 Ruiqiang You1,2
Ruiqiang You1,2 Yijun Kang
Yijun Kang- 1Jiangsu Key Laboratory for Bioresources of Saline Soils, Yancheng Teachers University, Yancheng, Jiangsu, China
- 2College of Environmental Science and Engineering, Yangzhou University, Yangzhou, China
The objective of this study was to investigate the impact of various chemical nitrogen fertilizers on the profile of antibiotic resistance genes (ARGs) in soil. A microcosm experiment was conducted with four treatments, including CK (control with no nitrogen), AN (ammonium nitrogen), NN (nitrate nitrogen), and ON (urea nitrogen), and the abundance of ARGs was assessed over a 30-day period using a metagenomic sequencing approach. The levels of core ARGs varied between 0.16 and 0.22 copies per cell across different treatments over time. The abundance of core ARGs in the ON treatment closely resembled that of the CK treatment, suggesting that environmentally friendly nitrogen fertilizers, particularly those in controlled release formulations, may be preferable. The core ARG abundance in the AN and NN treatments exhibited noticeable fluctuations over time. Overall, chemical nitrogen fertilizers had minimal effects on the core ARG profile as determined by principal component analysis and clustering analyses. Conversely, distinct and significant changes in bacterial communities were observed with the use of different nitrogen fertilizers. However, the influence of nitrogen fertilizers on the core ARGs is limited due to the unaffected potential bacterial hosts. Nitrogen-cycling-related genes (NCRGs), such as those involved in nitrogen-fixing (nifK, nifD, nifH) and denitrification (narG, napA, nirK, norB, nosZ) processes, exhibit a positive correlation with ARGs (rosA, mexF, bacA, vanS), indicating a potential risk of ARG proliferation during intense denitrification activities. This study indicates that the application of chemical nitrogen has a minimal effect on the abundance of ARGs in soil, thereby alleviating concerns regarding the potential accumulation of ARGs due to the use of chemical nitrogen fertilizers.
Introduction
The annual consumption of antibiotics in China reaches millions of kilograms (Zhang et al., 2015), with a significant portion being excreted into the environment by animals due to incomplete absorption and metabolism (Daghrir and Drogui, 2013). This phenomenon has led to the proliferation of antimicrobial resistance, driven by antibiotic resistance genes (ARGs) as a result of the improper use or excessive administration of antibiotics. The World Health Organization has identified this issue as a major threat to human health in the current century (Edwards et al., 2021). Despite composting and high temperature treatment, certain antibiotics and ARGs retain their biological activity upon entering the environment and can become more prevalent in soil through horizontal gene transfer (HGT) (Su et al., 2015; Wang et al., 2015; Kang et al., 2016), thereby contributing to the global dissemination of ARGs.
Agricultural soil serves as a significant locus for the occurrence and dissemination of ARGs due to the substantial influx of manure (Kang et al., 2018b; Hilaire et al., 2022), microplastic residues (Zhu et al., 2022), and the application of pesticides and hormones (García et al., 2020). Of these factors, fertilization emerges as paramount, given its role in modulating soil microbial compositions (Zeng et al., 2016; Shawver et al., 2021) and introducing heavy metals and biologically active compounds into the soil (Guo et al., 2018). There are persistent debates regarding the consequences of fertilization practices. Overall, the temporary application of composted manure may induce the proliferation of ARGs in the topsoil layer (Xie et al., 2018a; Kang et al., 2022), whereas prolonged use of manure may enhance the variety of ARGs without affecting their quantities (Wang F. et al., 2018). Nevertheless, research has indicated that the utilization of manure from cattle treated with antibiotics can have lasting effects on the soil resistome and microbial community structure for a minimum of 3 years (Shawver et al., 2021).
Controversies surrounding chemical fertilizers have been attributed to the increased indeterminacy of ARG profiles in chemically fertilized soils compared to organically fertilized soils (Liu W. et al., 2022). Studies have indicated that the application of nitrogen-phosphorus-potassium (NPK) fertilizers can lead to significant alterations in bacterial communities, while having a moderate impact on ARG diversity and abundance (Liu et al., 2017; Xie et al., 2018b; Wang F. et al., 2020). Furthermore, research has shown that the effects of chemical fertilizers on ARGs vary depending on soil type, with an increase in relative abundance observed in dryland soil and a decrease in paddy soil (Wang F. et al., 2018). In addition to these, numerous academic studies have demonstrated that the use of chemical fertilizers can increase the presence of ARGs and antibiotic-resistant pathogens in soil (Sun et al., 2019; Huang et al., 2021; Yang et al., 2022), thereby posing a potential threat to human health through plant endosphere transmission. Research has also suggested that N fertilization may be a more effective method than PK fertilizers in mitigating the risks of ARGs in soil exposed to reclaimed water irrigation (Cui et al., 2022). Additionally, scholars have highlighted the close association between ARGs and various N-cycling-related genes (NCRGs) (Wang M. et al., 2020), suggesting that potential hosts for ARGs may coexist with hosts for nitrate reduction genes (NRGs) (Hu et al., 2022). Moreover, the enforcement of denitrification has been found to contribute to a decrease in antibiotic and ARG levels (Sun et al., 2017; Wang L. et al., 2022), underscoring the importance of nitrogen fertilization in regulating soil ARGs.
The forms of nitrogen present have been identified as a factor influencing ARG profiles in both soil and plants. Sun et al. have demonstrated that the application of NH4+–N and NO3−–N may stimulate the abundances of ARGs in soil without significant differences (Sun et al., 2020). A study demonstrated that the positive impact of NH4+–N accumulation in soil was more pronounced than that of NO3−–N, although this trend was not observed in plants (Wang T. et al., 2022). A recent study has indicated that the concentration of NH4+–N may serve as a significant determinant of ARG profiles in soil (Wang et al., 2024). However, previous research has indicated that NRGs and ARGs coexist, suggesting that NO3−–N may exert a greater influence on promoting ARG abundances compared to NH4+–N (Hu et al., 2022). These differences may be attributable to the varied soil management practices employed during the field experiment. For instance, NH4+–N generally exhibits greater stability when associated with soil particles compared to NO3−–N (Zhuang, 1999), which could influence the concentrations of different nitrogen forms and the microbial utilization processes within the soil (Peacock et al., 2001). These factors are likely to exert differential effects on the soil physical and chemical properties, as well as on soil microbial communities (de Nijs et al., 2019; Alster et al., 2020), thereby ultimately impacting ARGs (Forsberg et al., 2014; Kang et al., 2018a). Therefore, to address potential variability in the field experiment, it is essential to conduct a controlled laboratory experiment to investigate possible alterations in ARG profiles resulting from different nitrogen sources.
The current study utilized metagenomic technology to assess the effects of three nitrogen sources, along with a control group, on soil ARGs over time. This research aimed to examine three principal objectives: (i) the potential variances in core ARGs profiles resulting from various forms of nitrogen application, (ii) the relationship between core ARGs and NCRGs, and (iii) potential mechanisms that may explain the divergent profiles of core ARGs. This research offers significant insights for the advancement of sustainable agricultural practices and environmental management strategies.
Materials and methods
Soil sampling
On March 17, 2023, soil samples were collected from a weed-infested area on the campus of Yancheng Teachers University, where no fertilization had occurred for a minimum of 3 years. The sandy loam soil exhibited characteristics such as a pH of 8.60 (1:1 soil to water ratio), 9.36 g/kg of organic matter, 2.89 mg/kg of ammonia nitrogen (NH4–N), and 25.00 mg/kg of nitrate nitrogen (NO3−–N). Prior to analysis, the soil was air-dried and passed through a 2 mm sieve.
Experimental design
A total of 36 Petri dishes with a diameter of 150 mm were prepared and filled with 40 g of soil each. Concentrated stocks of NaNO3, NH4Cl, and CO(NH2)2 were previously prepared separately and evenly added to three randomly selected dishes (replicates) at a final concentration of 100 mg N/kg. Additionally, nine of the 36 Petri dishes containing the same soils were supplemented with equal volumes of water as a reference to the N fertilization treatments. Therefore, four treatments were established: CK (no N addition), nitrate (NN), ammonia (AN), and urea (ON) treatments.
The Petri dishes were incubated at 25°C with 60% humidity for a period of 30 d. On days 0, 15, and 30 post-fertilization, approximately 1.0 g of mixed soils from three randomly selected points within each Petri dish were collected and preserved at −80°C. To minimize sampling-related disturbances, nine replicates were established for each treatment, with three of these replicates being randomly selected for sampling at varying time intervals.
Metagenomic sequencing and ARGs analysis
Microbial DNA was extracted from soil samples using a PowerSoil® DNA Isolation Kit and verified through agarose gel electrophoresis. Fragmentation of the DNA was achieved with a Covaris M220 sonicator, with selection of approximately 450 bp fragments for library construction. Subsequently, a paired-end library was generated utilizing the TruSeq PE Cluster Kit v3-cBot-HS and TruSeq SBS kit v.3-HS sequencing kit from Illumina. Alkali degeneration was employed to produce single-stranded DNA fragments. The treated samples underwent sequencing using an Illumina NovaSeq 6000 platform at Shanghai BIOZERON Biotechnology Co., Ltd. Raw sequences that were contaminated by adapters or contained more than 10% unknown nucleotides were excluded. Additionally, reads with abnormal nucleotides at the 5′ ends and those shorter than 75 base pairs after quality control were eliminated.
The resulting clean sequences were analyzed for ARGs using the ARG analysis pipeline (ARG-OAP, v2.2) with cutoff parameters of e-value ≤10−7, sequence identity ≥80%, and alignment length ≥25 amino acids (Yin et al., 2018). Reads were aligned against the Structured Antibiotic Resistance Genes (SARG, v2.2) and BacMet databases using BLASTX with recommended parameters, resulting in the annotation of reads as either ARG or metal resistance genes (MRG) (Ma et al., 2017). ARG types and subtypes were automatically identified using ARGs-OAP (v2.0), and their relative abundances were quantified as “ARG copy per cell” based on a specific equation (Jia et al., 2020; Yin et al., 2023):
Where, NARG-like reads represents the number of the ARG-like sequences matched with one specific ARG reference sequence; Lread is the read length (150 bp); LARG reference sequence is the nucleotide sequence length of the correspondingly specific ARG reference sequence (bp); Cell number is the estimated number of bacterial cells in each metagenomic dataset calculated by ARGs-OAP based on the searching results of 30 sets of essential single copy marker genes (Yin et al., 2018); n is the number of mapped ARG reference sequences belonging to the target ARG types/subtypes. Besides, the percentage (%) of individual ARG type/subtype was calculated as the ratio of the relative abundance of target ARG type/subtype versus the relative abundance of all ARGs in each sample. Furthermore, the trimmed reads underwent taxonomic classification using the Kraken2 (v2.0.6) database, which includes the NCBI reference nucleotide database (RefSeq) for various taxonomic levels (Wood and Salzberg, 2014).
Data analysis
In this study, the ARGs and mobile genetic elements (MGEs) were identified as core ARGs and core MGEs, defined as persistent ARGs present in all sites regardless of treatment over time. The raw data was analyzed using SPSS Statistics for Windows version 18.0 to calculate means and standard errors. A two-way analysis of variance (ANOVA) was conducted to examine the impact of nitrogen forms (NN, AN, and ON) and time (0 d, 15 d, and 30 d) on the levels of ARGs in different treatments. Subsequently, Bonferroni’s post-hoc tests were conducted to assess significant differences among treatments within a group corresponding to the x-axis at a significance level of 0.05. Principal component analysis (PCA) was utilized to analyze the relative abundance of ARGs and heatmaps were generated using the ClustVis online tool (Metsalu and Vilo, 2015). The column and line charts featured in this study were produced using Sigma Plot for Windows Version 10.0 (Systat Software, San Jose, CA). Furthermore, to investigate the potential correlation between core ARGs and NCRGs, NCRGs were annotated utilizing the integrative database NCycDB with an e-value cutoff of 10−7 (Tu et al., 2019).
Results
Profiles of core ARGs following N fertilizations over time
A total of 22 types and 381 subtypes of ARGs were identified. Among these, 101 core ARGs were consistently present in all treatments over time and were used for analysis. The abundance of ARGs in all treatments ranged from 0.16 to 0.22 copies per cell (Figure 1A). The trend of ARG abundance in the treatment with nitrogen fertilization was similar to that in the control group, and there was no significant difference between the two groups at the same sampling time (Figure 1B). Fertilizations with AN and NN have a significant impact on the abundance of ARGs over a 30-day period. Specifically, there is a notable decrease in ARG abundance on day 15 following fertilization with AN and NN, with a subsequent increase on day 30. The abundance of ARGs in the AN treatment on day 15 is significantly lower than that in the NN treatment (Figure 1B), although there is no statistical difference between the two treatments on day 30. Additionally, the changes in MGEs following nitrogen fertilizations are more pronounced than those in core ARGs (Supplementary Figures S1A,B). Nitrogen fertilizations lead to a decrease in MGE abundance on day 30 compared to day 0. MGE abundance in the ON treatment is lowest on both day 0 and day 30, while the AN treatment shows the lowest abundance on day 15.
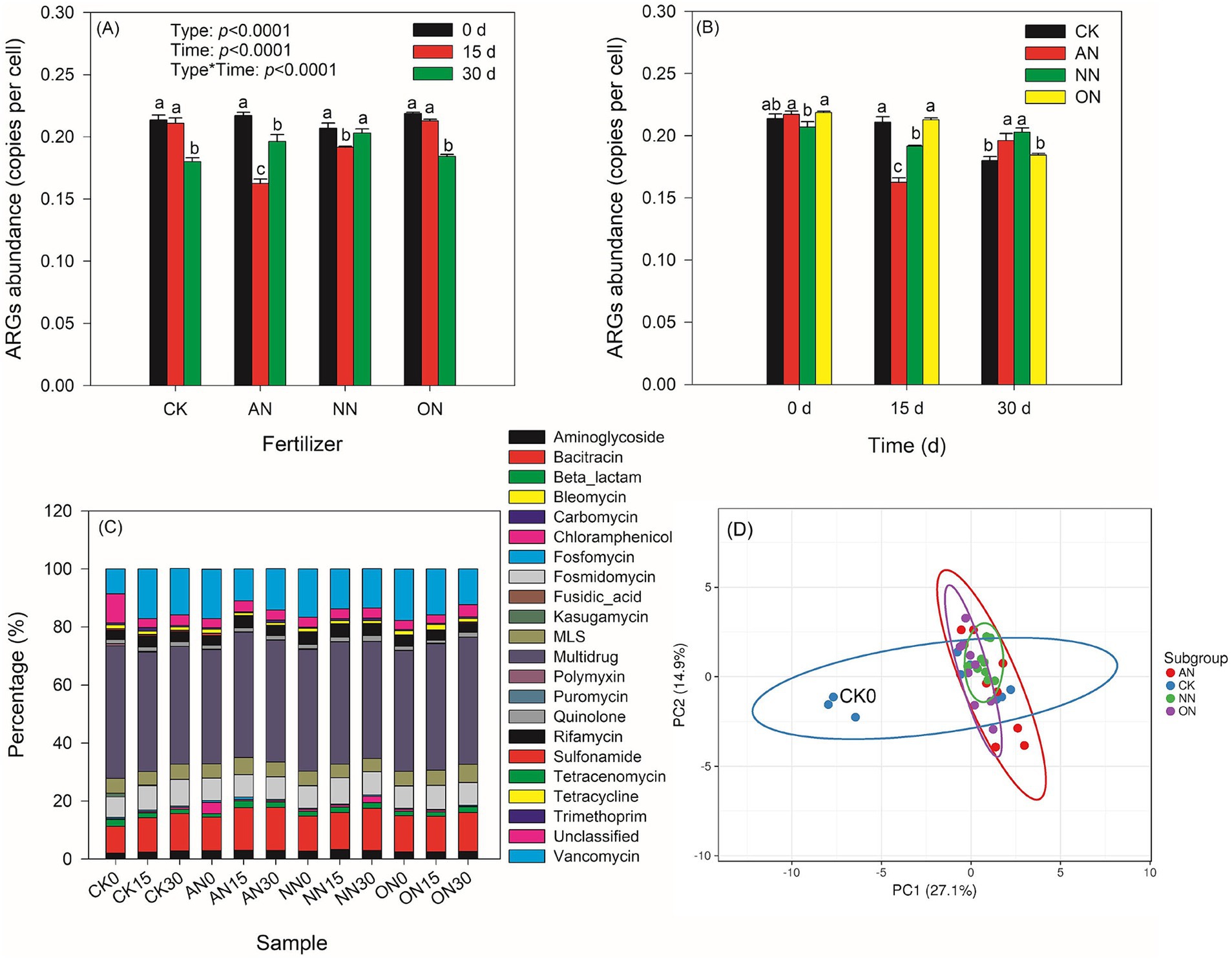
Figure 1. Effects of different chemical N fertilizers on core ARGs profiles with time in soil. Chemical N fertilizers have limited impacts on core ARGs abundances (A,B). Similar compositions of ARGs among treatments with time (C). Principal component analysis (PCA) shows that all the treatments can be categorized together except CK0, which is labelled in the panel (D). In panels (A,B) different letters represent significant differences within one group corresponding to the x-axis.
The prevalence of unclassified ARGs is significantly higher in the CK0 treatment compared to other treatments (Figure 1C). Notably, the percentages of Bacitracin and Vancomycin ARGs exhibit distinct differences among the treatments. PCA demonstrates that all treatments, except for CK0, exhibit similar ARG profiles (Figure 1D). CK treatment displays more pronounced changes over time compared to N fertilization treatments, which show convergent ARG profiles, particularly evident in the NN treatment. These findings are further supported by PCA analyses conducted over time and across treatments. These findings can be further confirmed through PCA over time or across different treatments. The lower explanations of PC1 in the N fertilization treatments compared to the CK (Supplementary Figures S2A–D) suggest that N fertilization leads to more consistent profiles of ARGs over time. Additionally, the decreasing explanation values of PC1 over time (Supplementary Figures S2E–H) indicate that time plays a crucial role in the convergence effects of N fertilization.
Potential bacterial hosts for ARGs
The abundance of ARGs exhibits a positive correlation with the quantities of ARG subtypes (Figure 2A), metal resistance genes (MRGs) (Figure 2B), and MGEs (Figure 2C). MGEs play a crucial role in facilitating the horizontal transfer of ARGs among bacterial strains. Consequently, the identification of bacterial species that demonstrate positive correlations with both ARGs and MGEs serves as a key criterion for screening potential hosts in this study. Specifically, among the six core MGEs examined, IS91, tniB, and tnpA exhibit positive correlations with certain bacterial species (Figure 2D). According to the aforementioned criteria, Paenibacillus, Lysobacter, Nocardioides, Mesorhizobium, Cupriavidus, Rhizobium, Arthrobacter, Mycolicibacterium, Agromyces, Mycobacterium, Massilia, Microvirga, Sinorhizobium, Ensifer, and Rhodococcus are considered potential bacterial hosts. These bacteria may harbor multiple ARGs within a single cell. For instance, Rhodococcus has been found to contain rosA, ksgA, mgtA, acrA, and other ARGs simultaneously. Among these potential hosts, Paenibacillus and Massilia are identified as potential human gut microbes based on a search of the Human Gut Microbiome Database (hGMB),1 and Mycobacterium and Rhodococcus are considered potential clinical pathogens based on searches conducted using GlobalRPh.2 Furthermore, the results of the clustering analysis indicate that the potential bacterial hosts can be grouped based on time rather than fertilizer types (Figure 3). Additionally, there are no discernible distinctions in potential hosts between days 15 and 30.
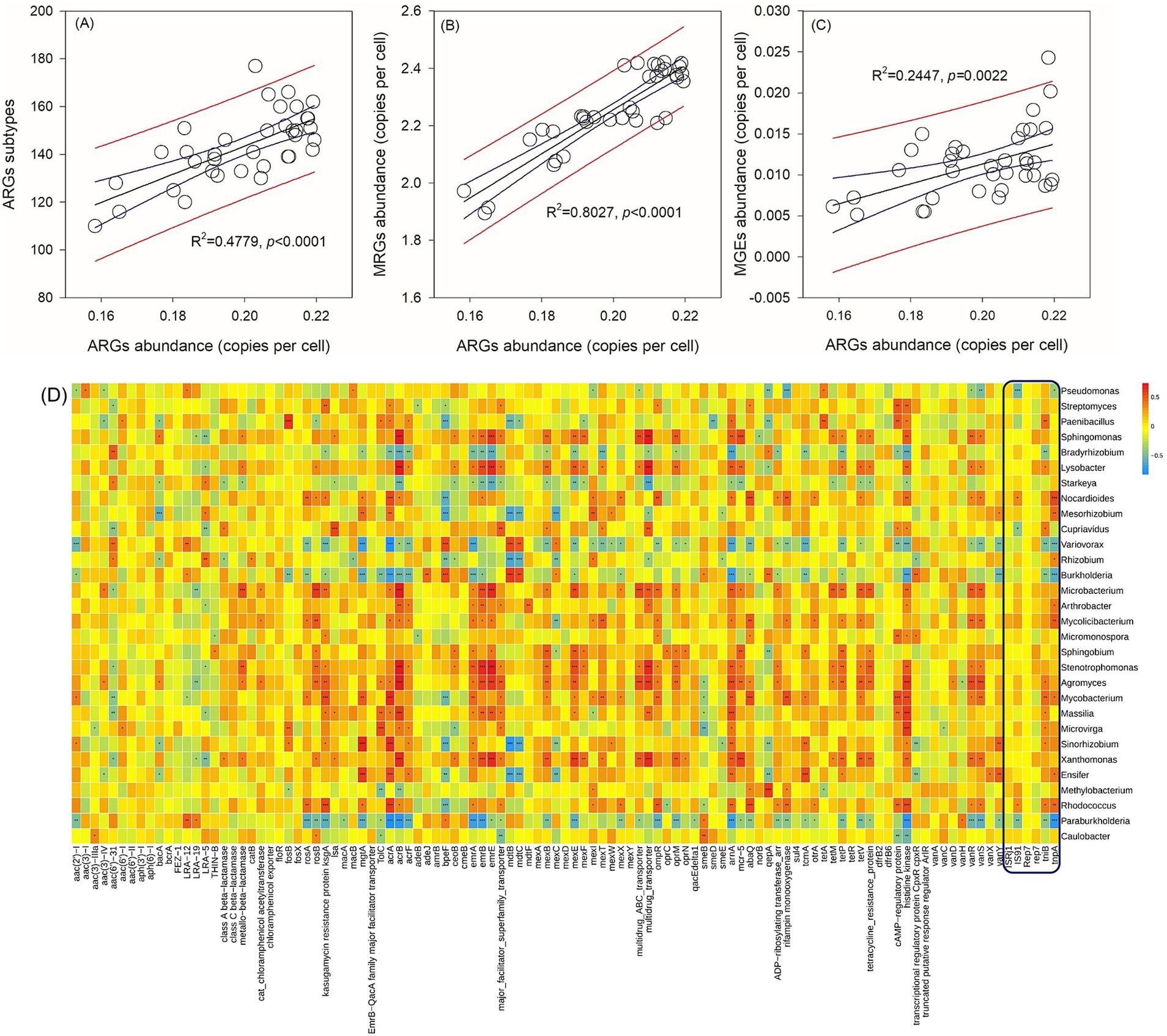
Figure 2. Correlations between core ARGs abundances and ARGs subtypes (A), core MRGs abundances (B), and core MGEs abundances (C). A correlation matrix between ARGs and bacterial genera (D). Different numbers of asterisk (*, **, ***) in panel (D) represent significant correlations using the Pearson’s correlation coefficient at p-values of 0.05, 0.01, and 0.001.
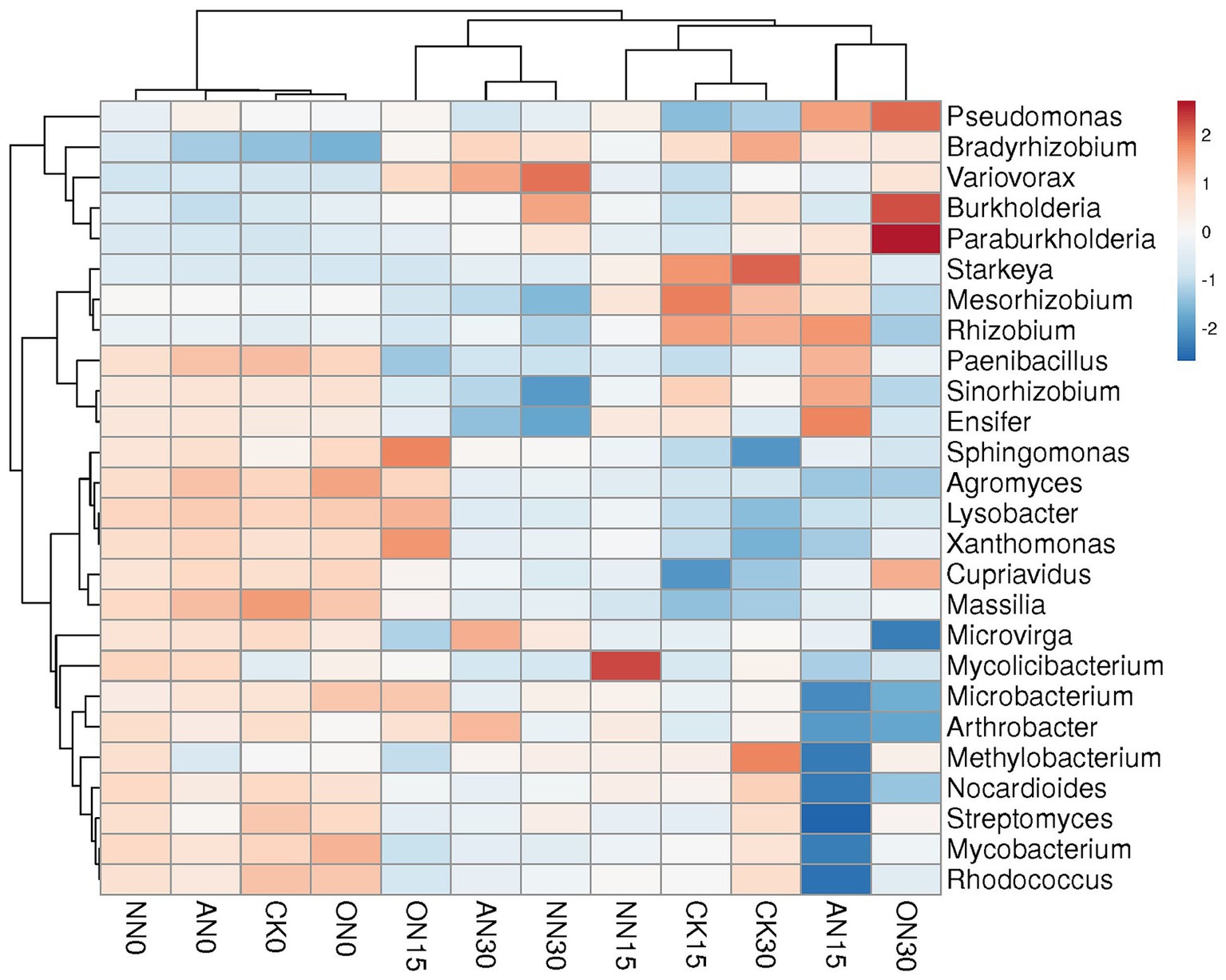
Figure 3. The heatmap of the potential bacterial hosts for core ARGs.
Changes in bacterial communities following N fertilizations with time
The bacterial communities across treatments can also be classified by time, with the exception of ON15 and CK30, which are grouped with those on day 0 and day 15, respectively (Figure 4). The upper left taxa, including Paenibacillus, Pseudomonas, and Cupriavidus, exhibit high abundance on day 0, which subsequently decreases over time (Figure 4). Conversely, Bradyrhizobium, Variovorax, and Burkholderia demonstrate relatively high abundance on day 30. The ON30 treatment displays the highest abundances of Burkholderia and Pseudomonas, while Variovorax is more abundant in NN30 and AN30. Starkeya emerges as the most abundant species in CK30. PCA reveals that all treatments can be clearly distinguished by time, with high explanatory values of PC1, albeit without significant differences (Supplementary Figures S4A–D). In contrast to ARGs, various nitrogen fertilizers have a significant impact on bacterial communities. The explanatory power of PC1 increases from 39.5% on day 0 (Supplementary Figure S3E) to 63.6% on day 15, allowing for clear differentiation of treatments through PCA (Supplementary Figure S3F). However, there is a slight decrease in explanatory power on day 30, where the AN and NN treatments are indistinguishable compared to day 15 (Supplementary Figure S3H). These findings suggest that both temporal factors and the type of fertilizer utilized can have a substantial influence on bacterial communities.
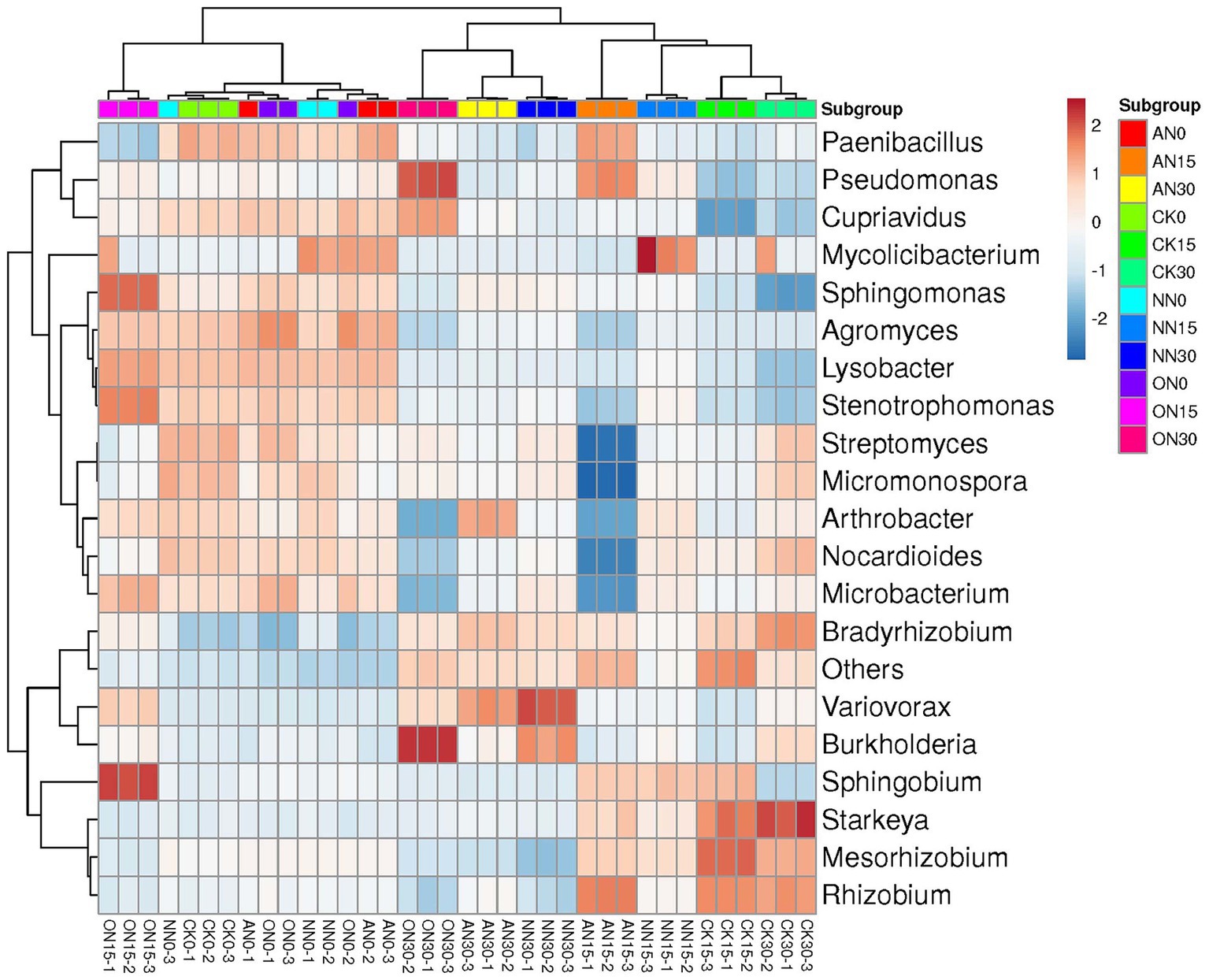
Figure 4. The heatmap of bacterial community using top 20 abundant genera. “Others” represents the remaining sum of genera.
The relationship of ARGs and NCRGs in soil
Most NCRGs, particularly NRGs, exhibit a significant correlation with ARGs (Figure 5). Within the classification of NRGs, two principal categories can be distinguished: nitrogen-fixing genes, such as nifK, nifD, and nifH, and denitrification genes, including narG, napA, nirK, norB, and nosZ. Notably, several ARGs such as rosA, rosB otrA, mexF, bacA, and vanS are found to be correlated with these NRGs. In order to assess reliability, two representative strains of potential bacterial hosts listed in Figure 3, specifically Pseudomonas aeruginosa PAO1 (NCBI Taxonomy ID 287) and Burkholderia pseudomallei (NCBI Taxonomy ID 28450), were examined in the Genome database of the National Center for Biotechnology Information (NCBI)3 following the retrieval of their annotated genomes. Analysis revealed the presence of ARGs mexB, oprM, bacA, mexF, mexD and NRGs napA, narJ, narH, narG within P. aeruginosa PAO1, all of which are documented in Figure 5. Similarly, emrB, bacA and narJ, narH, and narG were identified in B. pseudomallei.
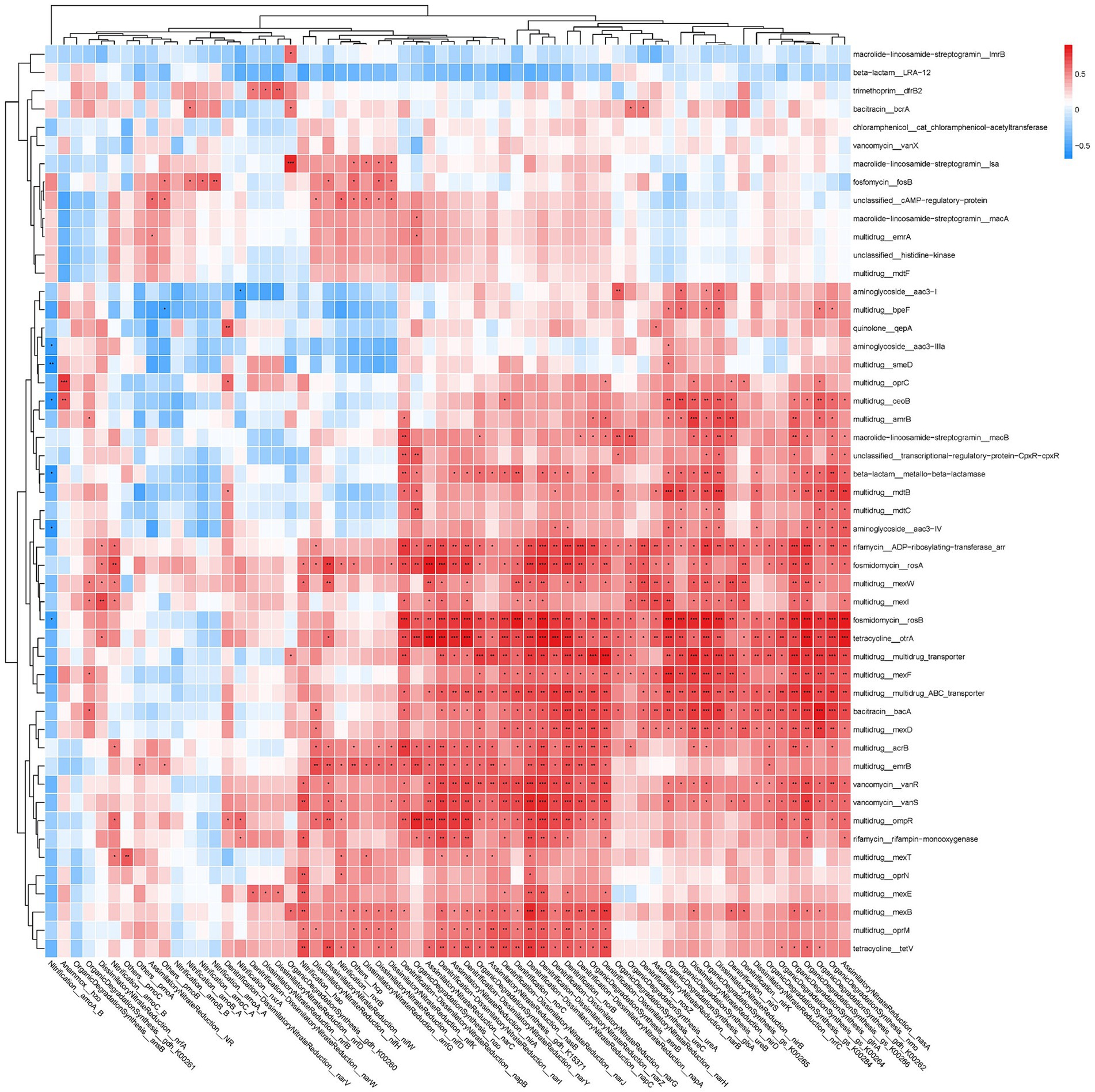
Figure 5. The correlation matrix between ARGs and nitrogen-cycling related genes (NCRGs). Different numbers of asterisk (*, **, ***) represent significant correlations using the Pearson’s correlation coefficient at p-values of 0.05, 0.01, and 0.001.
Discussion
Limited impact of N fertilization on the core ARGs profile in soil
There is ongoing debate regarding the effects of chemical fertilizer application on soil ARGs. A study demonstrated that a 15-year application of chemical fertilizers did not have a significant impact on the relative abundance of total ARGs (Wang F. et al., 2020). Additionally, a study showed that single nitrogen treatment reduced bacterial diversity and abundance, leading to the elimination of ARGs (Cui et al., 2022). However, other studies have suggested that N fertilizers can have a strong influence on the content of soil ARGs (Forsberg et al., 2014). A study demonstrated that the application of chemical fertilizer resulted in an increase in the relative abundance of ARGs in dryland soil, while decreasing their relative abundance in paddy soil (Wang F. et al., 2018). A recent studies employing metagenomic technology demonstrated that nitrogen limitation predominantly facilitated the dissemination of ARGs through HGT events (Liu et al., 2024). However, another study reported a contrasting result (Wang et al., 2024). These findings indicate that additional factors, beyond nitrogen fertilizers, may play crucial roles in the regulation of ARGs. Therefore, a control experiment is necessary to investigate the potential effects of nitrogen fertilizers on soil ARGs. The findings of a microcosmic experiment presented in this study reveal that chemical N fertilizers have varying impacts on the abundances of ARGs and MGEs, depending on their forms and the timing of sampling in the short term (30 days). While there were increases in the abundance of ARGs in treatments with AN and NN on day 30, these increases were not significantly higher compared to the CK on day 0. Overall, the application of chemical N fertilizers has limited effects on the accumulation of ARGs in soil. This finding is consistent with the composition of ARGs as reflected in PCA.
Numerous studies have indicated that the accumulation of ARGs in soil is primarily attributed to the regulation of bacterial communities by environmental factors, such as fertilizers (Liu et al., 2017; Han et al., 2018; Wang M. et al., 2018). Our research revealed that varying levels of nitrogen fertilization can lead to significant shifts in bacterial communities within a short period of 15 d, yet only have minimal effects on the abundance of ARGs. It is suggested that exogenous additives lacking ARGs and/or not participating in co-occurrence events may have limited influence on the levels of ARGs in soil. The nitrogen input rate utilized in this study was 100 mg/kg, equivalent to 2,250 kg N/ha. In the natural environment, nitrogen deposition can reach levels of up to 90 kg N/ha/year (Verma and Sagar, 2020; He et al., 2021), requiring a period of 25 years of continuous N deposition to match the rate of addition in the absence of additional N input. Previous research has indicated minimal fluctuations in ARGs during the soil maturation process over an 86-year period (Tang et al., 2021). Consequently, it is suggested that a 25-year period of N deposition may not lead to substantial variations in ARGs within the soil, potentially explaining the limited impacts observed in the present study.
Effects of N forms on ARGs profiles
Soil microbes exhibit a preference for NH4+–N over NO3−–N, contingent upon the specific types of carbon sources introduced (Romero et al., 2015). Contrary to previous findings, a study indicated that soil microbes did not exhibit a preference for either form of nitrogen (Harrison et al., 2007). NH4+–N levels experienced a rapid decline following application, stabilizing by day 15 across all treatments (Supplementary Figure S4A). Conversely, NO3−–N levels consistently remained highest in the NN treatment over time, persisting at elevated levels in both NN and ON treatments (Supplementary Figure S4B). This finding suggests that organisms with a preference for NH4+–N over NO3−–N are dominant species in this environment, likely due to limited available carbon in the soil. For instance, the prevalent species in this investigation (Romero et al., 2015), such as Pseudomonas (Zhang et al., 2019), Rhizobium (Ye et al., 2021), and Sphingomonas (Banik et al., 2007) have been shown to favor NH4+–N over NO3−–N. Therefore, the preference for N forms may contribute to significant alterations in the bacterial community. Despite this shift, there was no corresponding change in ARGs profiles. Generally, microbes exhibit a preference for inorganic N uptake over organic N in most instances (Moe, 2013). In the present investigation, the inclusion of urea yielded comparable outcomes to CK in terms of the abundance of ARGs, suggesting that urea may be a more environmentally benign N fertilizer option when compared to AN and NN. Moreover, given the relatively high N content and eco-friendly manufacturing process associated with CO2 fixation (Alper and Yuksel Orhan, 2017), controlled-release urea may serve as a viable chemical N fertilizer for agricultural applications (Tian et al., 2021).
The influence of nitrogen fertilizers on the abundance of ARGs exhibited variability. A study employing real-time quantitative PCR (qPCR) demonstrated that distinct forms of nitrogen (NH4+–N and NO3−–N) could affect ARG abundance; however, these forms were not identified as primary determinants (Wang T. et al., 2022). In a separate study, it was found that NH4+–N significantly increased ARG abundance using qPCR (Sun et al., 2020). Conversely, studies suggesting that chemical fertilizers had minimal effects on ARGs employed high-throughput quantitative PCR or metagenomic sequencing technologies (Xie et al., 2018b; Wang F. et al., 2020). Therefore, the variation in outcomes may be attributed to the divergent methodologies utilized. Conventional qPCR serves as an effective tool for promptly identifying and quantifying ARGs, facilitating the comparative analysis of antibiotic resistance, particularly for specific ARGs like tetracycline resistance genes (Su et al., 2017; Kang et al., 2022). However, this also presents a limitation of qPCR in terms of its restricted target detection capabilities. Besides, Due to the high diversity of DNA templates and complexity in environmental samples, the use of qPCR may lead to the overestimation or even false-positive detection of target genes, attributable to potential non-specific amplification (Li and Yan, 2021). By contrast, metagenomics is proposed as an effective tool for conducting a thorough investigation of environmental ARGs (Liu et al., 2019). Besides, it is strongly recommended that results be compared using consistent methodologies. For example, the abundance of the key MGE intI1 is often quantitatively assessed due to its potential involvement in HGT (Liu X. et al., 2022). However, other essential MGEs such as tnpA, tniB, and IS91 exhibit a higher co-occurrence with ARGs compared to intI1 in this study, consistent with previous findings (Li et al., 2020; Zhao et al., 2021). Such patterns can only be elucidated through the use of high-throughput technologies.
The relationship of ARGs and NCRGs in soil
Currently, there is a limited number of academic papers discussing the potential co-occurrence of ARGs with NCRGs. Previous studies have demonstrated positive correlations between the ratio of AOA/AOB amoA and soil available Cd and Cu contents (Li et al., 2009; Zhang et al., 2017). Furthermore, research has indicated that most ARGs, MRGs, and NRGs share the same host bacterial species in leachates (Wang et al., 2021). The co-occurrence of ARGs and MRGs is a common phenomenon, which is also reflected in the close relationship between ARGs and MRGs observed in this study. Our findings further demonstrated that numerous NCRGs (especially NRGs) exhibit a positive correlation with ARGs, which again suggests the existence of close relationships among ARGs, MRGs, and NRGs. A study demonstrated that a significant percentage (75.3–94.9%) of microorganisms carrying ARGs also possess NRGs, suggesting that nitrate reducing bacteria may serve as primary hosts for ARGs (Hu et al., 2022). Other studies have concluded that ARGs exhibit positive correlations with denitrification processes (Wu et al., 2017; Rahman et al., 2018; Wang et al., 2024). However, a study revealed a negative correlation between NRGs and ARG abundance (Sun et al., 2017). This discrepancy may be due to variations in methodology, as the use of qPCR for gene detection can yield differing results. Our previous research demonstrated a higher prevalence of tetracycline resistance genes in wet soils compared to dry soils (Kang et al., 2018a). It is established that denitrification processes are augmented in wet soils owing to elevated levels of NRGs (Sun et al., 2017), implying a positive correlation between ARGs and NRGs. A study confirmed that the moisture content of soil has an impact on the bioavailability of stress factors such as metals and antibiotics, leading to increased pressure in soil niches (Markowicz et al., 2021). Consequently, it is suggested that ARGs may enable denitrifying bacteria to better adapt to more challenging environments.
Conclusion
The prevalence of core ARGs in ON treatment shows similarity to CK treatment over time, suggesting that ON treatment, particularly in a controlled release dosage form, may serve as an environmentally sustainable N fertilizer. While chemical N fertilizers have minimal effects on the core ARGs profile as determined by PCA and clustering analyses, different N fertilizers can induce significant and distinct changes in bacterial communities. Nevertheless, the core ARGs’ potential bacterial hosts remain unaffected by N fertilizers, which may explain the limited impact on the core ARGs profile. The presence of nitrogen-fixing genes and denitrification genes has been found to be positively associated with ARGs such as rosA, mexF, bacA, vanS, etc., suggesting a potential risk of ARG proliferation in environments with high denitrification activity. Concerns regarding the accumulation of ARGs due to the use of chemical N fertilizers should be addressed.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
RY: Data curation, Investigation, Writing – original draft. YY: Formal analysis, Writing – original draft. MS: —. YZ: Methodology, Writing – original draft. JH: —. YK: Conceptualization, Funding acquisition, Software, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by the National Natural Science Foundation of China (42477388 and 41977112).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2024.1447782/full#supplementary-material
Footnotes
References
Alper, E., and Yuksel Orhan, O. (2017). CO2 utilization: developments in conversion processes. Petroleum 3, 109–126. doi: 10.1016/j.petlm.2016.11.003
Alster, C. J., Fischer, J. C., Allison, S. D., and Treseder, K. K. (2020). Embracing a new paradigm for temperature sensitivity of soil microbes. Glob. Change Biol. 26, 3221–3229. doi: 10.1111/gcb.15053
Banik, R., Santhiagu, A., and Upadhyay, S. (2007). Optimization of nutrients for gellan gum production by Sphingomonas paucimobilis ATCC-31461 in molasses based medium using response surface methodology. Bioresour. Technol. 98, 792–797. doi: 10.1016/j.biortech.2006.03.012
Cui, E., Fan, X., Hu, C., Neal, A. L., Cui, B., Liu, C., et al. (2022). Reduction effect of individual N, P, K fertilization on antibiotic resistance genes in reclaimed water irrigated soil. Ecotoxicol. Environ. Saf. 231:113185. doi: 10.1016/j.ecoenv.2022.113185
Daghrir, R., and Drogui, P. (2013). Tetracycline antibiotics in the environment: a review. Environ. Chem. Lett. 11, 209–227. doi: 10.1007/s10311-013-0404-8
de Nijs, E. A., Hicks, L. C., Leizeaga, A., Tietema, A., and Rousk, J. (2019). Soil microbial moisture dependences and responses to drying–rewetting: the legacy of 18 years drought. Glob. Change Biol. 25, 1005–1015. doi: 10.1111/gcb.14508
Edwards, L. A., Goldenberg, S. D., and Shawcross, D. L. (2021). Meeting the challenge of antimicrobial resistance in cirrhosis: the invisible threat that lies within. Gastroenterology 161, 413–415. doi: 10.1053/j.gastro.2021.05.043
Forsberg, K. J., Patel, S., Gibson, M. K., Lauber, C. L., Knight, R., Fierer, N., et al. (2014). Bacterial phylogeny structures soil resistomes across habitats. Nature 509, 612–616. doi: 10.1038/nature13377
García, J., García-Galán, M. J., Day, J. W., Boopathy, R., White, J. R., Wallace, S., et al. (2020). A review of emerging organic contaminants (EOCs), antibiotic resistant bacteria (ARB), and antibiotic resistance genes (ARGs) in the environment: increasing removal with wetlands and reducing environmental impacts. Bioresour. Technol. 307:123228. doi: 10.1016/j.biortech.2020.123228
Guo, T., Lou, C., Zhai, W., Tang, X., Hashmi, M. Z., Murtaza, R., et al. (2018). Increased occurrence of heavy metals, antibiotics and resistance genes in surface soil after long-term application of manure. Sci. Total Environ. 635, 995–1003. doi: 10.1016/j.scitotenv.2018.04.194
Han, X.-M., Hu, H.-W., Chen, Q.-L., Yang, L.-Y., Li, H.-L., Zhu, Y.-G., et al. (2018). Antibiotic resistance genes and associated bacterial communities in agricultural soils amended with different sources of animal manures. Soil Biol. Biochem. 126, 91–102. doi: 10.1016/j.soilbio.2018.08.018
Harrison, K. A., Bol, R., and Bardgett, R. D. (2007). Preferences for different nitrogen forms by coexisting plant species and soil microbes. Ecology 88, 989–999. doi: 10.1890/06-1018
He, W., Zhang, M., Jin, G., Sui, X., Zhang, T., and Song, F. (2021). Effects of nitrogen deposition on nitrogen-mineralizing enzyme activity and soil microbial community structure in a Korean pine plantation. Microb. Ecol. 81, 410–424. doi: 10.1007/s00248-020-01595-6
Hilaire, S. S., Chen, C., Pan, Z., Radolinski, J., Stewart, R. D., Maguire, R. O., et al. (2022). Subsurface manure injection reduces surface transport of antibiotic resistance genes but may create antibiotic resistance hotspots in soils. Environ. Sci. Technol. 56, 14972–14981. doi: 10.1021/acs.est.2c00981
Hu, J., Chen, Q., Zhong, S., Liu, Y., Gao, Q., Graham, E. B., et al. (2022). Insight into co-hosts of nitrate reduction genes and antibiotic resistance genes in an urban river of the Qinghai-Tibet plateau. Water Res. 225:119189. doi: 10.1016/j.watres.2022.119189
Huang, J., Mi, J., Yan, Q., Wen, X., Zhou, S., Wang, Y., et al. (2021). Animal manures application increases the abundances of antibiotic resistance genes in soil-lettuce system associated with shared bacterial distributions. Sci. Total Environ. 787:147667. doi: 10.1016/j.scitotenv.2021.147667
Jia, S., Bian, K., Shi, P., Ye, L., and Liu, C.-H. (2020). Metagenomic profiling of antibiotic resistance genes and their associations with bacterial community during multiple disinfection regimes in a full-scale drinking water treatment plant. Water Res. 176:115721. doi: 10.1016/j.watres.2020.115721
Kang, Y., Gu, X., Hao, Y., and Hu, J. (2016). Autoclave treatment of pig manure does not reduce the risk of transmission and transfer of tetracycline resistance genes in soil: successive determinations with soil column experiments. Environ. Sci. Pollut. Res. 23, 4551–4560. doi: 10.1007/s11356-015-5671-4
Kang, Y., Li, Q., Mei, L., Zhao, H., Bai, Y., Shen, M., et al. (2018a). Tetracycline resistance genes are more prevalent in wet soils than in dry soils. Ecotoxicol. Environ. Saf. 156, 337–343. doi: 10.1016/j.ecoenv.2018.03.041
Kang, Y., Li, Q., Yin, Z., Shen, M., Zhao, H., Bai, Y., et al. (2018b). High diversity and abundance of cultivable tetracycline-resistant bacteria in soil following pig manure application. Sci. Rep. 8:1489. doi: 10.1038/s41598-018-20050-8
Kang, Y., Xu, W., Zhang, Y., Tang, X., Bai, Y., and Hu, J. (2022). Bloom of tetracycline resistance genes in mudflats following fertilization is attributed to the increases in the shared potential hosts between soil and organic fertilizers. Environ. Sci. Pollut. Res. 29, 13292–13304. doi: 10.1007/s11356-021-16676-3
Li, D.-C., Gao, J.-F., Zhang, S.-J., Gao, Y.-Q., and Sun, L.-X. (2020). Emergence and spread patterns of antibiotic resistance genes during two different aerobic granular sludge cultivation processes. Environ. Int. 137:105540. doi: 10.1016/j.envint.2020.105540
Li, B., and Yan, T. (2021). Next generation sequencing reveals limitation of qPCR methods in quantifying emerging antibiotic resistance genes (ARGs) in the environment. Appl. Microbiol. Biotechnol. 105, 2925–2936. doi: 10.1007/s00253-021-11202-4
Li, X., Zhu, Y.-G., Cavagnaro, T. R., Chen, M., Sun, J., Chen, X., et al. (2009). Do ammonia-oxidizing archaea respond to soil cu contamination similarly asammonia-oxidizing bacteria? Plant Soil 324, 209–217. doi: 10.1007/s11104-009-9947-7
Liu, W., Cheng, Y., Guo, J., Duan, Y., Wang, S., Xu, Q., et al. (2022). Long-term manure inputs induce a deep selection on agroecosystem soil antibiotic resistome. J. Hazard. Mater. 436:129163. doi: 10.1016/j.jhazmat.2022.129163
Liu, P., Jia, S., He, X., Zhang, X., and Ye, L. (2017). Different impacts of manure and chemical fertilizers on bacterial community structure and antibiotic resistance genes in arable soils. Chemosphere 188, 455–464. doi: 10.1016/j.chemosphere.2017.08.162
Liu, Q., Jia, J., Hu, H., Li, X., Zhao, Y., and Wu, C. (2024). Nitrogen and phosphorus limitations promoted bacterial nitrate metabolism and propagation of antibiotic resistome in the phycosphere of Auxenochlorella pyrenoidosa. J. Hazard. Mater. 468:133786. doi: 10.1016/j.jhazmat.2024.133786
Liu, X., Wang, H., Li, L., Deng, C., Chen, Y., Ding, H., et al. (2022). Do microplastic biofilms promote the evolution and co-selection of antibiotic and metal resistance genes and their associations with bacterial communities under antibiotic and metal pressures? J. Hazard. Mater. 424:127285. doi: 10.1016/j.jhazmat.2021.127285
Liu, X., Xiao, P., Guo, Y., Liu, L., and Yang, J. (2019). The impacts of different high-throughput profiling approaches on the understanding of bacterial antibiotic resistance genes in a freshwater reservoir. Sci. Total Environ. 693:133585. doi: 10.1016/j.scitotenv.2019.133585
Ma, L., Li, B., Jiang, X.-T., Wang, Y.-L., Xia, Y., Li, A.-D., et al. (2017). Catalogue of antibiotic resistome and host-tracking in drinking water deciphered by a large scale survey. Microbiome 5:154. doi: 10.1186/s40168-017-0369-0
Markowicz, A., Bondarczuk, K., Cycoń, M., and Sułowicz, S. (2021). Land application of sewage sludge: response of soil microbial communities and potential spread of antibiotic resistance. Environ. Pollut. 271:116317. doi: 10.1016/j.envpol.2020.116317
Metsalu, T., and Vilo, J. (2015). ClustVis: a web tool for visualizing clustering of multivariate data using principal component analysis and heatmap. Nucleic Acids Res. 43, W566–W570. doi: 10.1093/nar/gkv468
Moe, L. A. (2013). Amino acids in the rhizosphere: from plants to microbes. Am. J. Bot. 100, 1692–1705. doi: 10.3732/ajb.1300033
Peacock, A. D., Mullen, M. D., Ringelberg, D. B., Tyler, D. D., Hedrick, D. B., Gale, P. M., et al. (2001). Soil microbial community responses to dairy manure or ammonium nitrate applications. Soil Biol. Biochem. 33, 1011–1019. doi: 10.1016/S0038-0717(01)00004-9
Rahman, M. M., Shan, J., Yang, P., Shang, X., Xia, Y., and Yan, X. (2018). Effects of long-term pig manure application on antibiotics, abundance of antibiotic resistance genes (ARGs), anammox and denitrification rates in paddy soils. Environ. Pollut. 240, 368–377. doi: 10.1016/j.envpol.2018.04.135
Romero, C. M., Engel, R., Chen, C., and Wallander, R. (2015). Microbial immobilization of Nitrogen-15 labelled ammonium and nitrate in an agricultural soil. Soil Sci. Soc. Am. J. 79, 595–602. doi: 10.2136/sssaj2014.08.0332
Shawver, S., Wepking, C., Ishii, S., Strickland, M. S., and Badgley, B. D. (2021). Application of manure from cattle administered antibiotics has sustained multi-year impacts on soil resistome and microbial community structure. Soil Biol. Biochem. 157:108252. doi: 10.1016/j.soilbio.2021.108252
Su, J.-Q., Cui, L., Chen, Q.-L., An, X.-L., and Zhu, Y.-G. (2017). Application of genomic technologies to measure and monitor antibiotic resistance in animals: genomic technologies to investigate AR in animals. Ann. N.Y. Acad. Sci. 1388, 121–135. doi: 10.1111/nyas.13296
Su, J.-Q., Wei, B., Ou-Yang, W.-Y., Huang, F.-Y., Zhao, Y., Xu, H.-J., et al. (2015). Antibiotic resistome and its association with bacterial communities during sewage sludge composting. Environ. Sci. Technol. 49, 7356–7363. doi: 10.1021/acs.est.5b01012
Sun, S., Lu, C., Liu, J., Williams, M. A., Yang, Z., Gao, Y., et al. (2020). Antibiotic resistance gene abundance and bacterial community structure in soils altered by ammonium and nitrate concentrations. Soil Biol. Biochem. 149:107965. doi: 10.1016/j.soilbio.2020.107965
Sun, Y., Qiu, T., Gao, M., Shi, M., Zhang, H., and Wang, X. (2019). Inorganic and organic fertilizers application enhanced antibiotic resistome in greenhouse soils growing vegetables. Ecotoxicol. Environ. Saf. 179, 24–30. doi: 10.1016/j.ecoenv.2019.04.039
Sun, M., Ye, M., Liu, K., Schwab, A. P., Liu, M., Jiao, J., et al. (2017). Dynamic interplay between microbial denitrification and antibiotic resistance under enhanced anoxic denitrification condition in soil. Environ. Pollut. 222, 583–591. doi: 10.1016/j.envpol.2016.10.015
Tang, X., Shen, M., Zhang, Y., Zhu, D., Wang, H., Zhao, Y., et al. (2021). The changes in antibiotic resistance genes during 86 years of the soil ripening process without anthropogenic activities. Chemosphere 266:128985. doi: 10.1016/j.chemosphere.2020.128985
Tian, X., Fan, Z., Zhao, Y., Sun, S., Li, T., Yu, N., et al. (2021). Controlled-release urea improved cotton productivity and nitrogen use efficiency in China: a meta-analysis. Agron. J. 113, 2251–2259. doi: 10.1002/agj2.20702
Tu, Q., Lin, L., Cheng, L., Deng, Y., and He, Z. (2019). NCycDB: a curated integrative database for fast and accurate metagenomic profiling of nitrogen cycling genes. Bioinformatics 35, 1040–1048. doi: 10.1093/bioinformatics/bty741
Verma, P., and Sagar, R. (2020). Effect of nitrogen (N) deposition on soil-N processes: a holistic approach. Sci. Rep. 10:10470. doi: 10.1038/s41598-020-67368-w
Wang, J., Ben, W., Zhang, Y., Yang, M., and Qiang, Z. (2015). Effects of thermophilic composting on oxytetracycline, sulfamethazine, and their corresponding resistance genes in swine manure. Environ. Sci. Proces. Impacts 17, 1654–1660. doi: 10.1039/c5em00132c
Wang, Y., Cai, J., Chen, X., Guo, B., Liu, J., Qiu, G., et al. (2024). The connection between the antibiotic resistome and nitrogen-cycling microorganisms in paddy soil is enhanced by application of chemical and plant-derived organic fertilizers. Environ. Res. 243:117880. doi: 10.1016/j.envres.2023.117880
Wang, F., Han, W., Chen, S., Dong, W., Qiao, M., Hu, C., et al. (2020). Fifteen-year application of manure and chemical fertilizers differently impacts soil ARGs and microbial community structure. Front. Microbiol. 11:62. doi: 10.3389/fmicb.2020.00062
Wang, M., Liu, P., Xiong, W., Zhou, Q., Wangxiao, J., Zeng, Z., et al. (2018). Fate of potential indicator antimicrobial resistance genes (ARGs) and bacterial community diversity in simulated manure-soil microcosms. Ecotoxicol. Environ. Saf. 147, 817–823. doi: 10.1016/j.ecoenv.2017.09.055
Wang, T., Sun, S., Xu, Y., Waigi, M. G., Odinga, E. S., Vasilyeva, G. K., et al. (2022). Nitrogen regulates the distribution of antibiotic resistance genes in the soil-vegetable system. Front. Microbiol. 13:848750. doi: 10.3389/fmicb.2022.848750
Wang, P., Wu, D., You, X., Su, Y., and Xie, B. (2021). Antibiotic and metal resistance genes are closely linked with nitrogen-processing functions in municipal solid waste landfills. J. Hazard. Mater. 403:123689. doi: 10.1016/j.jhazmat.2020.123689
Wang, M., Xie, X., Wang, M., Wu, J., Zhou, Q., and Sun, Y. (2020). The bacterial microbiota in florfenicol contaminated soils: the antibiotic resistome and the nitrogen cycle. Environ. Pollut. 259:113901. doi: 10.1016/j.envpol.2019.113901
Wang, F., Xu, M., Stedtfeld, R. D., Sheng, H., Fan, J., Liu, M., et al. (2018). Long-term effect of different fertilization and cropping systems on the soil antibiotic resistome. Environ. Sci. Technol. 52, 13037–13046. doi: 10.1021/acs.est.8b04330
Wang, L., Yang, C., Yang, Z., Chen, Y., Wang, F., Li, W., et al. (2022). Shortcut nitrification and denitrification shed light on simultaneous removal of conventional contaminants and antibiotic resistance genes (ARGs). J. Environ. Chem. Eng. 10:106925. doi: 10.1016/j.jece.2021.106925
Wood, D. E., and Salzberg, S. L. (2014). Kraken: ultrafast metagenomic sequence classification using exact alignments. Genome Biol. 15:R46. doi: 10.1186/gb-2014-15-3-r46
Wu, D., Chen, G., Zhang, X., Yang, K., and Xie, B. (2017). Change in microbial community in landfill refuse contaminated with antibiotics facilitates denitrification more than the increase in ARG over long-term. Sci. Rep. 7:41230. doi: 10.1038/srep41230
Xie, W.-Y., Shen, Q., and Zhao, F. J. (2018a). Antibiotics and antibiotic resistance from animal manures to soil: a review. Eur. J. Soil Sci. 69, 181–195. doi: 10.1111/ejss.12494
Xie, W.-Y., Yuan, S.-T., Xu, M.-G., Yang, X.-P., Shen, Q.-R., Zhang, W.-W., et al. (2018b). Long-term effects of manure and chemical fertilizers on soil antibiotic resistome. Soil Biol. Biochem. 122, 111–119. doi: 10.1016/j.soilbio.2018.04.009
Yang, F., Shen, S., Gao, W., Ma, Y., Han, B., Ding, Y., et al. (2022). Deciphering discriminative antibiotic resistance genes and pathogens in agricultural soil following chemical and organic fertilizer. J. Environ. Manag. 322:116110. doi: 10.1016/j.jenvman.2022.116110
Ye, L., Yang, P., Zeng, Y., Li, C., Jian, N., Wang, R., et al. (2021). Rhizobium symbiosis modulates the accumulation of arsenic in Medicago truncatula via nitrogen and NRT3.1-like genes regulated by ABA and linalool. J. Hazard. Mater. 415:125611. doi: 10.1016/j.jhazmat.2021.125611
Yin, X., Chen, X., Jiang, X.-T., Yang, Y., Li, B., Shum, M. H.-H., et al. (2023). Toward a universal unit for quantification of antibiotic resistance genes in environmental samples. Environ. Sci. Technol. 57, 9713–9721. doi: 10.1021/acs.est.3c00159
Yin, X., Jiang, X. T., Chai, B., Li, L., Yang, Y., Cole, J. R., et al. (2018). ARGs-OAP v2.0 with an expanded SARG database and hidden Markov models for enhancement characterization and quantification of antibiotic resistance genes in environmental metagenomes. Bioinformatics 34, 2263–2270. doi: 10.1093/bioinformatics/bty053
Zeng, J., Liu, X., Song, L., Lin, X., Zhang, H., Shen, C., et al. (2016). Nitrogen fertilization directly affects soil bacterial diversity and indirectly affects bacterial community composition. Soil Biol. Biochem. 92, 41–49. doi: 10.1016/j.soilbio.2015.09.018
Zhang, M., Bai, S. H., Tang, L., Zhang, Y., Teng, Y., and Xu, Z. (2017). Linking potential nitrification rates, nitrogen cycling genes and soil properties after remediating the agricultural soil contaminated with heavy metal and fungicide. Chemosphere 184, 892–899. doi: 10.1016/j.chemosphere.2017.06.081
Zhang, N., Chen, H., Lyu, Y., and Wang, Y. (2019). Nitrogen removal by a metal-resistant bacterium, Pseudomonas putida ZN1, capable of heterotrophic nitrification–aerobic denitrification. J. Chem. Technol. Biotechnol. 94, 1165–1175. doi: 10.1002/jctb.5863
Zhang, Q.-Q., Ying, G.-G., Pan, C.-G., Liu, Y.-S., and Zhao, J.-L. (2015). Comprehensive evaluation of antibiotics emission and fate in the river basins of China: source analysis, multimedia modeling, and linkage to bacterial resistance. Environ. Sci. Technol. 49, 6772–6782. doi: 10.1021/acs.est.5b00729
Zhao, R., Feng, J., Huang, J., Li, X., and Li, B. (2021). Reponses of microbial community and antibiotic resistance genes to the selection pressures of ampicillin, cephalexin and chloramphenicol in activated sludge reactors. Sci. Total Environ. 755:142632. doi: 10.1016/j.scitotenv.2020.142632
Zhu, D., Ma, J., Li, G., Rillig, M. C., and Zhu, Y.-G. (2022). Soil plastispheres as hotspots of antibiotic resistance genes and potential pathogens. ISME J. 16, 521–532. doi: 10.1038/s41396-021-01103-9
Keywords: antibiotics resistance genes, bacterial community, chemical nitrogen fertilizer, soil metagenomic sequencing, core resistome
Citation: You R, Yu Y, Shen M, Zhang Y, Hong J and Kang Y (2024) Applications of different forms of nitrogen fertilizers affect soil bacterial community but not core ARGs profile. Front. Microbiol. 15:1447782. doi: 10.3389/fmicb.2024.1447782
Edited by:
Abhijeet Mishra, University of Delhi, IndiaCopyright © 2024 You, Yu, Shen, Zhang, Hong and Kang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yijun Kang, eWprYW5nQHljdHUuZWR1LmNu