
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 26 February 2025
Sec. Biology of Archaea
Volume 16 - 2025 | https://doi.org/10.3389/fmicb.2025.1543464
This article is part of the Research TopicMolecular Biology of Archaea - 2024View all 3 articles
 A-L. Sailer1†
A-L. Sailer1† J. Brendel1†A. Chernev2,3S. König2,3T. Bischler4T. Gräfenhan4H. Urlaub2,3
J. Brendel1†A. Chernev2,3S. König2,3T. Bischler4T. Gräfenhan4H. Urlaub2,3 U. Gophna5
U. Gophna5 A. Marchfelder1*
A. Marchfelder1*CRISPR-Cas is a sophisticated defence system used by bacteria and archaea to fend off invaders. CRISPR-Cas systems vary in their Cas protein composition and have therefore been divided into different classes and types. Type I systems of bacteria have been shown to contain the small protein Cas11 as part of the interference complex known as Cascade. Here we show for the first time that an archaeal CRISPR-Cas type I-B system also contains a homolog of Cas11. The Cas11b protein, encoded by the cas8b gene in Haloferax volcanii, represents the first known case of an internal in-frame translation of an archaeal protein. Translation initiation at an internal methionine of the cas8b open reading frame results in synthesis of Cas11b. Cas11b is required for an effective CRISPR-Cas interference reaction, and in its absence fewer Cascade complexes are formed. Comparison of transcriptomes from wild type and a Cas11b-less strain shows that the depletion of Cas11b also results in differential transcript abundance of many genes, presumably affecting their regulation. Taken together, Cas11b is important for the defence reaction of the type I-B CRISPR-Cas system and seems to play an additional cellular role.
The CRISPR-Cas immune system is one of several systems evolved in bacteria and archaea to defend themselves against invaders (Hille et al., 2018; Koonin et al., 2017; Makarova et al., 2020). In contrast to other defence systems, the CRISPR-Cas system is special because it adapts to new viruses and makes the cell and its daughter cells immune to recurring attacks. Many different versions of the CRISPR-Cas systems have been found and described, however only a few of them have been characterised in detail (Makarova et al., 2020). All CRISPR-Cas systems have in common that they employ Cas proteins and short RNAs (known as crRNAs) for the defence reaction. They all use three stages to fend off the invader: adaptation, expression and interference (Koonin and Makarova, 2022). The adaptation step makes the CRISPR-Cas system more sophisticated than other defence systems since it renders the cell immune against new mobile genetic elements. Upon detection of the invader its DNA is degraded and a piece of it is integrated into the CRISPR locus. The integration of the invader DNA into the genome makes the defence hereditary. In the expression step the CRISPR RNA containing the information from all previous invaders, is transcribed and processed into the invader specific crRNAs, each being able to detect an invader during a recurring attack by sequence complementarity. In the last step, the interference step, the invader DNA is recognised by its specific crRNA and immediately degraded. This last step is very efficient ensuring cell survival.
One major difference between the many CRISPR-Cas systems found is the nature of the effector that detects and degrades the invader. In class 2 systems the effector is a single protein like Cas9, while class 1 systems use a multi-protein complex as effector (Makarova et al., 2020). There are three different class 1 types: type I, III and IV with type I systems occurring most frequently (Makarova et al., 2020). The major types are further subdivided into subtypes based on the nature of their Cas proteins. For type I these are subtypes I-A to I-G (Makarova et al., 2020), with I-B being the most abundant subtype. The effector complex of the type I systems is termed Cascade (CRISPR-associated complex for antiviral defence).
CRISPR-Cas systems are more prevalent in archaea, with about 80% encoding this system, compared to only 40% of bacteria (Makarova et al., 2020). The archaeon Haloferax volcanii has a single CRISPR-Cas system of type I-B, which has been studied in detail (Brendel et al., 2014; Fischer et al., 2012; Maier et al., 2017; Maier et al., 2013; Maier et al., 2019; Maier et al., 2015; Miezner et al., 2023). This system served as the basis for developing an efficient CRISPRi tool (Gophna et al., 2017; Schwarz et al., 2022; Stachler and Marchfelder, 2016). Its Cascade complex consists of Cas5 and Cas7 as well as a loosely associated Cas8b. Cas6b was copurified with the complex, but is not required for the interference, rendering it a non-integral part of the complex (Figure 1A) (Brendel et al., 2014; Maier et al., 2015). Type I-A and type I-E Cascade complexes contain an additional small subunit, Cas11, encoded by a separate gene (Figure 1B) (Makarova et al., 2020). Although Cas11 proteins from different subtypes lack significant sequence similarity (Makarova et al., 2020), they share structural features and bind to similar positions in Cas complexes (Hayes et al., 2016; Hu et al., 2022; Jackson et al., 2014; Jore et al., 2011; Lu et al., 2024; Mulepati et al., 2014; Wiedenheft et al., 2011). Cas11 is present in two copies in the type I-E Cascade complex and binds opposite the Cas7 multimers (Figure 1B) (Hayes et al., 2016; Jackson et al., 2014; Jore et al., 2011; Mulepati et al., 2014; Wiedenheft et al., 2011; Zhao et al., 2014). Structural analyses of the type I-A Cascade have revealed that five Cas11 subunits form the inner belly (Hu et al., 2022). However, data reported so far about the different Cas11 proteins suggest that despite their structural similarity they differ in their functional roles in interference (Liu and Doudna, 2020; Reeks et al., 2013; Tan et al., 2022).
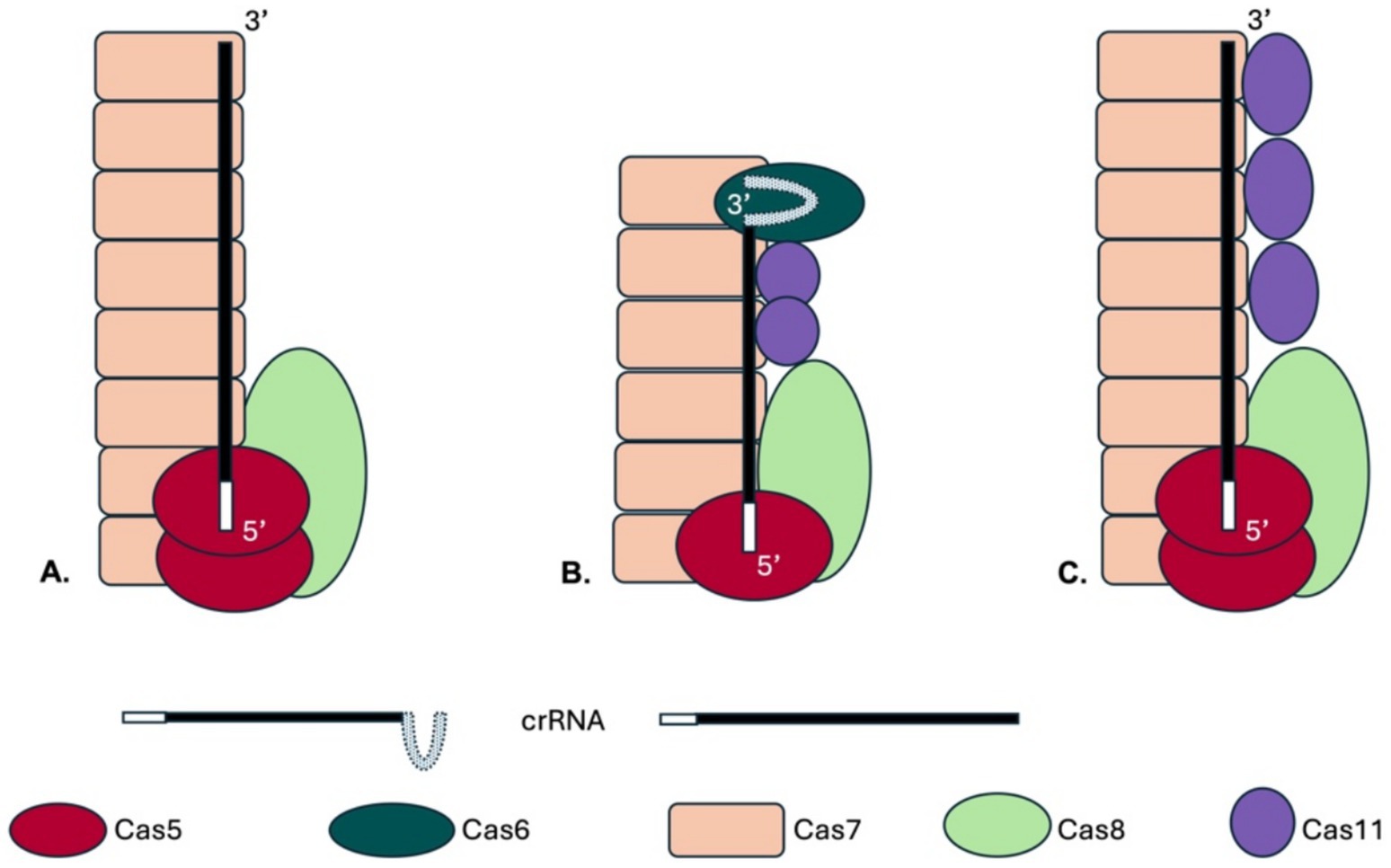
Figure 1. Schematic drawings of Cascade complexes. The type I CRISPR-Cas complexes contain a large subunit (Cas8 or Cas10), Cas7 proteins form the complex’s backbone and Cas5 binds at the crRNA 5′ end. With the exception of type I-F all Cascades contain a small subunit, the Cas11 protein. (A) A potential complex structure of the Haloferax Cascade complex is shown. The structure of the Haloferax Cascade complex has not yet been solved, but the composition of a purified Cascade was determined (Brendel et al., 2014). In this previous work potential Cas11b and Cas8b peptides could not be distinguished therefore no Cas11b is shown. The genomic location of the cas genes and CRISPR loci of the H. volcanii type I-B system is shown in Supplementary Figure S1. (B) The type I-E Cascade complex contains six Cas7 proteins, one Cas5, Cas6 and Cas8 each and two Cas11 proteins (in purple) (Jore et al., 2011; Wiedenheft et al., 2011). (C) A potential Haloferax Cascade structure including the here identified Cas11b is shown, the number of Cas11b proteins was assumed as found in the bacterial type I-B (Lu et al., 2024).
Recently, an internal translation initiation site was discovered in the cas10 gene of a bacterial type I-D system, resulting in synthesis of Cas11 (McBride et al., 2020). Similar internal translation initiation sites were subsequently discovered in the cas8 genes of the bacterial type I-B and I-C systems (Hussain et al., 2023; McBride et al., 2020; O’Brien et al., 2020; Tan et al., 2022). Since all reports so far concentrated on bacterial systems and the archaeal Haloferax type I-B CRISPR-Cas system does not contain a separate gene for a Cas11b protein, we investigated here whether the cas8b gene of H. volcanii (HVO_A0206) encodes a Cas11b protein. Having identified this protein we then studied how this Cas11b contributes to Cascade stability and function and to potential additional cellular functions beyond virus defence.
Strains, plasmids and primers used are listed in Supplementary Table S1. Hfx. volcanii H119 was used as parent strain (wild type strain) for all experiments. H119 is derived from wild type strain DS2 by removal of plasmid pHV2 and deletion of three genes that can be used as selection markers (ΔpyrE2, ΔtrpA, ΔleuB) (Allers et al., 2004). All Hfx. volcanii strains were grown aerobically with shaking (200 rpm) at 45°C in Hv-YPC (rich medium) or Hv-Cab media (minimal medium) with appropriate supplements if carrying any plasmid as described (Allers et al., 2004).
E. coli strains DH5α (Invitrogen, Thermo Fischer Scientific, Waltham, MA, United States) and GM121 (Allers et al., 2010) were grown aerobically at 37°C in 2YT medium (Miller, 1972) supplemented with 75 μg/mL ampicillin for selection.
The generation of plasmid pTA927-cas8b-FLAGN for expression of the Cas8b protein under control of the p.tna promotor carrying an N-terminal 3xFLAG-tag is described in Cass et al. (2015). Plasmid pTA927-cas8b-FLAGC for expression of the Cas8b protein under control of the p.tna promotor carrying a C-terminal 3xFLAG-tag, was generated by amplification of the cas8b gene using the oligonucleotides 5-Csh1-NdeI and 3-Cas8-SmaI and pTA927-cas8b-FLAGN as template, the resulting DNA fragment was digested with NdeI and SmaI and ligated into NdeI and SnaBI digested vector pTA927-FLAGC (Hadjeras et al., 2023), resulting in the final construct pTA927-cas8b-FLAGC. The plasmid for expressing FLAG-Cas8bM545A was generated by performing a mutagenesis PCR using the QuikChange II XL Site-Directed Mutagenesis Kit (Agilent Technologies) with the oligonucleotides 8M545A and 8M545Ar and pTA927-cas8b-FLAGC as template, resulting in the final construct pTA927-cas8bM545A-FLAGC. Plasmid pTA927-cas11b for expression of the Cas11b protein under the control of the p.tna promoter was generated as follows. The cas11b gene was amplified by PCR using oligonucleotides 5-Cas11-NdeI and 3-Cas11-XbaI and pTA927-cas8b-FLAGC as template. The resulting PCR product was cleaved with NdeI and XbaI and cloned into vector pTA927 (also cleaved with NdeI and XbaI) (Allers et al., 2010), resulting in the final construct pTA927-cas11b. To obtain the plasmid for expression of the mutated Cas8b protein (lacking the start codon for Cas11b) and the Cas11b from the same plasmid, the cas11b gene under the control of the p.tna promoter was amplified by PCR using the oligonucleotides 5-ptna-BamHI and 3-cas11-XbaI and pTA927-cas11b as template and cloned into the plasmid pTA927-cas8bM545A-FLAGC via the restriction sites BamHI and XbaI, resulting in the final construct pTA927-cas8bM545A-FLAGC-cas11b. The plasmid for generating the cas8bM545A mutant strain was generated as follows. The mutated cas8bM545A gene was amplified by PCR using the oligonucleotides 5-cas8-fw and 3-cas8 rev and pTA927-cas8bM545A-FLAGC as template. The existing plasmid pTA131-up.do (Csh1) (Cass et al., 2015), which contains the app. 500–550 base pairs upstream and downstream regions of the cas8b gene, was used as template in a PCR reaction using the oligonucleotides IP cas8 rev and IP cas8 fw to amplify the plasmid in an inverted PCR. The mutated cas8bM545A insert was then ligated into that vector, resulting in the final construct pTA131-up.cas8bM545A.do.
To generate the cas8bM545A mutant strain a ∆cas8b deletion strain (Cass et al., 2015; Stoll, 2013) was transformed with plasmid pTA131-up.cas8bM544A.do to integrate the modified gene into the cas8b gene locus using the pop-in/pop-out method (Allers et al., 2004; Bitan-Banin et al., 2003). Pop-out-candidates were validated by Southern blot analysis as follows (Supplementary Figure S2). 10 μg of isolated genomic DNA were digested with SacII and resulting fragments were separated by size on an 0.8% agarose gel and transferred to a nylon membrane Hybond™-N+ (GE Healthcare) via capillary blot. For detection of DNA fragments, two PCR generated probes were used: one for detection of the cas8b gene (Cas8_intern) and one for the downstream region (Cas8_do). Probes were labelled with [γ-32P]-dCTP using the DECAprime II DNA labelling kit (Thermo Fisher Scientific) and used as hybridisation probe.
SDS-PAGE-separated protein samples (Supplementary Figure S4) were processed as described by Shevchenko et al. (1996). The resulting peptides were loaded onto nano-HPLC (Dionex Ultimate 3,000 UHPLC Thermo Fischer Scientific) coupled with in-house packed C18 column (ReproSil-Pur 120 C18-AQ, 3 μm particle size, 75 μm inner diameter, 30 cm length, Dr. Maisch GmbH). The peptides were separated with a linear gradient of 11–42% buffer B (80% acetonitrile and 0.1% formic acid) at flow rate of 300 nL/min over 43 min gradient time. Eluting peptides were analysed by Q Exactive mass spectrometer (Thermo Fischer Scientific). The MS data was acquired by scanning the precursors in mass range from 350 to 1,600 m/z at a resolution of 70,000 at 200 m/z. Top 20 precursor ions were chosen for MS2 by using data-dependent acquisition (DDA) mode at a resolution of 17,500 at 200 m/z. Data analysis and search was performed using Mascot Software (2–3-02) and Scaffold 5.2.1 with Uniprot_Haloferax_volcanii database with 11,678 entries.
Peptide identification and quantification was done with MaxQuant version 2.2.0.0 (Cox and Mann, 2006) with default settings against Haloferax volcanii database accessed from UniProt at 24.01.2023 (UP000008243; 3,921 entries) and the sequence of the predicted Cas11b protein. The resulting peptides.txt table was used to extract raw intensity values for the selected peptides.
The invader plasmid pTA352-PAM3-P1.1 was used to test the interference activity of the endogenous CRISPR-Cas type I-B system (Supplementary Figure S3) (Brendel et al., 2014; Fischer et al., 2012). This plasmid is based on the Haloferax shuttle vector pTA352 (Norais et al., 2007) including spacer 1 of the CRISPR locus P1 and the PAM sequence TTC (PAM3) (Fischer et al., 2012). As a control reaction Haloferax cells were transformed with the vector pTA352 without insert. Plasmids were passaged through E. coli GM121 cells (to avoid methylation) and were then introduced into Haloferax cells using the PEG method (Allers et al., 2004; Cline et al., 1989). Four dilutions of the transformed cells (undiluted, dilutions 1:10, 1:100 and 1:1000) were spotted on plates with suitable selection medium (minimal medium containing tryptophane). To confirm active interference, H. volcanii cells were transformed at least three times with the plasmid invader or the control vector.
To analyse crRNA levels, H119 x pTA927, cas8bM545A x pTA927 and cas8bM545A x pTA927-Cas11 cells were grown in Hv-Cab supplemented with tryptophane until OD650 of 0.49–0.54. Total RNA was isolated using NucleoZol (Macherey-Nagel) and 10 μg of each culture were separated on an 8% denaturing polyacrylamide gel. RNA was transferred to a nylon membrane Hybond™-N+ (GE Healthcare) by electroblotting and the membrane was hybridised overnight at 50°C with oligo P1SP1, which was labelled with γ-ATP-32P using T4 polynucleotide kinase (Thermo Scientific). The same procedure was repeated with an oligo binding to the 5S rRNA as a loading control to normalise the P1SP1 signal. The P1SP1 probe detects spacer1 of the CRISPR locus P1.
Wild-type strain H119 and mutant strain cas8bM545A were transformed with pTA927-Cas7-N-FLAG. As a control H119 was transformed with pTA927-FLAGcontrol. Cells were grown in 500 mL Hv-Cab medium to OD650 of 0.78–0.86, then tryptophane was added to a final concentration of 3 M to induce expression of Cas7-N-FLAG. Cells were grown for 3 h, harvested and subsequently washed in enriched PBS (2.5 M NaCl, 150 mM MgCl2, 1x PBS). After lysis by ultrasonification in 10 mL Lysis Buffer (100 mM Tris–HCl pH 7.5, 10 mM EDTA, 150 mM NaCl), membranes and insoluble material were pelleted by centrifugation at 100,000 x g at 4°C for 1 h. The FLAG-tagged proteins and their interaction partners were further purified from the supernatant with Anti-FLAG M2 Affinity Gel (Sigma Aldrich), protein concentration was determined with Roti-Nanoquant (Carl Roth GmbH). 200 μg/sample or the complete fraction were filtered using an Amicon Ultra Centrifugation filter 50 kDa MWCO (Merck). The material retained on the filter was resuspended in 300 μL SEC-Buffer (1 M NaCl, 25 mM Tris–HCl pH = 7.0, 5 mM MgCl2), and then loaded on a SuperdexTM 200 Increase 10/300 GL column (Cytiva) with a flow-rate of 0.375 mL/min. The flow through was collected in 1 mL fractions and conductivity as well as absorption at 280 nm were measured. Proteins of the first fractions were separated on a 10% SDS-Gel and transferred to a nitrocellulose membrane (Roti®NC, Carl Roth GmbH) by semi-dry Western blot. The FLAG-tag containing proteins were visualised using an Anti-FLAG M2-mcAb-HRP antibody (Sigma Aldrich).
Five biological replicates of the wildtype strain H119 and the mutant strain cas8bM545A were grown in YPC at 45°C to an OD650 of 0.48–0.53. Total RNA was isolated using NucleoZol™ (Macherey-Nagel, Düren, Germany). A Turbo-DNase digest (Thermo Fisher Scientific) was performed followed by depletion of ribosomal RNA molecules using a commercial Pan-Archaea riboPOOL rRNA depletion kit (siTOOLs Biotech, dp-K024-000027). The ribo-depleted RNA samples were first fragmented using ultrasound (2 pulses of 30 s at 4°C). Then, an oligonucleotide adapter was ligated to the 3′ end of the RNA molecules. First-strand cDNA synthesis was performed using M-MLV reverse transcriptase with the 3′ adapter as primer. After purification, the 5’ Illumina TruSeq sequencing adapter was ligated to the 3′ end of the antisense cDNA. The resulting cDNA was PCR-amplified using a high-fidelity DNA polymerase and the barcoded TruSeq-libraries were pooled in approximately equimolar amounts. Sequencing of pooled libraries, spiked with PhiX control library, was performed at 7–14 million reads per sample in single-ended mode with 100 cycles on the NextSeq 2000 platform (Illumina). Demultiplexed FASTQ files were generated with bcl-convert v4.3.6 (Illumina). Raw sequencing reads were subjected to quality and adapter trimming via Cutadapt (Martin, 2011) v2.5 using a cutoff Phred score of 20 and discarding reads without any remaining bases (parameters: --nextseq-trim = 20 -m 1 -a AGATCGGAAGAGCACACGTCTGAACTCCAGTCAC). Afterwards, all reads longer than 11 nt were aligned to the Haloferax volcanii DS2 reference genome [RefSeq assembly accession: GCF_000025685.1 excluding plasmid pHV2 (NC_013965.1)] using the pipeline READemption (Förstner et al., 2014) v2.0.3 with segemehl v0.3.4 (Hoffmann et al., 2009) and an accuracy cut-off of 95% (parameters: -l 12 -a 95). READemption gene_quanti was applied to quantify aligned reads overlapping genomic features by at least 10 nt (–o 10) on the sense strand based on annotations (CDS, mature_transcript, rRNA, tRNA) originating from a detailed continuous manual curation effort (Laass et al., 2019). The annotation version from 28-MAR-2023 was used (annotated protein sequences in: https://doi.org/10.5281/zenodo.7794769), this version includes small ORFs reported in Hadjeras et al. (2023). Based on these counts, differential expression analysis was conducted via DESeq2 (Love et al., 2014) v1.24.0. Read counts were normalized by DESeq2 and fold-change shrinkage was conducted by setting the parameter betaPrior to TRUE. Differential expression was assumed at adjusted p-value after Benjamini-Hochberg correction (padj) < 0.05 and |log2FoldChange| ≥ 2 (Table 1).
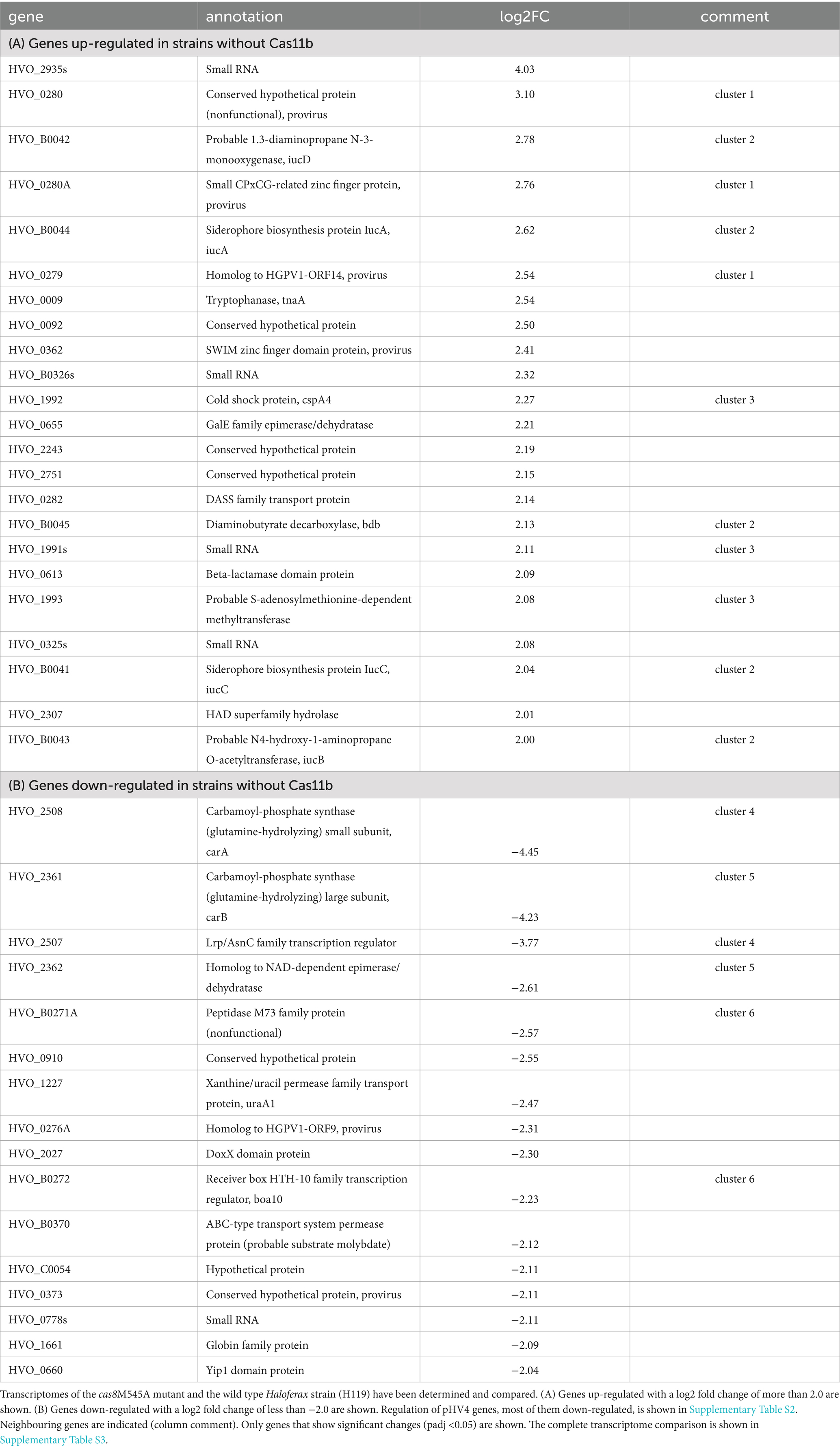
Table 1. Genes regulated in strains without Cas11b.
As initial approach to determine expression of a potential Cas11b protein from the cas8b gene we generated a plasmid for expression of a Cas8b-FLAG fusion protein. Upon expression of an N-FLAG-Cas8b fusion protein, only a single band was visible on the Western blot with anti-FLAG antibodies (Figures 2A,C). In contrast, two signals were detected upon expression of a C-FLAG-Cas8b fusion protein (Figures 2B,C), suggesting that a second protein, of about 29 kDa, is produced with a translation start in the 3′ part of the cas8b gene. Based on the sizes observed, the long protein probably corresponds to Cas8b (calculated molecular weight 92.8 kDa) while the smaller protein is likely to be Cas11b translated from the cas8b open reading frame starting at the ATG for Met545 (calculated molecular weight 28.9 kDa) (Figure 3).
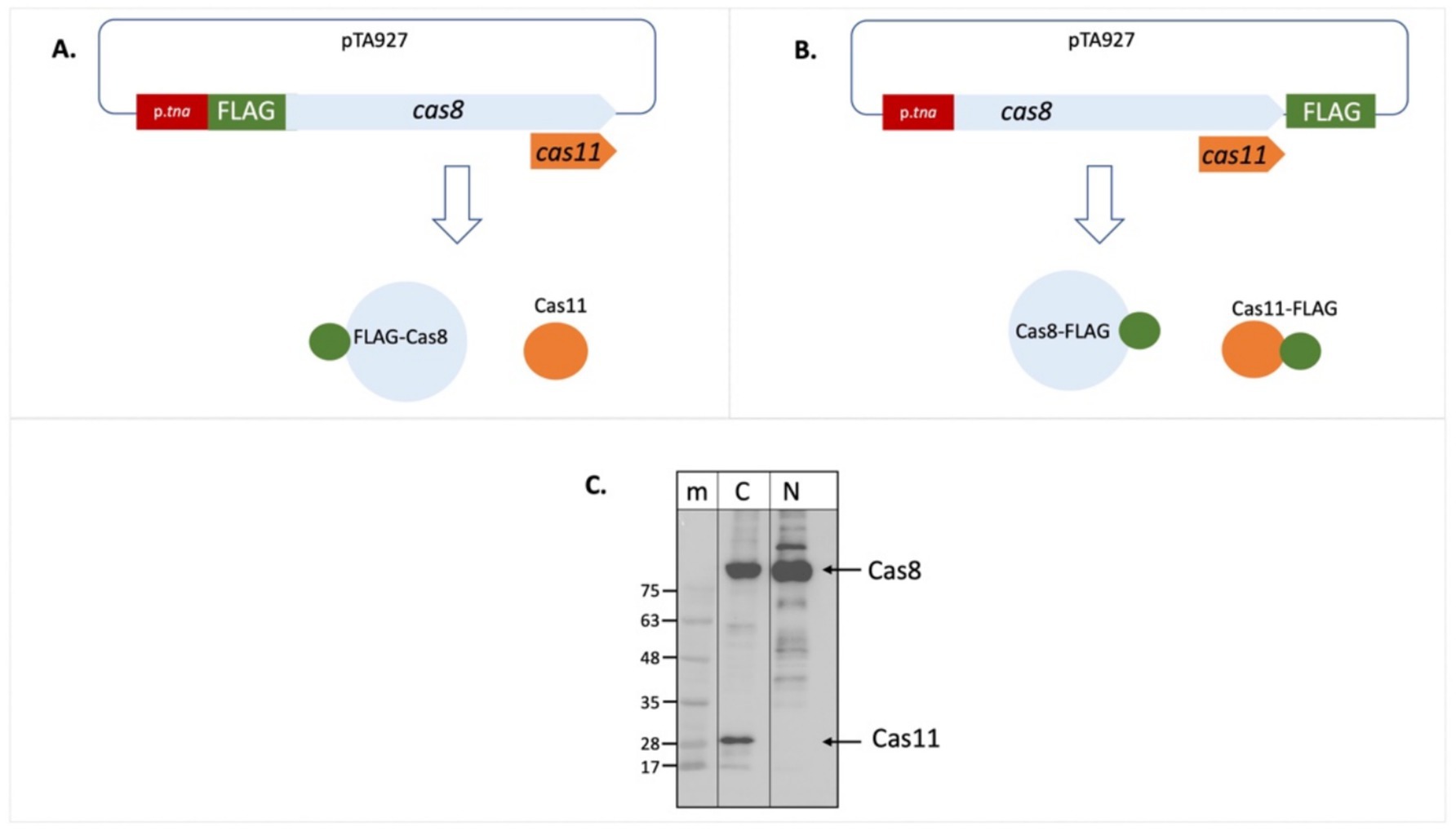
Figure 2. Cas11b is encoded in the cas8b gene. The cDNA for the FLAG tag was fused to the cas8b gene to allow expression of a Cas8b-FLAG protein. (A) Fusion to the N-terminus results in a FLAG-Cas8b protein and an untagged Cas11b protein (not visible in the Western). (B) Fusion to the C-terminus results in a Cas8b-FLAG protein as well as a Cas11b-FLAG protein. (C) Haloferax wild type cells (H119) were transformed with both constructs. Proteins were isolated and separated on SDS-PAGE and subsequently transferred to a Western membrane that was probed with an Anti-FLAG antibody. Lane m: protein size marker, sizes are given at the left in kDa, lane N: FLAG fused to the N-terminus of the protein, lane C: FLAG fused to the C-terminus of the protein. The calculated molecular weight of Cas8b is 79.4 kDa and of Cas11b (starting at Met545) is 25.5 kDa. Due to the high amount of acidic amino acids, halophilic proteins run generally slower on SDS-PAGE (Guan et al., 2015). Taking this into account, the calculated molecular weight of Cas8b is 92.8 kDa and of Cas11b (starting at Met545) is 28.9 kDa, their position in the gel is indicated with arrows at the right.
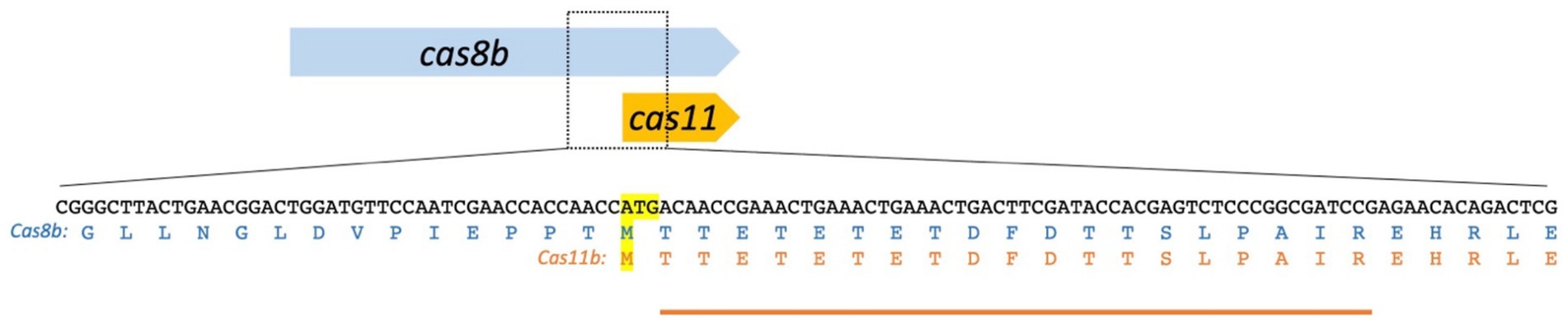
Figure 3. The cas8b gene encodes Cas8b and Cas11b. The Cas11b protein is encoded in the 3′ terminal third of the cas8b gene. The translation start ATG and the corresponding methionine residues [the internal Met545 of the Cas8b sequence (upper sequence) and the start codon of Cas11b (lower sequence)] are highlighted in yellow. The amino acid sequence of Cas8b and Cas11b is shown. The peptide identified in the mass spectrometry analysis is indicated by an orange line. The observed peptide starts with Thr546, consistent with the ATG for Met545 being used for translation initiation with subsequent cleavage of the N-terminal methionine, which is a common step in haloarchaeal N-terminal protein maturation.
Both proteins, the smaller one with 29 kDa and the larger one with 93 kDa, were analysed by mass spectrometry. The larger one was confirmed to be Cas8b, and there is strong evidence that the smaller protein is Cas11b (Figure 3; Supplementary Figure S4). A peptide starting at Thr546 was identified, strongly indicating that translation initiated at the ATG for Met545 with subsequent cleavage of the N-terminal Met, which is very often found in haloarchaea (Falb et al., 2006; Schulze et al., 2020). This peptide cannot be generated from Cas8b by tryptic cleavage, however it could be a result of non-specific fragmentation. Therefore, we wanted to obtain further support for a Cas11b translation initiation at Met545. To further test the hypothesis of an internal translation start and to prove that Met545 is the start codon of Cas11b, we mutated the Met545 codon (ATG) to Ala545 (GCG).
The mutated cas8bM545A gene was cloned as C-FLAG-fusion construct (Figure 4). Expression of the C-FLAG-cas8bM545A yielded only a single signal corresponding to a protein of about 93 kDa in the Western. Thus, the Met545Ala mutation prevents translation initiation for the Cas11b protein and therefore the 29 kDa protein cannot be detected in the Western blot. Taken together these data show that a Cas11b protein is translated from the cas8b gene with an internal in-frame translation start.
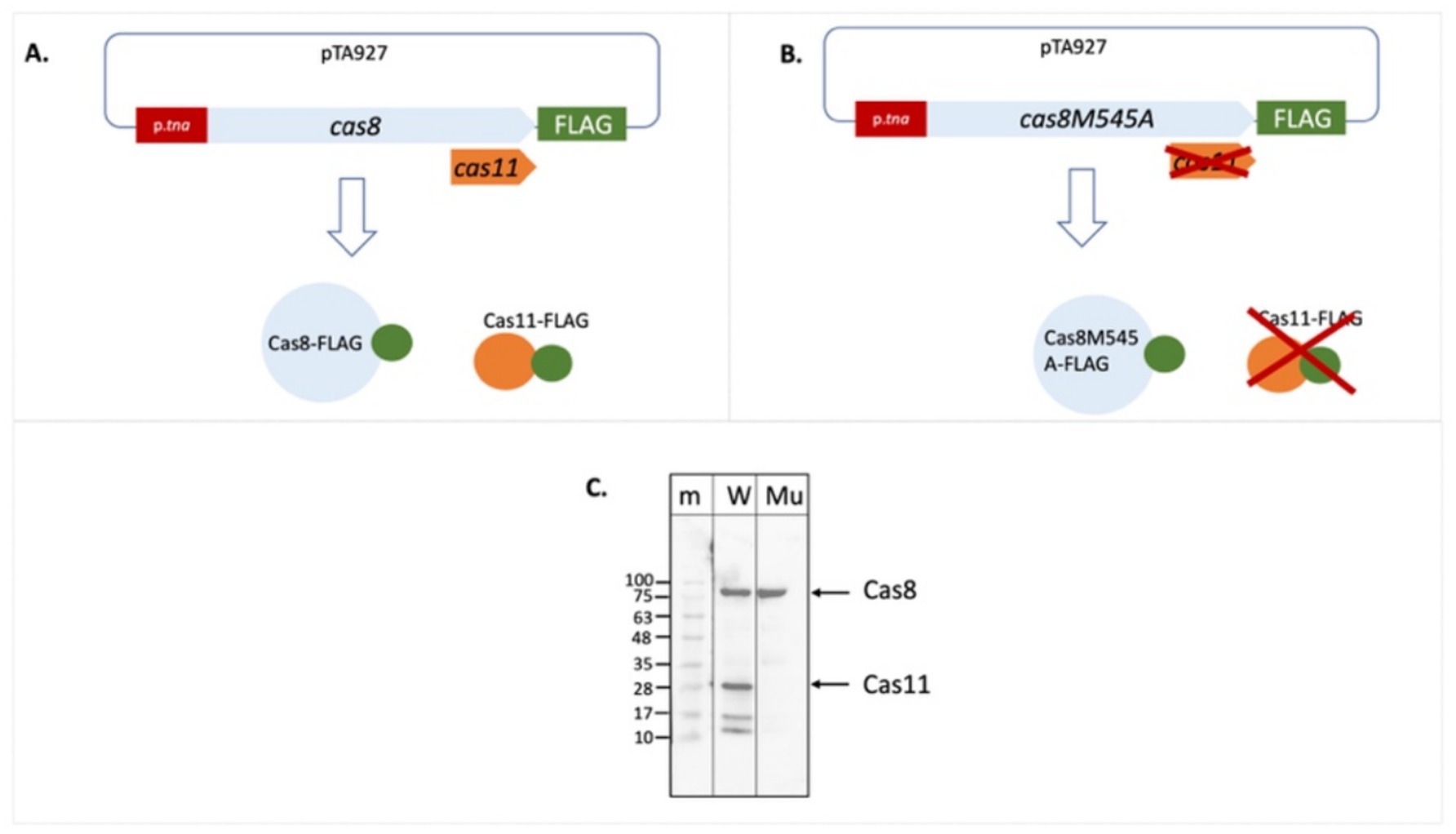
Figure 4. The Cas8bM545A mutation abolishes production of Cas11b. The FLAG cDNA was cloned in frame downstream of the wild type cas8b gene and also of the mutated cas8bM545 gene (A,B). Expression of this gene results in a Cas8b-FLAG fusion protein and potentially also in a Cas11b-FLAG fusion protein. (C) Haloferax cells were transformed with the plasmids, proteins were extracted and separated on a SDS-PAGE. After transfer of the proteins to a Western membrane the FLAG tag was identified with Anti-FLAG antibodies. Lane m: protein size marker, sizes are given at the left in kDa, lane W: C-FLAG-Cas8b and C-FLAG-Cas11b are expressed from the wild type cas8b gene, lane Mu: Only C-FLAG-Cas8b is expressed from the mutated cas8b gene. The calculated molecular weight of Cas8b is 92.8 kDa and of Cas11b (starting at Met545) is 28.9 kDa (taking into account the high percentage of acidic amino acids), their position in the gel is indicated with arrows at the right.
Next, we wanted to examine whether Cas11b is required for the defence activity of the CRISPR-Cas type I-B system. An effective interference assay using an invader plasmid was previously established for the Haloferax type I-B system (Fischer et al., 2012). A typical invader plasmid contains a protospacer flanked by a protospacer adjacent motif (PAM) that is detected by one of the crRNAs (Supplementary Figure S3) (Maier et al., 2019; Turgeman-Grott et al., 2019).
Here, we used an invader plasmid containing the functional PAM TTC and protospacer P1-1 that is detected by the Haloferax CRISPR-Cas system, since the first spacer from CRISPR locus P1 can base-pair with it (Fischer et al., 2012). To confirm that expression of the Cas8b-FLAG fusion protein expressed from a plasmid is functionally active in the Cascade complex we transformed a ∆cas8b strain with a plasmid expressing the Cas8b-C-FLAG fusion protein and subsequently with the invader plasmid. The invader assays clearly showed that the CRISPR-Cas system is active in destroying the invader plasmid (Figure 5A). In contrast, when the deletion strain ∆cas8b was transformed with a plasmid expressing the Cas8bM545A mutant, the interference reaction was not active anymore and there was no reduction of transformation efficiency evident compared to the control (Figure 5B). Thus, without Cas11b the interference reaction is far less efficient. Upon transformation of ∆cas8b with a plasmid expressing both, mutated Cas8b as well as Cas11b independently (∆cas8b x pTA927-cas8M545A_FLAGC-cas11), interference activity was recovered, and a reduction of transformation efficiency was again observed (Figure 5C). Thus, Cas11b is required for an efficient defence reaction.
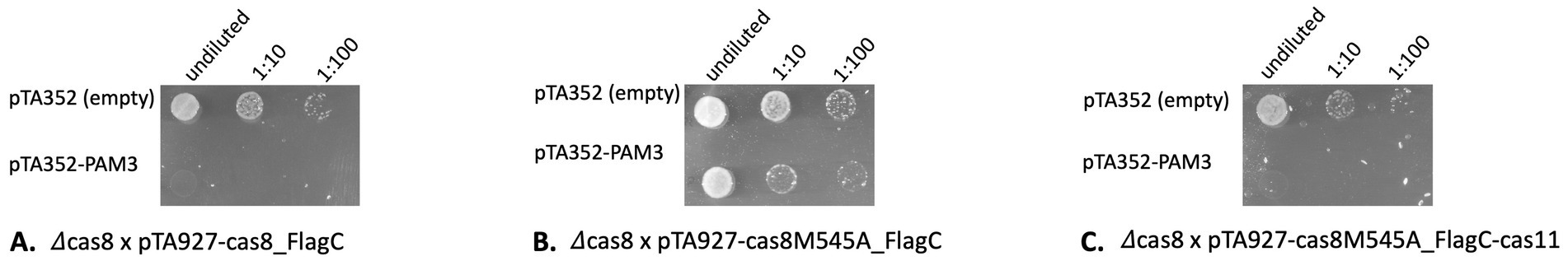
Figure 5. Without Cas11b the interference reaction is less efficient. Haloferax cells were transformed with an invader plasmid (pTA352-PAM3-P1.1) and plated on selective medium. Only cells that did not degrade the invader plasmid can grow. As control cells are transformed with the pTA352 plasmid. (A) Cells with Cas8b and Cas11b (∆cas8 x pTA927-cas8_FlagC) efficiently degrade the plasmid invader. (B) Cells with Cas8b but without Cas11b (∆cas8 x pTA927-cas8M545A_FlagC) cannot degrade the plasmid invader anymore. (C) When the mutant Cas8b and additionally Cas11b (∆cas8 x pTA927-cas8M545A_FlagC_FlagC-cas11) are present, cells efficiently degrade the plasmid invader.
To investigate further why Cas11b is important for efficient interference, we investigated the crRNA levels in a Cas11b less strain. The crRNA is one of the key players of the CRISPR-Cas system and essential for guiding the Cascade complex to the target. Previous studies showed that crRNAs are not stably maintained in H. volcanii cells when Cascade is not present, suggesting that these small RNAs are protected by binding to the Cas protein complex (Brendel et al., 2014). To analyse whether Cas11b is required for stable crRNA maintenance, we investigated the presence and levels of crRNAs in a Cas11b-deficient strain using northern blot analyses (Supplementary Figure S5), compared to wild type Haloferax cells transformed with plasmid pTA927 (without an insert) that express the native Cas8b and Cas11b from the natural cas8b gene. In these cells crRNAs were readily detected (Supplementary Figure S5). Haloferax mutant cells (cas8M545A), which express only the mutated Cas8b contained similar levels of crRNAs and complementation of H. volcanii mutant cells (cas8M545A) with pTA927-cas11b also showed wild type levels of crRNAs. These data show that crRNAs are not affected when Cas11b is missing.
Next, we wanted to investigate whether Cascade without Cas11b is still stable. Haloferax cells with Cas11b (H119 x pTA-Cas7-N-FLAG) and without Cas11b (cas8M545AxpTA-Cas7-N-FLAG) were grown, the soluble protein fraction was isolated, and proteins were separated on a gel filtration column after a FLAG immuno-purification step. Comparison of the complex levels between the two strains showed that in the absence of Cas11b, slightly less Cascade complexes were formed and eluted similarly to previous analyses (Brendel, 2014) (Supplementary Figure S6). Analysis of the fractions with Western blot confirmed the presence of Cas7-FLAG (Supplementary Figure S6).
We next wanted to determine whether the elimination of Cas11b affects the transcriptome of the cell. RNA was isolated from wild type cells (expressing Cas8b and Cas11b) and cas8M545A cells (expressing only the mutated Cas8bM545A but not Cas11b) and used for RNA-seq analyses. Many genes showed a change in expression (Table 1; Supplementary Table S3). Genes encoded on the megaplasmid pHV4 were especially affected (Supplementary Table S2): more than 100 genes located between HVO_A0279_A and HVO_A0405 were down-regulated, while only six pHV4 genes were up-regulated. Genes encoded on the proviruses Halfvol1 and Halfvol2 were also differentially expressed (down- as well as up-regulated). The expression of five small RNAs and 13 genes coding for proteins without known function also changed significantly.
The five neighbouring genes HVO_B0041-45 (iucC,D,B,A and bdb) were up-regulated with a log2 fold change of more than 2; these genes are believed to be involved in the siderophore biosynthetic pathway, which is important for iron acquisition in iron-poor environments (Blin et al., 2023; Sailer et al., 2024). Removal of Cas11b thus has an effect on this iron related pathway, probably relieving repression during growth in iron-replete medium. A strong up-regulation is observed for two genes (carA and carB) involved in carbamoyl-phosphate synthesis, which is used for pyrimidine synthesis, for example.
In Haloferax, Cas11b is produced from an internal in-frame translation initiation site that is present in the cas8b gene. To our knowledge, this is the first experimental proof for internal, in-frame translation in archaea. A ribosome profiling study performed with H. volcanii suggested potential internal in frame translation initiation sites (Gelsinger et al., 2020), but the Cas11b start site was not detected in this study and actual proof for an internal translation initiation was not shown. This is also the first report about an archaeal type I-B encoding Cas11b within the gene for the large subunit Cas8b. A short fragment of Cas8b was previously identified for the type I-B systems of Clostridium thermocellum und Methanococcus maripaludis, but at the time fragmentation of the Cas8b protein was suggested (Richter et al., 2017). Structural studies of other type I Cascade complexes showed that they contain varying numbers of Cas11 proteins: five (I-A), three (I-B) or two (I-C, I-D, I-E) (Hu et al., 2022; Jackson et al., 2014; Jore et al., 2011; Lu et al., 2024; McBride et al., 2020; Mulepati et al., 2014; O’Brien et al., 2020; Schwartz et al., 2022; Wiedenheft et al., 2011; Zhao et al., 2014). This suggests that for the I-B and I-D systems translation from the internal AUG must be three times and twice as effective as from the Cas8b AUG, respectively.
In contrast to bacterial mRNAs, haloarchaeal mRNAs do not always contain Shine-Dalgarno sequences and especially in H. volcanii the number of mRNAs containing a Shine-Dalgarno sequence is low (Kramer et al., 2014). Thus, a missing Shine-Dalgarno (SD) sequence does not contradict the presence of a translation start site. However, a potential SD sequence [GGATG instead of the standard GGAGG (Kramer et al., 2014)] can be identified upstream of the Cas11b M545 (Figure 6), and could explain how higher translation levels of Cas11b vs. Cas8b are achieved. Since the type I-B system is present in many haloarchaea (Vestergaard et al., 2014) and a Cas11b gene has not been identified in these organisms, an internal translation of a Cas11b protein from the cas8b gene might also occur in these haloarchaea.
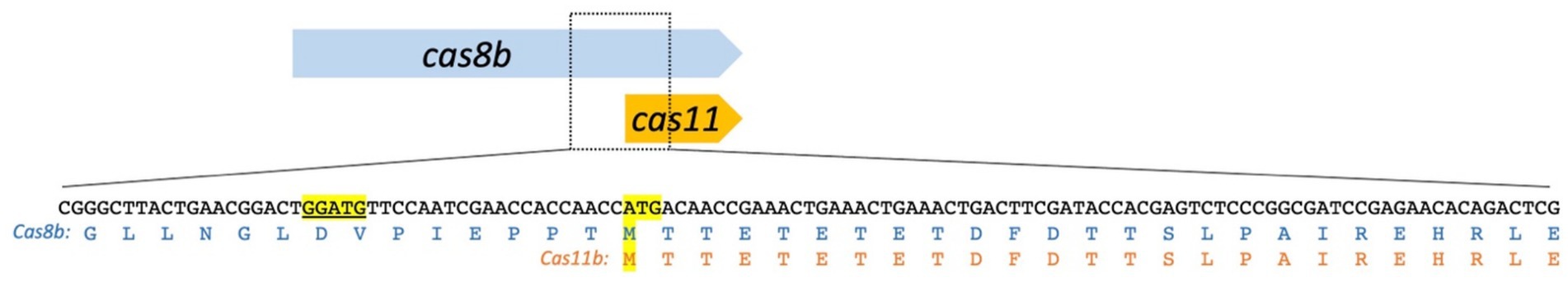
Figure 6. A potential Shine-Dalgarno sequence upstream of the Cas11b ATG. The Cas11b protein is encoded in the 3′ terminal third of the cas8b gene. The translation start ATG and the corresponding methionine residues (M545) are highlighted in yellow. The amino acid sequence of Cas8b and Cas11b is shown below the DNA sequence. A potential Shine-Dalgarno (SD) sequence is located upstream of the M545 ATG: GGATG [highlighted in yellow and underlined; the standard SD for Hfx. volcanii is: GGAGG (Kramer et al., 2014)].
Our data show that Cascade can form in Haloferax without Cas11b, which is in line with the data reported for the I-D Cascade complex (McBride et al., 2020) and for the Synechocystis sp. I-B Cascade (Tan et al., 2022). The amount of Cascade complexes without Cas11b reported here seems to be slightly lower than with Cas11b, but complexes are still able to assemble. In contrast type I-C Cascade does not form without Cas11c (Tan et al., 2022). Thus, for the Haloferax type I-B Cascade stability Cas11b is not essential. Recently an anti-CRISPR protein was found, that “chips away” the Cas7 subunits of a type I-F Cascade complex (Trost et al., 2024). Cascades from type I-F do not have Cas11, which probably makes the observed removal of Cas7 proteins easier for the anti-CRISPR protein.
The interference reaction is drastically reduced without Cas11b. Although Cascade complexes still form without Cas11b, they are not active in interference anymore. A loss in interference was also observed for the Leptospira interrogans I-B system upon depletion of the Cas11b (Hussain et al., 2023). Similar observations were made by Tan et al. while using the CRISPR-Cas3 systems for genome editing in eukaryotes: adding the Cas11 protein boosted the activity (Tan et al., 2022). Our previous data using the CRISPR interference tool (CRISPRi) show that in Haloferax CRISPRi is much more effective when the crRNAs used for CRISPRi are present in high concentrations (Stachler and Marchfelder, 2016). Thus, the concentration of crRNAs is an important factor for efficient Cascade activity. However, since a Cas11b free Cascade still binds and protects crRNAs effectively, the loss of interference is not due to lower crRNA concentrations.
McBride et al. showed for the bacterial I-D system, that Cascade without Cas11d does not bind to target DNA anymore. In E. coli, Cas11 helps to lock the non-target strand by tightening the binding of Cascade to the DNA (Hayes et al., 2016), and similar observations have been reported for other systems (Liu and Doudna, 2020; Xiao et al., 2017). Further experiments would have to reveal whether this is also true for the Haloferax Cas11b, being the reason why interference activity is reduced without Cas11b.
Taken together, the loss of interference activity might be due to inefficient interaction of the interference complex with the target DNA.
Comparison of transcriptomes from cells with and without Cas11b showed clear differences in expression of a plethora of genes. Absence of Cas11b resulted in down-regulation of more than a 100 pHV4-encoded genes in the region from HVO_A0279A-HVO_A0405. Since the CRISPR-Cas system is encoded on pHV4, one can speculate that many genes within this element have coevolved with that system and their RNA stability depends on Cas11b to some extent. A few provirus genes are also affected by the absence of Cas11b, which might be related to its role in the CRISPR-Cas defence function. Since the function of many of the regulated genes (four sRNAs and 13 genes for hypothetical proteins) is not known, we cannot deduce the functional impact of Cas11b here. Genes involved in siderophore and carbamoyl phosphate synthesis are regulated in cells without Cas11b, thus the siderophore as well as pyrimidine syntheses seem to be affected by a missing Cas11b.
Taken together the many changes in gene expression upon loss of Cas11b suggest a role of this Cas protein beyond the original CRISPR-Cas function in defence and is hinting at a key role of Cas11b in several pathways.
Transcriptome data was deposited in the European Nucleotide Archive (ENA) at EMBL-EBI under study accession number: PRJEB83033.
A-LS: Data curation, Investigation, Methodology, Validation, Visualization, Writing – review & editing. JB: Investigation, Methodology, Validation, Visualization, Writing – review & editing. AC: Data curation, Methodology, Validation, Writing – review & editing. SK: Data curation, Formal analysis, Investigation, Methodology, Validation, Writing – review & editing. TB: Data curation, Formal analysis, Methodology, Validation, Writing – review & editing. TG: Data curation, Formal analysis, Methodology, Resources, Validation, Writing – review & editing. HU: Data curation, Formal analysis, Funding acquisition, Methodology, Resources, Validation, Writing – review & editing. UG: Conceptualization, Data curation, Methodology, Writing – review & editing. AM: Conceptualization, Data curation, Funding acquisition, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Work in the laboratory of Anita Marchfelder was funded by the DFG (Ma1538/25-2 and in the frame of the DFG priority programme “CRISPR-Cas functions beyond defence” SPP2141, Ma1538/27-2). Uri Gophna was funded by the European Research Council (grant ERC-AdG 787514) and by the DFG priority programme “CRISPR-Cas functions beyond defence” SPP2141. The Core Unit Systems Medicine is partly funded (Z-6) by the Interdisciplinary Center for Clinical Research (IZKF) Würzburg.
We thank Manuela Weishaupt, Elena Katzowitsch and Panagiota Arampatzi for expert technical assistance and Lisa-Katharina Maier and Friedhelm Pfeiffer for constructive discussions and careful reading of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
The author(s) declare that no Gen AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2025.1543464/full#supplementary-material
Allers, T., Barak, S., Liddell, S., Wardell, K., and Mevarech, M. (2010). Improved strains and plasmid vectors for conditional overexpression of his-tagged proteins in Haloferax volcanii. Appl. Environ. Microbiol. 76, 1759–1769. doi: 10.1128/AEM.02670-09
Allers, T., Ngo, H. P., Mevarech, M., and Lloyd, R. G. (2004). Development of additional selectable markers for the halophilic archaeon Haloferax volcanii based on the leuB and trpA genes. Appl. Environ. Microbiol. 70, 943–953. doi: 10.1128/AEM.70.2.943-953.2004
Bitan-Banin, G., Ortenberg, R., and Mevarech, M. (2003). Development of a gene knockout system for the halophilic archaeon Haloferax volcanii by use of the pyrE gene. J. Bacteriol. 185, 772–778. doi: 10.1128/JB.185.3.772-778.2003
Blin, K., Shaw, S., Augustijn, H. E., Reitz, Z. L., Biermann, F., Alanjary, M., et al. (2023). antiSMASH 7.0: new and improved predictions for detection, regulation, chemical structures and visualisation. Nucleic Acids Res. 51, W46–w50. doi: 10.1093/nar/gkad344
Brendel, J. (2014). Charakterisierung der Prozessierungs- und Interferenzaktivität des CRISPR/Cas- Systems in Haloferax volcanii, Faculty of Natural Sciences. Ulm: Ulm University.
Brendel, J., Stoll, B., Lange, S. J., Sharma, K., Lenz, C., Stachler, A. E., et al. (2014). A complex of Cas proteins 5, 6, and 7 is required for the biogenesis and stability of clustered regularly interspaced short palindromic repeats (crispr)-derived rnas (crrnas) in Haloferax volcanii. J. Biol. Chem. 289, 7164–7177. doi: 10.1074/jbc.M113.508184
Cass, S. D., Haas, K. A., Stoll, B., Alkhnbashi, O., Sharma, K., Urlaub, H., et al. (2015). The role of Cas8 in type I CRISPR interference. Biosci. Rep. 35:e00197. doi: 10.1042/BSR20150043
Cline, S. W., Schalkwyk, L. C., and Doolittle, W. F. (1989). Transformation of the archaebacterium Halobacterium volcanii with genomic DNA. J. Bacteriol. 171, 4987–4991. doi: 10.1128/jb.171.9.4987-4991.1989
Cox, J., and Mann, M. (2006). MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat Biotechnol, 26, 1367–72. doi: 10.1038/nbt.1511
Falb, M., Aivaliotis, M., Garcia-Rizo, C., Bisle, B., Tebbe, A., Klein, C., et al. (2006). Archaeal N-terminal protein maturation commonly involves N-terminal acetylation: a large-scale proteomics survey. J. Mol. Biol. 362, 915–924. doi: 10.1016/j.jmb.2006.07.086
Fischer, S., Maier, L. K., Stoll, B., Brendel, J., Fischer, E., Pfeiffer, F., et al. (2012). An archaeal immune system can detect multiple protospacer adjacent motifs (PAMs) to target invader DNA. J. Biol. Chem. 287, 33351–33363. doi: 10.1074/jbc.M112.377002
Förstner, K. U., Vogel, J., and Sharma, C. M. (2014). READemption-a tool for the computational analysis of deep-sequencing-based transcriptome data. Bioinformatics 30, 3421–3423. doi: 10.1093/bioinformatics/btu533
Gelsinger, D. R., Dallon, E., Reddy, R., Mohammad, F., Buskirk Allen, R., and DiRuggiero, J. (2020). Ribosome profiling in archaea reveals leaderless translation, novel translational initiation sites, and ribosome pausing at single codon resolution. Nucleic Acids Res. 48, 5201–5216. doi: 10.1093/nar/gkaa304
Gophna, U., Allers, T., and Marchfelder, A. (2017). Finally, Archaea get their CRISPR-Cas toolbox. Trends Microbiol. 25, 430–432. doi: 10.1016/j.tim.2017.03.009
Guan, Y., Zhu, Q., Huang, D., Zhao, S., Jan Lo, L., and Peng, J. (2015). An equation to estimate the difference between theoretically predicted and SDS PAGE-displayed molecular weights for an acidic peptide. Sci. Rep. 5:13370. doi: 10.1038/srep13370
Hadjeras, L., Bartel, J., Maier, L. K., Maaß, S., Vogel, V., Svensson, S. L., et al. (2023). Revealing the small proteome of Haloferax volcanii by combining ribosome profiling and small-protein optimized mass spectrometry. Microlife 4:uqad001. doi: 10.1093/femsml/uqad001
Hayes, R. P., Xiao, Y., Ding, F., Van Erp, P. B., Rajashankar, K., Bailey, S., et al. (2016). Structural basis for promiscuous PAM recognition in type I-E Cascade from E. coli. Nature 530, 499–503. doi: 10.1038/nature16995
Hille, F., Richter, H., Wong, S. P., Bratovic, M., Ressel, S., and Charpentier, E. (2018). The biology of CRISPR-Cas: backward and forward. Cell 172, 1239–1259. doi: 10.1016/j.cell.2017.11.032
Hoffmann, S., Otto, C., Kurtz, S., Sharma, C. M., Khaitovich, P., Vogel, J., et al. (2009). Fast mapping of short sequences with mismatches, insertions and deletions using index structures. PLoS Comput. Biol. 5:e1000502. doi: 10.1371/journal.pcbi.1000502
Hu, C., Ni, D., Nam, K. H., Majumdar, S., McLean, J., Stahlberg, H., et al. (2022). Allosteric control of type I-A CRISPR-Cas3 complexes and establishment as effective nucleic acid detection and human genome editing tools. Mol. Cell 82, 2754–2768.e5. doi: 10.1016/j.molcel.2022.06.007
Hussain, M. S., Anand, V., and Kumar, M. (2023). Functional PAM sequence for DNA interference by CRISPR-Cas I-B system of Leptospira interrogans and the role of LinCas11b encoded within lincas8b. Int. J. Biol. Macromol. 237:124086. doi: 10.1016/j.ijbiomac.2023.124086
Jackson, R. N., Golden, S. M., van Erp, P. B., Carter, J., Westra, E. R., Brouns, S. J., et al. (2014). Crystal structure of the CRISPR RNA-guided surveillance complex from Escherichia coli. Science 345, 1473–1479. doi: 10.1126/science.1256328
Jore, M. M., Lundgren, M., van Duijn, E., Bultema, J. B., Westra, E. R., Waghmare, S. P., et al. (2011). Structural basis for CRISPR RNA-guided DNA recognition by Cascade. Nat. Struct. Mol. Biol. 18, 529–536. doi: 10.1038/nsmb.2019
Koonin, E. V., and Makarova, K. S. (2022). Evolutionary plasticity and functional versatility of CRISPR systems. PLoS Biol. 20:e3001481. doi: 10.1371/journal.pbio.3001481
Koonin, E. V., Makarova, K. S., and Zhang, F. (2017). Diversity, classification and evolution of CRISPR-Cas systems. Curr. Opin. Microbiol. 37, 67–78. doi: 10.1016/j.mib.2017.05.008
Kramer, P., Gäbel, K., Pfeiffer, F., and Soppa, J. (2014). Haloferax volcanii, a prokaryotic species that does not use the Shine Dalgarno mechanism for translation initiation at 5′-UTRs. PLoS One 9:e94979. doi: 10.1371/journal.pone.0094979
Laass, S., Monzon, V. A., Kliemt, J., Hammelmann, M., Pfeiffer, F., Förstner, K. U., et al. (2019). Characterization of the transcriptome of Haloferax volcanii, grown under four different conditions, with mixed RNA-Seq. PLoS One 14:e0215986. doi: 10.1371/journal.pone.0215986
Liu, T. Y., and Doudna, J. A. (2020). Chemistry of class 1 CRISPR-Cas effectors: binding, editing, and regulation. J. Biol. Chem. 295, 14473–14487. doi: 10.1074/jbc.REV120.007034
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15:550. doi: 10.1186/s13059-014-0550-8
Lu, M., Yu, C., Zhang, Y., Ju, W., Ye, Z., Hua, C., et al. (2024). Structure and genome editing of type I-B CRISPR-Cas. Nat. Commun. 15:4126. doi: 10.1038/s41467-024-48598-2
Maier, L-K, Alkhnbashi, O. S., Backofen, R., and Marchfelder, A. (2017). “CRISPR and Salty: CRISPR-Cas Systems in Haloarchaea” in RNA Metabolism and Gene Expression in Archaea. Ed. Clouet-d’Orval B (Springer International Publishing: Cham). 243–269.
Maier, L. K., Lange, S. J., Stoll, B., Haas, K. A., Fischer, S., Fischer, E., et al. (2013). Essential requirements for the detection and degradation of invaders by the Haloferax volcanii CRISPR/Cas system I-B. RNA Biol. 10, 865–874. doi: 10.4161/rna.24282
Maier, L. K., Stachler, A. E., Brendel, J., Stoll, B., Fischer, S., Haas, K. A., et al. (2019). The nuts and bolts of the Haloferax CRISPR-Cas system I-B. RNA Biol. 16, 469–480. doi: 10.1080/15476286.2018.1460994
Maier, L. K., Stachler, A. E., Saunders, S. J., Backofen, R., and Marchfelder, A. (2015). An active immune defense with a minimal CRISPR (clustered regularly interspaced short palindromic repeats) RNA and without the Cas6 protein. J. Biol. Chem. 290, 4192–4201. doi: 10.1074/jbc.M114.617506 Epub 612014 Dec 617515
Makarova, K. S., Wolf, Y. I., Iranzo, J., Shmakov, S. A., Alkhnbashi, O. S., Brouns, S. J. J., et al. (2020). Evolutionary classification of CRISPR–Cas systems: a burst of class 2 and derived variants. Nat. Rev. Microbiol. 18, 67–83. doi: 10.1038/s41579-019-0299-x
Martin, M. (2011). Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnetjournal; 17, No 1: Next Generation Sequencing Data Analysis. doi: 10.14806/ej.17.1.200
McBride, T. M., Schwartz, E. A., Kumar, A., Taylor, D. W., Fineran, P. C., and Fagerlund, R. D. (2020). Diverse CRISPR-Cas complexes require independent translation of small and large subunits from a single gene. Mol. Cell 80, 971–979.e7. doi: 10.1016/j.molcel.2020.11.003
Miezner, G., Turgeman-Grott, I., Zatopek, K. M., Gardner, A. F., Reshef, L., Choudhary, D. K., et al. (2023). An archaeal Cas3 protein facilitates rapid recovery from DNA damage. Microlife 4:uqad007. doi: 10.1093/femsml/uqad007
Miller, J. H. (1972). Experiments in molecular genetics. Cold Spring Harbour, N. Y: Cold Spring Harbour Laboratory Press.
Mulepati, S., Héroux, A., and Bailey, S. (2014). Crystal structure of a CRISPR RNA–guided surveillance complex bound to a ssDNA target. Science 345, 1479–1484. doi: 10.1126/science.1256996
Norais, C., Hawkins, M., Hartman, A. L., Eisen, J. A., Myllykallio, H., and Allers, T. (2007). Genetic and physical mapping of DNA replication origins in Haloferax volcanii. PLoS Genet. 3:e77. doi: 10.1371/journal.pgen.0030077
O’Brien, R. E., Santos, I. C., Wrapp, D., Bravo, J. P. K., Schwartz, E. A., Brodbelt, J. S., et al. (2020). Structural basis for assembly of non-canonical small subunits into type I-C Cascade. Nat. Commun. 11:5931. doi: 10.1038/s41467-020-19785-8
Reeks, J., Naismith, J. H., and White, M. F. (2013). CRISPR interference: a structural perspective. Biochem. J. 453, 155–166. doi: 10.1042/BJ20130316
Richter, H., Rompf, J., Wiegel, J., Rau, K., and Randau, L. (2017). Fragmentation of the CRISPR-Cas type I-B signature protein Cas8b. Biochim. Biophys. Acta Gen. Subj. 1861, 2993–3000. doi: 10.1016/j.bbagen.2017.02.026
Sailer, A. L., Jevtic, Z., Stoll, B., Wörtz, J., Sharma, K., Urlaub, H., et al. (2024). Iron starvation results in up-regulation of a probable Haloferax volcanii siderophore transporter. Front. Microbiol. 15:1422844. doi: 10.3389/fmicb.2024.1422844
Schulze, S, Adams, Z, Cerletti, M, de Castro, R., Ferreira-Cerca, S, Fufezan, C, et al. (2020). The archaeal proteome project advances knowledge about archaeal cell biology through comprehensive proteomics. Nat. Commun. 11:3145, doi: 10.1038/s41467-020-16784-7, PMCID: PMC7305310
Schwartz, E. A., McBride, T. M., Bravo, J. P. K., Wrapp, D., Fineran, P. C., Fagerlund, R. D., et al. (2022). Structural rearrangements allow nucleic acid discrimination by type I-D Cascade. Nat. Commun. 13:2829. doi: 10.1038/s41467-022-30402-8
Schwarz, T. S., Schreiber, S. S., and Marchfelder, A. (2022). CRISPR interference as a tool to repress gene expression in Haloferax volcanii. Methods Mol. Biol. 2522, 57–85. doi: 10.1007/978-1-0716-2445-6_4
Shevchenko, A., Wilm, M., Vorm, O., Jensen, O. N., Podtelejnikov, A. V., Neubauer, G., et al. (1996). A strategy for identifying gel-separated proteins in sequence databases by MS alone. Biochem. Soc. Trans. 24, 893–896. doi: 10.1042/bst0240893
Stachler, A. E., and Marchfelder, A. (2016). Gene repression in Haloarchaea using the CRISPR (clustered regularly interspaced short palindromic repeats)—Cas I-B system. J. Biol. Chem. 291, 15226–15242. doi: 10.1074/jbc.M116.724062
Stoll, B. (2013). Analyse des prokaryotischen Immunsystems CRISPR/Cas Typ I-B im archaealen Modellorganismus Haloferax volcanii, Biology II : Ulm University, Ulm, Germany.
Tan, R., Krueger, R. K., Gramelspacher, M. J., Zhou, X., Xiao, Y., Ke, A., et al. (2022). Cas11 enables genome engineering in human cells with compact CRISPR-Cas3 systems. Mol. Cell 82, 852–867.e5. doi: 10.1016/j.molcel.2021.12.032
Trost, C. N., Yang, J., Garcia, B., Hidalgo-Reyes, Y., Fung, B. C. M., Wang, J., et al. (2024). An anti-CRISPR that pulls apart a CRISPR–Cas complex. Nature 632, 375–382. doi: 10.1038/s41586-024-07642-3
Turgeman-Grott, I., Joseph, S., Marton, S., Eizenshtein, K., Naor, A., Soucy, S. M., et al. (2019). Pervasive acquisition of CRISPR memory driven by inter-species mating of archaea can limit gene transfer and influence speciation. Nat. Microbiol. 4, 177–186. doi: 10.1038/s41564-018-0302-8
Vestergaard, G., Garrett, R. A., and Shah, S. A. (2014). CRISPR adaptive immune systems of Archaea. RNA Biol. 11, 156–167. doi: 10.4161/rna.27990 Epub 22014 Feb 27997
Wiedenheft, B., Lander, G. C., Zhou, K., Jore, M. M., Brouns, S. J. J., van der Oost, J., et al. (2011). Structures of the RNA-guided surveillance complex from a bacterial immune system. Nature 477, 486–489. doi: 10.1038/nature10402
Xiao, Y., Luo, M., Hayes, R. P., Kim, J., Ng, S., Ding, F., et al. (2017). Structure basis for directional R-loop formation and substrate handover mechanisms in type I CRISPR-Cas system. Cell 170, 48–60.e11. doi: 10.1016/j.cell.2017.06.012
Keywords: archaea, Haloferax volcanii, CRISPR-Cas, type I-B, Cas11, in-frame translation
Citation: Sailer A-L, Brendel J, Chernev A, König S, Bischler T, Gräfenhan T, Urlaub H, Gophna U and Marchfelder A (2025) Internal in-frame translation generates Cas11b, which is important for effective interference in an archaeal CRISPR-Cas system. Front. Microbiol. 16:1543464. doi: 10.3389/fmicb.2025.1543464
Edited by:
Marleen van Wolferen, University of Freiburg, GermanyReviewed by:
Yulong Shen, Shandong University (Qingdao), ChinaCopyright © 2025 Sailer, Brendel, Chernev, König, Bischler, Gräfenhan, Urlaub, Gophna and Marchfelder. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: A. Marchfelder, YW5pdGEubWFyY2hmZWxkZXJAdW5pLXVsbS5kZQ==
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.