 Desh Deepak Ratna
Desh Deepak Ratna Tanner Chase Francis
Tanner Chase Francis- Department of Drug Discovery and Biomedical Sciences, College of Pharmacy, University of South Carolina, Columbia, SC, United States
The striatum is an integrated component of the basal ganglia responsible for associative learning and response. Besides the presence of the most abundant γ-aminobutyric acid (GABA-ergic) medium spiny neurons (MSNs), the striatum also contains distributed populations of cholinergic interneurons (ChIs), which bidirectionally communicate with many of these neuronal subtypes. Despite their sparse distribution, ChIs provide the largest source of acetylcholine (ACh) to striatal cells, have a prominent level of arborization and activity, and are potent modulators of striatal output and play prominent roles in plasticity underlying associative learning and reinforcement. Deviations from this tonic activity, including phasic bursts or pauses caused by region-selective excitatory input, neuromodulator, or neuropeptide release can exert strong influences on intrinsic activity and synaptic plasticity via diverse receptor signaling. Recent studies and new tools have allowed improved identification of factors driving or suppressing cholinergic activity, including peptides. This review aims to outline our current understanding of factors that control tonic and phasic ChI activity, specifically focusing on how neuromodulators and neuropeptides interact to facilitate or suppress phasic ChI responses underlying learning and plasticity.
1 Introduction
The striatum is the largest portion of the basal ganglia and provides information required for movement, motivation, and sensory processing. In rodents, the striatum is typically divided into the dorsal striatum (DSt) and ventral striatum composed of the nucleus accumbens (NAc) and olfactory tubercle (Chen et al., 2020). The striatum consists of primarily inhibitory GABAergic cells that receive significant excitatory, glutamatergic input from various regions including cortex, thalamus, and other limbic structures (Haber, 2011; Gerfen and Bolam, 2016; Hunnicutt et al., 2016). Additionally, they also receive local inputs releasing substance P and GABA (Blomeley and Bracci, 2008; Pickel et al., 2000; Francis et al., 2019; Tepper et al., 2004). The predominant GABAergic neurons are primary projection neurons of the striatum, known as spiny projection neurons or medium spiny neurons (MSNs), which integrate these excitatory, inhibitory, and modulatory inputs to produce movement and motivated action, and this activity is modulated by synaptic plasticity. MSNs require global and local modulatory signaling, notably dopamine (DA) and acetylcholine (ACh) signaling, to produce lasting excitatory and inhibitory plasticity to promote behavioral learning and response (Aosaki and Kawaguchi, 1996; Aosaki et al., 1998; Hunnicutt et al., 2016; Klawonn and Malenka, 2018). However, several peptides have recently been observed to strongly control local activity and response (Picciotto, 2008; Castro and Bruchas, 2019; Belilos et al., 2023). These modulatory signals from global and local sources converge to drive long-term potentiation (LTP), long-term depression (LTD) (Xu et al., 2020; Rana et al., 2022), and ensemble activity (Pennartz et al., 1994; O’Donnell, 1999), critical for striatal function and behavior.
The primary source of ACh is local cholinergic interneurons (ChIs), which make up only 1% of the total cells within the region. ChIs are widely distributed throughout the striatum and are found in low density. Despite the small density of cells, ChIs have large axonal and dendritic arbors, allowing them to influence activity by providing broad, synchronous signals to MSNs and other neuronal cell types in striatum such as neuropeptide Y (NPY)-expressing and tyrosine hydroxylase-expressing interneurons (English et al., 2012; Matamales et al., 2016; Kocaturk et al., 2022). ChI activity often anti-correlates with midbrain activity and DA release during a widely described “conditioned pause” response, which opens windows in which signaling can facilitate plasticity (Kimura et al., 1984; Aosaki et al., 1994a; Graybiel et al., 1994; Morris et al., 2004). To provide temporally locked release of ACh, this response relies on: (1) intrinsic mechanisms; (2) classical neurotransmitters that act on post-synaptic sites causing direct excitation or inhibition including glutamatergic and GABAergic neurons; and (3) neuromodulatory systems including dopamine, acetylcholine, and peptides. ChIs in both the dorsal and ventral striatum play key roles in regulating various behavioral functions, including motivation, reward, learning, feeding, pleasure, motor control, and decision-making (Haber, 2011; Warner-Schmidt et al., 2012; Atallah et al., 2014).
Novel tools such as neurotransmitter and neuromodulator sensors have provided a new understanding into the mechanisms of this phasic transmission. These studies, paired with current and previous pharmacological studies, have given deep insight into the necessity and sufficiency of ACh in controlling different forms of excitatory plasticity on MSNs and other cell types. Here, we review how ChIs are excited or inhibited to provide a basis for understanding what factors participate in phasic ChI responses underlying plasticity. In addition, we discuss a newly described mechanism for peptide control over these phasic responses and how this response facilitates plasticity in the striatum.
2 ChI properties
2.1 Morphology and distribution
ChIs are distinctive with large soma and aspiny dendrites with a high order of axonal arborizations (Pickel et al., 2000; Tepper and Bolam, 2004). These neurons can be distinguished by choline acetyltransferase (ChAT) and vesicular ACh transporter (VAChT) expression. ChIs can self-regulate local ACh through their expression of acetylcholinesterase (Gonzales and Smith, 2015). While the developmental origin of NAc and DSt ChIs arise from the septal epithelium during development, the DSt also contains ChIs arising from the medial ganglionic eminence and preoptic area, which may drive diversity in ChI function and activity (Allaway and Machold, 2017; Ahmed et al., 2019). In rodents, ChIs are higher in density in dorsolateral regions in the rostral portions of the striatum and tend to be higher in density in rostral regions, compared to more caudal regions (Meredith et al., 1989; Matamales et al., 2016; Gonzales and Smith, 2015).
2.2 Intrinsic control of ChI activity
ChIs are tonically active neurons spontaneously firing around 5–10 Hz and these tonic activity patterns are autonomous, without the need for any input (Wilson et al., 1990). ChI activity primarily releases ACh but a subset of ChIs have been documented to also express vGlut3 and directly release glutamate or facilitate glutamate release via presynaptic nicotinic receptors (Higley et al., 2011). ChIs display several distinct patterns both ex vivo and in vivo: bursting, irregular, and rhythmic firing or “pacemaking” firing (Goldberg and Wilson, 2005; Sharott et al., 2012; Shroff et al., 2023) (Figure 1A). In addition, mixed mode ChIs are observed in the NAc shell that differs across sex (Olson et al., 2024). The mode of these cells can change with the blockade of Cav2.2 calcium channels which activates a SK potassium channel current, causing the mode to change from rhythmic to bursting (Bennett et al., 2000; Goldberg and Wilson, 2005; Goldberg and Wilson, 2016). The tonic activity of ChIs relies on calcium channels, particularly Cav2.1 and 2.2, and several potassium conductances, including the BK and SK potassium currents which mediate the repolarization and medium afterhyperpolarization potential, respectively. Cav1 causes a slower afterhyperpolarization potential and inward rectifying potassium currents can prolong the hyperpolarization (Goldberg et al., 2009). ChIs experience a large voltage sag during inhibition, characteristic of the hyperpolarization-activated cyclic nucleotide-gated (HCN) channel-mediated Ih potassium current (Oswald et al., 2009), which drives rebound spiking (Figure 1B). In addition, the Kv7.2/7.3 mediated delayed rectifier IKr potassium current (Zhang et al., 2018) or Kv1 (Tubert et al., 2024) drives inhibition after burst firing (Figure 1B). This inhibition provides the ability to hyperpolarize and suppress ChI firing and may be a significant part of the inhibitory current in phasic pause responses (Zhang et al., 2018).
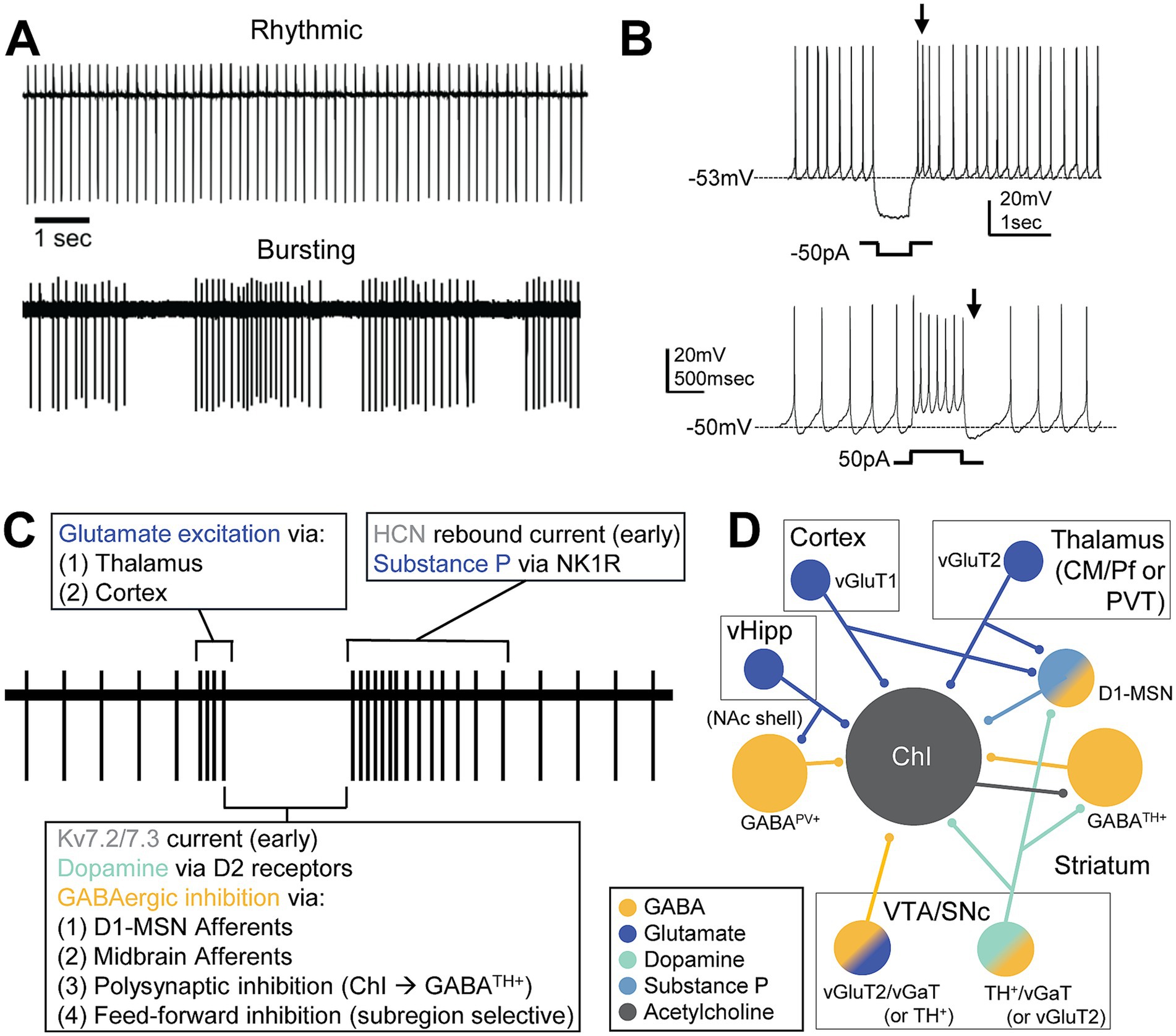
Figure 1. Firing pattern mechanisms of ChIs. (A) Ex vivo cell attached recordings showing rhythmic and bursting activity of ChIs. (B) Whole cell patch clamp of ChIs ex vivo shows rebound firing caused by Ih current after current injected to inhibit the ChI (top) and, after current is injected to excite the ChI, inhibition caused by an after hyperpolarization initiated by Cav1 channels and a Kir current followed by depolarization by HCN and persistent sodium currents (bottom). (C) Phases of the conditioned pause response and known mechanisms driving each phase. Glutamatergic excitation causes bursting activity followed by DA and GABA mediated pause or decrease in firing. Rebound firing occurs followed by extended excitation by the neuropeptide substance P. (D) Local and afferent inputs to ChIs that have been identified to be critical in the conditioned burst-pause-rebound response. Cortical and thalamic inputs activate both ChIs and MSNs via glutamate (dark blue). D1-MSNs activated by this input are partially responsible for the initial GABAergic inhibition (yellow) followed by the release of substance P (light blue). GABA interneurons, ChIs, and D1-MSNs receive midbrain dopamine (teal) which modulates ChI activity. vHipp inputs to GABAPV+ neurons promote feedforward inhibition to ChIs in the NAc shell. ChIs receive excitatory glutamatergic and inhibitory GABAergic input from midbrain VTA neurons. These neurons could be glutamate only, GABA only, DA only, or a mix which are intermixed and differentially distributed across midbrain sub-compartments. Abbreviations: centromedian/parafascicular nucleus of the thalamus (CM/Pf), cholinergic interneuron (ChI), dopamine 2 receptor (D2R), nucleus accumbens (NAc), neurokinin 1 receptor (NK1R), paraventricular thalamus (PVT), substantia nigra pars compacta (SNc), tyrosine hydroxylase (TH), ventral hippocampus (vHipp), ventral tegmental area (VTA), vesicular GABA transporter (vGAT), vesicular glutamate transporter (vGluT).
3 Neurotransmitter and neuromodulator contributions to ChI phasic activity
Despite the intrinsic firing patterns of the ChIs, synaptic input can play a prominent role in altering activity patterns and synchronization. In vivo, ChIs synchronize their firing in response to behaviorally relevant stimuli (Ravel et al., 1999; Nougaret and Ravel, 2015) and undergo a pause burst pattern termed the “conditioned pause response” (Schulz and Reynolds, 2013; Zhang and Cragg, 2017). This response is thought to be driven by many diverse afferent and local sources, as listed in Table 1.
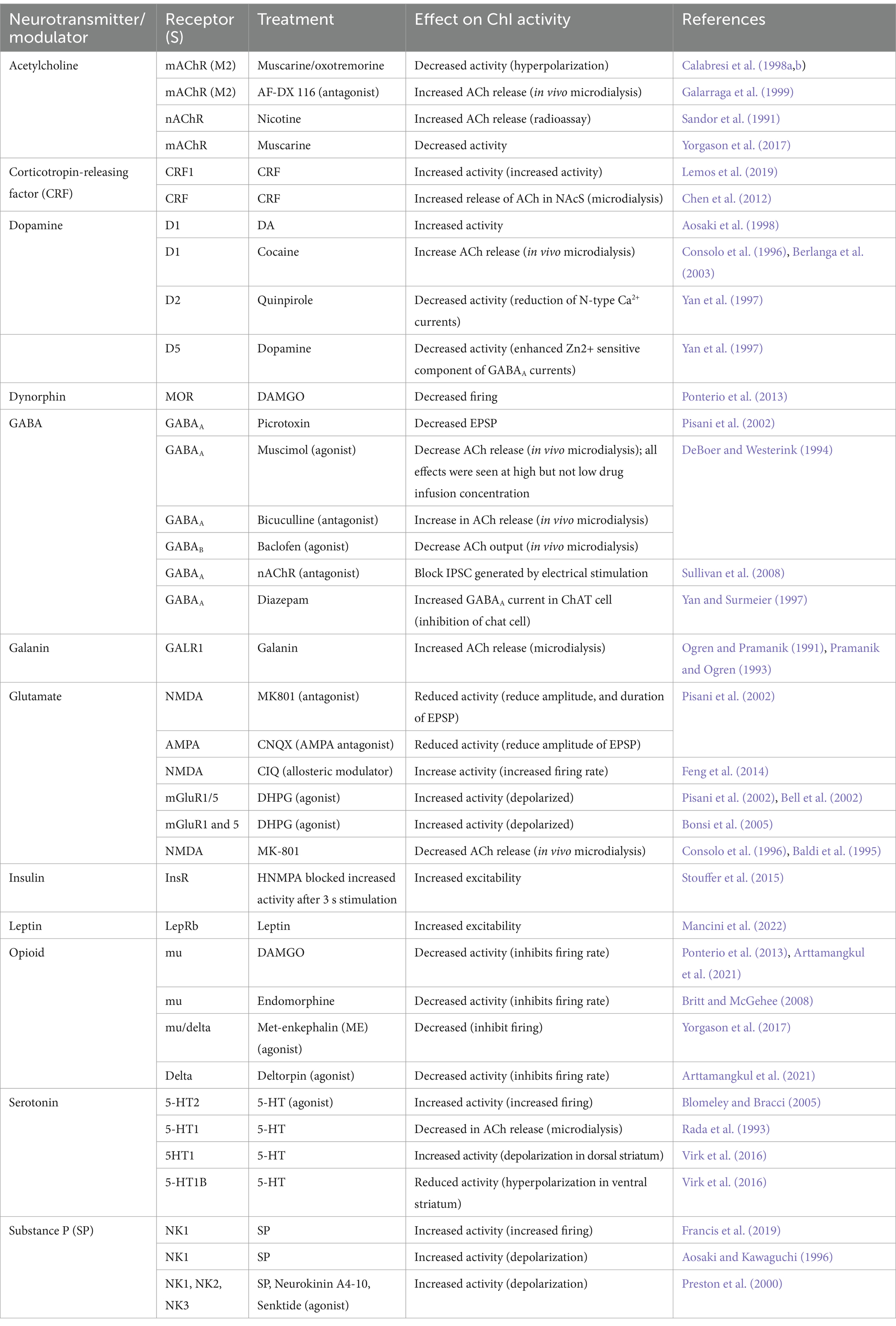
Table 1. Changes in striatal cholinergic interneuron activity by various neurotransmitters and neuromodulators.
Salient sensory stimuli, rewarding stimuli, and aversive stimuli during classical conditioning and instrumental tasks produce a stereotyped, multi-phasic, pause-rebound response in ChIs (Figure 1C) (Apicella et al., 1997; Morris et al., 2004; Schulz and Reynolds, 2013; Nougaret and Ravel, 2015; Zhang and Cragg, 2017). While salient stimuli alone can cause some ChIs to elicit this response, more ChIs are recruited after conditioning this stimulus to a rewarding or aversive outcome (Aosaki et al., 1994b; Nougaret and Ravel, 2015). Additionally, this response occurs in some ChIs, but not all, and may represent distinct sensory stimuli (Aosaki et al., 1994b; Apicella et al., 1997). However, the features of these responses appear consistent across these stimuli and are conserved across mammals. These responses have been observed in rodents as well as primates when assessing in vivo firing rates (Apicella et al., 1997; Ravel et al., 1999), calcium fluorescence signals from ChIs (Hegedüs et al., 2023), and ACh release via ACh sensor fluorescence (Gallo et al., 2022; Belilos et al., 2023). These conserved responses are significant because they underlie neuronal plasticity, which is important for learning, sensory discrimination, and motivated response (Zhang and Cragg, 2017). In addition, this pause-rebound is observed across all subregions of the striatum, with varying strength and duration of each signal (Duhne et al., 2024). The overarching mechanisms of this response have been debated but appear to involve multiple mechanisms, including several excitatory inputs to ChIs, intrinsic ChI electrophysiological properties, neuromodulators including DA (Paladini and Roeper, 2014; Goldberg and Wilson, 2016; Zhang and Cragg, 2017), and likely require other synaptic signaling factors including some peptides. While there are various ChI activity responses to sensory stimuli (Zhang and Cragg, 2017), here we discuss the classical burst-pause-rebound response and distinguish between factors that drive each phase. The mechanisms contributing to each behavior and subregion are summarized in Figures 1C,D and discussed in detail below.
3.1 Burst excitation via glutamate
ChIs are tonically active and continue to fire action potentials in the absence of any input (Bennett and Wilson, 1999). Glutamate receptor expression plays a significant role in promoting time-locked ChI activity and synchrony, influencing plasticity on local neurons and MSNs. Fast neurotransmission through AMPA receptors drives bursting activity in more tonic-firing populations (Colquhoun et al., 1992; Ding et al., 2010). All AMPA receptor subunits are expressed on ChIs (Calabresi et al., 1998a; Bell et al., 2002; Pisani et al., 2002; Deng et al., 2007) with subunits GluR1, GluR3, and GluR4 contributing to a high calcium permeability (Götz et al., 1997). ACh release is critically dependent on NMDA receptor activation on ChIs (de Jesus Aceves Buendia et al., 2019). In situ hybridization studies with double labeling reveal mRNA expression for NMDAR2D and lower levels of NMDAR2B and NMDAR1 subunits in ChIs (Landwehrmeyer et al., 1995). Additionally, single-cell PCR analysis and double-label in situ hybridization have detected low expression of NMDAR2A subunits (Standaert et al., 1999; Richardson et al., 2000). Allosteric modulation of NMDARs by bath application with CIQ (positive allosteric modulator for NMDA) enhances the spontaneous firing rate of ChIs (Feng et al., 2014).
Glutamatergic inputs to ChIs arise predominantly from the cortex and the thalamic parafascicular nucleus (PF) (Kemp and Powell, 1970; Scatton and Lehmann, 1982; McGeorge and Faull, 1989; Lapper and Bolam, 1992; Groenewegen and Berendse, 1994; Thomas et al., 2000; Ding et al., 2010, 2014; Smith et al., 2014; Guo et al., 2015; Klug et al., 2018; Assous and Tepper, 2019; Mandelbaum et al., 2019). These glutamatergic terminals can be differentiated by their origin: cortical terminals express vGluT1, while thalamic terminals express vGluT2 (Fremeau et al., 2004). In rats, the intralaminar thalamic nuclei serve as the primary origin of thalamostriatal projections (Berendse and Groenewegen, 1990; Erro et al., 2001; Van Der Werf et al., 2002) with substantial projections arising from centromedian-parafascicular (CM-PF) nuclei. Although both these nuclei send projections to ChIs, neuronal tracing in transgenic mice has shown that ChIs receive more projections from cortical neurons compared to the thalamus (Guo et al., 2015), but with less synaptic contact (Sadikot et al., 1992; Doig et al., 2014). Also, there is a difference in the synaptic localization onto the ChIs. Distal dendrites of ChIs receive synaptic input from cortical terminals. In contrast, soma and the dendritic shaft receive synaptic connections from thalamic terminals (Lapper and Bolam, 1992; Dimova et al., 1993; Sidibé and Smith, 1999; Thomas et al., 2000). Single-pulse electrical stimulation of cortical and thalamic neurons elicits different responses in ChIs. Cortical stimulation results in strong spiking activity, whereas thalamic stimulation induces weaker spiking. However, during trains of high-frequency stimulation, the probability of ChIs firing gradually decreases for cortical inputs but increases for thalamic inputs (Ding et al., 2014). This region-driven change in firing is driven by synaptic facilitation from thalamic inputs and accommodation from cortical inputs (Mamaligas et al., 2019).
Cortical and thalamic input can drive the initial excitation and may be responsible for the initial inhibitory pause via nicotinic DA release (Ding et al., 2010; Oldenburg and Ding, 2011). Cortical inputs are associated with the strongest initial ChI activity through direct excitation. Cortical stimulation can produce the initial excitation, pause, and rebound responses and facilitate DA release following excitation (Kosillo et al., 2016). Intralaminar thalamic inputs are implicated in the initial burst response through direct depolarization and activation of ChIs (Ding et al., 2010; English et al., 2012; Doig et al., 2014), and activity of these inputs are sustained through the initial phase of the pause. In particular, the CM/Pf region of the thalamus in primates and lateral and medial parafascicular thalamic nuclei are necessary for all three phases (Matsumoto et al., 2001; Yamanaka et al., 2018). Pauses and rebounds are attenuated after muscimol-mediated neuronal inactivation of these thalamic regions (Matsumoto et al., 2001). Despite this, the pause can occur in the absence of this initial excitation, indicating that multiple mechanisms may cause this excitation.
In the striatum, DA terminals also exhibit neurotransmitter co-release, including glutamate. In the medial ventral tegmental area (VTA) there are a variety of cell types with differential neurotransmitter release: glutamate only, glutamate with GABA, and GABA only (Miranda-Barrientos et al., 2021). DA neurons from this region tend to project to medial striatal subregions (Tecuapetla et al., 2010; Poulin et al., 2018; Chuhma et al., 2023; Voorn et al., 2004). The DA neurons in the lateral VTA are reported to co-release GABA with DA or DA alone (Kawano et al., 2006; Tritsch et al., 2012; Poulin et al., 2018; Steinkellner et al., 2018; Mingote et al., 2019). As evidence of this spatial segregation, midbrain glutamate inputs can excite ChIs in the striatum where the strongest excitation seen is in the medial shell region of the NAc (Chuhma et al., 2023). Glutamatergic input from VGluT2 expressing DA neurons in the medial portions of the VTA can produce burst excitation followed by a pause as well in the medial shell of the NAc (Chuhma et al., 2014). Some portions of the DSt defies this topology and it has been shown stimulation of midbrain inputs to this region excite ChIs (Straub et al., 2014; Mingote et al., 2019). This is driven by activation of metabotropic glutamate receptors (mGluRs) or D5 receptors which excite ChIs in the lateral DSt via TrpC3/7 channels (Chuhma et al., 2018). Overall, glutamate from multiple sources may contribute to these initial burst responses but is spatially differentiated by striatal sub-compartments.
3.2 Factors contributing to the pause
The mechanisms of the pause phase in ChI activity have been described and studied most prominently (Cragg, 2006; Brown et al., 2012; Zhang and Cragg, 2017; Zucca et al., 2018). This pause is a period of decreased or no ChI activity that is synchronous spatially across ChIs. This phenomenon requires several identified mechanisms, including DA inhibition, delayed rectification by intrinsic currents, and GABAergic inhibition (Sullivan et al., 2008; Aosaki et al., 2010; Brown et al., 2012; Tubert et al., 2024).
3.2.1 Dopamine
The most studied neuromodulator released in the striatum is DA. Dopaminergic inputs to the striatum primarily originate from the mesolimbic and nigrostriatal systems (Björklund and Dunnett, 2007; Nieh et al., 2013). These DA afferents are topographically arranged within the striatum. Projections from the VTA to NAc follow a medial-to-lateral topography where the medial shell of the NAc receives inputs from the posterior-medial VTA, while lateral shell and core receive inputs from the lateral VTA, with no overlap in the projecting VTA-DA neurons (Swanson, 1982; Beier et al., 2015; Luo et al., 2018; Breton et al., 2019; Mingote et al., 2019; Beier, 2022). The DA projections to the DSt, originate from the most lateral substantia nigra pars compacta (SNc) (Haber, 2011). DA signaling in the striatum occurs via both slow, long-lasting volume transmission and phasic release through discrete synaptic connections (Kubota et al., 1987; Hattori et al., 1991; Fuxe et al., 2010; Sulzer et al., 2016; Uchigashima et al., 2016; Liu and Kaeser, 2019).
Dopaminergic innervation of the striatum is well known to modulate both MSN and ChI activity (Bonsi et al., 2011; Lim et al., 2014; Mamaligas et al., 2019). ChIs are reported to co-express D2 and D5 receptors (Yan and Surmeier, 1996, 1997; Maurice et al., 2004; Castello et al., 2020). In situ hybridization and immunohistochemistry studies have shown co-localization of D2 receptors in ChAT-positive ChIs (Le Moine et al., 1990; Khan et al., 2000). D5 receptors are expressed in all striatal ChIs at levels significantly greater than those found in other striatal neurons. Electron microscopy and immunostaining studies further show that D5 receptors are localized to ChIs (Bergson et al., 1995; Calabresi et al., 2000; Khan et al., 2000; Rivera et al., 2002), where they are predominantly expressed in cell bodies and on primary dendrites (Centonze et al., 2003). In slice preparations completely lacking the D1 receptor, DA application depolarized ChIs suggesting that D5 receptors are responsible for increasing ChI activity (Centonze et al., 2003). This depolarization is through mGluR and D5 receptor-mediated activation of TrpC3/7 channels in the lateral DSt, and mGluR only in the medial DSt (Chuhma et al., 2018). This depolarization was also evident following D1/D5 receptor agonism which significantly suppressed the pause response independently of HCN channels (Tubert et al., 2024). In a DA-depleted state, inverse agonism of the D5 receptor restores the pause response. Overall, this suggests activity of the D5 receptor plays a prominent role in inhibiting the pause response in both DA-depleted and DA-intact conditions.
The inhibitory D2 receptor is more prominently expressed than the D5 receptor, leading to the predominantly inhibitory effects of DA on ChIs (Yan et al., 1997). In response to salient stimuli, a pause in the tonic activity of ChIs is observed, triggered by a burst of midbrain DA in response to reward-associated or aversion-associated cues and events (Schulz and Reynolds, 2013; Straub et al., 2014; Kim et al., 2019) via inhibitory D2 receptors (Ikarashi et al., 1997; Watanabe and Kimura, 1998). This aligns with findings that show D2 agonists reduce the firing frequency of ChIs (Maurice et al., 2004; Wang et al., 2013) and DA application significantly diminishes spontaneous firing rates of ChI neurons in vivo (Rolls et al., 1984). The pause in ACh release can be completely blocked in vivo by the D2 receptor antagonist sulpiride in the DSt (Watanabe and Kimura, 1998), and NAc (Gallo et al., 2022), and D2 receptor overexpression can enhance this response (Gallo et al., 2022). This outcome is also evident ex vivo with electrically stimulated pause responses being diminished by decreased DA expression via a 6-OHDA lesion (Sanchez et al., 2011), suppressing D2 signaling pharmacologically (Ding et al., 2010), or knocking out D2 receptors (Kharkwal et al., 2016). Ex vivo nigrostriatal afferents to DSt ChIs produce a D2 receptor-sensitive pause, and a single action potential elicited from SNc activation causes all described phases (Straub et al., 2014). In addition, blocking D2 receptor activation can allow for excitation. Suppression of D2 receptor activation with sulpiride promotes burst firing followed by a brief decrease in firing in most ChIs during phasic stimulation of SNc afferents via unmasking of mGluR signaling (Cai and Ford, 2018). Overall, this suggests that D2 receptor signaling is critically important for the pause response.
3.2.2 Acetylcholine
ChIs are the primary source of ACh in the striatum. ACh can directly control ChI activity and indirectly control signaling contributing to the pause response (Goldberg and Wilson, 2016). ACh released in the striatum, whether from local or external sources, acts through muscarinic ACh receptors (mAChRs) and nicotinic ACh receptors (nAChRs) (Oldenburg and Ding, 2011). In the striatum, mAChRs are present on ChIs (Weiner et al., 1990; Ding et al., 2006; Oldenburg and Ding, 2011), with M4 subtype being the most abundant, followed by M1 and M2 subtypes (Yasuda et al., 1993; Chapman et al., 2011). M2 and M4 AChRs are coupled to Gi/o inhibitory signaling and suppress voltage-activated calcium channel (Cav2) currents while enhancing inward rectifying potassium channel (Kir3) currents (Caulfield and Birdsall, 1998; Brown, 2018). M2 and M4 AChRs are expressed presynaptically on ChIs and function as autoreceptors to reduce ACh release (Hersch et al., 1994; Ding et al., 2006; Ztaou et al., 2016). Blocking M2 receptors increases ACh release in the striatum (Lachowicz et al., 2001) and muscarinic agonists inhibit ACh release not only in the striatal slices but also in hippocampal and cortical slice preparation (Zhang et al., 2002). In contrast, the genetic knockout mouse model reveals that autoinhibition of ACh release in striatum is mainly regulated by M4 receptor (Zhang et al., 2002).
nAChRs are expressed post-synaptically on ChIs or pre-synaptically on afferent terminals and are composed of various subunits that can form either homomeric or heteromeric complexes (Albuquerque et al., 2009; Dani, 2015; Ho et al., 2020). Alpha (α2-10) and beta (β2-4) are the most prominently expressed nAChR subtypes in the striatum (Zoli et al., 2002; Exley and Cragg, 2008; Gotti et al., 2009; Feduccia et al., 2012). Hybridization studies have shown that ChIs express α7- and β2- containing nAChRs, which play key roles in behaviors related to anxiety, social interactions, and exploration (Abbondanza et al., 2022). Deleting these receptors significantly alters several behavioral parameters, particularly as mentioned above (Abbondanza et al., 2022).
ACh and DA release are concurrently released and often covary. That is, when DA is released, ChI activity is decreased, resulting in low ACh release (Barbeau, 1962; Morris et al., 2004; Brown et al., 2012; Chuhma et al., 2014; Straub et al., 2014; Chantranupong et al., 2023). The temporal coordination of ACh and DA dynamics in striatum reflects the intrinsic rhythms of these neurotransmitters resulting in locally synchronized activity of the ChIs (Krok et al., 2023). Interaction between ACh and DA has been observed to play a key role in facilitating the pause response. The pause response is dependent on DA release, and nAChR activation can promote its release from DA terminals. Local ACh released modulates DA signaling through nAChRs and mAChRs located on DA axons in the striatum. Notably, DA release increases following selective and synchronous activation of ChIs via nAChRs (β2-containing) (Giorguieff et al., 1976; Clarke and Pert, 1985; Kulak et al., 1997; Wonnacott et al., 2000; Cachope et al., 2012; Exley et al., 2012; Threlfell et al., 2012; Koranda et al., 2014). In fact, activating cortical and thalamic inputs to the striatum promotes ChI activity, and increased ACh release can lead to more DA release (Threlfell et al., 2012; Kosillo et al., 2016). Glutamate released by ChIs may also play a secondary role in addition to the ACh-mediated presynaptic DA release in striatum (Wonnacott et al., 2000; Kljakic et al., 2017). AMPA receptors antagonist 2,3-dihydroxy-6-nitro-7-sulfamyl-benzo (f) quinoxaline (NBQX) has been shown to attenuate the amplitude of excitatory postsynaptic potentials in MSNs (Cachope et al., 2012), suggesting a multi-synaptic effect is responsible for ACh-mediated DA release. Interestingly, ACh release spatially scales DA release through ACh waves acting on nicotinic receptors (Matityahu et al., 2023). In the medial NAc, ChIs increase firing following photo-stimulation of DA terminals, whereas inhibitory effects are observed throughout most other striatal regions (Chuhma et al., 2023). These inhibitory effects on ChIs are mediated via a polysynaptic pathway involving inhibitory interneurons (Dorst et al., 2020). These connections are robust, as a single action potential can suppress the tonic activity of neighboring ChIs (Dorst et al., 2020). This inhibition is discussed further in section 3.2.2.
The striatum also receives cholinergic inputs from the pedunculopontine tegmentum nucleus (PPT) and laterodorsal tegmental nucleus (LDT), as confirmed by lesion studies (McGeer et al., 1971). Immunofluorescence labeling shows cholinergic innervation of both the striatum and NAc from PPN and LDT. The dorsolateral striatum receives dense innervation from rostral PPN, whereas the caudal PPN innervates small, sparsely distributed regions in the DSt (Dautan et al., 2014). ACh projections from PPN are predominantly restricted to the anterior striatum (Dautan et al., 2014), which also receives innervation from prefrontal cortical areas (Hunnicutt et al., 2016). However, retrograde tracing studies have revealed that a considerable proportion of these projections are non-cholinergic (Dautan et al., 2014; Klug et al., 2018) and are likely glutamatergic, synapsing more selectively on ChAT+ interneurons (Klug et al., 2018). This ACh source does not enhance DA release via nAChR activation (Brimblecombe and Cragg, 2015), a property attributed to ChI ACh release. The role of these inputs on the conditioned pause response is not known.
3.2.3 GABA
GABAergic input to ChIs is primarily local, driven by tonic GABAergic activity and local feedforward inhibition. GABAA receptors mediate inhibitory signaling from striatal GABAergic interneurons to ChIs (Gonzales et al., 2013; Sato et al., 2014). ChIs have been reported to co-express several GABAA receptor subunits, α3-4, β1-3, and γ1-3 subunits, as shown by RT-PCR analysis from individual ChIs (Persohn et al., 1992; Lim et al., 2014; Boccalaro et al., 2019).
ChIs receive extensive GABAergic inputs from MSN collaterals and GABAergic interneurons (Gonzales et al., 2013). Substance P expressing MSNs, also identified by DA 1 (D1) receptor expression, primarily synapse on ChIs. Enkephalin-positive terminals form few synapses on ChAT-positive neurons in rats, whereas substance P-positive terminals form symmetric synapses on cell bodies and proximal dendrites (Bolam et al., 1986; Martone et al., 1992; Kuramoto et al., 2007). This synaptic input from putative MSNs is different in primates. Using ultrastructural analysis with immunogold and localization along with peroxidase immunostaining, it has been shown that ChIs in primates receive GABAergic inputs, intra-striatal in origin, which are terminals from the direct (D1-MSNs) and indirect pathway (dopamine 2 (D2) expressing MSNs) (Gonzales et al., 2013). Substance P/D1-MSNs generate large inhibitory currents on ChIs (Francis et al., 2019) via GABAA and GABAB receptors (Martone et al., 1992; Kuramoto et al., 2007). Pauses mediated by D1-MSN activation and collateral inhibition produce a pause followed by long-lasting increases in activity that are dependent on substance P, neurokinin 1 receptor signaling (Belilos et al., 2023). It is possible that dopaminergic input to D1-MSNs and excitation via concomitant glutamatergic input and D1 receptor activation could enhance D1-MSN activation and inhibition of ChIs. However, blocking D1 receptor signaling in the DSt of primates was only able to suppress the pause response caused by a conditioned cue in a small subset of ChIs (Watanabe and Kimura, 1998), suggesting that this effect could be either selective to subtypes of ChIs or be region selective.
Local GABAergic inhibition is also sensitive to D2 receptor activity. ChIs participate in a large recurrent inhibitory network (Sullivan et al., 2008). Feedforward inhibition via polysynaptic connections from ChIs to tyrosine-hydroxylase positive (TH+) GABAergic interneurons (GABATH+) can cause inhibition between local ChIs (Sullivan et al., 2008; Dorst et al., 2020). This could facilitate spatially localized inhibition of ChIs following ChI bursting activity by glutamatergic input. Increased DA can suppress this inhibition through D2 receptors, which may suggest that DA levels tightly and spatiotemporally control somatic ChI inhibition and polysynaptic inhibition.
Feedforward inhibition through other GABAergic interneurons has been hypothesized to play a role in the pause response. Local GABAergic inhibition via electrical stimulation can shunt action potential firing in ChIs (Bennett and Wilson, 1998). In monkeys, CM thalamic neuron stimulation promotes GABAA receptor release of ACh, but through an unknown GABA source (Nanda et al., 2009). In the DSt, parvalbumin (PV+) interneurons and somatostatin (SOM+) have weak or no GABAA receptor-mediated connectivity to ChIs (Szydlowski et al., 2013), indicating these local interneuron populations likely do not participate in feed-forward inhibition or contribute to the ChI pause in this striatal sub-compartment. However, in the NAc shell, ventral hippocampal excitatory input to PV+ interneurons can promote strong inhibition of ChIs, while the paraventricular thalamus mainly excites ChIs (Baimel et al., 2022). Overall, the effect on the pause response is likely limited as the response is rapid and shows little to no modulation by dopamine, except in the case of polysynaptic inhibition by GABATH+ neurons.
The pause is likely partially driven by midbrain GABAergic inhibition of ChIs. Midbrain neurons can vary in their expression and co-expression. These cells can be DA-only, DA-GABAergic, GABAergic-only, and glutamatergic-GABAergic (Miranda-Barrientos et al., 2021; Chuhma et al., 2023). GABA is released from the midbrain across all striatal sub-compartments (Chuhma et al., 2023; Kim et al., 2023). In the NAc this response is varied. Optogenetic stimulation of midbrain GABAergic terminals induces inhibitory postsynaptic current (IPSC) in ChIs, effectively silencing tonic activity within the ventral NAc-shell, but not the dorsal NAc-shell (Brown et al., 2012; Al-Hasani et al., 2021). This pause in ChIs activity resembles the conditioned pause response and is reversed with a GABAA receptor antagonist (Brown et al., 2012; Dorst et al., 2020). In the DSt, GABA can also be loaded and released from DA neuron terminals (Tritsch and Sabatini, 2012; Nelson et al., 2014; Stensrud et al., 2014; Kim et al., 2023) or released from midbrain inputs. Midbrain input to ChIs is strong and prevalent (Chuhma et al., 2023). ChIs are excited by co-releasing midbrain inputs that release GABA and glutamate; however, the strength of the inhibition tends to dominate the response. Therefore, GABAergic input from the midbrain may contribute to this inhibition. This has been observed in vivo as well. In one study, stimulation of GABAergic midbrain neurons targets and directly inhibits ChIs in a pause-like response, facilitating stimulus-outcome learning (Brown et al., 2012). These studies would suggest that direct GABAergic inhibition plays a role in the pause response.
3.2.4 Intrinsic inhibition
This DA-mediated inhibition occurs in coordination with intrinsic ChI properties, particularly the slow inactivating IKr potassium currents, which are induced with the withdrawal of excitation (Zhang et al., 2018) and regulate thalamic-mediated pauses (McGuirt et al., 2022). However, this may only occur in the case of pauses that follow bursts and may only account for the initial period of inhibition (Oswald et al., 2009). In addition, the afterhyperpolarization potential could be prolonged by inhibition of the Ih current and sodium channels (Aosaki et al., 2010; Ding et al., 2010).
3.3 Rebound from pause
Rebound ChI spiking that occurs following the pause has received considerably less attention. The rebound can last up to seconds and also appears to rely on multiple mechanisms (Schulz and Reynolds, 2013). This response is not a consistently observed response across all behaviors and regions, though it is observed with most thalamic-induced pauses (Matsumoto et al., 2001; Yamanaka et al., 2018). While both appetitive and aversive conditioned stimuli produce the rebound response, aversive stimuli produce large rebound responses (Nougaret and Ravel, 2015).
3.3.1 Intrinsic properties
The initial cause of the rebound is likely due to intrinsic ChI mechanisms of inhibition-induced rebound (Figure 1B) driven by HCN channels that produce the Ih current (Zhang et al., 2018), among other currents. However, this does not describe the extensive duration of the response in some preparations, as has been shown in the DSt (Straub et al., 2014). Nigrostriatal afferents to the DSt can produce pause and more lasting burst response (rebound) responses, where the rebound response mechanism was not identified (Straub et al., 2014). Blockade of calcium channels by cadmium and action potentials by tetrodotoxin eliminated this response, indicating that the synaptic release of another substance is causing this excitation. Aversive stimuli tend to produce the largest and longest-lasting rebound responses, which have been observed to last up to seconds (Ravel et al., 2003; Belilos et al., 2023). Therefore, this lasting excitation would require an additional source.
3.3.2 Substance P
The peptide substance P has emerged as a potential candidate for the long-lasting excitation following the pause. Substance P is expressed at high levels within the striatum and is released in response to salient stimuli (Commons, 2010; Vanderah and Sandweiss, 2015; Belilos et al., 2023). Substance P modulates DA release (Brimblecombe and Cragg, 2015), its effect varies among ChI-dense regions around striosomes in the DSt (Brimblecombe and Cragg, 2015), and causes significant excitation of ChIs (Aosaki and Kawaguchi, 1996; Govindaiah et al., 2010; Francis et al., 2019) and increases ACh release (Guevara Guzman et al., 1993; Francis et al., 2019; Belilos et al., 2023). Of the three neurokinin (NK) receptors, the NK1 receptor has the highest affinity to substance P among other variants (Pennefather et al., 2004). The NK1 receptor is a Gq coupled, G-protein receptor expressed in ChIs (Bolam et al., 1986; Aubry et al., 1993; Richardson et al., 2000; Pérez et al., 2007) and localized to somatostatin interneurons and ChI dendrites and somas (Martone et al., 1992; Pickel et al., 2000; Francis et al., 2019). Substance P causes a change in ChI activity in several ways via direct and indirect effects. Direct binding to the NK1 receptor results in persistent yet unidentified sodium current. When substance P is in high concentration ex vivo (250 nM – 1 μM), it can lead to long-lasting excitation lasting for minutes (Francis et al., 2019). However, shorter excitation due to substance P has been observed in vivo at physiological concentrations (Belilos et al., 2023). Indirectly, substance P can also suppress GABAA receptor-mediated IPSCs, resulting in increased ACh release from ChIs (Arenas et al., 1991; Govindaiah et al., 2010). This resulting effect of increased ACh release through direct or indirect mechanisms has been shown to have excitatory (Pakhotin and Bracci, 2007; Belilos et al., 2023) and inhibitory effects (Blomeley and Bracci, 2005) on excitatory transmission through ACh neurons.
Recently, Belilos et al. demonstrated that substance P, which causes lasting excitation of ChIs (Francis et al., 2019), is responsible for rebound ACh release during aversive foot shock. In fact, the amplitude of the rebound scaled with the conditioned freezing response elicited by cue/foot shock pairings. However, the foot shock, not the conditioning itself, produced the release. Therefore, substance P could be partially responsible for this response when an animal encounters a salient, aversive stimulus. It is also possible that substance P, which is released in response to rewarding stimuli (Commons, 2010; Vanderah and Sandweiss, 2015), could also be accountable for the rebound response in these reward-related conditions as well. However, this has yet to be demonstrated. In fact, activating D1-MSNs at higher frequency stimulation produces both a pause through the inhibition of ChIs via GABAA and GABAB receptors followed by excitation via the NK1 receptor (Francis et al., 2019). It remains to be seen if other excitatory peptides or disinhibition mechanisms may also provide a source of excitation during the rebound. Other neuromodulators and peptides that could participate in the response are discussed below.
4 Other neurotransmitters and neuromodulators controlling cholinergic activity
ChIs express many other neurotransmitter, neuromodulator, and neuropeptide receptors, and their activity elevates or suppresses tonic spontaneous firing rates, fine-tuning striatal circuitry. In some cases, the functional effect on activity is not fully understood. However, unlike most other neuromodulators, peptides provide a broad signal and can signal through volume transmission, which can spatially synchronize the activity of ChIs and other neurons. While the role of these neuromodulators in the conditioned pause response is not known, these factors may be shown to be involved in phasic ChI responses in the future.
4.1 Serotonin
The raphe nuclei send dense serotonergic projections to the striatum (Steinbusch, 1981; Lavoie and Parent, 1990). ChIs express several serotonin [5-HydroxyTryptamine (5-HT)] receptor subtypes, including 5-HT1, 5-HT2C, 5-HT6, and 5HT7, which modulate potassium ion currents and enhance neuronal excitability (Rada et al., 1993; Blomeley and Bracci, 2005; Bonsi et al., 2007). Immunolabeling studies have detected greater expression of 5-HT1A and 5-HT1B on ChIs of the ventral striatum, with little to no detection of these receptors in the DSt. Conversely, 5-HT7 receptors are more abundantly expressed in the DSt, while 5-HT5A receptors show equal expression in both regions (Steinbusch, 1981; Virk et al., 2016). In rats, in vivo infusion of 5-HT in NAc results in decreased ACh release, which was blocked by 5-HT1 antagonist (Rada et al., 1993). Upon serotonin (5-HT) application, ChIs in the DSt are depolarized, whereas those in the ventral striatum show hyperpolarization (Virk et al., 2016). Serotonin-mediated ChI depolarization may be due to 5-HT2 receptors as bath application of selective agonist (α-methyl-5-HT) for 5-HT2 receptor significantly increased spontaneous firing in ChIs and is blocked by 5-HT2 antagonist (Blomeley and Bracci, 2005). Rapid serotonin release may provide another signal to directly depolarize or hyperpolarize ChIs during phasic responses, albeit in a region-selective and concentration-dependent manner.
4.2 Opioids
ChIs express the mu-opioid receptor (MOR) and delta-opioid receptor (DOR), members of the GPCR family. Activation of these results in decreased spontaneous firing, resulting in reduced ACh release (Mulder et al., 1984; Svingos et al., 2001; Jabourian et al., 2005; Pérez et al., 2007; Arttamangkul et al., 2021). The double immunostaining experiment indicates MOR expression is restricted to ChIs in limbic/prefrontal territory but not in the sensorimotor area of the DSt. MOR is also reported to have diurnal variation in its expression, which is higher in the afternoon (last phase of light period) than in the morning (start of light period) in rats (Jabourian et al., 2005), which corresponds to the sleep phase of the rodents. In situ hybridization of DOR mRNA is seen in ChIs in the striatum (Le Moine et al., 1994; Svingos et al., 1998; Heath et al., 2018). Substance P induces delta receptor accumulation in the somatic area of ChIs in the NAc shell via the activation of NK1R (Heath et al., 2018). In addition, a GFP-tagged DOR knock-in mouse showed functionally expressed DOR, indicating its postsynaptic localization on ChIs (Le Moine et al., 1994; Scherrer et al., 2006). The inhibition observed in the current–voltage relationship for MOR suggests that inward rectification mediated by the GIRK channel drives inhibition (Britt and McGehee, 2008). Receptor desensitization occurs following simultaneous activation of MOR and DOR, but agonist-induced internalization is observed only for DOR (Faget et al., 2012; Arttamangkul et al., 2021). This finding indicates that MOR and DOR receptors may function independently of one another and may be activated in certain conditions. In addition, dynorphin, which is released from D1-MSNs and acts on ChIs via the kappa opioid receptor (KOR) can excite ChIs at low concentrations (Crain and Shen, 1996), and high concentrations can inhibit ChIs (Gross et al., 1990). Overall, activation of these receptors within the striatum may act as a brake on ChI activity, especially when released at higher levels. However, rapid, phasic changes in ChI activity by opioids is still not well understood.
4.3 Corticotrophin releasing factor
CRF is released into NAc in response to salient environmental stimuli, such as stress and novel objects (Lemos et al., 2012; Holly et al., 2016; Williams et al., 2022). CRF acts via CRF receptor type-1 (CRF-R1) and type-2 (CRF-R2), which are broadly expressed across the brain (Van Pett et al., 2000). However, RNAscope in situ hybridization has shown that Crh 1 mRNA is expressed in ChAT-positive neurons, while Crh 2 is absent in these regions (Van Pett et al., 2000; Lemos et al., 2019). CRF is expressed in a heterogenous population of MSNs in the ventral portion of the striatum (Lemos et al., 2019) and can be released by collaterals (Eckenwiler et al., 2024). Notably, CRF mRNA expression is high in the NAc but lacking in the DSt (Lemos et al., 2019), suggesting that regional differences in the expression may contribute to behavioral regulation. Activation of CRF receptors leads to cAMP-dependent activation of ChIs, elevating ACh levels as demonstrated through in vivo microdialysis in the NAc (Chen et al., 2012). Bath application of CRF enhances the spontaneous firing of ChIs across different regions and has been observed to robustly increase ChI firing in the NAc shell (Lemos et al., 2019; Ingebretson et al., 2024) and DSt (Lemos et al., 2019). Therefore, CRF may be able to alter several components of the phasic response, including DA release, or provide direct excitation of ChIs like substance P.
4.4 Galanin, leptin, and insulin
Galanin, Leptin, and Insulin are peptides that interact and regulate several processes related to feeding (Morton et al., 2006; Wu et al., 2012; Myers and Olson, 2014; Le et al., 2020; Affinati et al., 2023; Dornbush and Aeddula, 2024). Galanin, a 29 (mouse) or 30 (human) amino acid peptide primarily co-released mainly from neuronal populations in the locus coeruleus region along with other regions, has been linked to stress regulation, and conditions like depression, anxiety, and substance abuse (Weiss et al., 2006; Kuteeva et al., 2008; Juhasz et al., 2014; Hökfelt et al., 2018; Genders et al., 2020). Its action is mediated through galanin receptors (GALRs) divided into three subtypes GALR1, GALR2, and GALR3 (Wang and Gustafson, 1998; Branchek et al., 2000). In situ hybridization studies have shown that GALR1 mRNA is expressed in the striatum, with a distribution pattern resembling that of ChIs in the NAc (Zachariou, 2000). Galanin can stimulate striatal ACh release via GALRs on ChIs, which could be mediated by GALR1 (Ogren and Pramanik, 1991; Pramanik and Ogren, 1993; Zachariou, 2000). This effect depends on the anesthesia state of the animal, as awake-behaving rodents displayed decreased ACh release (Antoniou et al., 1997).
Leptin, a 146 amino acid peptide hormone released from adipose tissue, activates leptin receptors (LepR), releasing DA via nAChR (β2 containing) mechanisms (Wada et al., 2014; Mancini et al., 2022), by increasing ChI excitability. ChIs also express insulin receptors on ChIs (Stouffer et al., 2015), and, unlike Leptin, insulin, a 51 amino acids peptide, is reported to be expressed by some neuronal populations in brain, but their projections to the striatum are not known (Eerola et al., 2022; Dakic et al., 2023). Regardless of the source, peripheral or central, insulin enhances ChI excitability in all striatal subcompartments (Patel et al., 2023). This enhanced activity can also cause DA release via nAChR signaling. Therefore, these peptides may be a candidate for activity during rebound, particularly in the context of feeding and reward.
4.5 Estrogen
Estrogen exerts its effects in the CNS primarily via receptors ERα and ERβ (Eyster, 2016). Sex-dependent activity patterns have been observed in ChIs in the NAc (Ingebretson et al., 2024), suggesting ChI tonic activity may be regulated by estrogen signaling. However, direct application of 17β-estradiol has minimal to no effect on ChI spontaneous firing activity (Kövesdi et al., 2023). Using RNAscope in situ hybridization, estrogen receptors have been detected in subpopulations of ChIs in the striatum (Kövesdi et al., 2023). Ultrastructural analysis further indicates that ERα is expressed on ChIs at lower levels (Almey et al., 2012). Although estrogen receptors are expressed on ChIs, estrogen interacts with the striatal dopaminergic system, indirectly influencing ACh’s local release (Euvrard et al., 1979).
5 Outcomes of altering ChI activity and ACh release
Temporal signaling allows for synchronous and broad cholinergic signaling through large areas of the striatum, which controls the activity of MSNs or facilitates short-term or long-term plasticity in MSNs, thus controlling responses to salient environmental stimuli. Temporally selective, acute release of ACh alters intrinsic excitability. During a synchronous ChI pause, less ACh is released leading to diminished post-synaptic M1 receptor signaling and MSNs undergo less additional depolarization in the upstate (Zucca et al., 2018). This depolarization suppression effectively diminishes activity of MSNs when the membrane potential is near the action potential threshold. In vivo, it has been shown that activation of ChIs disrupts spiking of MSNs by causing a suppression in activity (Crittenden et al., 2017), potentially through the same mechanism. Oppositely, repeated activation of ChIs can lead to a lasting, M1 receptor-dependent increase in MSN excitability (Lv et al., 2017), suggesting a bidirectional control of MSN excitability by M1 receptor signaling putatively through suppression or disinhibition of Kv4 potassium channel currents (Nakamura et al., 1997) or N- and L-type calcium channels (Perez-Burgos et al., 2008; Howe and Surmeier, 1995). Overall, these results indicate that M1 receptor activation by suppression or enhancement of ACh release can oppositely control intrinsic excitability of MSNs.
ACh has profound effects on excitation within the striatum and altering ChI activity phasically influences the outcome of excitatory input via dendritic excitability and long-term plasticity including LTP and LTD of excitatory transmission which is dependent on receptor subtype activation (Oldenburg and Ding, 2011). Phasic activity of ChIs can activate pre-synaptic M2 and M4 receptors which are expressed on glutamatergic afferents, suppressing glutamate release from cortical terminals on MSNs (Pancani et al., 2014; Hersch et al., 1994; Barral et al., 1999; Pakhotin and Bracci, 2007; Ding et al., 2010). Aligned with these findings, M2 receptor inhibition can enhance the expression of LTP on MSNs (Calabresi et al., 1998b). Other pre-synaptic ACh receptors can modulate glutamate release as well. Nicotinic receptor activation facilitates or causes glutamate release onto MSNs depending on the subunit (Howe et al., 2016; Assous, 2021). It remains unclear how M2, M4, and nicotinic receptor activation temporally locks glutamate release on MSNs to the phasic burst-pause-rebound response. However, it is known that nicotinic receptors can promote the release of glutamate on various GABAergic interneuron populations and therefore provide inhibition of MSNs. Pause excitation sequences lead to inhibition of MSNs in vivo and ex vivo via nAChR-mediated activation of NPY- Nerve growth factor (NGF) GABA neurons (English et al., 2012; Faust et al., 2016). Further, inhibition of ACh release during a pause suppresses cortical glutamate input. This excitatory suppression is due to a decrease in post-synaptic M1 receptor signaling which decreases the dendritic excitability of MSNs and suppress the size of excitatory responses (Wang et al., 2006). An increase in M1 receptor signaling, however, enhances dendritic excitability by suppressing Kir2 dendritic inhibition selectively in D2-MSNs (Shen et al., 2007) and may play a role in enhancing excitation to D2-MSNs during the thalamic-mediated burst-pause response (Ding et al., 2010). These outcomes provide M1, M2, M4, and nicotinic receptors with the ability to acutely modulate excitation during phasic bursts and pauses. It is still unclear how pre-synaptic and post-synaptic mechanisms coordinate glutamatergic transmission to provide an overall increase or decrease in glutamatergic signaling. Based on the current literature, we predict that post-synaptic signaling may play a larger role in long-term synaptic plasticity.
In line with changes in dendritic excitability, suppression of M1 receptor signaling restores endocannabinoid LTD induction even in the absence of D2 receptor signaling, which is critically involved in corticostriatal LTD (Wang et al., 2006). Induction of LTP in conditions where NMDA receptors are active at resting states (i.e., low magnesium conditions), requires M1 receptor signaling (Calabresi et al., 1999). Often, ACh release inversely covaries with DA release to promote plasticity on MSN subtypes. A coordinated decrease in ACh release in the DSt during the pause with matched DA release is sufficient to induce sensory-evoked LTP on MSNs in vivo (Reynolds et al., 2022). When ACh and DA release are mismatched, that is when ACh and DA are both high, MSNs can display sensory-evoked LTD. However, the mismatch may provide subregion-selective information and could vary across these regions (Duhne et al., 2024). Cell-type activation of mAChRs selects the expression of plasticity. In D1-MSNs, M4 receptor activation promotes excitatory LTD on D1-MSNs and blocks LTP (Shen et al., 2015). Pauses in DA during the putatively high activity of ACh have been shown to enhance spine formation and LTP during discrimination learning selectively on NAc D2-MSNs (Schulz and Reynolds, 2013; Iino et al., 2020), likely through muscarinic receptor activation (Calabresi et al., 1998b; Francis et al., 2019; Poppi et al., 2021). In addition, while the pause itself is important for inducing plasticity, the rebound facilitates plasticity in the NAc in response to aversive stimuli. The rebound correlates with aversive learning in mice, is substance P dependent, and facilitates LTP selectively on D2-MSNs (Belilos et al., 2023). These DA pauses and ACh mediated D2-MSN plasticity may be overlapping mechanisms to facilitate excitatory LTP on D2-MSNs. In contrast, pauses or transient decreases in ACh release can allow for corticostriatal LTP on D1-MSNs (Laverne et al., 2022). This suggests these responses could aid in facilitating cell-type selective excitatory plasticity on the major projection neurons of the striatum. Due to the multi-factor requirement of each phase, eliminating one molecule that participates in this response can have drastic effects on the response. This would provide precise, input-specific, cell-type specific, and striatal sub-region selective control over ACh-dependent plasticity in the striatum.
6 Future directions
The conditioned pause response is one of the key mechanisms ChIs can use to facilitate changes in MSN activity and plasticity. We summarize how glutamatergic inputs from cortical and thalamic inputs can facilitate an initial burst followed by rapid release of DA and GABA, mediating a pause response and a rebound mediated by intrinsic properties and peptide release. The mechanisms of the burst and pause are quite clear and very well studied. However, the rebound and what drives the extended activation of the rebound is much less well studied. Information about the temporal release of molecules, particularly across striatal sub-compartments, would provide a better understanding of how selective plasticity drives behavioral outcomes in response to phasic changes in ChI activity.
Along with collecting information about mechanisms controlling each phase of the conditioned pause response, we have discussed one peptide, substance P, which participates in mediating the rebound response (Belilos et al., 2023). Neuropeptides are known to modulate various aspects of the neuronal function, often in a context and time-dependent manner, suggesting that they could critically shape the response of ChIs to salient stimuli. The peptides we review here excite ChIs and can facilitate the release of DA. We suggest other neurotransmitters, neuromodulators, and neuropeptides may facilitate the long rebound observed following the pause, potentially in a context-dependent manner. This would require additional studies to assess how other peptides may play a temporal-dependent role in the phasic excitation of ChIs. These studies should assess neuropeptide release in coordination with classical neurotransmitter release on ChIs, including glutamate, which facilitates the release of these peptides by strongly stimulating local striatal neurons. Furthermore, understanding how other peptides that activate ChIs may be involved in the rebound response would provide a better understanding of how stimuli with different valences or contexts may enhance some of these responses over others.
Although few, striatal ChIs are critical for the striatal output, guiding short-term and long-term changes in projection neuron output. Yet, refined control of ChI activity remains partially understood. The studies described here indicate that altering the ChI firing rate is multifaceted, requiring a variety of intrinsic ChI properties and extrinsic neurotransmitters and neuromodulators. Furthermore, striatal subregion-specific differences and cell-type-specific differences are observed. Understanding the control of ChI activity provides a foundation for understanding how the striatum engages in associative responses and learning underlying learning, reward, and aversion.
Author contributions
DR: Writing – original draft, Writing – review & editing. TF: Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work is supported by grants from the National Institute on Mental Health at the National Institutes of Mental Health R00MH123673 (TCF) and R21MH135207 (TCF), the Brain and Behavior Research Foundation Early Investigator NARSAD award (TCF), and the College of Pharmacy at the University of South Carolina.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abbondanza, A., Bas, I. R., Modrak, M., Capek, M., Minich, J., Tyshkevich, A., et al. (2022). Nicotinic acetylcholine receptors expressed by striatal interneurons inhibit striatal activity and control striatal-dependent behaviors. J. Neurosci. 42, 2786–2803. doi: 10.1523/JNEUROSCI.1627-21.2022
Affinati, A. H., Elias, C. F., Olson, D. P., and Myers, M. G. (2023). “Brain regulation of feeding and energy homeostasis” in Metabolic syndrome: A comprehensive textbook (Cham: Springer International Publishing), 1–19.
Ahmed, N. Y., Knowles, R., and Dehorter, N. (2019). New insights into cholinergic neuron diversity. Front. Mol. Neurosci. 12:204. doi: 10.3389/fnmol.2019.00204
Albuquerque, E. X., Pereira, E. F. R., Alkondon, M., and Rogers, S. W. (2009). Mammalian icotinic acetylcholine receptors: from structure to function. Physiol. Rev. 89, 73–120. doi: 10.1152/physrev.00015.2008
Al-Hasani, R., Gowrishankar, R., Schmitz, G. P., Pedersen, C. E., Marcus, D. J., Shirley, S. E., et al. (2021). Ventral tegmental area GABAergic inhibition of cholinergic interneurons in the ventral nucleus accumbens shell promotes reward reinforcement. Nat. Neurosci. 24, 1414–1428. doi: 10.1038/s41593-021-00898-2
Allaway, K. C., and Machold, R. (2017). Developmental specification of forebrain cholinergic neurons. Dev. Biol. 421, 1–7. doi: 10.1016/j.ydbio.2016.11.007
Almey, A., Filardo, E. J., Milner, T. A., and Brake, W. G. (2012). Estrogen receptors are found in glia and at extranuclear neuronal sites in the dorsal striatum of female rats: evidence for cholinergic but not dopaminergic colocalization. Endocrinology 153, 5373–5383. doi: 10.1210/en.2012-1458
Antoniou, K., Kehr, J., Snitt, K., and Ogren, S. O. (1997). Differential effects of the neuropeptide galanin on striatal acetylcholine release in anaesthetized and awake rats. Br. J. Pharmacol. 121, 1180–1186. doi: 10.1038/sj.bjp.0701233
Aosaki, T., Graybiel, A. M., and Kimura, M. (1994a). Effect of the nigrostriatal dopamine system on acquired neural responses in the striatum of behaving monkeys. Science 265, 412–415. doi: 10.1126/science.8023166
Aosaki, T., and Kawaguchi, Y. (1996). Actions of substance P on rat neostriatal neurons in vitro. J. Neurosci. 16, 5141–5153. doi: 10.1523/JNEUROSCI.16-16-05141.1996
Aosaki, T., Kiuchi, K., and Kawaguchi, Y. (1998). Dopamine D1-like receptor activation excites rat striatal large aspiny neurons in vitro. J. Neurosci. 18, 5180–5190. doi: 10.1523/JNEUROSCI.18-14-05180.1998
Aosaki, T., Miura, M., Suzuki, T., Nishimura, K., and Masuda, M. (2010). Acetylcholine-dopamine balance hypothesis in the striatum: An update. Geriatr Gerontol Int 10, S148–S157. doi: 10.1111/j.1447-0594.2010.00588.x
Aosaki, T., Tsubokawa, H., Ishida, A., Watanabe, K., Graybiel, A., and Kimura, M. (1994b). Responses of tonically active neurons in the primate’s striatum undergo systematic changes during behavioral sensorimotor conditioning. J. Neurosci. 14, 3969–3984. doi: 10.1523/JNEUROSCI.14-06-03969.1994
Apicella, P., Legallet, E., and Trouche, E. (1997). Responses of tonically discharging neurons in the monkey striatum to primary rewards delivered during different behavioral states. Exp. Brain Res. 116, 456–466. doi: 10.1007/pl00005773
Arenas, E., Alberch, J., Perez-Navarro, E., Solsona, C., and Marsal, J. (1991). Neurokinin receptors differentially mediate endogenous acetylcholine release evoked by tachykinins in the neostriatum. J. Neurosci. 11, 2332–2338. doi: 10.1523/JNEUROSCI.11-08-02332.1991
Arttamangkul, S., Platt, E. J., Carroll, J., and Farrens, D. (2021). Functional independence of endogenous μ- and δ-opioid receptors co-expressed in cholinergic interneurons. eLife 10:e69740. doi: 10.7554/eLife.69740
Assous, M. (2021). Striatal cholinergic transmission: focus on nicotinic receptors' influence in striatal circuits. Eur. J. Neurosci. 53, 2421–2442. doi: 10.1111/ejn.15135
Assous, M., and Tepper, J. M. (2019). Cortical and thalamic inputs exert cell type-specific feedforward inhibition on striatal GABAergic interneurons. J. Neurosci. Res. 97, 1491–1502. doi: 10.1002/jnr.24444
Atallah, H. E., McCool, A. D., Howe, M. W., and Graybiel, A. M. (2014). Neurons in the ventral striatum exhibit cell-type-specific representations of outcome during learning. Neuron 82, 1145–1156. doi: 10.1016/j.neuron.2014.04.021
Aubry, J.-M., Schulz, M.-F., Pagliusi, S., Schulz, P., and Kiss, J. Z. (1993). Coexpression of dopamine D2 and substance P (neurokinin-1) receptor messenger RNAs by a subpopulation of cholinergic neurons in the rat striatum. Neuroscience 53, 417–424. doi: 10.1016/0306-4522(93)90205-t
Baimel, C., Jang, E., Scudder, S. L., Manoocheri, K., and Carter, A. G. (2022). Hippocampal-evoked inhibition of cholinergic interneurons in the nucleus accumbens. Cell Rep. 40:111042. doi: 10.1016/j.celrep.2022.111042
Baldi, G., Russi, G., Nannini, L., Vezzani, A., and Consolo, S. (1995). Trans-synaptic modulation of striatal ACh release in vivo by the Parafascicular thalamic nucleus. Eur. J. Neurosci. 7, 1117–1120. doi: 10.1111/j.1460-9568.1995.tb01100.x
Barbeau, A. (1962). The pathogenesis of Parkinson’s disease: a new hypothesis. Can. Med. Assoc. J. 87, 802–807
Barral, J., Galarraga, E., and Bargas, J. (1999). Muscarinic presynaptic inhibition of neostriatal glutamatergic afferents is mediated by Q-type Ca2+ channels. Brain Res. Bull. 49, 285–289. doi: 10.1016/S0361-9230(99)00061-1
Beier, K. (2022). Modified viral-genetic mapping reveals local and global connectivity relationships of ventral tegmental area dopamine cells. eLife 11:e76886. doi: 10.7554/eLife.76886
Beier, K. T., Steinberg, E. E., Deloach, K. E., Xie, S., Miyamichi, K., Schwarz, L., et al. (2015). Circuit architecture of VTA dopamine neurons revealed by systematic input-output mapping. Cell 162, 622–634. doi: 10.1016/j.cell.2015.07.015
Belilos, A., Gray, C., Sanders, C., Black, D., Mays, E., Richie, C., et al. (2023). Nucleus accumbens local circuit for cue-dependent aversive learning. Cell Rep. 42:113488. doi: 10.1016/j.celrep.2023.113488
Bell, M. I., Richardson, P. J., and Lee, K. (2002). Functional and molecular characterization of metabotropic glutamate receptors expressed in rat striatal cholinergic interneurones. J. Neurochem. 81, 142–149. doi: 10.1046/j.1471-4159.2002.00815.x
Bennett, B. D., Callaway, J. C., and Wilson, C. J. (2000). Intrinsic membrane properties underlying spontaneous tonic firing in neostriatal cholinergic interneurons. J. Neurosci. 20, 8493–8503. doi: 10.1523/JNEUROSCI.20-22-08493.2000
Bennett, B. D., and Wilson, C. J. (1998). Synaptic regulation of action potential timing in neostriatal cholinergic interneurons. J. Neurosci. 18, 8539–8549. doi: 10.1523/JNEUROSCI.18-20-08539.1998
Bennett, B. D., and Wilson, C. J. (1999). Spontaneous activity of neostriatal cholinergic interneurons in vitro. J. Neurosci. 19, 5586–5596. doi: 10.1523/JNEUROSCI.19-13-05586.1999
Berendse, H. W., and Groenewegen, H. J. (1990). Organization of the thalamostriatal projections in the rat, with special emphasis on the ventral striatum. J. Comp. Neurol. 299, 187–228. doi: 10.1002/cne.902990206
Bergson, C., Mrzljak, L., Smiley, J., Pappy, M., Levenson, R., and Goldman-Rakic, P. (1995). Regional, cellular, and subcellular variations in the distribution of D1 and D5 dopamine receptors in primate brain. J. Neurosci. 15, 7821–7836. doi: 10.1523/JNEUROSCI.15-12-07821.1995
Berlanga, M. L., Olsen, C. M., Chen, V., Ikegami, A., Herring, B. E., Duvauchelle, C. L., et al. (2003). Cholinergic interneurons of the nucleus accumbens and dorsal striatum are activated by the self-administration of cocaine. Neuroscience 120, 1149–1156. doi: 10.1016/s0306-4522(03)00378-6
Björklund, A., and Dunnett, S. B. (2007). Dopamine neuron systems in the brain: an update. Trends Neurosci. 30, 194–202. doi: 10.1016/j.tins.2007.03.006
Blomeley, C., and Bracci, E. (2005). Excitatory effects of serotonin on rat striatal cholinergic interneurones. J. Physiol. 569, 715–721. doi: 10.1113/jphysiol.2005.098269
Blomeley, C., and Bracci, E. (2008). Substance P depolarizes striatal projection neurons and facilitates their glutamatergic inputs. J. Physiol. 586, 2143–2155. doi: 10.1113/jphysiol.2007.148965
Boccalaro, I. L., Cristiá-Lara, L., Schwerdel, C., Fritschy, J. M., and Rubi, L. (2019). Cell type-specific distribution of GABAA receptor subtypes in the mouse dorsal striatum. J. Comp. Neurol. 527, 2030–2046. doi: 10.1002/cne.24665
Bolam, J. P., Ingham, C. A., Izzo, P. N., Levey, A. I., Rye, D. B., Smith, A. D., et al. (1986). Substance P-containing terminals in synaptic contact with cholinergic neurons in the neostriatum and basal forebrain: A double Immunocytochemical study in the rat. Brain Res. 397, 279–289. doi: 10.1016/0006-8993(86)90629-3
Bonsi, P., Cuomo, D., De Persis, C., Centonze, D., Bernardi, G., Calabresi, P., et al. (2005). Modulatory action of metabotropic glutamate receptor (mGluR) 5 on mGluR1 function in striatal cholinergic interneurons. Neuropharmacol 49, 104–13. doi: 10.1016/j.neuropharm.2005.05.012
Bonsi, P., Cuomo, D., Ding, J., Sciamanna, G., Ulrich, S., Tscherter, A., et al. (2007). Endogenous serotonin excites striatal cholinergic interneurons via the activation of 5-HT 2C, 5-HT6, and 5-HT7 serotonin receptors: implications for extrapyramidal side effects of serotonin reuptake inhibitors. Neuropsychopharmacology 32, 1840–1854. doi: 10.1038/sj.npp.1301294
Bonsi, P., Cuomo, D., Martella, G., Madeo, G., Schirinzi, T., Puglisi, F., et al. (2011). Centrality of striatal cholinergic transmission in basal ganglia function. Front. Neuroanat. 5:9268. doi: 10.3389/fnana.2011.00006
Branchek, T. A., Smith, K. E., Gerald, C., and Walker, M. W. (2000). Galanin receptor subtypes. Trends Pharmacol. Sci. 21, 109–117. doi: 10.1016/S0165-6147(00)01446-2
Breton, J. M., Charbit, A. R., Snyder, B. J., Fong, P. T. K., Dias, E. V., Himmels, P., et al. (2019). Relative contributions and mapping of ventral tegmental area dopamine and GABA neurons by projection target in the rat. J. Comp. Neurol. 527, 916–941. doi: 10.1002/cne.24572
Brimblecombe, K. R., and Cragg, S. J. (2015). Substance P weights striatal dopamine transmission differently within the striosome-matrix axis. J. Neurosci. 35, 9017–9023. doi: 10.1523/JNEUROSCI.0870-15.2015
Britt, J. P., and McGehee, D. S. (2008). Presynaptic opioid and nicotinic receptor modulation of dopamine overflow in the nucleus accumbens. J. Neurosci. 28, 1672–1681. doi: 10.1523/JNEUROSCI.4275-07.2008
Brown, D. A. (2018). Regulation of neural ion channels by muscarinic receptors. Neuropharmacology 136, 383–400. doi: 10.1016/j.neuropharm.2017.11.024
Brown, M. T. C., Tan, K. R., O’Connor, E. C., Nikonenko, I., Muller, D., and Lüscher, C. (2012). Ventral tegmental area GABA projections pause accumbal cholinergic interneurons to enhance associative learning. Nature 492, 452–456. doi: 10.1038/nature11657
Cachope, R., Mateo, Y., Mathur, B. N., Irving, J., Wang, H. L., Morales, M., et al. (2012). Selective activation of cholinergic interneurons enhances accumbal phasic dopamine release: setting the tone for reward processing. Cell Rep. 2, 33–41. doi: 10.1016/j.celrep.2012.05.011
Cai, Y., and Ford, C. P. (2018). Dopamine cells differentially regulate striatal cholinergic transmission across regions through corelease of dopamine and glutamate. Cell Rep. 25, 3148–3157.e3. doi: 10.1016/j.celrep.2018.11.053
Calabresi, P., Centonze, D., Gubellini, P., and Bernardi, G. (1999). Activation of M1-like muscarinic receptors is required for the induction of corticostriatal LTP. Neuropharmacology 38, 323–326. doi: 10.1016/s0028-3908(98)00199-3
Calabresi, P., Centonze, D., Gubellini, P., Pisani, A., and Bernardi, G. (1998a). Endogenous ACh enhances striatal NMDA-responses via M1-like muscarinic receptors and PKC activation. Eur. J. Neurosci. 10, 2887–2895. doi: 10.1111/j.1460-9568.1998.00294.x
Calabresi, P., Centonze, D., Gubellini, P., Pisani, A., and Bernardi, G. (1998b). Blockade of M2-like muscarinic receptors enhances long-term potentiation at corticostriatal synapses. Eur. J. Neurosci. 10, 3020–3023. doi: 10.1111/j.1460-9568.1998.00348.x
Calabresi, P., Centonze, D., Gubellini, P., Pisani, A., and Bernardi, G. (2000). Acetylcholine-mediated modulation of striatal function. Trends Neurosci. 23, 120–126. doi: 10.1016/S0166-2236(99)01501-5
Castello, J., Cortés, M., Malave, L., Kottmann, A., Sibley, D. R., Friedman, E., et al. (2020). The dopamine D5 receptor contributes to activation of cholinergic interneurons during L-DOPA induced dyskinesia. Sci. Rep. 10:2542. doi: 10.1038/s41598-020-59011-5
Castro, D. C., and Bruchas, M. R. (2019). A motivational and neuropeptidergic hub: anatomical and functional diversity within the nucleus accumbens shell. Neuron 102, 529–552. doi: 10.1016/j.neuron.2019.03.003
Caulfield, M. P., and Birdsall, N. J. (1998). International Union of Pharmacology. XVII. Classification of muscarinic acetylcholine receptors. Pharmacol. Rev. 50, 279–290. doi: 10.1016/S0031-6997(24)01362-0
Centonze, D., Grande, C., Usiello, A., Gubellini, P., Erbs, E., Martín, A. B., et al. (2003). Receptor subtypes involved in the pesynaptic and postsynaptic actions of dopamine on striatal interneurons. J. Neurosci. 23, 6245–6254. doi: 10.1523/JNEUROSCI.23-15-06245.2003
Chantranupong, L., Beron, C. C., Zimmer, J. A., Wen, M. J., Wang, W., and Sabatini, B. L. (2023). Dopamine and glutamate regulate striatal acetylcholine in decision-making. Nature 621, 577–585. doi: 10.1038/s41586-023-06492-9
Chapman, K. L., Vaswani, D., Hendry, N., Langmead, C. J., Kew, J. N. C., and Watson, J. M. (2011). The muscarinic M(4) receptor is the functionally predominant subtype in rat and mouse striatum as demonstrated using [(35)S] GTPγS binding. Eur. J. Pharmacol. 652, 1–6. doi: 10.1016/j.ejphar.2010.10.079
Chen, S. Y., Lu, K. M., Ko, H. A., Huang, T. H., Hao, J. H., Yan, Y. T., et al. (2020). Parcellation of the striatal complex into dorsal and ventral districts. PNAS 117, 7418–7429. doi: 10.1073/pnas.1921007117
Chen, Y. W., Rada, P. V., Bützler, B. P., Leibowitz, S. F., and Hoebel, B. G. (2012). Corticotropin-releasing factor in the nucleus accumbens shell induces swim depression, anxiety, and anhedonia along with changes in local dopamine/acetylcholine balance. Neuroscience 206, 155–166. doi: 10.1016/j.neuroscience.2011.12.009
Chuhma, N., Mingote, S., Moore, H., and Rayport, S. (2014). Dopamine neurons control striatal cholinergic neurons via regionally heterogeneous dopamine and glutamate signaling. Neuron 81, 901–912. doi: 10.1016/j.neuron.2013.12.027
Chuhma, N., Mingote, S., Yetnikoff, L., Kalmbach, A., Ma, T., Ztaou, S., et al. (2018). Dopamine neuron glutamate cotransmission evokes a delayed excitation in lateral dorsal striatal cholinergic interneurons. eLife 7:e39786. doi: 10.7554/eLife.39786
Chuhma, N., Oh, S. J., and Rayport, S. (2023). The dopamine neuron synaptic map in the striatum. Cell Rep. 42:112204. doi: 10.1016/j.celrep.2023.112204
Clarke, P. B., and Pert, A. (1985). Autoradiographic evidence for nicotine receptors on nigrostriatal and mesolimbic dopaminergic neurons. Brain Res. 348, 355–358. doi: 10.1016/0006-8993(85)90456-1
Colquhoun, D., Jonas, P., and Sakmann, B. (1992). Action of brief pulses of glutamate on AMPA/kainate receptors in patches from different neurons of rat hippocampal slices. J. Physiol. 458, 261–287. doi: 10.1113/jphysiol.1992.sp019417
Commons, K. G. (2010). Neuronal pathways linking substance P to drug addiction and stress. Brain Res. 1314, 175–182. doi: 10.1016/j.brainres.2009.11.014
Consolo, S., Baldi, G., Giorgi, S., and Nannini, L. (1996). The cerebral cortex and parafascicular thalamic nucleus facilitate in vivo acetylcholine release in the rat striatum through distinct glutamate receptor subtypes. Eur. J. Neurosci. 8, 2702–2710. doi: 10.1111/j.1460-9568.1996.tb01565.x
Cragg, S. J. (2006). Meaningful silences: how dopamine listens to the ACh pause. Trends Neurosci. 29, 125–131. doi: 10.1016/j.tins.2006.01.003
Crain, S. M., and Shen, K.-F. (1996). Modulatory effects of Gs-coupled excitatory opioid receptor functions on opioid analgesia, tolerance, and dependence*. Neurochem. Res. 21, 1347–1351. doi: 10.1007/BF02532375
Crittenden, J. R., Lacey, C. J., Weng, F.-J., Garrison, C. E., Gibson, D. J., Lin, Y., et al. (2017). Striatal cholinergic interneurons modulates spike-timing in striosomes and matrix by an amphetamine-sensitive mechanism. Front. Neuroanat. 11:20. doi: 10.3389/fnana.2017.00020
Dakic, T., Jevdjovic, T., Lakic, I., Ruzicic, A., Jasnic, N., Djurasevic, S., et al. (2023). The expression of insulin in the central nervous system: what have we learned so far? Int. J. Mol. Sci. 24:6586. doi: 10.3390/ijms24076586
Dani, J. A. (2015). Neuronal nicotinic acetylcholine receptor structure and function and response to nicotine. Int. Rev. Neurobiol. 124, 3–19. doi: 10.1016/bs.irn.2015.07.001
Dautan, D., Huerta-Ocampo, I., Witten, I. B., Deisseroth, K., Bolam, J. P., Gerdjikov, T., et al. (2014). A major external source of cholinergic innervation of the striatum and nucleus accumbens originates in the brainstem. J. Neurosci. 34, 4509–4518. doi: 10.1523/JNEUROSCI.5071-13.2014
DeBoer, P., and Westerink, B. H. (1994). GABAergic modulation of striatal cholinergic interneurons: an in vivo microdialysis study. J. Neurochem. 62, 70–75. doi: 10.1046/j.1471-4159.1994.62010070.x
de Jesus Aceves Buendia, J., Tiroshi, L., Chiu, W. H., and Goldberg, J. A. (2019). Selective remodeling of glutamatergic transmission to striatal cholinergic interneurons after dopamine depletion. Eur. J. Neurosci. 49, 824–833. doi: 10.1111/ejn.13715
Deng, Y. P., Xie, J. P., Wang, H. B., Lei, W. L., Chen, Q., and Reiner, A. (2007). Differential localization of the GluR1 and GluR2 subunits of the AMPA-type glutamate receptor among striatal neuron types in rats. J. Chem. Neuroanat. 33, 167–192. doi: 10.1016/j.jchemneu.2007.02.008
Dimova, R., Vuillet, J., Nieoullon, A., and Goff, L. K.-L. (1993). Ultrastructural features of the choline acetyltransferase-containing neurons and relationships with nigral dopaminergic and cortical afferent pathways in the rat striatum. Neuroscience 53, 1059–1071. doi: 10.1016/0306-4522(93)90489-3
Ding, J., An, D., Liao, W., Wu, G., Xu, Q., Zhou, D., et al. (2014). Abnormal functional connectivity density in psychogenic non-epileptic seizures. Epilepsy Res. 108, 1184–1194. doi: 10.1016/j.eplepsyres.2014.05.006
Ding, J. B., Guzman, J. N., Peterson, J. D., Goldberg, J. A., and Surmeier, D. J. (2010). Thalamic gating of corticostriatal signaling by cholinergic interneurons. Neuron 67, 294–307. doi: 10.1016/j.neuron.2010.06.017
Ding, J., Guzman, J. N., Tkatch, T., Chen, S., Goldberg, J. A., Ebert, P. J., et al. (2006). RGS4-dependent attenuation of M4 autoreceptor function in striatal cholinergic interneurons following dopamine depletion. Nat. Neurosci. 9, 832–842. doi: 10.1038/nn1700
Doig, N. M., Magill, P. J., Apicella, P., Bolam, J. P., and Sharott, A. (2014). Cortical and thalamic excitation mediate the multiphasic responses of striatal cholinergic interneurons to motivationally salient stimuli. J. Neurosci. 34, 3101–3117. doi: 10.1523/JNEUROSCI.4627-13.2014
Dornbush, S., and Aeddula, N. R. (2024). Physiology, leptin. In: StatPearls [internet]. Treasure Island, FL: StatPearls Publishing; (2024). Available at: http://www.ncbi.nlm.nih.gov/pubmed/8923829 (Accessed November 11, 2024).
Dorst, M. C., Tokarska, A., Zhou, M., Lee, K., Stagkourakis, S., Broberger, C., et al. (2020). Polysynaptic inhibition between striatal cholinergic interneurons shapes their network activity patterns in a dopamine-dependent manner. Nat. Commun. 11:5113. doi: 10.1038/s41467-020-18882-y
Duhne, M., Mohebi, A., Kim, K., Pelattini, L., and Berke, J. D. (2024). A mismatch between striatal cholinergic pauses and dopaminergic reward prediction errors. PNAS 121:e2410828121. doi: 10.1073/pnas.2410828121
Eckenwiler, E. A., Ingebretson, A. E., Stolley, J. J., Fusaro, M. A., Romportl, A. M., Ross, J. M., et al. (2024). CRF release from a unique subpopulation of accumbal neurons constrains action-outcome acquisition in reward learning. bioRxiv. doi: 10.1101/2023.11.16.567495
Eerola, K., Longo, F., Reinbothe, T. M., Richard, J. E., Shevchouk, O. T., López-Ferreras, L., et al. (2022). Hindbrain insulin controls feeding behavior. Mol. Metab. 66:101614. doi: 10.1016/j.molmet.2022.101614
English, D. F., Ibanez-Sandoval, O., Stark, E., Tecuapetla, F., Buzsáki, G., Deisseroth, K., et al. (2012). GABAergic circuits mediate the reinforcement-related signals of striatal cholinergic interneurons. Nat. Neurosci. 15, 123–130. doi: 10.1038/nn.2984
Erro, M., Lanciego, J., Arribas, J., and Giménez-Amaya, J. (2001). Striatal input from the ventrobasal complex of the rat thalamus. Histochem. Cell Biol. 115, 447–454. doi: 10.1007/s004180100273
Euvrard, C., Labrie, F., and Boissier, J. R. (1979). Effect of estrogen on changes in the activity of striatal cholinergic neurons induced by DA drugs. Brain Res. 169, 215–220. doi: 10.1016/0006-8993(79)90392-5
Exley, R., and Cragg, S. J. (2008). Presynaptic nicotinic receptors: a dynamic and diverse cholinergic filter of striatal dopamine neurotransmission. Br. J. Pharmacol. 153, S283–S297. doi: 10.1038/sj.bjp.0707510
Exley, R., McIntosh, J. M., Marks, M. J., Maskos, U., and Cragg, S. J. (2012). Striatal α5 nicotinic receptor subunit regulates dopamine transmission in dorsal striatum. J. Neurosci. 32, 2352–2356. doi: 10.1523/JNEUROSCI.4985-11.2012
Eyster, K. M. (2016). The estrogen receptors: An overview from different perspectives. New York, NY: Humana Press, 1–10.
Faget, L., Erbs, E., le Merrer, J., Scherrer, G., Matifas, A., Benturquia, N., et al. (2012). In vivo visualization of delta opioid receptors upon physiological activation uncovers a distinct internalization profile. J. Neurosci. 32, 7301–7310. doi: 10.1523/JNEUROSCI.0185-12.2012
Faust, T. W., Assous, M., Tepper, J. M., and Koós, T. (2016). Neostriatal GABAergic interneurons mediate cholinergic inhibition of spiny projection neurons. J. Neurosci. 36, 9505–9511. doi: 10.1523/JNEUROSCI.0466-16.2016
Feduccia, A. A., Chatterjee, S., and Bartlett, S. E. (2012). Neuronal nicotinic acetylcholine receptors: neuroplastic changes underlying alcohol and nicotine addictions. Front. Mol. Neurosci. 5:27779. doi: 10.3389/fnmol.2012.00083
Feng, Z. J., Zhang, X., and Chergui, K. (2014). Allosteric modulation of NMDA receptors alters neurotransmission in the striatum of a mouse model of Parkinson’s disease. Exp. Neurol. 255, 154–160. doi: 10.1016/j.expneurol.2014.03.001
Francis, T. C., Yano, H., Demarest, T. G., Shen, H., and Bonci, A. (2019). High-frequency activation of nucleus accumbens D1-MSNs drives excitatorypPotentiation on D2-MSNs. Neuron 103, 432–444.e3. doi: 10.1016/j.neuron.2019.05.031
Fremeau, R. T., Kam, K., Qureshi, T., Johnson, J., Copenhagen, D. R., Storm-Mathisen, J., et al. (2004). Vesicular glutamate transporters 1 and 2 target to functionally distinct synaptic release sites. Science 304, 1815–1819. doi: 10.1126/science.1097468
Fuxe, K., Dahlström, A. B., Jonsson, G., Marcellino, D., Guescini, M., Dam, M., et al. (2010). The discovery of central monoamine neurons gave volume transmission to the wired brain. Prog. Neurobiol. 90, 82–100. doi: 10.1016/j.pneurobio.2009.10.012
Galarraga, E., Hernandez-Lopez, S., Reyes, A., Miranda, I., Bermudez-Rattoni, F., Vilchis, C., et al. (1999). Cholinergic modulation of neostriatal output: a functional antagonism between different types of muscarinic receptors. J. Neurosci. 19, 3629–3638. doi: 10.1523/JNEUROSCI.19-09-03629.1999
Gallo, E. F., Greenwald, J., Yeisley, J., Teboul, E., Martyniuk, K. M., Villarin, J. M., et al. (2022). Dopamine D2 receptors modulate the cholinergic pause and inhibitory learning. Mol. Psychiatry 27, 1502–1514. doi: 10.1038/s41380-021-01364-y
Genders, S. G., Scheller, K. J., and Djouma, E. (2020). Neuropeptide modulation of addiction: focus on galanin. Neurosci. Biobehav. Rev. 110, 133–149. doi: 10.1016/j.neubiorev.2018.06.021
Gerfen, C. R., and Bolam, J. P. (2016). “The neuroanatomical Organization of the Basal Ganglia” in Handbook of basal ganglia structure and function. 2nd ed (Amsterdam: Elsevier/Academic Press), 3–32.
Giorguieff, M. F., Le Floc’h, M. L., Westfall, T. C., Glowinski, J., and Besson, M. J. (1976). Nicotinic effect of acetylcholine on the release of newly synthesized [3H] dopamine in rat striatal slices and cat caudate nucleus. Brain Res. 106, 117–131. doi: 10.1016/0006-8993(76)90077-9
Goldberg, J. A., Teagarden, M. A., Foehring, R. C., and Wilson, C. J. (2009). Nonequilibrium calcium dynamics regulate the autonomous firing pattern of rat striatal cholinergic interneurons. J. Neurosci. 29, 8396–8407. doi: 10.1523/JNEUROSCI.5582-08.2009
Goldberg, J. A., and Wilson, C. J. (2005). Control of spontaneous firing patterns by the selective coupling of calcium currents to calcium-activated potassium currents in striatal cholinergic interneurons. J. Neurosci. 25, 10230–10238. doi: 10.1523/JNEUROSCI.2734-05.2005
Goldberg, J. A., and Wilson, C. J. (2016). “The cholinergic interneuron of the striatum” in Handbook of basal ganglia structure and function. 2nd ed (Amsterdam: Elsevier/Academic Pres), 137–155.
Gonzales, K. K., Pare, J., Wichmann, T., and Smith, Y. (2013). GABAergic inputs from direct and indirect striatal projection neurons onto cholinergic interneurons in the primate putamen. J. Comp. Neurol. 521, 2502–2522. doi: 10.1002/cne.23295
Gonzales, K. K., and Smith, Y. (2015). Cholinergic interneurons in the dorsal and ventral striatum: anatomical and functional considerations in normal and diseased conditions. Ann. N. Y. Acad. Sci. 1349, 1–45. doi: 10.1111/nyas.12762
Gotti, C., Clementi, F., Fornari, A., Gaimarri, A., Guiducci, S., Manfredi, I., et al. (2009). Structural and functional diversity of native brain neuronal nicotinic receptors. Biochem. Pharmacol. 78, 703–711. doi: 10.1016/j.bcp.2009.05.024
Götz, T., Kraushaar, U., Geiger, J., Lübke, J., Berger, T., and Jonas, P. (1997). Functional properties of AMPA and NMDA receptors expressed in identified types of basal ganglia neurons. J. Neurosci. 17, 204–215. doi: 10.1523/JNEUROSCI.17-01-00204.1997
Govindaiah, G., Wang, Y., and Cox, C. L. (2010). Substance P selectively modulates GABAA receptor-mediated synaptic transmission in striatal cholinergic interneurons. Neuropharmacology 58, 413–422. doi: 10.1016/j.neuropharm.2009.09.011
Graybiel, A. M., Aosaki, T., Flaherty, A. W., and Kimura, M. (1994). The basal ganglia and adaptive motor control. Science 265, 1826–1831. doi: 10.1126/science.8091209
Groenewegen, H. J., and Berendse, H. W. (1994). The specificity of the ‘nonspecific’ midline and intralaminar thalamic nuclei. Trends Neurosci. 17, 52–57. doi: 10.1016/0166-2236(94)90074-4
Gross, R. A., Moisest, H. C., Uhlert, M. D., and Macdonald, R. L. (1990). Dynorphin A and cAMP-dependent protein kinase independently regulate neuronal calcium currents. Proc. Natl. Acad. Sci. USA 87, 7025–7029. doi: 10.1073/pnas.87.18.7025
Guevara Guzman, R., Kendrick, K. M., and Emson, P. C. (1993). Effect of substance P on acetylcholine and dopamine release in the rat striatum: a microdialysis study. Brain Res. 622, 147–154. doi: 10.1016/0006-8993(93)90813-3
Guo, Q., Wang, D., He, X., Feng, Q., Lin, R., Xu, F., et al. (2015). Whole-brain mapping of inputs to projection neurons and cholinergic interneurons in the dorsal striatum. PLoS One 10:e0123381. doi: 10.1371/journal.pone.0123381
Haber, S. N. (2011). Neuroanatomy of reward: A view from the ventral striatum. In: JA Gottfried, editor. Neurobiology of sensation and reward. Boca Raton (FL): CRC Press/Taylor & Francis. Available at: http://www.ncbi.nlm.nih.gov/pubmed/3972039 (Accessed November 10, 2024).
Hattori, T., Takada, M., Moriizumi, T., and Van Der Kooy, D. (1991). Single dopaminergic nigrostriatal neurons form two chemically distinct synaptic types: possible transmitter segregation within neurons. J. Comp. Neurol. 309, 391–401. doi: 10.1002/cne.903090308
Heath, E., Chieng, B., Christie, M. J., and Balleine, B. W. (2018). Substance P and dopamine interact to modulate the distribution of delta-opioid receptors on cholinergic interneurons in the striatum. Eur. J. Neurosci. 47, 1159–1173. doi: 10.1111/ejn.13750
Hegedüs, P., Sviatkó, K., Király, B., Martínez-Bellver, S., and Hangya, B. (2023). Cholinergic activity reflects reward expectations and predicts behavioral responses. iScience 26:26. doi: 10.1016/j.isci.2022.105814
Hersch, S., Gutekunst, C., Rees, H., Heilman, C., and Levey, A. (1994). Distribution of m1-m4 muscarinic receptor proteins in the rat striatum: light and electron microscopic immunocytochemistry using subtype- specific antibodies. J. Neurosci. 14, 3351–3363. doi: 10.1523/JNEUROSCI.14-05-03351.1994
Higley, M. J., Gittis, A. H., Oldenburg, I. A., Balthasar, N., Seal, R. P., Edwards, R. H., et al. (2011). Cholinergic interneurons mediate fast VGluT3-dependent glutamatergic transmission in the striatum. PLoS One 6:e19155. doi: 10.1371/journal.pone.0019155
Ho, T. N. T., Abraham, N., and Lewis, R. J. (2020). Structure-function of neuronal nicotinic acetylcholine receptor inhibitors derived from natural toxins. Front. Neurosci. 14:609005. doi: 10.3389/fnins.2020.609005
Hökfelt, T., Barde, S., Xu, Z. Q. D., Kuteeva, E., Rüegg, J., Le Maitre, E., et al. (2018). Neuropeptide and small transmitter coexistence: fundamental studies and relevance to mental illness. Front. Neural Circuits 12:106. doi: 10.3389/fncir.2018.00106
Holly, E. N., Boyson, C. O., Montagud-Romero, S., Stein, D. J., Gobrogge, K. L., DeBold, J. F., et al. (2016). Episodic social stress-escalated cocaine self-administration: role of phasic and tonic corticotropin releasing factor in the anterior and posterior ventral tegmental area. J. Neurosci. 36, 4093–4105. doi: 10.1523/JNEUROSCI.2232-15.2016
Howe, A. R., and Surmeier, D. J. (1995). Muscarinic receptors modulate N-, P-, and L-type Ca2+ currents in rat striatal neurons through parallel pathways. J. Neurosci. 15, 458–469. doi: 10.1523/JNEUROSCI.15-01-00458.1995
Howe, W. M., Young, D. A., Bekheet, G., and Kozak, R. (2016). Nicotinic receptor subtypes differentially modulate glutamate release in the dorsal medial striatum. Neurochem. Int. 100, 30–34. doi: 10.1016/j.neuint.2016.08.009
Hunnicutt, B. J., Jongbloets, B. C., Birdsong, W. T., Gertz, K. J., Zhong, H., and Mao, T. (2016). A comprehensive excitatory input map of the striatum reveals novel functional organization. eLife 5:e19103. doi: 10.7554/eLife.19103
Iino, Y., Sawada, T., Yamaguchi, K., Tajiri, M., Ishii, S., Kasai, H., et al. (2020). Dopamine D2 receptors in discrimination learning and spine enlargement. Nature 579, 555–560. doi: 10.1038/s41586-020-2115-1
Ikarashi, Y., Takahashi, A., Ishimaru, H., Arai, T., and Maruyama, Y. (1997). Suppression of cholinergic activity via the dopamine D2 receptor in the rat striatum. Neurochem. Int. 30, 191–197. doi: 10.1016/s0197-0186(96)00024-1
Ingebretson, A. E., Alonso-Caraballo, Y., Razidlo, J. A., and Lemos, J. C. (2024). Corticotropin releasing factor alters the functional diversity of accumbal cholinergic interneurons. J. Neurophysiol. 132, 403–417. doi: 10.1152/jn.00348.2023
Jabourian, M., Venance, L., Bourgoin, S., Ozon, S., Pérez, S., Godeheu, G., et al. (2005). Functional mu opioid receptors are expressed in cholinergic interneurons of the rat dorsal striatum: territorial specificity and diurnal variation. Eur. J. Neurosci. 21, 3301–3309. doi: 10.1111/j.1460-9568.2005.04154.x
Juhasz, G., Hullam, G., Eszlari, N., Gonda, X., Antal, P., Anderson, I. M., et al. (2014). Brain galanin system genes interact with life stresses in depression-related phenotypes. Proc. Natl. Acad. Sci. USA 111, E1666–E1673. doi: 10.1073/pnas.1403649111
Kawano, M., Kawasaki, A., Sakata-Haga, H., Fukui, Y., Kawano, H., Nogami, H., et al. (2006). Particular subpopulations of midbrain and hypothalamic dopamine neurons express vesicular glutamate transporter 2 in the rat brain. J. Comp. Neurol. 498, 581–592. doi: 10.1002/cne.21054
Kemp, J. M., and Powell, T. P. S. (1970). The cortico-striate projection in the monkey. Brain 93, 525–546. doi: 10.1093/brain/93.3.525
Khan, Z. U., Gutiérrez, A., Martı́n, R., Peñafiel, A., Rivera, A., and de la Calle, A. (2000). Dopamine D5 receptors of rat and human brain. Neuroscience 100, 689–699. doi: 10.1016/S0306-4522(00)00274-8
Kharkwal, G., Brami-Cherrier, K., Lizardi-Ortiz, J. E., Nelson, A. B., Ramos, M., Del Barrio, D., et al. (2016). Parkinsonism driven by antipsychotics originates from dopaminergic control of striatal cholinergic interneurons. Neuron 91, 67–78. doi: 10.1016/j.neuron.2016.06.014
Kim, T., Capps, R. A., Hamade, K. C., Barnett, W. H., Todorov, D. I., Latash, E. M., et al. (2019). The functional role of striatal cholinergic interneurons in reinforcement learning from computational perspective. Front. Neural Circuits 13:10. doi: 10.3389/fncir.2019.00010
Kim, H. J., Hwang, B., Reva, M., Lee, J., Lee, B. E., Lee, Y., et al. (2023). GABAergic-like dopamine synapses in the brain. Cell Rep. 42:113239. doi: 10.1016/j.celrep.2023.113239
Kimura, M., Rajkowski, J., and Evarts, E. (1984). Tonically discharging putamen neurons exhibit set-dependent responses. PNAS 81, 4998–5001. doi: 10.1073/pnas.81.15.4998
Klawonn, A. M., and Malenka, R. C. (2018). Nucleus Accumbens modulation in reward and aversion. Cold Spring Harb. Symp. Quant. Biol. 83, 119–129. doi: 10.1101/sqb.2018.83.037457
Kljakic, O., Janickova, H., Prado, V. F., and Prado, M. A. M. (2017). Cholinergic/glutamatergic co-transmission in striatal cholinergic interneurons: new mechanisms regulating striatal computation. J. Neurochem. 142, 90–102. doi: 10.1111/jnc.14003
Klug, J. R., Engelhardt, M. D., Cadman, C. N., Li, H., Smith, J. B., Ayala, S., et al. (2018). Differential inputs to striatal cholinergic and parvalbumin interneurons imply functional distinctions. eLife 7:e35657. doi: 10.7554/eLife.35657
Kocaturk, S., Guven, E. B., Shah, F., Tepper, J. M., and Assous, M. (2022). Cholinergic control of striatal GABAergic microcircuits. Cell Rep. 41:111531. doi: 10.1016/j.celrep.2022.111531
Koranda, J. L., Cone, J. J., McGehee, D. S., Roitman, M. F., Beeler, J. A., and Zhuang, X. (2014). Nicotinic receptors regulate the dynamic range of dopamine release in vivo. J. Neurophysiol. 111, 103–111. doi: 10.1152/jn.00269.2013
Kosillo, P., Zhang, Y. F., Threlfell, S., and Cragg, S. J. (2016). Cortical control of striatal dopamine transmission via striatal cholinergic interneurons. Cereb. Cortex 26, 4160–4169. doi: 10.1093/cercor/bhw252
Kövesdi, E., Udvarácz, I., Kecskés, A., Szőcs, S., Farkas, S., Faludi, P., et al. (2023). 17β-estradiol does not have a direct effect on the function of striatal cholinergic interneurons in adult mice in vitro. Front. Endocrinol. (Lausanne) 13:993552. doi: 10.3389/fendo.2022.993552
Krok, A. C., Maltese, M., Mistry, P., Miao, X., Li, Y., and Tritsch, N. X. (2023). Intrinsic dopamine and acetylcholine dynamics in the striatum of mice. Nature 621, 543–549. doi: 10.1038/s41586-023-05995-9
Kubota, Y., Inagaki, S., Shimada, S., Kito, S., Eckenstein, F., and Tohyama, M. (1987). Neostriatal cholinergic neurons receive direct synaptic inputs from dopaminergic axons. Brain Res. 413, 179–184. doi: 10.1016/0006-8993(87)90167-3
Kulak, J. M., Nguyen, T. A., Olivera, B. M., and McIntosh, J. M. (1997). α-Conotoxin MII blocks nicotine-stimulated dopamine release in rat striatal Synaptosomes. J. Neurosci. 17, 5263–5270. doi: 10.1523/JNEUROSCI.17-14-05263.1997
Kuramoto, E., Fujiyama, F., Unzai, T., Nakamura, K., Hioki, H., Furuta, T., et al. (2007). Metabotropic glutamate receptor 4-immunopositive terminals of medium-sized spiny neurons selectively form synapses with cholinergic interneurons in the rat neostriatum. J. Comp. Neurol. 500, 908–922. doi: 10.1002/cne.21216
Kuteeva, E., Hökfelt, T., Wardi, T., and Ögren, S. O. (2008). Galanin, galanin receptor subtypes and depression-like behaviour. Cell. Mol. Life Sci. 65, 1854–1863. doi: 10.1007/s00018-008-8160-9
Lachowicz, J. E., Duffy, R. A., Ruperto, V., Kozlowski, J., Zhou, G., Clader, J., et al. (2001). Facilitation of acetylcholine release and improvement in cognition by a selective M2 muscarinic antagonist, SCH 72788. Life Sci. 68, 2585–2592. doi: 10.1016/S0024-3205(01)01056-6
Landwehrmeyer, G., Standaert, D., Testa, C., Penney, J., and Young, A. (1995). NMDA receptor subunit mRNA expression by projection neurons and interneurons in rat striatum. J. Neurosci. 15, 5297–5307. doi: 10.1523/JNEUROSCI.15-07-05297.1995
Lapper, S. R., and Bolam, J. P. (1992). Input from the frontal cortex and the parafascicular nucleus to cholinergic interneurons in the dorsal striatum of the rat. Neuroscience 51, 533–545. doi: 10.1016/0306-4522(92)90293-B
Laverne, G., Pesce, J., Reynders, A., Combrisson, E., Gascon, E., Melon, C., et al. (2022). Cholinergic interneuron inhibition potentiates corticostriatal transmission in direct medium spiny neurons and rescues motor learning in parkinsonism. Cell Rep. 40:111034. doi: 10.1016/j.celrep.2022.111034
Lavoie, B., and Parent, A. (1990). Immunohistochemical study of the serotoninergic innervation of the basal ganglia in the squirrel monkey. J. Comp. Neurol. 299, 1–16. doi: 10.1002/cne.902990102
Le, B., Cheng, X., and Qu, S. (2020). Cooperative effects of galanin and leptin on alleviation of insulin resistance in adipose tissue of diabetic rats. J. Cell. Mol. Med. 24, 6773–6780. doi: 10.1111/jcmm.15328
Le Moine, C., Kieffer, B., Gaveriaux-Ruff, C., Befort, K., and Block, B. (1994). Delta-opioid receptor gene expression in the mouse forebrain: localization in cholinergic neurons of the striatum. Neuroscience 62, 635–640. doi: 10.1016/0306-4522(94)90464-2
Le Moine, C., Normand, E., Guitteny, A. F., Fouque, B., Teoule, R., and Bloch, B. (1990). Dopamine receptor gene expression by enkephalin neurons in rat forebrain. PNAS 87, 230–234. doi: 10.1073/pnas.87.1.230
Lemos, J. C., Shin, J. H., and Alvarez, V. A. (2019). Striatal cholinergic interneurons are a novel target of corticotropin releasing factor. J. Neurosci. 39, 5647–5661. doi: 10.1523/JNEUROSCI.0479-19.2019
Lemos, J. C., Wanat, M. J., Smith, J. S., Reyes, B. A. S., Hollon, N. G., Van Bockstaele, E. J., et al. (2012). Severe stress switches CRF action in the nucleus accumbens from appetitive to aversive. Nature 490, 402–406. doi: 10.1038/nature11436
Lim, S. A. O., Kang, U. J., and McGehee, D. S. (2014). Striatal cholinergic interneuron regulation and circuit effects. Front. Synaptic Neurosci. 6:22. doi: 10.3389/fnsyn.2014.00022
Liu, C., and Kaeser, P. S. (2019). Mechanisms and regulation of dopamine release. Curr. Opin. Neurobiol. 57, 46–53. doi: 10.1016/j.conb.2019.01.001
Luo, R., Uematsu, A., Weitemier, A., Aquili, L., Koivumaa, J., McHugh, T. J., et al. (2018). A dopaminergic switch for fear to safety transitions. Nat. Commun. 9:2483. doi: 10.1038/s41467-018-04784-7
Lv, X., Dickerson, J. W., Rook, J. M., Lindsley, C. W., Conn, P. J., and Xiang, Z. (2017). M1 muscarinic activation induces long-lasting increase in intrinsic excitability of striatal projection neurons. Neuropharmacology 118, 209–222. doi: 10.1016/j.neuropharm.2017.03.017
Mamaligas, A. A., Barcomb, K., and Ford, C. P. (2019). Cholinergic transmission at muscarinic synapses in the striatum is driven equally by cortical and thalamic inputs. Cell Rep. 28, 1003–1014.e3. doi: 10.1016/j.celrep.2019.06.077
Mancini, M., Patel, J. C., Affinati, A. H., Witkovsky, P., and Rice, M. E. (2022). Leptin promotes striatal dopamine release via cholinergic interneurons and regionally distinct signaling pathways. J. Neurosci. 42, 6668–6679. doi: 10.1523/JNEUROSCI.0238-22.2022
Mandelbaum, G., Taranda, J., Haynes, T. M., Hochbaum, D. R., Huang, K. W., Hyun, M., et al. (2019). Distinct cortical-thalamic-striatal circuits through the Parafascicular nucleus. Neuron 102, 636–652.e7. doi: 10.1016/j.neuron.2019.02.035
Martone, M. E., Armstrong, D. M., Young, S. J., and Groves, P. M. (1992). Ultrastructural examination of enkephalin and substance P input to cholinergic neurons within the rat neostriatum. Brain Res. 594, 253–262. doi: 10.1016/0006-8993(92)91132-X
Matamales, M., Götz, J., and Bertran-Gonzalez, J. (2016). Quantitative imaging of cholinergic interneurons reveals a distinctive spatial organization and a functional gradient across the mouse striatum. PLoS One 11:e0157682. doi: 10.1371/journal.pone.0157682
Matityahu, L., Gilin, N., Sarpong, G. A., Atamna, Y., Tiroshi, L., Tritsch, N. X., et al. (2023). Acetylcholine waves and dopamine release in the striatum. Nat. Commun. 14:6852. doi: 10.1038/s41467-023-42311-5
Matsumoto, N., Minamimoto, T., Graybiel, A. M., and Kimura, M. (2001). Neurons in the thalamic CM-pf complex supply striatal neurons with information about behaviorally significant sensory events. J. Neurophysiol. 85, 960–976. doi: 10.1152/jn.2001.85.2.960
Maurice, N., Mercer, J., Chan, C. S., Hernandez-Lopez, S., Held, J., Tkatch, T., et al. (2004). D2 dopamine receptor-mediated modulation of voltage-dependent Na+ channels reduces autonomous activity in striatal cholinergic interneurons. J. Neurosci. 24, 10289–10301. doi: 10.1523/JNEUROSCI.2155-04.2004
McGeer, P. L., McGeer, E. G., Fibiger, H. C., and Wickson, V. (1971). Neostriatal choline acetylase and cholinesterase following selective brain lesions. Brain Res. 35, 308–314. doi: 10.1016/0006-8993(71)90625-1
McGeorge, A. J., and Faull, R. L. M. (1989). The organization of the projection from the cerebral cortex to the striatum in the rat. Neuroscience 29, 503–537. doi: 10.1016/0306-4522(89)90128-0
McGuirt, A., Pigulevskiy, I., and Sulzer, D. (2022). Developmental regulation of thalamus-driven pauses in striatal cholinergic interneurons. iScience 25:25. doi: 10.1016/j.isci.2022.105332
Meredith, G. E., Blank, B., and Groenewegen, H. J. (1989). The distribution and compartmental organization of the cholinergic neurons in nucleus accumbens of the rat. Neuroscience 31, 327–345. doi: 10.1016/0306-4522(89)90377-1
Mingote, S., Amsellem, A., Kempf, A., Rayport, S., and Chuhma, N. (2019). Dopamine-glutamate neuron projections to the nucleus accumbens medial shell and behavioral switching. Neurochem. Int. 129:104482. doi: 10.1016/j.neuint.2019.104482
Miranda-Barrientos, J., Chambers, I., Mongia, S., Liu, B., Wang, H. L., Mateo-Semidey, G. E., et al. (2021). Ventral tegmental area GABA, glutamate, and glutamate-GABA neurons are heterogeneous in their electrophysiological and pharmacological properties. Eur. J. Neurosci. 54, 4061–4084. doi: 10.1111/ejn.15156
Morris, G., Arkadir, D., Nevet, A., Vaadia, E., and Bergman, H. (2004). Coincident but distinct messages of midbrain dopamine and striatal tonically active neurons. Neuron 43, 133–143. doi: 10.1016/j.neuron.2004.06.012
Morton, G. J., Cummings, D. E., Baskin, D. G., Barsh, G. S., and Schwartz, M. W. (2006). Central nervous system control of food intake and body weight. Nature 443, 289–295. doi: 10.1038/nature05026
Mulder, A. H., Wardeh, G., Hogenboom, F., and Frankhuyzen, A. L. (1984). κ and δ-opioid receptor agonists differentially inhibit striatal dopamine and acetylcholine release. Nature 308, 278–280. doi: 10.1038/308278a0
Myers, M. G., and Olson, D. P. (2014). SnapShot: neural pathways that control feeding. Cell Metab. 19, 732–732.e1. doi: 10.1016/j.cmet.2014.03.015
Nakamura, T. Y., Coetzee, W. A., De Miera, E. V.-S., Artman, M., and Rudy, B. (1997). Modulation of Kv4 channels, key components of rat ventricular transient outward K+ current, by PKC. Am. J. Phys. 273, H1775–H1786. doi: 10.1152/ajpheart.1997.273.4.H1775
Nanda, B., Galvan, A., Smith, Y., and Wichmann, T. (2009). Effects of stimulation of the centromedian nucleus of the thalamus on the activity of striatal cells in awake rhesus monkeys. Eur. J. Neurosci. 29, 588–598. doi: 10.1111/j.1460-9568.2008.06598.x
Nelson, A. B., Hammack, N., Yang, C. F., Shah, N. M., Seal, R. P., and Kreitzer, A. C. (2014). Striatal cholinergic interneurons drive GABA release from dopamine terminals. Neuron 82, 63–70. doi: 10.1016/j.neuron.2014.01.023
Nieh, E. H., Kim, S. Y., Namburi, P., and Tye, K. M. (2013). Optogenetic dissection of neural circuits underlying emotional valence and motivated behaviors. Brain Res. 1511, 73–92. doi: 10.1016/j.brainres.2012.11.001
Nougaret, S., and Ravel, S. (2015). Modulation of tonically active neurons of the monkey striatum by events carrying different force and reward information. J. Neurosci. 35, 15214–15226. doi: 10.1523/JNEUROSCI.0039-15.2015
O’Donnell, P. (1999). Ensemble coding in the nucleus accumbens. Psychobiology 27, 187–197. doi: 10.3758/BF03332113
Ogren, S. O., and Pramanik, A. (1991). Galanin stimulates acetylcholine release in the rat striatum. Neurosci. Lett. 128, 253–256. doi: 10.1016/0304-3940(91)90273-v
Oldenburg, I. A., and Ding, J. B. (2011). Cholinergic modulation of synaptic integration and dendritic excitability in the striatum. Curr. Opin. Neurobiol. 21, 425–432. doi: 10.1016/j.conb.2011.04.004
Olson, K. L., Ingebretson, A. E., Vogiatzoglou, E., Mermelstein, P. G., and Lemos, J. C. (2024). Cholinergic interneurons in the nucleus accumbens are a site of cellular convergence for corticotropin-releasing factor and estrogen regulation in male and female mice. Eur. J. Neurosci. 60, 4937–4953. doi: 10.1111/ejn.16477
Oswald, M. J., Oorschot, D. E., Schulz, J. M., Lipski, J., and Reynolds, J. N. J. (2009). IH current generates the afterhyperpolarisation following activation of subthreshold cortical synaptic inputs to striatal cholinergic interneurons. J. Physiol. 587, 5879–5897. doi: 10.1113/jphysiol.2009.177600
Pakhotin, P., and Bracci, E. (2007). Cholinergic interneurons control the excitatory input to the striatum. J. Neurosci. 27, 391–400. doi: 10.1523/JNEUROSCI.3709-06.2007
Paladini, C. A., and Roeper, J. (2014). Generating bursts (and pauses) in the dopaminemidbrain neurons. Neuroscience 282, 109–121. doi: 10.1016/j.neuroscience.2014.07.032
Pancani, T., Bolarinwa, C., Smith, Y., Lindsley, C. W., Conn, P. J., and Xiang, Z. (2014). M4 mAChR-mediated modulation of glutamatergic transmission at corticostriatal synapses. ACS Chem. Neurosci. 5, 318–324. doi: 10.1021/cn500003z
Patel, J. C., Carr, K. D., and Rice, M. E. (2023). Actions and consequences of insulin in the striatum. Biomol. Ther. 13:518. doi: 10.3390/biom13030518
Pennartz, C., Groenewegen, H., and Lopesdasilva, F. (1994). The nucleus accumbens as a complex of functionally distinct neuronal ensembles: An integration of behavioural, electrophysiological and anatomical data. Prog. Neurobiol. 42, 719–761. doi: 10.1016/0301-0082(94)90025-6
Pennefather, J. N., Lecci, A., Candenas, M. L., Patak, E., Pinto, F. M., and Maggi, C. A. (2004). Tachykinins and tachykinin receptors: A growing family. Life Sci. 74, 1445–1463. doi: 10.1016/j.lfs.2003.09.039
Pérez, S., Tierney, A., Deniau, J., and Kemel, M. (2007). Tachykinin regulation of cholinergic transmission in the limbic/prefrontal territory of the rat dorsal striatum: implication of new neurokinine 1-sensitive receptor binding site and interaction with enkephalin/mu opioid receptor transmission. J. Neurochem. 103, 2153–2163. doi: 10.1111/j.1471-4159.2007.04944.x
Perez-Burgos, A., Perez-Rosello, T., Salgado, H., Flores-Barrera, E., Prieto, G. A., Figueroa, A., et al. (2008). Muscarinic M(1) modulation of N and L types of calcium channels is mediated by protein kinase C in neostriatal neurons. Neuroscience 155, 1079–1097. doi: 10.1016/j.neuroscience.2008.06.047
Persohn, E., Malherbe, P., and Richards, J. G. (1992). Comparative molecular neuroanatomy of cloned GABAA receptor subunits in the rat CNS. J. Comp. Neurol. 326, 193–216. doi: 10.1002/cne.903260204
Picciotto, M. R. (2008). Galanin and addiction. Cell. Mol. Life Sci. 65, 1872–1879. doi: 10.1007/s00018-008-8151-x
Pickel, V. M., Douglas, J., Chan, J., Gamp, P. D., and Bunnett, N. W. (2000). Neurokinin 1 receptor distribution in cholinergic neurons and targets of substance P terminals in the rat nucleus accumbens. J. Comp. Neurol. 423, 500–511. doi: 10.1002/1096-9861(20000731)423:3<500::AID-CNE12>3.0.CO;2-9
Pisani, A., Bonsi, P., Catania, M. V., Giuffrida, R., Morari, M., Marti, M., et al. (2002). Metabotropic glutamate 2 receptors modulate synaptic inputs and calcium signals in striatal cholinergic interneurons. J. Neurosci. 22, 6176–6185. doi: 10.1523/JNEUROSCI.22-14-06176.2002
Ponterio, G., Tassone, A., Sciamanna, G., Riahi, E., Vanni, V., Bonsi, P., et al. (2013). Powerful inhibitory action of mu opioid receptors (MOR) on cholinergic interneuron excitability in the dorsal striatum. Neuropharmacology 75, 78–85. doi: 10.1016/j.neuropharm.2013.07.006
Poppi, L. A., Ho-Nguyen, K. T., Shi, A., Daut, C. T., and Tischfield, M. A. (2021). Recurrent implication of striatal cholinergic interneurons in a range of neurodevelopmental, neurodegenerative, and neuropsychiatric disorders. Cells 10:907. doi: 10.3390/cells10040907
Poulin, J. F., Caronia, G., Hofer, C., Cui, Q., Helm, B., Ramakrishnan, C., et al. (2018). Mapping projections of molecularly defined dopamine neuron subtypes using intersectional genetic approaches. Nat. Neurosci. 21, 1260–1271. doi: 10.1038/s41593-018-0203-4
Pramanik, A., and Ogren, S. O. (1993). Galanin stimulates striatal acetylcholine release via a mechanism unrelated to cholinergic receptor stimulation. Regul. Pept. 45, 353–362. doi: 10.1016/0167-0115(93)90361-b
Preston, Z., Lee, K., Widdowson, L., Richardson, P. J., and Pinnock, R. D. (2000). Tachykinins increase [3H] acetylcholine release in mouse striatum through multiple receptor subtypes. Neuroscience 95, 367–376. doi: 10.1016/s0306-4522(99)00440-6
Rada, P. V., Mark, G. P., and Hoebel, B. G. (1993). In vivo modulation of acetylcholine in the nucleus accumbens of freely moving rats: I. Inhibition by serotonin. Brain Res. 619, 98–104. doi: 10.1016/0006-8993(93)91600-W
Rana, T., Behl, T., Sehgal, A., Singh, S., Sharma, N., Abdeen, A., et al. (2022). Exploring the role of neuropeptides in depression and anxiety. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 114:110478. doi: 10.1016/j.pnpbp.2021.110478
Ravel, S., Legallet, E., and Apicella, P. (1999). Tonically active neurons in the monkey striatum do not preferentially respond to appetitive stimuli. Exp. Brain Res. 128, 531–534. doi: 10.1007/s002210050876
Ravel, S., Legallet, E., and Apicella, P. (2003). Responses of tonically active neurons in the monkey striatum discriminate between motivationally opposing stimuli. J. Neurosci. 23, 8489–8497. doi: 10.1523/JNEUROSCI.23-24-08489.2003
Reynolds, J. N. J., Avvisati, R., Dodson, P. D., Fisher, S. D., Oswald, M. J., Wickens, J. R., et al. (2022). Coincidence of cholinergic pauses, dopaminergic activation and depolarisation of spiny projection neurons drives synaptic plasticity in the striatum. Nat. Commun. 13:1296. doi: 10.1038/s41467-022-28950-0
Richardson, P. J., Dixon, A. K., Lee, K., Bell, M. I., Cox, P. J., Williams, R., et al. (2000). Correlating physiology with gene expression in striatal cholinergic neurones. J. Neurochem. 74, 839–846. doi: 10.1046/j.1471-4159.2000.740839.x
Rivera, A., Alberti, I., Martín, A. B., Narváez, J. A., De La Calle, A., and Moratalla, R. (2002). Molecular phenotype of rat striatal neurons expressing the dopamine D 5 receptor subtype. Eur. J. Neurosci. 16, 2049–2058. doi: 10.1046/j.1460-9568.2002.02280.x
Rolls, E. T., Thorpe, S. J., Boytim, M., Szabo, I., and Perrett, D. I. (1984). Responses of striatal neurons in the behaving monkey. 3. Effects of iontophoretically applied dopamine on normal responsiveness. Neuroscience 12, 1201–1212. doi: 10.1016/0306-4522(84)90014-9
Sadikot, A. F., Parent, A., and François, C. (1992). Efferent connections of the centromedian and parafascicular thalamic nuclei in the squirrel monkey: A PHA-L study of subcortical projections. J. Comp. Neurol. 315, 137–159. doi: 10.1002/cne.903150203
Sanchez, G., Rodriguez, M. J., Pomata, P., Rela, L., and Murer, M. G. (2011). Reduction of an afterhyperpolarization current increases excitability in striatal cholinergic interneurons in rat parkinsonism. J. Neurosci. 31, 6553–6564. doi: 10.1523/JNEUROSCI.6345-10.2011
Sandor, N. T., Zelles, T., Kiss, J., Sershen, H., Torocsik, A., Lajtha, A., et al. (1991). Effect of nicotine on dopaminergic-cholinergic interaction in the striatum. Brain Res. 567, 313–316. doi: 10.1016/0006-8993(91)90810-i
Sato, A., Sasaoka, T., Nishijo, T., and Momiyama, T. (2014). GABAergic synaptic transmission onto striatal cholinergic interneurons in dopamine D2 receptor knock-out mice. Neuroscience 263, 138–147. doi: 10.1016/j.neuroscience.2014.01.010
Scatton, B., and Lehmann, J. (1982). N-methyl-D-aspartate-type receptors mediate striatal 3H-acetylcholine release evoked by excitatory amino acids. Nature 297, 422–424. doi: 10.1038/297422a0
Scherrer, G., Tryoen-Tóth, P., Filliol, D., Matifas, A., Laustriat, D., Cao, Y. Q., et al. (2006). Mice Knockin mice expressing fluorescent δ-opioid receptors uncover G protein-coupled receptor dynamics in vivo. PNAS 103, 9691–9696. doi: 10.1073/pnas.0603359103
Schulz, J. M., and Reynolds, J. N. J. (2013). Pause and rebound: sensory control of cholinergic signaling in the striatum. Trends Neurosci. 36, 41–50. doi: 10.1016/j.tins.2012.09.006
Sharott, A., Doig, N. M., Mallet, N., and Magill, P. J. (2012). Relationships between the firing of identified striatal interneurons and spontaneous and driven cortical activities in vivo. J. Neurosci. 32, 13221–13236. doi: 10.1523/JNEUROSCI.2440-12.2012
Shen, W., Plotkin, J. L., Francardo, V., Ko, W. K., Xie, Z., Li, Q., et al. (2015). M4 muscarinic receptor signaling ameliorates striatal plasticity deficits in models of L-DOPA-induced dyskinesia. Neuron 88, 762–773. doi: 10.1016/j.neuron.2015.10.039
Shen, W., Tian, X., Day, M., Ulrich, S., Tkatch, T., Nathanson, N. M., et al. (2007). Cholinergic modulation of Kir2 channels selectively elevates dendritic excitability in striatopallidal neurons. Nat. Neurosci. 10, 1458–1466. doi: 10.1038/nn1972
Shroff, S. N., Lowet, E., Sridhar, S., Gritton, H. J., Abumuaileq, M., Tseng, H. A., et al. (2023). Striatal cholinergic interneuron membrane voltage tracks locomotor rhythms in mice. Nat. Commun. 14:3802. doi: 10.1038/s41467-023-39497-z
Sidibé, M., and Smith, Y. (1999). Thalamic inputs to striatal interneurons in monkeys: synaptic organization and co-localization of calcium binding proteins. Neuroscience 89, 1189–1208. doi: 10.1016/S0306-4522(98)00367-4
Smith, Y., Galvan, A., Ellender, T. J., Doig, N., Villalba, R. M., Huerta-Ocampo, I., et al. (2014). The thalamostriatal system in normal and diseased states. Front. Syst. Neurosci. 8:5. doi: 10.3389/fnsys.2014.00005
Standaert, D. G., Friberg, I. K., Landwehrmeyer, G. B., Young, A. B., and Penney, J. B. (1999). Expression of NMDA glutamate receptor subunit mRNAs in neurochemically identified projection and interneurons in the striatum of the rat. Brain Res. Mol. Brain Res. 64, 11–23. doi: 10.1016/S0169-328X(98)00293-9
Steinbusch, H. W. M. (1981). Distribution of serotonin-immunoreactivity in the central nervous system of the rat—cell bodies and terminals. Neuroscience 6, 557–618. doi: 10.1016/0306-4522(81)90146-9
Steinkellner, T., Zell, V., Farino, Z. J., Sonders, M. S., Villeneuve, M., Freyberg, R. J., et al. (2018). Role for VGLUT2 in selective vulnerability of midbrain dopamine neurons. J. Clin. Invest. 128, 774–788. doi: 10.1172/JCI95795
Stensrud, M. J., Puchades, M., and Gundersen, V. (2014). GABA is localized in dopaminergic synaptic vesicles in the rodent striatum. Brain Struct. Funct. 219, 1901–1912. doi: 10.1007/s00429-013-0609-4
Stouffer, M. A., Woods, C. A., Patel, J. C., Lee, C. R., Witkovsky, P., Bao, L., et al. (2015). Insulin enhances striatal dopamine release by activating cholinergic interneurons and thereby signals reward. Nat. Commun. 6:8543. doi: 10.1038/ncomms9543
Straub, C., Tritsch, N. X., Hagan, N. A., Gu, C., and Sabatini, B. L. (2014). Multiphasic modulation of cholinergic interneurons by nigrostriatal afferents. J. Neurosci. 34, 8557–8569. doi: 10.1523/JNEUROSCI.0589-14.2014
Sullivan, M. A., Chen, H., and Morikawa, H. (2008). Recurrent inhibitory network among striatal cholinergic interneurons. J. Neurosci. 28, 8682–8690. doi: 10.1523/JNEUROSCI.2411-08.2008
Sulzer, D., Cragg, S. J., and Rice, M. E. (2016). Striatal dopamine neurotransmission: regulation of release and uptake. Basal Ganglia 6, 123–148. doi: 10.1016/j.baga.2016.02.001
Svingos, A. L., Clarke, C. L., and Pickel, V. M. (1998). Cellular sites for activation of δ-opioid receptors in the rat nucleus accumbens shell: relationship with Met5-Enkephalin. J. Neurosci. 18, 1923–1933. doi: 10.1523/JNEUROSCI.18-05-01923.1998
Svingos, A. L., Colago, E. E. O., and Pickel, V. M. (2001). Vesicular acetylcholine transporter in the rat nucleus accumbens shell: subcellular distribution and association with μ-opioid receptors. Synapse 40, 184–192. doi: 10.1002/syn.1041
Swanson, L. W. (1982). The projections of the ventral tegmental area and adjacent regions: A combined fluorescent retrograde tracer and immunofluorescence study in the rat. Brain Res. Bull. 9, 321–353. doi: 10.1016/0361-9230(82)90145-9
Szydlowski, S. N., Pollak, D. I, Planert, H., Carlén, M., Meletis, K., and Silberberg, G. (2013). Target selectivity of feedforward inhibition by striatal fast-spiking interneurons. J. Neurosci. 33, 1678–1683. doi: 10.1523/JNEUROSCI.3572-12.2013
Tecuapetla, F., Patel, J. C., Xenias, H., English, D., Tadros, I., Shah, F., et al. (2010). Glutamatergic signaling by mesolimbic dopamine neurons in the nucleus accumbens. J. Neurosci. 30, 7105–7110. doi: 10.1523/JNEUROSCI.0265-10.2010
Tepper, J. M., and Bolam, J. P. (2004). Functional diversity and specificity of neostriatal interneurons. Curr. Opin. Neurobiol. 14, 685–692. doi: 10.1016/j.conb.2004.10.003
Tepper, J. M., Koos, T., and Wilson, C. J. (2004). GABAergic microcircuits in the neostriatum. Trends Neurosci. 27, 662–669. doi: 10.1016/j.tins.2004.08.007
Thomas, T. M., Smith, Y., Levey, A. I., and Hersch, S. M. (2000). Cortical inputs to m2-immunoreactive striatal interneurons in rat and monkey. Synapse 37, 252–261. doi: 10.1002/1098-2396(20000915)37:4<252::AID-SYN2>3.0.CO;2-A
Threlfell, S., Lalic, T., Platt, N. J., Jennings, K. A., Deisseroth, K., and Cragg, S. J. (2012). Striatal dopamine release is triggered by synchronized activity in cholinergic interneurons. Neuron 75, 58–64. doi: 10.1016/j.neuron.2012.04.038
Tritsch, N., Ding, J., and Sabatini, B. (2012). Dopaminergic neurons inhibit striatal output through non-canonical release of GABA. Nature 490, 262–266. doi: 10.1038/nature11466
Tritsch, N. X., and Sabatini, B. L. (2012). Dopaminergic modulation of synaptic transmission in cortex and striatum. Neuron 76, 33–50. doi: 10.1016/j.neuron.2012.09.023
Tubert, C., M Paz, R., Stahl, A. M., Rela, L., and Murer, M. G. (2024). 2024Striatal cholinergic interneuron pause response requires Kv1 channels, is absent in dyskinetic mice, and is restored by dopamine D5 receptor inverse agonism. eLife 13:RP102184. doi: 10.7554/eLife.102184.1
Uchigashima, M., Ohtsuka, T., Kobayashi, K., and Watanabe, M. (2016). Dopamine synapse is a neuroligin-2-mediated contact between dopaminergic presynaptic and GABAergic postsynaptic structures. Proc. Natl. Acad. Sci. USA 113, 4206–4211. doi: 10.1073/pnas.1514074113
Van Der Werf, Y. D., Witter, M. P., and Groenewegen, H. J. (2002). The intralaminar and midline nuclei of the thalamus. Anatomical and functional evidence for participation in processes of arousal and awareness. Brain Res. Brain Res. Rev. 39, 107–140. doi: 10.1016/s0165-0173(02)00181-9
Van Pett, K., Viau, V., Bittencourt, J. C., Chan, R. K. W., Li, H.-Y., Arias, C., et al. (2000). Distribution of mRNAs encoding CRF receptors in brain and pituitary of rat and mouse. J. Comp. Neurol. 428, 191–212. doi: 10.1002/1096-9861(20001211)428:2<191::AID-CNE1>3.0.CO;2-U
Vanderah, T., and Sandweiss, A. (2015). The pharmacology of neurokinin receptors in addiction: prospects for therapy. Subst. Abus. Rehabil. 6, 93–102. doi: 10.2147/sar.s70350
Virk, M. S., Sagi, Y., Medrihan, L., Leung, J., Kaplitt, M. G., and Greengard, P. (2016). Opposing roles for serotonin in cholinergic neurons of the ventral and dorsal striatum. Proc. Natl. Acad. Sci. USA 113, 734–739. doi: 10.1073/pnas.1524183113
Voorn, P., Vanderschuren, L. J. M. J., Groenewegen, H. J., Robbins, T. W., and Pennartz, C. M. A. (2004). Putting a spin on the dorsal-ventral divide of the striatum. Trends Neurosci. 27, 468–474. doi: 10.1016/j.tins.2004.06.006
Wada, N., Hirako, S., Takenoya, F., Kageyama, H., Okabe, M., and Shioda, S. (2014). Leptin and its receptors. J. Chem. Neuroanat. 61-62, 191–199. doi: 10.1016/j.jchemneu.2014.09.002
Wang, W., Darvas, M., Storey, G. P., Bamford, I. J., Gibbs, J. T., Palmiter, R. D., et al. (2013). Acetylcholine encodes long-lasting presynaptic plasticity at glutamatergic synapses in the dorsal striatum after repeated amphetamine exposure. J. Neurosci. 33, 10405–10426. doi: 10.1523/JNEUROSCI.0014-13.2013
Wang, S., and Gustafson, E. L. (1998). Galanin receptor subtypes. Drug News Perspect. 11, 458–468. Available at: http://www.ncbi.nlm.nih.gov/pubmed/15616674
Wang, Z., Kai, L., Day, M., Ronesi, J., Yin, H. H., Ding, J., et al. (2006). Dopaminergic control of corticostriatal long-term synaptic depression in medium spiny neurons is mediated by cholinergic interneurons. Neuron 50, 443–452. doi: 10.1016/j.neuron.2006.04.010
Warner-Schmidt, J. L., Schmidt, E. F., Marshall, J. J., Rubin, A. J., Arango-Lievano, M., Kaplitt, M. G., et al. (2012). Cholinergic interneurons in the nucleus accumbens regulate depression-like behavior. Proc. Natl. Acad. Sci. USA 109, 11360–11365. doi: 10.1073/pnas.1209293109
Watanabe, K., and Kimura, M. (1998). Dopamine receptor–mediated mechanisms involved in the expression of learned activity of primate striatal neurons. J. Neurophysiol. 79, 2568–2580. doi: 10.1152/jn.1998.79.5.2568
Weiner, D. M., Levey, A. I., and Brann, M. R. (1990). Expression of muscarinic acetylcholine and dopamine receptor mRNAs in rat basal ganglia. Proc. Natl. Acad. Sci. USA 87, 7050–7054. doi: 10.1073/pnas.87.18.7050
Weiss, A. P., Goff, D., Schacter, D. L., Ditman, T., Freudenreich, O., Henderson, D., et al. (2006). Fronto-hippocampal function during temporal context monitoring in schizophrenia. Biol. Psychiatry 60, 1268–1277. doi: 10.1016/j.biopsych.2006.06.025
Williams, R. G., Li, K. H., and Phillips, P. E. M. (2022). The influence of stress on decision-making: effects of CRF and dopamine antagonism in the nucleus accumbens. Front. Psych. 12:814218. doi: 10.3389/fpsyt.2021.814218
Wilson, C. J., Chang, H. T., and Kitai, S. T. (1990). Firing patterns and synaptic potentials of identified giant aspiny interneurons in the rat neostriatum. J. Neurosci. 10, 508–519. doi: 10.1523/JNEUROSCI.10-02-00508.1990
Wonnacott, S., Kaiser, S., Mogg, A., Soliakov, L., and Jones, I. W. (2000). Presynaptic nicotinic receptors modulating dopamine release in the rat striatum. Eur. J. Pharmacol. 393, 51–58. doi: 10.1016/s0014-2999(00)00005-4
Wu, Q., Clark, M. S., and Palmiter, R. D. (2012). Deciphering a neuronal circuit that mediates appetite. Nature 483, 594–597. doi: 10.1038/nature10899
Xu, L., Nan, J., and Lan, Y. (2020). The nucleus Accumbens: A common target in the comorbidity of depression and addiction. Front. Neural Circuits 14:37. doi: 10.3389/fncir.2020.00037
Yamanaka, K., Hori, Y., Minamimoto, T., Yamada, H., Matsumoto, N., Enomoto, K., et al. (2018). Roles of centromedian parafascicular nuclei of thalamus and cholinergic interneurons in the dorsal striatum in associative learning of environmental events. J. Neural Transm. 125, 501–513. doi: 10.1007/s00702-017-1713-z
Yan, Z., Song, W.-J., and Surmeier, D. J. (1997). D 2 dopamine receptors reduce N-type ca 2+ currents in rat neostriatal cholinergic interneurons through a membrane-delimited, protein-kinase-C-insensitive pathway. J. Neurophysiol. 77, 1003–1015. doi: 10.1152/jn.1997.77.2.1003
Yan, Z., and Surmeier, D. J. (1996). Muscarinic (m2/m4) receptors reduce N- and P-type Ca2+ currents in rat neostriatal cholinergic interneurons through a fast, membrane- delimited, G-protein pathway. J. Neurosci. 16, 2592–2604. doi: 10.1523/JNEUROSCI.16-08-02592.1996
Yan, Z., and Surmeier, D. J. (1997). D5 dopamine receptors enhance Zn2+−sensitive GABAA currents in striatal cholinergic interneurons through a PKA/PP1 cascade. Neuron 19, 1115–1126. doi: 10.1016/S0896-6273(00)80402-X
Yasuda, R. P., Ciesla, W., Flores, L. R., Wall, S. J., Li, M., Satkus, S. A., et al. (1993). Development of antisera selective for m4 and m5 muscarinic cholinergic receptors: distribution of m4 and m5 receptors in rat brain. Mol. Pharmacol. 43, 149–157. doi: 10.1016/S0026-895X(25)13594-3
Yorgason, J. T., Zeppenfeld, D. M., and Williams, J. T. (2017). Cholinergic interneurons underlie spontaneous dopamine release in nucleus Accumbens. J. Neurosci. 37, 2086–2096. doi: 10.1523/JNEUROSCI.3064-16.2017
Zachariou, V. (2000). Upregulation of galanin binding sites and GalR1 mRNA levels in the mouse locus coeruleus following chronic morphine treatments and precipitated morphine withdrawal. Neuropsychopharmacology 23, 127–137. doi: 10.1016/S0893-133X(00)00094-4
Zhang, W., Basile, A. S., Gomeza, J., Volpicelli, L. A., Levey, A. I., and Wess, J. (2002). Characterization of central inhibitory muscarinic autoreceptors by the use of muscarinic acetylcholine receptor knock-out mice. J. Neurosci. 22, 1709–1717. doi: 10.1523/JNEUROSCI.22-05-01709.2002
Zhang, Y.-F., and Cragg, S. J. (2017). Pauses in striatal cholinergic interneurons: what is revealed by their common themes and variations? Front. Syst. Neurosci. 11:296195. doi: 10.3389/fnsys.2017.00080
Zhang, Y. F., Reynolds, J. N. J., and Cragg, S. J. (2018). Pauses in cholinergic interneuron activity are driven by excitatory input and delayed rectification, with dopamine modulation. Neuron 98, 918–925.e3. doi: 10.1016/j.neuron.2018.04.027
Zoli, M., Moretti, M., Zanardi, A., McIntosh, J. M., Clementi, F., and Gotti, C. (2002). Identification of the nicotinic receptor subtypes expressed on dopaminergic terminals in the rat striatum. J. Neurosci. 22, 8785–8789. doi: 10.1523/JNEUROSCI.22-20-08785.2002
Ztaou, S., Maurice, N., Camon, J., Guiraudie-Capraz, G., Kerkerian-Le Goff, L., Beurrier, C., et al. (2016). Involvement of striatal cholinergic interneurons and M1 and M4 muscarinic receptors in motor symptoms of parkinson’s disease. J. Neurosci. 36, 9161–9172. doi: 10.1523/JNEUROSCI.0873-16.2016
Keywords: neuropeptides, neuromodulators, cholinergic interneurons, striatum, dopamine, acetylcholine, substance P, plasticity
Citation: Ratna D and Francis TC (2025) Extrinsic and intrinsic control of striatal cholinergic interneuron activity. Front. Mol. Neurosci. 18:1528419. doi: 10.3389/fnmol.2025.1528419
Edited by:
Zhe Jin, Uppsala University, SwedenReviewed by:
Margaret E. Rice, New York University, United StatesDavid Krolewski, University of Michigan, United States
Copyright © 2025 Ratna and Francis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tanner Chase Francis, ZnJhbmNpdEBtYWlsYm94LnNjLmVkdQ==