
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mol. Biosci. , 25 February 2025
Sec. Molecular Diagnostics and Therapeutics
Volume 12 - 2025 | https://doi.org/10.3389/fmolb.2025.1525103
This article is part of the Research Topic The Role of Natural and Synthetic Antioxidants in the Therapeutic Targeting of Oxidative Stress View all 3 articles
 Matteo Micucci1†
Matteo Micucci1† Federico Gianfanti1†
Federico Gianfanti1† Sabrina Donati Zeppa2
Sabrina Donati Zeppa2 Giosuè Annibalini1
Giosuè Annibalini1 Barbara Canonico1Fabiana Fanelli1
Barbara Canonico1Fabiana Fanelli1 Roberta Saltarelli1Riham Osman1Mariele Montanari1Daniele Lopez1Gemma Nasoni1
Roberta Saltarelli1Riham Osman1Mariele Montanari1Daniele Lopez1Gemma Nasoni1 Giovanna Panza1Erik Bargagni1Francesca Luchetti1Michele Retini1
Giovanna Panza1Erik Bargagni1Francesca Luchetti1Michele Retini1 Michele Mari1Giovanni Zappia2,3*Vilberto Stocchi2
Michele Mari1Giovanni Zappia2,3*Vilberto Stocchi2 Alessia Bartolacci1Sabrina Burattini1
Alessia Bartolacci1Sabrina Burattini1 Michela Battistelli1
Michela Battistelli1Background: Mitochondrial dysfunction and oxidative stress are central mechanisms in the progression of neurodegenerative diseases. This study first evaluated the toxicity of Q-Der (Q10-diacetate), a derivative of Coenzyme Q10, in HT22 hippocampal neurons under normal and oxidative stress conditions.
Methods: HT22 cells were treated with Q-Der at 2.5, 5 and 10 µM with and without rotenone. Mitochondrial superoxide production (Mitosox), gene expression (via qRT-PCR), and protein levels (via Western blot) were measured. Morphological analyses were performed using transmission (TEM) and scanning (SEM) electron microscopes.
Results: Q-Der significantly reduced mitochondrial superoxide levels, particularly at 5 μM, and upregulated key mitochondrial biogenesis genes, including PGC-1α and TFAM. Additionally, it restored the expression of MT-ND1 and MT-COI, which were downregulated by rotenone. Western blot results showed a significant recovery in CV-ATP5A (complex V) expression (p < 0.05), preserving mitochondrial ATP production. Morphological analyses further confirmed Q-Der’s ability to maintain cellular and mitochondrial structure under stress conditions.
Conclusion: These findings suggest that Q-Der is non-toxic under normal conditions and protects against oxidative stress, supporting its potential as a therapeutic agent for neurodegenerative diseases.
Neurodegeneration is an inflammatory response in the central nervous system (CNS). It represents a complex constellation of conditions characterized by the progressive decline in neurons’ structure, function, and viability. This pathological phenomenon is central to various disorders that collectively are known as neurodegenerative diseases (NDs). These conditions—such as Alzheimer’s disease (AD), Parkinson’s disease (PD), amyotrophic lateral sclerosis (ALS), Huntington’s disease (HD), and multiple sclerosis (MS)—are unified by their insidious onset and the inexorable deterioration of cognitive, motor, or sensory functions they inflict upon individuals.
These diseases share a gradual onset and a relentless progression of cognitive, motor, or sensory impairments, making them a leading cause of disability worldwide, particularly among older populations. As society ages, the prevalence of NDs is expected to increase, highlighting the critical need for enhanced understanding and development of innovative preventive strategies.
At the cellular level, neurodegeneration is characterised by a series of events including synaptic dysfunction, neuronal loss, and brain atrophy. These processes are driven by the neurons’ inability to sustain necessary biochemical and cellular operations, leading to apoptosis or other forms of cell death (Granatiero et al., 2019; Angelova and Abramov, 2018; Wilson et al., 2023). This results in the breakdown of neural networks and neurotransmission disruption, manifesting in symptoms depending on the affected brain regions. The molecular mechanisms at play, such as mitochondrial dysfunction, oxidative stress, and low-grade inflammation, are pivotal to understanding the pathology of NDs. Mitochondrial dysfunction compromises cellular energy metabolism, contributing to the cell death seen in NDs. Oxidative stress, resulting from an imbalance between reactive oxygen species (ROS) production and antioxidant defenses, further damages neurons (Lin and Beal, 2006). Key molecular players in these processes include oxidative stress induction and molecular networks related to neuroinflammation and cellular dysfunction, exacerbating neuronal damage (Sivandzade et al., 2019).
The environmental and lifestyle factors, including diet, physical activity, and exposure to toxins, further modulate the risk and progression of neurodegenerative diseases. Here, the role of diet emerges as both a potential risk factor and a protective ally against neurodegeneration (Solfrizzi et al., 2017; Zhang et al., 2021; Bianchi et al., 2021; Yang et al., 2021; Godos et al., 2024). Nutritional patterns rich in polyphenols and healthy fats can mitigate the risk of neurodegenerative diseases by influencing the molecular mechanisms of neurodegeneration. Diets that reduce oxidative stress, dampen low-grade inflammation, and improve mitochondrial function can significantly impact the progression of these diseases. Additionally, consistent consumption of fruits like berries and cherries, which are high in polyphenols, may contribute to maintaining cognitive health (Wang et al., 2023). Investigating phytocomplexes used in folk medicine could lead to the discovery of compounds with diverse preventive benefits across various diseases, including cancer (Micucci et al., 2024), cardiovascular issues (Micucci et al., 2020), and neurological disorders (Lapi et al., 2020).
The exploration of phytocomplexes derived from plants such as Acacia catechu (Chiaino et al., 2021), Castanea sativa (Brizi et al., 2016), Olea europea (Chiaino et al., 2020), and phytochemicals like flavonoids and curcumin have unveiled their potential to modulate critical molecular pathways involved in neurodegeneration. These substances can inhibit the activation of NF-κB, thereby reducing the production of pro-inflammatory cytokines such as IL-1 and IL-6. They also promote the activation of Nrf2, enhancing the cellular antioxidant response and protecting against oxidative damage (Abrahams et al., 2019; Calis et al., 2020; Farzaei et al., 2019). Indeed, generally, flavonoids lead to a reduction in ROS, regardless of their source (endogenous: mitochondria, peroxisomes, xanthine oxidase, Fenton reaction, NADPH oxidase, lipoxygenases, cytochrome P450 or exogenous: visible, UV and ionising radiation, chemotherapeutics) (Kicinska and Jarmuszkiewicz, 2020). Additionally, compounds naturally occurring in the human body, such as melatonin and Coenzyme Q10 (CoQ10), have also been studied and extracted, leading to the development of novel agents to mitigate oxidative stress and support mitochondrial function in neurodegenerative conditions (Areti et al., 2017; Bagheri et al., 2023).
Amidst these complex interactions, CoQ10 and its derivatives stand out for their dual role in mitochondrial function and antioxidant defence. Coenzyme q10 provides a promising avenue for neurodegerative disorders, acting on oxidative phosphorylation, which occurs in the mitochondria via the electron transport chain, the primary process responsible for ATP production (Kang et al., 2020). However, challenges in bioavailability and optimal dosing underscore the need for further research to harness CoQ10’s full potential in combating NDs.
This study introduces a pioneering in vitro comparison of a novel CoQ10 analog with CoQ10, focusing on their neuroprotective effects and underlying mechanisms in HT22 neuronal cells. Our investigation delves into mitochondrial ROS modulation, regulation of genes implicated in mitochondrial biogenesis and function (PGC-1ɑ, TFAM, MT-ND1, MT-COI, MT-COX5B), and OXPHOS subunit protein expression, aiming to illuminate the novel analog’s superior efficacy and mechanism profiles. Finally, SEM and TEM analyses suggest neuroprotective effects evidence by preserved mitochondrial integrity, reduced apoptotic cells, autophagic vacuoles, and cellular morphology in the treatment group compared to controls. This work represents a significant stride in overcoming CoQ10’s dosing challenges, potentially setting a new benchmark in neuroprotective strategies.
CoQ10 was obtained from Sigma-Aldrich (C9538). CoQ10 derivative (Q-Der) was kindly provided by Umolsystem Srl. Rotenone was used as the neurotoxin at a concentration of 5 µM. Dimethyl sulfoxide (DMSO) was used as a vehicle control. Other reagents included fetal bovine serum (FBS), L-glutamine (100 mM), and antibiotics (penicillin and streptomycin). All chemicals were sourced from Sigma-Aldrich and prepared according to the manufacturer’s instructions.
Mouse hippocampal neuronal cell line (HT22) was maintained in DMEM-Ham’s F12, supplemented with 10% fetal bovine serum, L-glutamine (100 mM), and 1% antibiotics (penicillin, streptomycin), and incubated in humidified 5% CO2 atmosphere at 37°C. At 80% confluence, cells were detached with trypsin-EDTA, washed, and sub cultivated in new flasks for 1–2 days before the experimental procedures. Neuroprotective effects were evaluated by pre-incubating cells with varying concentrations of Q10 and Q-Der for 3 or 24 h, followed by a 24-h exposure to 5 µM rotenone to induce neurotoxicity.
Cells were divided into six experimental groups, as reported in Figure 1:
− Control: cells treated with the vehicle (DMSO).
− Q10: cells treated with 5 µM Q10 for 24 h.
− Q-Der Low, Medium, High: cells treated with 2.5, 5, or 10 µM Q-Der for 24 h.
− Rotenone: cells exposed to 5 µM rotenone for 24 h to induce neurotoxicity.
− Q10 + rotenone: cells pre-treated with 5 µM Q10 for 3 or 24 h, followed by 24-h rotenone exposure.
− Q-Der + rotenone: cells pre-treated with 5 µM Q-Der for 3 or 24 h, followed by 24-h rotenone exposure.
Propidium Iodide (PI) staining was performed to assess cell viability. PI is a fluorescent dye that penetrates only cells with compromised membranes, indicating cell death. Control and rotenone-challenged cells treated with 2.5, 5, and 10 µM Q-Der at 24 h post-treatment, were incubated with PI for 10 minutes. PI fluorescence was measured using a flow cytometer (FACSCanto II). The percentage of PI-positive cells, representing dead or necrotic cells, was calculated for each experimental group.
Mitochondrial ROS production was measured using MitoSOX Red (Thermo Fischer, M36008), a dye specifically oxidized by mitochondrial superoxide. Control and rotenone-challenged HT22 cells treated with 2.5, 5, and 10 µM Q-Der for 24 h were incubated with 5 µM MitoSOX Red for 10 min at 37°C. Flow cytometry measured fluorescence intensity (excitation wavelength 561 nm, emission wavelength 610 nm). The results were expressed as percentages of MitoSOX positive cells.
Total RNA was isolated from treated and control cells using the Omega Bio-Tek E.Z.N.A.™ Total RNA Kit following the manufacturer’s protocol. RNA concentrations were measured using the SpectraMax® QuickDrop™ Micro-Volume Spectrophotometer. cDNA was synthesized from 500 ng of total RNA using the Takara PrimeScript™ RT Master Mix. Quantitative real-time PCR (qRT-PCR) was performed using the StepOnePlus™ Real-Time PCR System (Applied Biosystems) and PowerUp SYBR Green Master Mix. Primers used for PGC-1α, TFAM, MT-ND1, MT-COI, COX5B, and S16 were designed based on previous studies (see Table 1 for primer sequences). Relative gene expression was normalized to S16 as the internal control, and data were analyzed using the 2−ΔΔCT method.
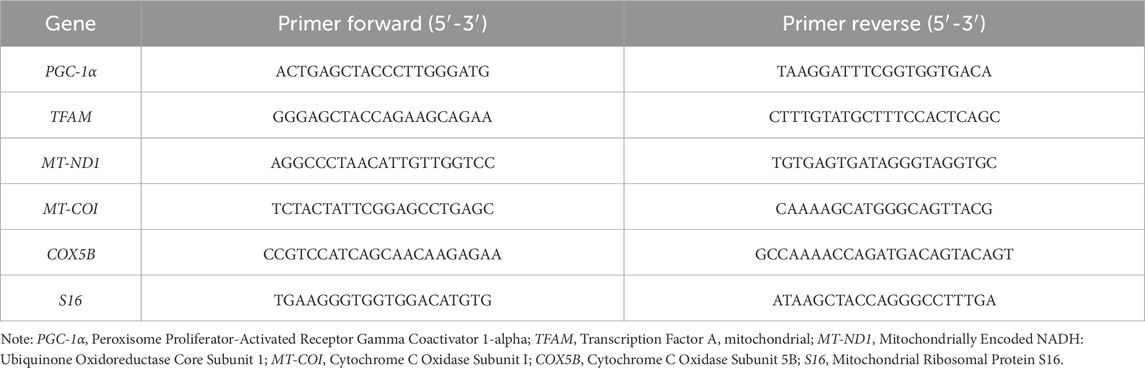
Table 1. Primers used in real-time RT-PCR quantification.
The target genes and the corresponding primer sequences used in qRT-PCR quantification are provided in Table 1.
Total protein extracts were obtained from the organic phase following the QIAzol protocol, solubilised in ISOT lysis buffer containing 8 M urea, 4% CHAPS, 65 mM DTE, 40 mM Tris base, 1 mM NaF, 1 mM Na3VO4, 1× complete protease inhibitor cocktail (Roche Diagnostics), then sonicated three times for 5 s on ice. After centrifugation at 12,000xg for 10 min, protein concentration was determined using Bradford colorimetric assay (Bio-Rad Laboratories) (Bradford, 1976). Equal amounts (30 μg) of total proteins were resolved in precast stain-free 4%–15% SDS polyacrylamide gels (Bio-Rad Laboratories, 4568084) and electrotransferred to polyvinylidene difluoride (PVDF) membranes (Bio-Rad Laboratories, 1620177) using the Power Blotter System semi-dry transfer device (ThermoFisher Scientific). Western blot analyses were performed using total oxidative phosphorylation (OXPHOS) cocktail from Invitrogen (458099) diluted 1:1,000 to detect individual complexes of the electron transport chain: CI-NDUFB8 (20 kDa), CII-SDHB (30 kDa), CIII-UQCRC2 (48 kDa), CIV-MTCO1 (40 kDa) and CV-ATP5A (55 kDa) (Welinder and Ekblad, 2011).
Control and treated HT22 hippocampal cells were directly processed on coverslips in Petri dishes. After careful washing with 0.1 M phosphate buffer, monolayers were fixed with 2.5% glutaraldehyde in the same buffer for 1 h. All the specimens were post-fixed with 1% OsO4 in 0.1 M phosphate buffer for 1 h. After alcohol dehydration, they were critical point dried, gold-sputtered, and observed with an ESEM scanning electron microscope (UMKC, Kansas City, MO, United States). For TEM analysis, HT22 hippocampal cells, growing adherent in flasks, were washed, immediately fixed “in situ” with 2.5% glutaraldehyde in 0.1 M phosphate buffer for 30 min, gently scraped, and centrifuged at 1,200 rpm. The pellets were fixed in 2.5% glutaraldehyde for an additional 30 min. All the specimens were post-fixed in 1% OsO4 for 1 h, alcohol dehydrated, and embedded in araldite (Salucci et al., 2015). Ultrathin sections of the embedded samples were cut using a Leica UC7 ultramicrotome, and thin sections were stained with uranyl acetate and lead citrate and then analyzed with a Philips CM10 transmission electron microscope.
For a holistic analysis of data derived from various assays discussed—including cell viability, mitochondrial functionality, and qRT-PCR for PGC-1α, TFAM, MT-ND1, MT-COI, and COX5B expression—statistical evaluation was meticulously designed to ensure the accuracy and reliability of the findings. Initially, all data were subjected to the Shapiro-Wilk test to assess normality, a critical step in determining the appropriate statistical methods for analysis. For data conforming to a normal distribution, analysis of variance (ANOVA) was employed to compare the mean values across different experimental and control groups. Subsequent post hoc analysis, utilizing Tukey’s and Sidak’s tests, helped identify specific groups showing statistically significant differences. In contrast, for datasets not meeting normal distribution criteria, the Kruskal-Wallis test was applied as a non-parametric alternative to ANOVA for comparing means across groups. Dunn’s post hoc test was then used for detailed pairwise comparisons to pinpoint where significant differences lay. The statistical analyses were performed using advanced software tools like GraphPad Prism, ensuring high precision and reliability. The significance threshold was consistently set at p < 0.05 across all tests. Results were reported as mean ± SD for parametric data sets or median with interquartile range for non-parametric datasets, appropriately representing the data’s central tendency and dispersion, from at least three independent experiments. Statistical significance was indicated as p < 0.05 (*), p < 0.01 (**), and p < 0.001 (***).
The results from the PI staining assay reveal distinct effects of Q-Der, both in the absence and presence of rotenone, on HT22 cell viability (Figure 2). Under baseline conditions, healthy cells treated with Q-Der at 5 µM show a reduction in PI-positive cells compared to the control. However, this decrease was not significant at 2.5 µM. These data suggest the in vitro safety profile of Q-Der under non-stressful conditions.
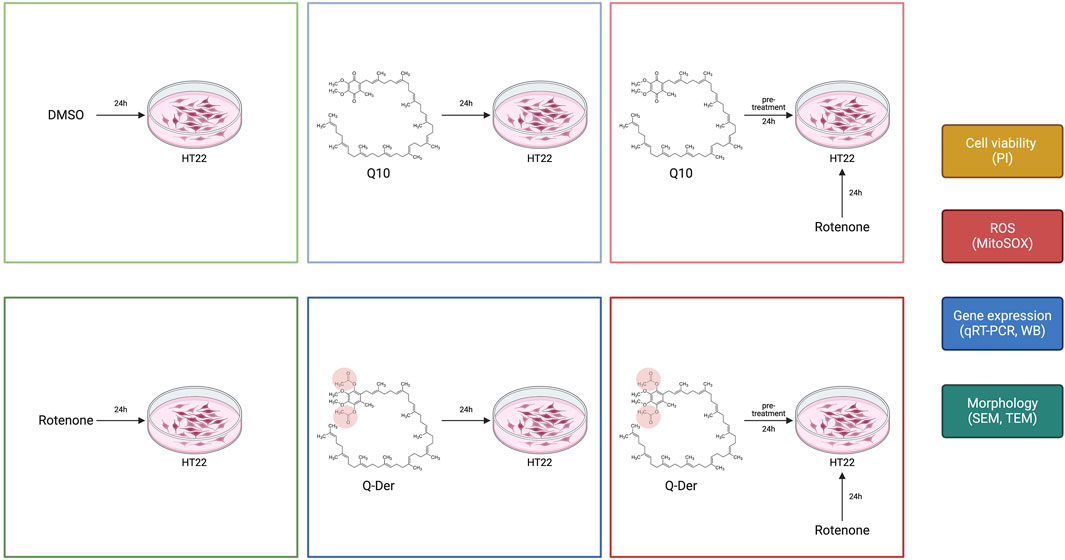
Figure 1. Design of the experimental procedure on HT22 cells in the different experimental conditions.
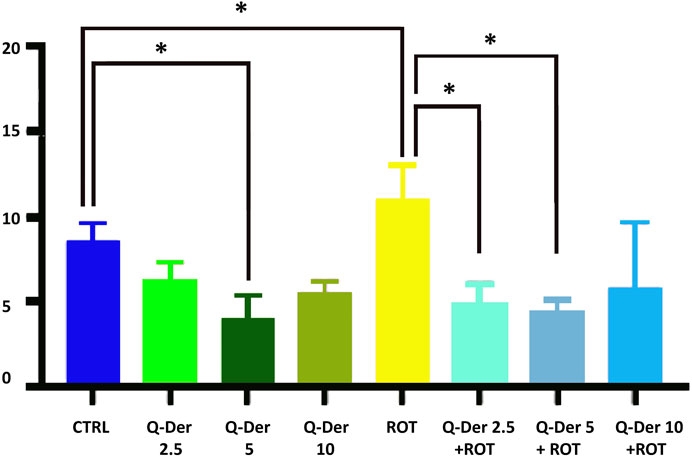
Figure 2. Q-Der effects on HT22 viability in healthy and rotenone-treated cells. The panel shows the percentage of PI positive cells, indicating cell death. Ordinary one-way ANOVA, Šidák’s multiple comparison, *P value <0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
The neurotoxic effects of rotenone were evident, as indicated by a marked increase in PI-positive cells relative to the control. Pre-treatment with Q-Der at both concentrations significantly attenuates this rotenone-induced effect. Notably, the reduction in PI-positive cells in the Q-Der pre-treated groups approaches levels observed in control cells, indicating a robust protective effect against rotenone-mediated cytotoxicity.
These findings underscore the in vitro efficacy of Q-Der in preserving cell viability under oxidative stress conditions, with its protective effects becoming more pronounced in the presence of rotenone compared to the modest impact observed in the absence of the neurotoxin.
The results of the MitoSOX Red assay, used to quantify mitochondrial superoxide production, demonstrate the efficacy of Q-Der in mitigating oxidative stress induced by rotenone. In the control group, mitochondrial superoxide levels remain at baseline, suggesting physiological mitochondrial function. Mitochondrial ROS levels were not modified in healthy cells treated with Q-Der at all concentrations. On the contrary, rotenone-induced a substantial elevation in superoxide production. Pre-treatment with Q-Der at 2.5 µM and 5 µM reduced the rotenone-induced increase of mitochondrial ROS levels, nearly restoring them to baseline. Q-Der at 10 µM reduces mitochondrial superoxide levels to a lesser extent than the lower doses, suggesting a potential dose-dependent plateau. These findings indicate that Q-Der effectively counters oxidative stress at optimal concentrations, with a strong statistical significance in the protective effects observed at 2.5 µM and 5 µM. This underscores its potential to preserve mitochondrial superoxide levels under induced oxidative stress (Figure 3).
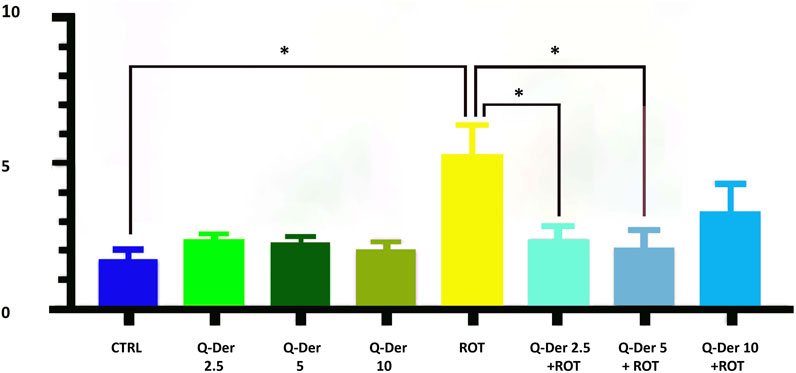
Figure 3. Evaluation of mitochondrial superoxide levels in control and rotenone-treated cells upon the treatment of 2.5, 5, and 10 µM Q-Der. Ordinary one-way ANOVA, Šidák’s multiple comparison, *P value <0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
The qRT-PCR results reveal significant insights into the impact of Q-Der on the expression of key mitochondrial genes under basal and rotenone-induced conditions. The genes analyzed include PGC-1α, TFAM, MT-ND1, MT-COI, and COX5B, integral to mitochondrial biogenesis, respiratory function, and oxidative phosphorylation.
In the absence of rotenone, Q-Der at 5 µM induced a noticeable upregulation of PGC-1α expression, while CoQ10 at the same concentration did not affect this parameter. The expression of TFAM, a gene essential for mitochondrial DNA maintenance, was not modified by CoQ10 or Q-Der treatment. As expected, rotenone markedly reduced TFAM expression, both CoQ10 and Q-Der partially antagonized this effect. MT-ND1 and MT-COI, which encode subunits of the mitochondrial respiratory chain complexes I and IV, showed a similar pattern: the active compounds partially restored their rotenone-decreased expression. Finally, COX5B, a nuclear-encoded Cytochrome C oxidase subunit (complex IV), increased its expression only in rotenone-challenged cells treated with CoQ10 and Q-Der, with Q-Der showing a higher potency.
The qRT-PCR analysis reveals that Q-Der is critical in promoting mitochondrial gene expression, particularly under oxidative stress conditions. The significant upregulation of PGC-1α, partial recovery of TFAM, and restoration of MT-ND1 and MT-COI expression highlight Q-Der’s efficacy in maintaining mitochondrial function and biogenesis (Figure 4).
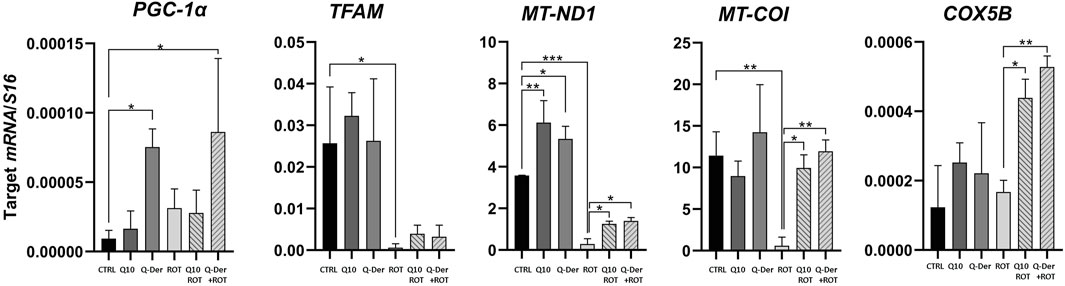
Figure 4. Mitochondrial-Related Gene expression. HT22 mRNA expression levels of PGC-1ɑ, TFAM, MT-ND1, MT-COI and COX5B in healthy and rotenone-treated cells challenged with CoQ10 and Q-Der at 5 μM *P < 0.05, **P < 0.01, ***P < 0.001.
The Western blot analysis of the OXPHOS complexes reveals key findings, particularly in relation to the expression of CI-NDUFB8 (complex I), CII-SDHB (complex II), CIII-UQCRC2 (complex III), and CV-ATP5A (complex V). Despite the range of experimental conditions, the analysis shows that the expression of CI-NDUFB8 remains unaffected by rotenone treatment. There is no statistically significant alteration in CI-NDUFB8 levels across all groups, including both Q-Der and CoQ10 pre-treatments. This stability indicates that rotenone does not exert a detectable impact on complex I subunit expression under these experimental conditions, and neither Q-Der nor Q10 significantly modifies CI-NDUFB8 levels. Similarly, the expression of CII-SDHB and CIII-UQCRC2 follows a comparable trend, with no statistical significant differences were observed between the control, rotenone, and pre-treatment groups. Although there is a slight trend toward recovery in the pre-treated groups, these changes do not meet the threshold for statistical significance. These observations suggest that while a minor biological effect may exist, it is not strong enough to be conclusive in terms of altering the expression of complex II and complex III subunits. The only subunit showing a statistically significant change is CV-ATP5A (complex V). Q-Der increased this subunit expression only in rotenone-treated cells, while CoQ10 determined a similar effect both in control and rotenone-treated cells (Figure 5).
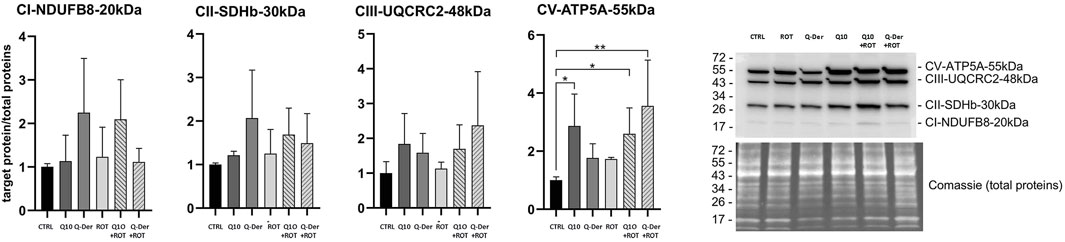
Figure 5. OXPHOS protein expression. Densitometric analyses of subunits of complexes I (CI-NDUFB8, 20 kDa), II (CII-SDHB, 30 kDa), III (CIII-UQCRC2, 48 kDa) and V (CV-ATP5A, 55 kDa) of the OXPHOS system normalised to total proteins. A representative Western blot image of OXPHOS and total proteins is shown on the right side of the figure.
In the control condition at 24 h, cell morphology appears well preserved, HT22 cells appear elongated, and sometimes, some show a slightly rounded shape Figures 6A–C. The cells appear confluent and generate a carpet by forming extracellular membrane protrusions that create bridges between adjacent cells.
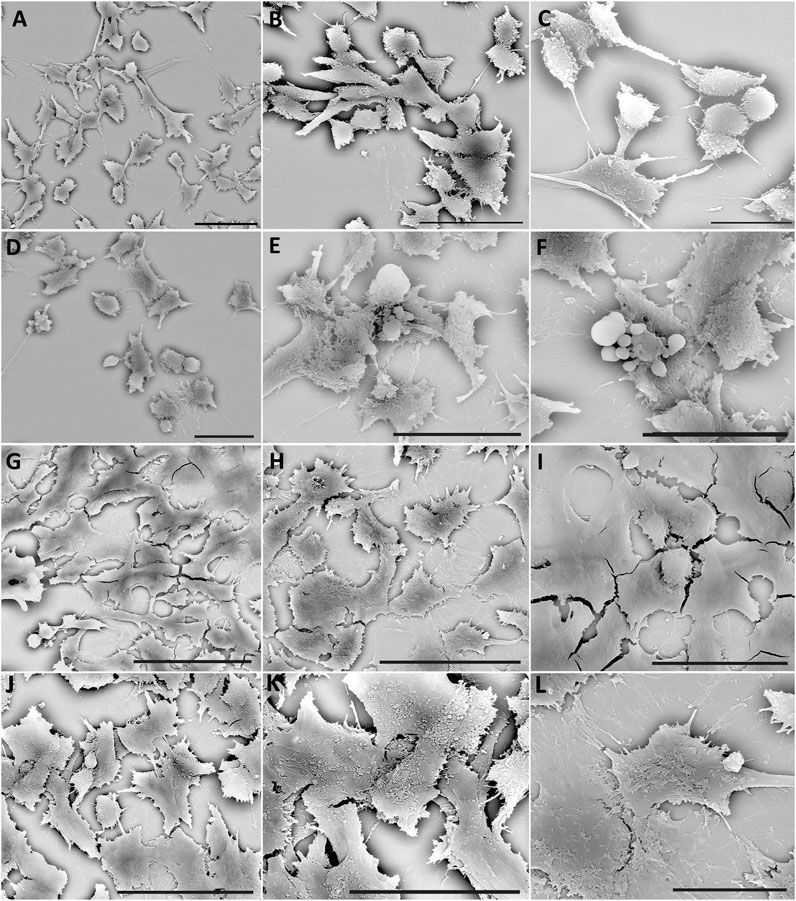
Figure 6. SEM of HT22 hippocampal cells. Control cells (A–C); cells treated with rotenone (5 µM) for 24 h (D–F); rotenone-treated cells pretreated with Q10 (5 µM) for 24 h (G–I), rotenone-treated cells pretreated with Q-Der (5 µM) for 24 h (J–L). (A, B, D–I), Bar = 20 µm; (C, J, K), Bar = 10 µm.
24 h of rotenone treatment induces a deep change in cell morphology; the HT22 appears predominantly rounded, probably due to a rearrangement of the cytoskeletal structure (Figures 6D–F).
The cells show fewer cytoplasmic protrusions, and a loss of extracellular junctions reduces cell carpet formation. Numerous blebs on the cellular membrane and the apoptotic bodies demonstrate the presence of numerous apoptotic cells.
After pre-treatment with Q10, the morphology is restored to the control’s; new cell junctions (Figures 6G–I) can be formed. Finally, pre-treatment with Q-Der shows the cells all flattened, more confluent, and with characteristic microvilli on the surface (Figures 6J–L). Both Q10 and Q-Der can reduce the percentage of apoptotic cells and induce the percentage of apoptotic cells and the preservation of cell communication.
For all experimental conditions we evaluate three grids. Every grid consisted of ten thin sections. Therefore, we performed the evaluation of the ultrastructural changes in 200–300 cells. The ultrastructural evaluation evidence good cell viability in control conditions (Figures 7A–C). HT22 cell morphology appeared elongated and confluent. After rotenone treatment, several rounded cells with blebs and apoptotic bodies appeared (Figures 7D–F). We can observe numerous autophagic vacuoles (G), with different sizes ranging from 200 to 500 nm. In the nucleus, we can observe chromatin condensation and a diffuse detachment of the nuclear membrane. Mitochondria appear swollen and heavily damaged; their maintained membrane is well preserved but appears well preserved while create are completely disrupted.
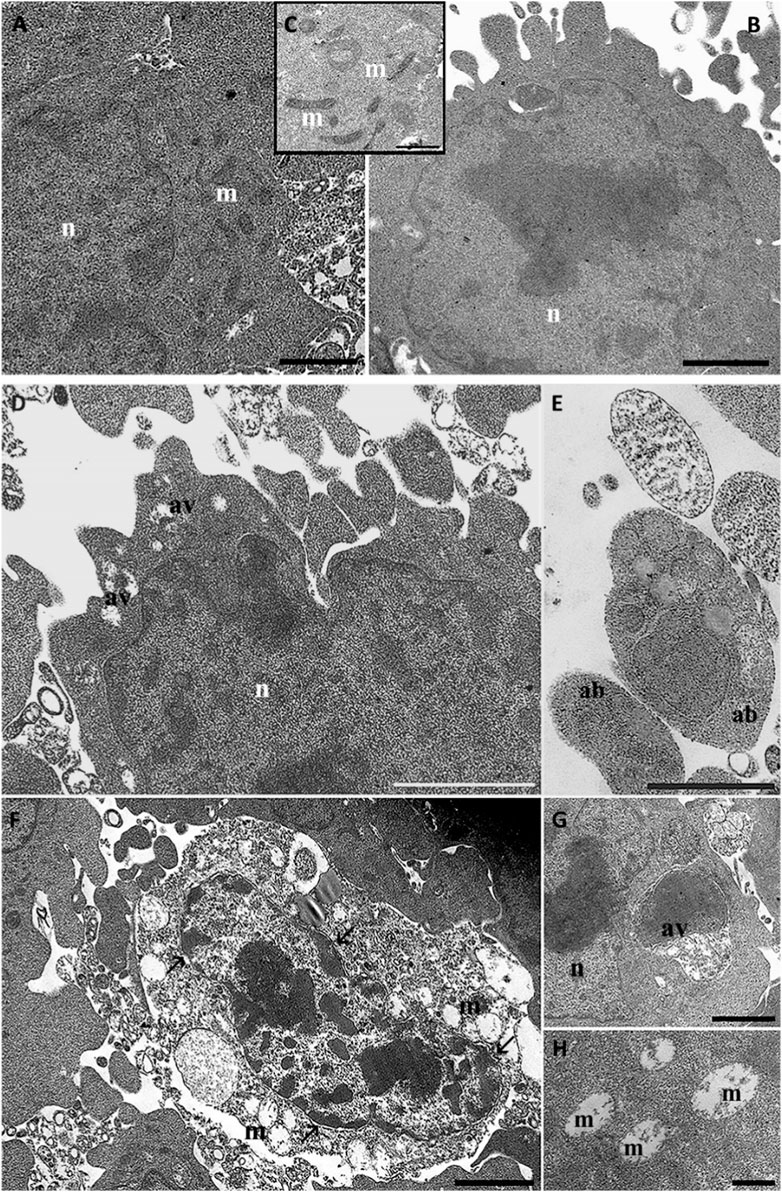
Figure 7. HT22 hippocampal cells at TEM. Control (A, B), cells treated with rotenone (5 µM) for 24 h (C–F, G). m: mitochondria; n: nucleus; ab: apoptotic bodies; av: autophagic vacuole; →: chromatin condensation. (A, B), Bar = 1 μm; (C, F), Bar = 500 nm; (D, E), Bar = 2 µm.
After Q10 treatment, the cells appeared confluent and showed well-preserved mitochondria (Figures 8A–C) comparable to the control conditions. A better mitochondria morphology appeared when cells were treated with Q10-der (Figures 8D–F). We can see well-preserved mitochondria that appear more numerous if compared pre-treatment to control or rotenone treatment. Mitochondria are present in hight number but smaller than control condition, suggesting a mitochondrial biogenesis.
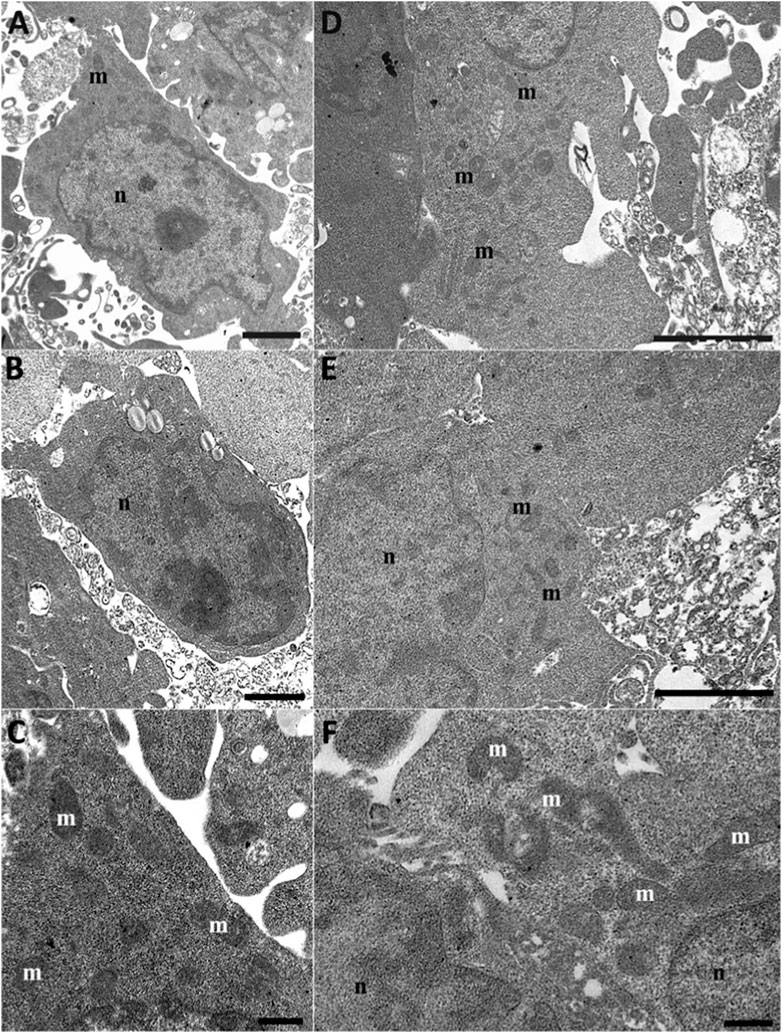
Figure 8. HT22 hippocampal cells at TEM. Rotenone-treated cells pretreated with Q10 (5 µM) for 24 h Q10 (A–C) and rotenone-treated cells pretreated with Q-Der (5 µM) for 24 h Q-der (D–F). m: mitochondria; n: nucleus. (A–C, E), Bar = 2 μm; (C, F), Bar = 500 nm.
The findings from this study highlight the significant protective effects of Q-Der on HT22 neuronal cells, particularly under conditions of oxidative stress induced by rotenone. This condition offers a targeted model for studying mitochondrial dysfunction, as it selectively inhibits Complex I of the electron transport chain, disrupting ATP synthesis and causing localized ROS generation (Menke et al., 2003; Moon et al., 2005). This inhibition leads to mitochondrial membrane depolarization, increased expression of mitochondrial fission markers and activation of caspase-3-dependent apoptotic pathways. These effects are highly relevant to neurodegenerative diseases like Parkinson’s disease, where mitochondrial dysfunction and selective dopaminergic neuronal death are central features (Moon et al., 2005; Li et al., 2017). Several stressors act on neurons through the affection of the same pathways targeted by rotenone. For example, H₂O₂ induces oxidative stress generating ROS across multiple cellular compartments, leading to generalized oxidative damage that is not specific to mitochondria. Despite this distinction, both rotenone and H₂O₂ share some key effects, including ROS generation, mTOR pathway suppression, apoptotic activation involving caspase-3 cleavage and PARP fragmentation. These shared mechanisms provide complementary insights into oxidative stress and neuroprotection, with rotenone being a specific tool for the study of mitochondria-dependent neurodegenerative pathologies (Panee et al., 2007; Zhou et al., 2015; Giraldo-Berrio et al., 2024; Roy et al., 2023; Millichap et al., 2024).
Our study focuses on rotenone-induced oxidative stress due to its mitochondria-specific effects. Our results add to a growing body of evidence that underscores the importance of targeting mitochondrial dysfunction in neurodegenerative diseases such as Parkinson’s and Alzheimer’s, where oxidative stress plays a pivotal role. Q-Der demonstrated substantial efficacy in reducing cell death, restoring mitochondrial superoxide levels, and modulating key gene expressions involved in mitochondrial biogenesis and respiration. While the protective effects of Q10 have been well-documented in similar models (Bhagavan and Chopra, 2006), the current data suggest that Q-Der may offer superior benefits, particularly in maintaining mitochondrial homeostasis under stress conditions.
Chemically, Q-Der differs from Coenzyme Q10 in that it carries two acetyl groups on its quinone ring, increasing its lipophilicity and potentially enhancing its stability against oxidation. While CoQ10 functions through rapid cycling between oxidized and reduced forms to neutralize reactive oxygen species, the addition of acetate groups in Q-Der is theorized to confer a protective effect on the molecule itself, reducing its reactivity to oxidative agents. This structural modification may support a prolonged antioxidant presence within the mitochondrial membrane, thus maintaining cellular protection over extended periods of oxidative stress. These properties underscore Q-Der’s potential as a neuroprotective agent, particularly in environments where mitochondrial stability and sustained antioxidant effects are essential.
A key aspect of this study is the ability of Q-Der to preserve mitochondrial function and ROS levels. Compared to Q10, Q-Der consistently showed a trend toward better protection against oxidative stress markers such as mitochondrial superoxide, although this trend did not reach statistical significance. This mirrors findings in other studies where derivatives of Q10, such as MitoQ and Ubidecarenone, have been shown to exhibit enhanced mitochondrial targeting and antioxidant properties compared to the parent compound (Smith and Murphy, 2010). MitoQ, a mitochondria-targeted form of CoQ10, has demonstrated strong efficacy in reducing mitochondrial ROS and improving bioenergetics by selectively accumulating in the mitochondrial membrane, facilitating more effective protection against oxidative damage (Cochemé et al., 2007). Although MitoQ and Q-Der share similarities in their mechanisms of action—both aim to enhance mitochondrial function by modulating ROS levels—the differences in their molecular structures likely contribute to their varying degrees of efficacy. For instance, MitoQ’s triphenylphosphonium cation facilitates its direct accumulation in the mitochondria, while Q-Der may rely more on its metabolic conversion within cells.
The qRT-PCR analysis further underscores the potential of Q-Der, particularly in its ability to upregulate key genes involved in mitochondrial biogenesis, such as PGC-1α and TFAM. This is consistent with literature indicating that compounds enhancing PGC-1α activity could promote mitochondrial biogenesis and repair, thereby offering neuroprotection (Wenz, 2009). The upregulation of PGC-1α and partial rescue of TFAM under rotenone-induced oxidative stress suggest that Q-Der helps restore mitochondrial integrity and function, which is crucial for maintaining ATP production and preventing cellular apoptosis. Comparatively, studies on other Q10 derivatives like Q10-hydroxydecyl benzoate (IDE) have shown similar mitochondrial-protective effects, particularly in preventing the loss of complex I and complex III activity, further highlighting the importance of preserving mitochondrial bioenergetics in disease models (Villalaín, 2004). However, Q-Der’s ability to modulate both mitochondrial biogenesis and complex V expression distinguishes it as a promising candidate for further exploration.
The Western blot analysis revealed that while there were no significant changes in the expression of CI-NDUFB8, CII-SDHB, or CIII-UQCRC2 across the treatment groups, Q-Der significantly increased CV-ATP5A (complex V) expression in rotenone-treated cells. This result is particularly noteworthy, as complex V is essential for ATP synthesis, and its dysfunction is a critical contributor to energy deficits observed in neurodegenerative diseases (Nicholls and Budd, 2000). The statistically significant recovery of CV-ATP5A in Q-Der-treated cells indicates that this novel compound effectively preserves mitochondrial ATP production capacity under oxidative stress conditions. This protective effect on ATP synthase has also been observed in studies using idebenone, a synthetic analog of CoQ10, which has shown promise in clinical trials for conditions such as Friedreich’s ataxia and Leber’s hereditary optic neuropathy (Schiff and Rustin, 2016; Lyseng-Williamson, 2016). While idebenone primarily targets complex I deficiency, the broader effect of Q-Der on many mitochondrial parameters suggests a more comprehensive mitochondrial support mechanism.
An interesting finding in our study is that, despite QDer improving numerous parameters of mitochondrial function, it does not appear to increase the levels of UQCRC2, a key component of Complex III of the electron transport chain. This result may seem to contrast with well-established CoQ10-mediated regulation of other mitochondrial markers (Yousef et al., 2019). One possible explanation is that QDer exerts its effects primarily on Complex I or V, leaving Complex III relatively unaffected. Additionally, UQCRC2 levels might already be saturated under basal conditions, leaving little room for further regulation. Studies on PARL-deficient models demonstrate selective defects in Complex III mediated by TTC19 instability without impacting other complexes of the ETC (Spinazzi et al., 2019).
Furthermore, the partial increase in Complex III activity observed in idebenone-treated models suggests that increased protein expression does not always correlate with functional recovery (Llewellyn et al., 2015). Similarly, the unchanged GSSG:GSH ratio in the cerebellum suggests that certain oxidative stress markers may resist modulation even under treatment.
Morphologically, the structural preservation observed through SEM and TEM further supports the molecular data. Even after rotenone exposure, cells pre-treated with Q-Der exhibited well-preserved cellular morphology and mitochondrial structure. This contrasts with the significant cytoskeletal and mitochondrial damage seen in rotenone-treated cells, aligning with the concept that maintaining mitochondrial integrity is crucial for cellular survival under oxidative stress. Studies on other derivatives, such as SkQ1 (a plastoquinone derivative), have also demonstrated the importance of targeting mitochondrial health to prevent structural damage in neurons, supporting the idea that mitochondrial-targeted therapies can significantly affect disease progression (Fedorov et al., 2022; Sacks et al., 2021). However, while SkQ1 is known for its anti-apoptotic properties, Q-Der offers a broader protective mechanism by reducing oxidative stress and enhancing mitochondrial gene expression, further solidifying its role as a potential neuroprotective agent.
In conclusion, the results of this study place Q-Der in a promising position as a neuroprotective agent aimed at preserving mitochondrial function, particularly in NDs. While other derivatives of Q10, such as MitoQ, idebenone, and SkQ1, have shown efficacy in specific mitochondrial pathways, Q-Der appears to provide a multifaceted approach, combining antioxidant effects, mitochondrial biogenesis and enhanced ATP production. The statistical significance observed in the restoration of CV-ATP5A expression and the trends toward improvement in mitochondrial superoxide levels and gene expression positions Q-Der as a strong candidate for further in vivo studies and eventual clinical translation. Future research should aim to explore the exact molecular pathways through which Q-Der exerts its effects and to compare its efficacy in vivo against other Q10 derivatives in models of neurodegeneration. Additionally, a deeper understanding of the pharmacokinetics and bioavailability of Q-Der relative to other mitochondrial-targeted therapies will be crucial in determining its therapeutic potential.
The present study provides compelling evidence of Q-Der’s neuroprotective efficacy in HT22 cells, highlighting its ability to attenuate rotenone-induced oxidative stress and preserve mitochondrial function. Q-Der’s effects were evident through significant reductions in mitochondrial superoxide production, enhanced expression of mitochondrial biogenesis genes (PGC-1α and TFAM), and the upregulation of complex V ATP synthase, contributing to sustained ATP synthesis under stress conditions. These findings support Q-Der as a possible candidate for further investigation in neurodegenerative disease models, particularly those characterized by mitochondrial dysfunction and oxidative stress.
While this study establishes Q-Der’s protective effects in vitro, its translational potential requires further examination. The exclusive use of HT22 cells limits applicability across broader neuronal models, and in vivo studies are essential to confirm bioavailability and metabolic stability. Future research should expand to diverse cellular models and focus on Q-Der’s pharmacokinetics to substantiate its therapeutic promise.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
MMi: Conceptualization, Funding acquisition, Writing - review and editing. FG: Investigation, Writing - original draft. SD: Investigation, Writing - original draft. GA: Investigation, Writing - original draft. BC: Investigation, Writing - original draft. FF: Investigation, Writing - original draft. RS: Investigation, Writing - original draft. RO: Investigation, Writing - original draft. MMo: Investigation, Writing - original draft. DL: Investigation, Writing - original draft. GN: Investigation, Writing - original draft. GP: Investigation, Writing - original draft. EB: Investigation, Writing - original draft. FL: Investigation, Writing - original draft. MR: Investigation, Writing - original draft. MMa: Investigation, Writing - original draft. GZ: Supervision, Writing - original draft. VS: Visualization, Writing - original draft. AB: Visualization, Writing - original draft. SB: Investigation, Writing - original draft. MB: Conceptualization, Investigation, Writing - original draft, Writing - review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This study was supported by the University of Urbino Carlo Bo.
The authors would like to thank Samuela Capellacci, University of Urbino, Carlo Bo.
Author ZG was employed by Umolsystem Srl.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmolb.2025.1525103/full#supplementary-material
Abrahams, S., Haylett, W. L., Johnson, G., Carr, J. A., and Bardien, S. (2019). Antioxidant effects of curcumin in models of neurodegeneration, aging, oxidative and nitrosative stress: a review. Neuroscience 406, 1–21. doi:10.1016/j.neuroscience.2019.02.020
Angelova, P. R., and Abramov, A. Y. (2018). Role of mitochondrial ROS in the brain: from physiology to neurodegeneration. FEBS Lett. 592 (5), 692–702. doi:10.1002/1873-3468.12964
Areti, A., Komirishetty, P., Akuthota, M., Malik, R. A., and Kumar, A. (2017). Melatonin prevents mitochondrial dysfunction and promotes neuroprotection by inducing autophagy during oxaliplatin-evoked peripheral neuropathy. J. Pineal. Res. 62. doi:10.1111/jpi.12393
Bagheri, S., Haddadi, R., Saki, S., Kourosh-Arami, M., Rashno, M., Mojaver, A., et al. (2023). Neuroprotective effects of coenzyme Q10 on neurological diseases: a review article. Front. Neurosci. 17, 1188839. doi:10.3389/fnins.2023.1188839
Bhagavan, H. N., and Chopra, R. K. (2006). Coenzyme Q10: absorption, tissue uptake, metabolism and pharmacokinetics. Free Radic. Res. 40, 445–453. doi:10.1080/10715760600617843
Bianchi, V. E., Herrera, P. F., and Laura, R. (2021). Effect of nutrition on neurodegenerative diseases. A systematic review. Nutr. Neurosci. 24 (10), 810–834. doi:10.1080/1028415X.2019.1681088
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254. doi:10.1006/abio.1976.9999
Brizi, C., Santulli, C., Micucci, M., Budriesi, R., Chiarini, A., Aldinucci, C., et al. (2016). Neuroprotective effects of Castanea sativa mill. Bark extract in human neuroblastoma cells subjected to oxidative stress. J. Cell. Biochem. 117, 510–520. doi:10.1002/jcb.25302
Calis, Z., Mogulkoc, R., and Baltaci, A. K. (2020). The roles of flavonols/flavonoids in neurodegeneration and neuroinflammation. Mini-Reviews Med. Chem. 20, 1475–1488. doi:10.2174/1389557519666190617150051
Chiaino, E., Micucci, M., Cosconati, S., Novellino, E., Budriesi, R., Chiarini, A., et al. (2020). Olive leaves and Hibiscus flowers extracts-based preparation protect brain from oxidative stress-induced injury. Antioxidants (Basel) 9, 806. doi:10.3390/antiox9090806
Chiaino, E., Stella, R., Peggion, C., Micucci, M., Budriesi, R., Mattioli, L. B., et al. (2021). Acacia catechu willd. Extract protects neuronal cells from oxidative stress-induced damage. Antioxidants (Basel) 11, 81. doi:10.3390/antiox11010081
Cochemé, H. M., Kelso, G. F., James, A. M., Ross, M. F., Trnka, J., Mahendiran, T., et al. (2007). Mitochondrial targeting of quinones: therapeutic implications. Mitochondrion 7, S94–S102. doi:10.1016/j.mito.2007.02.007
Farzaei, M. H., Singh, A. K., Kumar, R., Croley, C. R., Pandey, A. K., Coy-Barrera, E., et al. (2019). Targeting inflammation by flavonoids: novel therapeutic strategy for metabolic disorders. Int. J. Mol. Sci. 20, 4957. doi:10.3390/ijms20194957
Fedorov, A. V., Chelombitko, M. A., Chernyavskij, D. A., Galkin, I. I., Pletjushkina, O.Yu., Vasilieva, T. V., et al. (2022). Mitochondria-targeted antioxidant SkQ1 prevents the development of experimental colitis in mice and impairment of the barrier function of the intestinal epithelium. Cells 11, 3441. doi:10.3390/cells11213441
Giraldo-Berrio, D., Mendivil-Perez, M., Velez-Pardo, C., and Jimenez-Del-Rio, M. (2024). Rotenone induces a neuropathological phenotype in cholinergic-like neurons resembling Parkinson's disease dementia (PDD). Neurotox. Res. 42 (3), 28. doi:10.1007/s12640-024-00705-3
Godos, J., Micek, A., Mena, P., Del Rio, D., Galvano, F., Castellano, S., et al. (2024). Dietary (Poly)phenols and cognitive decline: a systematic review and meta-analysis of observational studies. Mol. Nutr. Food Res. 68 (1), e2300472. doi:10.1002/mnfr.202300472
Granatiero, V., Pacifici, M., Raffaello, A., De Stefani, D., and Rizzuto, R. (2019). Overexpression of mitochondrial calcium uniporter causes neuronal death. Oxid. Med. Cell Longev. 2019, 1681254. doi:10.1155/2019/1681254
Kang, L., Liu, S., Li, J., Tian, Y., Xue, Y., and Liu, X. (2020). The mitochondria-targeted anti-oxidant MitoQ protects against intervertebral disc degeneration by ameliorating mitochondrial dysfunction and redox imbalance. Cell. Prolif. 53, e12779. doi:10.1111/cpr.12779
Kicinska, A., and Jarmuszkiewicz, W. (2020). Flavonoids and mitochondria: activation of cytoprotective pathways? Molecules 25, 3060. doi:10.3390/molecules25133060
Lapi, D., Stornaiuolo, M., Sabatino, L., Sommella, E., Tenore, G., Daglia, M., et al. (2020). The pomace extract taurisolo protects rat brain from ischemia-reperfusion injury. Front. Cell. Neurosci. 14, 3. doi:10.3389/fncel.2020.00003
Li, H. N., Zimmerman, M., Milledge, G. Z., Hou, X. L., Cheng, J., Wang, Z. H., et al. (2017). Water-soluble coenzyme Q10 reduces rotenone-induced mitochondrial fission. Neurochem. Res. 42 (4), 1096–1103. doi:10.1007/s11064-016-2143-2
Lin, M. T., and Beal, M. F. (2006). Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 443, 787–795. doi:10.1038/nature05292
Llewellyn, K. J., Nalbandian, A., Gomez, A., Wei, D., Walker, N., and Kimonis, V. E. (2015). Administration of CoQ10 analogue ameliorates dysfunction of the mitochondrial respiratory chain in a mouse model of Angelman syndrome. Neurobiol. Dis. 76, 77–86. doi:10.1016/j.nbd.2015.01.005
Lyseng-Williamson, K. A. (2016). Idebenone: a review in Leber’s hereditary optic neuropathy. Drugs 76, 805–813. doi:10.1007/s40265-016-0574-3
Menke, T., Gille, G., Reber, F., Janetzky, B., Andler, W., Funk, R. H., et al. (2003). Coenzyme Q10 reduces the toxicity of rotenone in neuronal cultures by preserving the mitochondrial membrane potential. Biofactors 18 (1-4), 65–72. doi:10.1002/biof.5520180208
Micucci, M., Bolchi, C., Budriesi, R., Cevenini, M., Maroni, L., Capozza, S., et al. (2020). Antihypertensive phytocomplexes of proven efficacy and well-established use: mode of action and individual characterization of the active constituents. Phytochemistry 170, 112222. doi:10.1016/j.phytochem.2019.112222
Micucci, M., Xiang, B. Z., Ting, C. M., Kwan, H. Y., Mari, M., Retini, M., et al. (2024). Matching traditional Chinese medicine and western medicine-based research: advanced nutraceutical development for proactive gastric cancer prevention. World J. Gastrointest. Oncol. 16, 3798–3819. doi:10.4251/wjgo.v16.i9.3798
Millichap, L., Turton, N., Damiani, E., Marcheggiani, F., Orlando, P., Silvestri, S., et al. (2024). The effect of neuronal CoQ10 deficiency and mitochondrial dysfunction on a rotenone-induced neuronal cell model of Parkinson's disease. Int. J. Mol. Sci. 25 (12), 6622. doi:10.3390/ijms25126622
Moon, Y., Lee, K. H., Park, J. H., Geum, D., and Kim, K. (2005). Mitochondrial membrane depolarization and the selective death of dopaminergic neurons by rotenone: protective effect of coenzyme Q10. J. Neurochem. 93 (5), 1199–1208. doi:10.1111/j.1471-4159.2005.03112.x
Nicholls, D. G., and Budd, S. L. (2000). Mitochondria and neuronal survival. Physiol. Rev. 80, 315–360. doi:10.1152/physrev.2000.80.1.315
Panee, J., Liu, W., Nakamura, K., and Berry, M. J. (2007). The responses of HT22 cells to the blockade of mitochondrial complexes and potential protective effect of selenium supplementation. Int. J. Biol. Sci. 3 (5), 335–341. doi:10.7150/ijbs.3.335
Polster, B. M., and Fiskum, G. (2004). Mitochondrial mechanisms of neural cell apoptosis. J. Neurochem. 90 (6), 1281–1289. doi:10.1111/j.1471-4159.2004.02572.x
Roy, T., Chatterjee, A., and Swarnakar, S. (2023). Rotenone induced neurodegeneration is mediated via cytoskeleton degradation and necroptosis. Biochim. Biophys. Acta Mol. Cell Res. 1870 (3), 119417. doi:10.1016/j.bbamcr.2022.119417
Sacks, B., Onal, H., Martorana, R., Sehgal, A., Harvey, A., Wastella, C., et al. (2021). Mitochondrial targeted antioxidants, mitoquinone and SKQ1, not vitamin C, mitigate doxorubicin-induced damage in H9c2 myoblast: pretreatment vs. Co-treatment. BMC Pharmacol. Toxicol. 22, 49. doi:10.1186/s40360-021-00518-6
Salucci, S., Burattini, S., Battistelli, M., Buontempo, F., Canonico, B., Martelli, A. M., et al. (2015). Tyrosol prevents apoptosis in irradiated keratinocytes. J. Dermatol. Sci. 80, 61–68. doi:10.1016/j.jdermsci.2015.07.002
Schiff, M., and Rustin, P. (2016). Idebenone in Friedreich ataxia and Leber’s hereditary optic neuropathy: close mechanisms, similar therapy? Brain 139, e39. doi:10.1093/brain/aww085
Sivandzade, F., Prasad, S., Bhalerao, A., and Cucullo, L. (2019). NRF2 and NF-ҚB interplay in cerebrovascular and neurodegenerative disorders: molecular mechanisms and possible therapeutic approaches. Redox Biol. 21, 101059. doi:10.1016/j.redox.2018.11.017
Smith, R. A. J., and Murphy, M. P. (2010). Animal and human studies with the mitochondria-targeted antioxidant MitoQ. Ann. N. Y. Acad. Sci. 1201, 96–103. doi:10.1111/j.1749-6632.2010.05627.x
Solfrizzi, V., Custodero, C., Lozupone, M., Imbimbo, B. P., Valiani, V., Agosti, P., et al. (2017). Relationships of dietary patterns, foods, and Micro- and macronutrients with Alzheimer's disease and late-life cognitive disorders: a systematic review. J. Alzheimers Dis. 59 (3), 815–849. doi:10.3233/JAD-170248
Spinazzi, M., Radaelli, E., Horré, K., Arranz, A. M., Gounko, N. V., Agostinis, P., et al. (2019). PARL deficiency in mouse causes Complex III defects, coenzyme Q depletion, and Leigh-like syndrome. Proc. Natl. Acad. Sci. U. S. A. 116 (1), 277–286. doi:10.1073/pnas.1811938116
Villalaín, J. (2004). Location and interaction of idebenone and mitoquinone in a membrane similar to the inner mitochondrial membrane. Comparison with Ubiquinone 10. Free. Radic. Biol. Med. 222, 211–222. doi:10.1016/j.freeradbiomed.2024.06.017
Wang, Y., Haskell-Ramsay, C., Gallegos, J. L., and Lodge, J. K. (2023). Effects of chronic consumption of specific fruit (berries, cherries and citrus) on cognitive health: a systematic review and meta-analysis of randomised controlled trials. Eur. J. Clin. Nut. 77, 7–22. doi:10.1038/s41430-022-01138-x
Welinder, C., and Ekblad, L. (2011). Coomassie staining as loading control in Western blot analysis. J. Proteome Res. 10 (3), 1416–1419. doi:10.1021/pr1011476
Wenz, T. (2009). PGC-1alpha activation as a therapeutic approach in mitochondrial disease. IUBMB Life 61, 1051–1062. doi:10.1002/iub.261
Wilson, D. M., Cookson, M. R., Van Den Bosch, L., Zetterberg, H., Holtzman, D. M., and Dewachter, I. (2023). Hallmarks of neurodegenerative diseases. Cell 186 (4), 693–714. doi:10.1016/j.cell.2022.12.032
Yang, W., Cui, K., Li, X., Zhao, J., Zeng, Z., Song, R., et al. (2021). Effect of polyphenols on cognitive function: evidence from population-based studies and clinical trials. J. Nutr. Health Aging 25 (10), 1190–1204. doi:10.1007/s12603-021-1685-4
Yousef, A. O. S., A Fahad, A., Abdel Moneim, A. E., Metwally, D. M., El-Khadragy, M. F., and Kassab, R. B. (2019). The neuroprotective role of coenzyme Q10 against lead acetate-induced neurotoxicity is mediated by antioxidant, anti-inflammatory and anti-apoptotic activities. Int. J. Environ. Res. Public Health 16 (16), 2895. doi:10.3390/ijerph16162895
Zhang, H., Greenwood, D. C., Risch, H. A., Bunce, D., Hardie, L. J., and Cade, J. E. (2021). Meat consumption and risk of incident dementia: cohort study of 493,888 UK Biobank participants. Am. J. Clin. Nutr. 114 (1), 175–184. doi:10.1093/ajcn/nqab028
Keywords: coenzyme Q10, Q-Der, oxidative stress, mitochondrial dysfunction, rotenone, neuroprotection, HT22 cells, ATP synthesis
Citation: Micucci M, Gianfanti F, Donati Zeppa S, Annibalini G, Canonico B, Fanelli F, Saltarelli R, Osman R, Montanari M, Lopez D, Nasoni G, Panza G, Bargagni E, Luchetti F, Retini M, Mari M, Zappia G, Stocchi V, Bartolacci A, Burattini S and Battistelli M (2025) Q-Der: a next-generation CoQ10 analogue supercharging neuroprotection by combating oxidative stress and enhancing mitochondrial function. Front. Mol. Biosci. 12:1525103. doi: 10.3389/fmolb.2025.1525103
Received: 08 November 2024; Accepted: 30 January 2025;
Published: 25 February 2025.
Edited by:
Sara Spinelli, University of Messina, ItalyReviewed by:
Shane Austin, The University of the West Indies, Cave Hill, BarbadosCopyright © 2025 Micucci, Gianfanti, Donati Zeppa, Annibalini, Canonico, Fanelli, Saltarelli, Osman, Montanari, Lopez, Nasoni, Panza, Bargagni, Luchetti, Retini, Mari, Zappia, Stocchi, Bartolacci, Burattini and Battistelli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Giovanni Zappia, Z2lvdmFubmkuemFwcGlhQHVuaXJvbWE1Lml0
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.