 Micaël Carrier
Micaël Carrier Eva Šimončičová
Eva Šimončičová Marie-Kim St-Pierre
Marie-Kim St-Pierre Chloe McKee
Chloe McKee Marie-Ève Tremblay
Marie-Ève Tremblay- 1Axe Neurosciences, Centre de Recherche du CHU de Québec, Université Laval, Québec City, QC, Canada
- 2Division of Medical Sciences, University of Victoria, Victoria, BC, Canada
- 3Department of Molecular Medicine, Université Laval, Québec City, QC, Canada
- 4Department of Biology, University of Victoria, Victoria, BC, Canada
- 5Neurology and Neurosurgery Department, McGill University, Montreal, QC, Canada
- 6Department of Biochemistry and Molecular Biology, University of British Columbia, Vancouver, BC, Canada
The relationship between the central nervous system (CNS) and microglia is lifelong. Microglia originate in the embryonic yolk sac during development and populate the CNS before the blood-brain barrier forms. In the CNS, they constitute a self-renewing population. Although they represent up to 10% of all brain cells, we are only beginning to understand how much brain homeostasis relies on their physiological functions. Often compared to a double-edged sword, microglia hold the potential to exert neuroprotective roles that can also exacerbate neurodegeneration once compromised. Microglia can promote synaptic growth in addition to eliminating synapses that are less active. Synaptic loss, which is considered one of the best pathological correlates of cognitive decline, is a distinctive feature of major depressive disorder (MDD) and cognitive aging. Long-term psychological stress accelerates cellular aging and predisposes to various diseases, including MDD, and cognitive decline. Among the underlying mechanisms, stress-induced neuroinflammation alters microglial interactions with the surrounding parenchymal cells and exacerbates oxidative burden and cellular damage, hence inducing changes in microglia and neurons typical of cognitive aging. Focusing on microglial interactions with neurons and their synapses, this review discusses the disrupted communication between these cells, notably involving fractalkine signaling and the triggering receptor expressed on myeloid cells (TREM). Overall, chronic stress emerges as a key player in cellular aging by altering the microglial sensome, notably via fractalkine signaling deficiency. To study cellular aging, novel positron emission tomography radiotracers for TREM and the purinergic family of receptors show interest for human study.
Introduction
Since their discovery, microglia have been described as the resident macrophages of the central nervous system (CNS). They migrate from the yolk sac to the brain around embryonic day 9.5 in mice (Ginhoux et al., 2010) and can be observed for the first time in the developing human brain around the 4.5–5th week of gestation (Andjelkovic et al., 1998; Monier et al., 2006; Verney et al., 2010). Following their migration and colonization, microglia remain distributed unevenly between the gray and white matters and across individual brain regions (Lawson et al., 1990; Mittelbronn et al., 2001). Indeed, Lawson et al. (1990) described the distribution of microglia in adult mice and found that these immune cells ranged from 5 to 12% of all brain cells. In humans, microglial distribution, which was discovered to be greater in the white matter, varied from 0.3 to 16.9% of all brain cells depending on the region (Mittelbronn et al., 2001). Microglial numbers are then maintained through local self-renewal in physiological conditions, a phenomenon conserved across species (Lawson et al., 1992; Askew et al., 2017; Füger et al., 2017; Réu et al., 2017; Tay et al., 2017).
Microglia are categorized most notably by their morphological state (e.g., surveillant, bushy or hyper-ramified, ameboid, dystrophic) and molecular signature (Martinez and Gordon, 2014; Ransohoff, 2016; Stratoulias et al., 2019). While previously termed as “quiescent” or “resting,” surveillant microglia are quite dynamic; their ramified processes retract and extend, constantly surveying the parenchyma for environmental cues (Davalos et al., 2005; Nimmerjahn et al., 2005). This allows them to interact with numerous cellular elements including astrocytic and neuronal cell bodies, synapses, but also with the basement membrane of the brain vasculature (Tremblay et al., 2010; Bisht et al., 2016; Stogsdill and Eroglu, 2017; Joost et al., 2019; Matejuk and Ransohoff, 2020; Vainchtein and Molofsky, 2020). Microglia contacting neuronal cell bodies are known as “satellite” microglia (Wogram et al., 2016; Stratoulias et al., 2019), a position recently associated with the regulation of neuronal activity (Cserép et al., 2020). Another interesting aspect of microglia is their ability to quickly alter morphologically to appropriately respond to the functional needs of the brain. Once microglia detect CNS insults through local stimuli, they undergo drastic morphological transformations. Their morphology can range from ameboid to hyper-ramified states (Davalos et al., 2005; Nimmerjahn et al., 2005; Wake et al., 2009), tightly following changes in their transcriptome and proteome (Reynolds et al., 2008; Dungrawala et al., 2010; Zhurinsky et al., 2010; Marguerat and Bähler, 2012; Rangaraju et al., 2018; Bell et al., 2019; Miedema et al., 2020; Rayaprolu et al., 2020). This microglial remodeling is observed in conditions of psychological stress, where among various brain regions that include the prefrontal cortex (PFC) and hippocampus, microglia were found to dynamically transform their morphology, gene and protein expression, as well as function (Bordeleau et al., 2020; Picard et al., 2021). These microglial changes to stress become further exacerbated with aging (Tay et al., 2017). Psychological stress is well-known to accelerate cellular aging (Yegorov et al., 2020), hence potentially predisposing to various neuropsychiatric disorders and neurodegenerative diseases across the lifespan (Musazzi and Marrocco, 2016; Justice, 2018; Desmarais et al., 2020; Smith and Pollak, 2020).
Microglia entertain a particular relationship with neurons. Developmental studies taught us that microglia notably influence neurogenesis by controlling the maturation, density and migration of neuronal progenitors during the first weeks of life (Paolicelli and Gross, 2011; Bordeleau et al., 2019). Acting specifically on neurons, microglia can influence their axonal growth (Pont-Lezica et al., 2014) and ability to form synapses by inducing the genesis of filopodia at dendritic elements (Miyamoto et al., 2016). Microglia are key players in synaptic plasticity, both structural and functional, which allows them to modify the neuronal circuitry via synaptic pruning, synaptic stripping, the secretion of neurotrophic factors and regulation of synaptic activity (Ji et al., 2013a,b; Sipe et al., 2016; Zhou et al., 2019), phenomena confirmed in the olfactory bulb, hippocampus, as well as cerebral cortex in mice (Tremblay and Majewska, 2011; Kettenmann et al., 2013; Morris et al., 2013; Schafer et al., 2013; Milior et al., 2016; Miyamoto et al., 2016). Microglia can modulate learning and memory, including auditory-cued fear conditioning and novel object recognition. This was for instance shown in mice depleted of microglia, which displayed altered performance in these tasks (Parkhurst et al., 2013; Torres et al., 2016). During early postnatal development, microglia are also able to shape the visual cortex of mice in an experience-dependent manner (Tremblay et al., 2010). By sensing synaptic activity, microglia remove weaker signaling synapses in the visual thalamus (Schafer et al., 2013). This is an important concept, as the best correlate for cognitive decline is synaptic loss (Scheff et al., 2006; Jackson et al., 2019; Colom-Cadena et al., 2020). Reduced synaptic density in the PFC is observed upon chronic psychological stress (Csabai et al., 2018), although some studies also show region-specific synaptic growth, for instance in the amygdala, in association with an emotional memory component (Picard et al., 2021). These various roles of microglia in the healthy brain were additionally shown to differ between sexes (Bordeleau et al., 2019).
In this review, we describe recent discoveries on the bidirectional communication between microglia, neurons, and their synapses, as well as discuss how their interactions are altered upon psychological stress and in major depressive disorder (MDD), leading to accelerated brain and cognitive aging. Studies on these emerging mechanisms in humans by positron emission tomography (PET) imaging are also briefly mentioned because of their potential to investigate MDD. Previous investigations using PET imaging have shown decreased metabolites in the limbic lobe and basal ganglia of MDD patients (Su et al., 2014). Further investigation tracking microglia-neuron receptor/ligand communication could offer monitoring opportunity for disease progression and treatment response in MDD patients.
Microglia-Neuron Communication and Interaction in Health, Stress, and Aging
The communication between microglia and neurons is bidirectional (Tremblay, 2011; Eyo and Wu, 2013; Szepesi et al., 2018), essential for homeostatic function and implicated in neurodegenerative disorders (Sheridan and Murphy, 2013). It is achieved via the complex microglial sensome constituting a myriad of various key receptors that constantly receive signals from surrounding neurons (Hickman et al., 2013) as well as their synapses (Roumier et al., 2004, 2008; Bessis et al., 2007; Béchade et al., 2013; Cantaut-Belarif et al., 2017; Tay et al., 2017; Comer et al., 2020). Several essential molecular mechanisms such as fractalkine signaling, the classical complement pathway, purinergic signaling, and triggering receptor expressed on myeloid cells 2 (TREM2) are discussed below. Diverse types of signaling can influence each other, acting as on and off systems (Biber et al., 2007).
Fractalkine Signaling
A main mediator of this communication is signaling between the neuronal chemokine fractalkine (CX3CL1) and its unique receptor (CX3CR1) expressed on the surface of microglia (Harrison et al., 1998; Ransohoff and Perry, 2009; Paolicelli et al., 2014; Lauro et al., 2019). During development, neurons expressing CX3CL1 (Meucci et al., 1998) modulate microglial pruning of synapses and brain functional connectivity, thus participating to proper brain maturation and social behavior in mice (Zhan et al., 2014; Arnoux and Audinat, 2015; Gunner et al., 2019). In adult mice, loss of fractalkine signaling causes widespread deficits in glutamate release at hippocampal synapses, which are associated with defects of adult hippocampal neurogenesis as well as learning and memory (Maggi et al., 2009; Rogers et al., 2011; Paolicelli et al., 2014; Zhan et al., 2014; Basilico et al., 2019), while psychological stress is associated with lower CX3CL1-CX3CR1 signaling between neurons and microglia (Han et al., 2019b). Throughout life, fractalkine signaling plays a part in the stress response, as CX3CR1 knockout in mice delays or prevents the response to chronic stress (Hulshof et al., 2003; Wohleb et al., 2013; Milior et al., 2016; Rimmerman et al., 2017; Winkler et al., 2017). In aging, fractalkine signaling is important for the regulation of adult neurogenesis in the hippocampus, but not the olfactory bulb (Gemma et al., 2010; Reshef et al., 2017; Bolós et al., 2018). However, hippocampal fractalkine signaling decreases steadily over the course of aging in mice, where its consequences still remain unknown (Mecca et al., 2018). Moreover, young adult (2-month old) mice with knockout for CX3CR1 were demonstrated to have a microglial transcriptome resembling that of aged mice based on its expression of inflammatory genes, suggesting a protective role of fractalkine signaling in aging (Gyoneva et al., 2019).
Classical Complement Pathway
Complement signaling is an essential part of the innate immune system bearing the role of opsonization, the secretion of molecules enhancing phagocytosis, contributing to overall pathogen removal (Veerhuis et al., 2011). Growing evidence places the complement protein 3 (C3) as essential for proper brain development (Lee et al., 2019), considering that C3 guides microglial synaptic pruning (Schafer et al., 2012; Luchena et al., 2018). In aging rhesus monkeys, the expression of C1q protein, an upstream complex of C3, is increased within synaptic elements of the PFC, signaling to microglia which synapses need to be pruned (Datta et al., 2020). Evidence for an implication of the complement pathway in stress-related disorders shows increased C3 expression in the PFC of mouse models of stress-induced depressive-like disorder (Crider et al., 2018). Distress paradigm in mice has been associated with hyper-ramified microglia and a loss of dendritic spines, paralleled by an increased expression of C1q component in the medial PFC and hippocampus (Smith et al., 2019). These observations are in agreement with an initially beneficial immune response, as shown by other mouse studies, where chronic stress was found to initially increase microglial survival through colony stimulating factor 1 (CSF1) signaling, which additionally promoted neuronal remodeling via the complement pathway (Kreisel et al., 2014; Wohleb et al., 2018; Horchar and Wohleb, 2019). Prolonged stress reduced microglial numbers, hence preventing effective neurogenesis, but this deficit was rescued by stimulating CSF1 signaling (Kreisel et al., 2014).
Purinergic Signaling
Purinergic receptors, many of which are highly expressed by microglia, can be separated into two families, i.e., sensors of adenosine (P1) counting A1, A2A and A3 or nucleotides (P2), which include the receptors P2Y12, P2Y6, P2Y4, P2X4, P2X7, and pannexin 1 (Burnstock, 2013; Illes et al., 2020; Sanchez-Arias et al., 2021). During development and injury, P2Y12 is a key adenosine triphosphate (ATP)-sensitive receptor responsible for determining microglial process motility, an important aspect involved in phagocytosis of both cellular debris or synaptic elements, notably in the mouse visual system (Haynes et al., 2006; Eyo et al., 2014; Sipe et al., 2016). This process was also characterized at the microglial filopodia level, showing that these structures perform a nanoscale surveillance of the mouse brain in homeostasis (Bernier et al., 2019). As microglia sense ATP to put themselves in motion, they also communicate with neurons via the secretion of adenosine binding to adenosine A1 receptor (A1R), which is able to act as negative feedback to suppress neuronal activity by limiting synaptic transmission (Badimon et al., 2020). During prolonged stress exposure, P2X7 receptor is up-regulated in hippocampal and PFC microglia, initiating an inflammatory response via the NOD-, LRR- and pyrin domain-containing protein 3 (NLRP3) inflammasome (Franklin et al., 2018; Dias et al., 2021). Assembly of the NLRP3 inflammasome via the P2X7 receptor was shown in the rat hippocampus to take place after 3 weeks of chronic stress (Yue et al., 2017). The implication of purinergic receptors in aging is still elusive, but cognitive decline-related disease conditions involving inflammation, such as Alzheimer’s disease (AD), show a consistent up-regulation of P2X7 in microglia (Woods et al., 2016; Francistiová et al., 2020; Pietrowski et al., 2021).
Triggering Receptor Expressed on Myeloid Cells 2 Signaling
TREM2 is a cell surface receptor for putative ligands that include lipids, DNA and pro-inflammatory proteins such as apolipoprotein E and amyloid (β) oligomers, which activate downstream signaling pathways for cell survival, phagocytosis, metabolic fitness and cell motility (Ulland et al., 2017; Konishi and Kiyama, 2018). While TREM2 is a major pathology-induced immune signaling pathway (Deczkowska et al., 2020), it is also required for synaptic pruning during normal development (Filipello et al., 2018). Cell survival-regulating Wnt/β-catenin pathways that are up-regulated after TREM2 binding, notably by one of its ligands Transmembrane Protein 59 (TMEM59), bear great importance for microglia. Knockout of the receptor impairs microglial response in AD, leading to an exacerbated sensibility to stress-induced cell death (Zheng et al., 2017; Liu et al., 2020; McQuade et al., 2020), with reports highlighting the emergence of susceptibility only upon TREM2 haploinsufficiency (Sayed et al., 2018). Later in life, TREM2 sustains a key signaling pathway for microglia, allowing their clearance of myelin debris (Kober and Brett, 2017). This role in myelin recycling is also key in AD, as mutant microglia for TREM2 overly produce autophagic vesicles via a deficit in the mammalian target for rapamycin (mTOR) pathway, leading to AD pathologies in the 5XFAD mouse model (Ulland et al., 2017). Furthermore, aged TREM2 knockout mouse microglia show reduction of phagocytosis and sudden increase in cellular oxidative stress (Linnartz-Gerlach et al., 2019). Overall, TREM2 seems to be major signaling pathway throughout life and shows a great pathological risk when altered in aging.
Tyro3, Axl, and Mer Receptors
Tyro3, Axl, and Mer (TAM) receptors are part of the cellular tyrosine kinase (TK) signaling pathways of the brain (Lemke, 2013). Microglial TAM receptors comprised of Axl and MerTK were shown to be required for their functions during normal physiological conditions as well as injury, specifically their process motility and phagocytosis of apoptotic newborn neurons (Fourgeaud et al., 2016). This is important as TAM receptors can promote hippocampal neurogenesis by preventing the secretion of inflammatory cytokines, as shown in vitro using a triple (Tyro3, Axl, MerTK) knockout model for these receptors with cultured microglia (Ji et al., 2013c). During aging, the Axl pathway is implicated in amyloid β clearance, hence preventing cellular toxicity in mice (Zhang et al., 2018) and with amyloid plaque-associated microglia expressing Axl and MerTK (Savage et al., 2015). However, the role of microglial TAM receptors during stress and aging remains elusive.
Microglial Metabolic and Morphological Changes in Stress and Aging
Immune dysregulation crucially impacts an individual’s ability to cope with internal and external challenges, such as trauma, infection, but also psychological stress, altering future response to stress outcomes in life and promoting cellular aging (Godbout et al., 2005; Buchanan et al., 2010; O’Donovan et al., 2012; Ritzel et al., 2019). A combination of several mechanisms including cellular senescence, deficits in energetic metabolism, release of inflammatory mediators, dysregulation of protein degradation pathways, activation of DNA repair mechanisms and gut dysbiosis, were suggested to be involved in cellular aging (Franceschi and Campisi, 2014; Franceschi et al., 2018). Upon exposure to immune challenges, stress or pathological aging, microglia are presumed to undergo a priming process (Püntener et al., 2012; Frank et al., 2018; Keane et al., 2021) leading to an elevated basal inflammatory activity [i.e., production of tumor necrosis factor alpha (TNFα), interleukin (IL)-1 beta (IL-1β), IL-6] (Sierra et al., 2007; Frank et al., 2010; Marschallinger et al., 2020), further increasing the overall inflammation levels in the brain (Franceschi and Campisi, 2014). Recent evidence showed that microglial priming can be explained by the trained immunity hypothesis, in which a challenge trains microglia to a next insult by promoting their secretion of intracellular metabolites and changing their epigenetic program (Haley et al., 2019; Neher and Cunningham, 2019). These findings are in line with increased gene expression within the interferon (IFN)-pathways observed in rodent models of aging (Grabert et al., 2016; Raj et al., 2017), and result in an inflammatory environment for microglia and neurons (Figure 1; Carnevale et al., 2007).
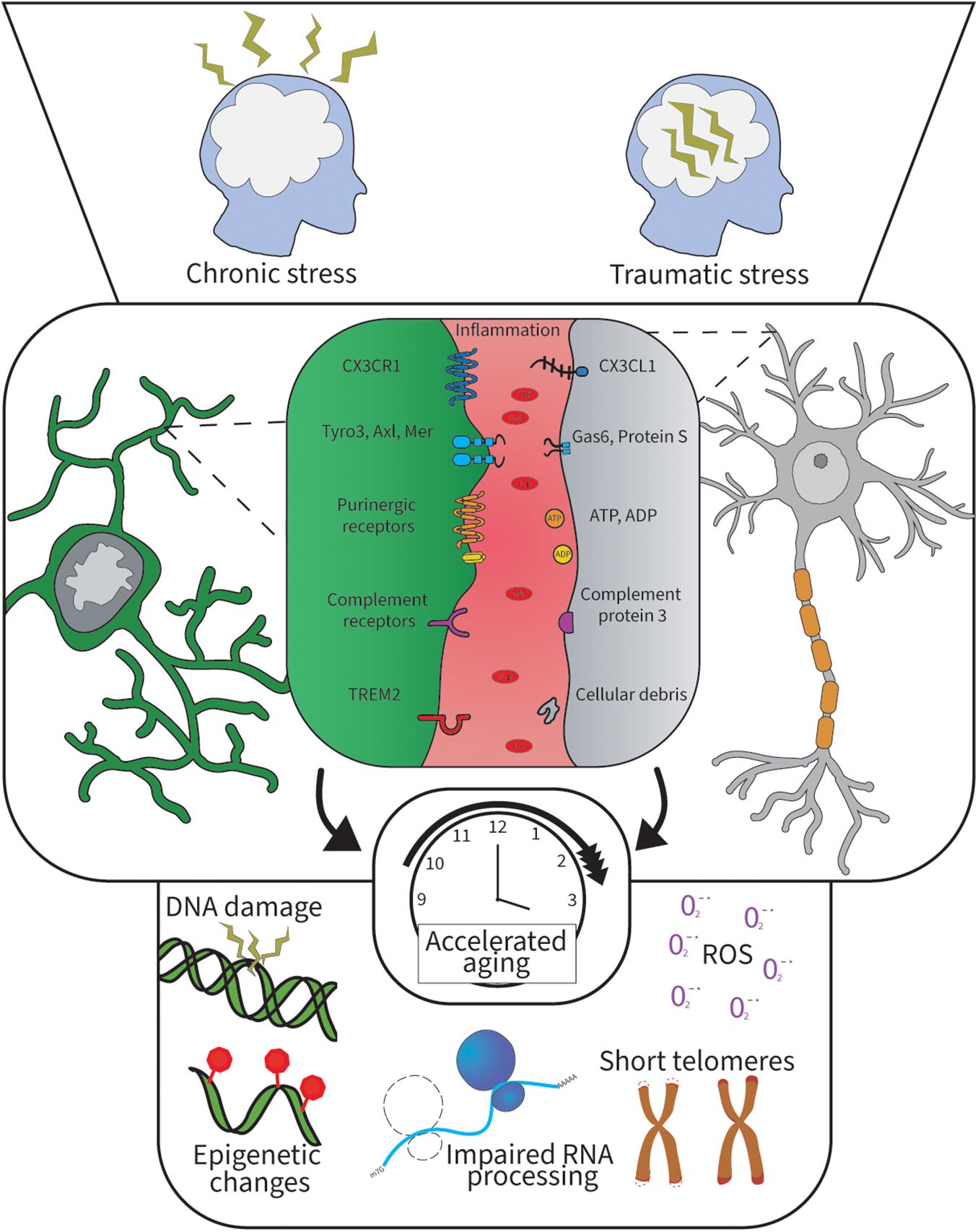
Figure 1. Psychological stress and inflammation alter microglia-neuron communication and lead to cellular aging. Psychological stress induces brain inflammation and oxidative stress resulting in cellular aging when prolonged. This hostile brain environment disrupts the microglia-neuron brain communication and leads to cognitive decline. The top panel illustrates two major types of stress lived by humans. The middle panel shows the different microglia-neuron signaling pathways in the brain and the key inflammatory factors identified in chronic stress. The bottom panel illustrates the different cellular aging processes happening when cells undergo cellular aging, which is often associated with inflammation and aging.
Microglial populations become more heterogeneous with aging as compared to the adult life stage (Olah et al., 2020; Delage et al., 2021). Concurrently, they display similarities with the early developmental period, a stage when microglia actively participate in the assembly and refinement of neural circuits (Schafer et al., 2012; Hammond et al., 2019). Parallel to an increase in the diversity of aging-related molecular signatures, aged microglia are morphologically altered when observed using light microscopy and tend to adopt more of an amoeboid-shaped phenotype. Alternatively, increased density of rod-shaped microglia (Bachstetter et al., 2017) or dystrophic microglia with reduced, gnarly processes have been described (Streit et al., 2004; Damani et al., 2011). These alterations may be of direct relevance to the functional irregularities reported in aged microglia. In this line of idea, transcriptomic investigation in human microglia reveals lower expression of CX3CR1 in aging (Galatro et al., 2017). Similar investigation in stressed microglia showed clustering of microglial population for genes related to inflammation and cytokine secretion (Lehmann et al., 2018), with the main ones illustrated in Figure 1. Furthermore, the overall microglial response to stimuli seems to be poorly managed with aging, suffering delays as well as prolonged duration, with potential detrimental consequences for the integrity of the aged CNS (Damani et al., 2011; Hefendehl et al., 2014; Jiang et al., 2020). Long-term sustenance of non-homeostatic microglial states in combination with morphological changes and loss of motility interfering with their optimal area surveillance (Damani et al., 2011; Hefendehl et al., 2014) may be contributing to an excessive accumulation of debris. Underlying mechanisms of the excessive debris accumulation may include increased lysosomal pH and compromised debris clearance notably through phagocytosis. This might be caused by their inability to protect and maintain the physiological function of neurons, synapses, myelin, etc. via trophic factors (Solé-Domènech et al., 2018; Pluvinage et al., 2019). Of note, the concept of senescence, a state when a cell enters division cycle arrest but preserves its metabolic activity and possesses a modified senescence-associated secretory phenotype (Coppé et al., 2010), has recently been a trending topic in the microglial field. Current evidence demonstrated that microglia can preserve their proliferative capacity even in aging as acutely extracted microglial cells from aged murine brains seem to lack up-regulation of senescence-associated markers, e.g., beta-galactosidase, as compared to long-term cultured microglia (Stojiljkovic et al., 2019; Hu et al., 2021). Previously, dystrophic morphological traits have been suggested for this phenotype (Streit et al., 2004). It is yet to be established whether senescent microglia are an independent microglial subtype of aged microglia or a state within the course of microglial aging.
Aged microglia possess elevated levels of mature lysosomal structures, undegradable lipofuscin granules and lipid bodies that are also larger in size (Sierra et al., 2007; Safaiyan et al., 2016; Marschallinger et al., 2020). Moreover, as was shown in a mouse model, aged microglia display reduced expression of the essential autophagy gene, Atg7 (Berglund et al., 2020), suggesting a limited intracellular maintenance. Metabolic failure occurs alongside impairments in the mitochondrial electron transport chain (ETC) during aging, identified by an altered/abnormal state of mitochondrial cristae ultrastructure, a marker of oxidative stress (Minhas et al., 2021). Upon exposure to challenges, microglia can undergo a switch from the efficient homeostatic oxidative metabolism (OXPHOS) to a faster ATP production via aerobic glycolysis, to sustain their elevated activity and increased coping strategies (Lauro and Limatola, 2020). In aging, this mechanism may become maladaptive, leading to the formation and accumulation of glycogen (a glucose storage), accompanied by a decreased mitochondrial respiration via the prostaglandin E2 (PGE2) and its receptor (EP2) pathway, as recently shown in mice (Minhas et al., 2021). Microglial proteomic analysis further suggested that this aging-related metabolic shift may relate to their utilization of alternate substrates, such as fatty acid, thus contributing to increased cholesterol trafficking with aging, which may result in neuronal dysfunction and cognitive decline (Flowers et al., 2017). Higher levels of neuronal cholesterol are considered detrimental, as myelinogenesis is slowed in aging, coincident with cytotoxic accumulation of cholesterol and cognitive decline (Kadish et al., 2009). On top of that, increase of intracellular iron, possibly related to an accumulation of lipofuscin, which is able to bind metals (Höhn et al., 2010), may be a contributing factor to aging-related oxidative imbalance and OXPHOS failure. As such, iron overload is considered one of the hallmarks of aged or senescent microglia (Angelova and Brown, 2019).
Several intracellular microglial properties were shown to be affected by aging (Cho et al., 2019). In agreement with observations made in other cell types, aged microglia show several signs of cellular aging such as epigenetic changes via the hypomethylation of IL-β associated with elevated Il-β transcript in microglia (Cho et al., 2015), which are described in Figure 1, along with an impaired DNA repair capability, altered transcription machinery and diminished ability of chromatin remodeling, as revealed by proteomic analysis of aged primary mouse microglia (Figure 1; Raj et al., 2014; Flowers et al., 2017; Costa et al., 2021). This inadequate microglial remodeling may underlie their impaired ability to dynamically change states according to the needs of the CNS during aging (Flowers et al., 2017). It is important to highlight that the telomere length is chemically affected by oxidative stress, notably via hydrogen peroxidase (H2O2), a reactive substrate that can cause prominent DNA damage (Oikawa and Kawanishi, 1999). Together, these cellular changes should be prevented or rescued to promote healthy aging (Depp et al., 2007; Wolkowitz et al., 2011), especially among key brain regions that are impacted by stress and aging.
Brain Hippocampus and Prefrontal Cortex: Key Regions Impacted by Stress and Aging
The Hippocampus
Hippocampal involvement in memory and learning has been widely studied across mouse, rat, non-human primate and human (Jarrard, 1993; Banta Lavenex and Lavenex, 2009; Voss et al., 2017; Zalucki et al., 2018). While still debated in human (Rodríguez-Iglesias et al., 2019), neurons in animal models are continuously produced throughout the lifespan among the olfactory bulb (Breton-Provencher et al., 2009) and hippocampus (Toda et al., 2019), while microglia also maintain their population by self-renewal (Tay et al., 2017). Several studies indicate that microglia may participate in the hippocampal-dependent learning and memory processes (Jarrard, 1993; Banta Lavenex and Lavenex, 2009; Parkhurst et al., 2013; Torres et al., 2016; Voss et al., 2017; Zalucki et al., 2018). For instance, microglial ablation via the CX3CR1-diphtheria toxin receptor reduces performance in object recognition test in rats (De Luca et al., 2020). The hippocampal CA1 makes projection to other brain regions also affected by stress, including the PFC (Rosene and Van Hoesen, 1977). This association seems to place the hippocampus as the gateway network particularly activated during learning tasks, as shown using magnetic resonance imaging (MRI) in human (Doyère et al., 1993; McEwen et al., 2016). It was also shown that stress resulting from sleep deprivation decreases hippocampal neurogenesis and induces depressive behavior (Murata et al., 2018; Han et al., 2019a). This contrasts with the ability of newborn neurons to contribute to stress resilience by inhibiting the hippocampal dentate gyrus mature granule cells activity as measured using electrophysiology (Anacker et al., 2018). The hippocampal functions are highly sensitive to accelerated aging, showing reduced cholinergic inputs and diminished neuronal activity rhythm in a mouse model of AD pathology (Rubio et al., 2012). Microglia regulate hippocampal neurogenesis in development (Pérez-Rodríguez et al., 2021) and throughout life (Reshef et al., 2017; Szepesi et al., 2018; Augusto-Oliveira et al., 2019; Bordeleau et al., 2019; Tan et al., 2020), in both health and neurodegeneration (De Lucia et al., 2016), via an IL-4 driven brain-derived neurotrophic factor (BDNF)-dependent mechanism (Zhang et al., 2021). Microglial influence on cognition could result from their roles in neurogenesis and at synapses. Indeed, these roles discussed in the section on fractalkine signaling can be performed notably via the secretion of inflammatory mediators and neurotrophic factors (Parkhurst et al., 2013; Sierra et al., 2014). Furthermore, microglia contribute to the control of synaptic activity and plasticity, constituting a quad-partite synapse alongside astrocytes, notably in the hippocampus (Bennett, 2007; Tremblay et al., 2011, 2014; Béchade et al., 2013; Schafer et al., 2013; Sierra and Tremblay, 2014).
The Prefrontal Cortex
The PFC is a key area affected by stress, with major impact on the behavior across species (Arnsten and Goldman-Rakic, 1998; Cerqueira et al., 2007; Holmes and Wellman, 2009; Myers-Schulz and Koenigs, 2012; Arnsten et al., 2015; McKlveen et al., 2015; Page et al., 2019). The stress response involves complex changes in neuron-microglia-astrocyte relationships, notably denoted by increased microglial phagocytosis of dendritic spines, as well as reduced astrocytic coverage areas in the PFC of rats exposed to chronic stress (Woodburn et al., 2021). This is in line with data showing an altered microglial morphology in rats after chronic stress, with females showing a greater proportion of primed microglia compared to males (Bollinger et al., 2016). Studies performed in aging are nevertheless lacking. Furthermore, PFC-dependent functions such as spatial working memory are affected by chronic stress in aged humans with MDD (Beats et al., 1996; Nebes et al., 2000) and in aged rats, and correlate with increase in staining intensity of ionized calcium binding adapter molecule 1 (Hinwood et al., 2012), a microglia/infiltrating macrophage marker (Imai et al., 1996). In a rat model of working memory deficiency, performance during the task was improved by administering minocycline (Hinwood et al., 2012), which normalizes microglial inflammatory and phagocytic functions (Mattei et al., 2014, 2017), potentially rescuing the homeostatic microglia-neuron crosstalk (Miao et al., 2018). As shown in the hippocampus during aging and in the PFC upon chronic stress, time is of the essence as studies show deleterious effects over time (Hanamsagar and Bilbo, 2017; Angelova and Brown, 2019), notably due to chronic inflammation acting in a negative feedback manner (Comer et al., 2020).
Microglial Involvement in Stress and Cognitive Decline Assessed in Human
In human, the cognitive abilities of aged or chronically stressed subjects can be assessed on multiple levels. One of them is cumulative knowledge (crystallized intelligence) including general knowledge with vocabulary and historical knowledge, the other being the cognitive processing (fluid intelligence) of information in order to complete a task (Paterniti et al., 2002; Wilson et al., 2013; Murman, 2015). Extensive work from Harada et al. (2013) highlighted deficits in processing speed when performing tasks, attention, memory and language with aging. These deficits were highly correlated with gray and white matter volume reductions revealed by MRI (Harada et al., 2013). A reduced executive control in aging can also be detected using the Wisconsin Card Sorting Task as an impairment often associated with function and volume diminution of the PFC and other frontal areas (Ballesteros et al., 2013). Patients with MDD experience deficits in attention, learning, as well as in a short-term and working memory (Lam et al., 2014). This is accompanied by evidence showing changes in white matter microstructures in adults with MDD including the cingulum and the hippocampus (van Velzen et al., 2020). There is then a clear link between cognitive deficit, stress and anatomical changes in the brain (Lam et al., 2014; van Velzen et al., 2020), but one might ask how this process occurs at the cellular level. Microglial activity can be measured with PET using the coupling of a radiotracer, most commonly [11C]PK11195 (Best et al., 2019) or [11C]PBR28 (Nettis et al., 2020), and its receptor Translocator Protein 18 kDa (TSPO) (Figure 2). TSPO has diverse cellular functions, and increases in its expression can signify impaired mitochondrial metabolism, increased oxidative stress, phagocytosis, and inflammation (Shoshan-Barmatz et al., 2019), particularly in glial cells (Pannell et al., 2020). Using this tracer, it was shown that elevated microglial PET activity parallels a decline in cognition with aging evaluated by cognitive processing (Parbo et al., 2017). Hence, the aging population is likely to suffer cognitive symptoms which can be triggered and/or escalated by chronic psychological stress and other environmental factors (Gonçalves de Andrade et al., 2021; Rizzo and Paolisso, 2021).
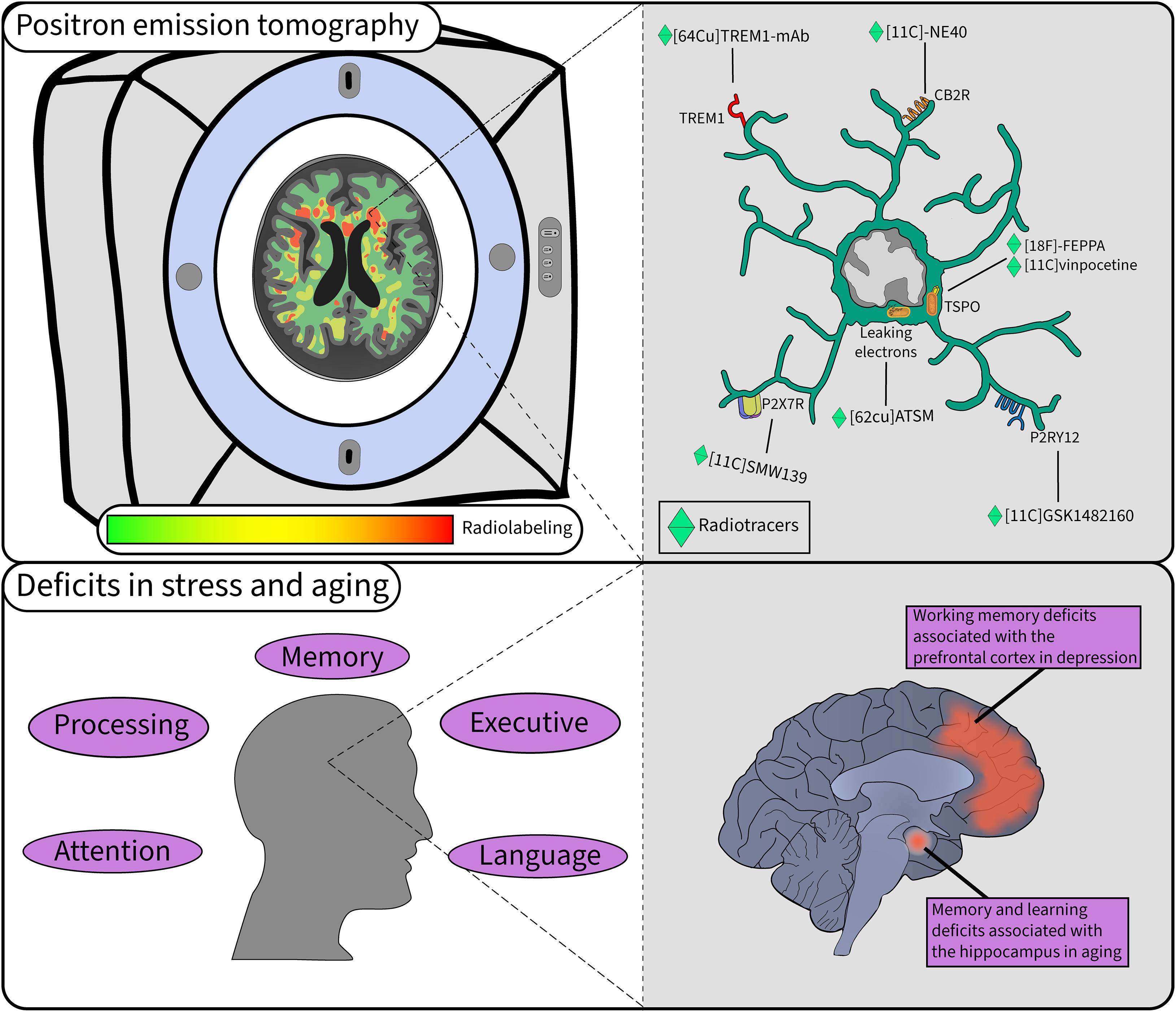
Figure 2. Microglial study using PET in stress and aging. PET is a widely used technique when investigating microglial activity in pathology. In the top left, microglial labeling of radiotracers is quantified to provide insight into their function in the human brain in a non-invasive manner. In the top right a myriad of tracers provide different information on the function of microglia in the brain. The bottom left panel highlights some of the cognitive deficits observed in aging that can be accelerated with chronic stress. The bottom right panel illustrates the key regions and deficits associated with stress and aging.
Measuring TSPO to infer glial activity is informative in various neurological diseases including AD, but also mild cognitive impairment (Hannestad et al., 2013; Holmes et al., 2018; Best et al., 2019; Kreisl et al., 2020; Meyer et al., 2020), and cognitive aging (Gulyás et al., 2011; Suridjan et al., 2014). Most studies have reported increased TSPO expression in patients with MDD compared to healthy controls, and higher levels were measured the longer MDD remained untreated (Meyer et al., 2020). PET studies on aging have reported mixed results (Repalli, 2014), with one using [18F]-FEPPA showing no increase in TSPO levels across the lifespan in healthy individuals (Suridjan et al., 2014), while another using [11C]vinpocetine found that TSPO expression increased linearly with age (Gulyás et al., 2011). A study by Ottoy et al. (2019) found that reduced 18F-fluorodeoxyglucose (18FDG) PET uptake, indicative of brain hypometabolism, was strongly associated with short-term cognitive decline and predicted a short-term transition to AD in mildly cognitively-impaired patients. Inconsistencies between studies may be in part due to limitations with TSPO radiotracers such as low signal-to-noise ratio or partial blood-brain barrier permeability (Kreisl et al., 2020). Moreover, limitations exist with the TSPO target itself, as it is not microglia-specific, being also expressed by astrocytes, endothelial cells and other myeloid cell types which can also be at play upon stress and in aging (Janssen et al., 2018; Carrier et al., 2020). Certain alternatives to TSPO, like the TREM and purinergic family of receptors, which are summarized in Figure 2, have gained interest in recent years to assess microglial function and their interactions with neurons in human (Janssen et al., 2018; Narayanaswami et al., 2018). Other than measuring mitochondrial activity, mitochondrial dysfunction can be imaged with 62Cu-ATSM [Cu-diacetyl-bis(N4-methylthiosemicarbzone)], which tags excessive electron buildup caused by leakage from malfunctioning ETC, suggesting an accumulation of reactive oxygen species (ROS) (Ikawa et al., 2020). ROS might be of particular interest as their cerebral levels are known to increase with stress and aging (Seo et al., 2012; Narayanaswami et al., 2018), limiting microglial ability to interact properly with neurons (Qin et al., 2002; Hur et al., 2010; Spencer et al., 2016; Munoz et al., 2017). Other investigations have developed and tested radiotracers targeting the myeloid cell marker CSF1R, able to track microglia in health and disease (Horti et al., 2019). However, there is more work required to expand the PET imaging radiotracer panel to understand human microglia and neuron crosstalk.
Discussion
Chronic stress was proposed to accelerate the process of cellular aging and promote sensitization or priming of microglia toward external stressors (Franklin et al., 2018; Rentscher et al., 2019). This over-sensitization by psychological stress is related to immunosenescence, i.e., the accumulation of oxidative stress causing deficit in responses revolving around the immune system (Gingrich, 2005; Deleidi et al., 2015) and preventing appropriate communication of microglia with the local environment, reported as hostile (Cornejo and von Bernhardi, 2016). The microglial sensome was identified as a key defective player during cognitive aging in the hippocampus and PFC (Kerr et al., 1991; Miller and O’Callaghan, 2005; Bauer, 2008; Bloss et al., 2010; Tay et al., 2018, 2019; Zannas et al., 2018). Figure 1 shows how psychological stress can alter the communication between microglia and neurons via inflammation leading to cellular aging of both neurons and microglia through multiple processes. In particular, aged microglia show morphological signs of dystrophy preventing their homeostatic function and often associated with neurodegenerative disease (Angelova and Brown, 2019; Savage et al., 2019; Swanson et al., 2020; Shahidehpour et al., 2021). Overall, microglia in pathological aging recruit different pathways compared to younger ages, with a predominant presence of inflammatory ones including TREM2, TAM and complement signaling (Flowers et al., 2017; Tay et al., 2017; Ulland et al., 2017; Pan et al., 2020; Keane et al., 2021). Microglial TREM2 has been particularly linked to aging, with 24 month old TREM2 knockout mice showing fewer and less phagocytic microglia based on a lower expression of CD68 in brain samples (Linnartz-Gerlach et al., 2019). Future studies are needed to investigate this change in TREM2 expression in humans experiencing chronic stress along the aging trajectory by PET imaging combined with a psychological cognitive assessment. Furthermore, the dynamics of purinergic receptors also require further investigation to determine if A1R is possibly affected in stress or aging, particularly if a disbalance in P2RY12/A1R may be at play.
There are also genetic predispositions known to accelerate aging by altering the microglial responses to stress. Genetic ablation of the protease cathepsin B (CatB) reduces oxidative stress in mice, thus improving cognitive performance with aging (Ni et al., 2019). Other types of stressors such as obesity-induced oxidative stress can accelerate the adverse effects of aging (Tarantini et al., 2018), resulting in increase of oxidative radicals in microglia which in turn contribute to the accelerating aging process (Raj et al., 2015). It is important to note that even if the role of microglia in oxidative stress is substantial, neurons in culture without glial cells can also show oxidative stress, meaning that glial cells are not solely responsible for brain cellular stress (Sompol et al., 2008; Hui et al., 2016). Indeed, neuronal inflammation is a major contributor to the Werner syndrome mouse model of premature aging, a neurological disorder associated with neuronal oxidative stress and microglial morphological changes (Hui et al., 2018). The overall outcome of impaired microglial aging may lead to impairments of synaptic plasticity, neurogenesis, accompanied by a loss of functions of other glial cells such as oligodendrocytes (Baror et al., 2019) and pathological neovascularization (Jiang et al., 2020). Oligodendrocytes are responsible for the CNS myelination and alteration of these cells contributes to memory deficits in aged mice (Wang et al., 2020). As these processes may play a role in aging-associated cognitive decline, targeting microglia presents an innovative direction of the therapeutic research. Potential therapies that could limit or rescue microglial aging, normalizing their key roles in learning and memory and the adaptation to environmental challenges, most notably include aerobic exercise and stress management in humans, also shown using wheel-running and outdoor living in mice (Tosato et al., 2007; Niraula et al., 2017; Yegorov et al., 2020). Dietary interventions like caloric restriction, in order to limit cellular cytotoxicity and promote anti-inflammatory activities, have also shown a great potential to promote healthy aging (Cheng et al., 2010). For instance, by inhibiting the mTOR pathway (López-Lluch and Navas, 2016) and thus potentially normalizing TREM2 signaling (Zhou et al., 2018), diet could lead to more efficient microglia when looking at the phagocytic ability and response ability to stress. Dietary intervention using ketogenic food made from soybean oil and cocoa butter is also beneficial in mice subjected to chronic stress (Guan et al., 2020; Morris et al., 2020). Similar dietary intervention is also demonstrating high potential to reducing mild cognitive impairment encountered in aging (Fortier et al., 2019, 2021). While we have presented preventive options, other curative therapies have shown interest. For instance, targeting survival signaling by blocking CSF1R was found to reduce Alzheimer’s-like pathology in mice (Olmos-Alonso et al., 2016). The monitoring of stress and aging in patients using PET imaging and psychological testing for cognitive decline seems like an interesting strategy to promote healthy aging, even more when paired with diet and healthy living.
Conclusion
This work aimed to focus on potential therapeutic targets and highlight preventive therapies that may be beneficial to limit the development of MDD and accelerated aging, as they are often less covered in the literature because of their long-term reward on the patient health. Using the communication pathways altered in stress and aging summarized in Figure 1, we can thus orient the development of new radiotracers and investigate pre-existing ones as shown in Figure 2, which would allow to monitor microglia-neuron communication in MDD. Some major regions of the brain are altered in stress and aging such as the hippocampus and the PFC, although other regions are also targeted like the insular lobe. Finally, this work proposes the long-term goal of preventing and identifying novel biomarkers for MDD and accelerated cognitive aging using translational research.
Author Contributions
MC was the principal manager of the review, wrote the discussion, section on human brain regions, and microglial-neuron communication, took care of the overall revision and formatting of the manuscript, and created the figures included in the manuscript. EŠ oversaw writing of the microglial metabolic changes in stress and aging section. CM oversaw redaction of the PET imaging section. M-KS-P was responsible for writing the introduction. M-ÈT oversaw the conceptual framework of the review while contributing significantly to the organization, design, and revision of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This research was supported by Natural Sciences and Engineering Research Council of Canada Discovery and Canadian Institute of Health Research Foundation grants awarded to M-ÈT. MC and M-KS-P were supported by doctoral training awards from Fonds de recherche du Québec–Santé. EŠ was a recipient of a graduate Branch Out Neurological Foundation scholarship. M-KS-P was also supported by a Canadian Institutes of Health Research doctorate scholarship. M-ÈT is a Canada Research Chair (Tier II) in Neurobiology of Aging and Cognition.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We are grateful to our colleagues for sharing their insights and contributing to our manuscript through their expertise and time, especially Ifeoluwa O. Awogbindin for his revision and proofing of the text.
References
Anacker, C., Luna, V. M., Stevens, G. S., Millette, A., Shores, R., Jimenez, J. C., et al. (2018). Hippocampal neurogenesis confers stress resilience by inhibiting the ventral dentate gyrus. Nature 559, 98–102. doi: 10.1038/s41586-018-0262-4
Andjelkovic, A. V., Nikolic, B., Pachter, J. S., and Zecevic, N. (1998). Macrophages/microglial cells in human central nervous system during development: an immunohistochemical study. Brain Res. 814, 13–25. doi: 10.1016/s0006-8993(98)00830-0
Angelova, D. M., and Brown, D. R. (2019). Microglia and the aging brain: are senescent microglia the key to neurodegeneration? J. Neurochem. 151, 676–688. doi: 10.1111/jnc.14860
Arnoux, I., and Audinat, E. (2015). Fractalkine signaling and microglia functions in the developing brain. Neural. Plast. 2015:689404. doi: 10.1155/2015/689404
Arnsten, A. F., and Goldman-Rakic, P. S. (1998). Noise stress impairs prefrontal cortical cognitive function in monkeys: evidence for a hyperdopaminergic mechanism. Arch. Gen. Psychiatry 55, 362–368. doi: 10.1001/archpsyc.55.4.362
Arnsten, A. F. T., Raskind, M. A., Taylor, F. B., and Connor, D. F. (2015). The effects of stress exposure on prefrontal cortex: translating basic research into successful treatments for post-traumatic stress disorder. Neurobiol. Stress 1, 89–99. doi: 10.1016/j.ynstr.2014.10.002
Askew, K., Li, K., Olmos-Alonso, A., Garcia-Moreno, F., Liang, Y., Richardson, P., et al. (2017). Coupled proliferation and apoptosis maintain the rapid turnover of microglia in the adult brain. Cell Rep. 18, 391–405. doi: 10.1016/j.celrep.2016.12.041
Augusto-Oliveira, M., Arrifano, G. P., Lopes-Araújo, A., Santos-Sacramento, L., Takeda, P. Y., Anthony, D. C., et al. (2019). What do microglia really do in healthy adult brain? Cells 8:1293. doi: 10.3390/cells8101293
Bachstetter, A. D., Ighodaro, E. T., Hassoun, Y., Aldeiri, D., Neltner, J. H., Patel, E., et al. (2017). Rod-shaped microglia morphology is associated with aging in 2 human autopsy series. Neurobiol. Aging 52, 98–105. doi: 10.1016/j.neurobiolaging.2016.12.028
Badimon, A., Strasburger, H. J., Ayata, P., Chen, X., Nair, A., Ikegami, A., et al. (2020). Negative feedback control of neuronal activity by microglia. Nature 586, 417–423. doi: 10.1038/s41586-020-2777-8
Ballesteros, S., Mayas, J., and Reales, J. M. (2013). Cognitive function in normal aging and in older adults with mild cognitive impairment. Psicothema 25, 18–24. doi: 10.7334/psicothema2012.181
Banta Lavenex, P., and Lavenex, P. (2009). Spatial memory and the monkey hippocampus: not all space is created equal. Hippocampus 19, 8–19. doi: 10.1002/hipo.20485
Baror, R., Neumann, B., Segel, M., Chalut, K. J., Fancy, S. P. J., Schafer, D. P., et al. (2019). Transforming growth factor-beta renders ageing microglia inhibitory to oligodendrocyte generation by CNS progenitors. Glia 67, 1374–1384. doi: 10.1002/glia.23612
Basilico, B., Pagani, F., Grimaldi, A., Cortese, B., Di Angelantonio, S., Weinhard, L., et al. (2019). Microglia shape presynaptic properties at developing glutamatergic synapses. Glia 67, 53–67. doi: 10.1002/glia.23508
Bauer, M. E. (2008). Chronic stress and immunosenescence: a review. Neuroimmunomodulation 15, 241–250. doi: 10.1159/000156467
Beats, B. C., Sahakian, B. J., and Levy, R. (1996). Cognitive performance in tests sensitive to frontal lobe dysfunction in the elderly depressed. Psychol. Med. 26, 591–603. doi: 10.1017/s0033291700035662
Béchade, C., Cantaut-Belarif, Y., and Bessis, A. (2013). Microglial control of neuronal activity. Front. Cell Neurosci. 7:32. doi: 10.3389/fncel.2013.00032
Bell, O. H., Copland, D. A., Ward, A., Nicholson, L. B., Lange, C. A. K., Chu, C. J., et al. (2019). Single Eye mRNA-Seq reveals normalisation of the retinal microglial transcriptome following acute inflammation. Front. Immunol. 10:3033. doi: 10.3389/fimmu.2019.03033
Bennett, M. R. (2007). Synaptic P2X7 receptor regenerative-loop hypothesis for depression. Aust. N Z J Psychiatry 41, 563–571. doi: 10.1080/00048670701399994
Berglund, R., Guerreiro-Cacais, A. O., Adzemovic, M. Z., Zeitelhofer, M., Lund, H., Ewing, E., et al. (2020). Microglial autophagy-associated phagocytosis is essential for recovery from neuroinflammation. Sci. Immunol. 5:5077. doi: 10.1126/sciimmunol.abb5077
Bernier, L.-P., Bohlen, C. J., York, E. M., Choi, H. B., Kamyabi, A., Dissing-Olesen, L., et al. (2019). Nanoscale surveillance of the brain by microglia via cAMP-Regulated Filopodia. Cell Rep. 27, 2895–2908. doi: 10.1016/j.celrep.2019.05.010
Bessis, A., Béchade, C., Bernard, D., and Roumier, A. (2007). Microglial control of neuronal death and synaptic properties. Glia 55, 233–238. doi: 10.1002/glia.20459
Best, L., Ghadery, C., Pavese, N., Tai, Y. F., and Strafella, A. P. (2019). New and Old TSPO PET radioligands for imaging brain microglial activation in neurodegenerative disease. Curr. Neurol. Neurosci. Rep. 19:24. doi: 10.1007/s11910-019-0934-y
Biber, K., Neumann, H., Inoue, K., and Boddeke, H. W. G. M. (2007). Neuronal “On” and “Off” signals control microglia. Trends Neurosci. 30, 596–602. doi: 10.1016/j.tins.2007.08.007
Bisht, K., Sharma, K. P., Lecours, C., Gabriela Sánchez, M., El Hajj, H., Milior, G., et al. (2016). Dark microglia: a new phenotype predominantly associated with pathological states. Glia 64, 826–839. doi: 10.1002/glia.22966
Bloss, E. B., Janssen, W. G., McEwen, B. S., and Morrison, J. H. (2010). Interactive effects of stress and aging on structural plasticity in the prefrontal cortex. J. Neurosci. 30, 6726–6731. doi: 10.1523/JNEUROSCI.0759-10.2010
Bollinger, J. L., Bergeon Burns, C. M., and Wellman, C. L. (2016). Differential effects of stress on microglial cell activation in male and female medial prefrontal cortex. Brain Behav. Immun. 52, 88–97. doi: 10.1016/j.bbi.2015.10.003
Bolós, M., Perea, J. R., Terreros-Roncal, J., Pallas-Bazarra, N., Jurado-Arjona, J., Ávila, J., et al. (2018). Absence of microglial CX3CR1 impairs the synaptic integration of adult-born hippocampal granule neurons. Brain Behav. Immun. 68, 76–89. doi: 10.1016/j.bbi.2017.10.002
Bordeleau, M., Carrier, M., Luheshi, G. N., and Tremblay, M. E. (2019). Microglia along sex lines: From brain colonization, maturation and function, to implication in neurodevelopmental disorders. Semin. Cell Dev. Biol. 94, 152–163. doi: 10.1016/j.semcdb.2019.06.001
Bordeleau, M., Lacabanne, C., Fernández, de Cossío, L., Vernoux, N., Savage, J. C., et al. (2020). Microglial and peripheral immune priming is partially sexually dimorphic in adolescent mouse offspring exposed to maternal high-fat diet. J. Neuroinflam. 17:264. doi: 10.1186/s12974-020-01914-1
Breton-Provencher, V., Lemasson, M., Peralta, M. R., and Saghatelyan, A. (2009). Interneurons produced in adulthood are required for the normal functioning of the olfactory bulb network and for the execution of selected olfactory behaviors. J. Neurosci. 29, 15245–15257. doi: 10.1523/JNEUROSCI.3606-09.2009
Buchanan, M. M., Hutchinson, M., Watkins, L. R., and Yin, H. (2010). Toll-like receptor 4 in CNS pathologies. J. Neurochem. 114, 13–27. doi: 10.1111/j.1471-4159.2010.06736.x
Burnstock, G. (2013). Introduction to purinergic signalling in the brain. Adv. Exp. Med. Biol. 986, 1–12. doi: 10.1007/978-94-007-4719-7_1
Cantaut-Belarif, Y., Antri, M., Pizzarelli, R., Colasse, S., Vaccari, I., Soares, S., et al. (2017). Microglia control the glycinergic but not the GABAergic synapses via prostaglandin E2 in the spinal cord. J. Cell Biol. 216, 2979–2989. doi: 10.1083/jcb.201607048
Carnevale, D., De Simone, R., and Minghetti, L. (2007). Microglia-neuron interaction in inflammatory and degenerative diseases: role of cholinergic and noradrenergic systems. CNS Neurol. Disord. Drug Targets 6, 388–397. doi: 10.2174/187152707783399193
Carrier, M., Robert, M. E., GonzalezIbanez, F., Desjardins, M., and Tremblay, M. E. (2020). Imaging the neuroimmune dynamics across space and time. Front. Neurosci. 14:903. doi: 10.3389/fnins.2020.00903
Cerqueira, J. J., Mailliet, F., Almeida, O. F. X., Jay, T. M., and Sousa, N. (2007). The prefrontal cortex as a key target of the maladaptive response to stress. J. Neurosci. 27, 2781–2787. doi: 10.1523/JNEUROSCI.4372-06.2007
Cheng, W.-H., Bohr, V. A., and de Cabo, R. (2010). Nutrition and aging. Mech. Ageing Dev. 131, 223–224. doi: 10.1016/j.mad.2010.03.011
Cho, C. E., Damle, S. S., Wancewicz, E. V., Mukhopadhyay, S., Hart, C. E., Mazur, C., et al. (2019). A modular analysis of microglia gene expression, insights into the aged phenotype. BMC Genomics 20:9. doi: 10.1186/s12864-019-5549-9
Cho, S.-H., Chen, J. A., Sayed, F., Ward, M. E., Gao, F., Nguyen, T. A., et al. (2015). SIRT1 deficiency in microglia contributes to cognitive decline in aging and neurodegeneration via epigenetic regulation of IL-1β. J. Neurosci. 35, 807–818. doi: 10.1523/JNEUROSCI.2939-14.2015
Colom-Cadena, M., Spires-Jones, T., Zetterberg, H., Blennow, K., Caggiano, A., DeKosky, S. T., et al. (2020). The clinical promise of biomarkers of synapse damage or loss in Alzheimer’s disease. Alzheimer’s Res. Ther. 12:21. doi: 10.1186/s13195-020-00588-4
Comer, A. L., Carrier, M., Tremblay, M. E., and Cruz-Martin, A. (2020). The inflamed brain in schizophrenia: the convergence of genetic and environmental risk factors that lead to uncontrolled neuroinflammation. Front. Cell Neurosci. 14:274. doi: 10.3389/fncel.2020.00274
Coppé, J.-P., Desprez, P.-Y., Krtolica, A., and Campisi, J. (2010). The senescence-associated secretory phenotype: the dark side of tumor suppression. Annu. Rev. Pathol. 5, 99–118. doi: 10.1146/annurev-pathol-121808-102144
Cornejo, F., and von Bernhardi, R. (2016). Age-Dependent Changes in the Activation and Regulation of Microglia in Glial Cells in Health and Disease of the CNS Advances in Experimental Medicine and Biology. New York, NY: Springer. doi: 10.1007/978-3-319-40764-7_10
Costa, J., Martins, S., Ferreira, P. A., Cardoso, A. M. S., Guedes, J. R., Peça, J., et al. (2021). The old guard: Age-related changes in microglia and their consequences. Mech. Ageing Dev. 197:111512. doi: 10.1016/j.mad.2021.111512
Crider, A., Feng, T., Pandya, C. D., Davis, T., Nair, A., Ahmed, A. O., et al. (2018). Complement component 3a receptor deficiency attenuates chronic stress-induced monocyte infiltration and depressive-like behavior. Brain Behav. Immun. 70, 246–256. doi: 10.1016/j.bbi.2018.03.004
Csabai, D., Wiborg, O., and Czéh, B. (2018). Reduced synapse and axon numbers in the prefrontal cortex of rats subjected to a chronic stress model for depression. Front. Cell. Neurosci. 12:24. doi: 10.3389/fncel.2018.00024
Cserép, C., Pósfai, B., Lénárt, N., Fekete, R., László, Z. I., Lele, Z., et al. (2020). Microglia monitor and protect neuronal function through specialized somatic purinergic junctions. Science 367, 528–537. doi: 10.1126/science.aax6752
Damani, M. R., Zhao, L., Fontainhas, A. M., Amaral, J., Fariss, R. N., and Wong, W. T. (2011). Age-related alterations in the dynamic behavior of microglia. Aging Cell 10, 263–276. doi: 10.1111/j.1474-9726.2010.00660.x
Datta, D., Leslie, S. N., Morozov, Y. M., Duque, A., Rakic, P., van Dyck, C. H., et al. (2020). Classical complement cascade initiating C1q protein within neurons in the aged rhesus macaque dorsolateral prefrontal cortex. J. Neuroinflam. 17:8. doi: 10.1186/s12974-019-1683-1
Davalos, D., Grutzendler, J., Yang, G., Kim, J. V., Zuo, Y., Jung, S., et al. (2005). ATP mediates rapid microglial response to local brain injury in vivo. Nat. Neurosci. 8, 752–758. doi: 10.1038/nn1472
De Luca, S. N., Soch, A., Sominsky, L., Nguyen, T.-X., Bosakhar, A., and Spencer, S. J. (2020). Glial remodeling enhances short-term memory performance in Wistar rats. J. Neuroinflam. 17:52. doi: 10.1186/s12974-020-1729-4
De Lucia, C., Rinchon, A., Olmos-Alonso, A., Riecken, K., Fehse, B., Boche, D., et al. (2016). Microglia regulate hippocampal neurogenesis during chronic neurodegeneration. Brain Behav. Immun. 55, 179–190. doi: 10.1016/j.bbi.2015.11.001
Deczkowska, A., Weiner, A., and Amit, I. (2020). The Physiology. Pathology, and Potential Therapeutic Applications of the TREM2 Signaling Pathway. Cell 181, 1207–1217. doi: 10.1016/j.cell.2020.05.003
Delage, C. I, Simoncicova, E., and Tremblay, M. E. (2021). Microglial heterogeneity in aging and Alzheimer’s disease: Is sex relevant? J. Pharmacol. Sci. 146, 169–181. doi: 10.1016/j.jphs.2021.03.006
Deleidi, M., Jäggle, M., and Rubino, G. (2015). Immune aging, dysmetabolism, and inflammation in neurological diseases. Front. Neurosci. 2015:172. doi: 10.3389/fnins.2015.00172
Depp, C. A., Glatt, S. J., and Jeste, D. V. (2007). Recent advances in research on successful or healthy aging. Curr. Psychiatry Rep. 9, 7–13. doi: 10.1007/s11920-007-0003-0
Desmarais, P., Weidman, D., Wassef, A., Bruneau, M.-A., Friedland, J., Bajsarowicz, P., et al. (2020). The interplay between post-traumatic stress disorder and dementia: a systematic review. Am. J. Geriatric. Psychiatry 28, 48–60. doi: 10.1016/j.jagp.2019.08.006
Dias, L., Lopes, C. R., Gonçalves, F. Q., Nunes, A., Pochmann, D., Machado, N. J., et al. (2021). Crosstalk Between ATP-P2X7 and Adenosine A2A receptors controlling neuroinflammation in rats subject to repeated restraint stress. Front. Cell Neurosci. 15:639322. doi: 10.3389/fncel.2021.639322
Doyère, V., Burette, F., Negro, C. R., and Laroche, S. (1993). Long-term potentiation of hippocampal afferents and efferents to prefrontal cortex: implications for associative learning. Neuropsychologia 31, 1031–1053. doi: 10.1016/0028-3932(93)90031-t
Dungrawala, H., Manukyan, A., and Schneider, B. L. (2010). Gene regulation: global transcription rates scale with size. Curr. Biol. 20, R979–R981. doi: 10.1016/j.cub.2010.09.064
Eyo, U. B., Peng, J., Swiatkowski, P., Mukherjee, A., Bispo, A., and Wu, L.-J. (2014). Neuronal hyperactivity recruits microglial processes via neuronal NMDA Receptors and Microglial P2Y12 Receptors after Status Epilepticus. J. Neurosci. 34, 10528–10540. doi: 10.1523/JNEUROSCI.0416-14.2014
Eyo, U. B., and Wu, L.-J. (2013). Bidirectional microglia-neuron communication in the healthy brain. Neural. Plast. 2013:456857. doi: 10.1155/2013/456857
Filipello, F., Morini, R., Corradini, I., Zerbi, V., Canzi, A., Michalski, B., et al. (2018). The Microglial Innate Immune Receptor TREM2 is required for synapse elimination and normal brain connectivity. Immunity 48, 979–991. doi: 10.1016/j.immuni.2018.04.016
Flowers, A., Bell-Temin, H., Jalloh, A., Stevens, S. M., and Bickford, P. C. (2017). Proteomic analysis of aged microglia: shifts in transcription, bioenergetics, and nutrient response. J. Neuroinflam. 14:96. doi: 10.1186/s12974-017-0840-7
Fortier, M., Castellano, C.-A., Croteau, E., Langlois, F., Bocti, C., St-Pierre, V., et al. (2019). A ketogenic drink improves brain energy and some measures of cognition in mild cognitive impairment. Alzheimers Dement. 15, 625–634. doi: 10.1016/j.jalz.2018.12.017
Fortier, M., Castellano, C.-A., St-Pierre, V., MyetteCote, E., Langlois, F., Roy, M., et al. (2021). A ketogenic drink improves cognition in mild cognitive impairment: Results of a 6-month RCT. Alzheimers Dement. 17, 543–552. doi: 10.1002/alz.12206
Fourgeaud, L., Través, P. G., Tufail, Y., Leal-Bailey, H., Lew, E. D., Burrola, P. G., et al. (2016). TAM receptors regulate multiple features of microglial physiology. Nature 532, 240–244. doi: 10.1038/nature17630
Franceschi, C., and Campisi, J. (2014). Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J. Gerontol. A Biol. Sci. Med. Sci. 69, S4–S9. doi: 10.1093/gerona/glu057
Franceschi, C., Garagnani, P., Parini, P., Giuliani, C., and Santoro, A. (2018). Inflammaging: a new immune-metabolic viewpoint for age-related diseases. Nat. Rev. Endocrinol. 14, 576–590. doi: 10.1038/s41574-018-0059-4
Francistiová, L., Bianchi, C., DiLauro, C., Sebastian-Serrano, A., deDiego-Garcia, L., Kobolak, J., et al. (2020). The Role of P2X7 Receptor in Alzheimer’s Disease. Front. Mol. Neurosci 13:94. doi: 10.3389/fnmol.2020.00094
Frank, M. G., Barrientos, R. M., Watkins, L. R., and Maier, S. F. (2010). Aging sensitizes rapidly isolated hippocampal microglia to LPS ex vivo. J. Neuroimmunol. 226, 181–184. doi: 10.1016/j.jneuroim.2010.05.022
Frank, M. G., Fonken, L. K., Annis, J. L., Watkins, L. R., and Maier, S. F. (2018). Stress disinhibits microglia via down-regulation of CD200R: A mechanism of neuroinflammatory priming. Brain Behav. Immun. 69, 62–73. doi: 10.1016/j.bbi.2017.11.001
Franklin, T. C., Wohleb, E. S., Zhang, Y., Fogaça, M., Hare, B., and Duman, R. S. (2018). Persistent increase in microglial RAGE contributes to chronic stress Induced priming of depressive-like behavior. Biol. Psychiatry 83, 50–60. doi: 10.1016/j.biopsych.2017.06.034
Füger, P., Hefendehl, J. K., Veeraraghavalu, K., Wendeln, A.-C., Schlosser, C., Obermüller, U., et al. (2017). Microglia turnover with aging and in an Alzheimer’s model via long-term in vivo single-cell imaging. Nat. Neurosci. 20, 1371–1376. doi: 10.1038/nn.4631
Galatro, T. F., Holtman, I. R., Lerario, A. M., Vainchtein, I. D., Brouwer, N., Sola, P. R., et al. (2017). Transcriptomic analysis of purified human cortical microglia reveals age-associated changes. Nat. Neurosci. 20, 1162–1171. doi: 10.1038/nn.4597
Gemma, C., Bachstetter, A. D., and Bickford, P. C. (2010). Neuron-Microglia Dialogue and Hippocampal Neurogenesis in the Aged Brain. Aging Dis. 1, 232–244.
Gingrich, J. A. (2005). Oxidative stress is the new stress. Nature Med. 11, 1281–1282. doi: 10.1038/nm1205-1281
Ginhoux, F., Greter, M., Leboeuf, M., Nandi, S., See, P., Gokhan, S., et al. (2010). Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science 330, 841–845. doi: 10.1126/science.1194637
Godbout, J. P., Chen, J., Abraham, J., Richwine, A. F., Berg, B. M., Kelley, K. W., et al. (2005). Exaggerated neuroinflammation and sickness behavior in aged mice following activation of the peripheral innate immune system. FASEB J. 19, 1329–1331. doi: 10.1096/fj.05-3776fje
GonçalvesdeAndrade, E., Simoncicova, E., Carrier, M., Vecchiarelli, H. A., Robert, M. E., and Tremblay, M. E. (2021). Microglia Fighting for Neurological and Mental Health: On the Central Nervous System Frontline of COVID-19 Pandemic. Front. Cell Neurosci. 15:647378. doi: 10.3389/fncel.2021.647378
Grabert, K., Michoel, T., Karavolos, M. H., Clohisey, S., Baillie, J. K., Stevens, M. P., et al. (2016). Microglial brain region-dependent diversity and selective regional sensitivities to aging. Nat. Neurosci. 19, 504–516. doi: 10.1038/nn.4222
Guan, Y.-F., Huang, G.-B., Xu, M.-D., Gao, F., Lin, S., Huang, J., et al. (2020). Anti-depression effects of ketogenic diet are mediated via the restoration of microglial activation and neuronal excitability in the lateral habenula. Brain Behav. Immun. 88, 748–762. doi: 10.1016/j.bbi.2020.05.032
Gulyás, B., Vas, A., Tóth, M., Takano, A., Varrone, A., Cselényi, Z., et al. (2011). Age and disease related changes in the translocator protein (TSPO) system in the human brain: positron emission tomography measurements with [11C]vinpocetine. Neuroimage 56, 1111–1121. doi: 10.1016/j.neuroimage.2011.02.020
Gunner, G., Cheadle, L., Johnson, K. M., Ayata, P., Badimon, A., Mondo, E., et al. (2019). Sensory lesioning induces microglial synapse elimination via ADAM10 and fractalkine signaling. Nat. Neurosci. 22, 1075–1088. doi: 10.1038/s41593-019-0419-y
Gyoneva, S., Hosur, R., Gosselin, D., Zhang, B., Ouyang, Z., Cotleur, A. C., et al. (2019). Cx3cr1-deficient microglia exhibit a premature aging transcriptome. Life Sci. Alliance 2:e201900453. doi: 10.26508/lsa.201900453
Haley, M. J., Brough, D., Quintin, J., and Allan, S. M. (2019). Microglial Priming as Trained Immunity in the Brain. Neuroscience 405, 47–54. doi: 10.1016/j.neuroscience.2017.12.039
Hammond, T. R., Dufort, C., Dissing-Olesen, L., Giera, S., Young, A., Wysoker, A., et al. (2019). Single-Cell RNA Sequencing of Microglia throughout the Mouse Lifespan and in the Injured Brain Reveals Complex Cell-State Changes. Immunity 50, 253–271. doi: 10.1016/j.immuni.2018.11.004
Han, X., Li, Q., Lan, X., El-Mufti, L., Ren, H., and Wang, J. (2019a). Microglial Depletion with Clodronate Liposomes Increases Proinflammatory Cytokine Levels. Induces Astrocyte Activation, and Damages Blood Vessel Integrity. Mol. Neurobiol. 56, 6184–6196. doi: 10.1007/s12035-019-1502-9
Han, Y., Zhang, L., Wang, Q., Zhang, D., Zhao, Q., Zhang, J., et al. (2019b). Minocycline inhibits microglial activation and alleviates depressive-like behaviors in male adolescent mice subjected to maternal separation. Psychoneuroendocrinology 107, 37–45. doi: 10.1016/j.psyneuen.2019.04.021
Hanamsagar, R., and Bilbo, S. D. (2017). Environment matters: microglia function and dysfunction in a changing world. Curr. Opin. Neurobiol. 47, 146–155. doi: 10.1016/j.conb.2017.10.007
Hannestad, J., DellaGioia, N., Gallezot, J.-D., Lim, K., Nabulsi, N., Esterlis, I., et al. (2013). The neuroinflammation marker translocator protein is not elevated in individuals with mild-to-moderate depression: a [11C]PBR28 PET study. Brain Behav. Immun. 33, 131–138. doi: 10.1016/j.bbi.2013.06.010
Harada, C. N., Natelson Love, M. C., and Triebel, K. (2013). Normal Cognitive Aging. Clin. Geriatr. Med. 29, 737–752. doi: 10.1016/j.cger.2013.07.002
Harrison, J. K., Jiang, Y., Chen, S., Xia, Y., Maciejewski, D., McNamara, R. K., et al. (1998). Role for neuronally derived fractalkine in mediating interactions between neurons and CX3CR1-expressing microglia. PNAS 95, 10896–10901.
Haynes, S. E., Hollopeter, G., Yang, G., Kurpius, D., Dailey, M. E., Gan, W.-B., et al. (2006). The P2Y12 receptor regulates microglial activation by extracellular nucleotides. Nat. Neurosci. 9, 1512–1519. doi: 10.1038/nn1805
Hefendehl, J. K., Neher, J. J., Sühs, R. B., Kohsaka, S., Skodras, A., and Jucker, M. (2014). Homeostatic and injury-induced microglia behavior in the aging brain. Aging Cell 13, 60–69. doi: 10.1111/acel.12149
Hickman, S. E., Kingery, N. D., Ohsumi, T. K., Borowsky, M. L., Wang, L., Means, T. K., et al. (2013). The microglial sensome revealed by direct RNA sequencing. Nat. Neurosci. 16, 1896–1905. doi: 10.1038/nn.3554
Hinwood, M., Morandini, J., Day, T. A., and Walker, F. R. (2012). Evidence that microglia mediate the neurobiological effects of chronic psychological stress on the medial prefrontal cortex. Cerebral. Cortex 22, 1442–1454. doi: 10.1093/cercor/bhr229
Höhn, A., Jung, T., Grimm, S., and Grune, T. (2010). Lipofuscin-bound iron is a major intracellular source of oxidants: role in senescent cells. Free Radic. Biol. Med. 48, 1100–1108. doi: 10.1016/j.freeradbiomed.2010.01.030
Holmes, A., and Wellman, C. L. (2009). Stress-induced prefrontal reorganization and executive dysfunction in rodents. Neurosci. Biobehav. Rev. 33, 773–783. doi: 10.1016/j.neubiorev.2008.11.005
Holmes, S. E., Hinz, R., Conen, S., Gregory, C. J., Matthews, J. C., Anton-Rodriguez, J. M., et al. (2018). Elevated translocator protein in anterior cingulate in major depression and a role for inflammation in suicidal thinking: a positron emission tomography study. Biol. Psychiatry 83, 61–69. doi: 10.1016/j.biopsych.2017.08.005
Horchar, M. J., and Wohleb, E. S. (2019). Glucocorticoid receptor antagonism prevents microglia-mediated neuronal remodeling and behavioral despair following chronic unpredictable stress. Brain Behav. Immun. 81, 329–340. doi: 10.1016/j.bbi.2019.06.030
Horti, A. G., Naik, R., Foss, C. A., Minn, I., Misheneva, V., Du, Y., et al. (2019). PET imaging of microglia by targeting macrophage colony-stimulating factor 1 receptor (CSF1R). Proc. Natl. Acad. Sci. USA 116, 1686–1691. doi: 10.1073/pnas.1812155116
Hu, Y., Fryatt, G. L., Ghorbani, M., Obst, J., Menassa, D. A., Martin-Estebane, M., et al. (2021). Replicative senescence dictates the emergence of disease-associated microglia and contributes to Aβ pathology. Cell Rep. 35:109228. doi: 10.1016/j.celrep.2021.109228
Hui, C. W., St-Pierre, M.-K., Detuncq, J., Aumailley, L., Dubois, M.-J., Couture, V., et al. (2018). Nonfunctional mutant Wrn protein leads to neurological deficits, neuronal stress, microglial alteration, and immune imbalance in a mouse model of Werner syndrome. Brain Behav. Immun. 73, 450–469. doi: 10.1016/j.bbi.2018.06.007
Hui, C. W., Zhang, Y., and Herrup, K. (2016). Non-Neuronal cells are required to mediate the effects of neuroinflammation: results from a neuron-enriched culture system. PLoS One 11:e0147134. doi: 10.1371/journal.pone.0147134
Hulshof, S., van Haastert, E. S., Kuipers, H. F., van den Elsen, P. J., De Groot, C. J., van der Valk, P., et al. (2003). CX3CL1 and CX3CR1 expression in human brain tissue: noninflammatory control versus multiple sclerosis. J. Neuropathol. Exp. Neurol. 62, 899–907. doi: 10.1093/jnen/62.9.899
Hur, J., Lee, P., Kim, M. J., Kim, Y., and Cho, Y.-W. (2010). Ischemia-activated microglia induces neuronal injury via activation of gp91phox NADPH oxidase. Biochem. Biophy. Res. Commun. 391, 1526–1530. doi: 10.1016/j.bbrc.2009.12.114
Ikawa, M., Okazawa, H., Nakamoto, Y., and Yoneda, M. (2020). PET imaging for oxidative stress in neurodegenerative disorders associated with mitochondrial dysfunction. Antioxidants 9:861. doi: 10.3390/antiox9090861
Illes, P., Rubini, P., Ulrich, H., Zhao, Y., and Tang, Y. (2020). Regulation of microglial functions by purinergic mechanisms in the healthy and diseased CNS. Cells 9:9051108. doi: 10.3390/cells9051108
Imai, Y., Ibata, I., Ito, D., Ohsawa, K., and Kohsaka, S. (1996). A novel gene iba1 in the major histocompatibility complex class III region encoding an EF hand protein expressed in a monocytic lineage. Biochem. Biophys. Res. Commun. 224, 855–862. doi: 10.1006/bbrc.1996.1112
Jackson, J., Jambrina, E., Li, J., Marston, H., Menzies, F., Phillips, K., et al. (2019). Targeting the Synapse in Alzheimer’s Disease. Front. Neurosci. 13:735. doi: 10.3389/fnins.2019.00735
Janssen, B., Vugts, D. J., Windhorst, A. D., and Mach, R. H. (2018). PET Imaging of Microglial Activation-Beyond Targeting TSPO. Molecules 23:23030607. doi: 10.3390/molecules23030607
Jarrard, L. E. (1993). On the role of the hippocampus in learning and memory in the rat. Behav. Neural Biol. 60, 9–26. doi: 10.1016/0163-1047(93)90664-4
Ji, K., Akgul, G., Wollmuth, L. P., and Tsirka, S. E. (2013a). Microglia actively regulate the number of functional synapses. PLoS One 8:e56293. doi: 10.1371/journal.pone.0056293
Ji, K., Miyauchi, J., and Tsirka, S. E. (2013b). Microglia: An Active Player in the Regulation of Synaptic Activity. Neural. Plasticity 2013:e627325. doi: 10.1155/2013/627325
Ji, R., Tian, S., Lu, H. J., Lu, Q., Zheng, Y., Wang, X., et al. (2013c). TAM receptors affect adult brain neurogenesis by negative regulation of microglial cell activation. J. Immunol. 191, 6165–6177. doi: 10.4049/jimmunol.1302229
Jiang, L., Mu, H., Xu, F., Xie, D., Su, W., Xu, J., et al. (2020). Transcriptomic and functional studies reveal undermined chemotactic and angiostimulatory properties of aged microglia during stroke recovery. J. Cereb. Blood Flow Metab. 40, S81–S97. doi: 10.1177/0271678X20902542
Joost, E., Jordão, M. J. C., Mages, B., Prinz, M., Bechmann, I., and Krueger, M. (2019). Microglia contribute to the glia limitans around arteries, capillaries and veins under physiological conditions, in a model of neuroinflammation and in human brain tissue. Brain Struct. Funct. 224, 1301–1314. doi: 10.1007/s00429-019-01834-8
Justice, N. J. (2018). The relationship between stress and Alzheimer’s disease. Neurobiol. Stress 8, 127–133. doi: 10.1016/j.ynstr.2018.04.002
Kadish, I., Thibault, O., Blalock, E. M., Chen, K.-C., Gant, J. C., Porter, N. M., et al. (2009). Hippocampal and cognitive aging across the lifespan: a bioenergetic shift precedes and increased cholesterol trafficking parallels memory impairment. J. Neurosci. 29, 1805–1816. doi: 10.1523/JNEUROSCI.4599-08.2009
Keane, L., Antignano, I., Riechers, S.-P., Zollinger, R., Dumas, A. A., Offermann, N., et al. (2021). mTOR-dependent translation amplifies microglia priming in aging mice. J. Clin. Invest. 131, 132727. doi: 10.1172/JCI132727
Kerr, D. S., Campbell, L. W., Applegate, M. D., Brodish, A., and Landfield, P. W. (1991). Chronic stress-induced acceleration of electrophysiologic and morphometric biomarkers of hippocampal aging. J. Neurosci. 11, 1316–1324.
Kettenmann, H., Kirchhoff, F., and Verkhratsky, A. (2013). Microglia: new roles for the synaptic stripper. Neuron 77, 10–18. doi: 10.1016/j.neuron.2012.12.023
Kober, D. L., and Brett, T. J. (2017). TREM2-ligand interactions in health and disease. J. Mol. Biol. 429, 1607–1629. doi: 10.1016/j.jmb.2017.04.004
Konishi, H., and Kiyama, H. (2018). Microglial TREM2/DAP12 Signaling: A Double-Edged Sword in Neural Diseases. Front. Cell Neurosci. 12:206. doi: 10.3389/fncel.2018.00206
Kreisel, T., Frank, M. G., Licht, T., Reshef, R., Ben-Menachem-Zidon, O., Baratta, M. V., et al. (2014). Dynamic microglial alterations underlie stress-induced depressive-like behavior and suppressed neurogenesis. Mol. Psychiatry 19, 699–709. doi: 10.1038/mp.2013.155
Kreisl, W. C., Kim, M.-J., Coughlin, J. M., Henter, I. D., Owen, D. R., and Innis, R. B. (2020). PET imaging of neuroinflammation in neurological disorders. Lancet. Neurol. 19, 940–950. doi: 10.1016/S1474-4422(20)30346-X
Lam, R. W., Kennedy, S. H., McIntyre, R. S., and Khullar, A. (2014). Cognitive Dysfunction in Major Depressive Disorder: Effects on Psychosocial Functioning and Implications for Treatment. Can. J. Psychiatry 59, 649–654.
Lauro, C., Chece, G., Monaco, L., Antonangeli, F., Peruzzi, G., Rinaldo, S., et al. (2019). Fractalkine Modulates Microglia Metabolism in Brain Ischemia. Front. Cell. Neurosci. 13:414. doi: 10.3389/fncel.2019.00414
Lauro, C., and Limatola, C. (2020). Metabolic Reprograming of Microglia in the Regulation of the Innate Inflammatory Response. Front. Immunol. 11:493. doi: 10.3389/fimmu.2020.00493
Lawson, L. J., Perry, V. H., Dri, P., and Gordon, S. (1990). Heterogeneity in the distribution and morphology of microglia in the normal adult mouse brain. Neuroscience 39, 151–170. doi: 10.1016/0306-4522(90)90229-w
Lawson, L. J., Perry, V. H., and Gordon, S. (1992). Turnover of resident microglia in the normal adult mouse brain. Neuroscience 48, 405–415. doi: 10.1016/0306-4522(92)90500-2
Lee, J. D., Coulthard, L. G., and Woodruff, T. M. (2019). Complement dysregulation in the central nervous system during development and disease. Semin. Immunol. 45:101340. doi: 10.1016/j.smim.2019.101340
Lehmann, M. L., Weigel, T. K., Cooper, H. A., Elkahloun, A. G., Kigar, S. L., and Herkenham, M. (2018). Decoding microglia responses to psychosocial stress reveals blood-brain barrier breakdown that may drive stress susceptibility. Sci. Rep. 8:11240. doi: 10.1038/s41598-018-28737-8
Lemke, G. (2013). Biology of the TAM receptors. Cold Spring Harb. Perspect Biol. 5:a009076. doi: 10.1101/cshperspect.a009076
Linnartz-Gerlach, B., Bodea, L.-G., Klaus, C., Ginolhac, A., Halder, R., Sinkkonen, L., et al. (2019). TREM2 triggers microglial density and age-related neuronal loss. Glia 67, 539–550. doi: 10.1002/glia.23563
Liu, Z., Ning, J., Zheng, X., Meng, J., Han, L., Zheng, H., et al. (2020). TMEM59 interacts with TREM2 and modulates TREM2-dependent microglial activities. Cell Death Dis. 11:678. doi: 10.1038/s41419-020-02874-3
López-Lluch, G., and Navas, P. (2016). Calorie restriction as an intervention in ageing. J. Physiol. 594, 2043–2060. doi: 10.1113/JP270543
Luchena, C., Zuazo-Ibarra, J., Alberdi, E., Matute, C., and Capetillo-Zarate, E. (2018). Contribution of neurons and glial cells to complement-mediated synapse removal during development, aging and in Alzheimer’s Disease. Med. Inflamm. 2018:414. doi: 10.1155/2018/2530414
Maggi, L., Trettel, F., Scianni, M., Bertollini, C., Eusebi, F., Fredholm, B. B., et al. (2009). LTP impairment by fractalkine/CX3CL1 in mouse hippocampus is mediated through the activity of adenosine receptor type 3 (A3R). J. Neuroimmunol. 215, 36–42. doi: 10.1016/j.jneuroim.2009.07.016
Marguerat, S., and Bähler, J. (2012). Coordinating genome expression with cell size. Trends Genet. 28, 560–565. doi: 10.1016/j.tig.2012.07.003
Marschallinger, J., Iram, T., Zardeneta, M., Lee, S. E., Lehallier, B., Haney, M. S., et al. (2020). Lipid-droplet-accumulating microglia represent a dysfunctional and proinflammatory state in the aging brain. Nat. Neurosci. 23, 194–208. doi: 10.1038/s41593-019-0566-1
Martinez, F. O., and Gordon, S. (2014). The M1 and M2 paradigm of macrophage activation: time for reassessment. F1000Prime Rep. 6:13. doi: 10.12703/P6-13
Matejuk, A., and Ransohoff, R. M. (2020). Crosstalk between astrocytes and microglia: an overview. Front. Immunol 11:1416. doi: 10.3389/fimmu.2020.01416
Mattei, D., Djodari-Irani, A., Hadar, R., Pelz, A., de Cossío, L. F., Goetz, T., et al. (2014). Minocycline rescues decrease in neurogenesis, increase in microglia cytokines and deficits in sensorimotor gating in an animal model of schizophrenia. Brain Behav. Immun. 38, 175–184. doi: 10.1016/j.bbi.2014.01.019
Mattei, D., Ivanov, A., Ferrai, C., Jordan, P., Guneykaya, D., Buonfiglioli, A., et al. (2017). Maternal immune activation results in complex microglial transcriptome signature in the adult offspring that is reversed by minocycline treatment. Transl. Psychiatry 7:e1120. doi: 10.1038/tp.2017.80
McEwen, B. S., Nasca, C., and Gray, J. D. (2016). Stress Effects on Neuronal Structure: Hippocampus. Amygdala, and Prefrontal Cortex. Neuropsychopharmacology 41, 3–23. doi: 10.1038/npp.2015.171
McKlveen, J. M., Myers, B., and Herman, J. P. (2015). The Medial Prefrontal Cortex: Coordinator of Autonomic. Neuroendocrine, and Behavioral Responses to Stress. J. Neuroendocrinol. 27, 446–456. doi: 10.1111/jne.12272
McQuade, A., Kang, Y. J., Hasselmann, J., Jairaman, A., Sotelo, A., Coburn, M., et al. (2020). Gene expression and functional deficits underlie TREM2-knockout microglia responses in human models of Alzheimer’s disease. Nat. Commun. 11:5370. doi: 10.1038/s41467-020-19227-5
Mecca, C., Giambanco, I., Donato, R., and Arcuri, C. (2018). Microglia and Aging: The Role of the TREM2–DAP12 and CX3CL1-CX3CR1 Axes. Int. J. Mol. Sci. 19:318. doi: 10.3390/ijms19010318
Meucci, O., Fatatis, A., Simen, A. A., Bushell, T. J., Gray, P. W., and Miller, R. J. (1998). Chemokines regulate hippocampal neuronal signaling and gp120 neurotoxicity. Proc. Natl. Acad. Sci. USA 95, 14500–14505. doi: 10.1073/pnas.95.24.14500
Meyer, J. H., Cervenka, S., Kim, M.-J., Kreisl, W. C., Henter, I. D., and Innis, R. B. (2020). Neuroinflammation in psychiatric disorders: PET imaging and promising new targets. Lancet. Psychiatry 7, 1064–1074. doi: 10.1016/S2215-0366(20)30255-8
Miao, H., Li, R., Han, C., Lu, X., and Zhang, H. (2018). Minocycline promotes posthemorrhagic neurogenesis via M2 microglia polarization via upregulation of the TrkB/BDNF pathway in rats. J. Neurophysiol. 120, 1307–1317. doi: 10.1152/jn.00234.2018
Miedema, A., Wijering, M. H. C., Eggen, B. J. L., and Kooistra, S. M. (2020). High-Resolution Transcriptomic and Proteomic Profiling of Heterogeneity of Brain-Derived Microglia in Multiple Sclerosis. Front. Mol. Neurosci. 13:583811. doi: 10.3389/fnmol.2020.583811
Milior, G., Lecours, C., Samson, L., Bisht, K., Poggini, S., Pagani, F., et al. (2016). Fractalkine receptor deficiency impairs microglial and neuronal responsiveness to chronic stress. Brain Behavior. Immun. 55, 114–125. doi: 10.1016/j.bbi.2015.07.024
Miller, D. B., and O’Callaghan, J. P. (2005). Aging, stress and the hippocampus. Ageing Res. Rev. 4, 123–140. doi: 10.1016/j.arr.2005.03.002
Minhas, P. S., Latif-Hernandez, A., McReynolds, M. R., Durairaj, A. S., Wang, Q., Rubin, A., et al. (2021). Restoring metabolism of myeloid cells reverses cognitive decline in ageing. Nature 590, 122–128. doi: 10.1038/s41586-020-03160-0
Mittelbronn, M., Dietz, K., Schluesener, H. J., and Meyermann, R. (2001). Local distribution of microglia in the normal adult human central nervous system differs by up to one order of magnitude. Acta Neuropathol. 101, 249–255. doi: 10.1007/s004010000284
Miyamoto, A., Wake, H., Ishikawa, A. W., Eto, K., Shibata, K., Murakoshi, H., et al. (2016). Microglia contact induces synapse formation in developing somatosensory cortex. Nat. Commun. 7:12540. doi: 10.1038/ncomms12540
Monier, A., Evrard, P., Gressens, P., and Verney, C. (2006). Distribution and differentiation of microglia in the human encephalon during the first two trimesters of gestation. J. Comp. Neurol. 499, 565–582. doi: 10.1002/cne.21123
Morris, G. P., Clark, I. A., Zinn, R., and Vissel, B. (2013). Microglia: A new frontier for synaptic plasticity, learning and memory, and neurodegenerative disease research. Neurobiol. Learn. Mem. 105, 40–53. doi: 10.1016/j.nlm.2013.07.002
Morris, G., Puri, B. K., Maes, M., Olive, L., Berk, M., and Carvalho, A. F. (2020). The role of microglia in neuroprogressive disorders: mechanisms and possible neurotherapeutic effects of induced ketosis. Prog. Neuro Psychopharmacol. Biol. Psychiatry 99:109858. doi: 10.1016/j.pnpbp.2020.109858
Munoz, F. M., Gao, R., Tian, Y., Henstenburg, B. A., Barrett, J. E., and Hu, H. (2017). Neuronal P2X7 receptor-induced reactive oxygen species production contributes to nociceptive behavior in mice. Sci. Rep. 7:3539. doi: 10.1038/s41598-017-03813-7
Murata, Y., Oka, A., Iseki, A., Mori, M., Ohe, K., Mine, K., et al. (2018). Prolonged sleep deprivation decreases cell proliferation and immature newborn neurons in both dorsal and ventral hippocampus of male rats. Neurosci. Res. 131, 45–51. doi: 10.1016/j.neures.2017.08.008
Murman, D. L. (2015). The impact of age on cognition. Semin. Hear 36, 111–121. doi: 10.1055/s-0035-1555115
Musazzi, L., and Marrocco, J. (2016). The many faces of stress: implications for neuropsychiatric Disorders. Neural. Plast. 2016:8389737. doi: 10.1155/2016/8389737
Myers-Schulz, B., and Koenigs, M. (2012). Functional anatomy of ventromedial prefrontal cortex: implications for mood and anxiety disorders. Mol. Psychiatry 17, 132–141. doi: 10.1038/mp.2011.88
Narayanaswami, V., Dahl, K., Bernard-Gauthier, V., Josephson, L., Cumming, P., and Vasdev, N. (2018). Emerging PET Radiotracers and Targets for Imaging of Neuroinflammation in Neurodegenerative Diseases: Outlook Beyond TSPO. Mol. Imag. 17:1536012118792317. doi: 10.1177/1536012118792317
Nebes, R. D., Butters, M. A., Mulsant, B. H., Pollock, B. G., Zmuda, M. D., Houck, P. R., et al. (2000). Decreased working memory and processing speed mediate cognitive impairment in geriatric depression. Psychol. Med. 30, 679–691. doi: 10.1017/s0033291799001968
Neher, J. J., and Cunningham, C. (2019). Priming Microglia for Innate Immune Memory in the Brain. Trends Immunol. 40, 358–374. doi: 10.1016/j.it.2019.02.001
Nettis, M. A., Veronese, M., Nikkheslat, N., Mariani, N., Lombardo, G., Sforzini, L., et al. (2020). PET imaging shows no changes in TSPO brain density after IFN-α immune challenge in healthy human volunteers. Transl. Psychiatry 10, 1–11. doi: 10.1038/s41398-020-0768-z
Ni, J., Wu, Z., Stoka, V., Meng, J., Hayashi, Y., Peters, C., et al. (2019). Increased expression and altered subcellular distribution of cathepsin B in microglia induce cognitive impairment through oxidative stress and inflammatory response in mice. Aging Cell 18:e12856. doi: 10.1111/acel.12856
Nimmerjahn, A., Kirchhoff, F., and Helmchen, F. (2005). Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science 308, 1314–1318. doi: 10.1126/science.1110647
Niraula, A., Sheridan, J. F., and Godbout, J. P. (2017). Microglia Priming with Aging and Stress. Neuropsychopharmacology 42, 318–333. doi: 10.1038/npp.2016.185
O’Donovan, A., Tomiyama, A. J., Lin, J., Puterman, E., Adler, N. E., Kemeny, M., et al. (2012). Stress appraisals and cellular aging: A key role for anticipatory threat in the relationship between psychological stress and telomere length. Brain Behav. Immun. 26, 573–579. doi: 10.1016/j.bbi.2012.01.007
Oikawa, S., and Kawanishi, S. (1999). Site-specific DNA damage at GGG sequence by oxidative stress may accelerate telomere shortening. FEBS Lett. 453, 365–368. doi: 10.1016/S0014-5793(99)00748-6
Olah, M., Menon, V., Habib, N., Taga, M. F., Ma, Y., Yung, C. J., et al. (2020). Single cell RNA sequencing of human microglia uncovers a subset associated with Alzheimer’s disease. Nat. Commun. 11:6129. doi: 10.1038/s41467-020-19737-2
Olmos-Alonso, A., Schetters, S. T. T., Sri, S., Askew, K., Mancuso, R., Vargas-Caballero, M., et al. (2016). Pharmacological targeting of CSF1R inhibits microglial proliferation and prevents the progression of Alzheimer’s-like pathology. Brain 139, 891–907. doi: 10.1093/brain/awv379
Ottoy, J., Niemantsverdriet, E., Verhaeghe, J., De Roeck, E., Struyfs, H., Somers, C., et al. (2019). Association of short-term cognitive decline and MCI-to-AD dementia conversion with CSF, MRI, amyloid- and 18F-FDG-PET imaging. Neuroimage Clin. 22:101771. doi: 10.1016/j.nicl.2019.101771
Page, C. E., Shepard, R., Heslin, K., and Coutellier, L. (2019). Prefrontal parvalbumin cells are sensitive to stress and mediate anxiety-related behaviors in female mice. Sci. Rep. 9:19772. doi: 10.1038/s41598-019-56424-9
Pan, J., Ma, N., Yu, B., Zhang, W., and Wan, J. (2020). Transcriptomic profiling of microglia and astrocytes throughout aging. J. Neuroinflam. 17:97. doi: 10.1186/s12974-020-01774-9
Pannell, M., Economopoulos, V., Wilson, T. C., Kersemans, V., Isenegger, P. G., Larkin, J. R., et al. (2020). Imaging of translocator protein upregulation is selective for pro-inflammatory polarized astrocytes and microglia. Glia 68, 280–297. doi: 10.1002/glia.23716
Paolicelli, R. C., Bisht, K., and Tremblay, M. E. (2014). Fractalkine regulation of microglial physiology and consequences on the brain and behavior. Front. Cell Neurosci. 8:129. doi: 10.3389/fncel.2014.00129
Paolicelli, R. C., and Gross, C. T. (2011). Microglia in development: linking brain wiring to brain environment. Neuron. Glia Biol. 7, 77–83. doi: 10.1017/S1740925X12000105
Parbo, P., Ismail, R., Hansen, K. V., Amidi, A., Mårup, F. H., Gottrup, H., et al. (2017). Brain inflammation accompanies amyloid in the majority of mild cognitive impairment cases due to Alzheimer’s disease. Brain 140, 2002–2011. doi: 10.1093/brain/awx120
Parkhurst, C. N., Yang, G., Ninan, I., Savas, J. N., Yates, J. R., Lafaille, J. J., et al. (2013). Microglia promote learning-dependent synapse formation through BDNF. Cell 155, 1596–1609. doi: 10.1016/j.cell.2013.11.030
Paterniti, S., Verdier-Taillefer, M.-H., Dufouil, C., and Alpérovitch, A. (2002). Depressive symptoms and cognitive decline in elderly people: Longitudinal study. Brit. J. Psychiatry 181, 406–410. doi: 10.1192/bjp.181.5.406
Pérez-Rodríguez, D. R., Blanco-Luquin, I., and Mendioroz, M. (2021). The Participation of Microglia in Neurogenesis: A Review. Brain Sci. 11:658. doi: 10.3390/brainsci11050658
Picard, K., St-Pierre, M.-K., Vecchiarelli, H. A., Bordeleau, M., and Tremblay, M. E. (2021). Neuroendocrine, neuroinflammatory and pathological outcomes of chronic stress: A story of microglial remodeling. Neurochem. Int. 145:104987. doi: 10.1016/j.neuint.2021.104987
Pietrowski, M. J., Gabr, A. A., Kozlov, S., Blum, D., Halle, A., and Carvalho, K. (2021). Glial Purinergic Signaling in Neurodegeneration. Front. Neurol. 12:654850. doi: 10.3389/fneur.2021.654850
Pluvinage, J. V., Haney, M. S., Smith, B. A. H., Sun, J., Iram, T., Bonanno, L., et al. (2019). CD22 blockade restores homeostatic microglial phagocytosis in ageing brains. Nature 568, 187–192. doi: 10.1038/s41586-019-1088-4
Pont-Lezica, L., Beumer, W., Colasse, S., Drexhage, H., Versnel, M., and Bessis, A. (2014). Microglia shape corpus callosum axon tract fasciculation: functional impact of prenatal inflammation. Eur. J. Neurosci. 39, 1551–1557. doi: 10.1111/ejn.12508
Püntener, U., Booth, S. G., Perry, V. H., and Teeling, J. L. (2012). Long-term impact of systemic bacterial infection on the cerebral vasculature and microglia. J. Neuroinflam. 9:146. doi: 10.1186/1742-2094-9-146
Qin, L., Liu, Y., Cooper, C., Liu, B., Wilson, B., and Hong, J.-S. (2002). Microglia enhance β-amyloid peptide-induced toxicity in cortical and mesencephalic neurons by producing reactive oxygen species. J. Neurochem. 83, 973–983. doi: 10.1046/j.1471-4159.2002.01210.x
Raj, D. D. A., Jaarsma, D., Holtman, I. R., Olah, M., Ferreira, F. M., Schaafsma, W., et al. (2014). Priming of microglia in a DNA-repair deficient model of accelerated aging. Neurobiol. Aging 35, 2147–2160. doi: 10.1016/j.neurobiolaging.2014.03.025
Raj, D. D. A., Moser, J., van der Pol, S. M. A., van Os, R. P., Holtman, I. R., Brouwer, N., et al. (2015). Enhanced microglial pro-inflammatory response to lipopolysaccharide correlates with brain infiltration and blood-brain barrier dysregulation in a mouse model of telomere shortening. Aging Cell 14, 1003–1013. doi: 10.1111/acel.12370
Raj, D., Yin, Z., Breur, M., Doorduin, J., Holtman, I. R., Olah, M., et al. (2017). Increased White Matter Inflammation in Aging- and Alzheimer’s Disease Brain. Front. Mol. Neurosci. 10:206. doi: 10.3389/fnmol.2017.00206
Rangaraju, S., Dammer, E. B., Raza, S. A., Rathakrishnan, P., Xiao, H., Gao, T., et al. (2018). Identification and therapeutic modulation of a pro-inflammatory subset of disease-associated-microglia in Alzheimer’s disease. Mol. Neurodegener. 13:24. doi: 10.1186/s13024-018-0254-8
Ransohoff, R. M. (2016). A polarizing question: do M1 and M2 microglia exist? Nat. Neurosci. 19, 987–991. doi: 10.1038/nn.4338
Ransohoff, R. M., and Perry, V. H. (2009). Microglial physiology: unique stimuli, specialized responses. Annu. Rev. Immunol. 27, 119–145. doi: 10.1146/annurev.immunol.021908.132528
Rayaprolu, S., Gao, T., Xiao, H., Ramesha, S., Weinstock, L. D., Shah, J., et al. (2020). Flow-cytometric microglial sorting coupled with quantitative proteomics identifies moesin as a highly-abundant microglial protein with relevance to Alzheimer’s disease. Mol. Neurodegener. 15:28. doi: 10.1186/s13024-020-00377-5
Rentscher, K. E., Carroll, J. E., Repetti, R. L., Cole, S. W., Reynolds, B. M., and Robles, T. F. (2019). Chronic stress exposure and daily stress appraisals relate to biological aging marker p16INK4a. Psychoneuroendocrinology 102, 139–148. doi: 10.1016/j.psyneuen.2018.12.006
Repalli, J. (2014). Translocator protein (TSPO) role in aging and Alzheimer’s disease. Curr. Aging Sci. 7, 168–175. doi: 10.2174/1874609808666141210103146
Reshef, R., Kudryavitskaya, E., Shani-Narkiss, H., Isaacson, B., Rimmerman, N., Mizrahi, A., et al. (2017). The role of microglia and their CX3CR1 signaling in adult neurogenesis in the olfactory bulb. Elife 6:30809. doi: 10.7554/eLife.30809
Réu, P., Khosravi, A., Bernard, S., Mold, J. E., Salehpour, M., Alkass, K., et al. (2017). The Lifespan and turnover of microglia in the human brain. Cell Rep. 20, 779–784. doi: 10.1016/j.celrep.2017.07.004
Reynolds, A. D., Glanzer, J. G., Kadiu, I., Ricardo-Dukelow, M., Chaudhuri, A., Ciborowski, P., et al. (2008). Nitrated alpha-synuclein-activated microglial profiling for Parkinson’s disease. J. Neurochem. 104, 1504–1525. doi: 10.1111/j.1471-4159.2007.05087.x
Rimmerman, N., Schottlender, N., Reshef, R., Dan-Goor, N., and Yirmiya, R. (2017). The hippocampal transcriptomic signature of stress resilience in mice with microglial fractalkine receptor (CX3CR1) deficiency. Brain Behav. Immun. 61, 184–196. doi: 10.1016/j.bbi.2016.11.023
Ritzel, R. M., Al Mamun, A., Crapser, J., Verma, R., Patel, A. R., Knight, B. E., et al. (2019). CD200-CD200R1 inhibitory signaling prevents spontaneous bacterial infection and promotes resolution of neuroinflammation and recovery after stroke. J. Neuroinflam. 16:40. doi: 10.1186/s12974-019-1426-3
Rizzo, M. R., and Paolisso, G. (2021). SARS-CoV-2 Emergency and Long-Term Cognitive Impairment in Older People. Aging Dis. 12, 345–352. doi: 10.14336/AD.2021.0109
Rodríguez-Iglesias, N., Sierra, A., and Valero, J. (2019). Rewiring of Memory Circuits: Connecting Adult Newborn Neurons With the Help of Microglia. Front. Cell Dev. Biol. 7:24. doi: 10.3389/fcell.2019.00024
Rogers, J. T., Morganti, J. M., Bachstetter, A. D., Hudson, C. E., Peters, M. M., Grimmig, B. A., et al. (2011). CX3CR1 deficiency leads to impairment of hippocampal cognitive function and synaptic plasticity. J. Neurosci. 31, 16241–16250. doi: 10.1523/JNEUROSCI.3667-11.2011
Rosene, D. L., and Van Hoesen, G. W. (1977). Hippocampal efferents reach widespread areas of cerebral cortex and amygdala in the rhesus monkey. Science 198, 315–317. doi: 10.1126/science.410102
Roumier, A., Béchade, C., Poncer, J.-C., Smalla, K.-H., Tomasello, E., Vivier, E., et al. (2004). Impaired synaptic function in the microglial KARAP/DAP12-deficient mouse. J. Neurosci. 24, 11421–11428. doi: 10.1523/JNEUROSCI.2251-04.2004
Roumier, A., Pascual, O., Béchade, C., Wakselman, S., Poncer, J.-C., Réal, E., et al. (2008). Prenatal activation of microglia induces delayed impairment of glutamatergic synaptic function. PLoS One 3:e2595. doi: 10.1371/journal.pone.0002595
Rubio, S. E., Vega-Flores, G., Martínez, A., Bosch, C., Pérez-Mediavilla, A., del Río, J., et al. (2012). Accelerated aging of the GABAergic septohippocampal pathway and decreased hippocampal rhythms in a mouse model of Alzheimer’s disease. FASEB J. 26, 4458–4467. doi: 10.1096/fj.12-208413
Safaiyan, S., Kannaiyan, N., Snaidero, N., Brioschi, S., Biber, K., Yona, S., et al. (2016). Age-related myelin degradation burdens the clearance function of microglia during aging. Nat. Neurosci. 19, 995–998. doi: 10.1038/nn.4325
Sanchez-Arias, J. C., van der Slagt, E., Vecchiarelli, H. A., Candlish, R. C., York, N., Young, P. A., et al. (2021). Purinergic signaling in nervous system health and disease: Focus on pannexin 1. Pharmacol. Ther. 225:107840. doi: 10.1016/j.pharmthera.2021.107840
Savage, J. C., Carrier, M., and Tremblay, M. E. (2019). Morphology of Microglia Across Contexts of Health and Disease. Methods Mol. Biol. 2034, 13–26. doi: 10.1007/978-1-4939-9658-2_2
Savage, J. C., Jay, T., Goduni, E., Quigley, C., Mariani, M. M., Malm, T., et al. (2015). Nuclear receptors license phagocytosis by trem2+ myeloid cells in mouse models of Alzheimer’s disease. J. Neurosci. 35, 6532–6543. doi: 10.1523/JNEUROSCI.4586-14.2015
Sayed, F. A., Telpoukhovskaia, M., Kodama, L., Li, Y., Zhou, Y., Le, D., et al. (2018). Differential effects of partial and complete loss of TREM2 on microglial injury response and tauopathy. Proc. Natl. Acad. Sci. USA 115, 10172–10177. doi: 10.1073/pnas.1811411115
Schafer, D. P., Lehrman, E. K., Kautzman, A. G., Koyama, R., Mardinly, A. R., Yamasaki, R., et al. (2012). Microglia Sculpt Postnatal Neural Circuits in an Activity and Complement-Dependent Manner. Neuron 74, 691–705. doi: 10.1016/j.neuron.2012.03.026
Schafer, D. P., Lehrman, E. K., and Stevens, B. (2013). The “quad-partite” synapse: microglia-synapse interactions in the developing and mature CNS. Glia 61, 24–36. doi: 10.1002/glia.22389
Scheff, S. W., Price, D. A., Schmitt, F. A., and Mufson, E. J. (2006). Hippocampal synaptic loss in early Alzheimer’s disease and mild cognitive impairment. Neurobiol. Aging 27, 1372–1384. doi: 10.1016/j.neurobiolaging.2005.09.012
Seo, J.-S., Park, J.-Y., Choi, J., Kim, T.-K., Shin, J.-H., Lee, J.-K., et al. (2012). NADPH oxidase mediates depressive behavior induced by chronic stress in mice. J. Neurosci. 32, 9690–9699. doi: 10.1523/JNEUROSCI.0794-12.2012
Shahidehpour, R. K., Higdon, R. E., Crawford, N. G., Neltner, J. H., Ighodaro, E. T., Patel, E., et al. (2021). Dystrophic microglia are associated with neurodegenerative disease and not healthy aging in the human brain. Neurobiol. Aging 99, 19–27. doi: 10.1016/j.neurobiolaging.2020.12.003
Sheridan, G. K., and Murphy, K. J. (2013). Neuron-glia crosstalk in health and disease: fractalkine and CX3CR1 take centre stage. Open Biol. 3:130181. doi: 10.1098/rsob.130181
Shoshan-Barmatz, V., Pittala, S., and Mizrachi, D. (2019). VDAC1 and the TSPO: Expression, Interactions, and Associated Functions in Health and Disease States. Int. J. Mol. Sci. 20:20133348. doi: 10.3390/ijms20133348
Sierra, A., Beccari, S., Diaz-Aparicio, I., Encinas, J. M., Comeau, S., and Tremblay, M. E. (2014). Surveillance, phagocytosis, and inflammation: how never-resting microglia influence adult hippocampal neurogenesis. Neural. Plast 2014:610343. doi: 10.1155/2014/610343
Sierra, A., Gottfried-Blackmore, A. C., McEwen, B. S., and Bulloch, K. (2007). Microglia derived from aging mice exhibit an altered inflammatory profile. Glia 55, 412–424. doi: 10.1002/glia.20468
Sierra, A., and Tremblay, M. E. (2014). Microglia in Health and Disease. (New York, NY: Springer), 3–6. doi: 10.1007/978-1-4939-1429-6_1
Sipe, G. O., Lowery, R. L., Tremblay, M. E., Kelly, E. A., Lamantia, C. E., and Majewska, A. K. (2016). Microglial P2Y12 is necessary for synaptic plasticity in mouse visual cortex. Nat. Commun. 7:10905. doi: 10.1038/ncomms10905
Smith, K. E., and Pollak, S. D. (2020). Early life stress and development: potential mechanisms for adverse outcomes. J. Neurodev. Disord. 12:34. doi: 10.1186/s11689-020-09337-y
Smith, K. L., Kassem, M. S., Clarke, D. J., Kuligowski, M. P., Bedoya-Pérez, M. A., Todd, S. M., et al. (2019). Microglial cell hyper-ramification and neuronal dendritic spine loss in the hippocampus and medial prefrontal cortex in a mouse model of PTSD. Brain Behav. Immun. 80, 889–899. doi: 10.1016/j.bbi.2019.05.042
Solé-Domènech, S., Rojas, A. V., Maisuradze, G. G., Scheraga, H. A., Lobel, P., and Maxfield, F. R. (2018). Lysosomal enzyme tripeptidyl peptidase 1 destabilizes fibrillar Aβ by multiple endoproteolytic cleavages within the β-sheet domain. Proc. Natl. Acad. Sci. USA 115, 1493–1498. doi: 10.1073/pnas.1719808115
Sompol, P., Ittarat, W., Tangpong, J., Chen, Y., Doubinskaia, I., Batinic-Haberle, I., et al. (2008). A neuronal model of Alzheimer’s disease: an insight into the mechanisms of oxidative stress-mediated mitochondrial injury. Neuroscience 153, 120–130. doi: 10.1016/j.neuroscience.2008.01.044
Spencer, N. G., Schilling, T., Miralles, F., and Eder, C. (2016). Mechanisms Underlying Interferon-γ-Induced Priming of Microglial Reactive Oxygen Species Production. PLoS One 11:e0162497. doi: 10.1371/journal.pone.0162497
Stogsdill, J. A., and Eroglu, C. (2017). The interplay between neurons and glia in synapse development and plasticity. Curr. Opin. Neurobiol. 42, 1–8. doi: 10.1016/j.conb.2016.09.016
Stojiljkovic, M. R., Ain, Q., Bondeva, T., Heller, R., Schmeer, C., and Witte, O. W. (2019). Phenotypic and functional differences between senescent and aged murine microglia. Neurobiol. Aging 74, 56–69. doi: 10.1016/j.neurobiolaging.2018.10.007
Stratoulias, V., Venero, J. L., Tremblay, M., and Joseph, B. (2019). Microglial subtypes: diversity within the microglial community. EMBO J. 38:1997. doi: 10.15252/embj.2019101997
Streit, W. J., Sammons, N. W., Kuhns, A. J., and Sparks, D. L. (2004). Dystrophic microglia in the aging human brain. Glia 45, 208–212. doi: 10.1002/glia.10319
Su, L., Cai, Y., Xu, Y., Dutt, A., Shi, S., and Bramon, E. (2014). Cerebral metabolism in major depressive disorder: a voxel-based meta-analysis of positron emission tomography studies. BMC Psychiatry 14:321. doi: 10.1186/s12888-014-0321-9
Suridjan, I., Rusjan, P. M., Voineskos, A. N., Selvanathan, T., Setiawan, E., Strafella, A. P., et al. (2014). Neuroinflammation in healthy aging: a PET study using a novel Translocator Protein 18kDa (TSPO) radioligand, [(18)F]-FEPPA. Neuroimage 84, 868–875. doi: 10.1016/j.neuroimage.2013.09.021
Swanson, M. E. V., Murray, H. C., Ryan, B., Faull, R. L. M., Dragunow, M., and Curtis, M. A. (2020). Quantitative immunohistochemical analysis of myeloid cell marker expression in human cortex captures microglia heterogeneity with anatomical context. Sci. Rep. 10:11693. doi: 10.1038/s41598-020-68086-z
Szepesi, Z., Manouchehrian, O., Bachiller, S., and Deierborg, T. (2018). Bidirectional Microglia-Neuron Communication in Health and Disease. Front. Cell Neurosci. 12:323. doi: 10.3389/fncel.2018.00323
Tan, Y.-L., Yuan, Y., and Tian, L. (2020). Microglial regional heterogeneity and its role in the brain. Mol. Psychiatry 25, 351–367. doi: 10.1038/s41380-019-0609-8
Tarantini, S., Valcarcel-Ares, M. N., Yabluchanskiy, A., Tucsek, Z., Hertelendy, P., Kiss, T., et al. (2018). Nrf2 Deficiency Exacerbates Obesity-Induced Oxidative Stress, Neurovascular Dysfunction, Blood-Brain Barrier Disruption, Neuroinflammation, Amyloidogenic Gene Expression, and Cognitive Decline in Mice, Mimicking the Aging Phenotype. J. Gerontol. A Biol. Sci. Med. Sci. 73, 853–863. doi: 10.1093/gerona/glx177
Tay, T. L., Béchade, C., D’Andrea, I., St-Pierre, M.-K., Henry, M. S., Roumier, A., et al. (2018). Microglia Gone Rogue: Impacts on Psychiatric Disorders across the Lifespan. Front. Mol. Neurosci. 10:421. doi: 10.3389/fnmol.2017.00421
Tay, T. L., Carrier, M., and Tremblay, M. E. (2019). Physiology of Microglia. Adv. Exp. Med. Biol. 1175, 129–148. doi: 10.1007/978-981-13-9913-8_6
Tay, T. L., Savage, J. C., Hui, C. W., Bisht, K., and Tremblay, M. E. (2017). Microglia across the lifespan: from origin to function in brain development, plasticity and cognition. J. Physiol. 595, 1929–1945. doi: 10.1113/JP272134
Toda, T., Parylak, S. L., Linker, S. B., and Gage, F. H. (2019). The role of adult hippocampal neurogenesis in brain health and disease. Mol. Psychiatry 24, 67–87. doi: 10.1038/s41380-018-0036-2
Torres, L., Danver, J., Ji, K., Miyauchi, J. T., Chen, D., Anderson, M. E., et al. (2016). Dynamic microglial modulation of spatial learning and social behavior. Brain Behav. Immun. 55, 6–16. doi: 10.1016/j.bbi.2015.09.001
Tosato, M., Zamboni, V., Ferrini, A., and Cesari, M. (2007). The aging process and potential interventions to extend life expectancy. Clin. Interv. Aging 2, 401–412.
Tremblay, M. E. (2011). The role of microglia at synapses in the healthy CNS: novel insights from recent imaging studies. Neuron. Glia Biol. 7, 67–76. doi: 10.1017/S1740925X12000038
Tremblay, M. E., Lowery, R. L., and Majewska, A. K. (2010). Microglial Interactions with Synapses Are Modulated by Visual Experience. PLoS Biol. 8:e1000527. doi: 10.1371/journal.pbio.1000527
Tremblay, M. E., and Majewska, A. K. (2011). A role for microglia in synaptic plasticity? Commun. Integr. Biol. 4, 220–222. doi: 10.4161/cib.4.2.14506
Tremblay, M. E., Paolicelli, R. C., Stevens, B., Wake, H., and Bessis, A. (2014). “Developing and Mature Synapses,” in Microglia in Health and Disease, eds M. E. Tremblay and A. Sierra (New York, NY: Springer), 223–248. doi: 10.1007/978-1-4939-1429-6_9
Tremblay, M. E., Stevens, B., Sierra, A., Wake, H., Bessis, A., and Nimmerjahn, A. (2011). The role of microglia in the healthy brain. J. Neurosci. 31, 16064–16069. doi: 10.1523/JNEUROSCI.4158-11.2011
Ulland, T. K., Song, W. M., Huang, S. C.-C., Ulrich, J. D., Sergushichev, A., Beatty, W. L., et al. (2017). TREM2 Maintains Microglial Metabolic Fitness in Alzheimer’s Disease. Cell 170, 649–663. doi: 10.1016/j.cell.2017.07.023
Vainchtein, I. D., and Molofsky, A. V. (2020). Astrocytes and Microglia: In Sickness and in Health. Trends Neurosci. 43, 144–154. doi: 10.1016/j.tins.2020.01.003
van Velzen, L. S., Kelly, S., Isaev, D., Aleman, A., Aftanas, L. I., Bauer, J., et al. (2020). White matter disturbances in major depressive disorder: a coordinated analysis across 20 international cohorts in the ENIGMA MDD working group. Mol. Psychiatry 25, 1511–1525. doi: 10.1038/s41380-019-0477-2
Veerhuis, R., Nielsen, H. M., and Tenner, A. J. (2011). Complement in the brain. Mol. Immunol. 48, 1592–1603. doi: 10.1016/j.molimm.2011.04.003
Verney, C., Monier, A., Fallet-Bianco, C., and Gressens, P. (2010). Early microglial colonization of the human forebrain and possible involvement in periventricular white-matter injury of preterm infants. J. Anat. 217, 436–448. doi: 10.1111/j.1469-7580.2010.01245.x
Voss, J. L., Bridge, D. J., Cohen, N. J., and Walker, J. A. (2017). A closer look at the hippocampus and memory. Trends Cogn. Sci. 21, 577–588. doi: 10.1016/j.tics.2017.05.008
Wake, H., Moorhouse, A. J., Jinno, S., Kohsaka, S., and Nabekura, J. (2009). Resting Microglia Directly Monitor the Functional State of Synapses In Vivo and Determine the Fate of Ischemic Terminals. J. Neurosci. 29, 3974–3980. doi: 10.1523/JNEUROSCI.4363-08.2009
Wang, F., Ren, S.-Y., Chen, J.-F., Liu, K., Li, R.-X., Li, Z.-F., et al. (2020). Myelin degeneration and diminished myelin renewal contribute to age-related deficits in memory. Nat. Neurosci. 23, 481–486. doi: 10.1038/s41593-020-0588-8
Wilson, C. B., McLaughlin, L. D., Nair, A., Ebenezer, P. J., Dange, R., and Francis, J. (2013). Inflammation and oxidative stress are elevated in the brain, blood, and adrenal glands during the progression of post-traumatic stress disorder in a predator exposure animal model. PLoS One 8:e76146. doi: 10.1371/journal.pone.0076146
Winkler, Z., Kuti, D., Ferenczi, S., Gulyas, K., Polyak, A., and Kovacs, K. J. (2017). Impaired microglia fractalkine signaling affects stress reaction and coping style in mice. Behav. Brain Res. 334, 119–128. doi: 10.1016/j.bbr.2017.07.023
Wogram, E., Wendt, S., Matyash, M., Pivneva, T., Draguhn, A., and Kettenmann, H. (2016). Satellite microglia show spontaneous electrical activity that is uncorrelated with activity of the attached neuron. Eur. J. Neurosci. 43, 1523–1534. doi: 10.1111/ejn.13256
Wohleb, E. S., Powell, N. D., Godbout, J. P., and Sheridan, J. F. (2013). Stress-induced recruitment of bone marrow-derived monocytes to the brain promotes anxiety-like behavior. J. Neurosci. 33, 13820–13833. doi: 10.1523/JNEUROSCI.1671-13.2013
Wohleb, E. S., Terwilliger, R., Duman, C. H., and Duman, R. S. (2018). Stress-induced neuronal colony stimulating factor 1 provokes microglia-mediated neuronal remodeling and depressive-like behavior. Biol. Psychiatry 83, 38–49. doi: 10.1016/j.biopsych.2017.05.026
Wolkowitz, O. M., Mellon, S. H., Epel, E. S., Lin, J., Dhabhar, F. S., Su, Y., et al. (2011). Leukocyte telomere length in major depression: correlations with chronicity. Inflammation and Oxidative Stress - Preliminary Findings. PLoS One 6:e17837. doi: 10.1371/journal.pone.0017837
Woodburn, S. C., Bollinger, J. L., and Wohleb, E. S. (2021). Synaptic and behavioral effects of chronic stress are linked to dynamic and sex-specific changes in microglia function and astrocyte dystrophy. Neurobiol. Stress 14:100312. doi: 10.1016/j.ynstr.2021.100312
Woods, L. T., Ajit, D., Camden, J. M., Erb, L., and Weisman, G. A. (2016). Purinergic Receptors as Potential Therapeutic Targets in Alzheimer’s Disease. Neuropharmacology 104, 169–179. doi: 10.1016/j.neuropharm.2015.10.031
Yegorov, Y. E., Poznyak, A. V., Nikiforov, N. G., Sobenin, I. A., and Orekhov, A. N. (2020). The Link between Chronic Stress and Accelerated Aging. Biomedicines 8:198. doi: 10.3390/biomedicines8070198
Yue, N., Huang, H., Zhu, X., Han, Q., Wang, Y., Li, B., et al. (2017). Activation of P2X7 receptor and NLRP3 inflammasome assembly in hippocampal glial cells mediates chronic stress-induced depressive-like behaviors. J. Neuroinflam. 14:102. doi: 10.1186/s12974-017-0865-y
Zalucki, O., Harkins, D., Harris, L., Burne, T. H. J., Gronostajski, R. M., and Piper, M. (2018). Analysis of hippocampal-dependent learning and memory behaviour in mice lacking Nfix from adult neural stem cells. BMC Res. Notes 11:564. doi: 10.1186/s13104-018-3652-7
Zannas, A. S., Arloth, J., Carrillo-Roa, T., Iurato, S., Röh, S., Ressler, K. J., et al. (2018). Correction to: Lifetime stress accelerates epigenetic aging in an urban. African American cohort: relevance of glucocorticoid signaling. Genome Biol. 19:61. doi: 10.1186/s13059-018-1441-1
Zhan, Y., Paolicelli, R. C., Sforazzini, F., Weinhard, L., Bolasco, G., Pagani, F., et al. (2014). Deficient neuron-microglia signaling results in impaired functional brain connectivity and social behavior. Nat. Neurosci. 17, 400–406. doi: 10.1038/nn.3641
Zhang, J., Culp, M. L., Craver, J. G., and Darley-Usmar, V. (2018). Mitochondrial function and autophagy: integrating proteotoxic, redox, and metabolic stress in Parkinson’s disease. J. Neurochem. 2018:14308. doi: 10.1111/jnc.14308
Zhang, J., Rong, P., Zhang, L., He, H., Zhou, T., Fan, Y., et al. (2021). IL4-driven microglia modulate stress resilience through BDNF-dependent neurogenesis. Sci. Adv. 7:eabb9888. doi: 10.1126/sciadv.abb9888
Zheng, H., Jia, L., Liu, C.-C., Rong, Z., Zhong, L., Yang, L., et al. (2017). TREM2 Promotes Microglial Survival by Activating Wnt/β-Catenin Pathway. J. Neurosci. 37, 1772–1784. doi: 10.1523/JNEUROSCI.2459-16.2017
Zhou, L.-J., Peng, J., Xu, Y.-N., Zeng, W.-J., Zhang, J., Wei, X., et al. (2019). Microglia are indispensable for synaptic plasticity in the spinal dorsal horn and chronic pain. Cell Rep. 27, 3844–3859. doi: 10.1016/j.celrep.2019.05.087
Zhou, Y., Ulland, T. K., and Colonna, M. (2018). TREM2-Dependent Effects on Microglia in Alzheimer’s Disease. Front. Aging Neurosci. 10:202. doi: 10.3389/fnagi.2018.00202
Keywords: microglia, neuron, synapse, cognitive aging, oxidative stress, chronic psychological stress, major depressive disorder
Citation: Carrier M, Šimončičová E, St-Pierre M-K, McKee C and Tremblay M-È (2021) Psychological Stress as a Risk Factor for Accelerated Cellular Aging and Cognitive Decline: The Involvement of Microglia-Neuron Crosstalk. Front. Mol. Neurosci. 14:749737. doi: 10.3389/fnmol.2021.749737
Received: 29 July 2021; Accepted: 16 September 2021;
Published: 04 November 2021.
Edited by:
Stefano Angiari, Medical University of Graz, AustriaReviewed by:
Valeria Carola, Sapienza University of Rome, ItalyMiles Herkenham, National Institute of Mental Health, National Institutes of Health (NIH), United States
Zili You, University of Electronic Science and Technology of China, China
Copyright © 2021 Carrier, Šimončičová, St-Pierre, McKee and Tremblay. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marie-Ève Tremblay, ZXZldHJlbWJsYXlAdXZpYy5jYQ==