
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Mol. Neurosci., 05 January 2024
Sec. Brain Disease Mechanisms
Volume 16 - 2023 | https://doi.org/10.3389/fnmol.2023.1279999
This article is part of the Research TopicCommunication Breakdown: Synaptic pathology in ALS/FTDView all 8 articles
 Karrthik Krishnamurthy*
Karrthik Krishnamurthy* Raj Kumar Pradhan
Raj Kumar PradhanAmyotrophic Lateral Sclerosis (ALS) and Frontotemporal Dementia (FTD) are debilitating neurodegenerative diseases with shared pathological features like transactive response DNA-binding protein of 43 kDa (TDP-43) inclusions and genetic mutations. Both diseases involve synaptic dysfunction, contributing to their clinical features. Synaptic biomarkers, representing proteins associated with synaptic function or structure, offer insights into disease mechanisms, progression, and treatment responses. These biomarkers can detect disease early, track its progression, and evaluate therapeutic efficacy. ALS is characterized by elevated neurofilament light chain (NfL) levels in cerebrospinal fluid (CSF) and blood, correlating with disease progression. TDP-43 is another key ALS biomarker, its mislocalization linked to synaptic dysfunction. In FTD, TDP-43 and tau proteins are studied as biomarkers. Synaptic biomarkers like neuronal pentraxins (NPs), including neuronal pentraxin 2 (NPTX2), and neuronal pentraxin receptor (NPTXR), offer insights into FTD pathology and cognitive decline. Advanced technologies, like machine learning (ML) and artificial intelligence (AI), aid biomarker discovery and drug development. Challenges in this research include technological limitations in detection, variability across patients, and translating findings from animal models. ML/AI can accelerate discovery by analyzing complex data and predicting disease outcomes. Synaptic biomarkers offer early disease detection, personalized treatment strategies, and insights into disease mechanisms. While challenges persist, technological advancements and interdisciplinary efforts promise to revolutionize the understanding and management of ALS and FTD. This review will explore the present comprehension of synaptic biomarkers in ALS and FTD and discuss their significance and emphasize the prospects and obstacles.
Amyotrophic Lateral Sclerosis (ALS) and Frontotemporal Dementia (FTD) are two serious neurodegenerative conditions that cause significant suffering and morbidity globally. ALS, also known as Lou Gehrig's disease, primarily affects the neurons responsible for voluntary muscle control, resulting in gradual muscle weakness and eventually respiratory failure (Zarei et al., 2015). In contrast, FTD is a group of disorders that cause a gradual loss of nerve cells in the brain's frontal and temporal lobes, leading to problems with language, behavior, and motor skills (Boxer and Miller, 2005; Wen et al., 2014). While FTD and ALS are distinct disorders, they share some common pathological features, including the presence of transactive response DNA-binding protein of 43 kDa (TDP-43) inclusions and similar genetic mutations (Lillo and Hodges, 2009). In particular, synaptic dysfunction is increasingly recognized as a key element in ALS/FTD pathogenesis (Laszlo et al., 2022; Mora and Allodi, 2023).
There is currently no cure for these diseases and limited treatment options are available, making it urgent to improve our understanding of their underlying mechanisms and identify new therapeutic targets. Biomarkers play a crucial role in providing insights into disease mechanisms, progression, and therapeutic responses. Lately, there has been a focus on a particular type of biomarker known as synaptic biomarkers. These biomarkers consist of proteins related to synaptic function or components of synaptic structure, which shed light on the integrity of synapses (Camporesi et al., 2020). Synapses are essential junctions where neurons communicate, playing a vital role in cognitive and motor functions (Batool et al., 2019). Any alterations that occur at the synaptic level can indicate the beginning and advancement of neurodegenerative diseases.
The exploration of synaptic biomarkers in ALS and FTD brings about both possibilities and obstacles. These biomarkers have the potential to detect the disease early, monitor its progression, and evaluate therapeutic responses. However, studying these biomarkers is not without challenges. From the limitations in detecting and measuring them to the difficulty of translating findings from animal models to humans, there are significant hurdles to overcome.
Synaptic dysfunction is increasingly recognized as a converging mechanism in various genetic forms of ALS and FTD (Ling et al., 2013; Krishnamurthy et al., 2021; Gelon et al., 2022). Several genes linked to familial forms of ALS and FTD encode proteins that have roles at the synapse or are associated with synaptic function (Sephton and Yu, 2015; Gan et al., 2018; Casci et al., 2019; Ionescu et al., 2023).
The TDP-43 protein has also been implicated in synaptic function (Ling, 2018; Strah et al., 2020; Broadhead et al., 2022). Abnormal aggregation of TDP-43 in the cytoplasm of neurons is a key pathological feature of most ALS cases and a subset of FTD cases (Mackenzie and Rademakers, 2008). In a study by Feiler et al. (2015), TDP-43 was shown to undergo synaptic intercellular transmission and prion-like seeding.
FUS is another RNA-binding protein associated with familial forms of ALS and FTD (Da Cruz and Cleveland, 2011; Daigle et al., 2016). It has been implicated in the regulation of synaptic activity and local protein translation at synapses (Sephton et al., 2014). Super-resolution imaging and CLIP-seq revealed that FUS localizes near the vesicle reserve pool at synapses, influencing synapse organization and plasticity. Increased synaptic FUS during early ALS disease in a mouse model led to altered GABAergic synapses, indicating that synaptic FUS accumulation may trigger neurodegeneration (Sahadevan et al., 2021).
The most common genetic cause of both familial ALS and FTD is a hexanucleotide repeat expansion in the C9orf72 gene (Balendra and Isaacs, 2018). Recently, in a zebrafish model with reduced C9orf72 function, synaptic vesicle trafficking and release at the neuromuscular junction were affected, shedding light on its role in synaptic function (Butti et al., 2021). Evidence from studies on C9orf72 models indicates pathological changes, including altered dendritic spines, enhanced excitotoxicity, and increased activity of extrasynaptic NMDA receptors, suggesting that synaptic disturbances may be involved in FTD (Huber et al., 2022).
While more commonly associated with Alzheimer's disease, mutations in the MAPT gene, which encodes the Tau protein, have been linked to some cases of FTD (Mackenzie and Neumann, 2016). Tau is known to regulate synaptic function and plasticity (Brunello et al., 2020; Robbins et al., 2021) and Tau pathology is associated with synaptic loss (Coomans et al., 2021; Wu et al., 2021).
In summary, multiple lines of evidence suggest that synaptic dysfunction may be a converging mechanism in various genetic forms of ALS and FTD. However, further research is needed to fully elucidate the roles of these and other synaptic proteins in disease pathogenesis and to understand how synaptic dysfunction contributes to the clinical features of these diseases.
Synaptic biomarkers represent quantifiable indicators of synaptic function or structure (Colom-Cadena et al., 2020). Synapses are critical for neurotransmission and are essential for the cognitive and motor functions of the CNS. Proteins involved in the transmission of signals across synapses, as well as those forming the synaptic architecture, can serve as synaptic biomarkers. Alterations in the concentration or activity of these biomarkers can offer valuable information on the functional integrity of synapses and the overall neuronal network (Camporesi et al., 2020).
The significance of synaptic biomarkers in understanding disease pathogenesis is multi-fold. First, synaptic dysfunction is often an early feature in many neurodegenerative diseases, preceding neuronal loss. Therefore, synaptic biomarkers can potentially provide early indicators of disease onset (Galasko et al., 2019). Second, they can shed light on the mechanisms of synaptic damage in disease conditions, offering a deeper understanding of disease pathogenesis (Taoufik et al., 2018). Finally, the correlation of synaptic biomarkers with disease progression can help to map the trajectory of the disease, contributing to better prognostic models. Synaptic biomarkers can play a critical role in the development of therapeutics for ALS and FTD (Krishnamurthy and Pasinelli, 2021; Das et al., 2023).
Our understanding of synaptic dysfunction in ALS is mainly based on post-mortem immunohistochemistry (Aousji et al., 2023) but this approach cannot be applied in living subjects. Synaptic biomarkers are studied in ALS either through non-invasive or invasive methods. Broadly these methods can be classified as imaging-based or fluid-based. Through these methods we can gain valuable insights into synaptic changes in living individuals with ALS/FTD and track disease progression over time (Sturmey and Malaspina, 2022; Mead et al., 2023). Table 1 presents an overview of various methods to study synaptic biomarkers in ALS/FTD, along with their advantages and limitations.
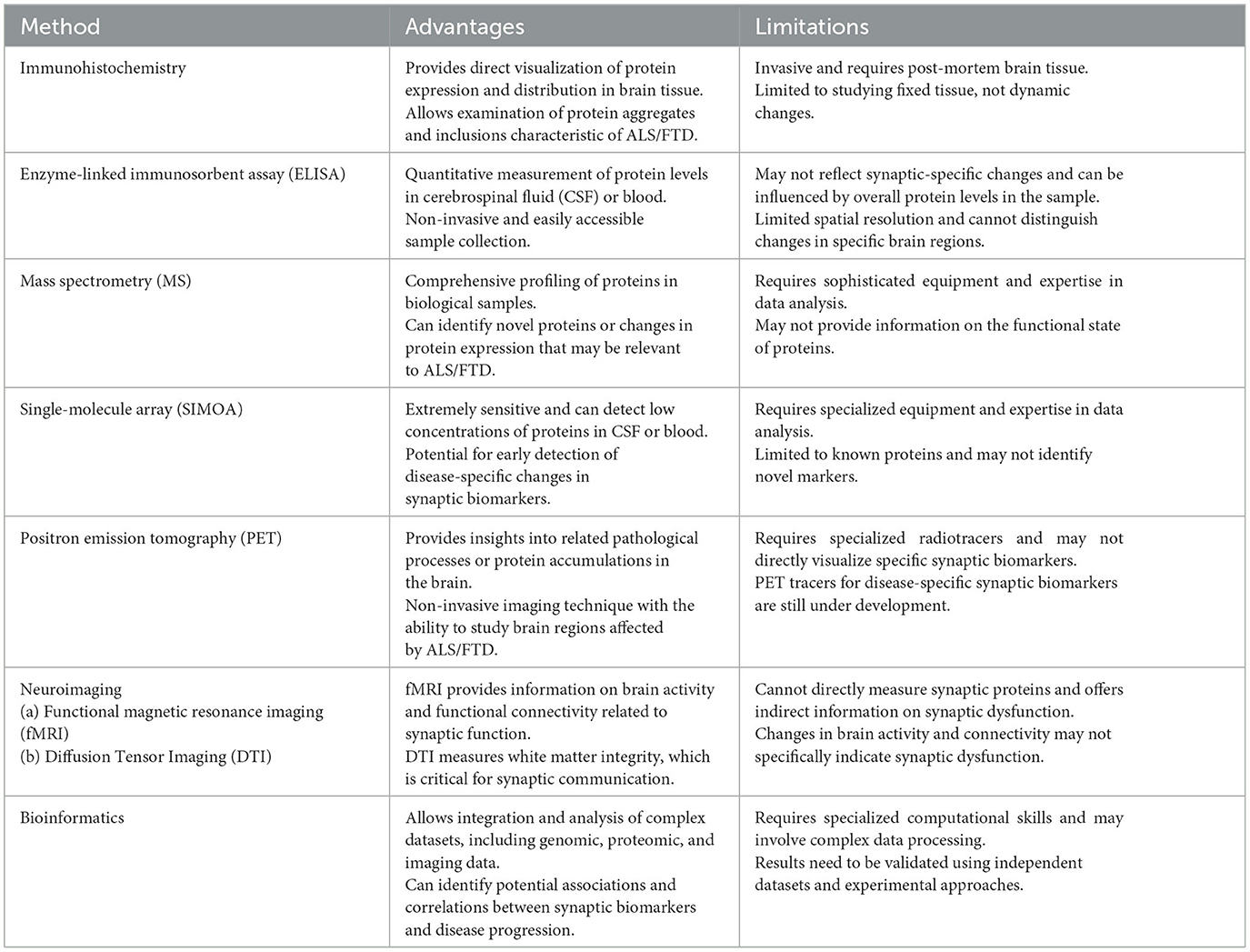
Table 1. Overview of methods to study synaptic biomarkers in ALS/FTD.
Neuroimaging techniques either report a specific biomarker or can reveal the overall status of synaptic structure and function in living subjects (Young et al., 2020). For example, Magnetic Resonance Imaging (MRI) can reveal changes in brain anatomy associated with synaptic loss or degeneration in ALS/FTD (Trojsi et al., 2012; Rajagopalan et al., 2013). Functional magnetic resonance imaging (fMRI) on the other hand can assess functional connectivity between brain regions related to synaptic activity (Peet et al., 2021). Diffusion Tensor Imaging (DTI) can investigate the integrity of white matter tracts connecting brain regions involved in synaptic transmission (Alruwaili et al., 2019). Positron Emission Tomography (PET) uses specific radiotracers to measure synaptic density or the presence of abnormal protein aggregates such as tau (Ni and Nitsch, 2022). PET imaging using radiotracers that bind to synaptic markers, such as synaptic vesicle glycoprotein 2A (SV2A) or synaptic density markers, can help visualize and quantify synaptic density and alterations in ALS/FTD (Serrano et al., 2022).
Fluid based biomarker analysis involves the use of bodily fluids such as blood and cerebrospinal fluid (CSF) that can be used to study levels of certain synaptic proteins or biomarkers that may correlate with synaptic dysfunction. Lumbar puncture allows the collection of CSF, which can be analyzed for synaptic biomarkers, neurotransmitters, or protein aggregates (Camporesi et al., 2020). Enzyme-Linked Immunosorbent Assay (ELISA) is one of the most commonly used technique used to study biomarkers associated with ALS and FTD allowing for the sensitive and quantitative measurement of specific proteins or molecules in biological samples. Magnetic Resonance Spectroscopy (MRS) allows researchers to measure the levels of various metabolites, including neurotransmitters like glutamate and GABA, which are essential for synaptic function (Pasanta et al., 2023). Another method of significant potential is targeted mass spectrometry (MS) which was recently utilized to explore 15 candidate synaptic biomarkers in CSF from patients with Parkinson's disease, corticobasal degeneration, progressive supranuclear palsy, multiple system atrophy, and Alzheimer's disease (Nilsson et al., 2023).
Numerous studies have been carried out to discover synaptic biomarkers that could be indicative of ALS. A growing body of studies suggests elevated levels of neurofilament light chain (NfL) in the CSF and blood of ALS patients. The levels of NfL were found to correlate with the progression of the disease, indicating that it could be a valuable biomarker for ALS (Sun et al., 2020; Dreger et al., 2021; Zhou et al., 2021; Thompson et al., 2022). The elevation of NfL is believed to reflect axonal damage resulting from motor neuron degeneration (Verde et al., 2021). Furthermore, TDP-43 has emerged as another important biomarker in ALS (Kasai et al., 2009). TDP-43 forms pathological inclusions in neurons in most ALS cases and its mislocalization may be associated with synaptic dysfunction (Lépine et al., 2023).
Research has shown the usefulness of synaptic biomarkers in understanding the progression and severity of ALS. For instance, individuals with higher NfL levels at diagnosis experienced a faster rate of disease progression (Lu et al., 2015; Sugimoto et al., 2020). This highlights the potential of this biomarker in monitoring disease progression and predicting prognosis. Additionally, evidence suggests that the amount of TDP-43 pathology is linked to the clinical symptoms of the disease, making it a promising biomarker for determining disease severity.
NfL and TDP-43 are two examples of biomarkers that are present in both ALS and other disorders like FTD (Katzeff et al., 2022). However, the particular characteristics of these markers, such as the pattern of TDP-43 pathology, suggest that they may be unique to ALS. Moreover, there are newly discovered potential synaptic biomarkers that could be specific to ALS, such as vesicle-associated membrane-protein-associated protein B (VAPB) which was absent in Parkinson's disease patient peripheral blood mononuclear cells (Cadoni et al., 2020). In a recent study, researchers used unbiased discovery-based methods along with targeted quantitative comparative analyses on CSF samples from ALS patients and healthy control individuals leading to the identification of 53 differential proteins between the two groups after CSF fractionation. These proteins included both previously known ones, validating the approach, and novel proteins with potential as new biomarkers (Oh et al., 2023). Next, the researchers used parallel reaction monitoring (PRM) mass spectrometry to further examine the identified proteins. Fifteen proteins were found to show significant differences between ALS and control groups. These proteins are: APOB, APP, CAMK2A, CHI3L1, CHIT1, CLSTN3, ERAP2, FSTL4, GPNMB, JCHAIN, L1CAM, NPTX2, SERPINA1, SERPINA3, and UCHL1. Among these CLSTN3, NPTX2, CAMK2A have synaptic roles. Further investigation is necessary to verify these biomarkers and comprehend their precise functions in ALS pathology.
As with ALS, several studies have aimed to identify potential synaptic biomarkers in FTD. Of these, TDP-43 has been the most studied, given its presence in neuronal inclusions in the majority of FTD cases (Neumann et al., 2006). Mislocalization and accumulation of TDP-43 in neurons are thought to disrupt synaptic function, potentially contributing to the cognitive and behavioral symptoms observed in FTD. Additionally, tau proteins, particularly in their hyperphosphorylated form, have been implicated in certain subtypes of FTD, known as tauopathies (Schraen-Maschke et al., 2008). The balance between phosphorylated tau (p-tau) and total tau (t-tau) in the CSF has been suggested as a potential biomarker for these cases (Hampel et al., 2010). Recent research has attempted to correlate synaptic biomarker levels with disease progression and severity in FTD. Interestingly, one study found that changes in CSF tau levels correlated with the progression of symptoms in tau-positive FTD, suggesting its potential role as a disease progression marker (Katzeff et al., 2022). More research is needed to understand how these and other potential synaptic biomarkers may correlate with disease progression and severity in FTD.
While FTD shares several potential synaptic biomarkers with ALS, such as TDP-43 and NfL, others are more unique to FTD. For example, tau proteins, especially in their hyperphosphorylated form, are particularly relevant to certain subtypes of FTD but not ALS (Bodea et al., 2016). Understanding these unique and common synaptic biomarkers can improve our understanding of these diseases, provide diagnostic and prognostic tools, and possibly highlight new therapeutic targets.
Neuronal pentraxins (NPs) are a family of proteins that play a crucial role in synaptic remodeling and homeostasis. They are named after the complement proteins due to their similar structure (Zhou et al., 2023). The family includes neuronal pentraxin 1 (NP1), neuronal pentraxin 2 (NPTX2), and neuronal pentraxin receptor (NPTXR). They have been shown to cluster at excitatory synapses and play a role in synaptic plasticity (Gómez de San José et al., 2022).
Lower levels of NPTX2 have been reported in the CSF of patients with FTD, suggesting a potential role as a synaptic biomarker (van der Ende et al., 2020). NPTXR, an integral component of the NP complex, is expressed primarily in neurons. Its expression level has been shown to correlate with cognitive function in FTD. Changes in NPTXR expression could therefore potentially serve as a marker of synaptic integrity in diseases like FTD (Sogorb-Esteve et al., 2022). In a recent study, Single-cell transcriptomics revealed a set of misregulated RNA targets shared by TDP-43 overexpressing neurons and patient brains with TDP-43 pathology, with the synaptic protein NPTX2 being consistently misaccumulated in ALS and FTD patient neurons providing a direct link between TDP-43 misregulation and NPTX2 accumulation (Hruska-Plochan et al., 2021).
One of the significant opportunities presented by synaptic biomarkers lies in their potential for early detection of ALS and FTD. Given that synaptic dysfunction often precedes neuronal loss, the presence of altered synaptic biomarkers could signal disease onset before the appearance of clinical symptoms. Additionally, biomarkers like NfL that correlate with disease progression could provide valuable prognostic information, guiding patient care and management (Swift et al., 2021; Vignaroli et al., 2023).
Synaptic biomarkers could pave the way for personalized medicine in ALS and FTD. By understanding how specific biomarkers correlate with disease progression and severity, treatments could be tailored based on a patient's unique biomarker profile (Das et al., 2023). This individualized approach could potentially lead to improved outcomes, mitigating disease progression and enhancing the quality of life for patients.
The study of synaptic biomarkers can also significantly contribute to drug development and clinical trials. By serving as objective measures of disease state or response to therapy, these biomarkers can provide critical insights into a drug's mechanism of action or efficacy (Colom-Cadena et al., 2020). For instance, reductions in biomarker levels following drug administration could indicate therapeutic effect, while no change might suggest a lack of efficacy. In this way, synaptic biomarkers can play a crucial role in the design and evaluation of clinical trials.
Perhaps one of the most exciting opportunities provided by synaptic biomarkers is the chance to further unravel the complex disease mechanisms underlying ALS and FTD. By exploring how these biomarkers change in response to disease progression and how they relate to one another, scientists can build a more complete picture of the molecular and cellular changes that drive these neurodegenerative diseases. This deeper understanding could, in turn, drive the development of new therapeutic strategies aimed at mitigating disease progression or even preventing disease onset. Table 2 provides a summary of the utility of synaptic biomarkers in therapeutic Development.
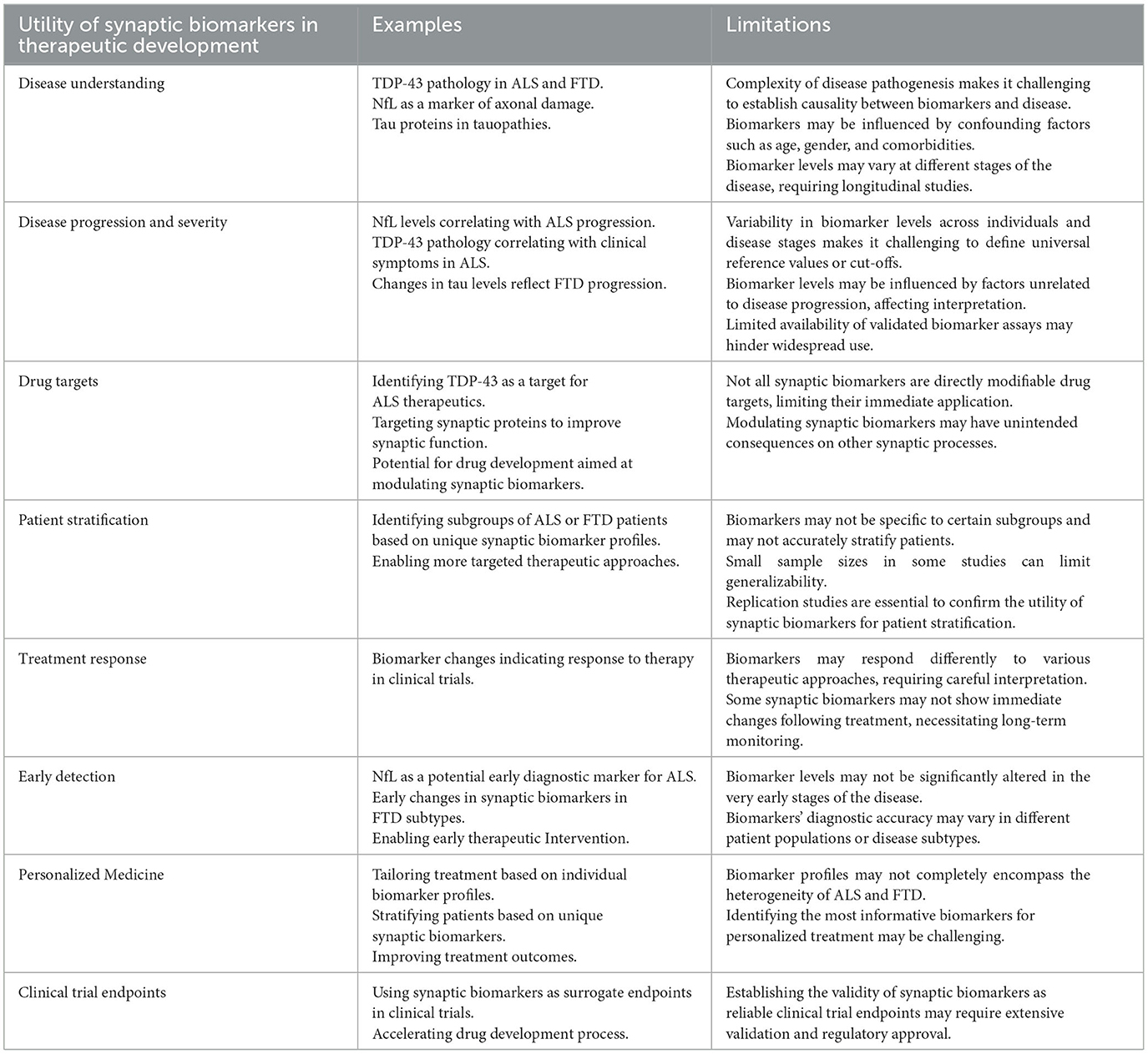
Table 2. Summary of the utility of synaptic biomarkers in therapeutic development.
One of the primary challenges in synaptic biomarker research is the technological limitation in detection and measurement. Synaptic biomarkers often require sensitive and specific assays for accurate detection. However, these biomarkers are usually present in minute quantities, and their levels may vary based on various factors, including the time of sample collection, sample handling, and patient-specific factors. This necessitates the development of more sensitive and robust assays for their detection (Klyucherev et al., 2022).
There is considerable variability in biomarker levels across different patients and stages of diseases, adding to the complexity of interpreting these markers. Factors such as genetic heterogeneity, disease subtype, age, gender, and comorbidities can influence biomarker levels, making it challenging to establish universally applicable cut-offs or reference values. Moreover, understanding how biomarker levels change over the course of the disease and correlating these changes with clinical progression is a complex task (Mayeux, 2004; McDermott et al., 2013).
While animal models have greatly contributed to our understanding of ALS and FTD, translating these findings to humans has been challenging. The pathophysiology of these diseases in animal models may not perfectly mimic the human condition, and hence, the biomarkers identified in these models may not be relevant in humans. Thus, findings from animal studies must be interpreted cautiously and validated in human populations (Bonifacino et al., 2021).
Many biomarker studies are based on small sample sizes, and their findings may not be generalizable to larger, more diverse patient populations. Therefore, there is a pressing need for larger, multi-center studies to validate these markers. Additionally, replication studies are crucial to ensure the reliability of these findings (Freidlin et al., 2010). As biomarker research is a rapidly evolving field, continuous re-evaluation and updating of these markers based on the latest evidence is necessary.
ML and AI have the potential to significantly accelerate synaptic biomarker discovery and therapeutic development for ALS and FTD. These advanced computational approaches can process large datasets and identify patterns and relationships that may be difficult to uncover with traditional methods.
For example, ML/AI algorithms can analyze complex multi-omics data, including genomic, proteomic, and neuroimaging data, to find novel synaptic biomarkers associated with ALS/FTD (Grollemund et al., 2019). By integrating diverse datasets, ML/AI models can reveal previously unknown connections between genes, proteins, and disease features, as has been shown in cancer research (Arjmand et al., 2022).
ML/AI techniques can also analyze clinical, imaging, and biomarker data to predict disease progression and estimate the likelihood of developing ALS/FTD (Rajagopalan et al., 2023). Early and accurate diagnosis and prognosis can inform personalized treatment strategies to improve patient outcomes.
Additionally, ML/AI can efficiently screen chemical libraries to identify potential ALS/FTD drug candidates and repurpose existing medications that may have therapeutic effects (Vatansever et al., 2021). By integrating molecular and genetic data, ML/AI can predict the most promising targets involved in synaptic dysfunction to prioritize for drug development. ML/AI can also optimize clinical trial design, including patient selection and treatment dosing, to increase the probability of success (Davenport and Kalakota, 2019). Analyzing patient-specific data with AI algorithms allows for personalized medicine by predicting individual responses to treatments.
Finally, ML/AI can quantify synaptic density and structural changes in the brain from neuroimaging data to track disease progression and monitor treatment efficacy (Boyle et al., 2021; Monsour et al., 2022). In summary, ML/AI has diverse applications that can accelerate nearly every aspect of ALS/FTD research and drug development, from biomarker discovery to personalized therapeutics.
Emerging technologies offer new avenues for improving the detection and measurement of synaptic biomarkers. Advances in genomics, proteomics, and metabolomics, coupled with powerful bioinformatics tools, are enabling the identification and validation of biomarkers at an unprecedented scale and speed. Additionally, the development of more sensitive and specific immunoassays, such as single-molecule array (SIMOA) technology, can greatly enhance our ability to detect minute quantities of these biomarkers (Wilson et al., 2016; Wu et al., 2022). Continued technological innovation will undoubtedly play a crucial role in the future of synaptic biomarker research.
Looking forward, there are several exciting new areas of synaptic biomarker research. For instance, exploring the interplay between synaptic biomarkers and other pathophysiological processes, such as inflammation or oxidative stress, could provide novel insights into disease mechanisms. Additionally, the emerging field of neuroimaging biomarkers could complement biochemical biomarker studies, providing a more holistic picture of disease progression. Lastly, research into biomarkers of synaptic resilience or compensatory mechanisms could open new avenues for therapeutic intervention.
Addressing the current challenges in synaptic biomarker research could significantly shape the future direction of this field. For example, overcoming technological limitations could allow for the detection of previously unidentified biomarkers or more accurate quantification of known ones. Tackling variability issues could lead to the development of personalized biomarker profiles, providing more precise diagnostic and prognostic information. Successfully translating findings from animal models to humans could accelerate the clinical application of these biomarkers, and large-scale validation studies could solidify their place in clinical practice. Thus, the future of synaptic biomarker research is contingent upon overcoming the present challenges, offering a promising and exciting path forward.
KK: Conceptualization, Resources, Visualization, Writing—original draft, Writing—review & editing. RP: Resources, Visualization, Software, Writing—review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. Publication made possible in part by generous support from the Thomas Jefferson University Open Access Fund.
Author would like to thank the global ALS/FTD patient community for the inspiration.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Alruwaili, A. R., Pannek, K., Henderson, R. D., Gray, M., Kurniawan, N. D., and McCombe, P. A. (2019). Tract integrity in amyotrophic lateral sclerosis: 6–month evaluation using MR diffusion tensor imaging. BMC Med. Imag. 19, 19. doi: 10.1186/s12880-019-0319-3
Aousji, O., Feldengut, S., Antonucci, S., Schön, M., Boeckers, T. M., Matschke, J., et al. (2023). Patterns of synaptic loss in human amyotrophic lateral sclerosis spinal cord: a clinicopathological study. Acta Neuropathol. Communic. 11, 120. doi: 10.1186/s40478-023-01616-8
Arjmand, B., Hamidpour, S. K., Tayanloo-Beik, A., Goodarzi, P., Aghayan, H. R., Adibi, H., et al. (2022). Machine learning: a new prospect in multi-omics data analysis of cancer. Front. Genet. 13, 824451. doi: 10.3389/fgene.2022.824451
Balendra, R., and Isaacs, A. M. (2018). C9orf72-mediated ALS and FTD: multiple pathways to disease. Nat. Rev. Neurol. 14, 544–558. doi: 10.1038/s41582-018-0047-2
Batool, S., Raza, H., Zaidi, J., Riaz, S., Hasan, S., and Syed, N. I. (2019). Synapse formation: from cellular and molecular mechanisms to neurodevelopmental and neurodegenerative disorders. J. Neurophysiol. 121, 1381–1397. doi: 10.1152/jn.00833.2018
Bodea, L., Eckert, A., Ittner, L. M., Piguet, O., and Götz, J. (2016). Tau physiology and pathomechanisms in frontotemporal lobar degeneration. J. Neurochem. 138, 71–94. doi: 10.1111/jnc.13600
Bonifacino, T., Zerbo, R. A., Balbi, M., Torazza, C., Frumento, G., Fedele, E., et al. (2021). Nearly 30 years of animal models to study amyotrophic lateral sclerosis: a historical overview and future perspectives. Int. J. Mol. Sci. 22, 12236. doi: 10.3390/ijms222212236
Boxer, A. L., and Miller, B. L. (2005). Clinical features of frontotemporal dementia. Alzheimer Dis. Assoc. Disor. 19, S3. doi: 10.1097/01.wad.0000183086.99691.91
Boyle, A. J., Gaudet, V. C., Black, S. E., Vasdev, N., Rosa-Neto, P., and Zukotynski, K. A. (2021). Artificial intelligence for molecular neuroimaging. Ann. Transl. Med. 9, 822. doi: 10.21037/atm-20-6220
Broadhead, M. J., Doucet, K., Kantelberg, O., Zhu, F., Grant, S. G., Horrocks, M. H., et al. (2022). Synaptic expression of TAR-DNA-binding protein 43 in the mouse spinal cord determined using super-resolution Microscopy. 29, 610. doi: 10.1101/2022.08.29.505610
Brunello, C. A., Merezhko, M., Uronen, R.-L., and Huttunen, H. J. (2020). Mechanisms of secretion and spreading of pathological tau protein. Cell. Mol. Life Sci. 77, 1721–1744. doi: 10.1007/s00018-019-03349-1
Butti, Z., Pan, Y. E., Giacomotto, J., and Patten, S. A. (2021). Reduced C9orf72 function leads to defective synaptic vesicle release and neuromuscular dysfunction in zebrafish. Commun. Biol. 4, 1–16. doi: 10.1038/s42003-021-02302-y
Cadoni, M. P. L., Biggio, M. L., Arru, G., Secchi, G., Orr,ù, N., Clemente, M. G., et al. (2020). VAPB ER-aggregates, a possible new biomarker in ALS pathology. Cells 9, 164. doi: 10.3390/cells9010164
Camporesi, E., Nilsson, J., Brinkmalm, A., Becker, B., Ashton, N. J., Blennow, K., et al. (2020). Fluid biomarkers for synaptic dysfunction and loss. Biomark Insights 15, 1177271920950319. doi: 10.1177/1177271920950319
Casci, I., Krishnamurthy, K., Kour, S., Tripathy, V., Ramesh, N., Anderson, E. N., et al. (2019). Muscleblind acts as a modifier of FUS toxicity by modulating stress granule dynamics and SMN localization. Nat. Commun. 10, 5583. doi: 10.1038/s41467-019-13383-z
Colom-Cadena, M., Spires-Jones, T., Zetterberg, H., Blennow, K., Caggiano, A., DeKosky, S. T., et al. (2020). The clinical promise of biomarkers of synapse damage or loss in Alzheimer's disease. Alzheimer's Res. Ther. 12, 21. doi: 10.1186/s13195-020-00588-4
Coomans, E. M., Schoonhoven, D. N., Tuncel, H., Verfaillie, S. C. J., Wolters, E. E., Boellaard, R., et al. (2021). In vivo tau pathology is associated with synaptic loss and altered synaptic function. Alzheimer's Res. Ther. 13, 35. doi: 10.1186/s13195-021-00772-0
Da Cruz, S., and Cleveland, D. W. (2011). Understanding the role of TDP-43 and FUS/TLS in ALS and beyond. Curr. Opin. Neurobiol. 21, 904–919. doi: 10.1016/j.conb.2011.05.029
Daigle, J. G., Krishnamurthy, K., Ramesh, N., Casci, I., Monaghan, J., McAvoy, K., et al. (2016). Pur-alpha regulates cytoplasmic stress granule dynamics and ameliorates FUS toxicity. Acta Neuropathol. 131, 605–620. doi: 10.1007/s00401-015-1530-0
Das, S., Goossens, J., Jacobs, D., Dewit, N., Pijnenburg, Y. A. L., In ‘t Veld, S. G. J. G., et al. (2023). Synaptic biomarkers in the cerebrospinal fluid associate differentially with classical neuronal biomarkers in patients with Alzheimer's disease and frontotemporal dementia. Alzheimer's Res. Ther. 15, 62. doi: 10.1186/s13195-023-01212-x
Davenport, T., and Kalakota, R. (2019). The potential for artificial intelligence in healthcare. Fut. Healthc. J. 6, 94–98. doi: 10.7861/futurehosp.6-2-94
Dreger, M., Steinbach, R., Gaur, N., Metzner, K., Stubendorff, B., Witte, O. W., et al. (2021). Cerebrospinal fluid neurofilament light chain (NfL) predicts disease aggressiveness in amyotrophic lateral sclerosis: an application of the D50 disease progression model. Front. Neurosci. 15, 651651. doi: 10.3389/fnins.2021.651651
Feiler, M. S., Strobel, B., Freischmidt, A., Helferich, A. M., Kappel, J., Brewer, B. M., et al. (2015). TDP-43 is intercellularly transmitted across axon terminals. J. Cell Biol. 211, 897–911. doi: 10.1083/jcb.201504057
Freidlin, B., McShane, L. M., and Korn, E. L. (2010). Randomized clinical trials with biomarkers: design issues. J. Natl. Cancer Inst. 102, 152–160. doi: 10.1093/jnci/djp477
Galasko, D., Xiao, M., Xu, D., Smirnov, D., Salmon, D. P., Dewit, N., et al. (2019). Synaptic biomarkers in CSF aid in diagnosis, correlate with cognition and predict progression in MCI and Alzheimer's disease. Alzheimers Dem. 5, 871–882. doi: 10.1016/j.trci.2019.11.002
Gan, L., Cookson, M. R., Petrucelli, L., and La Spada, A. R. (2018). Converging pathways in neurodegeneration, from genetics to mechanisms. Nat. Neurosci. 21, 1300–1309. doi: 10.1038/s41593-018-0237-7
Gelon, P. A., Dutchak, P. A., and Sephton, C. F. (2022). Synaptic dysfunction in ALS and FTD: anatomical and molecular changes provide insights into mechanisms of disease. Front. Mol. Neurosci. 15, 1000183. doi: 10.3389/fnmol.2022.1000183
Gómez de San José, N., Massa, F., Halbgebauer, S., Oeckl, P., Steinacker, P., and Otto, M. (2022). Neuronal pentraxins as biomarkers of synaptic activity: from physiological functions to pathological changes in neurodegeneration. J. Neural. Transm. 129, 207–230. doi: 10.1007/s00702-021-02411-2
Grollemund, V., Pradat, P.-F., Querin, G., Delbot, F., Le Chat, G., Pradat-Peyre, J.-F., et al. (2019). Machine learning in amyotrophic lateral sclerosis: achievements, pitfalls, and future directions. Front. Neurosci. 13, 135. doi: 10.3389/fnins.2019.00135
Hampel, H., Blennow, K., Shaw, L. M., Hoessler, Y. C., Zetterberg, H., and Trojanowski, J. Q. (2010). Total and phosphorylated tau protein as biological markers of Alzheimer's disease. Exp. Gerontol. 45, 30. doi: 10.1016/j.exger.2009.10.010
Hruska-Plochan, M., Betz, K. M., Ronchi, S., Wiersma, V. I., Maniecka, Z., Hock, E.-M., et al. (2021). Human neural networks with sparse TDP-43 pathology reveal NPTX2 misregulation in ALS/FTLD. bioRxiv, 2021-12. doi: 10.1101/2021.12.08.471089
Huber, N., Hoffmann, D., Giniatullina, R., Rostalski, H., Leskel,ä, S., Takalo, M., et al. (2022). C9orf72 hexanucleotide repeat expansion leads to altered neuronal and dendritic spine morphology and synaptic dysfunction. Neurobiol. Dis. 162, 105584. doi: 10.1016/j.nbd.2021.105584
Ionescu, A., Altman, T., and Perlson, E. (2023). Looking for answers far away from the soma—the (un)known axonal functions of TDP-43, and their contribution to early NMJ disruption in ALS. Molec. Neurodegener. 18, 35. doi: 10.1186/s13024-023-00623-6
Kasai, T., Tokuda, T., Ishigami, N., Sasayama, H., Foulds, P., Mitchell, D. J., et al. (2009). Increased TDP-43 protein in cerebrospinal fluid of patients with amyotrophic lateral sclerosis. Acta Neuropathol. 117, 55–62. doi: 10.1007/s00401-008-0456-1
Katzeff, J. S., Bright, F., Phan, K., Kril, J. J., Ittner, L. M., Kassiou, M., et al. (2022). Biomarker discovery and development for frontotemporal dementia and amyotrophic lateral sclerosis. Brain 145, 1598–1609. doi: 10.1093/brain/awac077
Klyucherev, T. O., Olszewski, P., Shalimova, A. A., Chubarev, V. N., Tarasov, V. V., Attwood, M. M., et al. (2022). Advances in the development of new biomarkers for Alzheimer's disease. Transl. Neurodegen. 11, 25. doi: 10.1186/s40035-022-00296-z
Krishnamurthy, K., and Pasinelli, P. (2021). Synaptic dysfunction in ALS/FTD- Therapeutic strategies and novel biomarkers. J. Neurosci. Res. 99, 1499–1503. doi: 10.1002/jnr.24824
Krishnamurthy, K., Trotti, D., Pasinelli, P., and Jensen, B. (2021). Real-time fluorescent measurement of synaptic functions in models of amyotrophic lateral sclerosis. JoVE. 173, e62813. doi: 10.3791/62813-v
Laszlo, Z. I., Hindley, N., Sanchez Avila, A., Kline, R. A., Eaton, S. L., Lamont, D. J., et al. (2022). Synaptic proteomics reveal distinct molecular signatures of cognitive change and C9ORF72 repeat expansion in the human ALS cortex. Acta Neuropathol. Commun. 10, 156. doi: 10.1186/s40478-022-01455-z
Lépine, S., Nauleau-Javaudin, A., Deneault, E., Chen, C. X.-Q., Abdian, N., Franco-Flores, A. K., et al. (2023). Homozygous ALS-linked mutations in TARDBP/TDP-43 lead to progressive synaptic dysfunction in human iPSC-derived motor neurons. bioRxiv, 2023-03. doi: 10.1101/2023.03.22.533562
Lillo, P., and Hodges, J. R. (2009). Frontotemporal dementia and motor neurone disease: overlapping clinic-pathological disorders. J. Clin. Neurosci. 16, 1131–1135. doi: 10.1016/j.jocn.2009.03.005
Ling, S.-C. (2018). Synaptic paths to neurodegeneration: the emerging role of TDP-43 and FUS in synaptic functions. Neural Plast. 2018, e8413496. doi: 10.1155/2018/8413496
Ling, S.-C., Polymenidou, M., and Cleveland, D. W. (2013). Converging mechanisms in ALS and FTD: disrupted RNA and protein homeostasis. Neuron 79, 416–438. doi: 10.1016/j.neuron.2013.07.033
Lu, C.-H., Macdonald-Wallis, C., Gray, E., Pearce, N., Petzold, A., Norgren, N., et al. (2015). Neurofilament light chain: a prognostic biomarker in amyotrophic lateral sclerosis. Neurology 84, 2247–2257. doi: 10.1212/WNL.0000000000001642
Mackenzie, I. R., and Rademakers, R. (2008). The role of transactive response DNA-binding protein-43 in amyotrophic lateral sclerosis and frontotemporal dementia. Curr. Opin. Neurol. 21, 693. doi: 10.1097/WCO.0b013e3283168d1d
Mackenzie, I. R. A., and Neumann, M. (2016). Molecular neuropathology of frontotemporal dementia: insights into disease mechanisms from postmortem studies. J. Neurochem. 138, 54–70. doi: 10.1111/jnc.13588
Mayeux, R. (2004). Biomarkers: potential uses and limitations. NeuroRx 1, 182–188. doi: 10.1602/neurorx.1.2.182
McDermott, J. E., Wang, J., Mitchell, H., Webb-Robertson, B.-J., Hafen, R., Ramey, J., et al. (2013). Challenges in biomarker discovery: combining expert insights with statistical analysis of complex omics data. Expert. Opin. Med. Diagn. 7, 37–51. doi: 10.1517/17530059.2012.718329
Mead, R. J., Shan, N., Reiser, H. J., Marshall, F., and Shaw, P. J. (2023). Amyotrophic lateral sclerosis: a neurodegenerative disorder poised for successful therapeutic translation. Nat. Rev. Drug. Discov. 22, 185–212. doi: 10.1038/s41573-022-00612-2
Monsour, R., Dutta, M., Mohamed, A.-Z., Borkowski, A., and Viswanadhan, N. A. (2022). Neuroimaging in the era of artificial intelligence: current applications. Fed. Pract. 39, S14–S20. doi: 10.12788/fp.0231
Mora, S., and Allodi, I. (2023). Neural circuit and synaptic dysfunctions in ALS-FTD pathology. Front. Neural Circ. 17, 1208876. doi: 10.3389/fncir.2023.1208876
Neumann, M., Sampathu, D. M., Kwong, L. K., Truax, A. C., Micsenyi, M. C., Chou, T. T., et al. (2006). Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science 314, 130–133. doi: 10.1126/science.1134108
Ni, R., and Nitsch, R. M. (2022). Recent developments in positron emission tomography tracers for proteinopathies imaging in dementia. Front. Aging Neurosci. 13, 751897. doi: 10.3389/fnagi.2021.751897
Nilsson, J., Cousins, K. A. Q., Gobom, J., Portelius, E., Chen-Plotkin, A., Shaw, L. M., et al. (2023). Cerebrospinal fluid biomarker panel of synaptic dysfunction in Alzheimer's disease and other neurodegenerative disorders. Alzheimer's Dem. 19, 1775–1784. doi: 10.1002/alz.12809
Oh, S., Jang, Y., and Na, C. H. (2023). Discovery of biomarkers for amyotrophic lateral sclerosis from human cerebrospinal fluid using mass-spectrometry-based proteomics. Biomedicines 11, 1250. doi: 10.3390/biomedicines11051250
Pasanta, D., He, J. L., Ford, T., Oeltzschner, G., Lythgoe, D. J., and Puts, N. A. (2023). Functional MRS studies of GABA and Glutamate/Glx – a systematic review and meta-analysis. Neurosci. Biobehav. Rev. 144, 104940. doi: 10.1016/j.neubiorev.2022.104940
Peet, B. T., Spina, S., Mundada, N., and La Joie, R. (2021). Neuroimaging in frontotemporal dementia: heterogeneity and relationships with underlying neuropathology. Neurotherapeutics 18, 728–752. doi: 10.1007/s13311-021-01101-x
Rajagopalan, V., Chaitanya, K. G., and Pioro, E. P. (2023). Quantitative brain MRI metrics distinguish four different ALS phenotypes: a machine learning based study. Diagnostics 13, 1521. doi: 10.3390/diagnostics13091521
Rajagopalan, V., Liu, Z., Allexandre, D., Zhang, L., Wang, X.-F., Pioro, E. P., et al. (2013). Brain white matter shape changes in amyotrophic lateral sclerosis (ALS): a fractal dimension study. PLoS ONE 8, e73614. doi: 10.1371/journal.pone.0073614
Robbins, M., Clayton, E., and Kaminski Schierle, G. S. (2021). Synaptic tau: a pathological or physiological phenomenon? Acta Neuropathol. Commun. 9, 149. doi: 10.1186/s40478-021-01246-y
Sahadevan, S., Hembach, K. M., Tantardini, E., Pérez-Berlanga, M., Hruska-Plochan, M., Megat, S., et al. (2021). Synaptic FUS accumulation triggers early misregulation of synaptic RNAs in a mouse model of ALS. Nat. Commun. 12, 3027. doi: 10.1038/s41467-021-23188-8
Schraen-Maschke, S., Sergeant, N., Dhaenens, C.-M., Bombois, S., Deramecourt, V., Caillet-Boudin, M.-L., et al. (2008). Tau as a biomarker of neurodegenerative diseases. Biomark. Med. 2, 363–384. doi: 10.2217/17520363.2.4.363
Sephton, C. F., Tang, A. A., Kulkarni, A., West, J., Brooks, M., Stubblefield, J. J., et al. (2014). Activity-dependent FUS dysregulation disrupts synaptic homeostasis. Proc. Natl. Acad. Sci. 111, E4769–E4778. doi: 10.1073/pnas.1406162111
Sephton, C. F., and Yu, G. (2015). The function of RNA-binding proteins at the synapse: implications for neurodegeneration. Cell. Mol. Life Sci. 72, 3621–3635. doi: 10.1007/s00018-015-1943-x
Serrano, M. E., Kim, E., Petrinovic, M. M., Turkheimer, F., and Cash, D. (2022). Imaging synaptic density: the next holy grail of neuroscience? Front. Neurosci. 16, 796129. doi: 10.3389/fnins.2022.796129
Sogorb-Esteve, A., Nilsson, J., Swift, I. J., Heller, C., Bocchetta, M., Russell, L. L., et al. (2022). Differential impairment of cerebrospinal fluid synaptic biomarkers in the genetic forms of frontotemporal dementia. Alzheimer's Res. Ther. 14, 118. doi: 10.1186/s13195-022-01042-3
Strah, N., Romano, G., Introna, C., Klima, R., Marzullo, M., Ciapponi, L., et al. (2020). TDP-43 promotes the formation of neuromuscular synapses through the regulation of Disc-large expression in Drosophila skeletal muscles. BMC Biol. 18, 34. doi: 10.1186/s12915-020-00767-7
Sturmey, E., and Malaspina, A. (2022). Blood biomarkers in ALS: challenges, applications and novel frontiers. Acta Neurol. Scandin. 146, 375–388. doi: 10.1111/ane.13698
Sugimoto, K., Han, Y., Song, Y., and Gao, Y. (2020). Correlational analysis of ALS progression and serum NfL measured by simoa assay in Chinese patients. Front. Neurol. 11, 579094. doi: 10.3389/fneur.2020.579094
Sun, Q., Zhao, X., Li, S., Yang, F., Wang, H., Cui, F., et al. (2020). CSF neurofilament light chain elevation predicts ALS severity and progression. Front. Neurol. 11, 919. doi: 10.3389/fneur.2020.00919
Swift, I. J., Sogorb-Esteve, A., Heller, C., Synofzik, M., Otto, M., Graff, C., et al. (2021). Fluid biomarkers in frontotemporal dementia: past, present and future. J. Neurol. Neurosurg. Psychiat. 92, 204–215. doi: 10.1136/jnnp-2020-323520
Taoufik, E., Kouroupi, G., Zygogianni, O., and Matsas, R. (2018). Synaptic dysfunction in neurodegenerative and neurodevelopmental diseases: an overview of induced pluripotent stem-cell-based disease models. Open Biol. 8, 180138. doi: 10.1098/rsob.180138
Thompson, A. G., Gray, E., Verber, N., Bobeva, Y., Lombardi, V., Shepheard, S. R., et al. (2022). Multicentre appraisal of amyotrophic lateral sclerosis biofluid biomarkers shows primacy of blood neurofilament light chain. Brain Commun. 4, fcac029. doi: 10.1093/braincomms/fcac029
Trojsi, F., Monsurr,ò, M. R., Esposito, F., and Tedeschi, G. (2012). Widespread structural and functional connectivity changes in amyotrophic lateral sclerosis: insights from advanced neuroimaging research. Neural. Plasticity 2012, e473538. doi: 10.1155/2012/473538
van der Ende, E. L., Xiao, M., Xu, D., Poos, J. M., Panman, J. L., Jiskoot, L. C., et al. (2020). Neuronal pentraxin 2: a synapse-derived CSF biomarker in genetic frontotemporal dementia. J. Neurol. Neurosurg. Psychiat. 91, 612–621. doi: 10.1136/jnnp-2019-322493
Vatansever, S., Schlessinger, A., Wacker, D., Kaniskan, H. Ü., Jin, J., Zhou, M.-M., et al. (2021). Artificial intelligence and machine learning-aided drug discovery in central nervous system diseases: state-of-the-arts and future directions. Med. Res. Rev. 41, 1427–1473. doi: 10.1002/med.21764
Verde, F., Otto, M., and Silani, V. (2021). Neurofilament light chain as biomarker for amyotrophic lateral sclerosis and frontotemporal dementia. Front. Neurosci. 15, 679199. doi: 10.3389/fnins.2021.679199
Vignaroli, F., Mele, A., Tondo, G., De Giorgis, V., Manfredi, M., Comi, C., et al. (2023). The need for biomarkers in the ALS–FTD spectrum: a clinical point of view on the role of proteomics. Proteomes 11, 1. doi: 10.3390/proteomes11010001
Wen, X., Tan, W., Westergard, T., Krishnamurthy, K., Markandaiah, S. S., Shi, Y., et al. (2014). Antisense proline-arginine RAN dipeptides linked to C9ORF72-ALS/FTD form toxic nuclear aggregates that initiate in vitro and in vivo neuronal death. Neuron 84, 1213–1225. doi: 10.1016/j.neuron.2014.12.010
Wilson, D. H., Rissin, D. M., Kan, C. W., Fournier, D. R., Piech, T., Campbell, T. G., et al. (2016). The simoa HD-1 analyzer: a novel fully automated digital immunoassay analyzer with single-molecule sensitivity and multiplexing. SLAS Technol. 21, 533–547. doi: 10.1177/2211068215589580
Wu, C., Dougan, T. J., and Walt, D. R. (2022). High-throughput, high-multiplex digital protein detection with attomolar sensitivity. ACS Nano 16, 1025–1035. doi: 10.1021/acsnano.1c08675
Wu, M., Zhang, M., Yin, X., Chen, K., Hu, Z., Zhou, Q., et al. (2021). The role of pathological tau in synaptic dysfunction in Alzheimer's diseases. Transl. Neurodeg. 10, 45. doi: 10.1186/s40035-021-00270-1
Young, P. N. E., Estarellas, M., Coomans, E., Srikrishna, M., Beaumont, H., Maass, A., et al. (2020). Imaging biomarkers in neurodegeneration: current and future practices. Alzheimer's Res. Ther. 12, 49. doi: 10.1186/s13195-020-00612-7
Zarei, S., Carr, K., Reiley, L., Diaz, K., Guerra, O., Altamirano, P. F., et al. (2015). A comprehensive review of amyotrophic lateral sclerosis. Surg. Neurol. Int. 6, 171. doi: 10.4103/2152-7806.169561
Zhou, J., Wade, S. D., Graykowski, D., Xiao, M.-F., Zhao, B., Giannini, L. A. A., et al. (2023). The neuronal pentraxin Nptx2 regulates complement activity and restrains microglia-mediated synapse loss in neurodegeneration. Sci. Transl. Med. 15, eadf0141. doi: 10.1126/scitranslmed.adf0141
Keywords: amyotrophic lateral sclerosis, frontotemporal dementia, synaptic dysfunction, synaptic biomarkers, TDP-43, neuronal pentraxin, C9orf72, artificial intelligence
Citation: Krishnamurthy K and Pradhan RK (2024) Emerging perspectives of synaptic biomarkers in ALS and FTD. Front. Mol. Neurosci. 16:1279999. doi: 10.3389/fnmol.2023.1279999
Received: 19 August 2023; Accepted: 01 December 2023;
Published: 05 January 2024.
Edited by:
Chris Henstridge, University of Dundee, United KingdomReviewed by:
Aitana Sogorb-Esteve, University College London, United KingdomCopyright © 2024 Krishnamurthy and Pradhan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Karrthik Krishnamurthy, a3hrMDU2QGplZmZlcnNvbi5lZHU=; a3Jpc2huYWJpb0BnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.