 Jakub Sawicki
Jakub Sawicki- 1Potsdam Institute for Climate Impact Research, Potsdam, Germany
- 2Institut für Mathematik, Technische Universität Berlin, Berlin, Germany
- 3Institut für Theoretische Physik, Technische Universität Berlin, Berlin, Germany
- 4Institut LOESER, Leipzig, Germany
- 5Bernstein Center for Computational Neuroscience Berlin, Humboldt-Universität, Berlin, Germany
In this study, we provide a dynamical systems perspective to the modelling of pathological states induced by tumors or infection. A unified disease model is established using the innate immune system as the reference point. We propose a two-layer network model for carcinogenesis and sepsis based upon the interaction of parenchymal cells and immune cells via cytokines, and the co-evolutionary dynamics of parenchymal, immune cells, and cytokines. Our aim is to show that the complex cellular cooperation between parenchyma and stroma (immune layer) in the physiological and pathological case can be qualitatively and functionally described by a simple paradigmatic model of phase oscillators. By this, we explain carcinogenesis, tumor progression, and sepsis by destabilization of the healthy homeostatic state (frequency synchronized), and emergence of a pathological state (desynchronized or multifrequency cluster). The coupled dynamics of parenchymal cells (metabolism) and nonspecific immune cells (reaction of innate immune system) are represented by nodes of a duplex layer. The cytokine interaction is modeled by adaptive coupling weights between the nodes representing the immune cells (with fast adaptation time scale) and the parenchymal cells (slow adaptation time scale) and between the pairs of parenchymal and immune cells in the duplex network (fixed bidirectional coupling). Thereby, carcinogenesis, organ dysfunction in sepsis, and recurrence risk can be described in a correct functional context.
1 Introduction
Tumors and sepsis are diseases of different genesis. They have very different time scales and the therapies are completely different. But the age incidence, the risk factors, the outcome, and the temporal recurrence behavior are similar. This justifies the attempt to describe tumor disease and sepsis with a unified disease model. A shift of the paradigm is proposed by choosing the nonspecific innate immune system as the reference point for both diseases Löser (2020). The innate immune system interacts with mutant cells and pathogens. Its role is to maintain the integrity of the organism by actively eliminating foreign organisms, degrading the organism’s own damaged cells, and activating and coordinating wound healing. It can ward off about 99% of all infections Bomans et al. (2018); it arose with the beginning of multicellular life, and has grown and protected life to this day Male et al. (2012); Rich and Chaplin (2019). It is ubiquitously present in the organism, is responsive without significant dead time, and is based on dynamically balanced activator-inhibitor mechanisms. Its regulation is essentially decentrally organized. The innate immune system includes humoral and cellular components, the endothelium, and the tissue stroma. With the multitude of components interacting in the innate immune system, it is organized in a complex way and has a broad response spectrum.
The innate immune system is not completely resistant to disturbances; it can be brought to dysregulation. Tumor disease and sepsis are two prominent examples of this behavior. Although tumor disease and sepsis correspond to different medical conditions, after the initial stages a relatively uniform course develops Weinberg (2014), Singer et al. (2016). The variability of tumor disease results from the initial genetic state of the tumor cells, their subsequent mutations, epithelial-mesenchymal transition, and interactions with the innate immune system Longo (2011). Recently, in case of cancer the crosstalk of tumor cells with immune cells has been investigated in more detail Zhang et al. (2021). In sepsis, the innate immune system is activated by infection and the clinical course is determined by the individual patient’s initial condition. In both cases, the innate immune system can be brought to dysregulation and develop its own clinical picture such as cachexia, coagulation disorders, and organ failure. In the case of tumor disease, this is compounded by space-occupying lesions, tissue invasion and destruction through the nonphysiological production of proliferation factors, cytokines, and chemokines. Possibly, not only the cytokine concentration, the cytokine mix but also the gradient of their increase is responsible for the extent of dysregulation of the innate immune system and thus for the disease consequences Morán et al. (2013), Altan-Bonnet and Mukherjee (2019).
The unified disease model with the innate immune system as reference point is the basis for our modeling approach in terms of nonlinear dynamics of complex networks. Note that this is not a biochemical or genetic or cellular or tissue model of carcogenesis, as has been reviewed elsewhere Vineis et al. (2010), but it rather describes tumor and sepsis and recurrence risk in a dynamical functional context. Complex networks are an ubiquitous paradigm in nature and technology, with a wide field of applications ranging from physics, chemistry, biology, neuroscience, to engineering, and socio-economic systems. Of particular interest are adaptive networks, where the connectivity changes in time, for instance, in chemical or biochemical systems Jain and Krishna (2001), where the reaction rates adapt dynamically depending on the variables of the system, or in neuronal synaptic plasticity Markram et al. (1997), Abbott and Nelson (2000), Meisel and Gross (2009), Lücken et al. (2016), in epidemics Gross et al. (2006), and in biological or social systems Gross and Blasius (2008). Another focus of recent research in network science are multilayer networks, which are systems interconnected through different types of links De Domenico et al. (2013), Boccaletti et al. (2014), Kivelä et al. (2014), De Domenico et al. (2015). A special case of multilayer networks are multiplex topologies, where each layer contains the same set of nodes, and only pairwise connections between corresponding nodes from neighboring layers exist Zhang et al. (2015), Maksimenko et al. (2016), Andrzejak et al. (2017), Leyva et al. (2017), Sawicki et al. (2018), Nikitin et al. (2019), Omelchenko et al. (2019), Rybalova et al. (2019), Drauschke et al. (2020), Berner et al. (2021a), Sawicki et al. (2021), Shepelev et al. (2021).
Synchronization is an important feature of the dynamics in networks of coupled nonlinear oscillators Pikovsky et al. (2001), Strogatz (2001), Albert and Barabási (2002), Newman (2003), Boccaletti et al. (2018), Berner (2021). Various synchronization patterns are known such as cluster synchronization where the network splits into groups of synchronous elements Dahms et al. (2012), or partial synchronization patterns such as chimera states where the system splits into coexisting domains of coherent (synchronized) and incoherent (desynchronized) states Kuramoto and Battogtokh (2002), Abrams and Strogatz (2004), Panaggio and Abrams (2015), Sawicki (2019), Schöll (2020), Schöll et al. (2020). These patterns were also explored in adaptive networks Seliger et al. (2002), Aoki and Aoyagi (2009), Timms and English (2014), Kasatkin et al. (2017), Berner et al. (2019b), Berner et al. (2021b), Berner et al. (2021d), and in particular in adaptive two-layer networks of phase oscillators Kasatkin and Nekorkin (2018), Berner et al. (2020b). Moreover, the role of synchronization is an important aspect in the field of network physiology, where multi-component physiological systems continuously interact in an integrated network to coordinate their functions Bashan et al. (2012), Ivanov and Bartsch (2014), Bartsch et al. (2015), Moorman et al. (2016), Lin et al. (2016). There are empirical studies dealing with the structural organization and functional complexity of human organism which demonstrate phase-synchronization as well as phase transitions Chen et al. (2006), Xu et al. (2006), Ivanov et al. (2009), Bartsch et al. (2012) between different modes of synchronization in real physiological systems. In case of complex diseases, the progression from a healthy to sick state can be abrupt and may cause a critical transition Chen et al. (2012), Liu et al. (2012), Liu et al. (2013a), Liu et al. (2013b).
In this article, we propose a two-layer network model for carcinogenesis and sepsis based upon the interaction of parenchymal cells and immune cells via cytokines and the co-evolutionary dynamics of parenchymal, immune cells, and cytokines. Parenchyma is the bulk of functional substance in an organ, in contrast to the stroma, which refers to the structural tissue of organs or structures. In many organs the parenchyma consists of epithelial cells. We stress that our model is not a detailed model of organs or of biochemical processes but a functional model of dynamic interactions. Thus, cytokines are not modeled in terms of concentrations but rather in terms of the cytokine-mediated information flow between nodes within each layer and between the parenchymal and the immune layer, describing the cytokine activity. In the following, we refer to the stroma as the immune layer. The article is organized as follows: In Section 2, we provide a brief overview of the physiology of tumor disease and sepsis. The functional network model is introduced in Section 3. Here, we discuss all variables, parameters, and their physiological meaning. Moreover, the methods and measures used for the subsequent numerical analysis are introduced and highlighted in their physiological context. In Section 4, we discuss various dynamical scenarios of the model simulations of tumor disease that are observed in the presence of pathological cells. Section 5 presents analogous computer simulations for sepsis. The results are summarized in Section 6.
2 Physiological Description
2.1 Initial Situation
The function of the innate immune system is ensured in a site-independent manner by the coordinated activity of humoral and cellular components. Coordination also occurs via cytokines. With new cytokine sources building up over individual lifetimes, the stability of regulatory behavior in the innate immune system changes. Contributing systemic factors include inflammaging Franceschi and Campisi (2014), Calder et al. (2017), activation of the coagulation system Tragl (1999), Franceschi and Campisi (2014), increase of body fat Fasshauer et al. (2004), Gaillard (2007), Chovatiya and Medzhitov (2014), lack of exercise Elisia et al. (2017), Fulop et al. (2018), age-related normal fibrosis Beneke (1971), Nemetschek (1971), chronic inflammation as local factors Virchow (1978), or smoking via initiation of local hypoxia Brunkhorst et al. (2018). The innate immune system exhibits pro-inflammatory activation. The number of individual variables suggest a wide range of activation levels, which still depend on cytokine gene polymorphisms, among other factors. Physiological cytokine production and additional pro-inflammatory cytokine activity (pattern, concentration, emitters involved, gradient of increase) determine the activation level of the innate immune system.
2.2 Tumor Disease
In tumor disease, mutant cells interact with macrophages of the innate immune system localized in the stroma. Mutated cells are genetically altered and genetically unstable parenchymal cells. They are a new cell entity with altered regulatory behavior. In interaction with the normal stroma (immune layer), e.g. in young healthy individuals, mutant cells do not find survival conditions. They persist silently or trigger apoptosis via tissue surveillance Löser (2018). Mutant cells in an area pre-damaged by chronic inflammation receive cytokines from the activated stroma, undergo an epithelial-mesenchymal transition Lamouille et al. (2014), Chockley and Keshamouni (2016), can proliferate, disseminate, and reprogram normal macrophages into tumor-associated macrophages Wu and Zhou (2009), Karlsson et al. (2017), Mantovani et al. (2017). They help to ensure oxygen and nutrient supply to the tumor. Tumor cells, tumor-associated macrophages, and normal stromal cells communicate via cytokines. The communication strength depends on tumor cell genetics and stroma activation. It increases with the tumor cell mass and the mass of specific tumor stroma. A mutually activating circular process starts between the tumor cells and the innate immune system. A specific cytokine microclimate is formed around the developing tumor, the size of which grows with the increase in tumor mass. Cytokines enter the lymph nodes with the lymph and enable the tumor cells that have floated there to proliferate. Lymphogenic metastasis begins, continuing downstream in additional lymph nodes according to the same pattern. Organ-specific metastatic patterns are formed Walther (1948). The circulating cytokine concentration increases and triggers the general symptoms of tumor disease such as cachexia Arends et al. (2015) and finally the lethal coagulopathy, lung failure, organ failure or lethal space-occupying lesions and tissue destruction. The postulate of Lewis Thomas Thomas (1972) also applies to tumor disease, according to which the interactions with the organism triggered by the tumor cells are responsible for the disease, i.e., tumor growth, enabling malignant cells to become invasive and destructive, recurrence, and finally induce the lethal general symptoms. Hematogenous metastasis may start when the disseminated tumor cells are adequately supplied with growth factors (cytokines) via further activation of the innate immune system.
2.3 Sepsis
The initial conditions for the transition of an infection into sepsis are analogous to the survival conditions of mutated cells in the tissue, a pre-activated innate immune system. Sepsis is triggered when the innate immune system no longer succeeds in locally fixing invading pathogens, but allows them to enter the organism. There they interact with the already pre-activated innate immune system. The immune system is activated systemically. Antibiosis or surgical sanitation can reduce or eliminate the pathogen load. Depending on the type of reaction, the pro-inflammatory response is stopped or it continues to escalate in varying degrees Seymour et al. (2019), for which also cytokine gene polymorphisms are responsible Majetschak and Schade (2001), Hotchkiss et al. (2016), Thomas (2020a), Thomas (2020b). Pro-inflammatory cytokines act on endothelial cells and the coagulation system. Microthrombi amplify inflammation through secretion of growth factors and cytokines Hotchkiss et al. (2016). The endothelial permeabilization barrier is opened, fluid retention in tissues occurs, blood pressure drops, and must be stabilized with fluid administration Brunkhorst et al. (2018). Prolonged oxygen diffusion pathways and the resulting tissue hypoxia trigger cytokine release causing a “second hit”, resulting in metabolic changes in the parenchyma Bomans et al. (2018). Organ failure, particularly of the kidneys, lungs, and liver is imminent. Endogenous molecules with immunogenic effects, especially mitochondrial DNA Franceschi et al. (2018), released e.g. after trauma, burns, pancreatitis or surgery, can have the same effect as exogenous pathogens.
2.4 Relapse
Tumors and sepsis have a recurrence behavior, the frequency of which correlates with the stage of the primary disease. In both diseases, the activated innate immune system is an initial prerequisite for the primary disease, which is brought to dysregulation in the course of the disease. After tumor removal and elimination of infection, the activation status reduces but remains at least at the level before disease onset. Thus, cytokines are produced for remaining tumor cells as a result of pro-inflammatory activation Gastpar (1982), Eichinger and Lechner (2004), Arends et al. (2015), Lippman (2016), which promote their proliferation and allow the organization of the tumor stroma. The process kinetics is determined by the proliferation rate of tumor cells, the availability of cytokines, and by the ability of tumor cells to organize their own stroma with connection to the blood supply. After clearance of the systemic infection, the innate immune system has an activation status at least equivalent to that before septic shock. The infection itself appears to have triggered irreversible elements of trained immunity Bomans et al. (2018). Surviving patients after septic shock thus die in 50% within the first two post-sepsis years from persistent and secondary nosocomial infections, tumors, or cardiac failure Prescott et al. (2016).
3 Model
3.1 Schematic Model for Tumor Disease and Sepsis
Organic tissue consisting of parenchymal cells and immune cells is shown schematically in Figure 1. We depict the initial and final configurations for tumor disease and sepsis on a tissue element. The tissue element consists of the epithelial parenchyma, the basal membrane, and the stroma. The parenchyma is the organ-specific functional layer. The basal membrane separates the parenchyma and stroma and is made of collagen of type IV. In the stroma, blood supply, lymphatic drainage, and immune response occur. The stroma consists of an extracellular matrix and embedded cells that do not form a solid association. The extracellular matrix is structurally composed of collagen, glycoproteins, proteoglycans, and water. Cells in the stroma are resident fibroblasts and fat cells, and mobile cells (macrophages, mast cells, granulocytes, and plasma cells).
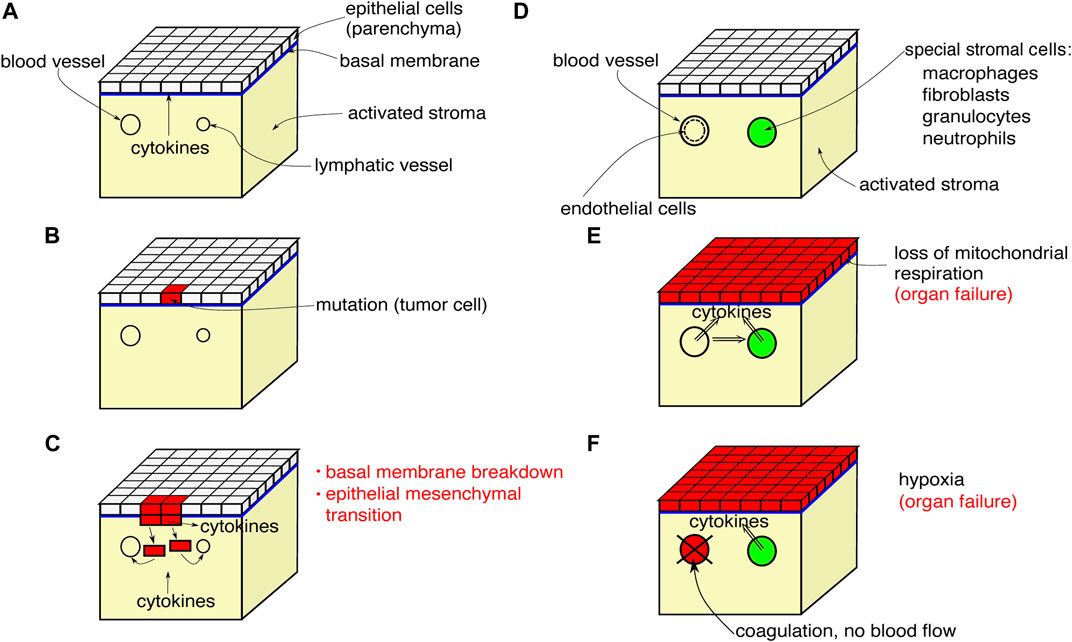
FIGURE 1. Scheme of a tissue element showing the initial progression of tumor disease (A–C), the initial configuration for sepsis (D) and the final configurations found for both tumor disease and sepsis (E),(F).
The initial process of carcinogenesis is shown in Figures 1A–C. In Figure 1A, the parenchyma is normal, the stroma is inflammatorily activated. In Figure 1B, a mutation of a tumor cell has occurred in the parenchyma, the inflammatory activation of the stroma continues. Finally in Figure 1C, the mutant cells proliferate, break through the basal membrane and migrate into the blood and lymphatic vessels, and cytokines are emitted.
Figures 1D–F shows the systemic effects of the final process of tumor disease and sepsis: Figure 1D depicts a tissue element that is not directly affected by tumor, metastasis, or primary inflammation. The stroma exhibits inflammatory activation. In Figure 1E, a systemic cytokine storm is occurring. Cytokine production by the stromal cells is additionally stimulated. The parenchyma responds to the primary and secondary cytokine storm by uncoupling mitochondrial respiration and switching to aerobic glycolysis. The energy supply is no longer sufficient for organ-specific cellular functions and the organ fails. Figure 1F shows a variant of Figure 1E. The cytokine storm activates the endothelium, blood coagulation is activated, oxygen transport breaks down, and hypoxia arises. The stromal cells respond with cytokine release. Both together, the hypoxia and the cytokine release, lead to the collapse of organ-specific cell functions in the parenchyma and the organ fails.
3.2 Dynamical Two-Layer Network Model
The unified disease model with the reference point given by the innate immune system is the basis for our model, which includes disease-specific initial conditions, mutant cells for tumor disease, and infection-driven cytokine dysregulation. A volume element of tissue consisting of parenchyma, basal membrane and stroma is used as a model for tumor disease and sepsis, describing the functional interactions between parenchyma (organ tissue) and stroma (immune layer). We represent the network layer of parenchymal cells (superscript 1) by N phase oscillators
The use of phase oscillators for the functional modeling of the interacting parenchymal cells and immune cells is motivated by the fact that phase oscillator networks are a paradigmatic model for collective coherent and incoherent dynamics. As discussed in detail in Section 3.3, healthy cells and tumor cells differ by their metabolic activity, i.e., tumor cells are less energy-efficient and thus have a faster cellular metabolism, which is reflected in our phase oscillator model by a higher frequency. Thus in the healthy homeostatic equilibrium state all parenchymal cells have the same lower frequency, while the pathological state splits into two clusters with different frequencies, healthy, and unhealthy, i.e., a multifrequency cluster. The healthy state is assumed to be characterized by regular periodic, fully synchronized dynamics of the phase oscillators, i.e., all cells show the same collective frequency of cellular metabolism. The pathological state is described by multifrequency clusters with different frequencies, i.e., the pathological cells in the parenchyma attain a higher frequency, while the healthy cells are still frequency-synchronized with the “healthy” frequency. This loss of synchrony reflects the fact that in case of tumor disease, the malignant mutation basically leads to a loss of performance of the parenchymal cells. As a consequence, they are no longer fully coordinated, which leads to a loss of proliferation and apoptosis control Longo (2011), Weinberg (2014). Such a pathological condition leads also to an alteration of the metabolic activity of the immune cells Coussens and Werb (2002), Heerboth et al. (2015), Chockley and Keshamouni (2016), Porporato (2016), Razak et al. (2018), Greten and Grivennikov (2019). In case of sepsis, i.e., a systemic inflammation, cells belonging to the innate immune system produce cytokines in an unregulated way affecting the parenchyma or microcirculation leading to organ failure Singer et al. (2016), Matsumoto et al. (2018) which is also associated in our model with the loss of synchrony.
A general multiplex network with two layers each consisting of N identical adaptively coupled phase oscillators is described by
where
The phase lag parameter β of the adaptation function
3.3 Methodology and Measures
Heterogeneous dynamics, e.g., multifrequency clusters and tumor growth, may arise through the dynamic interaction of parenchymal cells, immune cells, and localized cytokine activity in a self-organized self-adaptive manner, even if the system parameters in the layers are chosen uniformly, i.e., homogeneous Berner et al. (2019a), Berner et al. (2020a). Heterogeneity can enter the parameters of oscillator networks in various ways. Very prominent are heterogeneities in the natural frequencies of the oscillators or in the connectivity structure Acebrón et al. (2005). Heterogeneous frequencies ωi are used to model pathological parenchymal cells which occur by spontaneous random mutations. We assume that all healthy cells possess the same natural frequency ωi = ωh, and a fraction r of randomly chosen pathological cells possess the natural frequency ωi = ωp. The characterization of tumor cells via their metabolic properties Longo (2011), Weinberg (2014) has revealed a difference in the metabolic activity between tumor and healthy cells Warburg et al. (1924). Warburg demonstrated that cells after a malignant mutation obtain their energy via aerobic glycolysis, which provides only 4 mol ATP/mol glucose, in contrast to normal cells whose metabolism is based upon mitochondrial breathing yielding 36 mol ATP/mol glucose. Thus tumor cells have a metabolic efficiency of only about 10% of healthy cells, and therefore they need more glucose. Other general differences in metabolic activity between tumor cells and healthy cells include proliferation rate, apoptosis rate, initiation of angiogenesis, tissue invasion, or metastatic capacity. Here, quantitative differences exist between different organ tumors. Ultimately, any change in metabolic performance, regardless of which metabolic branch is affected (structural, energetic, cell division, or special metabolism), can be conceived as a frequency change.
Using numerical simulations, we study whether the healthy state (frequency synchronized) is persistent also in the presence of a few pathological cells. Under certain conditions depending on various parameters (age, inflammaging, chronic inflammation, other basic diseases, obesity, smoking, lack of exercise), an unregulated cytokine expression and hence a possibly lethal tumor can occur. In these cases, the healthy (synchronized) state is not persistent anymore against the perturbation by the heterogeneity (tumor cells). For our study, the cytokine adaptivity parameter (which we call age parameter) β and the fraction r are considered as the main model parameters to account for the various system conditions. In the case of sepsis, we introduce a fixed initial perturbation of the cytokine activity in the immune layer representing a systemic immune response, while keeping the natural frequencies uniform. Similar to the tumor disease, we study the effect of this initial system perturbation on the emergence of the healthy state in dependence of the age parameter β. For our simulations we have used a Runge–Kutta method of order 4 with a fixed stepsize of Δt = 0.05 and simulation time of Ts = 2000 time units where we have discarded the first 1,000 time units to account for transient dynamics.
In order to quantitatively characterize the dynamic collective state of the two-layer network, we introduce several measures. First, we use the mean phase velocities of the oscillators j in both layers μ = 1, 2
with averaging time window T, and the spatially averaged mean phase velocity for each layer
Furthermore, for both layers μ = 1, 2 we calculate the ensemble average sμ (ensemble size NE with ensemble elements E) of the standard deviation
and the ensemble average of the corresponding normalized standard deviation
In systems of adaptively coupled phase oscillators one often encounters frequency-synchronized dynamics. If the frequency is the same, the phases may still exhibit different behavior. They may either be all the same (complete in-phase synchronization) or they may be phase-locked such that each phase oscillator oscillates with the same frequency but a fixed, time-independent phase difference. A special case is a splay state, where the phase differences of all oscillators average out, for instance if the relative phase of the jth oscillator is 2πj/N, j = 1, … , N. In systems of the form Eqs. 1, 2, it is possible to find in-phase synchronization and splay states, and they may be interpreted as different quality of synchronization Berner et al. (2020b), Berner et al. (2021c). In our set-up a splay state is interpreted as a more vulnerable collective state (synchronization of parenchyma and immune layer) where small perturbations can quickly lead to partial or complete desynchronization.
3.4 Physiological Interpretation of Model Parameters
Table 1 gives an overview of the dynamical variables, parameters and measures of the model. In the right column the physiological meaning of all quantities is given in a concise manner. In more detail, the quantities of the mathematical model which are listed in Table 1 have the following physiological correspondence:
1) Variable ϕi mimics the metabolic activity of a parenchymal or immune cell i as a universal oscillatory phase variable. Here our aim is not to describe physiological processes on a detailed biochemical level, e.g., C-reactive protein (CRP) production after inflammatory activation, etc.
2) Variable κij describes the cytokine activity mediating information flow between cells j and i. It stands for the susceptibility of communications between cells via cytokines. Cytokines have pleiotropic properties, i.e., they can exert different, even contrary, effects depending on the initial situation and in combination with other cytokines. Therefore we model them by an adaptive coupling strength. Reaction strength of cytokines is due to individually different gene polymorphisms.
3) Parameter α models the phase lag of the intralayer cell-to-cell coupling. Its origin is the time delay from the activation of a defined metabolic branch until the mediator release; within the inflammatory cascade, characteristic time delays exist for all reaction sub-steps starting from the triggering of the inflammatory reaction, for example, the reaction time of the individual liver cells starting from the input signal until the maximum blood concentration of interleukin IL-6 (2 h) and CRP (12 h) is reached.
4) Parameter β governs the adaptivity (or plasticity) rule of the cytokines. It mimicks a systemic sum parameter which may account for different influences such as physiological changes due to age in the extracellular matrix, inflammaging, systemic and local inflammatory baseline, adiposity, pre-existing illness, physical inactivity, nutritional influence, and others. In case of tumor disease, it can include the malignancy grade of tumor cells. For the sake of brevity, we call this parameter the age parameter.
5) Parameter ω denotes the natural frequency of the basic metabolic activity of a single cell. For instance, it can stand for the minor CRP production in a normal healthy state.
6) Parameters ϵ1 and ϵ2 denote the inverse relaxation times (half-life) of the cytokines in the parenchymal and immune layer, respectively.
7) Parameter r denotes the fraction of mutant (pathological) parenchymal cells in the tissue element, and is implemented in the model as the fraction of cells with a deviating (pathological) natural frequency ωp.
8) Parameter aij denotes the genetically fixed intercellular communication pathways between parenchymal cells, they are wired by fixed cell-to-cell connections. In contrast, communication of parenchymal cells via cytokines and of cells within the immune layer runs in an open communication channel, which is controlled self-adaptively, after Shannon (1948).
9) Parameter σ denotes the interlayer coupling strength between the parenchymal and immune cells. It can be due to a mass transfer, e.g., cytokine expression of macrophages in the immune layer, and signal transfer into the parenchyma: macrophages invade the extracellular matrix and penetrate the basal membrane to the parenchyma.
10) The mean phase velocity
11) The ensemble-averaged standard deviation of the mean phase velocities s1, s2 is a measure which characterizes the inhomogeneity of metabolic activity within the parenchyma and the stroma (immune layer), respectively. It assumes non-zero values if the respective layer is not frequency-synchronized and splits into multifrequency clusters, e.g., a healthy cluster with one frequency and a pathological cluster with another frequency, and thus is a measure of pathogenicity in case of the parenchymal layer, or activation in case of the immune layer.
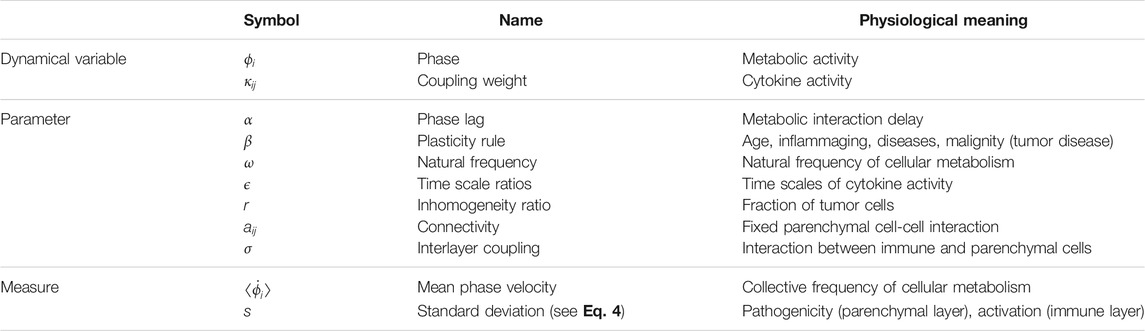
TABLE 1. Transcription table for the dynamical variables, parameters, and measures of the model (superscripts referring to layers μ = 1 and μ = 2 omitted).
4 Tumor Disease
In this section, we present exemplary computer simulations of our model to demonstrate different dynamic scenarios which this model can produce already in its simplest form. The model has not been refined or optimized with respect to the parameters, but our concern here is to display simulations which can describe principally different evolutions and outcomes of tumor disease. We assume N = 200, ϵ1 = 0.03, ϵ2 = 0.3,
Our system exhibits various dynamic patterns resembling healthy and pathological states. The results are depicted in Figure 2, where the mean phase velocities
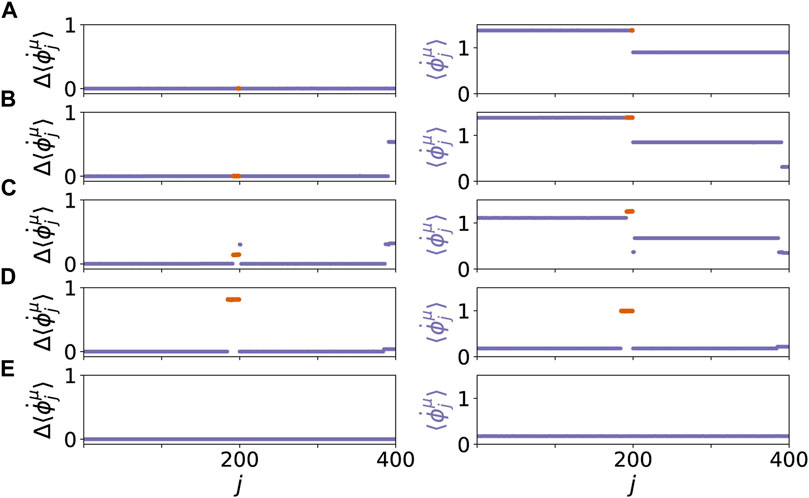
FIGURE 2. Dynamical scenarios for healthy states without clusters and pathological states with multifrequency clusters for different values of β and r. The left panel shows the absolute differences of the mean phase velocities
Figure 3 shows details of these scenario for the same values of age parameter β and tumor size r. The left and right columns show snapshots of the cytokine matrices
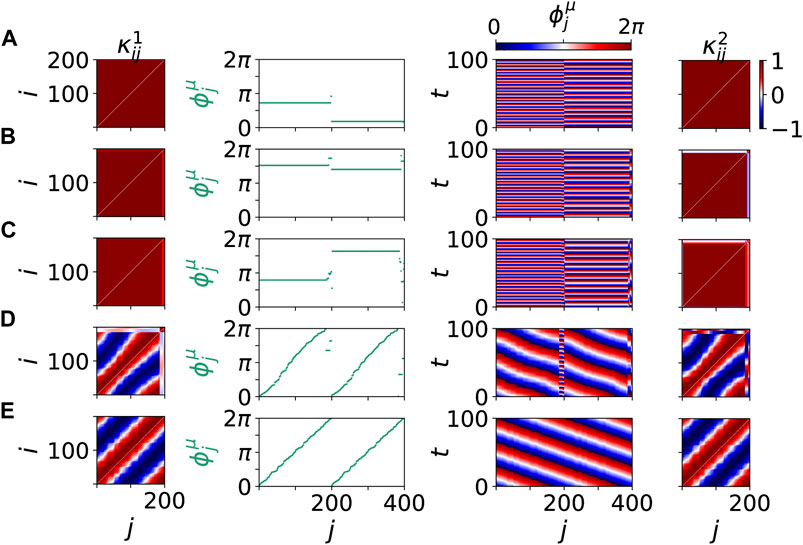
FIGURE 3. Details of dynamical scenarios for healthy states without clusters (A),(B),(E) and pathological states with multifrequency clusters (C),(D) for different values of β and r chosen as in Figure 2. The left and right columns show snapshots of cytokine matrices
A map of regimes in the parameter plane of (β, r) is shown in Figures 4A,B, where the ensemble average sμ of the standard deviation of the mean phase velocities is plotted for the parenchymal and immune layer, respectively. Bright colors correspond to large deviations of the frequencies (mean phase velocities), and hence to pathological two-cluster states. Their enhancement with increasing age parameter and tumor size can be clearly seen. In Figures 4C,D, a comparison between the ensemble average of the standard deviation and the normalized standard deviation is depicted. In comparison to the standard deviation, the normalized standard deviation shows higher values of the ensemble average s1 in case of splay states (see Figure 4C for β > 0.6π). On the contrary, the small bulge of s1 at β = 0.5π is less pronounced. Figure 5 depicts the comparison between the normalized standard deviation of the parenchymal and immune layer. It clearly shows the strong increase of the frequency deviation in the parenchyma associated with the pathogenicity of the tumor with increasing age parameter, while the much smaller bulge in the immune layer is associated with the immune layer activation before the tumor becomes clearly visible.
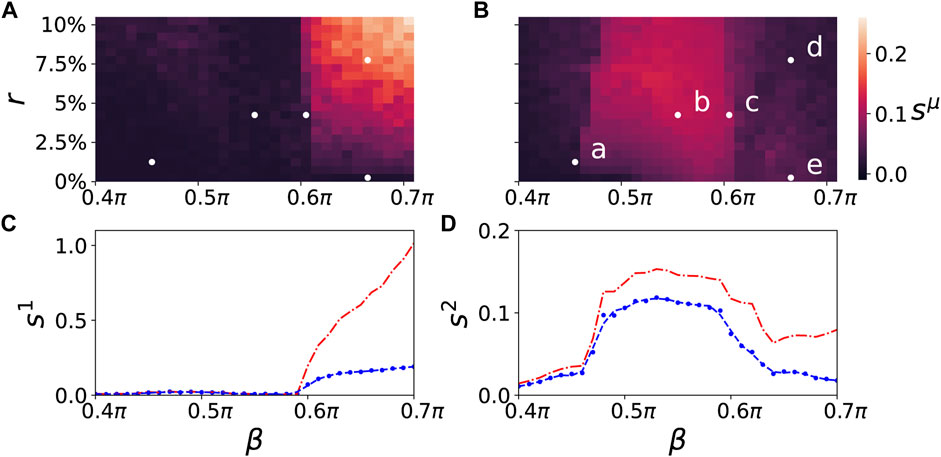
FIGURE 4. Map of regimes: ensemble average sμ of the standard deviation of the mean phase velocities in the parameter plane of age parameter β and tumor size r) (ensemble size is NE = 50) for the parenchymal (A) and immune layer (B), respectively. Bright colors correspond to the formation of multifrequency clusters. The dynamics of the points marked (a)–(e) in the upper right panel are shown in Figures 2, 3A–E. The lower panels depict the ensemble average sμ (blue) in dependence of β for a fixed value of r = 7.5% for the parenchymal (C) and immune layer (D), respectively. The dashed line is a regression curve for an ensemble size of NE = 200. For comparison the ensemble average sμ of the standard deviation (blue curves) and the normalized standard deviation (red curves) of the spatially averaged mean phase velocities is shown in dependence of β for the parenchymal (C) and immune layer (D), respectively. Other simulation parameters as in Figure 2.
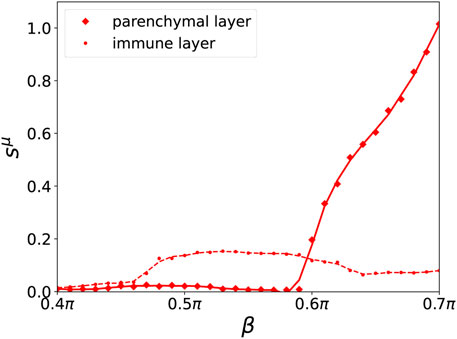
FIGURE 5. Ensemble average sμ of the normalized standard deviation of the mean phase velocities in dependence of β for a fixed value of r = 7.5% for the parenchymal (solid curve, diamonds) and immune layer (broken curve, dots), respectively for an ensemble size of NE = 200. A cubic Savitzky–Golay filter has been applied to smooth the curves. Other simulation parameters as in Figure 2.
5 Sepsis
In our model we focus on the stage of sepsis which is characterized by generalized inflammation. Depending on the individual subject and the local cellular situation of cytokine and fluid influx, either de-escalation of the inflammatory reaction and restoration of homeostasis, or organ failure occurs. Sepsis is usually preceded by a pre-septic perturbation of the parenchyma, e.g., by a wound which is infected by germs. This perturbation is terminated after a while by blocking off the wound by blood coagulation and eventually healing. Under “normal” conditions, the system returns to a healthy state; however, if sepsis occurs, an inflammatory immune response triggered by the infection spreads across the whole body (cytokine storm). As a consequence, the immune activity may invade large parts of the body through blood vessels and lead to severe organ failure and death. Whether sepsis terminates in a septic shock with severe consequences for the patient depends crucially on the ability of the immune and parenchymal system to regain homeostasis. As in the case of the tumor disease, the ability for returning to a healthy state, i.e., a frequency-synchronized state, is subsequently analyzed using the systemic sum parameter (age parameter) β.
For the simulations, we assume that a dysregulation of the cytokine activity in the immune system has already occurred due to a systemic immune response. The dysregulation of the cytokine activity is modeled by an initial perturbation of the cytokine activity matrix
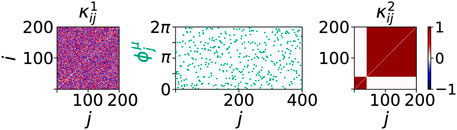
FIGURE 6. Initial conditions of sepsis: Cytokine dysregulation expressed by a cluster structure of the cytokine activity matrix
In the following, we present simulation results for different choices of the age parameter β showing that after an initial cytokine perturbation in the immune layer either the healthy frequency-synchronized state is restored, or the whole system goes to a desynchronized or multifrequency cluster state.
Figure 7 shows five representative dynamical scenarios that are induced by pathological initial cytokine activity in the immune layer. For an age parameter β = 0.5π in Figure 7A, we observe that the system relaxes to a healthy state, i.e., frequency synchronized state, after an initial immune layer perturbation. Hence, the pro-inflammatory response is stopped. A transient of 1,000 time units is discarded before the temporal average over T = 1,000 is taken in the mean phase velocities
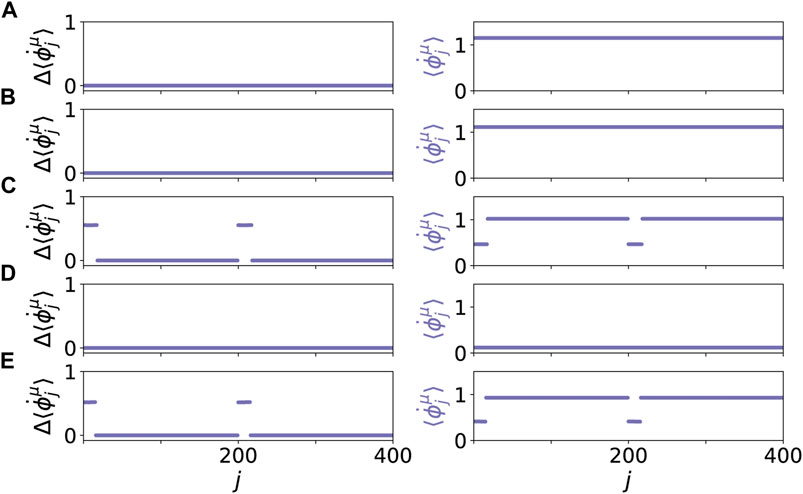
FIGURE 7. Dynamical scenarios for healthy states without clusters and pathological states with multifrequency clusters for different values of β. The left panels show the absolute differences of the mean phase velocities
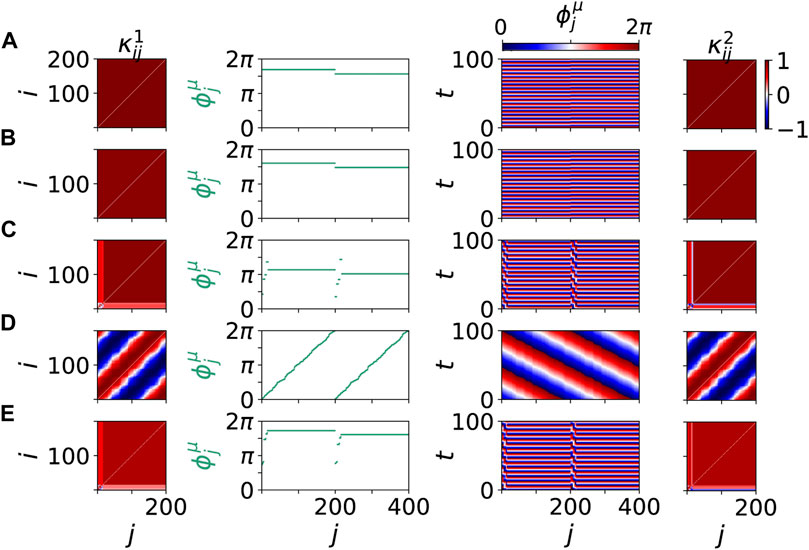
FIGURE 8. Details of dynamical scenarios for healthy states without clusters (A),(B),(D) and pathological states with multifrequency clusters (C),(E) for different values of β chosen as in Figure 7. The left and right columns show snapshots of cytokine activity matrices
For all presented scenarios in Figures 7, 8, we note that the frequencies of the parenchymal and the immune layer are locked. The emergence of the dynamical phenomenon of locking Pikovsky et al. (2001) can be explained by the increased value of the interlayer coupling strength σ compared to the simulation of the tumor disease, see Section 4. Locking can be interpreted as the dynamical manifestation of the physiological observation that the immune system is taking control of the whole system in case of sepsis. For smaller interlayer coupling strength this locking cannot be observed, and hence desynchronization induced by a pathologically activated immune system is not possible.
The dynamical scenarios discussed in this section are very similar to the observations presented for tumor disease. Additionally, in agreement with the results found for tumor disease, the desynchronization of the parenchymal layer due to an initially activated immune layer is observed for higher age parameter β with increasing probability. We note that also here resilient and vulnerable healthy states may coexist with pathological desynchronized states. The increased desynchronization probability with increasing β is reflected in the increasing ensemble average of the normalized standard deviation of the mean phase velocities in Figure 9.
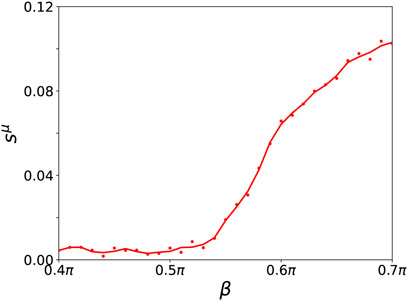
FIGURE 9. Ensemble average sμ of the normalized standard deviation of the mean phase velocities in dependence of β for a fixed value of σ = 1 for the parenchymal and immune layer (both curves coincide) for an ensemble size of NE = 200. A cubic Savitzky-Golay filter has been applied to smooth the curve. Other simulation parameters as in Figure 7.
6 Conclusion
In this article, we have proposed a two-layer network model for carcinogenesis and sepsis based upon the interaction of parenchymal cells and immune cells via cytokines and the co-evolutionary dynamics of parenchymal, immune cells and cytokines. Certain parallels between cancer and infectious disease have been unveiled in the medical sciences, however, little is understood about the underlying mechanism behind these very similar pathological states induced by tumor and sepsis, respectively Hotchkiss and Moldawer (2014). With this study, we propose a novel paradigm of unified functional modeling of tumor disease and sepsis from the complex dynamical network perspective by choosing the nonspecific innate immune system as the reference point for both diseases. The proposed model is not a detailed model of organs but a functional model of dynamic interactions. Here, cytokine activity is described as information flow within and between the parenchymal and the immune layer. In particular, the communication between cells of the parenchyma, and between cells of the immune layer is modeled by adaptive coupling weights. This approach is complementary to works modeling the cytokine concentrations as additional dynamical species Yiu et al. (2012). Thus our perspective accounts for the cytokine activation rather than their physical mass.
In this paper we have presented the simplest form of a two-layer network model based upon adaptively coupled phase oscillators. Although many simplifying assumptions have been made, this model can already capture essential unifying features of tumor disease and sepsis. Two important tunable model parameters have been identified, i.e., the tumor size r denoting the fraction of mutant (pathological) parenchymal cells which have a deviating natural frequency of single-cell basic metabolic activity, and an adaptivity parameter β (called age parameter) summarizing various physiological conditions like age, inflammaging, adiposity, pre-existing illness, physical inactivity, nutritional influence, and malignancy in case of tumor disease. The healthy system is modeled as a completely frequency-synchronized homeostatic state of metabolic activity in both the parenchyma and the immune layer, while in the pathological case the parenchyma splits into two frequency clusters with different collective frequency, one corresponding to the healthy part, and one corresponding to organ failure. The desynchronization of the parenchymal layer is observed for higher age parameter β with increasing probability both in case of tumor disease and sepsis.
To characterize the dynamical patterns resulting from this model, we have introduced the temporally averaged mean phase velocities of the cells as collective frequencies of metabolism, and their ensemble-averaged standard deviation sμ as a measure of pathogenicity in case of the parenchymal layer (μ = 1), and as a measure of metabolic activation in case of the immune layer (μ = 2). Thus s1 ≠ 0 is an immediate indicator of a pathological state. Further, we have discussed the initial perturbation for both types of pathological conditions, i.e., tumor and sepsis. For the analysis of the tumor disease, we assume heterogeneity in the natural frequency distribution of the parenchymal layer to account for the fact that mutated parenchymal cells may possess a faster metabolism than healthy parenchymal cells. For sepsis, we consider, however, a homogeneous network of oscillator with an initially activated cytokine matrix representing a systemic immune response. Moreover, for sepsis, we assume a high coupling strength between the parenchymal and the immune layer caused by the pro-inflammatory immune response.
In case of tumor disease, our simulations show that for small values of the age parameter β, as representative for young and healthy subjects, almost all simulations lead to a healthy state even for a relatively high fraction r of mutated cells. Thus the system is robust against random mutations of parenchymal cells. For increasing age parameter, the immune layer turns out to be more activated in order to keep the parenchymal layer synchronized. Here, only for a low fraction of mutated cells r the healthy state can be maintained. For higher values of the fraction r the immune layer is not able anymore to keep the parenchyma synchronized, and a pathological desynchronized state emerges. For high values of the age parameter the immune layer is completely dysregulated, and a severe pathological state of tumor disease is almost inevitable. One might speculate that in the in-phase scenario the pathological cluster (tumor) may be removed (by surgery) so that the healthy state is restored, whereas in the splay scenario the non-tumor cluster is already so strongly perturbed that removal of the tumor may not bring the system back to the healthy state.
In analogy with the study of tumor disease, we have demonstrated various dynamical scenarios for different age parameters in case of sepsis. Also here, the age parameter β has been shown to be critical for the system to return to the healthy state. For specifically chosen parameter values, close to those chosen for the examples of tumor disease, we have visualized the emergence of different synchronized and desynchronized states. Furthermore, we have found parameter regimes for which healthy states may coexist with pathological desynchronized states, i.e., it depends upon the random initial conditions which of the two states is asymptotically reached. Further systematic studies should investigate the detailed dependence upon the initial perturbation of the cytokin activity matrix in the immune layer, i.e., the initial cluster and its size, and upon the choice of the interlayer coupling strength. We hypothesize that the interlayer coupling strength σ may be used to model functionally the progression of sepsis where an increased σ may be interpreted as an ongoing in-stream of cytokines from the blood vessels into the stroma. For both tumor disease and sepsis our model represents only a first step towards more detailed and systematic investigations.
In summary, a unified disease model has been established using the innate immune system as the reference point. Diseases such as tumorigenesis and sepsis have been modeled by studying the dynamic functional interactions between parenchyma and stroma. For this purpose, a unified system of dynamic equations has been developed, in which only three different parameters, the age parameter, the number of mutated cells, and the initial cytokine activation are varied. Thereby, carcinogenesis, organ dysfunction in sepsis, and recurrence risk can be described in a correct functional context. Our studies open a perspective for further dynamic disease modeling. For instance, more detailed models, including disease progression may be developed in future studies.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author Contributions
JS and RB performed the numerical simulations and analyzed the simulated data. The idea of unified treatment of tumor disease and sepsis was due to TL. The idea of an adaptive two-layer network model where the cytokine activity is associated with adaptive coupling weights was due to ES. All authors designed the study and contributed to the preparation of the article. All the authors have read and approved the final article.
Funding
This work was supported by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation, project Nos. 429685422 and 440145547) and the Open Access Publication Fund of TU Berlin.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abbott, L. F., and Nelson, S. B. (2000). Synaptic Plasticity: Taming the Beast. Nat. Neurosci. 3, 1178–1183. doi:10.1038/81453
Abdol Razak, N., Jones, G., Bhandari, M., Berndt, M., and Metharom, P. (2018). Cancer-associated Thrombosis: An Overview of Mechanisms, Risk Factors, and Treatment. Cancers 10, 380. doi:10.3390/cancers10100380
Abrams, D. M., and Strogatz, S. H. (2004). Chimera States for Coupled Oscillators. Phys. Rev. Lett. 93, 174102. doi:10.1103/physrevlett.93.174102
Acebrón, J. A., Bonilla, L. L., Pérez Vicente, C. J., Ritort, F., and Spigler, R. (2005). The Kuramoto Model: A Simple Paradigm for Synchronization Phenomena. Rev. Mod. Phys. 77, 137–185. doi:10.1103/revmodphys.77.137
Albert, R., and Barabási, A.-L. (2002). Statistical Mechanics of Complex Networks. Rev. Mod. Phys. 74, 47–97. doi:10.1103/revmodphys.74.47
Altan-Bonnet, G., and Mukherjee, R. (2019). Cytokine-mediated Communication: a Quantitative Appraisal of Immune Complexity. Nat. Rev. Immunol. 19, 205–217. doi:10.1038/s41577-019-0131-x
Andrzejak, R. G., Ruzzene, G., and Malvestio, I. (2017). Generalized Synchronization between Chimera States. Chaos 27, 053114. doi:10.1063/1.4983841
Aoki, T., and Aoyagi, T. (2009). Co-evolution of Phases and Connection Strengths in a Network of Phase Oscillators. Phys. Rev. Lett. 102, 034101. doi:10.1103/physrevlett.102.034101
Aoki, T. (2015). Self-organization of a Recurrent Network under Ongoing Synaptic Plasticity. Neural Networks 62, 11–19. doi:10.1016/j.neunet.2014.05.024
Arends, J., Bertz, H., Bischoff, S., Fietkau, R., Herrmann, H., Holm, E., et al. (2015). Klinische Ernährung in der Onkologie. Aktuel. Ernährungsmed. 40, 1. doi:10.1055/s-0035-1552741
Bartsch, R. P., Liu, K. K. L., Bashan, A., and Ivanov, P. C. (2015). Network Physiology: How Organ Systems Dynamically Interact. PLoS One 10, e0142143. doi:10.1371/journal.pone.0142143
Bartsch, R. P., Schumann, A. Y., Kantelhardt, J. W., Penzel, T., and Ivanov, P. C. (2012). Phase Transitions in Physiologic Coupling. Proc. Natl. Acad. Sci. 109, 10181–10186. doi:10.1073/pnas.1204568109
Bashan, A., Bartsch, R. P., Kantelhardt, J. W., Havlin, S., and Ivanov, P. C. (2012). Network Physiology Reveals Relations between Network Topology and Physiological Function. Nat. Commun. 3, 702. doi:10.1038/ncomms1705
Beneke, G. (1971). Altersabhängige Veränderung des Kollagens und der Bindegewebszellen. Stuttgart, Germany, Schattauer Verlag, 1–37. chap. 1 Altern und Entwicklung.
Berner, R., Fialkowski, J., Kasatkin, D., Nekorkin, V., Yanchuk, S., and Schöll, E. (2019a). Hierarchical Frequency Clusters in Adaptive Networks of Phase Oscillators. Chaos 29, 103134. doi:10.1063/1.5097835
Berner, R., Mehrmann, V., Schöll, E., and Yanchuk, S. (2021a). The Multiplex Decomposition: An Analytic Framework for Multilayer Dynamical Networks. SIAM J. Appl. Dyn. Syst. 20, 1752–1772. doi:10.1137/21m1406180
Berner, R. (2021). Patterns of Synchrony in Complex Networks of Adaptively Coupled Oscillators. Cham: Springer. Springer Theses. doi:10.1007/978-3-030-74938-5
Berner, R., Polanska, A., Schöll, E., and Yanchuk, S. (2020a). Solitary States in Adaptive Nonlocal Oscillator Networks. Eur. Phys. J. Spec. Top. 229, 2183–2203. doi:10.1140/epjst/e2020-900253-0
Berner, R., Sawicki, J., and Schöll, E. (2020b). Birth and Stabilization of Phase Clusters by Multiplexing of Adaptive Networks. Phys. Rev. Lett. 124, 088301. doi:10.1103/physrevlett.124.088301
Berner, R., Schöll, E., and Yanchuk, S. (2019b). Multiclusters in Networks of Adaptively Coupled Phase Oscillators. SIAM J. Appl. Dyn. Syst. 18, 2227–2266. doi:10.1137/18m1210150
Berner, R., Vock, S., Schöll, E., and Yanchuk, S. (2021b). Desynchronization Transitions in Adaptive Networks. Phys. Rev. Lett. 126, 028301. doi:10.1103/physrevlett.126.028301
Berner, R., Yanchuk, S., Maistrenko, Y., and Schöll, E. (2021c). Generalized Splay States in Phase Oscillator Networks. Chaos 31, 073128. doi:10.1063/5.0056664
Berner, R., Yanchuk, S., and Schöll, E. (2021d). What Adaptive Neuronal Networks Teach Us about Power Grids. Phys. Rev. E 103, 042315. doi:10.1103/physreve.103.042315
Boccaletti, S., Bianconi, G., Criado, R., del Genio, C. I., Gómez-Gardeñes, J., Romance, M., et al. (2014). The Structure and Dynamics of Multilayer Networks. Phys. Rep. 544, 1–122. doi:10.1016/j.physrep.2014.07.001
Boccaletti, S., Pisarchik, A. N., del Genio, C. I., and Amann, A. (2018). Synchronization: From Coupled Systems to Complex Networks. Cambridge: Cambridge University Press.
Bomans, K., Schenz, J., Sztwiertnia, I., Schaack, D., Weigand, M. A., and Uhle, F. (2018). Sepsis Induces a Long-Lasting State of Trained Immunity in Bone Marrow Monocytes. Front. Immunol. 9, 2685. doi:10.3389/fimmu.2018.02685
Brunkhorst, F. M., Weigand, M., Pletz, M., Gastmeier, P., Lemmen, S. W., Meier-Hellmann, A., et al. (2018). S3-Leitlinie Sepsis - Prävention, Diagnose, Therapie und Nachsorge.
Calder, P. C., Bosco, N., Bourdet-Sicard, R., Capuron, L., Delzenne, N., Doré, J., et al. (2017). Health Relevance of the Modification of Low Grade Inflammation in Ageing (Inflammageing) and the Role of Nutrition. Ageing Res. Rev. 40, 95–119. doi:10.1016/j.arr.2017.09.001
Caporale, N., and Dan, Y. (2008). Spike Timing-dependent Plasticity: A Hebbian Learning Rule. Annu. Rev. Neurosci. 31, 25–46. doi:10.1146/annurev.neuro.31.060407.125639
Chen, L., Liu, R., Liu, Z.-P., Li, M., and Aihara, K. (2012). Detecting Early-Warning Signals for Sudden Deterioration of Complex Diseases by Dynamical Network Biomarkers. Sci. Rep. 2, 342. doi:10.1038/srep00342
Chen, Z., Hu, K., Stanley, H. E., Novak, V., and Ivanov, P. C. (2006). Cross-correlation of Instantaneous Phase Increments in Pressure-Flow Fluctuations: Applications to Cerebral Autoregulation. Phys. Rev. E 73, 031915. doi:10.1103/physreve.73.031915
Chockley, P. J., and Keshamouni, V. G. (2016). Immunological Consequences of Epithelial-Mesenchymal Transition in Tumor Progression. J.I. 197, 691–698. doi:10.4049/jimmunol.1600458
Chovatiya, R., and Medzhitov, R. (2014). Stress, Inflammation, and Defense of Homeostasis. Mol. Cel 54, 281–288. doi:10.1016/j.molcel.2014.03.030
Coussens, L. M., and Werb, Z. (2002). Inflammation and Cancer. Nature 420, 860–867. doi:10.1038/nature01322
Dahms, T., Lehnert, J., and Schöll, E. (2012). Cluster and Group Synchronization in Delay-Coupled Networks. Phys. Rev. E 86, 016202. doi:10.1103/physreve.86.016202
De Domenico, M., Nicosia, V., Arenas, A., and Latora, V. (2015). Structural Reducibility of Multilayer Networks. Nat. Commun. 6, 6864. doi:10.1038/ncomms7864
De Domenico, M., Solé-Ribalta, A., Cozzo, E., Kivelä, M., Moreno, Y., Porter, M. A., et al. (2013). Mathematical Formulation of Multilayer Networks. Phys. Rev. X 3, 041022. doi:10.1103/physrevx.3.041022
Drauschke, F., Sawicki, J., Berner, R., Omelchenko, I., and Schöll, E. (2020). Effect of Topology upon Relay Synchronization in Triplex Neuronal Networks. Chaos 30, 051104. doi:10.1063/5.0008341
Eichinger, S., and Lechner, K. (2004). Hämorrhagische und thromboembolische Komplikationen bei malignen Erkrankungen. Springer, 799–809. chap. 34. doi:10.1007/978-3-662-06670-6_34
Elisia, I., Lam, V., Hofs, E., Li, M. Y., Hay, M., Cho, B., et al. (2017). Effect of Age on Chronic Inflammation and Responsiveness to Bacterial and Viral Challenges. PLoS One 12, e0188881. doi:10.1371/journal.pone.0188881
Fasshauer, M., Klein, J., Blüher, M., and Paschke, R. (2004). Adipokine: Mögliches bindeglied zwischen insulinresistenz und adipositas. Dtsch. Ärztebl. Int. 101, 3491.
Franceschi, C., and Campisi, J. (2014). Chronic Inflammation (Inflammaging) and its Potential Contribution to Age-Associated Diseases. Journals Gerontol. Ser. A: Biol. Sci. Med. Sci. 69, S4–S9. doi:10.1093/gerona/glu057
Franceschi, C., Garagnani, P., Parini, P., Giuliani, C., and Santoro, A. (2018). Inflammaging: a New Immune-Metabolic Viewpoint for Age-Related Diseases. Nat. Rev. Endocrinol. 14, 576–590. doi:10.1038/s41574-018-0059-4
Fulop, T., Larbi, A., Dupuis, G., Le Page, A., Frost, E. H., Cohen, A. A., et al. (2018). Immunosenescence and Inflamm-Aging as Two Sides of the Same coin: Friends or Foes. Front. Immunol. 8, 1960. doi:10.3389/fimmu.2017.01960
Gaillard, R. C. (2007). Adipozyten: Endokrine Hochleistungsfabriken. Schweiz Ärzteztg 10, 163–167. doi:10.4414/cvm.2007.01248
Gastpar, H. (1982). Die Beeinflussung der Metastasierung über Gerinnungsmechanismen. Thieme, 118–128. chap. unb.
Greten, F. R., and Grivennikov, S. I. (2019). Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 51, 27–41. doi:10.1016/j.immuni.2019.06.025
Gross, T., and Blasius, B. (2008). Adaptive Coevolutionary Networks: a Review. J. R. Soc. Interf. 5, 259–271. doi:10.1098/rsif.2007.1229
Gross, T., D’Lima, C. J. D., and Blasius, B. (2006). Epidemic Dynamics on an Adaptive Network. Phys. Rev. Lett. 96, 208701. doi:10.1103/physrevlett.96.208701
Heerboth, S., Housman, G., Leary, M., Longacre, M., Byler, S., Lapinska, K., et al. (2015). EMT and Tumor Metastasis. Clin. Translational Med. 4, 6. doi:10.1186/s40169-015-0048-3
Hoppensteadt, F. C., and Izhikevich, E. M. (1996). Synaptic Organizations and Dynamical Properties of Weakly Connected Neural Oscillators Ii. Learning Phase Information. Biol. Cybernetics 75, 129–135. doi:10.1007/s004220050280
Hotchkiss, R. S., Moldawer, L. L., Opal, S. M., Reinhart, K., Turnbull, I. R., and Vincent, J.-L. (2016). Sepsis and Septic Shock. Nat. Rev. Dis. Primers 2, 16045. doi:10.1038/nrdp.2016.45
Hotchkiss, R. S., and Moldawer, L. L. (2014). Parallels between Cancer and Infectious Disease. N. Engl. J. Med. 371, 380–383. doi:10.1056/nejmcibr1404664
Ivanov, P. C., Bartsch, R. P., and Bartsch, R. P. (2009). Maternal-fetal Heartbeat Phase Synchronization. Proc. Natl. Acad. Sci. 106, 13641–13642. doi:10.1073/pnas.0906987106
Ivanov, P. C., and Bartsch, R. P. (2014). Network Physiology: Mapping Interactions between Networks of Physiologic Networks. Springer, 203–222. chap. 10. Networks of Networks: The Last Frontier of Complexity.
Jain, S., and Krishna, S. (2001). A Model for the Emergence of Cooperation, Interdependence, and Structure in Evolving Networks. Proc. Natl. Acad. Sci. 98, 543–547. doi:10.1073/pnas.98.2.543
Junqueira, L. C., Carneiro, J., and Kelley, R. O. (1995). Basic Histology. New York City: McGraw Hill Education.
Karlsson, M. C., Gonzalez, S. F., Welin, J., and Fuxe, J. (2017). Epithelial-mesenchymal Transition in Cancer Metastasis through the Lymphatic System. Mol. Oncol. 11, 781–791. doi:10.1002/1878-0261.12092
Kasatkin, D. V., and Nekorkin, V. I. (2018). Synchronization of Chimera States in a Multiplex System of Phase Oscillators with Adaptive Couplings. Chaos 28, 093115. doi:10.1063/1.5031681
Kasatkin, D. V., Yanchuk, S., Schöll, E., and Nekorkin, V. I. (2017). Self-organized Emergence of Multilayer Structure and Chimera States in Dynamical Networks with Adaptive Couplings. Phys. Rev. E 96, 062211. doi:10.1103/physreve.96.062211
Kivela, M., Arenas, A., Barthelemy, M., Gleeson, J. P., Moreno, Y., and Porter, M. A. (2014). Multilayer Networks. J. Complex Networks 2, 203–271. doi:10.1093/comnet/cnu016
Kuehn, C. (2015). Multiple Time Scale Dynamics. Cham: Springer. doi:10.1007/978-3-319-12316-5 Multiple Time Scale Dynamics.
Kuramoto, Y., and Battogtokh, D. (2002). Coexistence of Coherence and Incoherence in Nonlocally Coupled Phase Oscillators. Nonlin. Phen. Complex Sys 5, 380–385.
Lamouille, S., Xu, J., and Derynck, R. (2014). Molecular Mechanisms of Epithelial-Mesenchymal Transition. Nat. Rev. Mol. Cel Biol. 15, 178–196. doi:10.1038/nrm3758
Leyva, I., Sevilla-Escoboza, R., Sendiña-Nadal, I., Gutiérrez, R., Buldú, J. M., and Boccaletti, S. (2017). Inter-layer Synchronization in Non-identical Multi-Layer Networks. Sci. Rep. 7, 45475. doi:10.1038/srep45475
Lin, A., Liu, K. K. L., Bartsch, R. P., and Ivanov, P. C. (2016). Delay-correlation Landscape Reveals Characteristic Time Delays of Brain Rhythms and Heart Interactions. Phil. Trans. R. Soc. A. 374, 20150182. doi:10.1098/rsta.2015.0182
Liu, R., Aihara, K., and Chen, L. (2013a). Dynamical Network Biomarkers for Identifying Critical Transitions and Their Driving Networks of Biologic Processes. Quant Biol. 1, 105–114. doi:10.1007/s40484-013-0008-0
Liu, R., Li, M., Liu, Z.-P., Wu, J., Chen, L., and Aihara, K. (2012). Identifying Critical Transitions and Their Leading Biomolecular Networks in Complex Diseases. Sci. Rep. 2, 813. doi:10.1038/srep00813
Liu, R., Wang, X., Aihara, K., and Chen, L. (2013b). Early Diagnosis of Complex Diseases by Molecular Biomarkers, Network Biomarkers, and Dynamical Betwork Biomarkers. Med. Res. Rev. 34 (3), 455–478. doi:10.1002/med.21293
Löser, T. (2020). Aspects of Tumor Progression. Med. Hypotheses 144, 110157. doi:10.1016/j.mehy.2020.110157
Löser, T. (2018). Process Analysis of Carcinogenesis: Concept Derivation of the Tissue Function "preservation of a Homogeneous Gene Expression". Theor. Biosci. 137, 85–97. doi:10.1007/s12064-017-0256-z
Lücken, L., Popovych, O. V., Tass, P. A., and Yanchuk, S. (2016). Noise-enhanced Coupling between Two Oscillators with Long-Term Plasticity. Phys. Rev. E 93, 032210. doi:10.1103/physreve.93.032210
Madadi Asl, M., Valizadeh, A., and Tass, P. A. (2018). Dendritic and Axonal Propagation Delays May Shape Neuronal Networks with Plastic Synapses. Front. Physiol. 9, 1849. doi:10.3389/fphys.2018.01849
Maistrenko, Y. L., Lysyansky, B., Hauptmann, C., Burylko, O., and Tass, P. A. (2007). Multistability in the Kuramoto Model with Synaptic Plasticity. Phys. Rev. E 75, 066207. doi:10.1103/physreve.75.066207
Majetschak, M., and Schade, V. A. (2001). Mechanismen der inflammatorischen Wirtsantwort bei schweren Infektionen. Thieme Stuttgart, New York: Intensivmedizin.
Maksimenko, V. A., Makarov, V. V., Bera, B. K., Ghosh, D., Dana, S. K., Goremyko, M. V., et al. (2016). Excitation and Suppression of Chimera States by Multiplexing. Phys. Rev. E 94, 052205. doi:10.1103/physreve.94.052205
Male, D., Brostoff, J., Roth, D., and Roitt, I. (2012). Immunology. 8th edn. Philadelphia: Saunders.
Mantovani, A., Marchesi, F., Malesci, A., Laghi, L., and Allavena, P. (2017). Tumour-associated Macrophages as Treatment Targets in Oncology. Nat. Rev. Clin. Oncol. 14, 399–416. doi:10.1038/nrclinonc.2016.217
Markram, H., Lübke, J., Frotscher, M., and Sakmann, B. (1997). Regulation of Synaptic Efficacy by Coincidence of Postsynaptic APs and EPSPs. Science 275, 213–215. doi:10.1126/science.275.5297.213
Matsumoto, H., Ogura, H., Shimizu, K., Ikeda, M., Hirose, T., Matsuura, H., et al. (2018). The Clinical Importance of a Cytokine Network in the Acute Phase of Sepsis. Sci. Rep. 8, 13995. doi:10.1038/s41598-018-32275-8
Meisel, C., and Gross, T. (2009). Adaptive Self-Organization in a Realistic Neural Network Model. Phys. Rev. E 80, 061917. doi:10.1103/physreve.80.061917
Moorman, J. R., Lake, D. E., and Ivanov, P. C. (2016). Early Detection of Sepsis-A Role for Network Physiology. Crit. Care Med. 44, e312–e313. doi:10.1097/ccm.0000000000001548
Morán, G. A. G., Parra-Medina, R., Cardona, A. G., Quintero-Ronderos, P., and Rodríguez, É. G. (2013). “Cytokines, Chemokines and Growth Factors,” in Autoimmunity: From Bench to Bedside. Editors J. M. Anaya, Y. Shoenfeld, A. Rojas-Villarraga, R. A. Levy, and R. Cervera (Bogota, Colombia: El Rosario University Press), 133–168. chap. 9.
Nemetschek, T. (1971). Altersabhängige Abläufe am Kollagen in. Basel, Switzerland, Schattauer Verlag, 38–68. vol. 3 of Altern und Entwicklung, chap. 3.
Newman, M. E. J. (2003). The Structure and Function of Complex Networks. SIAM Rev. 45, 167–256. doi:10.1137/s003614450310.1137/s003614450342480
Nikitin, D., Omelchenko, I., Zakharova, A., Avetyan, M., Fradkov, A. L., and Schöll, E. (2019). Complex Partial Synchronization Patterns in Networks of Delay-Coupled Neurons. Phil. Trans. R. Soc. A. 377, 20180128. doi:10.1098/rsta.2018.0128
Omelchenko, I., Hülser, T., Zakharova, A., and Schöll, E. (2019). Control of Chimera States in Multilayer Networks. Front. Appl. Math. Stat. 4, 67. doi:10.3389/fams.2018.00067
Panaggio, M. J., and Abrams, D. M. (2015). Chimera States: Coexistence of Coherence and Incoherence in Networks of Coupled Oscillators. Nonlinearity 28, R67–R87. doi:10.1088/0951-7715/28/3/r67
Pikovsky, A., Rosenblum, M., and Kurths, J. (2001). Synchronization: A Universal Concept in Nonlinear Sciences. 1st edn. Cambridge: Cambridge University Press.
Popovych, O. V., Yanchuk, S., and Tass, P. A. (2013). Self-organized Noise Resistance of Oscillatory Neural Networks with Spike Timing-dependent Plasticity. Sci. Rep. 3, 2926. doi:10.1038/srep02926
Porporato, P. E. (2016). Understanding Cachexia as a Cancer Metabolism Syndrome. Oncogenesis 5, e200. doi:10.1038/oncsis.2016.3
Prescott, H. C., Osterholzer, J. J., Langa, K. M., Angus, D. C., and Iwashyna, T. J. (2016). Late Mortality after Sepsis: Propensity Matched Cohort Study. BMJ 353, i2375. doi:10.1136/bmj.i2375
Rich, R. R., and Chaplin, D. D. (2019). The Human Immune Response. Amsterdam: Elsevier, 3–17. chap. 1. doi:10.1016/b978-0-7020-6896-6.00001-6 The Human Immune Response.
Röhr, V., Berner, R., Lameu, E. L., Popovych, O. V., and Yanchuk, S. (2019). Frequency Cluster Formation and Slow Oscillations in Neural Populations with Plasticity. PLoS ONE 14, e0225094. doi:10.1371/journal.pone.0225094
Rybalova, E. V., Vadivasova, T. E., Strelkova, G. I., Anishchenko, V. S., and Zakharova, A. S. (2019). Forced Synchronization of a Multilayer Heterogeneous Network of Chaotic Maps in the Chimera State Mode. Chaos 29, 033134. doi:10.1063/1.5090184
Sakaguchi, H., and Kuramoto, Y. (1986). A Soluble Active Rotater Model Showing Phase Transitions via Mutual Entertainment. Prog. Theor. Phys. 76, 576–581. doi:10.1143/ptp.76.576
Sawicki, J. (2019). Delay Controlled Partial Synchronization in Complex Networks. Heidelberg: Springer. Springer Theses. doi:10.1007/978-3-030-34076-6_5
Sawicki, J., Koulen, J. M., and Schöll, E. (2021). Synchronization Scenarios in Three-Layer Networks with a Hub. Chaos 31, 073131. doi:10.1063/5.0055835
Sawicki, J., Omelchenko, I., Zakharova, A., and Schöll, E. (2018). Delay Controls Chimera Relay Synchronization in Multiplex Networks. Phys. Rev. E 98, 062224. doi:10.1103/physreve.98.062224
Schöll, E. (2020). Chimeras in Physics and Biology: Synchronization and Desynchronization of Rhythms. Nova Acta Leopoldina 425, 67–95. Invited contribution. doi:10.26164/LEOPOLDINA_10_00275
Schöll, E., Zakharova, A., and Andrzejak, R. G. (2020). “Chimera States in Complex Networks,” in Research Topic, Front. Appl. Math. Stat. (Lausanne: Frontiers Media SA). Ebook. doi:10.3389/978-2-88963-311-1
Seliger, P., Young, S. C., and Tsimring, L. S. (2002). Plasticity and Learning in a Network of Coupled Phase Oscillators. Phys. Rev. E 65, 041906. doi:10.1103/physreve.65.041906
Seymour, C. W., Kennedy, J. N., Wang, S., Chang, C.-C. H., Elliott, C. F., Xu, Z., et al. (2019). Derivation, Validation, and Potential Treatment Implications of Novel Clinical Phenotypes for Sepsis. JAMA 321, 2003. doi:10.1001/jama.2019.5791
Shannon, C. E. (1948). A Mathematical Theory of Communication. Bell Syst. Tech. J. 27, 379–423. doi:10.1002/j.1538-7305.1948.tb01338.x
Shepelev, I. A., Muni, S. S., Schöll, E., and Strelkova, G. I. (2021). Repulsive Inter-layer Coupling Induces Anti-phase Synchronization. Chaos 31, 063116. doi:10.1063/5.0054770
Singer, M., Deutschman, C. S., Seymour, C. W., Shankar-Hari, M., Annane, D., Bauer, M., et al. (2016). The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 315, 801. doi:10.1001/jama.2016.0287
Thomas, L. (2020a). Labor und Diagnose. Available at: https://www.clinical-laboratory-diagnostics-2020.com/.
Thomas, L. (2020b). Clinical Laboratory Diagnostics. Available at: https://www.clinical-laboratory-diagnostics-2020.com/.
Timms, L., and English, L. Q. (2014). Synchronization in Phase-Coupled Kuramoto Oscillator Networks with Axonal Delay and Synaptic Plasticity. Phys. Rev. E 89, 032906. doi:10.1103/physreve.89.032906
Vineis, P., Schatzkin, A., and Potter, J. D. (2010). Models of Carcinogenesis: an Overview. Carcinogenesis 31, 1703–1709. doi:10.1093/carcin/bgq087
Warburg, O., Posener, K., and Negelein, E. (1924). Über den Stoffwechsel der Carcinomzelle. Biochem. Z. 152, 309.
Wu, Y., and Zhou, B. P. (2009). Inflammation: a Driving Force Speeds Cancer Metastasis. Cell Cycle 8, 3267–3273. doi:10.4161/cc.8.20.9699
Xu, L., Chen, Z., Hu, K., Stanley, H. E., and Ivanov, P. C. (2006). Spurious Detection of Phase Synchronization in Coupled Nonlinear Oscillators. Phys. Rev. E 73, 065201. doi:10.1103/physreve.73.065201
Yiu, H. H., Graham, A. L., and Stengel, R. F. (2012). Dynamics of a Cytokine Storm. PLoS ONE 7, e45027–15. doi:10.1371/journal.pone.0045027
Zhang, X., Boccaletti, S., Guan, S., and Liu, Z. (2015). Explosive Synchronization in Adaptive and Multilayer Networks. Phys. Rev. Lett. 114, 038701. doi:10.1103/physrevlett.114.038701
Keywords: adaptive networks, cluster synchronization, coupled oscillators, pattern formation, sepsis, tumor disease, cytokine activity
Citation: Sawicki J, Berner R, Löser T and Schöll E (2022) Modeling Tumor Disease and Sepsis by Networks of Adaptively Coupled Phase Oscillators. Front. Netw. Physiol. 1:730385. doi: 10.3389/fnetp.2021.730385
Received: 24 June 2021; Accepted: 19 November 2021;
Published: 17 January 2022.
Edited by:
Alexander Neiman, Ohio University, United StatesReviewed by:
Justus Alfred Kromer, Stanford University, United StatesAlexey Zaikin, University College London, United Kingdom
Copyright © 2022 Sawicki, Berner, Löser and Schöll. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eckehard Schöll, schoell@physik.tu-berlin.de