 Zhuanzhuan Fan1†Jiali Wang1†Dandan Lv1Shangbin Li1
Zhuanzhuan Fan1†Jiali Wang1†Dandan Lv1Shangbin Li1 Yuan Miao1,2Mengjun Hu1Donghui Wu3Fengying Liu4*
Yuan Miao1,2Mengjun Hu1Donghui Wu3Fengying Liu4* Dong Wang1,2*
Dong Wang1,2*- 1International Joint Research Laboratory of Global Change Ecology, School of Life Sciences, Henan University, Kaifeng, China
- 2Henan Dabieshan National Field Observation and Research Station of Forest Ecosystem, Henan University, Kaifeng, China
- 3School of Landscape Architecture, Beijing Forestry University, Beijing, China
- 4Laboratory of Resources and Applied Microbiology, School of Life Sciences, Henan University, Kaifeng, China
The conversion of cropland to orchards is one of the main measures of the Grain for Green Program for soil and water conservation and ecosystem function maintenance in the eastern Loess Plateau, China. However, the patterns and influencing forces of soil multifunctionality during the conversion from cropland to orchard remain unclear. This study evaluated the responses and regulating factors of soil multifunctionality following the conversion of cropland to pomegranate (Punica granatum L.) orchard along a 10-year chronosequence. Results showed that the conversion of cropland to pomegranate trees significantly increased the L-leucine aminopeptidase enzyme activity from 4.77 to 17.69 nmol g−1 h−1. The 10-year pomegranate stand exhibited the highest nitrogen (N) cycle multifunctionality. The N cycle multifunctionality was positively correlated with soil dissolved organic carbon (C) content, soil available phosphorus content, microbial biomass C content, phospholipid fatty acid, and soil feature index (All p < 0.05). Structural equation modeling suggested that the increased N cycle multifunctionality was attributed to soil feature index rather than soil microbial C content and phospholipid fatty acid. Land-use change did not affect soil C cycle, phosphorus cycle, or soil multifunctionality. Overall, our findings reveal that cropland conversion to orchards significantly enhances soil N cycle multifunctionality, highlighting the soil feature index’s role in maintaining soil function. The conversion from cropland to orchards, which has economic benefits and increases soil N cycle multifunctionality, is an effective approach of the Grain for Green Program in the Loess Plateau.
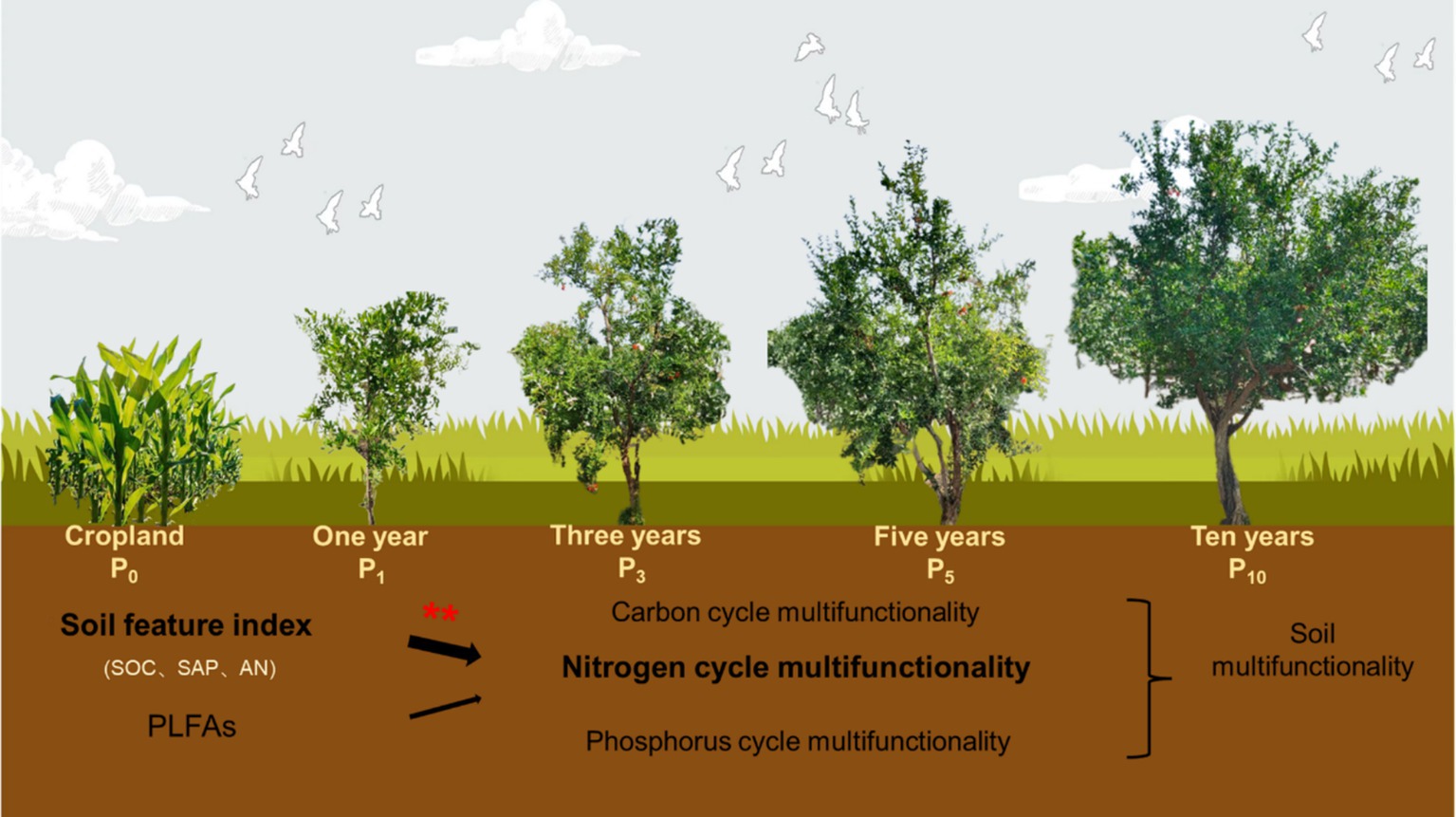
Graphical Abstract. Pomegranate forest stand age increased soil nitrogen cycle multifunctionality through soil physicochemical properties rather than microbial community.
1 Introduction
Since 1999, the “Grain for Green” Program, i.e., converting degraded land to plantations, secondary forests, and grasslands, has been implemented in the Loess Plateau, China, to prevent soil erosion, enhance soil nutrient content, and preserve the multiple soil functions (Wang et al., 2014; Liao et al., 2016). The “Grain for Green” project and other soil conservation projects have significantly enhanced ecosystem services, including carbon sequestration and net primary production on the Loess Plateau (Zhang et al., 2023). Ecological restoration projects are the effective strategy to enhance the local ecological environment and ecosystem service functions (Li et al., 2023). Soil multifunctionality refers to the ability of an ecosystem to provide a variety of soil functions and services simultaneously, including maintaining system health, productivity and sustainability, and nutrient cycling (Delgado-Baquerizo et al., 2020; Li et al., 2022; Yang et al., 2023; Jia et al., 2024). The conversion of cropland to orchards, a key approach in the “Grain for Green” project, has economic benefits and can conserve soil and water (Liao et al., 2018). However, the effect of conversion of cropland to orchards on the soil multifunctionality remains unclear, limiting the accurate assessment of ecological and environmental benefits of the “Grain for Green” project (Daelemans et al., 2023). Thus, it is necessary to investigate the effects of the conversion from cropland to orchard on soil multifunctionality to aid in effectively assessing the ecological recovery in the Loess Plateau.
Soil multifunctionality, serves as a practical indicator for assessing the interplay among various soil environmental factors, which respond sensitively to land use changes within agroecosystems (Delgado-Baquerizo et al., 2020). Soil enzymes, as the primary drivers of soil nutrient cycling, are key indicators of soil functioning and are often used to quantify soil multifunctionality (Lei et al., 2023). Note that standing age exerts dramatic but inconsistent influences on the enzyme activity associated with multifunctionality (Luo et al., 2017). Multifunctionality increases significantly with the successional stage in the natural secondary forests of the Wuyi Mountains, China (Shi et al., 2021). Soil multifunctionality increased by 3, 8, and 10% after 6, 15, and 30 years of restoration in the riparian zones of sugarcane production landscapes, respectively (Bieluczyk et al., 2023). Furthermore, compared to abandoned cropland, natural vegetation succession led to a fivefold increase in soil multifunctionality after 35 years of agricultural restoration in Russia (Ovsepyan et al., 2020). Phosphorus cycling multifunctionality has been observed to increase with stand age across the Franz Josef time series, while carbon and nitrogen cycle multifunctionality decreased with age in the organic soils (Allison et al., 2007). In addition, multifunctionality initially increases and subsequently decreases over time in tea plantations in southeastern China (Xue et al., 2006). Despite numerous experiments conducted on land use change in the agriculture ecosystem, the impacts of stand age on the soil multifunctionality of the conversion from cropland to orchards are uncertain. Therefore, examining the responses of soil multifunctionality to stand age is required to accurately assessing the factors influencing soil functions in orchards.
Land use change plays a critical role in ecosystem services and human well-being, and affects soil multifunctionality through multiple pathways (Allan et al., 2015; Yang et al., 2023). Soil microbes are essential components of the Earth’s biodiversity and significantly affect the element cycling and nutrient supply (Allison et al., 2007; Vázquez et al., 2020). Land use change regulates soil multifunctionality by alterations in soil microbial community structure and diversity (Smith et al., 2015). Yang et al. (2023) also showed divergent responses of belowground multitrophic organisms to land-use changes, and the potential role of microbial modules in forecasting soil multifunctionality. This is effectively assessed through the analysis of phospholipid fatty acids, a key indicator of microbial biomass and composition (Shi et al., 2021). It has also been shown that land use change improves soil multifunctionality by increasing the complexity of the soil polytrophic network on the Loess Plateau (He et al., 2023). Furthermore, soil nutrients, in addition to soil microbial diversity, are also regulating soil multifunctionality (Nazaries et al., 2021; Yang et al., 2024). Soil available nutrients and soil feature index are the main drivers of soil multifunctionality in the North China Plain (Jia et al., 2022). Soil multifunctionality is driven by a combination of soil nutrients and microorganisms, with the interaction of these factors explaining a greater variation in soil multifunctionality than any single factor alone (Wang et al., 2022; Wang et al., 2023). Overall, these findings indicate that the effects of land use change on soil multifunctionality are complex and multifaceted, which are likely to be determined by soil nutrients and microorganisms (Lei et al., 2023). Therefore, it is critical to investigate the relative roles of soil nutrient features and microbial attributes in the response of soil multifunctionality to land use change.
Pomegranate (Punica granatum L.) production reached 1.7 million tons in China in 2017. Pomegranate is widely cultivated in the Loess Plateau due to the abundant sunshine, good ventilation, and significant diurnal temperature differences (Wang S. L. et al., 2021). Planting pomegranates not only inhibits soil erosion and promotes soil fertility but also improves soil functions (Liao et al., 2018; Zheng et al., 2022). However, studies on how long-term stand age affects soil multifunctionality in pomegranate orchards are limited, especially in the semi-humid region of the Loess Plateau. To close these knowledge gaps, we selected four orchards of different stand ages (1, 3, 5, and 10 years) and cropland to comprehensively analyze soil nutrients, enzyme activities, microbial communities, and soil multifunctionality. This study aimed to address two questions: (1) How did soil multifunctionality respond to pomegranate stand age? and (2) What are the underlying mechanisms of soil multifunctionality in response to pomegranate stand age? Our findings would contribute to the understanding of the response of soil multifunctionality to conversion of cropland to orchards and its mechanisms, which in turn would benefit orchards development.
2 Materials and methods
2.1 Site description
This study was conducted in the eastern Loess Plateau (34 °59′ 65′′ N, 113 °25′ 05′′ E, 100 m a.s.l), in Henan Province, China (Figure 1). The region has a temperate continental monsoon climate. According to the data from the China National Meteorological Information Center from 2010 to 2020, the average annual temperature was 14.3°C, with January temperatures dropping as low as −9.6°C and July reaching up to 42.5°C. The average annual precipitation was 631.4 mm, of which 65% occurred from July to September. The main soil type was classified as sandy soil. The main crops cultivated are wheat (Triticum aestivum L.) and maize (Zea mays Linn).
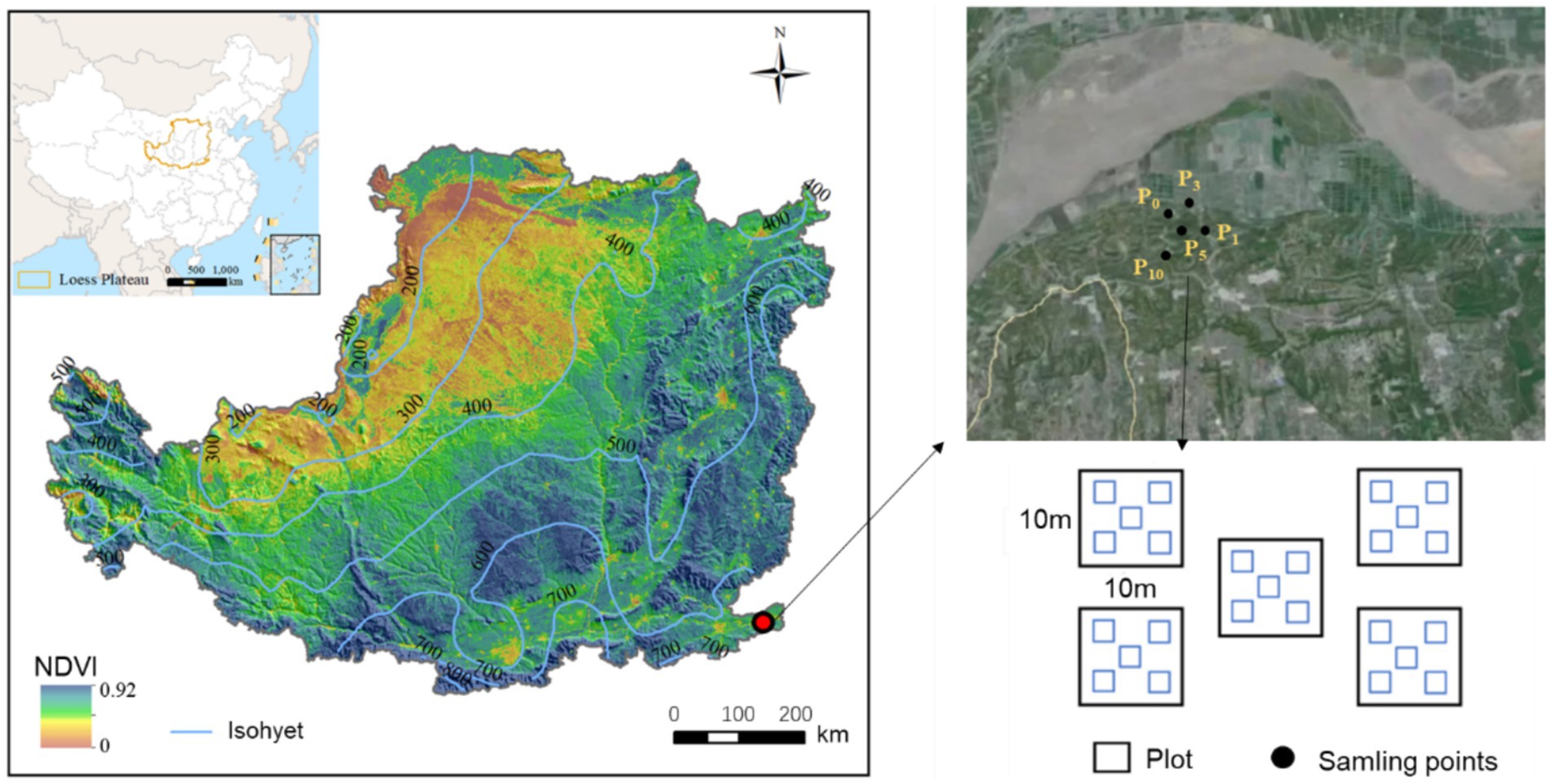
Figure 1. Experimental sitemap illustrating the location in the eastern part of the Loess Plateau, China. Black dots represent sample sites of the control (P0), pomegranate plantation of 1 year (P1), 3 years (P3), 5 years (P5), and 10 years (P10).
2.2 Experimental design
We selected five sites for the experiment, namely, cropland as a control (P0), and pomegranates with a stand age of 1 year (P1), 3 years (P3), 5 years (P5), and 10 years (P10). Five 10 m × 10 m plots were selected at each site, and the distance between two plots was larger than 100 m to ensure the independence of every plot. Each treatment included five repetitions. A single farmer owned the entire study site and applied consistent management practices across the area. These practices included uniform irrigation frequency and rate, the application of 1.5 t ha−1 of inorganic fertilizer and 3.0 t ha−1 of organic fertilizer inputs annually, and the implementation of no-tillage and weed control measures.
2.3 Soil sampling
In September 2020, soil samples were taken and mixed in the center and four corners of each plot by a 5 cm diameter soil drilling sampler at a depth range of 0–10 cm at each site. First, visible roots and other debris were removed from the soil sample. The soil samples were then passed through a 2 mm sieve. A total of 25 mixed soil samples were stored in a − 20°C refrigerator and divided into two subsamples to measure soil physical and chemical properties and determine soil enzyme activity, respectively.
2.4 Laboratory analysis
The soil enzyme activities of β-1, 4-glucosidase (BG), β-xylosidase (BX), β-cellobiohydrolase (BCE), L-leucine aminopeptidase (LAP), β-1, 4-N-acetylglucosaminidase (NAG) and acid phosphatase (AP) enzyme activities were measured from 1 g of soil to represent carbon, nitrogen, and phosphorus cycle multifunctionality, respectively (Chen et al., 2019; Hu et al., 2023). Dissolved organic carbon content (DOC) extracted with 0.5 M K2 SO4 of fresh soil sample was measured with a total Organic Carbon analyzer (vario TOC cube, Elementar, Germany). Soil available nitrogen content (AN) was determined using automated flow injection after the extraction of soil samples with 1 M KCl (Wang et al., 2022). Soil available phosphorus content (SAP) was extracted with 0.5 M of NaHCO3 and measured with an ultra-violet spectrophotometer (Fiske and Subbarow, 1925). Phospholipid fatty acids (PLFAs) were measured following the method described in Bossio and Scow (1998) and were analyzed separately using a gas chromatograph with a flame ionization detector (GC6890, Agilent Technologies, Bracknell, United Kingdom). The microbial biomass carbon (MBC) and microbial biomass nitrogen (MBN) contents were estimated by the fumigation-extraction method (Brookes et al., 1985).
2.5 Statistical analysis
To quantify soil multifunctionality (SMF), we used a method based on the calculation of the Z-scores of six key enzymes. In particular, the magnitude of the soil function is converted into a Z score and averaged to quantify the multifunctionality index (Lucas-Borja and Delgado-Baquerizo, 2019). We used the mean of the Z-scores of BG, BX and BCE to represent carbon cycle multifunctionality (CCM), the mean of the Z-scores of LAP and NAG to represent nitrogen cycle multifunctionality (NCM), and the Z-scores of LAP to represent phosphorus cycle multifunctionality (PCM). Following this, we calculated SMF using the average of the CCM, NCM, and PCM values.
Principal component analysis was used to calculate soil feature index (SFI) with three variables, namely, DOC, AN, and SAP (Masto et al., 2008). The DOC, AN, and SAP data were standardized using Z-scores before analysis. Using principal component analysis, two principal components were generated, with our study focusing on principal component one. The weight of the principal component one was multiplied by the Z-score normalized DOC, AN, and SAP data to calculate the values of SFI for each sample.
We used one-way analysis of variance (ANOVA) and least significant difference (LSD) to test for differences in soil nutrients, enzyme activity, PLFAs and multifunctionality among the five treatments. Prior to ANOVA, we performed a Shapiro–Wilk test to ensure that the experimental data conformed to a normal distribution to meet the assumptions of the statistical analysis. Pearson correlation analysis was performed between soil nutrients, PLFAs, enzyme activity and multifunctionality. Structural equation modeling (SEM) was used to estimate the direct and indirect effects of stand age, SFI, MBC, and PLFAs on NCM, based on a priori expectations of reasonable causal relationships. The difference was considered significant at the 0.05 level. These statistical analyses were carried out with R Studio (version 4.2.2, R Core Team).
3 Results
3.1 Soil nutrient and microbial biomass
The conversion of cropland to orchards exerted significant impacts on DOC, AN, SAP, MBC, PLFAs, and SFI (p < 0.05; Table 1; Figures 2, 3). The DOC of P10 increased by 56.2% compared to P0. The AN initially decreased and subsequently increased with the variations in planting time, peaking at P1. The SAP values first decreased and then increased after the conversion from cropland to orchards. The SAP peaked at P10 and increased by 133.0% compared to P0. The lowest MBC was observed at P0 and the highest at P1. The values of PLFAs at P10 were significantly higher than those at P0, P1, P3, and P5. The SFI exhibited a decreasing and subsequently increasing trend with stand ages, peaking at P10.
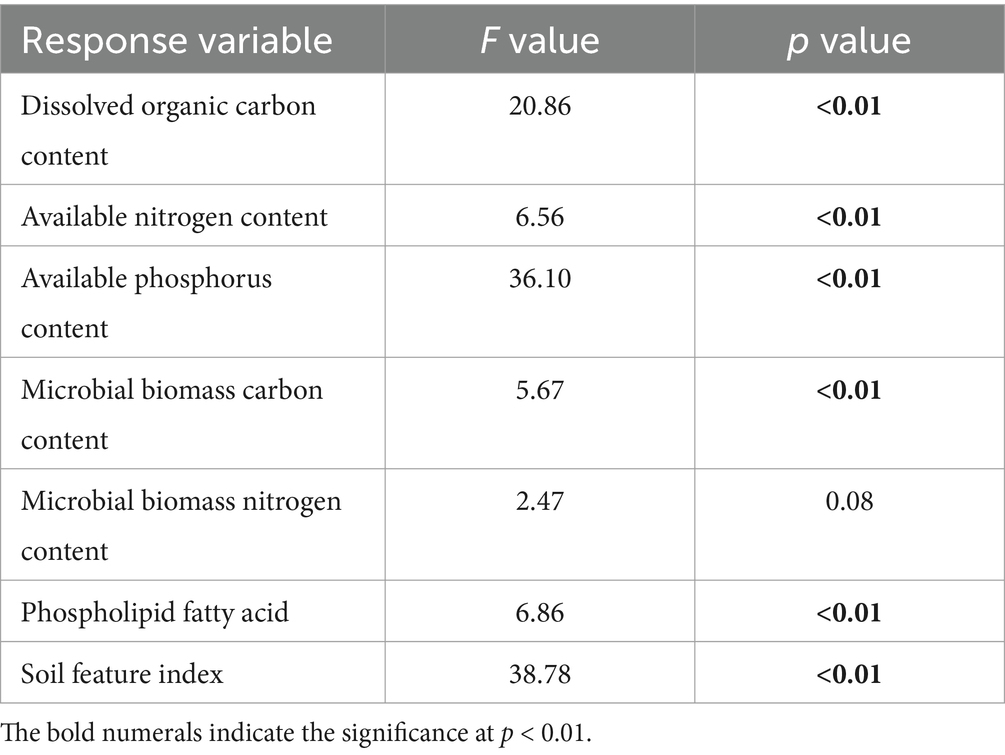
Table 1. Results of one-way analysis of variance (ANOVA) on the effects of pomegranate plantation age on dissolved organic carbon content, available nitrogen content, available phosphorus content, microbial biomass carbon content, microbial biomass nitrogen content, phospholipid fatty acid, and soil feature index.
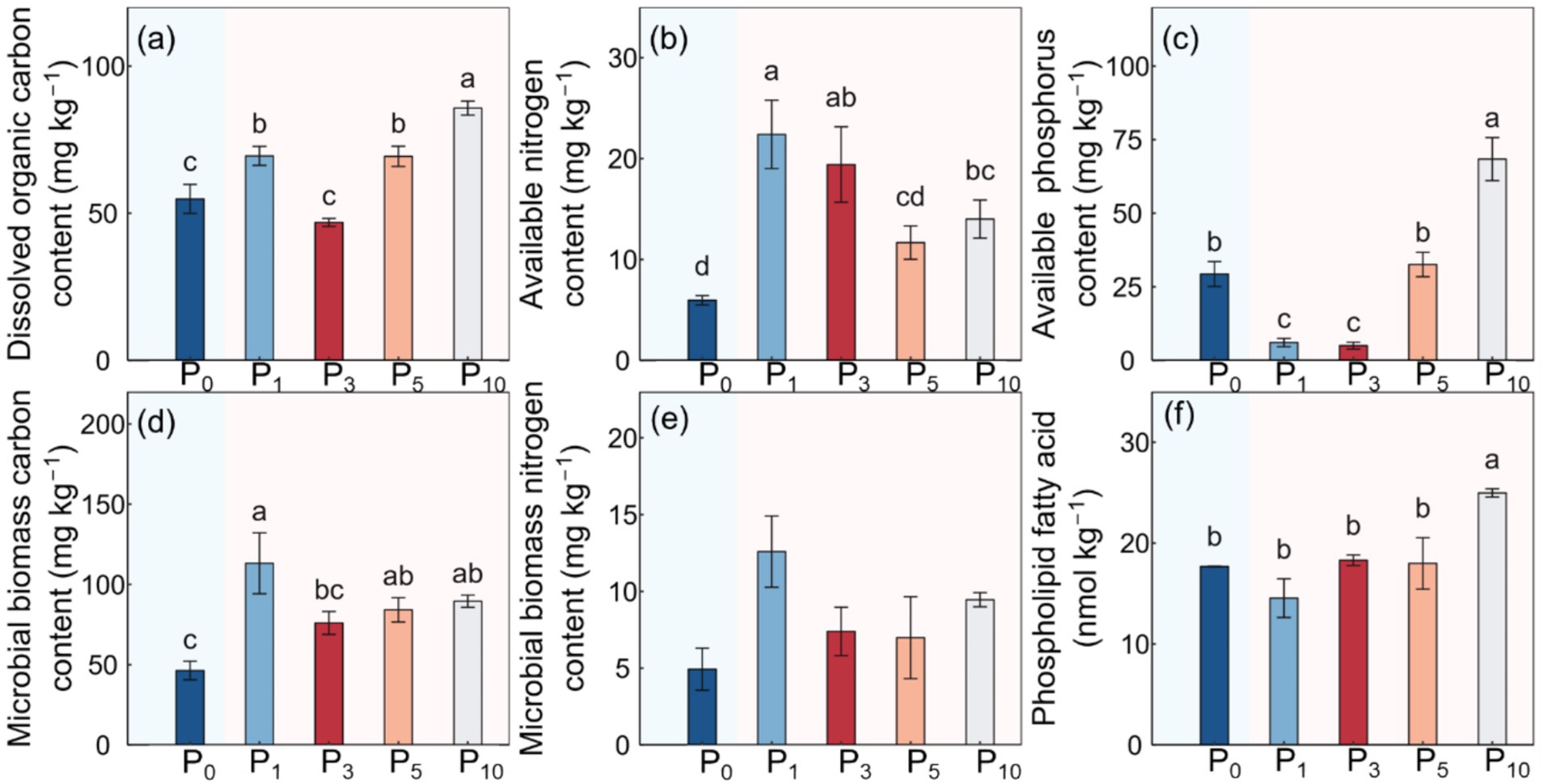
Figure 2. Soil dissolved organic carbon content (a), available nitrogen content (b), available phosphorus content (c), microbial biomass carbon content (d), microbial biomass nitrogen content (e), and phospholipid fatty acid (f) under the control (P0), pomegranate plantation of 1 year (P1), 3 years (P3), 5 years (P5), and 10 years (P10). The data are presented as the mean ± stand error. Different letters above the error bars indicate significant differences among the five treatments at p < 0.05 according to LSD tests.
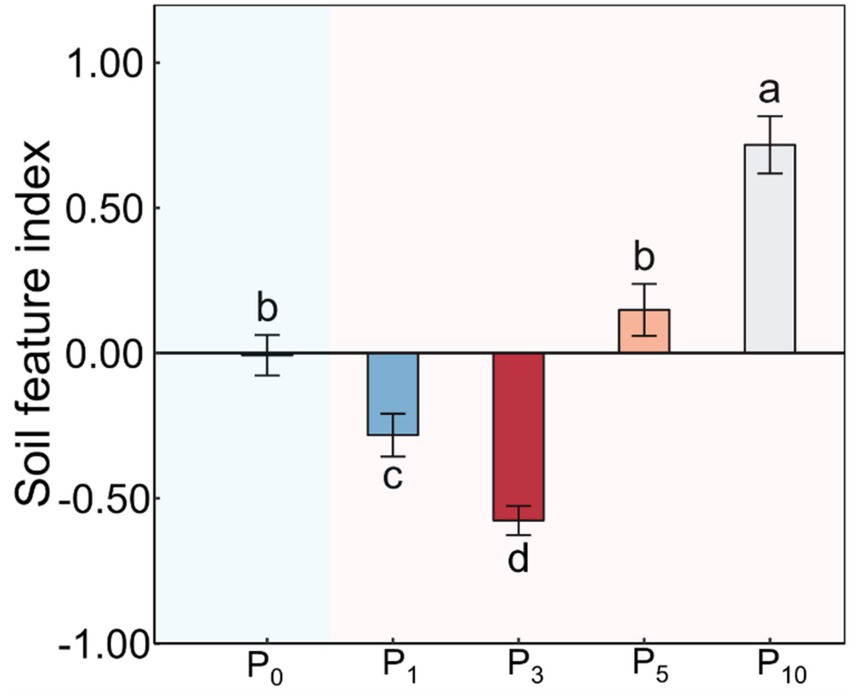
Figure 3. Soil feature index under the control (P0), pomegranate plantation of 1 year (P1), 3 years (P3), 5 years (P5), and 10 years (P10). The data are presented as the mean ± stand error. Different letters above the error bars indicate significant differences among the five treatments at p < 0.05 according to LSD tests.
3.2 Soil enzyme activities
The transition from cropland to orchards significantly affected the potential activities of BG (p < 0.05) and LAP (p < 0.01), but did not affect the potential activities of BX, BCE, NAG or AP (Table 2; Figure 4). The BG was minimized under P1 and maximized under P5. The LAP was the lowest under P0 and the highest under P10, with the latter determined to be 270.9% higher than that of cropland.
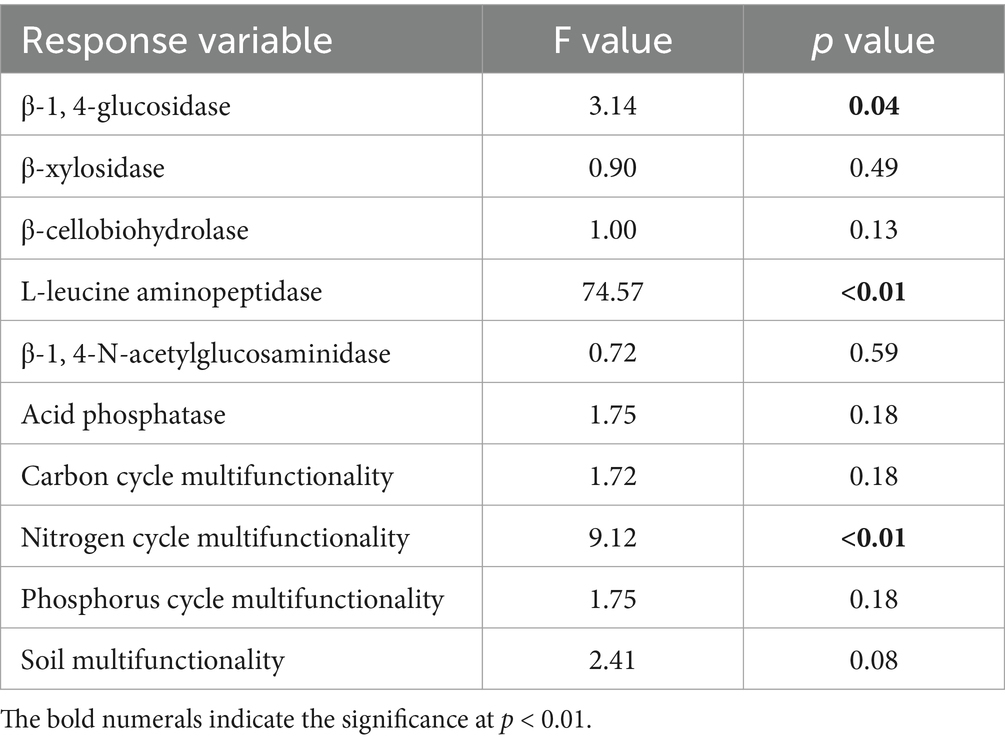
Table 2. Results of one-way analysis of variance (ANOVA) on the effects of pomegranate plantation age on soil β-1, 4-glucosidase, β-xylosidase, β-cellobiohydrolase, L-leucine aminopeptidase, β-1, 4-N-acetylglucosaminidase, acid phosphatase enzyme activities, carbon cycle multifunctionality, nitrogen cycle multifunctionality, phosphorus cycle multifunctionality, and soil multifunctionality.
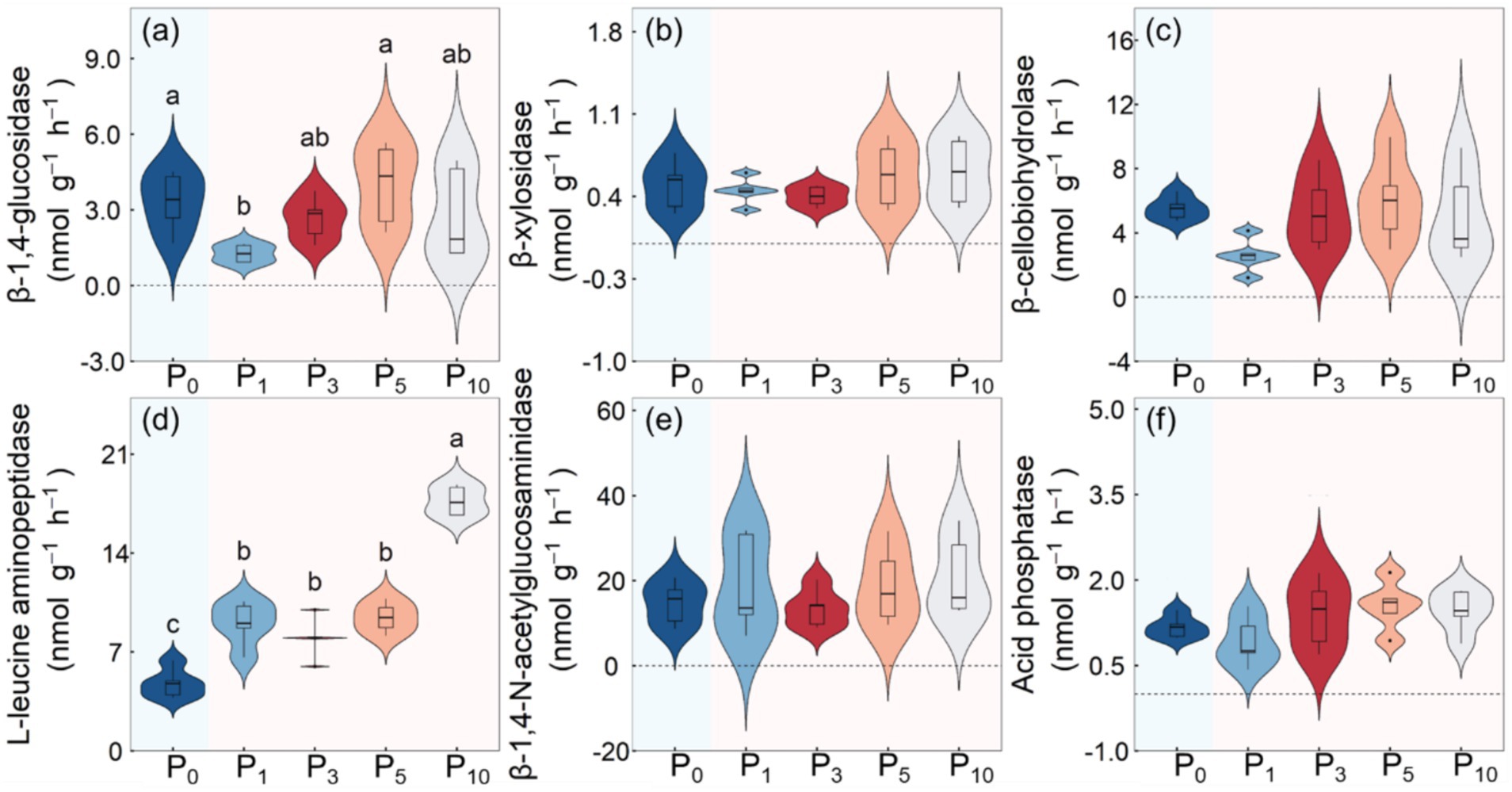
Figure 4. Soil β-1, 4-glucosidase (a), β-xylosidase (b), β-cellobiohydrolase (c), L-leucine aminopeptidase (d), β-1, 4-N-acetylglucosaminidase (e), and acid phosphatasese (f) enzyme activities under the control (P0), pomegranate plantation of 1 year (P1), 3 years (P3), 5 years (P5), and 10 years (P10). The different letters indicate the significant differences among the five treatments at p < 0.05 according to LSD tests.
3.3 Soil multifunctionality
The NCM was significantly higher after the conversion of cropland to orchards (p < 0.01, Table 2; Figure 5). The NCM increased with stand age and peaked at P10, but there was no significant difference between P1, P3, and P5. The CCM, PCM, and SMF were not significantly different among the five treatments.
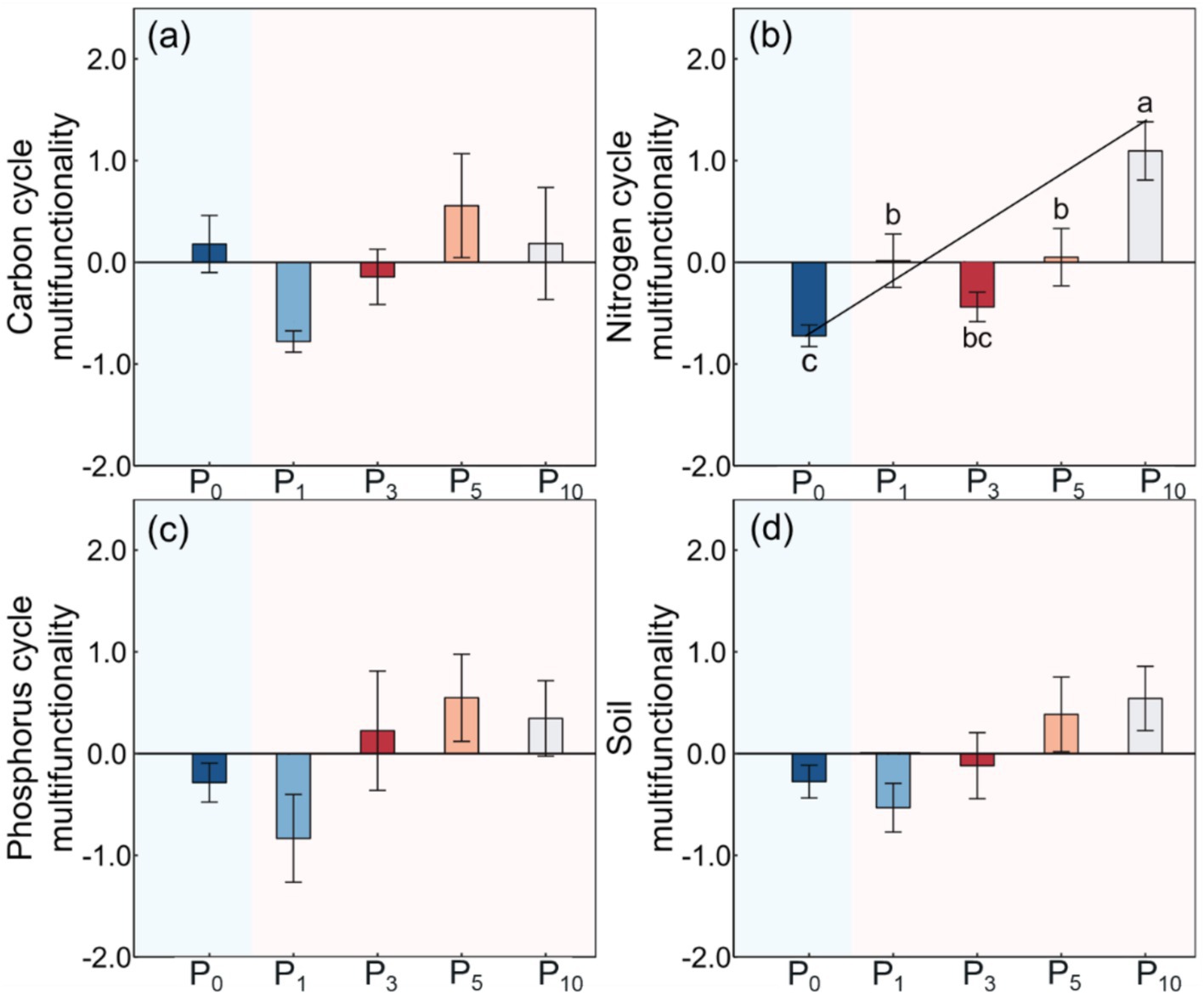
Figure 5. Soil carbon cycle multifunctionality (a), nitrogen cycle multifunctionality (b), phosphorus cycle multifunctionality (c), and soil multifunctionality (d) under the control (P0), pomegranate plantation of 1 year (P1), 2 years (P3), 5 years (P5), and 10 years (P10). The data are presented as the mean ± stand error. Different letters above the error bars indicate significant differences among the five treatments at p < 0.05 according to LSD test.
3.4 Relationships between multifunctionality and soil properties, PLFAs, and SFI
Pearson’s correlation analysis revealed that the LAP exhibited significant positive correlations with DOC (r = 0.78, p < 0.01), SAP (r = 0.67, p < 0.01), PLFAs (r = 0.57, p < 0.01), and SFI (r = 0.69, p < 0.01; Figure 6A). Moreover, the NAG presented a significant positive correlation with MBC (r = 0.55, p < 0.01) and MBN (r = 0.43, p < 0.05). The NCM was significantly positively correlated with DOC (r = 0.70, p < 0.01), SAP (r = 0.57, p < 0.01), MBC (r = 0.54, p < 0.01), PLFAs (r = 0.40, p < 0.05), and SFI (r = 0.57, p < 0.01; Figure 6B). The SMF was positively and significantly correlated with SAP (r = 0.43, p < 0.05). The SEM was performed to compare the effects of stand age on NCM in pomegranate trees (Figure 7, Fisher’s C = 13.062 with p-value = 0.011 and on 4 degrees of freedom). Our analysis revealed that SFI, MBC content, and PLFA pathways accounted for 68% of the overall variation in NCM. Furthermore, SFI was observed to enhance NCM rather than soil microbial (p < 0.01).
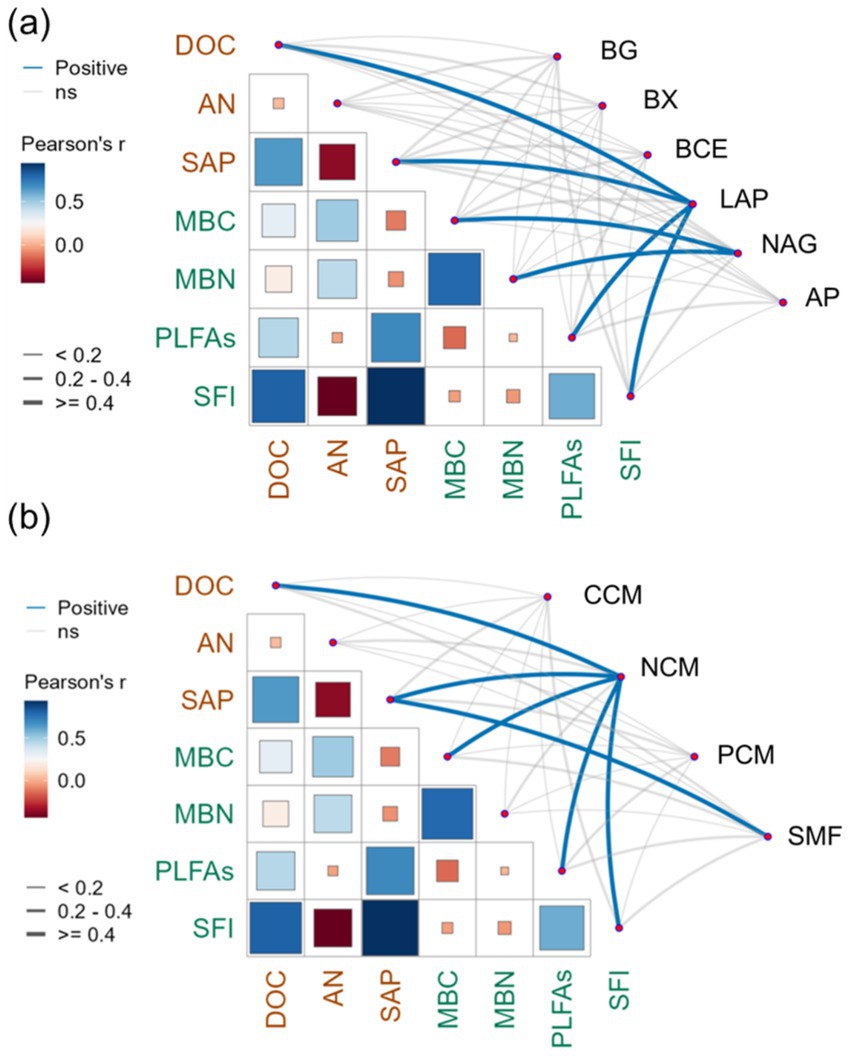
Figure 6. Pearson correlation matrix analysis between soil dissolved organic carbon content (DOC), available nitrogen content (AN), available phosphorus content (SAP), soil feature index (SFI), microbial biomass carbon content (MBC), microbial biomass nitrogen content (MBN), phospholipid fatty acid (PLFAs), soil β-1, 4-glucosidase (BG), β-xylosidase (BX), β-cellobiohydrolase (BCE), L-leucine aminopeptidase (LAP), β-1, 4-N-acetylglucosaminidase (NAG), acid phosphatase enzyme (AP) activities (a), carbon cycle multifunctionality (CCM), nitrogen cycle multifunctionality (NCM), phosphorus cycle multifunctionality (PCM), and soil multifunctionality (SMF) (b). Blue indicates a positive correlation and red a negative correlation. The depth of the color represents the r value (correlation coefficient). The thicker the line, the stronger the correlation. Blue line represents a significant positive correlation.
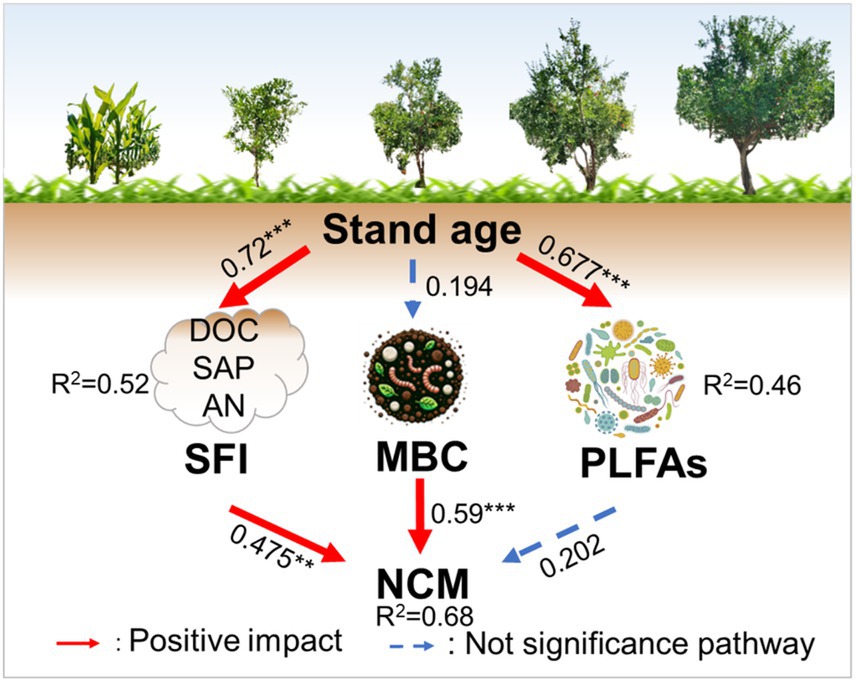
Figure 7. Structural equation modeling (SEM) of stand age effects on the soil feature index (SFI), microbial carbon content (MBC), phospholipid fatty acid (PLFAs) and nitrogen cycle multifunctionality (NCM). Red arrows indicate positive effects, solid arrows indicate operationally significant, and dashed arrows indicate insignificant. Arrow thickness represents the magnitude of the path coefficients; **, p < 0.01. ***, p < 0.001.
4 Discussion
4.1 Shifts in soil nutrients and the microbial community with stand age
Land use change alters nutrient content and microbiological properties in various ecosystems such as grassland, forest, and agroecosystems (Lucas-Borja et al., 2016; Yang et al., 2020). The DOC content increases with vegetation restoration in the semi-arid region of the Loess Plateau (Deng et al., 2019). However, our study showed that DOC decreases and subsequently increases with age. This discrepancy might be explained by regional climatic differences, as the higher rainfall in the semi-humid region of our study site compared to the semi-arid regions in other studies may lead to distinct patterns of soil nutrient dynamics (Wang et al., 2023). Increased rainfall can enhance leaching of organic carbon in the early stages of vegetation restoration, reducing DOC levels, while in later stages, improved plant growth and microbial activity may replenish soil DOC, contributing to its eventual increase (Deng et al., 2019). The AN and AP contents decreased and then increased after the conversion to orchards, which is consistent with the results along the vegetation restoration gradient in the Loess Plateau (Deng et al., 2019). Pomegranate trees affect soil properties by influencing litter, inter-root secretions, and fertilizer accumulation (Zheng et al., 2017). Compared to cropland, soil microorganisms and soil organic matter in pomegranate orchards have gradually adapted to their environment, leading to an increase in the rate of nitrogen decomposition and release (Yang et al., 2020). The observed trends of DOC and AN contents can be attributed to the intensified competition among plants and the increased demand for essential nutrients during the mid-growth period of pomegranates (Lei et al., 2023). Soil nutrient content was highest in the 10-year orchard, probably linked to the enhancement in litter inputs and nutrient cycle function (Wang et al., 2022). When pomegranate trees mature, their nutrient demand decreases as tree growth slows down, and plant residues return to the soil, thereby increasing soil nutrient content (Wang C. Q. et al., 2021). Furthermore, stopping tillage may help soil organic carbon content rise by lowering soil disturbance and soil organic carbon mineralization, which results in a rise in DOC content (Liao et al., 2018). The SFI increased significantly with the age of the pomegranate orchards, which is consistent with the study on natural forest ecosystems in Spain (Lucas-Borja et al., 2016). Mature stands typically have more efficient nutrient cycle systems that contribute to soil fertility, which in turn affects soil feature index (Wang et al., 2022). SFI refers to the capacity to sustain biological productivity, maintain environmental quality, and support nutrient cycling (Wang D. et al., 2021). Our results revealed that conversion from cropland to orchards increased the available nutrients and SFI, which were related to the responses of plant and soil properties to land use change.
Soil microbial communities not only constitute the largest part of global biological diversity, but also play a central role in the C, N, and P cycles (Zhang et al., 2017; Liao et al., 2018). The DOC content is the principal factor influencing soil microbial community diversity, suggesting a close relationship between soil microbes and soil nutrients (Yao et al., 2006). Soil microbial biomass and activity are sensitive indicators of the SFI and are crucial to the soil function of orchards (Qiang et al., 2020). The MBC increased with the stand age, which was consistent with the result obtained in one vegetation restoration process in central China (Wang et al., 2018). This increase is mainly attributed to the large amounts of litter produced by deciduous species, which enhances microbial metabolism and self-synthesis (Lu et al., 2022). In our study, PLFAs increased with the pomegranate stand age. The increase in PLFAs was attributed to increased soil nutrient availability, particularly at the initial stage (Zhong et al., 2018), reflecting the close relationship between the soil microbial community and the soil nutrient environment.
4.2 Shifts in NCM with stand age
The long-term presence of vegetation plays a crucial role in promoting soil functions related to soil nutrients (Wang et al., 2023). The enzyme activities involved in the nitrogen cycle remain relatively stable as tree age increases in the sloping citrus orchards of southwestern China (Qiang et al., 2020). In contrast, we observed the enzyme activities of the nitrogen cycle continue to increase with the plantation time. This difference could potentially be attributed to the distinct plant species and ecosystem types under investigation (Weand et al., 2010; Lucas-Borja et al., 2016). In addition, soil texture and topographic factors affect soil enzyme activity by altering temperature and humidity (Lu et al., 2022; Ma et al., 2022). The NCM significantly increases with the stand age, which is consistent with observations in apple orchards in the Losses Plateau (Wu et al., 2024) and in grassland restoration chronosequence (Guo et al., 2021). This is attributed to the fact that plant growth demands more nitrogen than what can be restored to the soil via the litter decomposition after the conversion of the cropland to pomegranate trees (Yang et al., 2020). Our findings are in line with Yang et al. (2020), who reported a gradual upward trend in NCM with time by vegetation growth. The NCM was positively correlated with DOC and SAP, suggesting that multiple abiotic factors, either jointly or individually, mediate soil nitrogen cycle multifunctionality (Jia et al., 2022). There was a significant positive correlation between NCM and SFI, indicating the key role of improving SFI in maintaining soil multifunctionality (Jia et al., 2022). In summary, improving the SFI is essential to promoting NCM.
In addition to soil nutrients, microorganisms are a major component in SMF (Guo et al., 2021; Shi et al., 2021; Li et al., 2024). Increased soil nutrient content and availability promote the proliferation of soil microorganisms, accelerate the geochemical cycle of soil nutrients, and ultimately improve soil functions (Fry et al., 2018; Vázquez et al., 2020; Ma et al., 2022). Microorganisms use energy to increase relative inputs, lifting the substrate limitation of soil enzymes and increasing enzyme activity of the nitrogen cycle (Tischer et al., 2015; Guo et al., 2021). Moreover, the NCM exhibits a significant positive correlation with both PLFAs and MBC, highlighting the vital role of microbes in maintaining soil function. Microenvironments with a higher SFI significantly stimulate microbial activity, which increases microbial abundance and diversity (Lei et al., 2023). Thereby transforming these microenvironments into hotspots for the decomposition of soil organic matter and nutrient cycle, potentially enhancing SMF by SFI (Jia et al., 2022; Wang et al., 2022).
Soil functions are driven by multiple factors, including changes in soil nutrient contents and biodiversity (Guo et al., 2021). The SEM revealed a positive direct influence of the SFI on soil NCM rather than the PLFAs. This suggests that soil NCM is primarily determined by soil nutrients rather than soil microbial community. However, microbial diversity regulates the SMF during natural secondary succession in the Wuyi Mountain, China (Shi et al., 2021). This difference could be due to the unique biological and environmental dynamics specific to the pomegranate tree population (Xu et al., 2023). Vegetation restoration reduces microbial nitrogen limitation, enhancing soil ecosystem functions. Consequently, improved nitrogen management in orchards can significantly benefit agroecosystems (Li et al., 2024).
4.3 Shifts in CCM, PCM, and SMF
In our study, we found that land use change and stand age did not significantly affect soil CCM. Among the key enzymes (BG, BX, and BCE), only the activity of BG was significantly influenced by stand age. This finding agrees with Wang C. Q. et al. (2021), who reported that BG activity varies with stand age in Cunninghamia lanceolate (Lamb.) Hook plantations. However, Qiang et al. (2020) observed that higher enzyme activities were associated with increased CCM along stand age in citrus orchards. In contrast, Wang et al. (2022) reported a decrease in soil enzyme activity with stand age over a 30-year plantation chronosequence. These differences highlight that although the activities of single enzymes such as BG differ, these differences are not sufficient to affect CCM as quantified by multiple enzymes (Qiang et al., 2020). The findings discrepancies could be attributed to cross-scale effects influenced by factors such as orchard type, soil composition, and climatic conditions (Chen et al., 2023). Additionally, our study, focusing on the early stages of the orchard, may not capture the long-term effects observed in other studies (Qiang et al., 2020). These varying results highlight the complexity of soil CCM drivers and the need for further research across different agroforestry systems.
The soil PCM increased with stand age in Eucalyptus grandis W. plantations in Brazil (Lino et al., 2016). However, stand age did not have a significant effect on soil PCM in pomegranate plantations. The observed differences may be due to the unique ecological conditions of the Loess Plateau, which require a longer time for the restoration of multiple soil functions (Wan et al., 2017). A similar conclusion was obtained in a previous study conducted in a Douglas fir plantation in Italy, which showed that AP activity did not reflect changes in soil development with stand age (Vittori Antisari et al., 2018). This may be due to the long stand-age period, giving time for the establishment of a new equilibrium in the soil (Lino et al., 2016). It is worth noting that these findings are based on specific areas of each stand ages, further investigations across a wider range of sites with greater stand ages are needed to confirm these results (Vittori Antisari et al., 2018).
Furthermore, stand age did not affect SMF in the pomegranate orchards in this study region. Pomegranate forests in the Loess Plateau region are not equivalent to a natural forest, as they have lower levels of vegetation complexity, nutrient content, and SMF (Shi et al., 2021; Wang et al., 2022; Li et al., 2024). The positive effect of mean annual precipitation on SMF is observed to be mediated by the soil moisture in the Tibetan Plateau (Jing et al., 2015). Restoration 30-year grassland supports twice soil function than the restoration 5-year grassland in a semi-arid temperate steppe in the Loess Plateau (Guo et al., 2021). The enhancement of SMF is a long process that requires decades to reach the next critical stage of development (Lucas-Borja and Delgado-Baquerizo, 2019). In this study, SMF showed an increasing trend in the P3, P5, and P10 treatments compared to the control. This may be due to the shortage of pomegranate trees, which may not have reached the tipping or turning point (Wan et al., 2017). Therefore, further investigation of older pomegranate trees is necessary to clarify the potential dynamics and patterns associated with SMF.
4.4 Applications and uncertainties
The conversion of cropland to orchard enhanced soil DOC content, AN content, SFI, and LAP activity (Liao et al., 2016). Overall, soil NCM improved as the pomegranate stand age progressed. In addition, the establishment of orchards not only improves SFI and NCM but also brings significant economic benefits and supports the prosperity of the local agricultural economy (Liao et al., 2018). Therefore, the conversion of cropland to orchards, a key aspect of the “Grain for Green” program, is particularly beneficial in the eastern Loess Plateau (Qian et al., 2014).
Our study found that stand age had no significant effect on CCM, PCM, or SMF, revealing an initial decrease and then a gradually increasing trend. The oldest trees in this study were only 10 years old, while the Chinese prickly ash orchard (Zanthoxylum bungeanum Maxim.) stands reached 35 years old (Liao et al., 2016). In addition, in a study conducted in the Mediterranean region, the maximum stand age of Spanish black pine (Pinus nigra Ar. ssp. salzmannii) forests was found to be 120 years (Lucas-Borja et al., 2016). These examples suggest that longer time periods are often required to fully capture the impact of land-use changes on soil processes. Furthermore, enzyme activity’s contribution to soil functioning varies between younger orchards and those between 20 and 40 years old (Zheng et al., 2022).
The restoration of SMF is a long process, potentially requiring more than 15 years to complete (Tian et al., 2022). Long-term monitoring is indispensable for assessing restoration success and guiding adaptive management strategies (Resch et al., 2021). Therefore, while our study provides valuable initial insights, further research is needed to evaluate the long-term impacts of land-use change on SMF, particularly with respect to larger spatial and temporal scales (Wu et al., 2024).
5 Conclusion
Our study provides insights into the effects of cropland conversion to orchards on soil multifunctionality in the Loess Plateau. After cropland to orchard conversion, soil nitrogen cycle multifunctionality increased with stand age. The SEM revealed that the soil feature index was the primary factor influencing soil nitrogen cycle multifunctionality, rather than the microbial community. Our results indicate a non-significant increasing trend in the carbon cycle, phosphorus cycle, and soil multifunctionality. Long-term monitoring of different stand ages in orchards is critical to accurately assessing soil multifunctionality. This finding is conducive to understanding the soil nutrients mediated nitrogen cycle multifunctionality mechanism along the stand ages in the orchard. These results are also useful for identifying the advantages of long-time orchard plantations and their impact on soil multifunctionality.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Author contributions
ZF: Writing – original draft, Data curation, Investigation. JW: Data curation, Writing – review & editing. DL: Investigation, Writing – review & editing. SL: Software, Writing – review & editing. YM: Conceptualization, Funding acquisition, Writing – review & editing. MH: Conceptualization, Writing – review & editing. FL: Conceptualization, Data curation, Writing – original draft. DW: Conceptualization, Funding acquisition, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This study was funded by the National Natural Science Foundation of China (NSFC32371672, 32130066), the Natural Science Foundation of Henan, China (No. 242300421102), Science and Technology Think Tank Research Topics of Henan, China (No. HNKJZK-2024-57B).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
BG, β-1, 4-glucosidase; BX, β-xylosidase; BCE, β-cellobiohydrolase; LAP, L-leucine aminopeptidase; NAG, β-1, 4-N-acetylglucosaminidase; AP, Acid phosphatase; DOC, Dissolved organic carbon content; AN, Available nitrogen content; SAP, Available phosphorus content; SFI, Soil feature index; PLFAs, Phospholipid fatty acids; MBC, Microbial biomass carbon; MBN, Microbial biomass nitrogen; SMF, Soil multifunctionality; CCM, Carbon cycle multifunctionality; NCM, Nitrogen cycle multifunctionality.
References
Allan, E., Manning, P., Alt, F., Binkenstein, J., Blaser, S., Bluthgen, N., et al. (2015). Land use intensification alters ecosystem multifunctionality via loss of biodiversity and changes to functional composition. Ecol. Lett. 18, 834–843. doi: 10.1111/ele.12469
Allison, V. J., Condron, L. M., Peltzer, D. A., Richardson, S. J., and Turner, B. L. (2007). Changes in enzyme activities and soil microbial community composition along carbon and nutrient gradients at the Franz Josef chronosequence, New Zealand. Soil Biol. Biochem. 39, 1770–1781. doi: 10.1016/j.soilbio.2007.02.006
Bieluczyk, W., Merloti, L. F., Cherubin, M. R., Mendes, L. W., Bendassolli, J. A., Rodrigues, R. R., et al. (2023). Forest restoration rehabilitates soil multifunctionality in riparian zones of sugarcane production landscapes. Sci. Total Environ. 888:164175. doi: 10.1016/j.scitotenv.2023.164175
Bossio, D. A., and Scow, K. M. (1998). Impacts of carbon and flooding on soil microbial communities: phospholipid fatty acid profiles and substrate utilization patterns. Microb. Ecol. 35, 265–278. doi: 10.1007/s002489900082
Brookes, P. C., Landman, A., Pruden, G., and Jenkinson, D. S. (1985). Chloroform fumigation and the release of soil nitrogen: a rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 17, 837–842. doi: 10.1016/0038-0717(85)90144-0
Chen, Q. L., Ding, J., Zhu, D., Hu, H. W., Delgado-Baquerizo, M., Ma, Y. B., et al. (2019). Rare microbial taxa as the major drivers of ecosystem multifunctionality in long-term fertilized soils. Soil Biol. Biochem. 141:107686. doi: 10.1016/j.soilbio.2019.107686
Chen, J., Zhang, Y., Kuzyakov, Y., Wang, D., and Olesen, J. E. (2023). Challenges in upscaling laboratory studies to ecosystems in soil microbiology research. Glob. Chang. Biol. 29, 569–574. doi: 10.1111/gcb.16537
Daelemans, R., Hulsmans, E., Fockaert, L., Vranken, L., De Bruyn, L., and Honnay, O. (2023). Agroecosystem multifunctionality of apple orchards in relation to agricultural management and landscape context. Ecol. Indic. 154:110496. doi: 10.1016/j.ecolind.2023.110496
Delgado-Baquerizo, M., Reich, P. B., Trivedi, C., Eldridge, D. J., Abades, S., Alfaro, F. D., et al. (2020). Multiple elements of soil biodiversity drive ecosystem functions across biomes. Nat. Ecol. Evol. 4, 210–220. doi: 10.1038/s41559-019-1084-y
Deng, L., Peng, C. H., Huang, C. B., Wang, K. B., Liu, Q. Y., Liu, Y. L., et al. (2019). Drivers of soil microbial metabolic limitation changes along a vegetation restoration gradient on the loess plateau, China. Geoderma 353, 188–200. doi: 10.1016/j.geoderma.2019.06.037
Fiske, C. H., and Subbarow, Y. (1925). The colorimetric determination of phosphorus. J. Biol. Chem. 66, 375–400. doi: 10.1016/s0021-9258(18)84756-1
Fry, E. L., Savage, J., Hall, A. L., Oakley, S., Pritchard, W. J., Ostle, N. J., et al. (2018). Soil multifunctionality and drought resistance are determined by plant structural traits in restoring grassland. Ecology 99, 2260–2271. doi: 10.1002/ecy.2437
Guo, Y. Q., Xu, T. Q., Cheng, J. M., Wei, G. H., and Lin, Y. B. (2021). Above-and belowground biodiversity drives soil multifunctionality along a long-term grassland restoration chronosequence. Sci. Total Environ. 772:145010. doi: 10.1016/j.scitotenv.2021.145010
He, J., Zhang, J. H., Wang, J. Y., Dong, Z. H., Meng, Z. X., Xu, R., et al. (2023). Natural restoration enhances soil multitrophic network complexity and ecosystem functions in the loess plateau. Catena 226:107059. doi: 10.1016/j.catena.2023.107059
Hu, M. J., Wang, J. L., Lu, L. L., Shao, P. S., Zhou, Z. X., Wang, D., et al. (2023). Post-fire soil extracellular enzyme activities in subtropical–warm temperate climate transitional forests. Land Degrad. Dev. 34, 1973–1983. doi: 10.1002/ldr.4582
Jia, J. Y., Hu, G. Z., Ni, G., Xie, M. X., Li, R. P., Wang, G. Z., et al. (2024). Bacteria drive soil multifunctionality while fungi are effective only at low pathogen abundance. Sci. Total Environ. 906:167596. doi: 10.1016/j.scitotenv.2023.167596
Jia, R., Zhou, J., Chu, J. C., Shahbaz, M., Yang, Y. D., Jones, D. L., et al. (2022). Insights into the associations between soil quality and ecosystem multifunctionality driven by fertilization management: a case study from the North China plain. J. Clean. Prod. 362:132265. doi: 10.1016/j.jclepro.2022.132265
Jing, X., Sanders, N. J., Shi, Y., Chu, H. Y., Classen, A. T., Zhao, K., et al. (2015). The links between ecosystem multifunctionality and above-and belowground biodiversity are mediated by climate. Nat. Commun. 6:8159. doi: 10.1038/ncomms9159
Lei, J. J., Cao, Y. X., Wang, J., Chen, Y. Z., Peng, Y. Y., Shao, Q. W., et al. (2023). Soil nutrients, enzyme activities, and microbial communities along a chronosequence of Chinese fir plantations in subtropical China. Plan. Theory 12:101931. doi: 10.3390/plants12101931
Li, W. Q., Liu, Y. J., Guo, Z. R., Xiao, P. Y., Li, T. X., Xiao, T., et al. (2024). Impacts of rice terrace abandonment on soil ecosystem multifunctionality: insights into reduced microbial diversity and exacerbated nitrogen limitation. J. Clean. Prod. 437:140731. doi: 10.1016/j.jclepro.2024.140731
Li, J. J., Yang, L., Fan, M. C., and Shangguan, Z. P. (2022). Plantation vegetation restoration enhances the relationship between rhizosphere microbial diversity and soil multifunctionality. Land Degrad. Dev. 33, 3630–3640. doi: 10.1002/ldr.4414
Li, W. W., Zhou, J. L., Xu, Z. Y., Liang, Y. K., Shi, J. W., and Zhao, X. N. (2023). Climate impact greater on vegetation NPP but human enhance benefits after the grain for green program in loess plateau. Ecol. Indic. 157:111201. doi: 10.1016/j.ecolind.2023.111201
Liao, H. K., Long, J., and Li, J. (2016). Conversion of cropland to Chinese prickly ash orchard affects soil organic carbon dynamics in a karst region of Southwest China. Nutr. Cycl. Agroecosyst. 104, 15–23. doi: 10.1007/s10705-015-9754-x
Liao, H. K., Zheng, C. L., Li, J., and Long, J. (2018). Dynamics of soil microbial recovery from cropland to orchard along a 20-year chronosequence in a degraded karst ecosystem. Sci. Total Environ. 639, 1051–1059. doi: 10.1016/j.scitotenv.2018.05.246
Lino, I. A. N., dos Santos, V. M., Costa Escobar, I. E., Alves da Silva, D. K., Ferreira de Araújo, A. S., and Costa Maia, L. (2016). Soil enzymatic activity in Eucalyptus grandis plantations of different ages. Land Degrad. Dev. 27, 77–82. doi: 10.1002/ldr.2454
Lu, Z. X., Wang, P., Ou, H. B., Wei, S. X., Wu, L. C., Jiang, Y., et al. (2022). Effects of different vegetation restoration on soil nutrients, enzyme activities, and microbial communities in degraded karst landscapes in Southwest China. For. Ecol. Manag. 508:120002. doi: 10.1016/j.foreco.2021.120002
Lucas-Borja, M. E., and Delgado-Baquerizo, M. (2019). Plant diversity and soil stoichiometry regulates the changes in multifunctionality during pine temperate forest secondary succession. Sci. Total Environ. 697:134204. doi: 10.1016/j.scitotenv.2019.134204
Lucas-Borja, M. E., Hedo, J., Cerdá, A., Candel-Pérez, D., and Viñegla, B. (2016). Unravelling the importance of forest age stand and forest structure driving microbiological soil properties, enzymatic activities and soil nutrients content in Mediterranean Spanish black pine (Pinus nigra Ar. Ssp. salzmannii) Forest. Sci. Total Environ. 562, 145–154. doi: 10.1016/j.scitotenv.2016.03.160
Luo, L., Meng, H., and Gu, J. D. (2017). Microbial extracellular enzymes in biogeochemical cycling of ecosystems. J. Environ. Manag. 197, 539–549. doi: 10.1016/j.jenvman.2017.04.023
Ma, H. Y., Zhou, J., Ge, J. Y., Nie, J. W., Zhao, J., Xue, Z. Q., et al. (2022). Intercropping improves soil ecosystem multifunctionality through enhanced available nutrients but depends on regional factors. Plant Soil 480, 71–84. doi: 10.1007/s11104-022-05554-7
Masto, R. E., Chhonkar, P. K., Singh, D., and Patra, A. K. (2008). Alternative soil quality indices for evaluating the effect of intensive cropping, fertilisation and manuring for 31 years in the semi-arid soils of India. Environ. Monit. Assess. 136, 419–435. doi: 10.1007/s10661-007-9697-z
Nazaries, L., Singh, B. P., Sarker, J. R., Fang, Y., Klein, M., and Singh, B. K. (2021). The response of soil multifunctionality to agricultural management practices can be predicted by key soil abiotic and biotic properties. Agric. Ecosyst. Environ. 307:107206. doi: 10.1016/j.agee.2020.107206
Ovsepyan, L., Kurganova, I., Lopes de Gerenyu, V., and Kuzyakov, Y. (2020). Conversion of cropland to natural vegetation boosts microbial and enzyme activities in soil. Sci. Total Environ. 743:140829. doi: 10.1016/j.scitotenv.2020.140829
Qian, X., Gu, J., Sun, W., Li, Y. D., Fu, Q. X., Wang, X. J., et al. (2014). Changes in the soil nutrient levels, enzyme activities, microbial community function, and structure during apple orchard maturation. Appl. Soil Ecol. 77, 18–25. doi: 10.1016/j.apsoil.2014.01.003
Qiang, W., Yang, B., Liu, Y., Qi, K. B., Yang, T. H., and Pang, X. Y. (2020). Effects of reclamation age on soil microbial communities and enzymatic activities in the sloping citrus orchards of southwestern China. Appl. Soil Ecol. 152:103566. doi: 10.1016/j.apsoil.2020.103566
Resch, M. C., Schütz, M., Buchmann, N., Frey, B., Graf, U., van der Putten, W. H., et al. (2021). Evaluating long-term success in grassland restoration: an ecosystem multifunctionality approach. Ecol. Appl. 31:e02271. doi: 10.1002/eap.2271
Shi, X. Z., Wang, J. Q., Lucas-Borja, M. E., Wang, Z. Y., Li, X., and Huang, Z. Q. (2021). Microbial diversity regulates ecosystem multifunctionality during natural secondary succession. J. Appl. Ecol. 58, 2833–2842. doi: 10.1111/1365-2664.14015
Smith, A. P., Marín-Spiotta, E., and Balser, T. (2015). Successional and seasonal variations in soil and litter microbial community structure and function during tropical postagricultural forest regeneration: a multiyear study. Glob. Chang. Biol. 21, 3532–3547. doi: 10.1111/gcb.12947
Tian, D. S., Xiang, Y. Z., Seabloom, E., Chen, H. Y. H., Wang, J. S., Yu, G. R., et al. (2022). Ecosystem restoration and belowground multifunctionality: a network view. Ecol. Appl. 32:e2575. doi: 10.1002/eap.2575
Tischer, A., Blagodatskaya, E., and Hamer, U. (2015). Microbial community structure and resource availability drive the catalytic efficiency of soil enzymes under land-use change conditions. Soil Biol. Biochem. 89, 226–237. doi: 10.1016/j.soilbio.2015.07.011
Vázquez, E., Benito, M., Espejo, R., and Teutscherova, N. (2020). Response of soil properties and microbial indicators to land use change in an acid soil under Mediterranean conditions. Catena 189:104486. doi: 10.1016/j.catena.2020.104486
Vittori Antisari, L., Papp, R., Vianello, G., and Marinari, S. (2018). Effects of Douglas fir stand age on soil chemical properties, nutrient dynamics, and enzyme activity: a case study in northern Apennines, Italy. Forests 9:100641. doi: 10.3390/f9100641
Wan, S. Z., Gu, H. J., Yang, Q. P., Hu, X. F., Fang, X. M., Singh, A. N., et al. (2017). Long-term fertilization increases soil nutrient accumulations but decreases biological activity in navel orange orchards of subtropical China. J. Soils Sediments 17, 2346–2356. doi: 10.1007/s11368-016-1439-9
Wang, D., Huang, X. D., Qiao, N., Geng, Q. L., Liu, Y. Z., Song, H. Q., et al. (2021). Effects of mowing and fertilization on soil quality in a semiarid grassland of North China. Land Degrad. Dev. 32, 1656–1666. doi: 10.1002/ldr.3783
Wang, J. Q., Shi, X. Z., Lucas-Borja, M. E., Lam, S. K., Wang, Z. Y., and Huang, Z. Q. (2022). Plants, soil properties and microbes directly and positively drive ecosystem multifunctionality in a plantation chronosequence. Land Degrad. Dev. 33, 3049–3057. doi: 10.1002/ldr.4371
Wang, D., Wu, G. L., Zhu, Y. J., and Shi, Z. H. (2014). Grazing exclusion effects on above-and below-ground C and N pools of typical grassland on the loess plateau (China). Catena 123, 113–120. doi: 10.1016/j.catena.2014.07.018
Wang, C. Q., Xue, L., and Jiao, R. Z. (2021). Soil organic carbon fractions, C-cycling associated hydrolytic enzymes, and microbial carbon metabolism vary with stand age in Cunninghamia lanceolate (lamb.) hook plantations. For. Ecol. Manag. 482:118887. doi: 10.1016/j.foreco.2020.118887
Wang, S. L., Yan, X. Y., Wang, D., Ahammad Siddique, I., Chen, J., Xu, Q., et al. (2021). Soil microbial community based on PLFA profiles in an age sequence of pomegranate plantation in the middle Yellow River floodplain. Diversity 13:090408. doi: 10.3390/d13090408
Wang, C., Yang, Q. N., Zhang, C., Zhang, X. L., Chen, J., and Liu, K. X. (2023). Vegetation restoration of abandoned cropland improves soil ecosystem multifunctionality through alleviating nitrogen-limitation in the China Danxia. Front. Plant Sci. 14:116179. doi: 10.3389/fpls.2023.1116179
Wang, D., Zhang, B., Zhu, L. L., Yang, Y. S., and Li, M. M. (2018). Soil and vegetation development along a 10-year restoration chronosequence in tailing dams in the Xiaoqinling gold region of Central China. Catena 167, 250–256. doi: 10.1016/j.catena.2018.05.007
Weand, M. P., Arthur, M. A., Lovett, G. M., McCulley, R. L., and Weathers, K. C. (2010). Effects of tree species and N additions on forest floor microbial communities and extracellular enzyme activities. Soil Biol. Biochem. 42, 2161–2173. doi: 10.1016/j.soilbio.2010.08.012
Wu, G. L., Liu, Y. F., Wang, D., and Zhao, J. X. (2024). Divergent successions to shrubs- and forbs-dominated meadows decrease ecosystem multifunctionality of hillside alpine meadow. Catena 236:107718. doi: 10.1016/j.catena.2023.107718
Wu, L. K., Ren, C. Y., Jiang, H., Zhang, W. Y., Chen, N., Zhao, X. N., et al. (2024). Land abandonment transforms soil microbiome stability and functional profiles in apple orchards of the Chinese losses plateau. Sci. Total Environ. 906:167556. doi: 10.1016/j.scitotenv.2023.167556
Xu, Z. Y., Hu, Z. H., Jiao, S., Bell, S. M., Xu, Q., Ma, L. L., et al. (2023). Depth-dependent effects of tree species identity on soil microbial community characteristics and multifunctionality. Sci. Total Environ. 878:162972. doi: 10.1016/j.scitotenv.2023.162972
Xue, D., Yao, H. Y., and Huang, C. Y. (2006). Microbial biomass, N mineralization and nitrification, enzyme activities, and microbial community diversity in tea orchard soils. Plant Soil 288, 319–331. doi: 10.1007/s11104-006-9123-2
Yang, Y., Chai, Y. B., Xie, H. J., Zhang, L., Zhang, Z. M., Yang, X., et al. (2023). Responses of soil microbial diversity, network complexity and multifunctionality to three land-use changes. Sci. Total Environ. 859:160225. doi: 10.1016/j.scitotenv.2022.160255
Yang, Y., Liang, C., Li, T., Cheng, H., and An, S. S. (2020). Soil extracellular enzyme stoichiometry reflects the shift from P- to N-limitation of microorganisms with grassland restoration. Soil Biol. Biochem. 149:107928. doi: 10.1016/j.soilbio.2020.107928
Yang, W., Qu, G. P., Kelly, A. R., Wu, G. L., and Zhao, J. X. (2024). Positive effects of leguminous shrub encroachment on multiple ecosystem functions of alpine meadows and steppes greatly depended on increasing soil nutrient. Catena 236:107745. doi: 10.1016/j.catena.2023.107745
Yao, H. Y., Bowman, D., and Shi, W. (2006). Soil microbial community structure and diversity in a turfgrass chronosequence: land-use change versus turfgrass management. Appl. Soil Ecol. 34, 209–218. doi: 10.1016/j.apsoil.2006.01.009
Zhang, W. W., Lu, Z. T., Yang, K., and Zhu, J. J. (2017). Impacts of conversion from secondary forests to larch plantations on the structure and function of microbial communities. Appl. Soil Ecol. 111, 73–83. doi: 10.1016/j.apsoil.2016.11.019
Zhang, J. X., Wang, Y. F., Sun, J., Chen, J. H., Zhang, J. T., Wang, D., et al. (2023). Interactions between the grain-for-green program and check dams increased vegetation carbon sequestration in the Yanhe basin, Loess Plateau. Land Degrad Dev. 34, 2310–2321. doi: 10.1002/ldr.4609
Zheng, J. F., Chen, J. H., Pan, G. X., Wang, G. M., Liu, X. Y., Zhang, X. H., et al. (2017). A long-term hybrid poplar plantation on cropland reduces soil organic carbon mineralization and shifts microbial community abundance and composition. Appl. Soil Ecol. 111, 94–104. doi: 10.1016/j.apsoil.2016.11.017
Zheng, J. Y., Wang, L., Zhao, J. S., Niu, Y. H., Xiao, H. B., Wang, Z., et al. (2022). Forty-year-old orchards promote carbon storage by changing aggregate-associated enzyme activities and microbial communities. Catena 213:106195. doi: 10.1016/j.catena.2022.106195
Keywords: enzyme activity, pomegranate trees, soil multifunctionality, land use change, the Loess Plateau
Citation: Fan Z, Wang J, Lv D, Li S, Miao Y, Hu M, Wu D, Liu F and Wang D (2024) Effects of cropland-to-orchard conversion on soil multifunctionality, particularly nitrogen cycling in the eastern Loess Plateau. Front. Microbiol. 15:1471329. doi: 10.3389/fmicb.2024.1471329
Edited by:
Juejie Yang, Beijing Forestry University, ChinaCopyright © 2024 Fan, Wang, Lv, Li, Miao, Hu, Wu, Liu and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fengying Liu, ZmVuZ3lpbmdsaXVAMTI2LmNvbQ==; Dong Wang, d2FuZ2RvbmcxOTg4MjAwNUAxNjMuY29t
†These authors have contributed equally to this work