
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Neurol. , 16 January 2019
Sec. Movement Disorders
Volume 9 - 2018 | https://doi.org/10.3389/fneur.2018.01117
 Felipe Franco da Graça1
Felipe Franco da Graça1 Thiago Junqueira Ribeiro de Rezende1
Thiago Junqueira Ribeiro de Rezende1 Luiz Felipe Rocha Vasconcellos2José Luiz Pedroso3
Luiz Felipe Rocha Vasconcellos2José Luiz Pedroso3 Orlando Graziani P. Barsottini3
Orlando Graziani P. Barsottini3 Marcondes C. França Jr.1*
Marcondes C. França Jr.1*Hereditary spastic paraplegias (HSP) are a large group of genetic diseases characterized by progressive degeneration of the long tracts of the spinal cord, namely the corticospinal tracts and dorsal columns. Genotypic and phenotypic heterogeneity is a hallmark of this group of diseases, which makes proper diagnosis and management often challenging. In this scenario, magnetic resonance imaging (MRI) emerges as a valuable tool to assist in the exclusion of mimicking disorders and in the detailed phenotypic characterization. Some neuroradiological signs have been reported in specific subtypes of HSP and are therefore helpful to guide genetic testing/interpretation. In addition, advanced MRI techniques enable detection of subtle structural abnormalities not visible on routine scans in the spinal cord and brain of subjects with HSP. In particular, quantitative spinal cord morphometry and diffusion tensor imaging look promising tools to uncover the pathophysiology and to track progression of these diseases. In the current review article, we discuss the current use and future perspectives of MRI in the context of HSP.
Hereditary spastic paraplegias (HSP) are a large group of genetic diseases characterized by progressive degeneration of the long tracts of the spinal cord, namely the corticospinal tracts and dorsal columns. Patients typically present with lower limb-predominant spasticity and weakness leading to gait abnormalities. Sensory deficits and urinary complaints are also frequently found (1). In the realm of neurogenetics, HSP is perhaps the condition with the most striking genetic heterogeneity. It may segregate as an autosomal dominant, autosomal recessive or X-linked trait. There are now around 70 loci and 60 genes associated to different forms of HSP (2). For the practicing clinician, this represents a diagnostic challenge and poses difficulty for proper therapeutic management as well as genetic counseling.
Neuroimaging is a powerful tool that enables the structural and functional assessment of the central nervous system (CNS). In particular, advanced techniques of magnetic resonance imaging (MRI) are able to provide detailed microstructural and biochemical information of the CNS; they have proven useful to uncover abnormalities in closely related heredodegenerative diseases, such as spinocerebellar ataxias and amyotrophic lateral sclerosis (3, 4). In the context of HSPs, MRI may assist in the phenotypic characterization and therefore help in the genetic testing approach (5). More recently, some studies using advanced and quantitative techniques have shown microstructural white matter abnormalities in HSP, not detectable with routine MRI sequences (6, 7). These results brought novel insights into the pathophysiology of HSPs and raised the possibility of using MRI as a biomarker to track disease progression. In this review article, we will discuss the available data concerning neuroimaging in HSP.
The precise diagnosis of HSPs, considering the great genetic variability with often similar phenotypes, is challenging. However, as with any neurological disease, a combination of detailed history, assessment of inheritance pattern, accompanying symptoms and physical examination is paramount to reduce the number of hypotheses and to optimize genetic testing (8).
In patients with spastic paraparesis, MRI is essential, initially to rule out usual causes of paraplegia such as compressive, inflammatory, infectious, or vascular myelopathies. Once the diagnosis of HSP is the most likely, neuroimaging may help in establishing the subtype.
The evaluation of spinal cord images, although very useful to rule out differential diagnoses, will rarely provide relevant tips for establishing the HSP subtype. Considering the pathophysiology of this group of diseases (involving axonal degeneration of long motor tracts), it is easy to understand why spinal cord volumetric reduction is found in most patients—Figure 1 (9, 10). Studies looking at the cervical and thoracic spinal cord cross-sectional areas in patients with ADP HSP highlighted significant alterations in all subtypes evaluated (SPG3A, 4, 6, and 8), particularly for SPG 6 and SPG 8 (without direct correlation between the degree of atrophy and clinical deficits) (9). Nevertheless, studies with SPG11, an AR subtype of HSP, demonstrated a direct correlation between disease duration and the severity of spinal cord volumetric reduction (6). Such atrophy, however, is not always evident in visual analyses and even if it is, this characteristic is not exclusive for HSPs and may also be found, for example, in acquired motor neuron diseases (4). Anecdotal reports of spinal cord abnormalities in HSPs have been also described. For example, prominent atrophy of the gracile fasciculus leading to visible enlargement of the ependymal canal in patients with AR HSP was reported (without specifying the subtype) (10). A Single patient with SPG 56 and hydromyelia revealed in MRI scans was described, suggesting that this finding was part of the general clinical picture and not the cause of paraparesis itself (11).
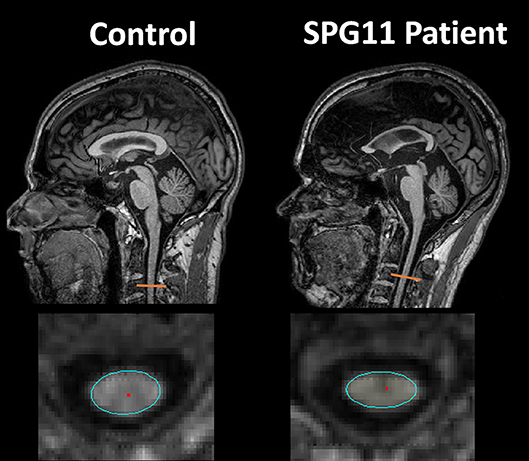
Figure 1. Sagittal and axial T1 weighted images showing cervical spinal cord atrophy and antero-posterior flattening in a patient with SPG11 (right column) compared to a healthy control (left column).
Unlike spinal cord imaging, brain MRI may show peculiar findings that may provide relevant clues for a specific genetic subtype. The presence of a thin corpus callosum (TCC) in a complicated AR HSP phenotype has been used to characterize a certain group of patients (HSP-TCC) (12). It was first considered that this MRI pattern was specific for SPG11 (Figure 2B), but it is now clear that other genotypes may share the same imaging profile. A comprehensive genetic evaluation of an Italian cohort of patients with HSP-TCC (n = 61) was performed and found SPG11 to be the most frequent subtype (26.2%), followed by SPG15 (14.8%), SPG35 (5%), and SPG48 (3%) (13). Other less frequent causes of HSP-TCC are SPG 4, 7, 18, 21, 46, 47, 49, and 54 (8). A new mutation for the SPG3A gene also presented with TCC (14).
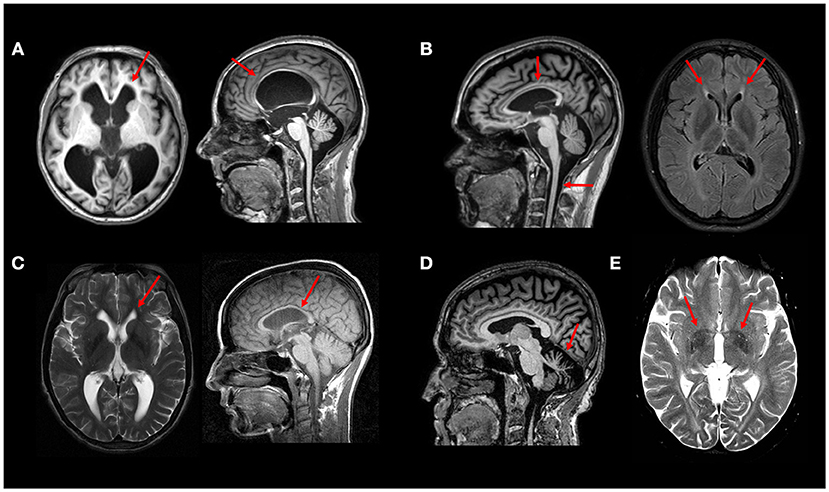
Figure 2. Typical MRI findings in distinct HSP subtypes—(A) Axial and sagittal T1W images showing hydrocephalus due to aqueductal stenosis in a patient with SPG4. This finding is rather unusual in SPG4, but relatively frequent in SPG1 patients. (B) Sagittal T1W image showing thin corpus callosum and spinal cord atrophy in a patient with SPG11; Axial FLAIR image showing the “ear-of-the-lynx” sign in the same patient. (C) Thin corpus callosum (T1W image) and subtle “ear-of-the-lynx” sign (T2W image) in a patient with SPG15. (D) Sagittal T1W image revealing marked cerebellar atrophy in a patient with SPG7. (E) White matter hyperintensities and T2 hypointense signal at the globi pallidi in a 21-years old patient with SPG35 (T2W image). All images were obtained during routine clinical care of these patients at UNICAMP and UNIFESP hospitals.
SPG11 itself has the most classic neuroimaging finding in this group of diseases (15). The so-called “ear-of-the-lynx sign” (Figure 2B) is characterized by an abnormality at the forceps minor of the corpus callosum (genum fibers), which appears hyperintense on T2-FLAIR-weighted and hypointense on T1-weighted images (16). This radiological sign may also be present in another form of complicated AR HSP: SPG 15 (Figure 2C). The visual evaluation of T2-FLAIR-weighted images has sensitivity and specificity for these 2 HSP subtypes as high as 94 and 97%, respectively (Pascual et al. Accepted). In a compatible clinical setting, this should raise a strong suspicion for the above described subtypes, even in the early stages of the disease.
SPG1, also known as MASA syndrome (Mental retardation, Aphasia, Shuffling gait, and Adducted thumbs) is a disease allelic to X-linked aqueductal stenosis or hydrocephalus and may also present with enlarged ventricles. Therefore, in a patient with X-linked inheritance and hydrocephalus, SPG1 should be the main hypothesis (17). Hydrocephalus has been also rarely described in SPG4 patients (as did subarachnoid cysts) Figure 2A (18, 19).
SPG2, on the other hand, is also X-linked and shares phenotypic similarities with an allelic disease, Pelizaeus Merzbacher disease (PMD). Although leukoencephalopathy is a much more evident feature in PMD, it may be also present in SPG2 (20). Multifocal areas of T2 white matter hyperintensities are also a possible presenting feature (sometimes they may even resemble those seen in multiple sclerosis) (21). This last MRI pattern has been reported in some AR-HSP subtypes, such as SPG 5, 21, and 35 (22–24). Other forms of autosomal recessive HSP with white matter disease include SPG11, SPG22, SPG26, SPG44, SPG45, SPG47, SPG50, SPG51, SPG52, SPG54, SPG56, SPG63, SPG64, and SPG 67 (25).
Considering the frequent association of extrapyramidal symptoms in patients with complicated HSPs, changes in the basal ganglia have been conspicuously described, mainly as volumetric abnormalities. Visually perceptible changes, although rare, exist and may assist in the diagnostic definition. In particular, one should remember that some HSP subtypes have basal ganglia iron deposition revealed as hypointense signal lesions in T2, T2* or Susceptibility weighted images. Patients with SPG28, SPG35, and SPG43 (24–27) may present NBIA phenotype with T2 hypointense signal at the globus pallidus (Figure 2E).
It is worth remembering, however, that other NBIAs also manifest with spasticity and similar imaging findings, and should be considered in the differential diagnosis. In addition to structural changes in the basal ganglia (BG), reduced dopaminergic binding with TRODAT has been described in SPG11 and SPG7 patients with complicated HSP phenotype and dopa-responsive parkinsonism (28, 29). Besides BG abnormalities, SPG7 may also present marked and diffuse cerebellar atrophy—Figure 2D (30).
Finally, neuroimaging may assist in the differential diagnosis of other diseases that are not included in the HSP group, but which may manifest with a predominance of spastic symptoms. The presence of a marked cerebellar atrophy with AD history may raise the suspicion for spinocerebellar ataxias (SCA), and some may manifest with predominantly paraparetic forms such as SCA3 (Machado-Joseph Disease) and SCA 7 (31). If cerebellar atrophy is associated with superior vermis atrophy and linear hypointensities in the pons, ARSACS is a possibility—Figure 3A (32). Cerebrotendinous Xantomatosis in adults may also display paraparetic symptoms and MRI abnormalities reveal diffuse atrophy, white matter signal changes and focal peri-dentate cerebellar lesions (Figure 3C); increased lactate with decreased n-acetylaspartate are also seen in MR spectroscopy (33, 34). X-Linked adrenomyeloneuropathy is also a differential diagnosis even in women (about 20% of female carriers develop spastic paraparesis in middle age); typical findings are predominantly posterior white matter T2 hyperintensities and reduced spinal cord volume are key features—Figure 3B (35). Adult-onset Langerhans cell histiocytosis (LCH) is a rare proliferative disorder with neurologic symptoms, including cerebellar ataxia and spastic paraparesis (36). LCH brain MRI can demonstrate globus pallidus/dentate nucleus T1 hyperintensity as well as brainstem and cerebellum T2 hyperintensity.
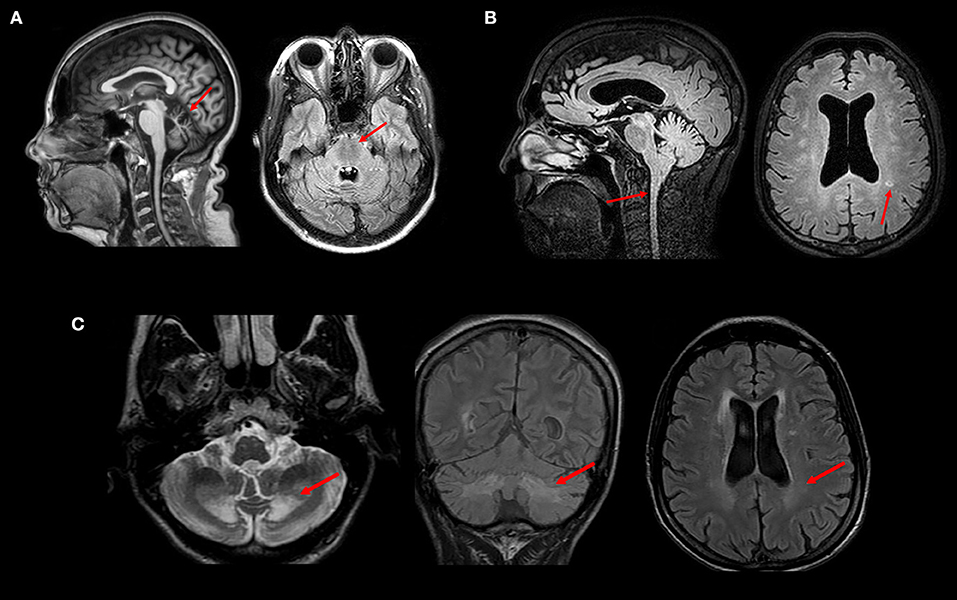
Figure 3. MRI and clinical findings in patients with inherited diseases that may mimic HSP—(A) Superior vermis atrophy (T1W image) and linear hypointensities (FLAIR image) in the pons in a patient with ARSACS. (B) Spinal cord atrophy and white matter hyperintensities (FLAIR images) in a patient with Adrenomyeloneuropathy. (C) Peridentate and periventricular white matter signal changes (FLAIR and T2W images) in a patient with Cerebrotendinous Xanthomatosis. All images were obtained during routine clinical care of these patients at UNICAMP and UNIFESP hospitals.
Despite the above discussed neuroimaging clues for HSP diagnosis, (and the several new descriptions being published in the area) (37), these findings are rarely sensitive or specific. So that their absence should not exclude a diagnostic hypothesis of HSP raised by clinical findings. One must always proceed to diagnostic genetic testing in cases of high clinical suspicion. Different neuroimaging features and their related genetic HSP subtypes herein described are summarized in Table 1, Figure 4 is a flowchart to guide genetic testing for HSP based on MRI findings.
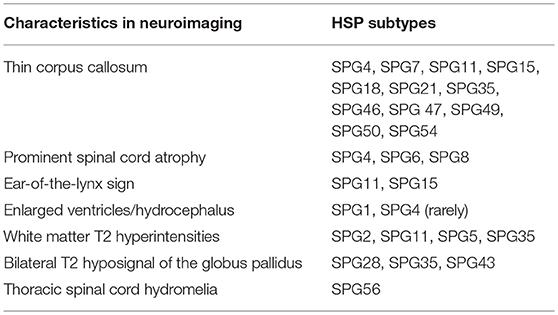
Table 1. Neuroimaging findings in different HSP subtypes.
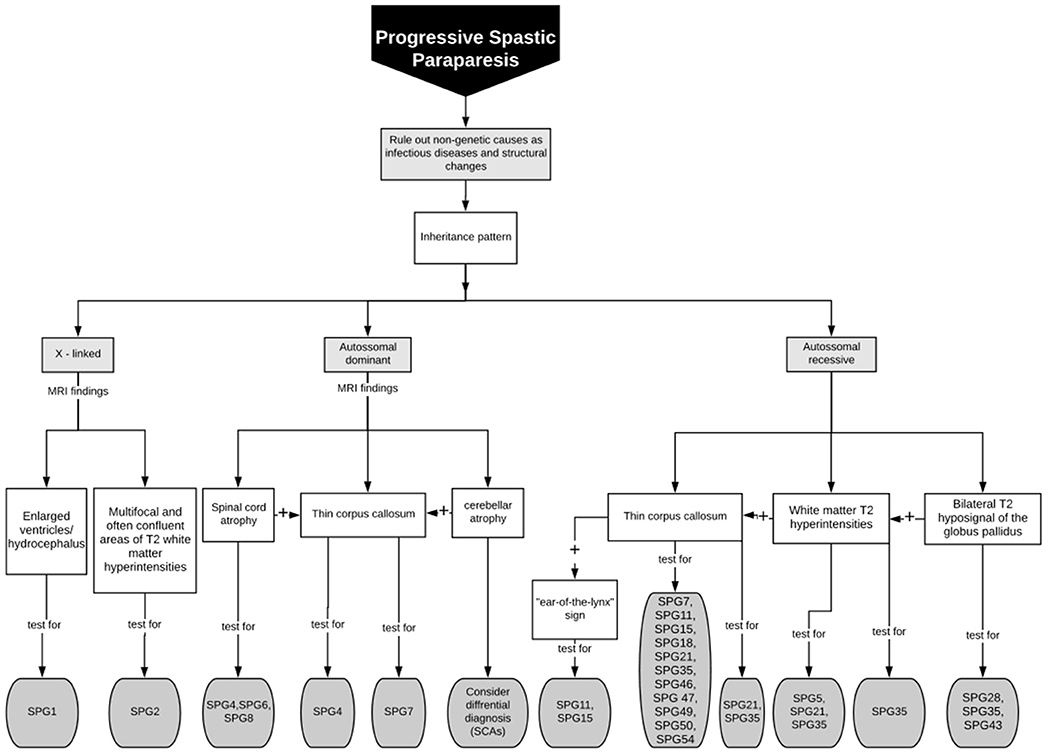
Figure 4. Flowchart to guide genetic testing for HSP based on MRI findings.
The widely variable clinical presentation of patients with HSP suggests that damage is not restricted to the corticospinal tracts in most subtypes of the disease. In this scenario, studies with advanced MRI have been used to investigate the real extension of cerebral damage in the disease (6, 7, 38–47). Such techniques are much more sensitive to capture subtle anatomical abnormalities than pure visual analyses. Moreover, they enable assessment of the CNS in three-dimensions, quantitatively and non-invasively. There are not many image-based studies in HSP and most of them relied upon small cohorts. These reports looked mostly at cerebral structural abnormalities, but some of them also investigated spinal cord morphometry (9, 42, 43). Very few assessed functional abnormalities in HSP (38, 46, 47).
Gray matter abnormalities in HSP—including the cortex and basal ganglia—were assessed using different techniques, such as voxel-based morphometry (VBM, n = 3), cortical thickness analyses (FreeSurfer, n = 1) and atlas-based segmentation volumetry (n = 1). The white matter assessment was performed either through volumetric analysis or through diffusion tensor imaging. To assess the WM volume, two studies used the VBM approach. In contrast, the diffusion images ware assessed using tractography (n = 2) and tract-based spatial statistics (n = 3). Only 5 studies evaluated simultaneously cerebral GM and WM in the same cohort of patients (6, 7, 40, 42, 43).
Advanced neuroimaging techniques are also useful to identify the distinct patterns of damage across the whole phenotypic and genotypic spectrum of HSPs. This might give us relevant insights into the genotype-phenotype correlations and ultimately into the pathophysiology of these disorders. Most studies split HSPs into pure and complicated forms in an attempt to identify different structural signatures for these two subgroups. The former group is characterized by widespread WM abnormalities, but essentially preserved cerebral GM (7, 37, 42). In contrast, the complicated forms, beyond WM damage, show GM volumetric reduction in the basal ganglia and cerebral cortex, particularly at the precentral, paracentral, cingulate, and parahippocampal gyri (6, 43). Overall, these imaging findings are in line with the few pathological reports available (48, 49). At this point, basal ganglia damage appears to be specific for complicated HSP and helps to explain some phenotypic features found in this subgroup, such as parkinsonism and dystonia (Figure 5). Cognitive decline, often severe, is frequent in complicated HSPs and is also related to cerebral GM atrophy. The absence of GM damage in pure HSP also explains why cognitive functions tend to be preserved in this subtype of the disease. Some patients with SPG4—a pure subtype of HSP—develop major cognitive deficits (50). The neuroanatomical substrate for such deficits still deserve investigation, but may be related to subcortical WM involvement, which is prominent in SPG4 (Figure 5).
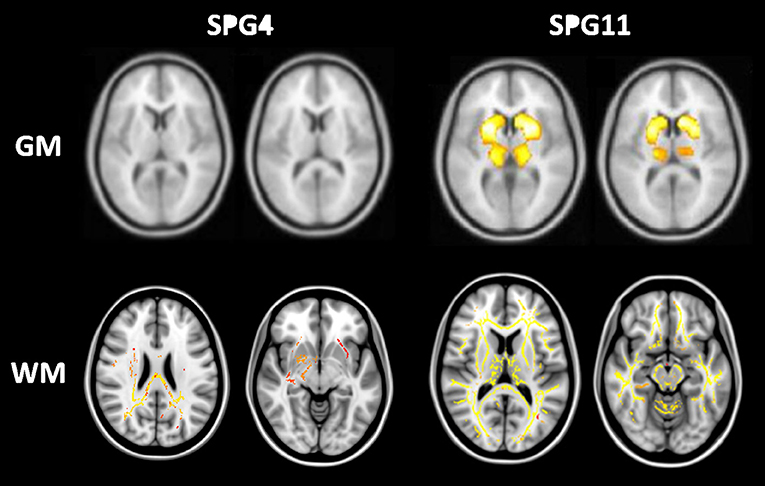
Figure 5. Structural signature for pure (SPG4) and complicated (SPG11) subtypes of HSP stratified for cerebral gray (GM-upper lane) and white matter (WM-lower lane). Regions highlighted in red-yellow are those found to be abnormal in each subtype of the disease. GM and WM results were obtained after comparison with matched healthy controls using voxel-based morphometry and tract-based spatial statistics, respectively. Adapted from references (40) and (42) with permission of the journals.
There are very few imaging studies looking at structural abnormalities in specific genetic subtypes of HSP. As mentioned previously, most authors combined heterogeneous series of patients with different molecular diagnoses and then divided them into pure vs. complicated forms. This was mostly due to the rarity of HSPs in general. More recently, a couple of papers came out with reasonably large cohort sizes of SPG4 and especially SPG11 (6, 7, 38, 42, 51). In SPG4, results largely overlap those reported for pure HSP in general—widespread WM damage (diffusion and volumetric damage) with mild or absent cerebral GM atrophy (7, 42, 51). Interestingly, the mutational profile at SPAST seems to play a role in neuroradiological findings. Indeed, Rezende et al. (42) identified that SPG4 patients with missense mutations had more severe corticospinal tract diffusivity abnormalities than patients with non-sense/frameshift variants (42). This finding is quite interesting because both subgroups were matched for age and disease duration. Distinct neurophysiologic results (somatosensory evoked potentials) were also found in patients with missense and nonsense SPAST mutations (52). In a recent publication of MRI findings and clinical correlates in a multicentric Brazilian cohort of 25 patients with SPG11, authors found widespread cerebral WM (diffusion and volumetric damage) and deep GM abnormalities in these subjects (6). Basal ganglia atrophy correlated with cognitive deficits and also motor function in these subjects. Surprisingly, no clinical correlate for WM damage was found. Furthermore, GM, but not WM, damage was significantly correlated with disease duration. Overall, these results indicate that KIAA1840 mutations impact cerebral GM and WM in distinct ways.
Functional imaging offers the opportunity to investigate how cerebral physiology changes in patients with HSP. To tackle this question, functional MRI (fMRI) and nuclear medicine techniques, such as PET or SPECT, have been used (43, 44, 49–60). The first one relies upon dynamic measurement of the BOLD signal using specific MRI acquisition sequences, whereas the others depend on the detection of radiation emitted by labeled radioligands.
There are 3 studies using fMRI in subjects with HSP (39, 46, 47). In 2 of them, authors employed a motor task to investigate how different cerebral regions are activated in the disease (39, 47). Both studies found abnormal primary motor cortex activation in patients with HSP in comparison to healthy controls; surprisingly, one study found increased whereas the other found reduced activation. In a resting state fMRI study, Liao et al recently reported abnormal functional connectivity between middle frontal and orbitofrontal gyri in patients with SPG4 (49). These authors also found that the amplitude of low frequency fluctuations of the BOLD signal at the right precentral gyrus was increased in the disease. This last parameter was significantly correlated with disease severity.
Cerebral metabolism in HSP was assessed in a few studies using PET and the [18F]-Fluorodeoxyglucose tracer—Table 2 (53–63). Cortical hypometabolism was the usual finding, but the regions affected were rather heterogeneous across the different reports. Such discrepancy is possibly due to the small sample sizes (most studies were single case reports or small series). Parkinsonism and dystonia are now recognized as part of the phenotypic spectrum of some HSP subtypes (29). Hence, some authors employed dopamine transporter (DAT) radiotracers to evaluate the nigrostriatal pathway in the disease using either PET or SPECT (Table 2). In a recent publication, Faber et al. found reduced bilateral striatal DAT uptake to be a universal phenomenon in a cohort of 22 patients with SPG11 (Figure 6). Even patients without parkinsonism had clearly abnormal results (63).
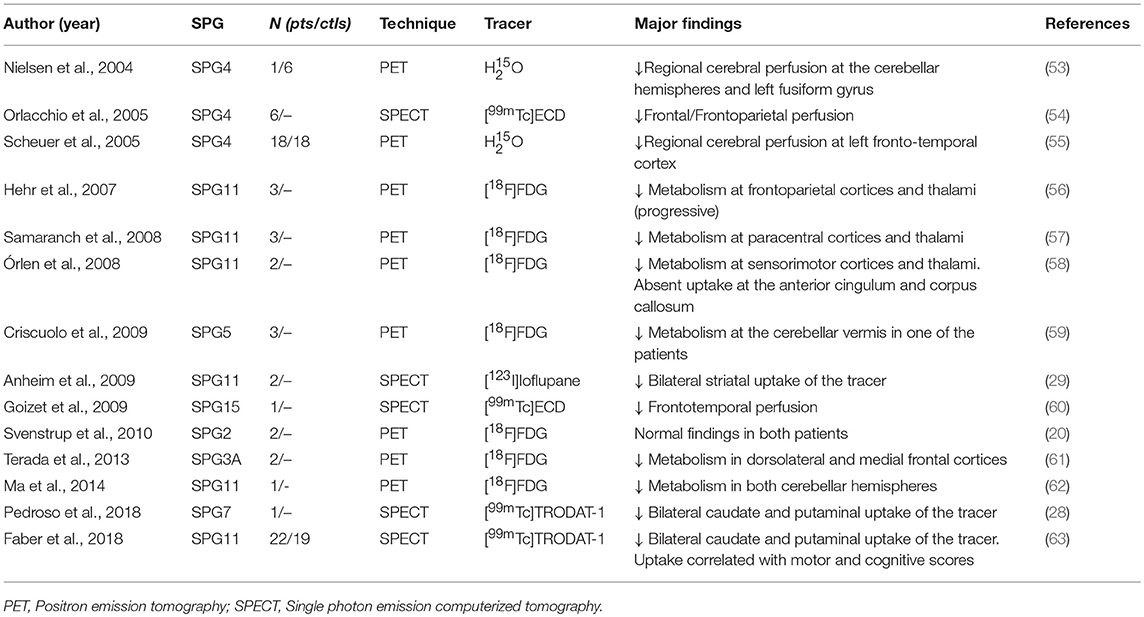
Table 2. Studies using nuclear medicine techniques in different HSP subtypes.
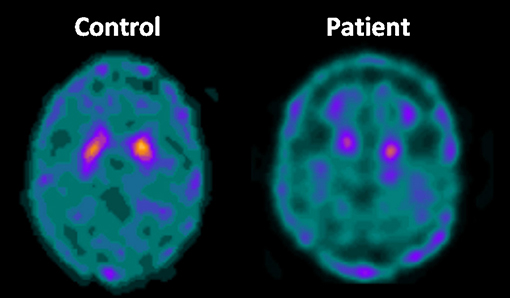
Figure 6. Axial DAT-scan with 99mTc-TRODAT showing reduced striatal uptake in a patient with SPG11 in comparison to a healthy control.
Magnetic resonance spectroscopy is another imaging technique that enables the biochemical characterization of disease-related cerebral abnormalities. The most frequently reported abnormality in patients with HSP is reduction of N-acetylaspartate (a marker for neuronal/axonal function and viability) in the subcortical WM, but other findings have been also described—Table 3 (64–69).
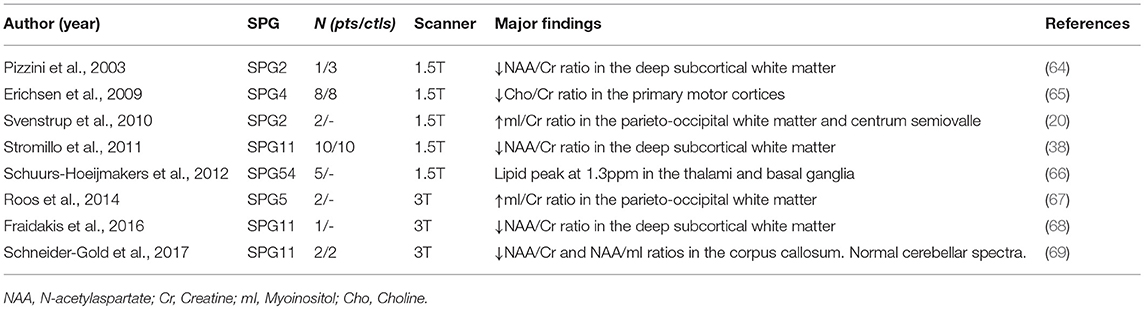
Table 3. Studies using proton magnetic resonance spectroscopy (1H-MRS) in different HSP subtypes.
The spinal cord is perhaps the major target of degeneration in HSPs and MRI studies published so far found it to be atrophic but without antero-posterior flattening in pure and complicated forms of the disease (6, 9, 42, 43, 70). Most reports assessed cervical spinal cord cross sectional area, but at least one also reported reduced thoracic dimensions (70). Postmortem studies corroborate these neuroimaging findings (48, 49). Faber et al. (6) recently found in a large SPG11-cohort that spinal cord area had an inverse correlation with disease duration (6). This argues in favor of a degenerative process taking place. However, it is possible that some sort of developmental issue also occurs. In a closely related heredodegenerative disorder—Friedreich's ataxia-imaging and pathological data support the later hypothesis (71, 72). Further studies in young patients with HSP should be done to check whether the same holds true in this group of diseases.
As basic research progresses and potential disease-modifying treatments become available, biomarkers will be needed to assist in the design of clinical trials for HSPs. In this context, some MRI-derived metrics look promising because they presented significant correlation with clinical data (disease duration and severity). In terms of cerebral abnormalities, candidates appear to be different for pure and complicated HSP subtypes. Most studies with larger cohorts of pure HSP found disease severity, expressed by the spastic paraplegia rating scale—SPRS (73), to be significantly correlated with diffusivity parameters at the anterior limb of internal capsule and corpus callosum, particularly the genu and body (42, 43). Such correlations are not found in complicated HSPs. In this last group, two studies with large SPG11 samples reported independently that the volume of certain cerebral regions correlates with disease-related disability (6, 38). Faber et al. indeed found that precentral/paracentral cortex thickness and deep nuclei volumes correlated not only with SPRS scores but also with disease duration in SPG11 (6). In line with that, the same authors showed that striatal DAT uptake also correlated inversely with disease severity and duration in SPG11 (63).
Spinal cord morphometry is another potential neuroimaging marker for HSPs. The first published studies failed to identify correlations between spinal cord area and disease severity (9, 42, 43, 70). However, this may be related to the small and rather heterogeneous HSP cohorts evaluated at the time. A recent study with a representative cohort of subjects with SPG11 revealed that cervical spinal cord area correlates inversely with SPRS scores and disease duration (6).
There are few studies devoted to characterize neuroimaging abnormalities in HSP so far. Despite that, currently available data strongly support MRI as an important tool to assist in the exclusion of HSP-mimics, to guide genetic testing and to understand the pathophysiology of the disease. For the near future, much work still has to be done in order to fully appreciate how neuroimaging can help us to manage HSP. Considering the rarity of individual HSP subtypes, we should design multicentric studies to obtain representative and homogeneous cohorts of patients. It is rather probable that each specific HSP has its own structural signature (beyond CST damage), but this can only be clarified with properly designed studies.
HSP are very slowly progressive disorders, in such a way that clinical scales have poor sensitivity to track longitudinal changes. For such a rare disease, this greatly limits the design and execution of randomized clinical trials based solely upon clinical tools. So, the potential role of MRI as a state biomarker for HSP should be further explored. Nevertheless, we will require technical and quantitative harmonization between centers before attempting collaborative, multicenter natural history studies. This is certainly a challenging task, but recent efforts done for similar rare neurodegenerative disorders, such as ALS, are encouraging (74). Another relevant question is to identify the most promising neuroimaging markers. At this point, spinal cord morphometry, basal ganglia volumetry and corpus callosum diffusivity emerge as promising candidates, but they still need to be evaluated prospectively. Novel technical advances in spinal cord imaging now enable automated measurements of diffusivity parameters and separate GM vs. WM cross-sectional areas (75). For HSPs, these are other interesting candidates to be investigated.
FdG, TdR, JP, OB, MF, and LV conceived the research project. FdG, TdR, JP, OB, and MF organized the research project, reviewed, and critique the statistical analysis. FdG, TdR, MF, and LV executed the research project and wrote the first draft of the manuscript. FdG, TdR, and MF designed the statistical analysis and executed the statistical analysis. FdG, TdR, JP, OB, and MF. FdG, TdR, MF, and LV. JP, OB, and MF reviewed and critique the manuscript.
TdR receives a Post-Doctoral scholarship from FAPESP. MF received research grants from CNPq (grant 303914/2014-9) and FAPESP (grant 2013/01766-7).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
1. Fink JK, Hedera P. Hereditary spastic paraplegia: genetic heterogeneity and genotype-phenotype correlation. Semin Neurol. (1999) 19:301–9. doi: 10.1055/s-2008-1040846
2. Klebe S, Stevanin G, Depienne C. Clinical and genetic heterogeneity in hereditary spastic paraplegias: from SPG1 to SPG72 and still counting. Rev Neurol. (2015) 171:505–30. doi: 10.1016/j.neurol.2015.02.017
3. Reetz K, Costa AS, Mirzazade S, Lehmann A, Juzek A, Rakowicz M, et al. Genotype specific patterns of atrophy progression are more sensitive than clinical decline in SCA1, SCA3 and SCA6. Brain (2013) 136:905–17. doi: 10.1093/brain/aws369
4. de Albuquerque M, Branco LM, Rezende TJ, de Andrade HM, Nucci A, França MC Jr. Longitudinal evaluation of cerebral and spinal cord damage in Amyotrophic Lateral Sclerosis. Neuroimage Clin. (2017) 14:269–76. doi: 10.1016/j.nicl.2017.01.024
5. Paganoni S, Atassi N. Upper motor neuron disorders hereditary spastic paraplegia and primary lateral sclerosis. Oxf Med Online (2017).
6. Faber I, Martinez ARM, de Rezende TJRD, Martins CR Jr, Martins MP, Lourenço CM, et al. SPG11 mutations cause widespread white matter and basal ganglia abnormalities, but restricted cortical damage. Neuroimage Clin. (2018) 19:848–57. doi: 10.1016/j.nicl.2018.05.031
7. Lindig T, Bender B, Hauser TK, Mang S, Schweikardt D, Klose U, et al. Gray and white matter alterations in hereditary spastic paraplegia type SPG4 and clinical correlations. J Neurol. (2015) 262:1961–71. doi: 10.1007/s00415-015-7791-7
8. Faber I, Servelhere KR, Martinez ARM, D'abreu A, Lopes-Cendes I Jr, França MC Jr. Clinical features and management of hereditary spastic paraplegia. Arq Neuro Psiquiatr. (2014) 72:219–26. doi: 10.1590/0004-282X20130248
9. Hedera P, Eldevik OP, Maly P, Rainier S, Fink JK. Spinal cord magnetic resonance imaging in autosomal dominant hereditary spastic paraplegia. Neuroradiology (2005) 47:730–4. doi: 10.1007/s00234-005-1415-3
10. Hourani R, El-Hajj T, Barada W, Hourani M, Yamout BI. MR imaging findings in autosomal recessive hereditary spastic paraplegia. Am J Neuroradiol. (2009) 30:936–40. doi: 10.3174/ajnr.A1483
11. Masciullo M, Tessa A, Perazza S, Santorelli F, Perna A, Silvestri G. Hereditary spastic paraplegia: novel mutations and expansion of the phenotype variability in SPG56. Eur J Pediatr Neurol. (2016) 20:444–8. doi: 10.1016/j.ejpn.2016.02.001
12. França MC Jr, D'Abreu A, Maurer-Morelli CV, Seccolin R, Appenzeller S, Alessio A, et al. Prospective neuroimaging study in hereditary spastic paraplegia with thin corpus callosum. Mov Disord. (2007) 22:1556–62. doi: 10.1002/mds.21480
13. Pensato V, Castellotti B, Gellera C, Pareyson D, Ciano C, Nanetti L, et al. Overlapping phenotypes in complex spastic paraplegias SPG11, SPG15, SPG35 and SPG48. Brain (2014)137:1907–20. doi: 10.1093/brain/awu121
14. Orlacchio A, Montieri P, Babalini C, Gaudiello F, Bernardi G, Kawarai T. Late-onset hereditary spastic paraplegia with thin corpus callosum caused by a new SPG3A mutation. J Neurol. (2011) 258:1361–3. doi: 10.1007/s00415-011-5934-z
15. Crimella C, Arnoldi A, Crippa F, Mostacciuolo ML, Boaretto F, Sironi M, et al. Point mutations and a large intragenic deletion in SPG11 in complicated spastic paraplegia without thin corpus callosum. J Med Genet. (2009) 46:345–51. doi: 10.1136/jmg.2008.063321
16. Riverol M, Samaranch L, Pascual B, Pastor P, Irigoyen J, Pastor M, et al. Forceps minor region signal abnormality “Ears of the Lynx”: an early MRI finding in spastic paraparesis with thin corpus callosum and mutations in the Spatacsin Gene(SPG11)on chromosome 15. J Neuroimaging (2009) 19:52–60. doi: 10.1111/j.1552-6569.2008.00327.x
17. Schrander-Stumpel C, Legius E, Fryns JP, Cassiman JJ. MASA syndrome: new clinical features and linkage analysis using DNA probes. J Med Genet. (1990) 27:688–92. doi: 10.1136/jmg.27.11.688
18. Scuderi C, Fichera M, Calabrese G, Elia M, Amato C, Savio M, et al. Posterior fossa abnormalities in hereditary spastic paraparesis with spastin mutations. J Neurol Neurosurg Psychiatry (2009) 80:440–3. doi: 10.1136/jnnp.2008.154807
19. Orlacchio A, Gaudiello F, Totaro A, Floris R, St George-Hyslop PH, Bernardi G, et al. A new SPG4 mutation in a variant form of spastic paraplegia with congenital arachnoid cysts. Neurology (2004) 62:1875–8. doi: 10.1212/01.WNL.0000125324.32082.D9
20. Svenstrup K, Giraud G, Boespflug-Tanguy O, Danielsen ER, Thomsen C, Rasmussen K, et al. Hereditary spastic paraplegia caused by the PLP1 'rumpshaker mutation'. J Neurol Neurosurg Psychiatry (2010) 81:666–72. doi: 10.1136/jnnp.2009.180315
21. Rubegni A, Battisti C, Tessa A, Cerase A, Doccini S, Malandrini A, et al. SPG2 mimicking multiple sclerosis in a family identified using next generation sequencing. J Neurol Sci. (2017) 375:198–202. doi: 10.1016/j.jns.2017.01.069
22. Biancheri R, Ciccolella M, Rossi A, Tessa A, Cassandrini D, Minetti C, et al. White matter lesions in spastic paraplegia with mutations in SPG5/CYP7B1. Neuromuscul Disord. (2009) 19:62–5. doi: 10.1016/j.nmd.2008.10.009
23. Simpson MA, Cross H, Proukakis C, Pryde A, Hershberger R, Chatonnet A, et al. Maspardin is mutated in mast syndrome, a complicated form of hereditary spastic paraplegia associated with dementia. Am J Hum Genet. (2003) 73:1147–56. doi: 10.1086/379522
24. Pedroso JL, Handfas BW, Abrahão A, Kok F, Barsottini OG, Oliveira AS. Fatty acid 2-hydroxylase deficiency: clinical features and brain iron accumulation. Neurology (2015) 84:960–1. doi: 10.1212/WNL.0000000000001316
25. Hedera P. Hereditary myelopathies. Continuum. (2018) 24:523–50. doi: 10.1212/CON.0000000000000580
26. Landouré G, Zhu PP, Lourenço CM, Johnson JO, Toro C, Bricceno KV, et al. Hereditary Spastic Paraplegia Type 43 (SPG43) is caused by mutation in c19orf12. Hum Mut. (2013) 34:1357–60. doi: 10.1002/humu.22378
27. Dard R, Meyniel C, Touitou V, Stevanin G, Lamari F, Durr A, et al. Mutations in DDHD1, encoding a phospholipase A1, is a novel cause of retinopathy and neurodegeneration with brain iron accumulation. Eur J Med Genet. (2017) 60:639–42. doi: 10.1016/j.ejmg.2017.08.015
28. Pedroso JL, Vale TC, Bueno FL, Marussi VHR, Amaral LLFD, França MC, et al. SPG7 with parkinsonism responsive to levodopa and dopaminergic deficit. Parkinsonism Relat Disord. (2018) 47:88–90. doi: 10.1016/j.parkreldis.2017.12.004
29. Anheim M, Lagier-Tourenne C, Stevanin G, Fleury M, Durr A, Namer IJ, et al. SPG11 spastic paraplegia. A new cause of juvenile parkinsonism. J Neurol. (2009) 256:104–8. doi: 10.1007/s00415-009-0083-3
30. Klebe S, Depienne C, Gerber S, Challe G, Anheim M, Charles P, et al. Spastic paraplegia gene 7 in patients with spasticity and/or optic neuropathy. Brain (2012) 135:2980–93. doi: 10.1093/brain/aws240
31. Teive HAG, Iwamoto FM, Camargo CH, Lopes-Cendes I, Werneck LC. Machado-Joseph disease versus hereditary spastic paraplegia: case report. Arq Neuro Psiquiatr. (2001) 59:809–11. doi: 10.1590/S0004-282X2001000500030
32. Synofzik M, Soehn AS, Gburek-Augustat J, Schicks J, Karle KN, Schüle R, et al. Autosomal recessive spastic ataxia of Charlevoix Saguenay (ARSACS): expanding the genetic, clinical and imaging spectrum. Orphanet J Rare Dis. (2013) 8:41. doi: 10.1186/1750-1172-8-41
33. Androdias G, Vukusic S, Gignoux L, Boespflug-Tanguy O, Acquaviva C, Zabot MT, et al. Leukodystrophy with a cerebellar cystic aspect and intracranial atherosclerosis: an atypical presentation of cerebrotendinous xanthomatosis. J Neurol. (2011) 259:364–66. doi: 10.1007/s00415-011-6167-x
34. De Stefano N, Dotti MT, Mortilla M, Federico A. Magnetic resonance imaging and spectroscopic changes in brains of patients with cerebrotendinous xanthomatosis. Brain (2001) 124:121–31. doi: 10.1093/brain/124.1.121
35. de Beer M, Engelen M, van Geel BM. Frequent occurrence of cerebral demyelination in adrenomyeloneuropathy. Neurology (2014) 83:2227–31. doi: 10.1212/WNL.0000000000001074
36. Laurencikas E, Gavhed D, Stålemark H, van't Hooft I, Prayer D, Grois N, et al. Incidence and pattern of radiological central nervous system Langerhans cell histiocytosis in children: a population based study. Pediatr Blood Cancer (2011) 56:250–7. doi: 10.1002/pbc.22791
37. Orlacchio A, Patrono C, Gaudiello F, Rocchi C, Moschella V, Floris R, et al. Silver syndrome variant of hereditary spastic paraplegia: a locus to 4p and allelism with SPG4. Neurology (2008) 70:1959–66. doi: 10.1212/01.wnl.0000294330.27058.61
38. Stromillo ML, Malandrini A, Dotti MT, Battaglini M, Borgogni F, Tessa A, et al. Structural and metabolic damage in brains of patients with SPG11-related spastic paraplegia as detected by quantitative MRI. J Neurol. (2011) 258:2240–7. doi: 10.1007/s00415-011-6106-x
39. Tomberg T, Braschinsky M, Rannikmäe K, Kepler J, Kepler K, Kõrv J, et al. Functional MRI of the cortical sensorimotor system in patients with hereditary spastic paraplegia. Spinal Cord (2012) 50:885–90. doi: 10.1038/sc.2012.70
40. França MC Jr, Yasuda CL, Pereira FRS, D'Abreu A, Lopes-Ramos CM, Rosa MV, et al. White and grey matter abnormalities in patients withSPG11mutations. J Neurol Neurosurg Psychiatry (2012) 83:828–33. doi: 10.1136/jnnp-2011-300129
41. Oguz KK, Sanverdi E, Has A, Temucin C, Turk S, Doerschner K. Tract-based spatial statistics of diffusion tensor imaging in hereditary spastic paraplegia with thin corpus callosum reveals widespread white matter changes. Diagn Interv Radiol. (2012) 19:181–6. doi: 10.5152/dir.2013.046
42. Rezende TJR, Albuquerque MD, Lamas GM, Martinez ARM, Campos BM, Casseb RF, et al. Multimodal MRI-based study in patients with SPG4 mutations. PloS ONE (2015) 10:e0117666. doi: 10.1371/journal.pone.0117666
43. Agosta F, Scarlato M, Spinelli EG, Canu E, Benedetti S, Bassi MT, et al. Hereditary spastic paraplegia: beyond clinical phenotypes toward a unified pattern of central nervous system damage. Radiology (2015) 276:207–18. doi: 10.1148/radiol.14141715
44. Aghakhanyan G, Martinuzzi A, Frijia F, Vavla M, Hlavata H, Baratto A, et al. Brain white matter involvement in hereditary spastic paraplegias: analysis with multiple diffusion tensor indices. Am J Neuroradiol. (2014) 35:1533–8. doi: 10.3174/ajnr.A3897
45. Martinuzzi A, Montanaro D, Vavla M, Paparella G, Bonanni P, Musumeci O, et al. Clinical and paraclinical indicators of motor system impairment in hereditary spastic paraplegia: a pilot study. PloS ONE (2016) 11:e0153283. doi: 10.1371/journal.pone.0153283
46. Liao X, Huang M, Xing W, Wu X, Liao W, Wang X, et al. Resting state fMRI studies in SPG4-linked hereditary spastic paraplegia. J Neurol Sci. (2018) 384:1–6. doi: 10.1016/j.jns.2017.10.048
47. Koritnik B, Azam S, Knific J, Zidar J. Functional changes of the cortical motor system in hereditary spastic paraparesis. Acta Neurol Scand. (2009) 120:182–90. doi: 10.1111/j.1600-0404.2008.01143.x
48. Mc Monagle P, Byrne P, Burke T, Parfrey N, Hutchinson M. Clinical and pathologic findings in hereditary spastic paraparesis with spastin mutation. Neurology (2001) 56:139. doi: 10.1212/WNL.56.1.139
49. Kuru S, Sakai M, Konagaya M, Yoshida M, Hashizume Y. Autopsy case of hereditary spastic paraplegia with thin corpus callosum showing severe gliosis in the cerebral white matter. Neuropathology (2005) 25:346–52. doi: 10.1111/j.1440-1789.2005.00620.x
50. Murphy S, Gorman G, Beetz C, Byrne P, Dytko M, McMonagle P, et al. Dementia in SPG4 hereditary spastic paraplegia: clinical, genetic, and neuropathologic evidence. Neurology (2009) 73:378–84. doi: 10.1212/WNL.0b013e3181b04c6c
51. Duning T, Warnecke T, Schirmacher A, Schiffbauer H, Lohmann H, Mohammadi S, et al. Specific pattern of early white-matter changes in pure hereditary spastic paraplegia. Mov Disord. (2010) 25:1986–92. doi: 10.1002/mds.23211
52. Karle KN, Schüle R, Klebe S, Otto S, Frischholz C, Liepelt-Scarfone I, et al. Electrophysiological characterisation of motor and sensory tracts in patients with hereditary spastic paraplegia (HSP). Orphanet J Rare Dis. (2013) 8:158. doi: 10.1186/1750-1172-8-158
53. Nielsen JE, Johnsen B, Koefoed P, Scheuer KH, Grønbech-Jensen M, Law I, et al. Hereditary spastic paraplegia with cerebellar ataxia: a complex phenotype associated with a new SPG4 gene mutation. Eur J Neurol. (2004) 11:817–24. doi: 10.1111/j.1468-1331.2004.00888.x
54. Orlacchio A, Kawarai T, Gaudiello F, Totaro A, Schillaci O, Stefani A, et al. Clinical and genetic study of a large SPG4 Italian family. Mov Disord. (2005) 20:1055–9. doi: 10.1002/mds.20494
55. Scheuer KH, Nielsen JE, Krabbe K, Simonsen C, Koefoed P, Sørensen SA, et al. Reduced regional cerebral blood flow in SPG4-linked hereditary spastic paraplegia. J Neurol Sci. (2005) 235:23–32. doi: 10.1016/j.jns.2005.03.051
56. Hehr U, Bauer P, Winner B, Schule R, Olmez A, Koehler W, et al. Long-term course and mutational spectrum of spatacsin-linked spastic paraplegia. Ann Neurol. (2007) 62:656–65. doi: 10.1002/ana.21310
57. Samaranch L, Riverol M, Masdeu JC, Lorenzo E, Vidal-Taboada JM, Irigoyen J, et al. SPG11 compound mutations in spastic paraparesis with thin corpus callosum. Neurology (2008) 71:332–6. doi: 10.1212/01.wnl.0000319646.23052.d1
58. Orlén H, Melberg A, Raininko R, Kumlien E, Entesarian M, Söderberg P, et al. SPG11 mutations cause Kjellin syndrome, a hereditary spastic paraplegia with thin corpus callosum and central retinal degeneration. Am J Med Genet B Neuropsychiatr Genet. (2009) 150B:984–92. doi: 10.1002/ajmg.b.30928
59. Criscuolo C, Filla A, Coppola G, Rinaldi C, Carbone R, Pinto S, et al. Two novel CYP7B1 mutations in Italian families with SPG5: a clinical and genetic study. J Neurol. (2009) 256:1252–7. doi: 10.1007/s00415-009-5109-3
60. Goizet C, Boukhris A, Maltete D, Guyant-Maréchal L, Truchetto J, Mundwiller E, et al. SPG15 is the second most common cause of hereditary spastic paraplegia with thin corpus callosum. Neurology (2009) 73:1111–9. doi: 10.1212/WNL.0b013e3181bacf59
61. Terada T, Kono S, Ouchi Y, Yoshida K, Hamaya Y, Kanaoka S, et al. SPG3A-linked hereditary spastic paraplegia associated with cerebral glucose hypometabolism. Ann Nucl Med. (2013) 27:303–8. doi: 10.1007/s12149-012-0673-5
62. Ma J, Xiong L, Chang Y, Jing X, Huang W, Hu B, et al. Novel mutations c.[5121_5122insAG]+[6859C>T] of the SPG11 gene associated with cerebellum hypometabolism in a Chinese case of hereditary spastic paraplegia with thin corpus callosum. Parkinsonism Relat Disord. (2014) 20:256–9. doi: 10.1016/j.parkreldis.2013.11.004
63. Faber I, Martinez ARM, Martins CR Jr, Maia ML, Souza JP, Lourenço CM, et al. SPG11-related parkinsonism: clinical profile, molecular imaging and L-dopa response. Mov Disord. (2018) 33:1650–6. doi: 10.1002/mds.27491
64. Pizzini F, Fatemi AS, Barker PB, Nagae-Poetscher LM, Horská A, Zimmerman AW, et al. Proton MR spectroscopic imaging in Pelizaeus-Merzbacher disease. AJNR Am J Neuroradiol. (2003) 24:1683–9. doi: 10.1177/197140090301600320
65. Erichsen AK, Server A, Landrø NI, Sandvik L, Tallaksen CM. Proton magnetic resonance spectroscopy and cognition in patients with spastin mutations. J Neurol Sci. (2009) 277:124–9. doi: 10.1016/j.jns.2008.10.030
66. Schuurs-Hoeijmakers JH, Geraghty MT, Kamsteeg EJ, Ben-Salem S, de Bot ST, Nijhof B, et al. Mutations in DDHD2, encoding an intracellular phospholipase A(1), cause a recessive form of complex hereditary spastic paraplegia. Am J Hum Genet. (2012) 91:1073–81. doi: 10.1016/j.ajhg.2012.10.017
67. Roos P, Svenstrup K, Danielsen ER, Thomsen C, Nielsen JE. CYP7B1: novel mutations and magnetic resonance spectroscopy abnormalities in hereditary spastic paraplegia type 5A. Acta Neurol Scand. (2014) 129:330–4. doi: 10.1111/ane.12188
68. Fraidakis MJ, Brunetti M, Blackstone C, Filippi M, Chiò A. Novel compound heterozygous spatacsin mutations in a Greek kindred with hereditary spastic paraplegia SPG11 and dementia. Neurodegener Dis. (2016) 16:373–81. doi: 10.1159/000444715
69. Schneider-Gold C, Dekomien G, Regensburger M, Schneider R, Trampe N, Krogias C, et al. Monozygotic twins with a new compound heterozygous SPG11 mutation and different disease expression. J Neurol Sci. (2017) 381:265–8. doi: 10.1016/j.jns.2017.09.005
70. Sperfeld AD, Baumgartner A, Kassubek J. Magnetic resonance investigation of the upper spinal cord in pure and complicated hereditary spastic paraparesis. Eur Neurol. (2005) 54:181–5. doi: 10.1159/000090294
71. Koeppen AH, Becker AB, Qian J, Feustel PJ. Friedreich Ataxia: hypoplasia of spinal cord and dorsal root ganglia. J Neuropathol Exp Neurol. (2017) 76:101–8. doi: 10.1093/jnen/nlw111
72. Mascalchi M, Bianchi A, Ciulli S, Ginestroni A, Aiello M, Dotti MT, et al. Lower medulla hypoplasia in Friedreich ataxia: MR imaging confirmation 140 years later. J Neurol. (2017) 264:1526–8. doi: 10.1007/s00415-017-8542-8
73. Schüle R, Holland-Letz T, Klimpe S, Kassubek J, Klopstock T, Mall V, et al. The Spastic Paraplegia Rating Scale (SPRS): a reliable and valid measure of disease severity. Neurology (2006) 67:430–4. doi: 10.1212/01.wnl.0000228242.53336.90
74. Müller HP, Turner MR, Grosskreutz J, Abrahams S, Bede P, Govind V, et al. A large-scale multicentre cerebral diffusion tensor imaging study in amyotrophic lateral sclerosis. J Neurol Neurosurg Psychiatry (2016) 87:570–9. doi: 10.1136/jnnp-2015-311952
Keywords: MRI, hereditary spastic paraplegia, diagnosis, DTI, spinal cord
Citation: da Graça FF, de Rezende TJR, Vasconcellos LFR, Pedroso JL, Barsottini OGP and França MC Jr (2019) Neuroimaging in Hereditary Spastic Paraplegias: Current Use and Future Perspectives. Front. Neurol. 9:1117. doi: 10.3389/fneur.2018.01117
Received: 31 July 2018; Accepted: 05 December 2018;
Published: 16 January 2019.
Edited by:
Giuseppe De Michele, University of Naples Federico II, ItalyReviewed by:
Filippo M. Santorelli, Fondazione Stella Maris (IRCCS), ItalyCopyright © 2019 da Graça, de Rezende, Vasconcellos, Pedroso, Barsottini and França. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marcondes C. França Jr., bWNmcmFuY2FqckB1b2wuY29tLmJy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.