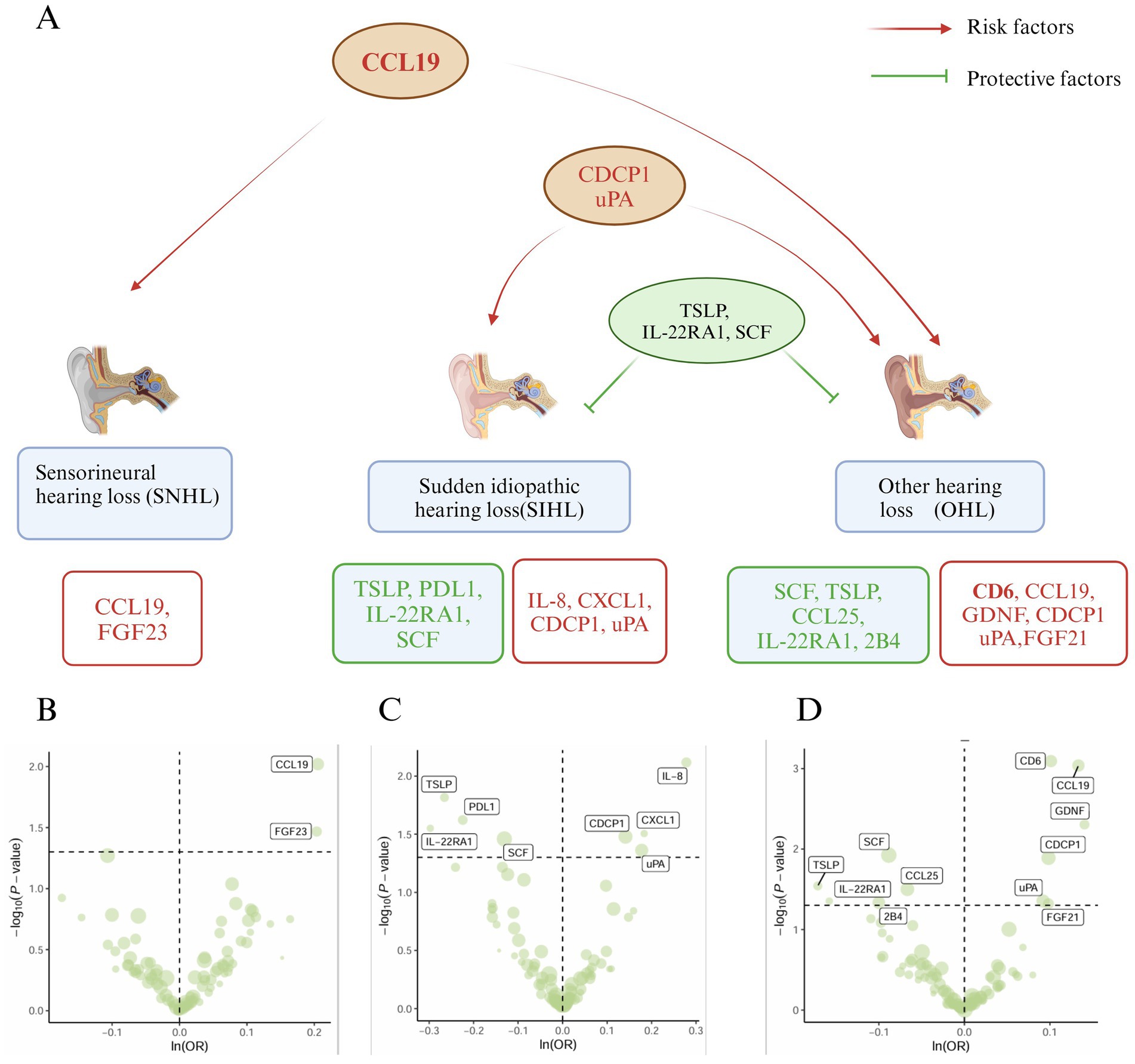
Figure 2. Mendelian analysis results of inflammatory proteins and hearing loss (SNHL, SIHL, OHL). (A) Relationship plot of inflammatory proteins and hearing loss (SNHL, SIHL, OHL). (B) Volcano plot of risk proteins (CCL19, FGF23) associated with SNHL. (C) Volcano plot of protective proteins (TSLP, PDL1, IL-22RA1, SCF) and risk proteins (IL-8, CXCL1, CDCP1, uPA) associated with SIHL. (D) Volcano plot of protective proteins (SCF, TSLP, CCL25, IL-22RA1, 2B4) and risk proteins (CD6, CCL19, GDNF, CDCP1, uPA, FGF21) associated with OHL.
Eleven inflammatory proteins were associated with OHL, with five being protective factors: SCF (β = −0.09, 95% CI = −0.16 to −0.01, p = 0.01), TSLP (β = −0.17, 95% CI = −0.33 to −0.02, p = 0.029), CCL25 (β = −0.07, 95% CI = −0.13 to −0.01, p = 0.031), IL-22RA1 (β = −0.16, 95% CI = −0.31 to −0.004, p = 0.044), 2B4 (β = −0.10, 95% CI = −0.20 to −0.002, p = 0.046). Six proteins were identified as risk factors for OHL: CD6 (β = 0.10, 95% CI = 0.04–0.16, p = 0.0008, pFDR = 0.04), CCL19 (β = 0.13, 95% CI = 0.05–0.21, p = 0.0009, pFDR = 0.04), GDNF (β = 0.14, 95% CI = 0.04–0.24, p = 0.005), CDCP1 (β = 0.10, 95% CI = 0.02–0.18, p = 0.01), uPA (β = 0.09, 95% CI = 0.002–0.18, p = 0.044), FGF21 (β = 0.10, 95% CI = 0.001–0.196, p = 0.047) (Figure 2D). Among these, CD6 and CCL19 exhibited higher significance (pFDR < 0.05) (Supplementary Table S5A). The F-statistics for the MR studies of SNHL, SIHL, and OHL were greater than 10 (Supplementary Tables S3B–S5B). The simple mode, weighted mode, weighted median, and MR-Egger methods generally supported the results of the IVW method (Supplementary Figures S2–S4), and the p-value of the intercept term in MR-Egger regression >0.05, no horizontal pleiotropy was observed (Supplementary Tables S3C–S5C). The heterogeneity analysis is provided in Supplementary Tables S3D–S5D, and the leave-one-out analysis results (Supplementary Figures S5, S6) indicated the robustness of our study.
Six inflammatory proteins (CCL19, CDCP1, uPA, TSLP, IL-22RA1, and SCF) associated with two or more types of hearing loss were further validated in the validation set “hearing loss, difficulty in hearing” (ebi-a-GCST90018857) (Supplementary Table S6A), and the results are presented in Figure 3. The weighted mode, weighted median, MR Egger, and IVW methods all supported the promoting effect of CCL19 on hearing loss (p < 0.05) (Figure 3A). It is worth noting that CDCP1 (pIVW = 0.069) (Figure 3B) was not associated with hearing loss, and no associations were found between the remaining inflammatory proteins (uPA, TSLP, IL-22RA1, and SCF) and hearing loss in the validation set (Figure 3C). The F-statistics were greater than 10 (Supplementary Table S6B), and no horizontal pleiotropy was observed (Supplementary Table S6C).
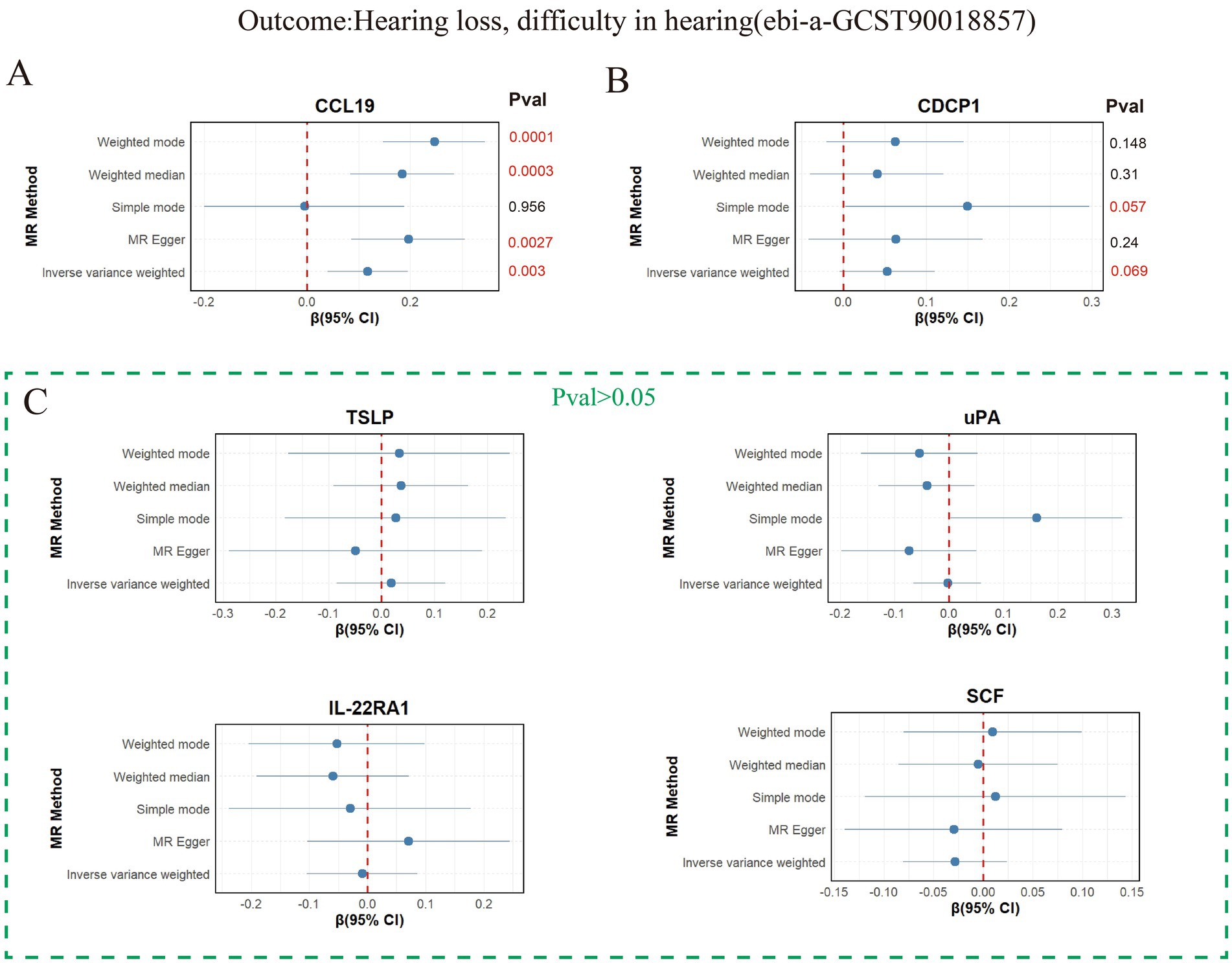
Figure 3. Mendelian analysis results of 6 inflammatory proteins and hearing loss, difficulty in hearing. (A) Mendelian analysis results of CCL19 and hearing loss, difficulty in hearing using five methods (weighted mode, weighted median, MR Egger, IVW, and simple mode). (B) Mendelian analysis results of CDCP119 and hearing loss, difficulty in hearing. (C) Mendelian analysis results of (uPA, TSLP, IL-22RA1, SCF) and hearing loss, difficulty in hearing.
3.2 Functional annotation, gene-based association and tissue analyses results
After gene annotation of SNPs related to SIHL-associated proteins, we identified 62 genes, including CDCP1 (Figure 4A). Detailed gene information is provided in Supplementary Table S7, and mapped gene-related information is available in Supplementary Table S8. Further “GENE2FUNC” enrichment analysis revealed that the biological processes are primarily associated with lipid metabolism, including protein-lipid complex subunit organization, protein complex remodeling, regulation of plasma lipoprotein particle levels, and positive regulation of lipid transport (Figure 4B). Lipid metabolism plays a crucial role in auditory function. A study showed that the loss of Bcl2 in the auditory cortex affects lipid metabolism, leading to decreased synaptic function and neurodegeneration (38). This finding aligns with our enrichment analysis results, further supporting the potential role of lipid metabolism in SIHL development.
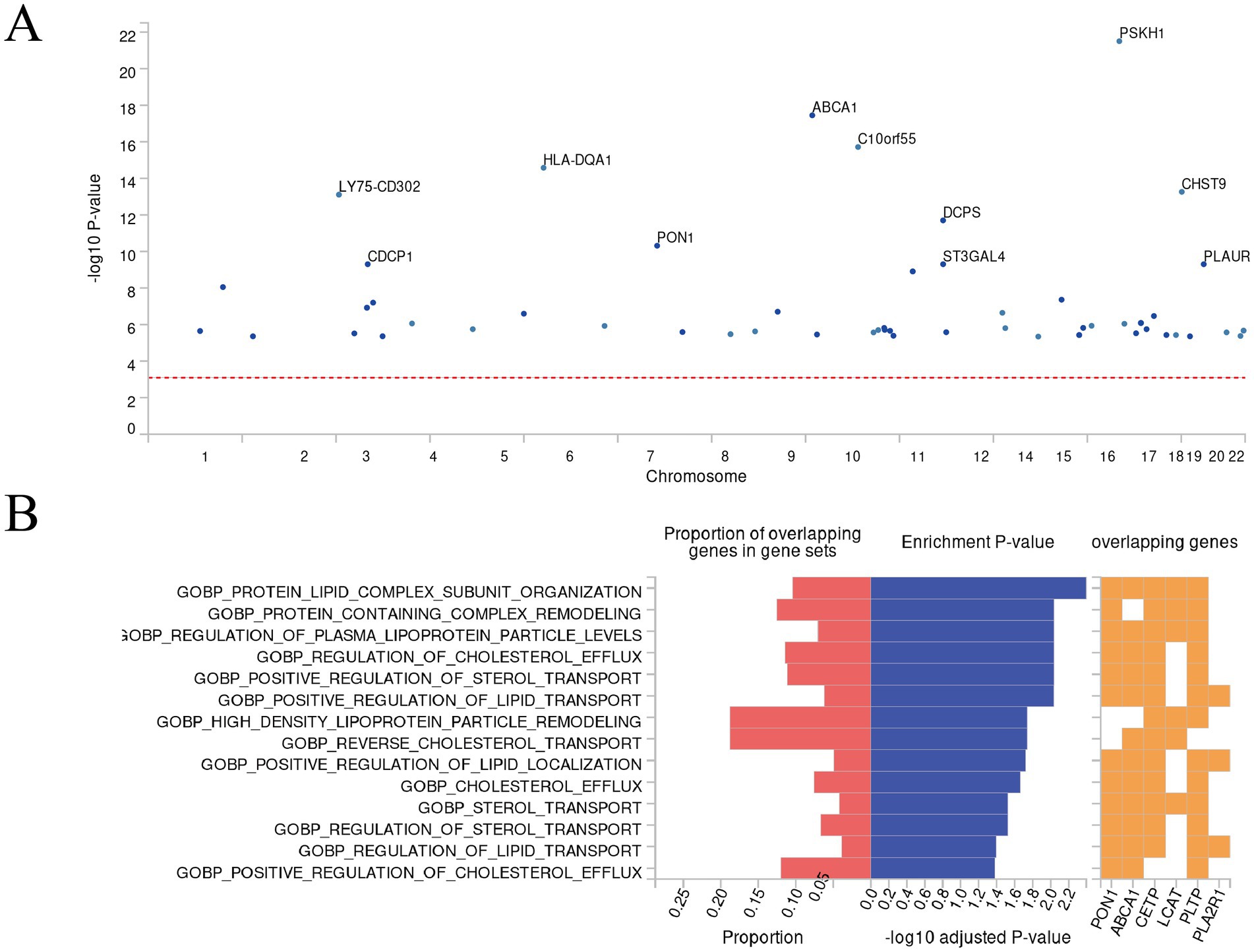
Figure 4. Results of FUMA’s “SNP2GENE” and “GENE2FUNC” analysis. (A) Manhattan plot. This Manhattan plot displays the gene-based test results computed by MAGMA. Input SNPs were mapped to 62 protein-coding genes, with the top 11 genes highlighted in the plot. The genome-wide significance threshold (red dashed line) was set at p = 0.05/62 = 8.065 ×10−4. (B) Biological processes enrichment plot. This functional enrichment plot, generated by FUMA’s GENE2FUNC analysis, illustrates the enrichment of GO biological processes (MsigDB) for a set of genes. The plot presents the proportion of overlapping genes in gene sets, enrichment p-values, and the specific overlapping genes. The left x-axis shows the proportion of overlapping genes in gene sets, while the right x-axis displays the −log10 adjusted p-value. The y-axis lists the GO biological processes. Red bars represent the proportion of overlapping genes in gene sets, and blue bars indicate the −log10 adjusted p-value. The orange grid highlights the overlapping genes for each process.
GWAS catalog reported genes enrichment analysis showed that the identified genes (Supplementary Figure S7) are enriched in gene sets related to schizophrenia, serum lipid metabolism (including triglycerides), and C-reactive protein (Supplementary Table S9). Previous research has suggested that hearing impairment is a risk factor for mental health-related disorders, potentially due to factors such as loneliness and reduced theory of mind (39). Additionally, a cross-sectional study found that lipid and C-reactive protein levels are risk factors for hearing loss in older adults, with high dietary cholesterol intake increasing the risk of hearing loss (40).
Tissue enrichment analysis results showed that these genes are primarily expressed in the brain amygdala, cervical spinal cord, and spleen (Supplementary Figure S8). The amygdala, as a crucial region for emotional processing and auditory information integration, warrants further investigation in its association with SIHL. The enrichment in the spleen may reflect the potential role of the immune system in SIHL development. These findings provide new perspectives for understanding the molecular mechanisms of SIHL and point towards directions for future research and potential therapeutic strategy development.
3.3 Multi-omics network analysis results
The multi-omics dendrogram (Figure 5) presents a “protein-miRNA-transcription factor” network after screening by MR analysis. CDCP1 protein, located at the top of the dendrogram, is a key node in this network and is associated with various downstream proteins. CCL19, TSLP, and other proteins occupy intermediary positions in the network.
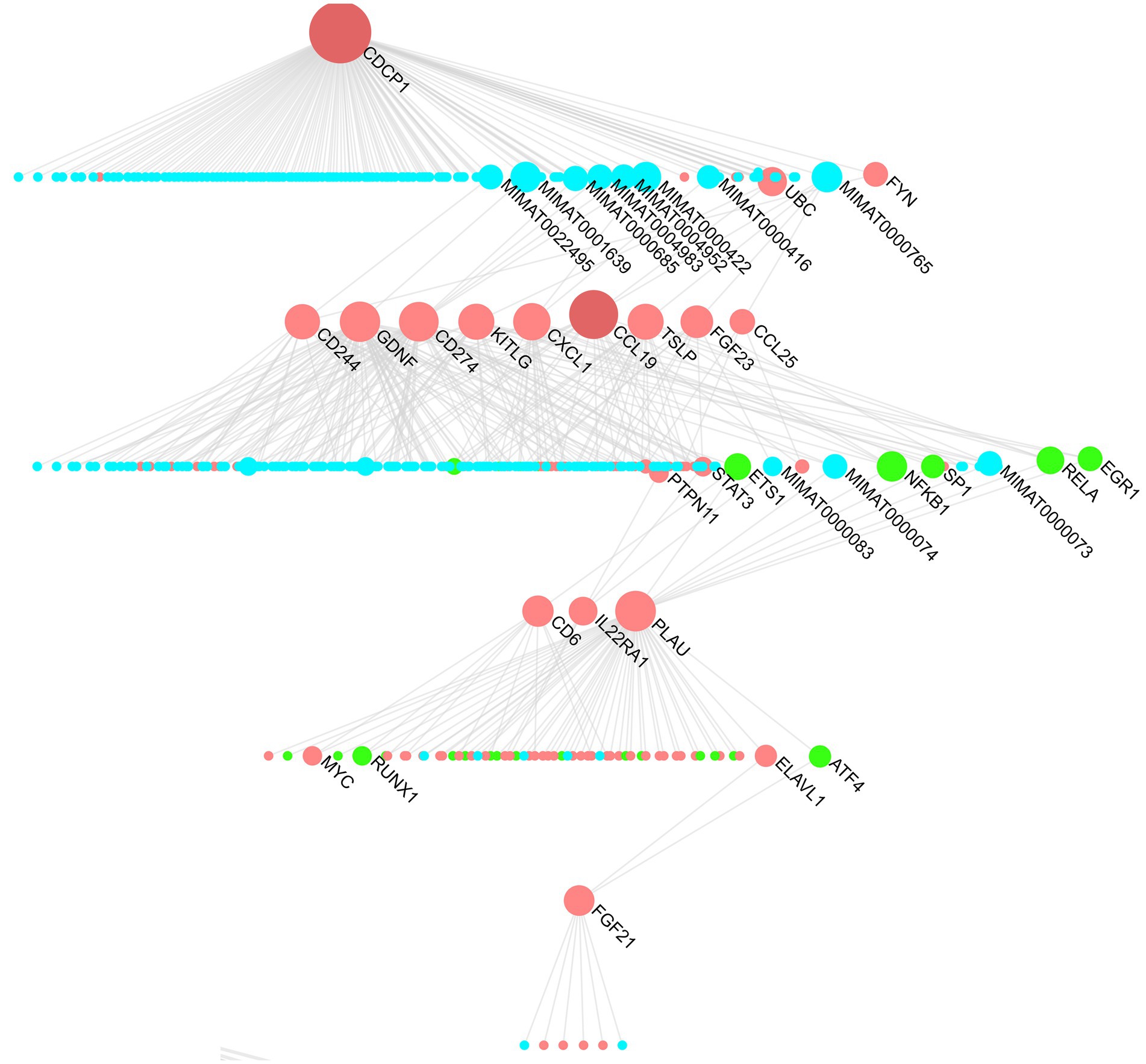
Figure 5. “Protein-miRNA-transcription factor” hierarchy for hearing-related proteins. Blue represents miRNAs, red represents proteins, and green represents transcription factors.
The “protein-miRNA-transcription factor” network reveals the key regulatory miRNA factors (MIMAT0022495, MIMAT0001639, etc.) that link CDCP1 protein with downstream proteins (CCL19, TSLP, CCL25, FGF23, etc.). These shared miRNA regulatory factors may mediate common regulatory mechanisms that play crucial roles in diseases. For instance, MIMAT0000422 (hsa-miR-124-3p) and MIMAT0000416 (hsa-miR-1-3p) have been implicated in neurodegenerative diseases. Expression of hsa-miR-124-3p is downregulated in Alzheimer’s disease (41), while both hsa-miR-124-3p and hsa-miR-1-3p are downregulated in cerebellar neurodegeneration (42). These neurodegenerative changes may lead to hearing loss (43, 44).
In addition to identifying key miRNAs, the “protein-miRNA-transcription factor” network also identified critical proteins and transcription factors such as ubiquitin-conjugating enzyme E2 C (UBC), early growth response 1 (EGR1), signal transducer and activator of transcription 3 (STAT3), nuclear factor kappa-B (NF-κB), and the MYC gene. Studies have shown that the STAT3 pathway is associated with various ototoxic hearing losses, while the NF-κB pathway promotes ARHL. Caffeine can improve this process by downregulating the NF-κB inflammatory pathway (45).
3.4 Drug repurposing prediction results
To identify potential drug repurposing opportunities, we performed drug target analysis on the core proteins CCL19, CDCP1, and TSLP. The “CoreMine” database revealed six drugs targeting CCL19, including recombinant human 6Ckine, recombinant beta chemokine, recombinant interleukin-7, interleukin-12, recombinant tumor necrosis factor-beta, and recombinant human macrophage inflammatory protein-1 beta. Two of these drugs were found to be in clinical trials according to the DrugBank database. Interleukin-7 has been used in trials investigating the treatment of metastatic breast cancer (46). Additionally, cetuximab and recombinant interleukin-12 are being used in a trial treating patients with recurrent, metastatic, or unresectable squamous cell carcinoma of the head and neck (detailed information available in DrugBank). Traditional Chinese medicines targeting CCL19 include Chuanxiong Rhizoma, Uncariae Ramulus Cumuncis, etc. (Figure 6A).
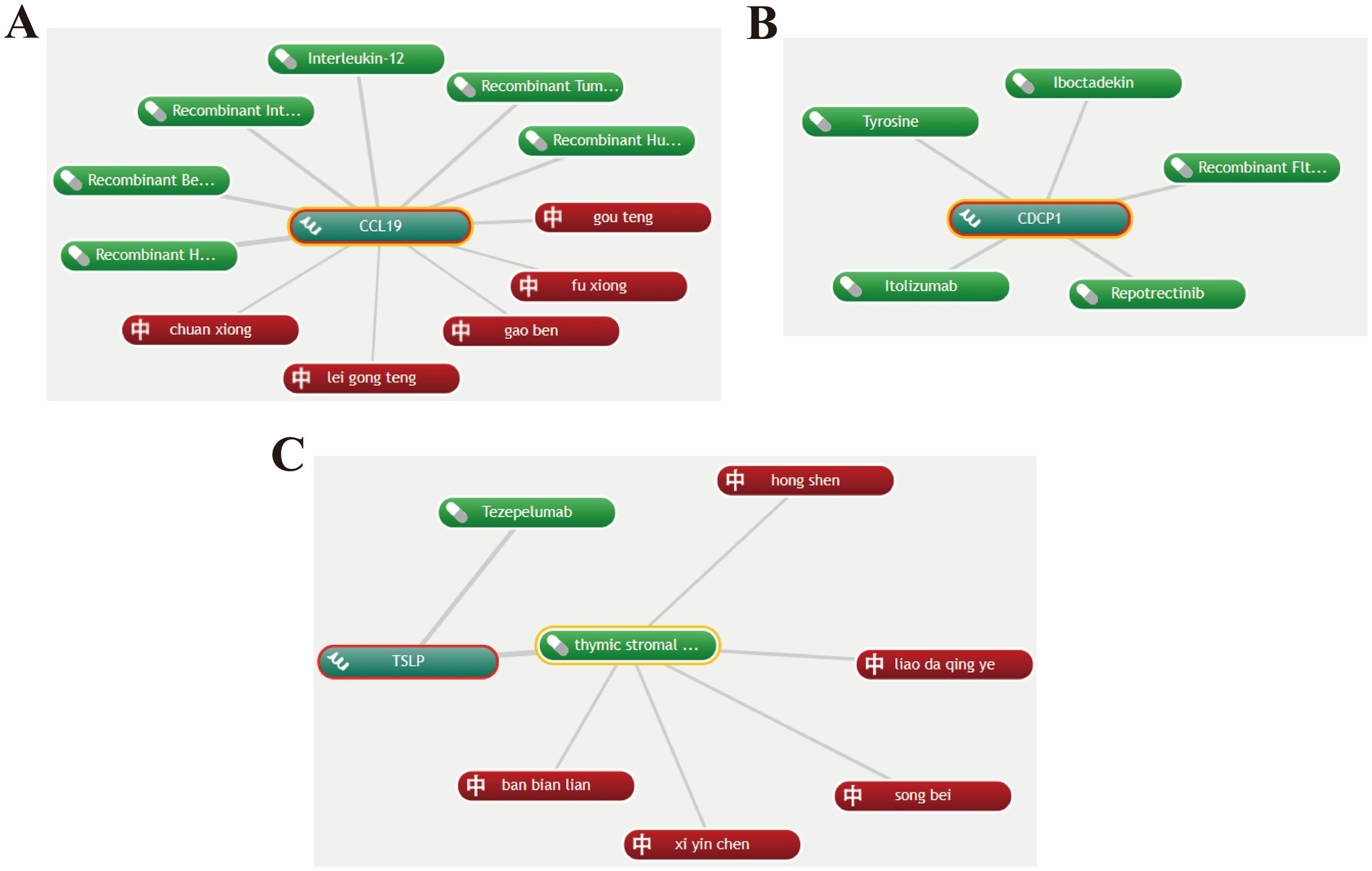
Figure 6. Potential drugs targeting CCL19, CDCP1 and TSLP. (A) Potential drugs targeting CCL19. (B) Potential drugs targeting CDCP1. (C) Potential drugs targeting TSLP. Green represents Western medicines, and red represents Chinese herbal medicines.
Based on the drug repurposing results from “CoreMine,” the drugs targeting CDCP1 include tyrosine, iboctadekin, itolizumab, repotrectinib, and recombinant Flt3 ligand. Among these, tyrosine can act as a mood elevator and antidepressant, while also improving memory and increasing mental alertness (47, 48) (Figure 6B).
Tezepelumab, a human monoclonal IgG2λ antibody that blocks TSLP, is the primary drug targeting TSLP and is mainly used for the treatment of asthma (49). Traditional Chinese medicines targeting TSLP include Carthami flos and Lobeliae chinensis Herba (Figure 6C).
3.5 Molecular docking results
After identifying traditional Chinese medicines targeting CCL19 through “CoreMine,” including Chuanxiong Rhizoma, Uncariae Ramulus Cumuncis, Ligustici Rhizoma Et Radix, and Tripterygii Radix, molecular docking analysis was performed to assess the binding affinity of their active components with CCL19 (Supplementary Table S10). The results revealed promising interactions for several compounds: perlolyrine (−7.0 kcal/mol), rhynchophylline (−6.5 kcal/mol), triptolide (−5.7 kcal/mol), and sitosterol (−7.2 kcal/mol), as illustrated in Figure 7. These binding energies indicate stable interactions between the compounds and CCL19, with lower values suggesting stronger potential drug effects at the target site. Notably, binding energies below −7.0 kcal/mol are generally considered to indicate strong binding affinity and potential biological activity (50). Moreover, the drugs we identified exhibit promising binding affinity with CDCP1 and TSLP (Supplementary Table S11). These findings provide insights into the potential efficacy of these TCM components in modulating inflammatory proteins-related pathways involved in hearing loss, laying a foundation for further investigation into their therapeutic potential and mechanisms of action in addressing hearing impairments.
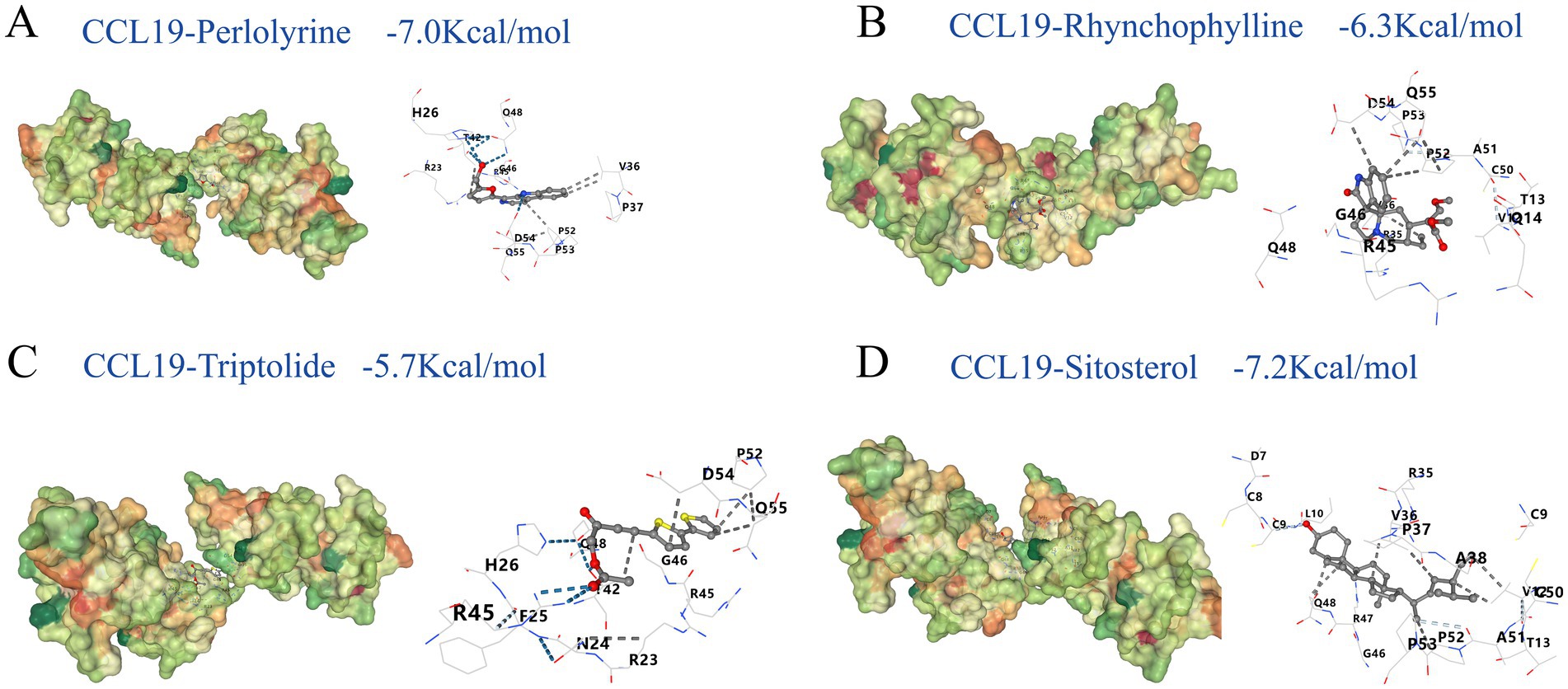
Figure 7. Molecular docking results. This figure shows the molecular docking results of perlolyrine (A), rhynchophylline (B), triptolide (C) and sitosterol (D) with CCL19. The binding energies are −7.0, −6.3, −5.7, −7.2 kcal/mol for A–D, respectively. In each subfigure, the 3D structure displays the interaction between the ligand and the protein, with key amino acid residues and interaction types labeled. The blue dashed lines represent hydrogen bonds; white dashed lines represent weak hydrogen bonds; black dashed lines represent hydrophobic contacts.
4 Discussion
Our MR analysis revealed CCL19 as a significant risk factor for both SNHL and OHL (Figure 2). This association was further validated in datasets focusing on hearing loss and difficulty in hearing (Figure 3A), demonstrating robust reliability. CCL19 is a chemokine that serves as a crucial mediator in immune responses and inflammatory processes (51). The immune homeostasis of the inner ear is predominantly maintained by resident macrophages distributed across various cochlear structures, including the spiral ligament, spiral ganglion, and stria vascularis. These macrophages serve multiple functions: they detect and eliminate tissue debris and pathogens through phagocytosis, while simultaneously orchestrating inflammatory responses and tissue repair by secreting inflammatory cytokines and chemokines (52). The interaction between CCL19 and CCR7 receptors on macrophage surfaces triggers downstream signaling cascades, thereby modulating macrophage chemotaxis, migration, and functional polarization (53). Accumulating evidence strongly supports CCL19’s role as a risk factor in hearing loss development. Experimental studies have demonstrated that noise exposure significantly elevates CCL19 expression in GRAIL wild-type mice, whereas GRAIL gene deletion reduces CCL19 levels and subsequently attenuates hearing loss, suggesting CCL19 as a promising therapeutic target for noise-induced hearing loss (54). Further supporting evidence comes from gene expression analyses of rat endolymphatic sac (ES), which revealed that dysregulation of CCL19-related genes may contribute to autoimmune inner ear disorders (55). Moreover, CCL19 has been implicated in inner ear aging processes. Through comprehensive comparative analyses of age-related differential gene expression and aging signatures in mouse cochlea, researchers identified CCL19 as a high-risk gene associated with age-related hearing decline (56).
Notably, CCL2 plays a pivotal role in the pathogenesis of hearing loss. Following cochlear injury, such as noise exposure, CCL2 levels are significantly elevated in the organ of Corti and surrounding inner ear tissues (57). Further investigations have demonstrated that anti-inflammatory medications effectively reduce CCL2 levels and ameliorate hearing loss (58, 59). The involvement of inflammatory and immune responses in cochlear damage is evidenced by the upregulation of inflammation/immune-related genes and increased immune cell infiltration following various types of injury (60). The mitochondrial toxin 3-nitropropionic acid (3-NP) induces acute mitochondrial dysfunction and activates the IL-6/CCL2 inflammatory pathway, leading to secondary inflammatory responses in the cochlear lateral wall. CCL2 has emerged as a potential therapeutic target for treating injuries resulting from acute mitochondrial dysfunction in the cochlear lateral wall (61). Additionally, studies have observed upregulation of CCL2-related genes in aged mouse cochlea, suggesting its association with age-related hearing loss (62).
Our MR analysis identified CDCP1 as a risk factor and TSLP as a protective factor for both SIHL and OHL (Figure 2). Although previous research found no significant association between CDCP1 and persistent tinnitus (63), the impact of CDCP1 on hearing remains largely unexplored. Our multi-omics analysis (Figure 5) reveals that CDCP1 regulates the downstream transcription factor STAT3 (64), which differentially modulates inflammatory and apoptotic signaling in the cochlea and regulates TRPV1 channels (65). TRPV1 has been established as a mediator for aminoglycoside entry into hair cells and is implicated in cisplatin-induced ototoxicity (66, 67). Suppression of oxidative stress or inflammation reduces TRPV1 channel expression, thereby preventing cochlear damage and hearing loss (65). Furthermore, studies have demonstrated that apelin-13 and capsaicin can protect against cisplatin-induced ototoxicity through STAT3 regulation (68, 69), suggesting the need for further investigation into the relationship between STAT3 and hearing loss, particularly in the context of ototoxicity. Research on TSLP’s role in hearing loss is limited; however, studies have shown significantly elevated TSLP levels in children with chronic serous otitis media, where it initiates and maintains local inflammatory responses in the Eustachian tube and may contribute to middle ear effusion in non-atopic patients (70).
Our functional enrichment analysis revealed that SIHL-associated proteins were significantly enriched in lipid metabolism pathways (including serum triglycerides) (Figure 4B) and schizophrenia-related pathways (Supplementary Table S9). The multi-omics network analysis (Figure 5) further demonstrated the role of inflammation-related miRNAs in neurodegenerative diseases. High-fat diets can induce systemic inflammation (71), which compromises blood-brain barrier (BBB) permeability and facilitates inflammatory factor infiltration into the brain, potentially leading to neurodegeneration (72, 73). Our tissue analysis identified enrichment of these inflammatory factors in the amygdala (Supplementary Figure S8), a region crucial for anxiety, post-traumatic stress disorder (PTSD), various psychological disorders, and auditory processing. The release of inflammatory factors can impair emotional regulation in the amygdala, increasing the risk of anxiety and depression, thereby exacerbating hearing loss (74). Chronic inflammation may also disrupt the amygdala’s stress response system, elevating the risk of neurodegenerative diseases such as Alzheimer’s disease (75, 76). Additionally, serum triglycerides can affect cochlear blood supply (77), compromise blood-labyrinth barrier integrity, leading to reduced oxygen supply to the inner ear and subsequent ischemic damage to hair cells (78). Our study highlights the connections between inflammation and various types of hearing loss, exploring the crosstalk between inflammation, downstream signaling pathways, and miRNAs, as well as their localization in the brain, thereby advancing our understanding of inflammation-mediated hearing loss mechanisms. Furthermore, we identified potential small-molecule drugs targeting these proteins (Figure 7), potentially offering new therapeutic strategies for hearing loss treatment.
The superiority of this study lies in its comprehensive investigation of the relationship between inflammation and hearing loss. Unlike previous studies that focused on a limited number of inflammatory markers (such as CRP and IL-6) (13, 79), this study expanded the scope to 91 inflammatory proteins. The study benefits from a comprehensive proteomic profile, a diverse set of outcomes (SNHL, SIHL, and OHL), minimal confounding and bias, and validation using a large cohort dataset, resulting in more robust findings and a more complete network of inflammation and hearing loss. Furthermore, Functional annotation, tissue analyses, and multi-omics network analysis provide further insights into the disease mechanisms. Drug repurposing and molecular docking analysis also offers potential directions for clinical treatment.
However, this study has some limitations. The population in this study primarily consists of Europeans, and further generalization should be made with caution. Due to the limitations of the available data, we were unable to further subdivide SNHL into age-related hearing loss and noise-induced hearing loss (80), which we believe would greatly benefit personalized treatment and should be a direction for future research. Although this study emphasizes the key role of CCL19 in hearing loss, current research findings suggest that further investigation of the CCL family, especially CCL2, is necessary. While Mendelian randomization can largely avoid the influence of confounding factors on the outcomes, further validation through animal experiments and clinical trials is still crucial.
5 Conclusion
This study investigated the causal relationships between various inflammatory proteins (including CCL19, CDCP1, and TSLP) and multiple types of hearing loss (SNHL, SIHL, and OHL). The research focused on downstream inflammatory signaling pathways and miRNA crosstalk, integrating enrichment and tissue analyses to construct an “inflammatory protein-hearing loss” network. Mechanisms by which inflammatory proteins lead to hearing loss were explored, and small molecule drugs targeting these proteins were identified. Future research directions encompass more refined analyses for different types of hearing loss, in-depth exploration of the relationship between CCL family members (particularly CCL2) and hearing loss, and prospective studies in both animal models and human populations. These efforts will enhance our understanding of the pathogenesis of hearing loss and provide additional leads for drug development.
Data availability statement
The datasets presented in this study can be found in online repositories. The FinnGen database data are available at https://www.finngen.fi/en/access_results and the GWAS summary datasets are available at https://gwas.mrcieu.ac.uk/.
Author contributions
JZ: Data curation, Visualization, Writing – original draft. TG: Investigation, Writing – review & editing. YC: Writing – review & editing. XW: Writing – review & editing. LW: Writing – review & editing. HX: Validation, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This research was supported by National Key Research and Development Program (2018YFC1704104) and Sichuan Province Science Popularization Training Project (2022JDKP0018).
Acknowledgments
The authors would like to express our gratitude to the FinnGen database for providing the data, and to the editors and reviewers for their helpful comments on the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fneur.2024.1422241/full#supplementary-material
Abbreviations
ARHL, Age-related hearing loss; SNHL, Sensorineural hearing loss; ROS, Reactive oxygen species; MR, Mendelian randomization; SIHL, Sudden idiopathic hearing loss; OHL, Other hearing loss; FUMA, Functional mapping and annotation of genome-wide association studies; SNPs, Single-nucleotide polymorphisms; LD, Linkage disequilibrium; MAF, Minor allele frequency; IVW, Inverse variance weighted; FDR, False discovery rate; NCBI, National Center for Biotechnology Information; TCMs, Traditional Chinese medicines; UBC, Enzyme E2 C; EGR1, Early growth response 1; STAT3, Signal transducer and activator of transcription 3; NF-κB, Nuclear factor kappa-B; TSLP, Thymic stromal lymphopoietin; TRPV1, Transient receptor potential vanilloid 1.
Footnotes
1. ^https://www.finngen.fi/en/access_results
3. ^https://www.ncbi.nlm.nih.gov/
References
1. GBD 2015 Disease and Injury Incidence and Prevalence Collaborators . Global, regional, and national incidence, prevalence, and years lived with disability for 310 diseases and injuries, 1990–2015: a systematic analysis for the Global Burden of Disease Study 2015. Lancet. (2016) 388:1545–602. doi: 10.1016/s0140-6736(16)31678-6
2. Kalinec, GM, Webster, P, Lim, DJ, and Kalinec, F. A cochlear cell line as an in vitro system for drug ototoxicity screening. Audiol Neurootol. (2003) 8:177–89. doi: 10.1159/000071059
3. Hahn, R, and Avraham, KB. Gene therapy for inherited hearing loss: updates and remaining challenges. Audiol Res. (2023) 13:952–66. doi: 10.3390/audiolres13060083
4. Basner, M, Babisch, W, Davis, A, Brink, M, Clark, C, Janssen, S, et al. Auditory and non-auditory effects of noise on health. Lancet. (2014) 383:1325–32. doi: 10.1016/s0140-6736(13)61613-x
5. Cohen, BE, Durstenfeld, A, and Roehm, PC. Viral causes of hearing loss: a review for hearing health professionals. Trends Hear. (2014) 18:18. doi: 10.1177/2331216514541361
6. Yamasoba, T, Lin, FR, Someya, S, Kashio, A, Sakamoto, T, and Kondo, K. Current concepts in age-related hearing loss: epidemiology and mechanistic pathways. Hear Res. (2013) 303:30–8. doi: 10.1016/j.heares.2013.01.021
7. Cunningham, LL, and Tucci, DL. Hearing loss in adults. N Engl J Med. (2017) 377:2465–73. doi: 10.1056/NEJMra1616601
8. Lanvers-Kaminsky, C, Zehnhoff-Dinnesen, AA, Parfitt, R, and Ciarimboli, G. Drug-induced ototoxicity: mechanisms, pharmacogenetics, and protective strategies. Clin Pharmacol Ther. (2017) 101:491–500. doi: 10.1002/cpt.603
9. Fujioka, M, Kanzaki, S, Okano, HJ, Masuda, M, Ogawa, K, and Okano, H. Proinflammatory cytokines expression in noise-induced damaged cochlea. J Neurosci Res. (2006) 83:575–83. doi: 10.1002/jnr.20764
10. Woo, JI, Pan, H, Oh, S, Lim, DJ, and Moon, SK. Spiral ligament fibrocyte-derived MCP-1/CCL2 contributes to inner ear inflammation secondary to nontypeable H. influenzae-induced otitis media. BMC Infect Dis. (2010) 10:314. doi: 10.1186/1471-2334-10-314
11. Yang, W, Zhao, X, Chai, R, and Fan, J. Progress on mechanisms of age-related hearing loss. Front Neurosci. (2023) 17:1253574. doi: 10.3389/fnins.2023.1253574
12. Kaur, T, Borse, V, Sheth, S, Sheehan, K, Ghosh, S, Tupal, S, et al. Adenosine A1 receptor protects against cisplatin ototoxicity by suppressing the NOX3/STAT1 inflammatory pathway in the cochlea. J Neurosci. (2016) 36:3962–77. doi: 10.1523/jneurosci.3111-15.2016
13. Gupta, S, Curhan, SG, and Curhan, GC. Biomarkers of systemic inflammation and risk of incident hearing loss. Ear Hear. (2019) 40:981–9. doi: 10.1097/aud.0000000000000678
14. Lawlor, DA, Harbord, RM, Sterne, JA, Timpson, N, and Davey Smith, G. Mendelian randomization: using genes as instruments for making causal inferences in epidemiology. Stat Med. (2008) 27:1133–63. doi: 10.1002/sim.3034
15. Burgess, S, Small, DS, and Thompson, SG. A review of instrumental variable estimators for Mendelian randomization. Stat Methods Med Res. (2017) 26:2333–55. doi: 10.1177/0962280215597579
16. Zhang, L, Chen, J, Zhong, S, Luo, J, and Tang, P. Causal associations of white blood cell count and sudden sensorineural hearing loss: a bidirectional and multivariable Mendelian randomization study. Front Neurol. (2024) 15:1387244. doi: 10.3389/fneur.2024.1387244
17. Ni, T, Shen, Z, Lu, X, Shi, H, Xie, H, Yang, S, et al. No causal relationship serum lipids on age-related hearing loss based on Mendelian randomized evidence. Hear Res. (2024) 453:109128. doi: 10.1016/j.heares.2024.109128
18. Wang, Y, Nie, J, Yan, K, Wang, J, Wang, X, and Zhao, Y. Inflammatory diet, gut microbiota and sensorineural hearing loss: a cross-sectional and Mendelian randomization study. Front Nutr. (2024) 11:1458484. doi: 10.3389/fnut.2024.1458484
19. Yan, J, Wu, L, Zheng, M, Lv, Y, Jiang, F, Gao, W, et al. Mendelian randomization study reveals a predicted relationship between sensorineural hearing loss and mitochondrial proteins. Otol Neurotol. (2024) 45:e655–63. doi: 10.1097/mao.0000000000004266
20. Skrivankova, VW, Richmond, RC, Woolf, BAR, Yarmolinsky, J, Davies, NM, Swanson, SA, et al. Strengthening the reporting of observational studies in epidemiology using Mendelian randomization: the STROBE-MR statement. JAMA. (2021) 326:1614–21. doi: 10.1001/jama.2021.18236
21. Zhao, JH, Stacey, D, Eriksson, N, Macdonald-Dunlop, E, Hedman, ÅK, Kalnapenkis, A, et al. Genetics of circulating inflammatory proteins identifies drivers of immune-mediated disease risk and therapeutic targets. Nat Immunol. (2023) 24:1540–51. doi: 10.1038/s41590-023-01588-w
22. Kurki, MI, Karjalainen, J, Palta, P, Sipilä, TP, Kristiansson, K, Donner, KM, et al. FinnGen provides genetic insights from a well-phenotyped isolated population. Nature. (2023) 613:508–18. doi: 10.1038/s41586-022-05473-8
23. Sakaue, S, Kanai, M, Tanigawa, Y, Karjalainen, J, Kurki, M, Koshiba, S, et al. A cross-population atlas of genetic associations for 220 human phenotypes. Nat Genet. (2021) 53:1415–24. doi: 10.1038/s41588-021-00931-x
24. Verbanck, M, Chen, CY, Neale, B, and Do, R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat Genet. (2018) 50:693–8. doi: 10.1038/s41588-018-0099-7
25. Vlaic, BA, Vlaic, A, Russo, IR, Colli, L, Bruford, MW, Odagiu, A, et al. Analysis of genetic diversity in Romanian Carpatina goats using SNP genotyping data. Animals. (2024) 14:560. doi: 10.3390/ani14040560
26. Papadimitriou, N, Dimou, N, Tsilidis, KK, Banbury, B, Martin, RM, Lewis, SJ, et al. Physical activity and risks of breast and colorectal cancer: a Mendelian randomisation analysis. Nat Commun. (2020) 11:597. doi: 10.1038/s41467-020-14389-8
27. EPIC-InterAct ConsortiumBurgess, S, Scott, RA, Timpson, NJ, Davey Smith, G, and Thompson, SG. Using published data in Mendelian randomization: a blueprint for efficient identification of causal risk factors. Eur J Epidemiol. (2015) 30:543–52. doi: 10.1007/s10654-015-0011-z
28. Burgess, S, Butterworth, A, and Thompson, SG. Mendelian randomization analysis with multiple genetic variants using summarized data. Genet Epidemiol. (2013) 37:658–65. doi: 10.1002/gepi.21758
29. Watanabe, K, Taskesen, E, van Bochoven, A, and Posthuma, D. Functional mapping and annotation of genetic associations with FUMA. Nat Commun. (2017) 8:1826. doi: 10.1038/s41467-017-01261-5
30. Fadista, J, Manning, AK, Florez, JC, and Groop, L. The (in)famous GWAS p-value threshold revisited and updated for low-frequency variants. Eur J Hum Genet. (2016) 24:1202–5. doi: 10.1038/ejhg.2015.269
31. Panagiotou, OA, and Ioannidis, JPAGenome-Wide Significance Project. What should the genome-wide significance threshold be? Empirical replication of borderline genetic associations. Int J Epidemiol. (2011) 41:273–86. doi: 10.1093/ije/dyr178
32. Elvsåshagen, T, Shadrin, A, Frei, O, van der Meer, D, Bahrami, S, Kumar, VJ, et al. The genetic architecture of the human thalamus and its overlap with ten common brain disorders. Nat Commun. (2021) 12:2909. doi: 10.1038/s41467-021-23175-z
33. Xia, J, Gill, EE, and Hancock, RE. NetworkAnalyst for statistical, visual and network-based meta-analysis of gene expression data. Nat Protoc. (2015) 10:823–44. doi: 10.1038/nprot.2015.052
34. Zhou, G, and Xia, J. Using OmicsNet for network integration and 3D visualization. Curr Protoc Bioinformatics. (2019) 65:e69. doi: 10.1002/cpbi.69
35. Jenssen, TK, Lægreid, A, Komorowski, J, and Hovig, E. A literature network of human genes for high-throughput analysis of gene expression. Nat Genet. (2001) 28:21–8. doi: 10.1038/ng0501-21
36. Ru, J, Li, P, Wang, J, Zhou, W, Li, B, Huang, C, et al. TCMSP: a database of systems pharmacology for drug discovery from herbal medicines. J Cheminform. (2014) 6:13. doi: 10.1186/1758-2946-6-13
37. Liu, Y, Yang, X, Gan, J, Chen, S, Xiao, ZX, and Cao, Y. CB-Dock2: improved protein-ligand blind docking by integrating cavity detection, docking and homologous template fitting. Nucleic Acids Res. (2022) 50:W159–w164. doi: 10.1093/nar/gkac394
38. Liu, Y, Zhang, H, Fan, C, Liu, F, Li, S, Li, J, et al. Potential role of Bcl2 in lipid metabolism and synaptic dysfunction of age-related hearing loss. Neurobiol Dis. (2023) 187:106320. doi: 10.1016/j.nbd.2023.106320
39. Linszen, MM, Brouwer, RM, Heringa, SM, and Sommer, IE. Increased risk of psychosis in patients with hearing impairment: review and meta-analyses. Neurosci Biobehav Rev. (2016) 62:1–20. doi: 10.1016/j.neubiorev.2015.12.012
40. Simpson, AN, Matthews, LJ, and Dubno, JR. Lipid and C-reactive protein levels as risk factors for hearing loss in older adults. Otolaryngol Head Neck Surg. (2013) 148:664–70. doi: 10.1177/0194599812473936
41. Lukiw, WJ . Micro-RNA speciation in fetal, adult and Alzheimer’s disease hippocampus. Neuroreport. (2007) 18:297–300. doi: 10.1097/WNR.0b013e3280148e8b
42. Schaefer, A, O’Carroll, D, Tan, CL, Hillman, D, Sugimori, M, Llinas, R, et al. Cerebellar neurodegeneration in the absence of microRNAs. J Exp Med. (2007) 204:1553–8. doi: 10.1084/jem.20070823
43. Oluwole, OG, James, K, Yalcouye, A, and Wonkam, A. Hearing loss and brain disorders: a review of multiple pathologies. Open Med. (2022) 17:61–9. doi: 10.1515/med-2021-0402
44. Brewster, KK, Hu, MC, Zilcha-Mano, S, Stein, A, Brown, PJ, Wall, MM, et al. Age-related hearing loss, late-life depression, and risk for incident dementia in older adults. J Gerontol A. (2021) 76:827–34. doi: 10.1093/gerona/glaa242
45. Zhang, X, Cao, R, Li, C, Zhao, H, Zhang, R, Che, J, et al. Caffeine ameliorates age-related hearing loss by downregulating the inflammatory pathway in mice. Otol Neurotol. (2024) 45:227–37. doi: 10.1097/mao.0000000000004098
46. Francois, B, Jeannet, R, Daix, T, Walton, AH, Shotwell, MS, Unsinger, J, et al. Interleukin-7 restores lymphocytes in septic shock: the IRIS-7 randomized clinical trial. JCI Insight. (2018) 3:e98960. doi: 10.1172/jci.insight.98960
47. Yao, H, Wang, X, Chi, J, Chen, H, Liu, Y, Yang, J, et al. Exploring novel antidepressants targeting G protein-coupled receptors and key membrane receptors based on molecular structures. Molecules. (2024) 29:964. doi: 10.3390/molecules29050964
48. Fernstrom, JD, and Fernstrom, MH. Tyrosine, phenylalanine, and catecholamine synthesis and function in the brain. J Nutr. (2007) 137:1539S–47S. doi: 10.1093/jn/137.6.1539S
49. Pelaia, C, Pelaia, G, Crimi, C, Maglio, A, Gallelli, L, Terracciano, R, et al. Tezepelumab: a potential new biological therapy for severe refractory asthma. Int J Mol Sci. (2021) 22. doi: 10.3390/ijms22094369
50. Wang, Y, Yuan, Y, Wang, W, He, Y, Zhong, H, Zhou, X, et al. Mechanisms underlying the therapeutic effects of Qingfeiyin in treating acute lung injury based on GEO datasets, network pharmacology and molecular docking. Comput Biol Med. (2022) 145:105454. doi: 10.1016/j.compbiomed.2022.105454
51. Pickens, SR, Chamberlain, ND, Volin, MV, Pope, RM, Mandelin, AM II, and Shahrara, S. Characterization of CCL19 and CCL21 in rheumatoid arthritis. Arthritis Rheum. (2011) 63:914–22. doi: 10.1002/art.30232
52. Li, P, Qian, T, and Sun, S. Spatial architecture of the cochlear immune microenvironment in noise-induced and age-related sensorineural hearing loss. Int Immunopharmacol. (2023) 114:109488. doi: 10.1016/j.intimp.2022.109488
53. Förster, R, Davalos-Misslitz, AC, and Rot, A. CCR7 and its ligands: balancing immunity and tolerance. Nat Rev Immunol. (2008) 8:362–71. doi: 10.1038/nri2297
54. Chuang, KF, Wang, CH, Chen, HK, Lin, YY, Lin, CH, Lin, YC, et al. GRAIL gene knockout mice protect against aging-related and noise-induced hearing loss. J Chin Med Assoc. (2023) 86:1101–8. doi: 10.1097/jcma.0000000000001005
55. Zhang, J, Wang, N, and Xu, A. Screening of genes associated with inflammatory responses in the endolymphatic sac reveals underlying mechanisms for autoimmune inner ear diseases. Exp Ther Med. (2018) 16:2460–70. doi: 10.3892/etm.2018.6479
56. Sun, G, Zheng, Y, Fu, X, Zhang, W, Ren, J, Ma, S, et al. Single-cell transcriptomic atlas of mouse cochlear aging. Protein Cell. (2022) 14:180–201. doi: 10.1093/procel/pwac058
57. Tan, WJ, Thorne, PR, and Vlajkovic, SM. Characterisation of cochlear inflammation in mice following acute and chronic noise exposure. Histochem Cell Biol. (2016) 146:219–30. doi: 10.1007/s00418-016-1436-5
58. Frye, MD, Ryan, AF, and Kurabi, A. Inflammation associated with noise-induced hearing loss. J Acoust Soc Am. (2019) 146:4020–32. doi: 10.1121/1.5132545
59. Prasad, KN, and Bondy, SC. Increased oxidative stress, inflammation, and glutamate: potential preventive and therapeutic targets for hearing disorders. Mech Ageing Dev. (2020) 185:111191. doi: 10.1016/j.mad.2019.111191
60. Fujioka, M, Okano, H, and Ogawa, K. Inflammatory and immune responses in the cochlea: potential therapeutic targets for sensorineural hearing loss. Front Pharmacol. (2014) 5:287. doi: 10.3389/fphar.2014.00287
61. Fujioka, M, Okamoto, Y, Shinden, S, Okano, HJ, Okano, H, Ogawa, K, et al. Pharmacological inhibition of cochlear mitochondrial respiratory chain induces secondary inflammation in the lateral wall: a potential therapeutic target for sensorineural hearing loss. PLoS One. (2014) 9:e90089. doi: 10.1371/journal.pone.0090089
62. Su, Z, Xiong, H, Liu, Y, Pang, J, Lin, H, Zhang, W, et al. Transcriptomic analysis highlights cochlear inflammation associated with age-related hearing loss in C57BL/6 mice using next generation sequencing. PeerJ. (2020) 8:e9737. doi: 10.7717/peerj.9737
63. Cederroth, CR, Hong, MG, Freydin, MB, Edvall, NK, Trpchevska, N, Jarach, C, et al. Screening for circulating inflammatory proteins does not reveal plasma biomarkers of constant tinnitus. J Assoc Res Otolaryngol. (2023) 24:593–606. doi: 10.1007/s10162-023-00920-3
64. Kajiwara, K, Chen, P-K, Abe, Y, Okuda, S, Kon, S, Adachi, J, et al. Src activation in lipid rafts confers epithelial cells with invasive potential to escape from apical extrusion during cell competition. Curr Biol. (2022) 32:3460–3476.e6. doi: 10.1016/j.cub.2022.06.038
65. Ramkumar, V, Sheth, S, Dhukhwa, A, Al Aameri, R, Rybak, L, and Mukherjea, D. Transient receptor potential channels and auditory functions. Antioxid Redox Signal. (2022) 36:1158–70. doi: 10.1089/ars.2021.0191
66. Wang, X, Zhou, Y, Wang, D, Wang, Y, Zhou, Z, Ma, X, et al. Cisplatin-induced ototoxicity: from signaling network to therapeutic targets. Biomed Pharmacother. (2023) 157:114045. doi: 10.1016/j.biopha.2022.114045
67. Tan, WJT, and Vlajkovic, SM. Molecular characteristics of cisplatin-induced ototoxicity and therapeutic interventions. Int J Mol Sci. (2023) 24. doi: 10.3390/ijms242216545
68. Yin, H, Sun, Y, Ya, B, Guo, Y, Zhao, H, Zhang, L, et al. Apelin-13 protects against cisplatin-induced ototoxicity by inhibiting apoptosis and regulating STAT1 and STAT3. Arch Toxicol. (2023) 97:2477–93. doi: 10.1007/s00204-023-03544-x
69. Bhatta, P, Dhukhwa, A, Sheehan, K, RFH, AA, Borse, V, Ghosh, S, et al. Capsaicin protects against cisplatin ototoxicity by changing the STAT3/STAT1 ratio and activating cannabinoid (CB2) receptors in the cochlea. Sci Rep. (2019) 9:4131. doi: 10.1038/s41598-019-40425-9
70. Kumral, TL, Dikker, O, Yıldırım, G, Karaketir, S, Altındağ, C, Çakın, MC, et al. The role of thymic stromal lymphopoietin in the development of chronic otitis media with effusion. Eur Arch Otorrinolaringol. (2022) 279:1937–42. doi: 10.1007/s00405-021-06995-z
71. Bach Knudsen, KE, Lærke, HN, Hedemann, MS, Nielsen, TS, Ingerslev, AK, Gundelund Nielsen, DS, et al. Impact of diet-modulated butyrate production on intestinal barrier function and inflammation. Nutrients. (2018) 10:1499. doi: 10.3390/nu10101499
72. Nation, DA, Sweeney, MD, Montagne, A, Sagare, AP, D’Orazio, LM, Pachicano, M, et al. Blood-brain barrier breakdown is an early biomarker of human cognitive dysfunction. Nat Med. (2019) 25:270–6. doi: 10.1038/s41591-018-0297-y
73. Al-Bachari, S, Naish, JH, Parker, GJM, Emsley, HCA, and Parkes, LM. Blood-brain barrier leakage is increased in Parkinson’s disease. Front Physiol. (2020) 11:593026. doi: 10.3389/fphys.2020.593026
74. Felger, JC . Imaging the role of inflammation in mood and anxiety-related disorders. Curr Neuropharmacol. (2018) 16:533–58. doi: 10.2174/1570159x15666171123201142
75. Johnson, JD, Barnard, DF, Kulp, AC, and Mehta, DM. Neuroendocrine regulation of brain cytokines after psychological stress. J Endocr Soc. (2019) 3:1302–20. doi: 10.1210/js.2019-00053
76. Leng, F, and Edison, P. Neuroinflammation and microglial activation in Alzheimer disease: where do we go from here? Nat Rev Neurol. (2021) 17:157–72. doi: 10.1038/s41582-020-00435-y
77. Wang, Y, Liu, H, Nie, X, Lu, N, Yan, S, Wang, X, et al. L-shaped association of triglyceride glucose index and sensorineural hearing loss: results from a cross-sectional study and Mendelian randomization analysis. Front Endocrinol. (2024) 15:1339731. doi: 10.3389/fendo.2024.1339731
78. Shi, X . Physiopathology of the cochlear microcirculation. Hear Res. (2011) 282:10–24. doi: 10.1016/j.heares.2011.08.006
79. Lassale, C, Vullo, P, Cadar, D, Batty, GD, Steptoe, A, and Zaninotto, P. Association of inflammatory markers with hearing impairment: the English longitudinal study of ageing. Brain Behav Immun. (2020) 83:112–9. doi: 10.1016/j.bbi.2019.09.020
Keywords: sensorineural hearing loss, sudden idiopathic hearing loss, Mendelian randomization, inflammation, multi-omics
Citation: Zhang J, Guo T, Chen Y, Wang X, Wu L and Xie H (2024) Investigating the causal relationship between inflammation and multiple types of hearing loss: a multi-omics approach combining Mendelian randomization and molecular docking. Front. Neurol. 15:1422241. doi: 10.3389/fneur.2024.1422241
Edited by:
Qingyin Zheng, Case Western Reserve University, United StatesReviewed by:
Jun Wang, The First Affiliated Hospital of Nanchang University, ChinaHongyang Wang, People’s Liberation Army General Hospital, China
Yan Sun, Yantai Yuhuangding Hospital, China
Copyright © 2024 Zhang, Guo, Chen, Wang, Wu and Xie. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hui Xie, d2FuZ3hpZS1jdHVAMTYzLmNvbQ==