 Xiao-Li Wei
Xiao-Li Wei Tian-Qi Luo
Tian-Qi Luo Jia-Ning Li
Jia-Ning Li Zhi-Cheng Xue
Zhi-Cheng Xue Yun Wang
Yun Wang You Zhang5
You Zhang5 Chuan Peng
Chuan Peng
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mol. Biosci. , 30 June 2021
Sec. Molecular Diagnostics and Therapeutics
Volume 8 - 2021 | https://doi.org/10.3389/fmolb.2021.691143
This article is part of the Research Topic Biomarkers and Therapeutic Targets of Reprogrammed Tumor Metabolism View all 12 articles
Background: Dysregulation of lipid metabolism plays important roles in the tumorigenesis and progression of gastric cancer (GC). The present study aimed to establish a prognostic model based on the lipid metabolism–related genes in GC patients.
Materials and Methods: Two GC datasets from the Gene Expression Atlas, GSE62254 (n = 300) and GSE26942 (n = 217), were used as training and validation cohorts to establish a risk predictive scoring model. The efficacy of this model was assessed by ROC analysis. The association of the risk predictive scores with patient characteristics and immune cell subtypes was evaluated. A nomogram was constructed based on the risk predictive score model and other prognostic factors.
Results: A risk predictive score model was established based on the expression of 19 lipid metabolism–related genes (LPL, IPMK, PLCB3, CDIPT, PIK3CA, DPM2, PIGZ, GPD2, GPX3, LTC4S, CYP1A2, GALC, SGMS1, SMPD2, SMPD3, FUT6, ST3GAL1, B4GALNT1, and ACADS). The time-dependent ROC analysis revealed that the risk predictive score model was stable and robust. Patients with high risk scores had significantly unfavorable overall survival compared with those with low risk scores in both the training and validation cohorts. A higher risk score was associated with more aggressive features, including a higher tumor grade, a more advanced TNM stage, and diffuse type of Lauren classification of GC. Moreover, distinct immune cell subtypes and signaling pathways were found between the high–risk and low–risk score groups. A nomogram containing patients’ age, tumor stage, adjuvant chemotherapy, and the risk predictive score could accurately predict the survival probability of patients at 1, 3, and 5 years.
Conclusion: A novel 19-gene risk predictive score model was developed based on the lipid metabolism–related genes, which could be a potential prognostic indicator and therapeutic target of GC.
Gastric cancer (GC) is one of the leading causes of cancer-related death worldwide, ranking the third in males and the fifth in females (Bray et al., 2018). Current treatments of GC, including surgery, chemotherapy, and targeted regimens, improve the survival of patients to some extent (Johnston and Beckman, 2019). New prognostic biomarkers remain needed to lower risk, stratify patients, and guide future research for potential new therapeutic targets.
Deregulation of lipid metabolism has a critical role in the promotion of tumorigenesis and tumor progression (Röhrig and Schulze, 2016; Yu et al., 2018; Yang et al., 2020; Esposito et al., 2019). It also participates in the regulation of T cell function, including T cell proliferation and differentiation (Lochner et al., 2015; Raud et al., 2018). Dysregulation of lipid metabolism contributes to various aspects of tumor growth (Lochner et al., 2015; Raud et al., 2018). Lipoproteins, high lipid droplets, and excessive cholesteryl ester storage are hallmarks of aggressiveness of cancers (Yue et al., 2014; Liu et al., 2017). Therefore, targeting deregulated lipid metabolism is a promising strategy for cancer treatment (Liu et al., 2017; Iannelli et al., 2018).
GC progression is closely associated with alterations of lipid metabolism. A low level of serum high-density lipoprotein predicted a high risk of GC development, a high rate of lymphatic and vascular invasion, an advanced nodal metastasis, and a poor prognosis in patients with GC (Guo et al., 2007; Tamura et al., 2012; Nam et al., 2019). Adipocytes and fatty acids fueled metastasis and conferred a poor prognosis of GC (Duan et al., 2016; Tan et al., 2018; Jiang et al., 2019). Various lipid metabolites and genes involved in lipid metabolism also shared some roles in GC tumorigenesis or progression (Abbassi-Ghadi et al., 2013; Tao et al., 2019; Huang et al., 2020; Zhang et al., 2020). For example, adipocytes promoted peritoneal metastasis of GC through reprogramming of fatty acid metabolism mediated by phosphatidylinositol transfer protein, cytoplasmic 1 (PITPNC1) (Tan et al., 2018). Enhanced fatty acid carnitinylation and oxidation mediated by carnitine palmitoyltransferase 1C (CPT1C) promoted proliferative ability of GC (Chen et al., 2020).
The mechanisms of deregulation of lipid metabolism in cancers are complicated, including alteration in pathways involved in de novo lipogenesis, lipid uptake, lipid storage, and lipolysis and generating enhanced synthesis, uptake, consumption, and storage of fatty acids (Liu et al., 2017). However, an overall view of the prognostic value of lipid metabolism–related genes in GC remained to be explored (Liu et al., 2017). Identification of genes associated with clinical outcomes is important for further research in this area. In the current study, lipid metabolism–related gene sets were extracted and analyzed for their prognostic value in patients with GC. A novel lipid metabolism–related gene panel was developed and validated for its capability of predicting patient outcomes.
Two GEO (Gene Expression Omnibus, https://www.ncbi.nlm.nih.gov/geo/) datasets, GSE62254 and GSE26942, were used for analyses. Patients eligible for analyses were as follows: 1) histologically diagnosed with gastric adenocarcinoma, 2) having available mRNA expression data, and having 3) available complete clinical and survival information. There were 300 and 217 patients in the GSE62254 and GSE26942 datasets, respectively. 17 patients were excluded from the GSE26942 dataset due to not meeting the inclusion criteria, including 12 cases of normal tissue, 3 cases of gastric stromal tumor, and 2 cases without available survival information. Finally, the 300-patient cohort from the GSE62254 dataset was used as the training cohort for our risk predictive score model development, and the 200-patient cohort from the GSE26942 dataset was used as the validation cohort. The risk predictive score was also validated in another two public GC datasets, including the TCGA dataset (n = 350) and GSE84437 dataset (n = 432).
The normalized gene expression data of the GSE62254 dataset were downloaded from GEO. Prognosis relevant genes from lipid metabolism–related gene sets were identified using the “survival” package. All the lipid metabolism–related genes were subjected to the univariate Cox regression model, and 63 genes were identified to be associated with overall survival (OS). The 63 genes were further subjected to the LASSO Cox regression model analysis using the glmnet package, and then 19 genes were selected for construction of the risk prognostic scoring system. Calculation of risk scores was performed using the generated coefficients and corresponding expression. According to the risk scores, patients were classified into low-risk and high-risk groups with a cut-off value (risk score = −3.793587), which best stratified patients with different OS.
The same model and coefficients in the training cohort were applied in the validation cohort (GSE26942 dataset). The normalized gene expression data of GSE26942 were downloaded. The efficacy of risk score prognostic classification was evaluated by ROC analysis with the timeROC package. The survival analysis was conducted as mentioned above and was also validated with another two gene sets (TCGA GC and GSE84437).
The timeROC package of R software was applied to perform the time-dependent receiver-operating characteristic curve (ROC) analysis. Survival analysis with Kaplan–Meier plots and the log-rank test, and the univariate and multivariate Cox hazard model were performed. GO and KEGG functional enrichment analyses were conducted through the R package clusterProfiler.
The expression matrices of GSE62254 and GSE26942 datasets were uploaded to CIBERSORT to determine the tumor-infiltrating immune cell fractions, which were calculated according to the LM22 signature with 1,000 permutations, as described previously (Newman et al., 2015). The genes and the immune cell markers are listed in Supplementary File 1. The immune cell subtype fractions were compared between the low–risk and high–risk score groups.
Independent prognostic factors were identified through univariate and multivariate Cox regression analysis. The independent prognostic factors were used to construct the prognostic nomogram, which assessed the OS probability at 1, 3, and 5 years with the “rms” package in R software.
Student’s t-test, the chi-squared test, and the Mann–Whitney U test were used to examine the differences between groups. Spearman analysis was adopted to assess the correlation between gene expression levels. All calculations were performed with R 3.5.3 software (http://www.R-project.org). A two-tailed p value < 0.05 was considered statistically significant.
Human lipid metabolism–related pathways were downloaded from the KEGG (https://www.kegg.jp/) database. 13 lipid metabolism–related pathways were included for analysis (Supplementary Table S1). The characteristics of patients in the GSE62254 and GSE26942 datasets are listed in Supplementary Table S2. The study procedures are shown in Supplementary Figure S1.
The risk prognostic model was developed using the training dataset (GSE62254). The univariate Cox regression model was used to identify genes with prognostic relevance for overall survival (OS). As a result, 63 genes were found to have statistically significant relevance with OS, and their correlations with each other were validated (Supplementary Figure S2A). The LASSO algorithm was used to reduce overfitting and construct the model. The LASSO algorithm is known to have the following features: it is simple and visible, can reduce variance through reduction of coefficients, and can increase interpretability and decrease overfitting through eliminating irrelevant variables. The risk score was built using the LASSO Cox regression model (Supplementary Figures S2B,S2C). 19 genes were selected for the construction of the risk prognostic scoring system (Supplementary Table S3). The correlations between these genes are shown in Supplementary Figure 2D. The risk score was calculated as follows based on the 19 genes:
Risk score = (0.100 * LPL) + (−0.374 * IPMK) + (−0.122 * PLCB3) + (0.311 * CDIPT) + (0.146 * PIK3CA) + (−0.263 * DPM2) + (0.310 * PIGZ) + (−0.594 * GPD2) + (0.043 * GPX3) + (0.077 * LTC4S) + (−0.782 * CYP1A2) + (−0.102 * GALC) + (−0.189 * SGMS1) + (−0.061 * SMPD2) + (−0.184 * SMPD3) + (−0.081 * FUT6) + (0.130 * ST3GAL1) + (0.549 * B4GALNT1) + (−0.040 * ACADS).
The GSE26942 dataset was adopted as the validation dataset. With a cut-off value of −3.793587, which stratified patients into two groups with the largest OS difference, patients were classified into low-risk and high-risk groups. The difference between the low-risk and high-risk groups in OS was statistically significant in the training dataset, the validation dataset, and both datasets combined. Kaplan–Meier curves are displayed in Figures 1A–C. Our constructed risk score also had significant prognostic relevance when validated with another two gene sets, TCGA GC and GSE84437 (Supplementary Figure S3). Subgroup analyses of Kaplan–Meier curves stratified by adjuvant chemotherapy (no/yes) and TNM stage (I + II/III + IV) in the combined dataset are displayed in Supplementary Figure S4. Time-dependent ROC analysis for the risk prognostic model at 1, 3, and 5 years is shown in Figures 1D–F. The area under the curve (AUC) was, respectively, 0.74 (95% CI: 0.67–0.82), 0.78 (95% CI: 0.73–0.83), and 0.78 (95% CI: 0.73–0.83) at 1, 3, and 5 years in the training dataset. The AUC was, respectively, 0.61 (95% CI: 0.50–0.72), 0.60 (95% CI: 0.51–0.68), and 0.63 (95% CI: 0.53–0.73) at 1, 3, and 5 years in the validation dataset and 0.69 (95% CI: 0.63–0.75), 0.71 (95% CI: 0.67–0.76), and 0.73 (95% CI: 0.68–0.77) at 1, 3, and 5 years in the combined dataset.
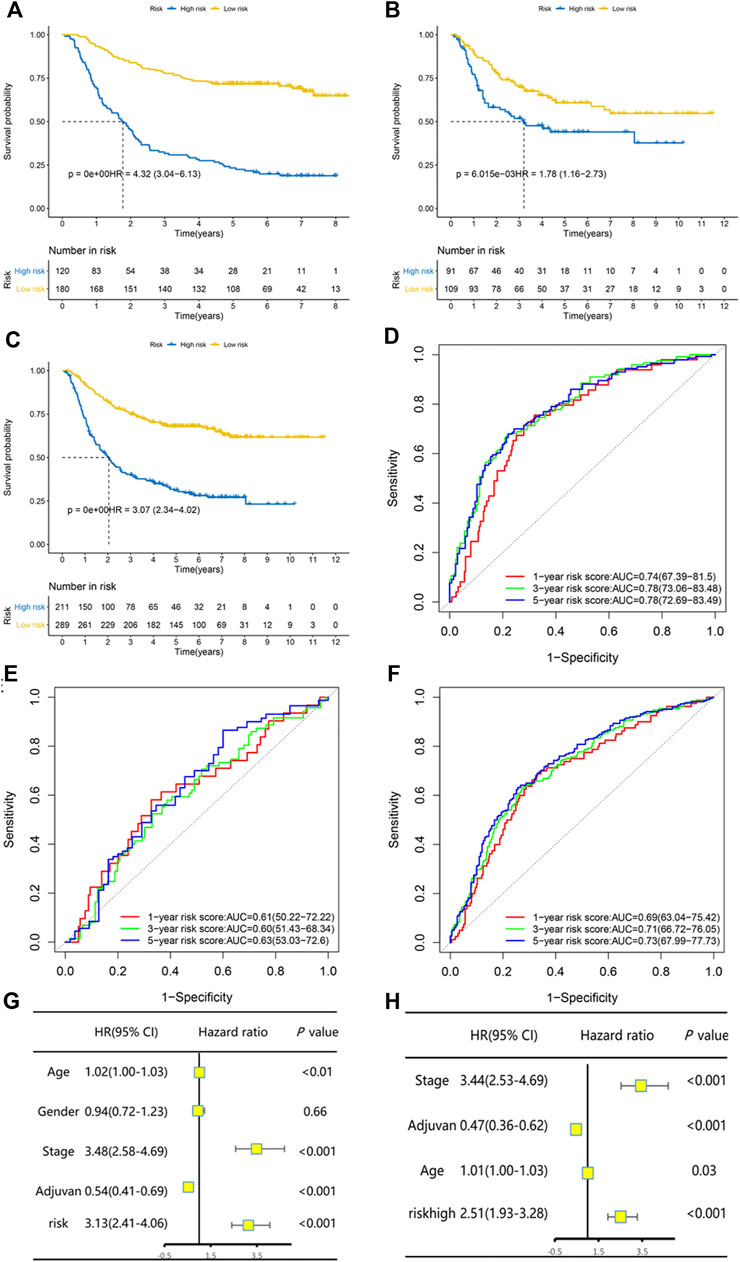
FIGURE 1. The risk predictive score model had high efficacy of prediction in the training set, the validation set, and the combination of both datasets. Kaplan–Meier curves of overall survival stratified by risk score (low/high) in the training set (A), validation set (B), and both datasets (C). Time-dependent ROC analysis for the risk predictive model at 1, 3, and 5 years in the training set (D), validation set (E), and both datasets (F). Univariate and multivariate Cox regression analysis for overall survival in both datasets (G–H).
Univariate and multivariate Cox regression analysis in the combined dataset showed that the risk prognostic score model was an independent and significant prognostic factor for OS (Figures 1G,H). Patients with a high risk score had significantly worse OS (HR and 95% CI: 2.51 [1.93–3.28]) after being adjusted by other independent prognostic factors. The continuous patient risk score, survival state, and expression of the 19 genes of both datasets are shown in Supplementary Figure S5.
We compared the risk score between patients with different clinical characteristics. The results showed that the risk score between patients with different age (<60 or ≥60 years) was comparative (p = 0.11), so it was between males and females (p = 0.84). Tumors with more aggressive features generally had a higher risk score than those with less aggressive features. In particular, patients with a higher grade (G3) tumor had a higher risk score than those with a lower grade (G1/G2) tumor (p < 0.001, Figure 2A). Patients in stage Ⅲ/Ⅳ had a higher risk score than those in stage I/II (p < 0.001, Figure 2B). In terms of Lauren classification, diffused tumors had a higher risk score compared with the intestinal tumors (p < 0.001, Figure 2C). In addition, tumors located in the gastric body had a higher risk score than those located in the gastric antrum (p = 0.02, Figure 2D).
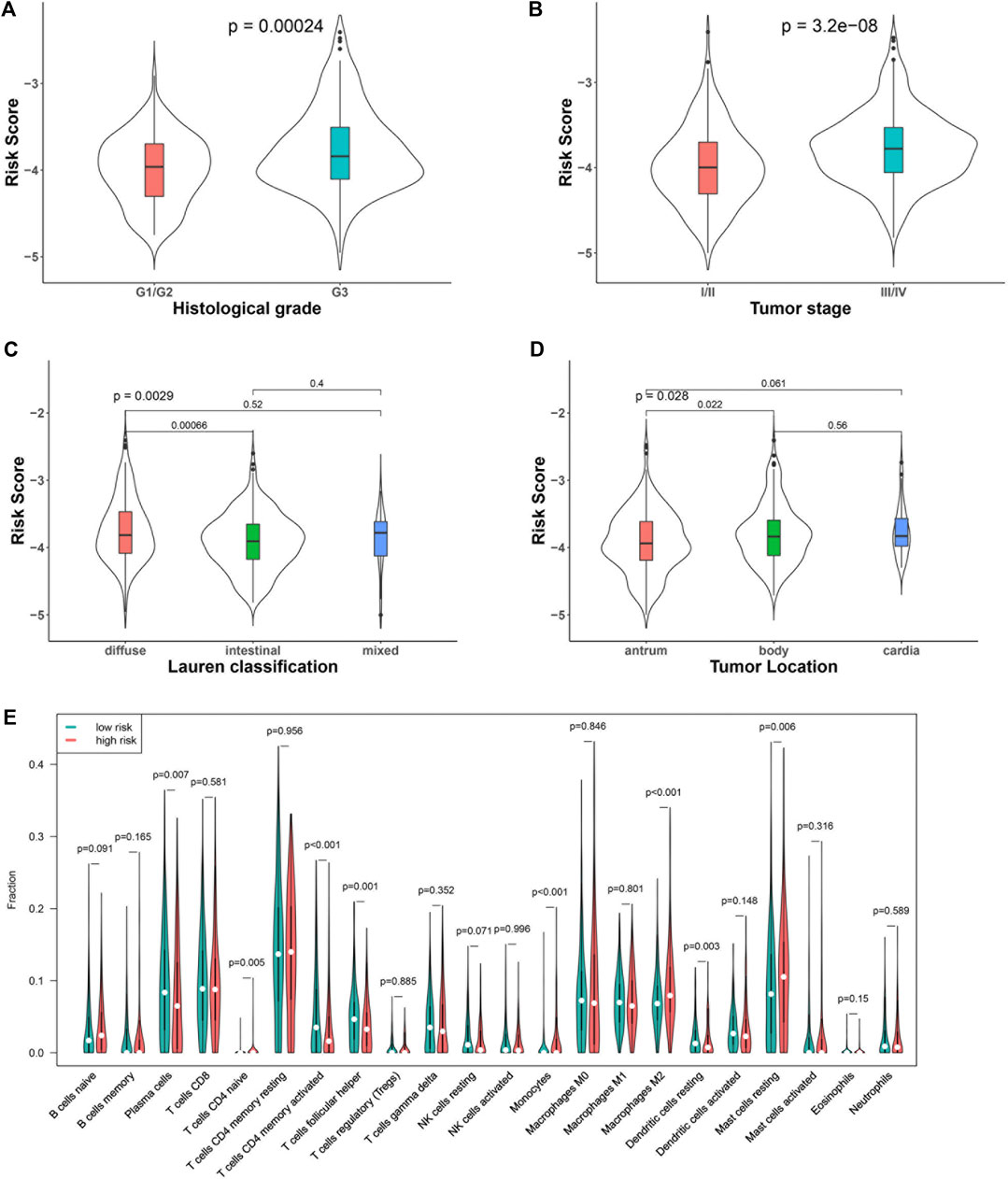
FIGURE 2. Association of the risk score with clinical characteristics and immune cells: tumor grade (A), stage (B), Lauren classification (C), tumor location (D), and immune cell subtypes (E).
We analyzed the percentage of 22 immune cell subtypes in the tumors of both datasets and compared their level between patients with low and high risk scores (Figure 2E). The levels of some immune cells, including plasma cells (p = 0.007), activated CD4 memory cells (p < 0.001), follicular helper T cells (p = 0.001), and resting dendritic cells, were significantly lower in the high–risk score group (p = 0.003), while some immune cells, including naïve CD4 T cells (p = 0.005), monocytes (p < 0.001), M2 macrophages (p < 0.001), and resting mast cells, were higher in the high–risk score group (p = 0.006).
Functional analysis of differentially expressed genes was performed with KEGG and GO functional enrichment analyses. The top 10 GO genes were found to be associated with the biological process (BP), cellular component (CC), and molecular function (MF) (Figure 3A). These genes are associated with positive regulation of cell development, focal adhesion, cell-substrate adherens junction, extracellular matrix structural constituent, growth factor binding, etc. The top 20 enriched pathways are shown in Figure 3B, with the focal adhesion signaling pathway as the most significantly differently expressed pathway. Some other cancer-related pathways, including the Wnt signaling pathway, PI3K–Akt signaling pathway, and MAPK signaling pathway, were also significantly enriched in the high–risk score group.
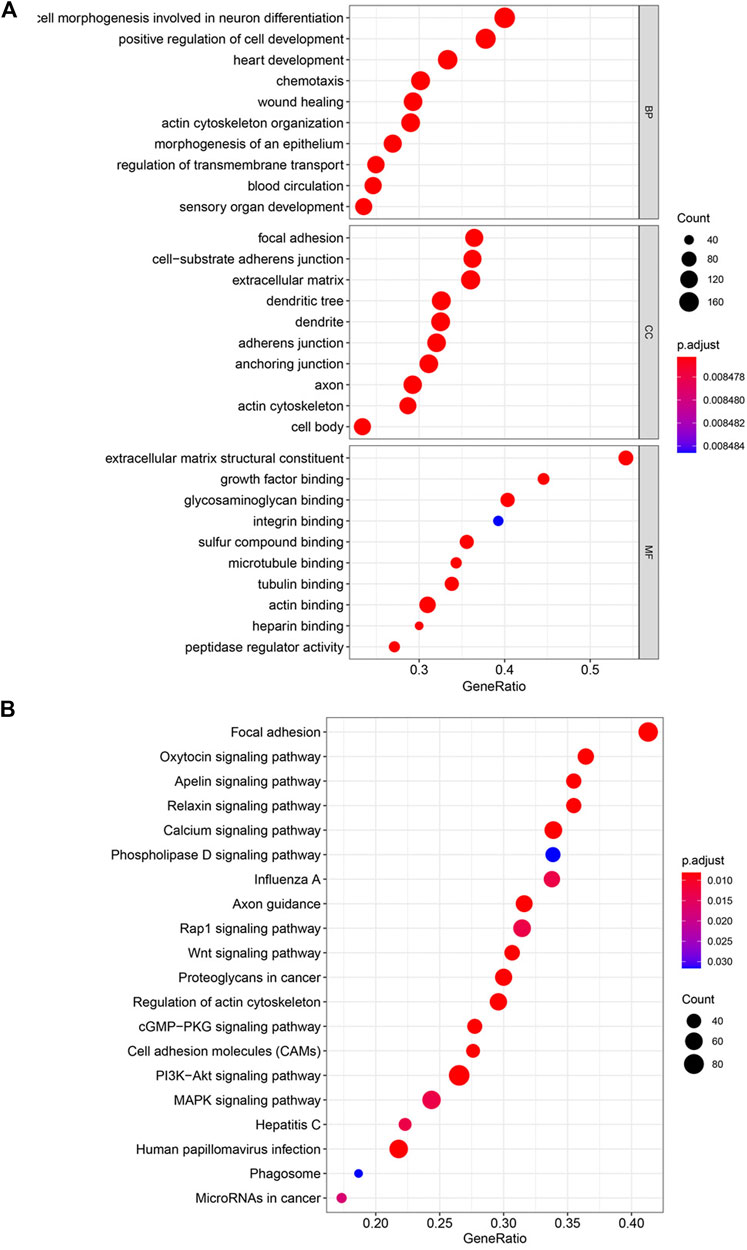
FIGURE 3. Enriched pathways by risk score with KEGG and GO functional enrichment analyses. (A) Top 10 GO terms in the biological process (BP), cellular component (CC), and molecular function (MF). (B) Top 20 enriched pathways in KEGG.
Factors identified by univariate and multivariate Cox regression analysis as independent and significant prognostic factors were applied in the construction of a nomogram model (Figure 4A). As shown in Figures 1G,H, those factors included patients’ age, tumor stage, adjuvant chemotherapy, and the risk prognostic score. The predictive accuracy of the nomogram at 1 year (AUC = 0.79, Figure 4B), 3 years (AUC = 0.82, Figure 4C), and 5 years (AUC = 0.81, Figure 4D) was calculated and assessed by ROC analysis. The comparisons between the 1-, 3-, and 5-year nomogram models and the ideal model are shown in Figures 4E–G that displayed consistent indices and indicated relatively high accuracy of the nomogram models. The decision curve analysis (DCA) showed high predicting efficiency of the nomogram for the 3- and 5-year overall survival in the training dataset, the validation dataset, and the combination of both datasets (Supplementary Figures S6A–S6F).
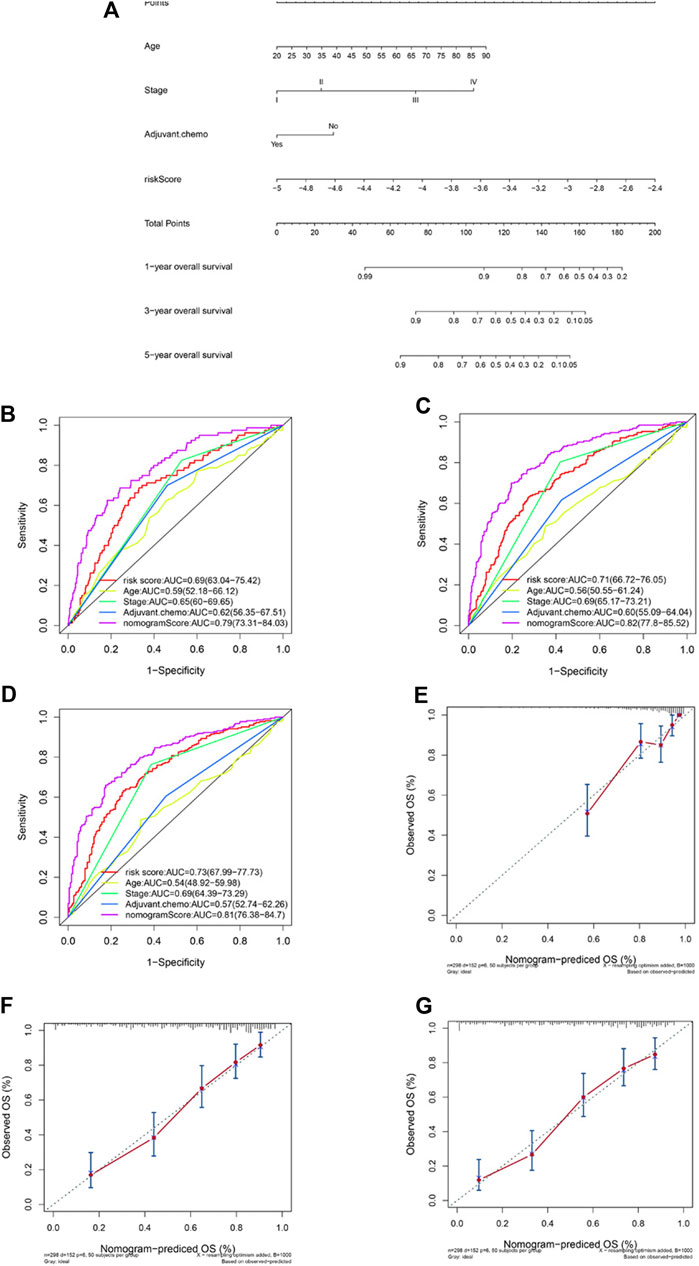
FIGURE 4. Construction of a nomogram based on the risk predictive score and other prognostic factors. Nomogram constructed based on the risk score and three other factors (A). ROC analysis comparing the AUCs at survival of 1 year (B), 3 years (C), and 5 years (D) among the nomogram, risk score model, age, stage, and adjuvant chemotherapy. Comparisons between the predicted overall survival of 1-year (E), 3-year (F), and 5-year (G) nomogram models and the ideal model.
GC has long been recognized as a recalcitrant cancer for its high incidence, high mortality, aggressive behavior, refractory traits, and poor prognosis (Van Cutsem et al., 2016). Identification of genetic factors that drive tumor progression and contribute to unfavorable outcomes is of key importance, for both improving patient care and developing potential therapeutics. As one of the most important basic metabolites, lipid has been demonstrated to be involved in the development and progression of malignancies in recent years, including GC (Huang et al., 2020). Although some studies have suggested important roles of lipid metabolites and lipid metabolism–related genes in GC (Huang et al., 2020), no reports have given an overall view of the prognostic value of lipid metabolism–related genes in GC.
In this study, we develop a novel prognostic scoring model based on the expression of lipid metabolism–related genes in gastric cancer. We used independent datasets from GEO to construct and validate the risk predictive scoring system containing 19 lipid metabolism–related genes. This scoring system was demonstrated to be efficient in predicting patient survival by ROC analysis. Patients had a significant and remarkable difference of OS between the high–risk and low–risk score groups. We further generated a nomogram integrating the risk predictive scoring system and three other prognostic factors (patients’ age, TNM stage, and adjuvant chemotherapy) that improved the efficiency of prognostic value of the nomogram and accurately predicted the 1-, 3-, and 5-year OS of GC patients in the GEO datasets.
The risk predictive score calculated with our scoring system was significantly associated with the aggressiveness of GC. Patients with a higher grade tumor and in an advanced stage were shown to have a higher risk score, suggesting that dysregulation of lipid metabolism not just was associated with cancer progression in GC but also served as a driving factor for the aggressiveness of GC. Some of the genes included in our risk scoring system had been found to be involved in cancers, such as lipoprotein lipase (LPL) (Zaidi et al., 2013), phosphate 3-kinase catalytic subunit alpha (PIK3CA) (Arafeh and Samuels, 2019), mitochondrial glycerol-3-phosphate dehydrogenase (GPD2) (Singh, 2014), leukotriene C4 synthase (LTC4S) (Halvorsen et al., 2014), galactosylceramidase (GALC) (Halvorsen et al., 2014), sphingomyelin phosphodiesterase 3 (SMPD3) (Singh et al., 2014), fucosyltransferase 6 (FUT6) (Singh et al., 2014), and β-galactoside α-2,3-sialyltransferase-1 (ST3Gal1) (Wu et al., 2018). Except for PIK3CA, which was found to be frequently altered in GC and was associated with unfavorable prognosis (Cancer Genome Atlas Research Network, 2014; Kim et al., 2017), although most of these genes were less studied, studies have suggested some roles of them in GC. LPL, which encodes lipoprotein lipase, a key enzyme in triglyceride metabolism, was reported to promote the progression of GC (Chang et al., 2017). Phospholipase C beta 3 (PLCB3), which encodes a member of the phosphoinositide phospholipase C beta enzyme family, was found to be one of the critical altered genes involved in aristolochic acid–induced gastric benign or malignant tumors (Wang et al., 2020). The polymorphism of cytochrome P450 family 1 subfamily A member 2 (CYP1A2) was repeatedly found to be associated with GC risk (Xue et al., 2014) and could modulate susceptibility to GC in patients with Helicobacter pylori infection (Ghoshal et al., 2014). Our study presented evidence for the prognostic value of these genes in GC, demonstrating their potential to be targets for anti-GC therapeutic research and development in the future.
Interestingly, the present study also revealed that patients with high and low risk scores had distinct features in tumor-infiltrating immune cells. Patients with high risk scores had significantly reduced number of plasma cells, activated CD4 memory cells, follicular helper T cells, and resting dendritic cells and increased number of naïve CD4 T cells, monocytes, M2 macrophages, and resting mast cells. Activated CD4 memory cells were associated with favorable outcomes in patients with cervical cancer (Ju et al., 2020) and favorable outcomes after radiotherapy in patients with multiple cancers (Ju et al., 2020; Wen et al., 2020). In gastric cancer, patients with low risk scores had increased number of activated CD4 memory cells and had superior prognosis (Zhao et al., 2020). Dendritic cells are specialized antigen-presenting cells which are key to the initiation of immune responses, including anti-tumor immune responses (Wculek et al., 2020). The increased number of dendritic cells induced by neoadjuvant chemotherapy was reported to be related to improved survival in GC (Hu et al., 2014). Naïve CD4 T cells were developed to form Treg cells in the tumor microenvironment and predicted poor prognosis in breast cancer (Su et al., 2017). M2 macrophages are a well-known tumor-promoting immunosuppressive cell type, and they have been proposed as a therapeutic target in GC (Gambardella et al., 2020). Immunotherapy has been established as a novel treatment in GC, but as monotherapy, PD-1 antibodies have limited benefit because the majority of patients do not respond (Xie et al., 2021). Novel combination options with immunotherapy are in great need in GC. Lipid metabolism not only impacts the proliferation and migration of tumor cells but also shapes the immuno-microenvironment by affecting the recruitment and function of tumor-infiltrating immune cells (He et al., 2021). In our study, patients with high risk scores had an immunosuppressive tumor microenvironment, indicating a possible role of treatments targeting lipid metabolism–related genes with immunotherapy in GC.
The major limitation of the present study was the lack of validation in larger patient cohorts from multicenter real-world clinical practice. Thus, the risk predictive score was still far from being able to be used in clinical practice. Another important limitation was that most of the genes used to construct this risk predictive score model were scarcely investigated in cancers. In addition, we did not perform the basic experiment to validate their roles and related mechanisms in GC cells. The biological mechanism was unclear and needed further experimental validation. However, as a prognostic risk score, our model was repeatedly validated and achieved consensus results, so the conclusions of our study are still convincing despite the lack of experimental validation of each gene’s role in GC.
In the present study, a novel lipid metabolism–related gene-based risk predictive score model was constructed and validated in datasets of patients with GC. This risk predictive scoring system could efficiently predict patient outcomes and had significant correlation with immune cell subtypes. A nomogram containing the risk score was generated, and it improved the prognostic predictive value of the current TNM staging system. This study will be helpful in biomarker and therapeutics development for GC patients.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
The study followed the Declaration of Helsinki and was approved by the Clinical Research Ethics Committee of Sun Yat-sen University Cancer Center. Because of the retrospective nature of the study, patient consent for inclusion was waived.
CP, Y-BC, and X-LW designed the study. T-QL, J-NL, and Z-CX retrieved the data and conducted analysis. T-QL, YW, and YZ drew the tables and figures. X-LW, T-QL, and J-NL wrote the manuscript. All authors read and approved the manuscript.
This study was supported by the Guangdong Basic and Applied Basic Research Foundation (2019A1515110171).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors would like to thank the authors who submitted the related data on the GEO website.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmolb.2021.691143/full#supplementary-material
Supplementary Figure 1 | Flowchart of the study. Two GEO datasets, GSE62254 and GSE26942, were used as the training and validation datasets for the risk predictive score model construction. Further comparisons and establishment of a nomogram based on the risk scores were conducted.
Supplementary Figure 2 | Construction of a risk predictive score model based on lipid metabolism–related genes. 63 prognostic relevant genes in lipid metabolism–related pathways were screened (A). The risk predictive score system was constructed using the LASSO Cox regression model (B,C). Correlation between the 19 selected genes (D).
Supplementary Figure 3 | Kaplan–Meier curves of overall survival stratified by risk score (low/high) in another two datasets: TCGA GC dataset (A) and GSE84437 dataset (B).
Supplementary Figure 4 | Subgroup analyses of Kaplan–Meier curves for overall survival stratified by adjuvant chemotherapy (no/yes) and TNM stage (I + II/III + IV) in the combined dataset. Adjuvant chemotherapy—no (A), adjuvant chemotherapy—yes (B), TNM stage—I + II (C), and TNM stage—III + IV (D).
Supplementary Figure 5 | Expression of 19 genes (A), continuous patient risk score (B), and survival state (C) in both datasets.
Supplementary Figure 6 | Decision curve analysis (DCA) for 3-year OS and 5-year OS. DCA for 3-year OS in the training dataset (A), validation dataset (B), and both datasets (C); DCA for 5-year OS in the training dataset (D), validation dataset (E), and both datasets (F).
Abbassi-Ghadi, N., Kumar, S., Huang, J., Goldin, R., Takats, Z., and Hanna, G. B. (2013). Metabolomic Profiling of Oesophago-Gastric Cancer: a Systematic Review. Eur. J. Cancer 49, 3625–3637. doi:10.1016/j.ejca.2013.07.004
Arafeh, R., and Samuels, Y. (2019). PIK3CA in Cancer: The Past 30 Years. Semin. Cancer Biol. 59, 36–49. doi:10.1016/j.semcancer.2019.02.002
Bray, F., Ferlay, J., Soerjomataram, I., Siegel, R. L., Torre, L. A., and Jemal, A. (2018). Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA: A Cancer J. Clinicians 68, 394–424. doi:10.3322/caac.21492
Chang, W.-C., Huang, S.-F., Lee, Y.-M., Lai, H.-C., Cheng, B.-H., Cheng, W.-C., et al. (2017). Cholesterol Import and Steroidogenesis Are Biosignatures for Gastric Cancer Patient Survival. Oncotarget 8, 692–704. doi:10.18632/oncotarget.13524
Chen, T., Wu, G., Hu, H., and Wu, C. (2020). Enhanced Fatty Acid Oxidation Mediated by CPT1C Promotes Gastric Cancer Progression. J. Gastrointest. Oncol. 11, 695–707. doi:10.21037/jgo-20-157
Cancer Genome Atlas Research Network 2014). Comprehensive Molecular Characterization of Gastric Adenocarcinoma. Nature. 513, 202–209.doi:10.1038/nature13480
Duan, J., Sun, L., Huang, H., Wu, Z., Wang, L., and Liao, W. (2016). Overexpression of Fatty Acid Synthase Predicts a Poor Prognosis for Human Gastric Cancer. Mol. Med. Rep. 13, 3027–3035. doi:10.3892/mmr.2016.4902
Esposito, M., Mondal, N., Greco, T. M., Wei, Y., Spadazzi, C., Lin, S.-C., et al. (2019). Bone Vascular Niche E-Selectin Induces Mesenchymal-Epithelial Transition and Wnt Activation in Cancer Cells to Promote Bone Metastasis. Nat. Cel Biol 21, 627–639. doi:10.1038/s41556-019-0309-2
Gambardella, V., Castillo, J., Tarazona, N., Gimeno-Valiente, F., Martínez-Ciarpaglini, C., Cabeza-Segura, M., et al. (2020). The Role of Tumor-Associated Macrophages in Gastric Cancer Development and Their Potential as a Therapeutic Target. Cancer Treat. Rev. 86, 102015. doi:10.1016/j.ctrv.2020.102015
Ghoshal, U., Tripathi, S., Kumar, S., Mittal, B., Chourasia, D., Kumari, N., et al. (2014). Genetic Polymorphism of Cytochrome P450 (CYP) 1A1, CYP1A2, and CYP2E1 Genes Modulate Susceptibility to Gastric Cancer in Patients with Helicobacter pylori Infection. Gastric Cancer 17, 226–234. doi:10.1007/s10120-013-0269-3
Guo, E., Chen, L., Xie, Q., Chen, J., Tang, Z., and Wu, Y. (2007). Serum HDL-C as a Potential Biomarker for Nodal Stages in Gastric Cancer. Ann. Surg. Oncol. 14, 2528–2534. doi:10.1245/s10434-007-9401-0
Halvorsen, A. R., Helland, A., Fleischer, T., Haug, K. M., Grenaker Alnæs, G. I., Nebdal, D., et al. (2014). Differential DNA Methylation Analysis of Breast Cancer Reveals the Impact of Immune Signaling in Radiation Therapy. Int. J. Cancer 135, 2085–2095. doi:10.1002/ijc.28862
He, S., Cai, T., Yuan, J., Zheng, X., and Yang, W. (2021). Lipid Metabolism in Tumor-Infiltrating T Cells. Adv. Exp. Med. Biol. 1316, 149–167. doi:10.1007/978-981-33-6785-2_10
Hu, M., Li, K., Maskey, N., Xu, Z., Peng, C., Wang, B., et al. (2014). Decreased Intratumoral Foxp3 Tregs and Increased Dendritic Cell Density by Neoadjuvant Chemotherapy Associated with Favorable Prognosis in Advanced Gastric Cancer. Int. J. Clin. Exp. Pathol. 7, 4685–4694.
Huang, S., Guo, Y., Li, Z., Zhang, Y., Zhou, T., You, W., et al. (2020). A Systematic Review of Metabolomic Profiling of Gastric Cancer and Esophageal Cancer. Cancer Biol. Med. 17, 181–198. doi:10.20892/j.issn.2095-3941.2019.0348
Iannelli, F., Lombardi, R., Milone, M. R., Pucci, B., De Rienzo, S., Budillon, A., et al. (2018). Targeting Mevalonate Pathway in Cancer Treatment: Repurposing of Statins. Pra 13, 184–200. doi:10.2174/1574892812666171129141211
Jiang, M., Wu, N., Xu, B., Chu, Y., Li, X., Su, S., et al. (2019). Fatty Acid-Induced CD36 Expression via O-GlcNAcylation Drives Gastric Cancer Metastasis. Theranostics 9, 5359–5373. doi:10.7150/thno.34024
Johnston, F. M., and Beckman, M. (2019). Updates on Management of Gastric Cancer. Curr. Oncol. Rep. 21, 67. doi:10.1007/s11912-019-0820-4
Ju, M., Qi, A., Bi, J., Zhao, L., Jiang, L., Zhang, Q., et al. (2020). A Five‐mRNA Signature Associated with post‐translational Modifications Can Better Predict Recurrence and Survival in Cervical Cancer. J. Cel Mol Med 24, 6283–6297. doi:10.1111/jcmm.15270
Kim, J.-W., Lee, H. S., Nam, K. H., Ahn, S., Kim, J. W., Ahn, S.-H., et al. (2017). PIK3CA Mutations Are Associated with Increased Tumor Aggressiveness and Akt Activation in Gastric Cancer. Oncotarget 8, 90948–90958. doi:10.18632/oncotarget.18770
Liu, Q., Luo, Q., Halim, A., and Song, G. (2017). Targeting Lipid Metabolism of Cancer Cells: A Promising Therapeutic Strategy for Cancer. Cancer Lett. 401, 39–45. doi:10.1016/j.canlet.2017.05.002
Lochner, M., Berod, L., and Sparwasser, T. (2015). Fatty Acid Metabolism in the Regulation of T Cell Function. Trends Immunol. 36, 81–91. doi:10.1016/j.it.2014.12.005
Nam, S. Y., Park, B. J., Nam, J. H., and Kook, M.-C. (2019). Effect of Helicobacter pylori Eradication and High-Density Lipoprotein on the Risk of De Novo Gastric Cancer Development. Gastrointest. Endosc. 90, 448–456. doi:10.1016/j.gie.2019.04.232
Newman, A. M., Liu, C. L., Green, M. R., Gentles, A. J., Feng, W., Xu, Y., et al. (2015). Robust Enumeration of Cell Subsets from Tissue Expression Profiles. Nat. Methods 12, 453–457. doi:10.1038/nmeth.3337
Raud, B., McGuire, P. J., Jones, R. G., Sparwasser, T., and Berod, L. (2018). Fatty Acid Metabolism in CD8+T Cell Memory: Challenging Current Concepts. Immunol. Rev. 283, 213–231. doi:10.1111/imr.12655
Röhrig, F., and Schulze, A. (2016). The Multifaceted Roles of Fatty Acid Synthesis in Cancer. Nat. Rev. Cancer 16, 732–749. doi:10.1038/nrc.2016.89
Singh, G. (2014). Mitochondrial FAD-Linked Glycerol-3-Phosphate Dehydrogenase: A Target for Cancer Therapeutics. Pharmaceuticals 7, 192–206. doi:10.3390/ph7020192
Singh, R., Pochampally, R., Watabe, K., Lu, Z., and Mo, Y.-Y. (2014). Exosome-mediated Transfer of miR-10b Promotes Cell Invasion in Breast Cancer. Mol. Cancer 13, 256. doi:10.1186/1476-4598-13-256
Su, S., Liao, J., Liu, J., Huang, D., He, C., Chen, F., et al. (2017). Blocking the Recruitment of Naive CD4+ T Cells Reverses Immunosuppression in Breast Cancer. Cell Res 27, 461–482. doi:10.1038/cr.2017.34
Tamura, T., Inagawa, S., Hisakura, K., Enomoto, T., and Ohkohchi, N. (2012). Evaluation of Serum High-Density Lipoprotein Cholesterol Levels as a Prognostic Factor in Gastric Cancer Patients. J. Gastroenterol. Hepatol. 27, 1635–1640. doi:10.1111/j.1440-1746.2012.07189.x
Tan, Y., Lin, K., Zhao, Y., Wu, Q., Chen, D., Wang, J., et al. (2018). Adipocytes Fuel Gastric Cancer Omental Metastasis via PITPNC1-Mediated Fatty Acid Metabolic Reprogramming. Theranostics 8, 5452–5468. doi:10.7150/thno.28219
Tao, L., Yu, H., Liang, R., Jia, R., Wang, J., Jiang, K., et al. (2019). Rev-erbα Inhibits Proliferation by Reducing Glycolytic Flux and Pentose Phosphate Pathway in Human Gastric Cancer Cells. Oncogenesis 8, 57. doi:10.1038/s41389-019-0168-5
Van Cutsem, E., Sagaert, X., Topal, B., Haustermans, K., and Prenen, H. (2016). Gastric Cancer. The Lancet 388, 2654–2664. doi:10.1016/s0140-6736(16)30354-3
Wang, L., Li, C., Tian, J., Liu, J., Zhao, Y., Yi, Y., et al. (2020). Genome-wide Transcriptional Analysis of Aristolochia Manshuriensis Induced Gastric Carcinoma. Pharm. Biol. 58, 98–106. doi:10.1080/13880209.2019.1710219
Wculek, S. K., Cueto, F. J., Mujal, A. M., Melero, I., Krummel, M. F., and Sancho, D. (2020). Dendritic Cells in Cancer Immunology and Immunotherapy. Nat. Rev. Immunol. 20, 7–24. doi:10.1038/s41577-019-0210-z
Wen, P., Gao, Y., Chen, B., Qi, X., Hu, G., Xu, A., et al. (2020). Pan-Cancer Analysis of Radiotherapy Benefits and Immune Infiltration in Multiple Human Cancers. Cancers (Basel) 12. doi:10.3390/cancers12040957
Wu, X., Zhao, J., Ruan, Y., Sun, L., Xu, C., and Jiang, H. (2018). Sialyltransferase ST3GAL1 Promotes Cell Migration, Invasion, and TGF-Β1-Induced EMT and Confers Paclitaxel Resistance in Ovarian Cancer. Cell Death Dis 9, 1102. doi:10.1038/s41419-018-1101-0
Xie, J., Fu, L., and Jin, L. (2021). Immunotherapy of Gastric Cancer: Past, Future Perspective and Challenges. Pathol. - Res. Pract. 218, 153322. doi:10.1016/j.prp.2020.153322
Xue, H., Lu, Y., Xue, Z., Lin, B., Chen, J., Tang, F., et al. (2014). The Effect of CYP1A1 and CYP1A2 Polymorphisms on Gastric Cancer Risk Among Different Ethnicities: a Systematic Review and Meta-Analysis. Tumor Biol. 35, 4741–4756. doi:10.1007/s13277-014-1620-y
Yang, M., Jiang, Z., Yao, G., Wang, Z., Sun, J., Qin, H., et al. (2020). GALC Triggers Tumorigenicity of Colorectal Cancer via Senescent Fibroblasts. Front. Oncol. 10, 380. doi:10.3389/fonc.2020.00380
Yu, X. H., Ren, X. H., Liang, X. H., and Tang, Y. L. (2018). Roles of Fatty Acid Metabolism in Tumourigenesis: Beyond Providing Nutrition (Review). Mol. Med. Rep. 18, 5307–5316. doi:10.3892/mmr.2018.9577
Yue, S., Li, J., Lee, S.-Y., Lee, H. J., Shao, T., Song, B., et al. (2014). Cholesteryl Ester Accumulation Induced by PTEN Loss and PI3K/AKT Activation Underlies Human Prostate Cancer Aggressiveness. Cel Metab. 19, 393–406. doi:10.1016/j.cmet.2014.01.019
Zaidi, N., Lupien, L., Kuemmerle, N. B., Kinlaw, W. B., Swinnen, J. V., and Smans, K. (2013). Lipogenesis and Lipolysis: the Pathways Exploited by the Cancer Cells to Acquire Fatty Acids. Prog. Lipid Res. 52, 585–589. doi:10.1016/j.plipres.2013.08.005
Zhang, H., Deng, T., Liu, R., Ning, T., Yang, H., Liu, D., et al. (2020). CAF Secreted miR-522 Suppresses Ferroptosis and Promotes Acquired Chemo-Resistance in Gastric Cancer. Mol. Cancer 19, 43. doi:10.1186/s12943-020-01168-8
Keywords: gastric cancer, prognostic model, lipid metabolism, nomogram, gene expression omnibus dataset, gene panel
Citation: Wei X-L, Luo T-Q, Li J-N, Xue Z-C, Wang Y, Zhang Y, Chen Y-B and Peng C (2021) Development and Validation of a Prognostic Classifier Based on Lipid Metabolism–Related Genes in Gastric Cancer. Front. Mol. Biosci. 8:691143. doi: 10.3389/fmolb.2021.691143
Received: 05 April 2021; Accepted: 07 June 2021;
Published: 30 June 2021.
Edited by:
Zhe-Sheng Chen, St. John’s University, United StatesReviewed by:
Yingyan Yu, Shanghai Jiao Tong University, ChinaCopyright © 2021 Wei, Luo, Li, Xue, Wang, Zhang, Chen and Peng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ying-Bo Chen, Y2hlbnliQHN5c3VjYy5vcmcuY24=; Chuan Peng, cGVuZ2NodWFuQHN5c3VjYy5vcmcuY24=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.