
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 03 July 2024
Sec. Microbial Physiology and Metabolism
Volume 15 - 2024 | https://doi.org/10.3389/fmicb.2024.1426882
 Tim Böer1
Tim Böer1 Miriam Antonia Schüler1
Miriam Antonia Schüler1 Alina Lüschen1Lena Eysell1Jannina Dröge1Melanie Heinemann1Lisa Engelhardt2
Alina Lüschen1Lena Eysell1Jannina Dröge1Melanie Heinemann1Lisa Engelhardt2 Mirko Basen2
Mirko Basen2 Rolf Daniel1
Rolf Daniel1 Anja Poehlein1*
Anja Poehlein1*Due to their metabolic versatility in substrate utilization, acetogenic bacteria represent industrially significant production platforms for biotechnological applications such as syngas fermentation, microbial electrosynthesis or transformation of one-carbon substrates. However, acetogenic strains from the genera Terrisporobacter and Acetoanaerobium remained poorly investigated for biotechnological applications. We report the isolation and characterization of four acetogenic Terrisporobacter strains and one Acetoanaerobium strain. All Terrisporobacter isolates showed a characteristic growth pattern under a H2 + CO2 atmosphere. An initial heterotrophic growth phase was followed by a stationary growth phase, where continuous acetate production was indicative of H2-dependent acetogenesis. One of the novel Terrisporobacter isolates obtained from compost (strain COMT) additionally produced ethanol besides acetate in the stationary growth phase in H2-supplemented cultures. Genomic and physiological characterizations showed that strain COMT represented a novel Terrisporobacter species and the name Terrisporobacter vanillatitrophus is proposed (=DSM 116160T = CCOS 2104T). Phylogenomic analysis of the novel isolates and reference strains implied the reclassification of the T. petrolearius/T. hibernicus phylogenomic cluster to the species T. petrolearius and of the A. noterae/A. sticklandii phylogenomic cluster to the species A. sticklandii. Furthermore, we provide first insights into active prophages of acetogens from the genera Terrisporobacter and Acetoanaerobium.
The genus Terrisporobacter was described in 2014 with Terrisporobacter glycolicus as the type species (Gerritsen et al., 2014). T. glycolicus DSM 1288T (formerly, Clostridium glycolicum) was the first Terrisporobacter isolate obtained in 1962 from the mud of a stagnant pond near the Chesapeake and Ohio canal (United States). Cells were described as strict anaerobic, motile, Gram-positive and endospore-forming rods. The bacterium was of particular interest due to its ethylene glycol (Gaston and Stadtman, 1963) and cinnamic acid degrading capabilities (Chamkha et al., 2001). In 1991, the first acetogenic Terrisporobacter species was described, Terrisporobacter mayombei DSM 6539T (formerly, Clostridium mayombei). The bacterium was isolated from the gut of the soil-feeding termite Cubitermes speciosus and was shown to grow acetogenically with H2 + CO2 (Kane et al., 1991). In addition to T. mayombei, the only other acetogenic Terrisporobacter isolates described comprised T. glycolicus DSM 13865 isolated from sewage sludge (Ohwaki and Hungate, 1977) and ATCC 29797 isolated from roots of sea grass (Küsel et al., 2001). The species Terrisporobacter petrolearius JCM 19845T, isolated in 2015 from a petroleum reservoir sample from an oilfield in Shengli (China) (Deng et al., 2015), and Terrisporobacter hibernicus NCTC 14625T, isolated in 2023 from bovine feces, were both described as non-acetogenic species (Mitchell et al., 2023). The genus Terrisporobacter has been found strongly associated with manure (Chen et al., 2022) and was shown to represent a key player in the degradation of manure by fermenting carbohydrates to H2 + CO2 and acetate (Budianto and Yasin, 2024). Correspondingly, biotechnological applications of bacteria from the genus Terrisporobacter have focused on their H2-producing capabilities. In contrast, biotechnological applications based on acetogenesis have hitherto been neglected. As CO is not being utilized by acetogenic Terrisporobacter strains an application as biocatalyst for the transformation of CO-rich industrial waste gases is unlikely. Bacteria from the genus Terrisporobacter are most likely to be employed as biocatalysts in the process of microbial electrosynthesis (MES) similar to the acetogenic genus Sporomusa (Aryal et al., 2017; Madjarov et al., 2022). The MES biotechnology allows to fix CO2 via acetogenesis and to transform electricity into biocommodities by extracellular electron transfer (Thulluru et al., 2023). The electricigenic potential of Terrisporobacter cells was already indicated by their positive correlation to bioelectricity generation in microbial fuel cells (Li et al., 2018) and their presence at the cathode of MES cells (Wang et al., 2023). Although every Terrisporobacter genome sequenced to date fulfilled the genetic prerequisites for acetogenesis, the conversion of H2 + CO2 to acetate was demonstrated only for a few strains. Non-acetogenic Terrisporobacter strains were hypothesized to have retained the Wood-Ljungdahl pathway (WLP) as an electron sink during the fermentation of amino acids via the Stickland reaction similar to Clostridioides difficile (Köpke et al., 2013; Gencic and Grahame, 2020; Böer et al., 2023). The genus Acetoanaerobium was formed in 1985 with the description of the acetogen Acetoanaerobium noterae ATCC 35199T. The bacterium was described to ferment H2 + CO2 to acetate as the sole fermentation product (Sleat et al., 1985). The first bacterium isolated from the genus Acetoanaerobium was the non-acetogenic bacterium Acetoanaerobium sticklandii DSM 519T (formerly, Clostridium sticklandii) in 1956 from a formate enrichment culture (Stadtman and McClung, 1957). A. sticklandii was reclassified in 2016 to the genus Acetoanaerobium (Galperin et al., 2016). In 2015, a third Acetoanaerobium species was described, Acetoanaerobium pronyense. A. pronyense DSM 27512T was isolated from a carbonate chimney of the Prony Hydrothermal Field in Prony Bay (New Caledonia) and together with A. sticklandii described to not transform H2 + CO2 to acetate (Bes et al., 2015). Similar to the genus Terrisporobacter, the biotechnological applications of bacteria from the genus Acetonaerobium were restricted to the degradation of protein-rich wastes to H2. A. sticklandii was shown to ferment amino acids to acetate, butyrate, acetone, butanol and ethanol as products (Sangavai and Chellapandi, 2019). Similar to Terrisporobacter, Acetoanaerobium noterae is a potential electrotroph as it was enriched at the cathode of MES cells (Wang et al., 2023). In this study, we present novel isolates of the genus Terrisporobacter and Acetoanaerobium and assess their potential for biotechnological applications by genomic and physiological characterizations.
We performed enrichments with different environmental samples comprising digested sludge sampled from a wastewater treatment plant in 2019 and 2022, horse feces, river marsh sediment and compost. Environmental samples were suspended (50% w/v) in PBS-buffer (NaCl, 8 g/L; KCl, 0.2 g/L; Na2HPO4, 1.42 g/L; KH2PO4, 0.24 g/L) by shaking at 50 rpm for 1 h on a horizontal shaker (Adolf Kühner, Birsfelden, Swiss). Enrichment cultures were inoculated with 20 μL of this solution. For the enrichments with digested sludge from 2019, horse feces and river marsh sediment, the inocula were additionally pasteurized at 80°C for 10 min. H2-supplemented enrichment cultures were performed with the digested sludge sampled in 2019, horse feces and marsh sediment using a modified DSM 311c medium substituting sulfate salts with chloride salts, exchanging casitone with peptone and omitting fructose. The detailed recipe of the medium is provided in Supplementary Table S1. The medium (100 mL) was dispensed in sterile anaerobic 500 mL Afnor flasks (Ochs, Bovenden, Germany). The atmosphere of the flasks was changed from N2/CO2 (80%/20%) to H2/CO2 (66%/33%) by gassing for 2 min and adding an overpressure of 2 bar. H2-supplemented enrichment cultures were incubated at 35°C for 7 days. Subsequently, isolates were obtained by plating 50 μL of the enrichment cultures to agar plates (1.5% w/v agar) of the modified DSM 311c medium supplemented with 60 mM glucose in an anaerobic chamber. Colonies were randomly picked after 2 days of incubation at 35°C and transferred to fresh agar plates with a dilution streak. Colonies were picked from the dilution streaks and an initial taxonomic classification of the isolates was performed by colony PCR. Therefore, single colonies were suspended in 100 μL PBS-buffer and the 16S rRNA gene sequence was amplified with a DreamTaq polymerase (Thermo Fisher Scientific, Waltham, MA, United States) using 0.5 μL of the dissolved colonies as the template and the primers 08F (5′-AGAGTTTGATCCTGGC-3′) and 1504R (5′-TACCTTGTTACGACTT-3′) following the standard instructions of the manufacturer. The PCR product was purified using the Master Pure Complete DNA & RNA Purification Kit (Epicentre, Madison, United States) and Sanger sequenced by Seqlab (Microsynth, Göttingen, Germany). Isolates identified to be affiliated to bacterial genera described to contain acetogenic strains were chosen for further investigations. Another enrichment was performed with digested sludge sampled in 2022 using the modified DSM 311c medium supplemented with additional 30 mM dimethylglycine (DMG) and omitting peptone. The enrichment with DMG was incubated at 35°C for 4 days and the isolation was performed as described for the H2-supplemented enrichments. The compost enrichment was conducted with Brain Heart Infusion (BHI) broth (Oxoid, Hampshire, United Kingdom) supplemented with the CDMN selective supplement (Code: SR0173) (Oxoid, Hampshire, United Kingdom) containing yeast extract (5 g/L), L-Cys-HCl (0.05 g/L), moxalactam (32 mg/L) and norfloxacin (12 mg/L) (CDMN-medium) (Aspinall and Hutchinson, 1992). The CDMN enrichment cultures were incubated at 37°C and the subsequent isolation was performed with CDMN-medium and 1.5% w/v agar performing the same dilution streak method as described for the H2-supplemented enrichments.
DNA isolation and Illumina sequencing were performed as described by Böer et al. (2023). Nanopore sequencing was performed by library preparation with 1.5 μg high-molecular weight DNA using the ligation sequencing kit 1D (SQK-LSK109) and the native barcode expansion kit (EXP-NBD104) as recommended by the manufacturer (Oxford Nanopore, Oxford, United Kingdom). Nanopore sequencing was conducted for 72 h employing the MinION device Mk1B, the SpotON flow cell R9.4.1 and the MinKNOW software (v21.6.0) following the instructions of the manufacturer (Oxford Nanopore). Demultiplexing and base calling of Nanopore sequencing data were performed with the Guppy software in HAC mode (v5.0.11). Long-read de novo genome assemblies were performed following the suggested Trycycler workflow for bacterial genome assemblies by Wick et al. (2021). Briefly, quality control and adapter trimming of paired-end Illumina sequences was performed with Fastp (v0.23.4) (Chen et al., 2018) and Trimmomatic (v0.39; LEADING: 3, TRAILING: 3, SLIDINGWINDOW:4:15, MINLEN:50) (Bolger et al., 2014). Porechop (v0.2.4) was used for adapter trimming of Nanopore reads, which were subsequently filtered with Filtlong (v0.2.1) to a minimal read length of 1 kb discarding the worst 5% of sequences in terms of quality. Nanopore reads were subsampled 24 times with Trycycler (v0.5.4) (Wick et al., 2021) and eight subsets each were used as input for the assemblers Flye (v2.9.2) (Kolmogorov et al., 2019), Canu (v2.2) (Koren et al., 2017) and Raven (v1.8.3) (Vaser and Šikić, 2021). Assemblies were combined to a single consensus sequence with Trycycler and the consensus sequence was polished with the full Nanopore data using Medaka (v1.10.0) and finally polished with the processed short-read sequences using Polypolish (v0.5.0) (Wick and Holt, 2022). Genomes were reorientated to start with the dnaA gene using the fixstart function of Circlator (v1.5.5) (Hunt et al., 2015). Genome annotations were performed with Prokka (v1.14.6) (Seemann, 2014) and quality assessment of the final genome assemblies was conducted with BUSCO (v5.5.0) (Manni et al., 2021) using the reference datasets clostridia_odb for Terrisporobacter isolates and clostridiales_odb for the Acetoanaerobium isolate. Phylogenomic analysis of isolates was performed with genomes of the genus Terrisporobacter and Acetoanaerobium using pyani (v0.2.12) (Pritchard et al., 2016) and the MUMmer alignment option (Marçais et al., 2018) determining average nucleotide identities (ANIm). Additionally, digital DNA–DNA hybridization values (dDDH) of the formula d0, d4 and d6 were obtained by using the Genome to Genome Distance Calculator (GGDC) (v3.0) (Meier-Kolthoff et al., 2022). Assembly accession numbers of analyzed reference genomes are listed in Supplementary Table S2. Bacterial phylogenomic trees were obtained by using the Type (Strain) Genome Server (TYGS) (Meier-Kolthoff and Göker, 2019). Unique orthologous groups (OGs) were identified by orthology analysis using Proteinortho (v6.3.0) (Lechner et al., 2011) with an identity threshold of ≥50% and an e-value of ≤1e−10. Unique OGs and ANIm/dDDH heatmaps were visualized in R with the ggplot2 package (v3.4.1) (Wickham, 2016). Sequencing results from the prophage activity screening were mapped on to the respective reference genomes with bowtie2 (v2.5.0) (Langmead and Salzberg, 2012) and visualized with the BRIG software (v0.95) (Alikhan et al., 2011). Mapping files from bowtie2 were converted to coverage graphs with the respective BRIG module using a window size of 1 and setting the maximum coverage graph value to 1,000 (for isolates ELB, COM and DSL2) or to 4,900 (isolate DSL). Predictions for genomic islands and prophages were added from PHASTEST (Wishart et al., 2023) and IslandViewer4 (Bertelli et al., 2017). Regions showing activity in the screening were extracted as nucleotide sequences and annotated with Pharokka (Bouras et al., 2023). Phage genomes were rotated to begin with the large terminase subunit using the corresponding Pharokka function. NCBI Virus and PhageScope were employed as phage reference databases, the latter database was also used as a tool for the taxonomy, host and lifestyle predictions of phage genomes (Wang et al., 2024). Viral phylogenomic trees were obtained by using the Virus Classification and Tree Building Online Resource (VICTOR) (Meier-Kolthoff and Göker, 2017).
Obtained isolates were routinely cultivated in the modified DSM 311c medium with glucose (30 mM) and inoculated to a start OD600 of 0.1. Growth temperature optima of the isolates were investigated first by incubation at temperatures ranging from 10 to 55°C in 5°C intervals. NaCl and pH optima were investigated subsequently at the respective temperature optimum of the strains. NaCl optima were determined by cultivation with added NaCl concentrations ranging from 0 to 7% in 1% intervals and additionally at 0.1%. pH optima were obtained by setting the pH to 5.5 to 9 in 0.5 intervals using HCl and NaOH. Gram-staining was carried out after 12 h of incubation during the exponential growth phase following the Claus protocol (Claus, 1992). Substrate utilization was evaluated by cultivation in Hungate tubes adding substrates from sterile anaerobic stock solutions in a final concentration of 30 mM. Vanillate (5 mM), syringate (5 mM) or starch (0.5 g/L) were added to the media before autoclaving. Growth was measured from triplicate cultures in Hungate tubes after 22 h of incubation at the temperature optimum of the respective strain. Basal growth was assessed by inoculating medium without substrate as control and substrate utilization was considered positive when substrate addition yielded at least an increase in OD600 of 0.05. Substrates not being used in the initial 22 h of incubation were continuously assessed for four more days before substrate utilization was considered negative. Utilization of H2 + CO2 of the novel isolates was assessed by cultivation of the isolates as described for the H2-supplemented enrichment cultures. Cultures with 2 bar overpressure under a N2 + CO2 atmosphere were used as a control. Growth was measured after 17, 24, 41, 48, 65, 72, 89, and 96 h of incubation under the optimal physiological conditions determined for the respective strain. Fermentation products were analyzed by gas chromatography as described by Baum et al. (2024). Cellular fatty acid analysis was performed with the identification service provided by the German Collection of Microorganisms and Cell Cultures (DSMZ, Braunschweig. Germany). The prophage activity screening for spontaneously released prophages was performed with all novel isolates grown to the late exponential phase. Bacterial cells were removed by centrifugation at 2,600 g for 5 min and subsequent sterile filtration of the supernatant with a 0.45 μm filter (Sarstedt, Nümbrecht, Germany). Remaining cells and host DNA were removed by the addition of 20 μL of lysozyme (100 mg/mL) (Serva, Heidelberg, Germany), 20 μL of DNase I (1 U/μL) (Epicentre, Madison, United States) and 20 μL of RNase A (5 μg/μL) (Epicentre, Madison, United States) and incubation at 37°C overnight. The following day the solution was centrifuged at 36,000 g for 2 h at room temperature in a DU PONT OTD 50B ultracentrifuge using the Ti-60 rotor (Thermo Fisher Scientific, Waltham, United States). The supernatant was discarded and the resulting phage pellet was suspended in 100 μL of TKM-buffer (10 mM Tris–HCl (pH 7.5), 10 mM KCl and 1 mM MgCl2) and stored at 4°C. Prior to the DNA isolation, DNase I was inactivated by incubation at 75°C for 10 min. DNA isolation from the dissolved phage particles and sequencing was performed with the protocol as described for bacterial genome sequencing. A description of the general principle of prophage activity screenings by sequencing of particle-protected DNA can be found in Hertel et al. (2015) and Schüler et al. (2024).
Enrichments were performed with the inocula listed in Table 1. We obtained one isolate from each of the five different inocula used in this study. Based on the initially analyzed 16S rRNA gene sequences the isolates DSL, ELB, HSE, and COM were associated to the genus Terrisporobacter and the isolate DSL2 was associated to the genus Acetoanaerobium.

Table 1. Isolates obtained from the different environmental samples and the corresponding enrichment and isolation strategy.
Genome sequencing and annotation data of the isolated organisms are summarized in Table 2. The assemblies for the isolates HSE, ELB, DSL, and COM yielded chromosomes with a GC-content of 29% and sizes ranging from 4.015–4.115 Mb. Additionally, circular plasmids with sizes ranging from 5.581–48.861 kb could be found. Assembly of the DSL2 genome yielded one circular replicon with a GC-content of 34% and a size of 2.633 Mb.
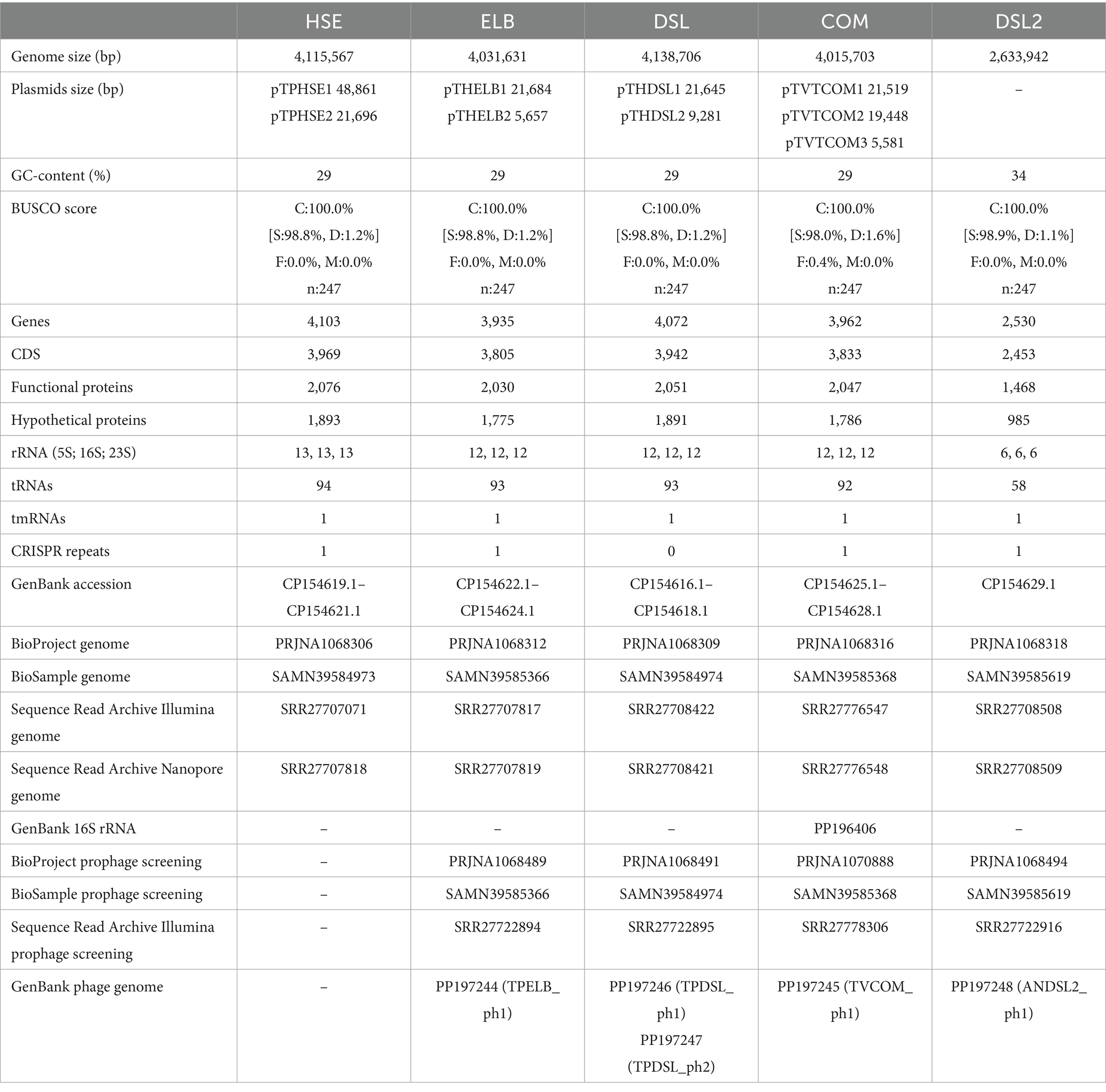
Table 2. Genome and annotation data of the obtained isolates.
The genomes of isolates HSE, ELB, DSL, and COM were compared to genome sequences of the genus Terrisporobacter (Figure 1A) and that of DSL2 to genomes of Acetoanaerobium (Figure 1B) by ANIm and dDDH (d4) analyses in order to obtain a phylogenomic classification. Complete GGDC results including the three dDDH formulas d0, d4, d6 and their respective confidence intervals are summarized in Supplementary Table S3. The ELB and DSL genomes were more identical to the type strain of T. hibernicus genome (ANIm: 98.4 and 98.9%, dDDH (d4): 85.3 and 89.4%) than to the type strain genome of T. petrolearius (ANIm: 96.3 and 96.3%, dDDH (d4): 68.7 and 68.8%). The HSE genome showed similar identities to the type strains genomes of T. hibernicus (ANIm: 96.4%, dDDH (d4): 69.9%) and T. petrolearius (ANIm: 96%, dDDH (d4): 66.6%). The genome of the COM isolate showed highest sequence identity to the type strain genome of T. mayombei DSM 6539T (ANIm: 92.8%, dDDH (d4): 49.3%) and T. glycolicus DSM 1288T (ANIm: 92.1%, dDDH (d4): 46.3%). The DSL2 genome was most identical to the type strain genome of A. noterae (ANIm: 97.7%, dDDH (d4): 78.6%). The genome of A. sticklandii DSM 519T showed the highest sequence identities to that of the A. noterae type strain (ANIm: 96.5%, dDDH (d4): 70.0%) and the DSL2 isolate (ANIm: 96.5%, dDDH (d4): 70.1%). Whole genome-based phylograms created by TYGS identified T. mayombei DSM 6539T as the closest relative of the COM isolate and assigned the novel Terrisporobacter isolates HSE, ELB and DSL in a phylogenomic group together with the strains T. glycolicus FS03, WW3900, KPPR-9, T. petrolearius UN03-225, JCM 19845T and T. hibernicus NCTC 14625T (Figure 2A). The Acetonaerobium isolate DSL2 was placed into a phylogenomic group with A. noterae ATCC 35199T and A. sticklandii DSM 519T (Figure 2B).
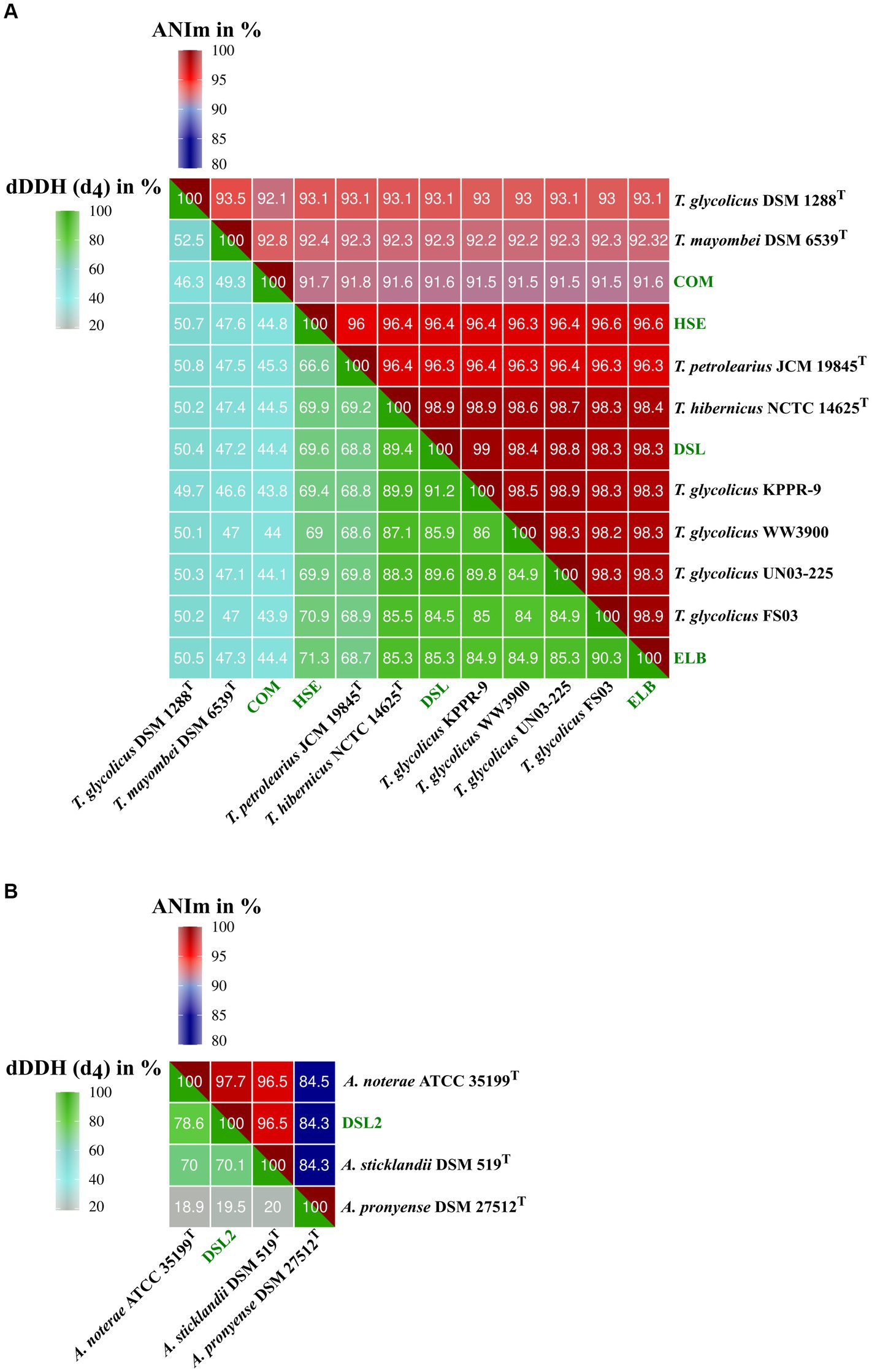
Figure 1. Phylogenomic analysis of novel Terrisporobacter and Acetoanaerobium isolates. The heatmaps visualize ANIm and dDDH values for the genomes of Terrisporobacter isolates HSE, ELB, DSL and COM (A) and the Acetoanaerobium isolate DSL2 genome (B). The top part of the heatmaps visualize the ANIm percentage identity values from 80–100% and the lower part of the heatmaps visualize dDDH values (d4) from 20–100%. The legend for the color code is shown on top (ANIm) and to the left (dDDH). Novel isolates from this study are highlighted in green.
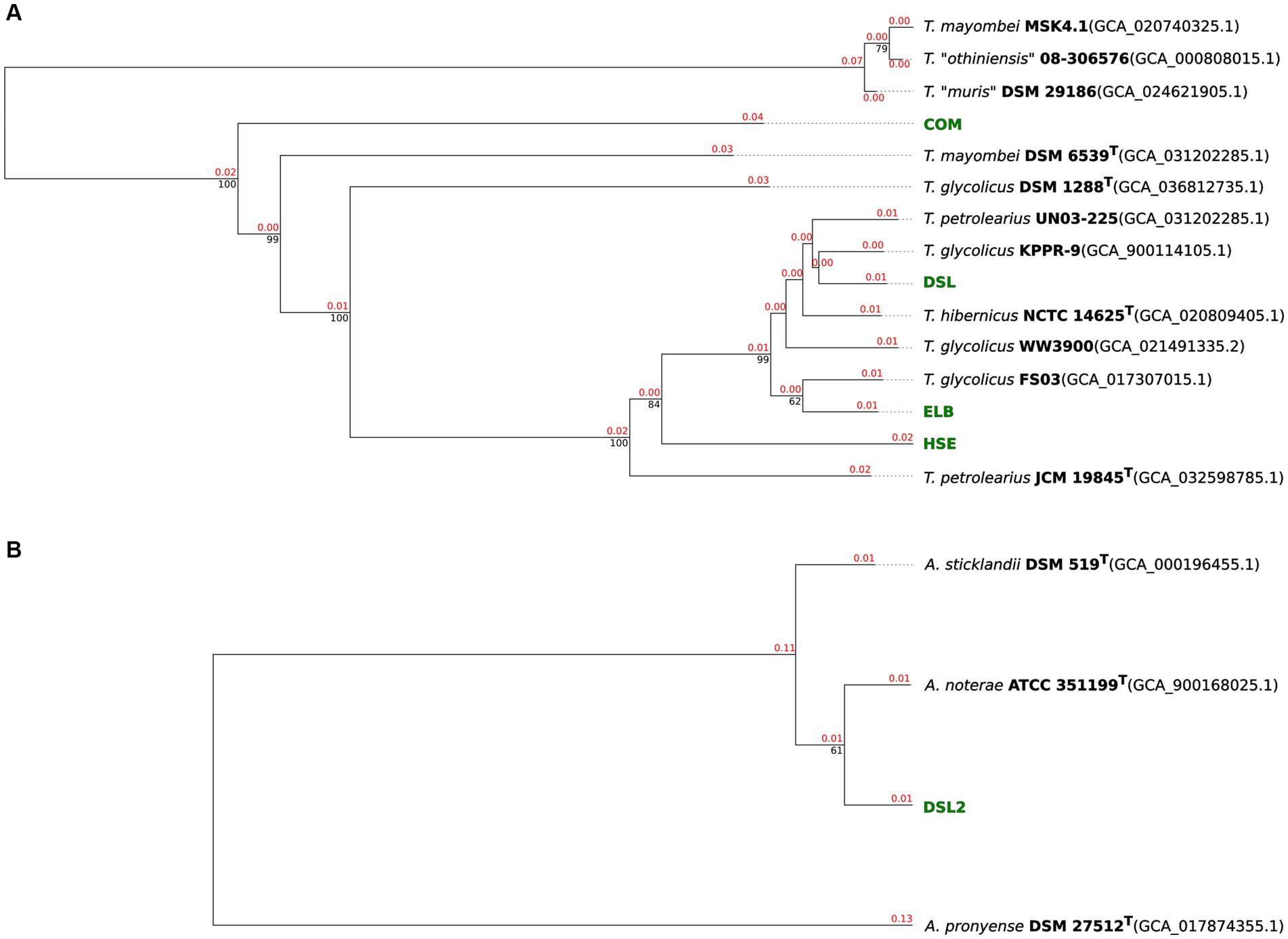
Figure 2. Phylograms of novel Terrisporobacter and Acetoanaerobium isolates. Whole-genome sequence-based phylogenetic tree of novel Terrisporobacter isolates (A). Branch lengths are scaled in terms of GBDP distance formula d5. Red numbers above branches are GBDP pseudo-bootstrap support values >60% from 100 replications, with an average branch support of 73.9%. Black numbers below branches are confidence scores. The tree was rooted at the midpoint. Whole-genome sequence-based phylogenetic tree of the novel Acetoanaerobium isolate (B). Branch lengths are scaled in terms of GBDP distance formula d5. Red numbers above branches are GBDP pseudo-bootstrap support values >60% from 100 replications, with an average branch support of 61.0%. Black numbers below branches are confidence scores. The tree was rooted at the midpoint.
After species assignment of all novel isolates, we performed a pan/core genome analysis to identify potential novel physiological traits. Encoded proteins of Terrisporobacter reference genomes were clustered into orthologous groups (OGs) and compared to the novel Terrisporobacter isolates HSE, ELB, DSL, and COM (Figure 3A). Similarly, the OGs of encoded proteins by Acetoanaerobium reference genomes were compared to those of the DSL2 genome (Figure 3B). OGs detected in only one genome were assigned as unique OGs and colored corresponding to the species assignments of our phylogenomic classification. The genus Terrisporobacter formed a pan genome of 9,867 OGs with a core genome of 800 OGs. All Terrisporobacter genomes encoded a WLP-gene cluster, a potential HDCR-complex, an electron-bifurcating hydrogenase (HydABC), an electron-bifurcating Sporomusa-type Nfn transhydrogenase (StnABC) and a Rnf-complex (RnfABCDEG). The highest number of unique OGs were detected for isolate COM (599), followed by T. mayombei MSK4.1 (398), T. “muris” DSM 29186 (382), T. petrolearius JCM 19845T (336), T. glycolicus DSM 1288T (275), isolate HSE (263) and isolate DSL (229). The lowest number of unique OGs were identified for T. glycolicus WW3900 (171), T. hibernicus NCTC14625T (162), isolate ELB (148), T. “othiniensis” 08-306,576 (135), T. petrolearius UN03 (132), T. glycolicus KPPR-9 (125) and T. glycolicus FS03 (94). The genome of isolate COM contained unique OGs for ArgBCDJ (TVTCOM_02650-02680), as well as ArgG (TVTCOM_21250), ArgF (TVTCOM_20250) and ArgH (TVTCOM_31940). Furthermore, four unique OGs were identified encoding the subunits PorABCD (TVTCOM_25810-25840) of a pyruvate ferredoxin oxidoreductase (PFOR). Other unique OGs encoded by the COM genome comprised subunits of a dissimilatory sulfite reductase (TVTCOM_04940-04960), proteins for the utilization of mannitol (TVTCOM_07370-07410) and 1,2-propanediol (TVTCOM_08460-08620), the subunits GrdH and GrdI of a betaine reductase (TVTCOM_13260-13270), proteins for the import of aliphatic sulfonates (TVTCOM_13580-13600 and TVTCOM_33140-33160), and proteins for the import and export of siderophores for the acquisition of iron (TVTCOM_18490-18530). The plasmid pTVTCOM2 encoded unique uviA and uviB genes (TVTCOM_39450-39460). The pan genome of the genus Acetoanaerobium comprised 4,591 OGs including the core genome of 1,387 OGs. All Acetoanaerobium genomes encoded an electron-bifurcating hydrogenase with three subunits (HydABC), a Rnf-complex and a PFOR gene. With the exception of A. pronyense DSM 27512T, every genome encoded a WLP-gene cluster and an electron-bifurcating Sporomusa-type Nfn transhydrogenase (StnABC). The highest number of unique OGs was identified for A. pronyense DSM 27512T (1,335) followed by A. noterae ATCC 35199T (337), A. sticklandii DSM 519T (333) and isolate DSL2 (146).
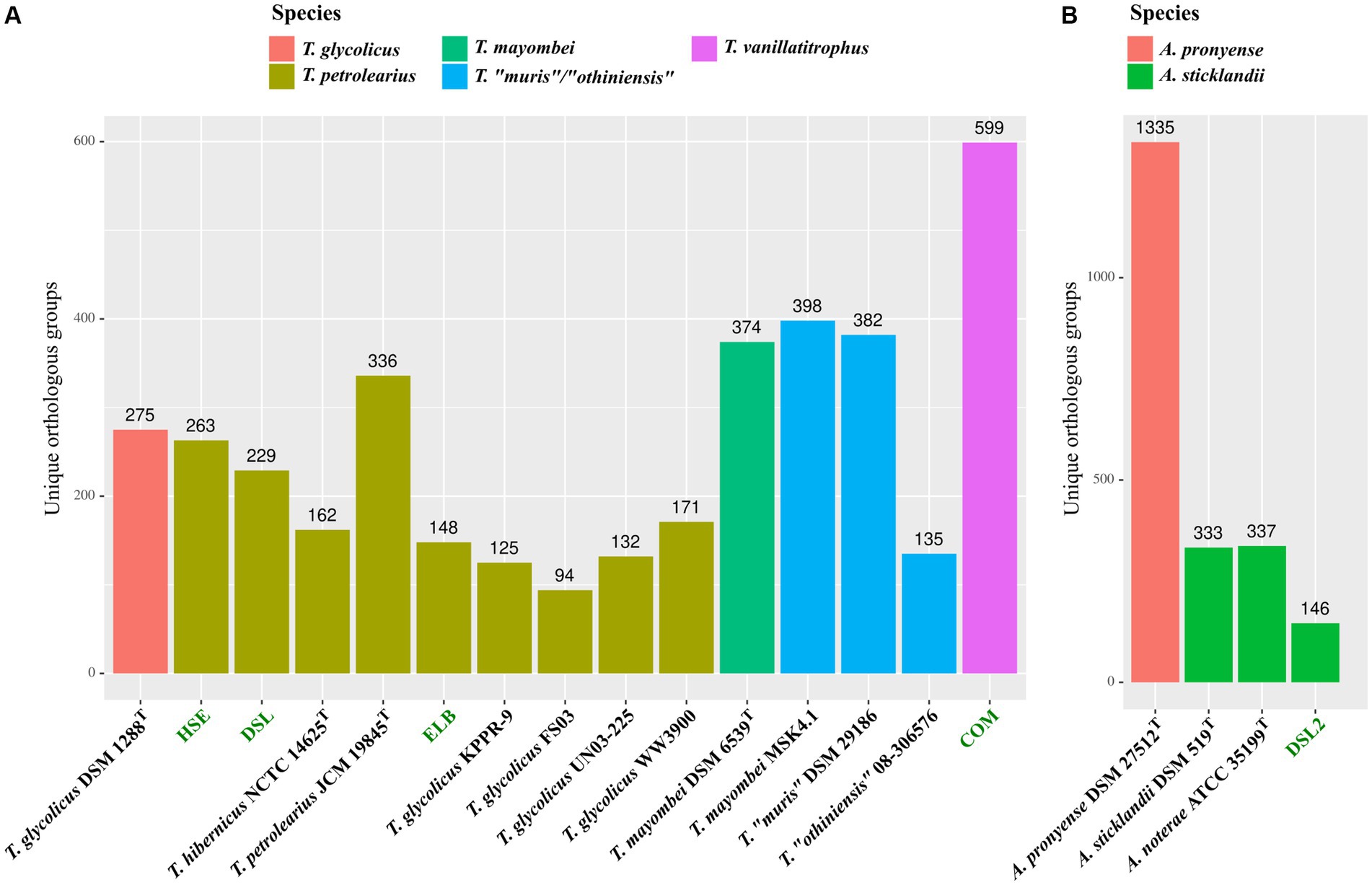
Figure 3. Pan/core genome analysis of novel isolates. Detected unique OGs in the novel and reference Terrisporobacter (A) and Acetoanaerobium (B) genomes colored by species. Novel isolates from this study are highlighted in green and the species color code is shown on the top.
All novel isolates grew only under strict anoxic conditions and no growth occurred when yeast extract was omitted. Growth of the isolates HSE, ELB, DSL and COM was also considerably improved by the addition of peptone. The rod-shaped cells stained Gram-positive and endospore formation was observed by light microscopy. In contrast, the rod-shaped cells of the DSL2 isolate stained Gram-negative and endospore formation was not detected by light microscopy. Cellular fatty acid analysis of isolate COM is summarized in Supplementary Table S4. The predominant cellular fatty acids identified were C16:0 (19%), C16:0 DMA (12.4%), C16:1 ∆9 (7.3%) and C16:1 ∆7 (6%).
Isolate-specific growth optima are shown in Table 3. All isolates grew at temperatures between 20–50°C. HSE and ELB showed the lowest temperature optimum at 25°C, while the DSL isolate grew best at 30°C. The highest temperature optima were determined for the COM and DSL2 isolate at 35°C. The DSL and DSL2 isolates tolerated added NaCl concentrations between 0 to 5% and 0 to 7%, thereby showing the broadest range of NaCl tolerance. However, all isolates grew optimally at low added NaCl concentrations ranging from 0 to 0.1%. With a pH of 8.5 and 8, the HSE and ELB isolate showed the highest pH optima in comparison to the other isolates growing optimally at pH 7 and 7.5. The substrate utilization tests yielded specific characteristics for every isolate (Table 4). The isolates HSE, ELB, DSL and COM all utilized fructose, glucose, sorbitol, maltose, raffinose and melezitose. None of these isolates utilized arabinose, ribose, galactose, mannose, rhamnose, glucuronic acid, myo-inositol, lactose, melibiose, dextran, starch, pyruvate, lactate, malate, citrate, DMG, betaine, syringate, methanol or ethanolamine. ELB was the only isolate utilizing trehalose for growth, while DSL was the only isolate growing with glycerol. Xylose fermentation was only observed for the DSL and COM isolates. Mannitol, cellobiose, formate, vanillate and 1,2-propanediol utilization were identified as unique substrates utilized by the COM isolate. Glucose was fermented to acetate and ethanol as the only fermentation products by the COM isolate. The isolate DSL2 utilized glucose, maltose, fructose, sorbitol, sucrose, trehalose, melibiose, raffinose, melezitose and dextrin for growth.
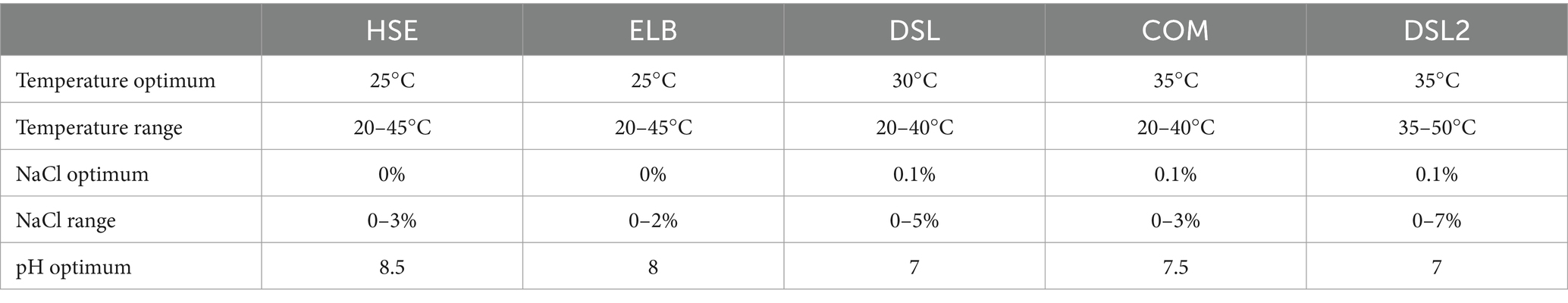
Table 3. Determined physiological optima of the novel Terrisporobacter isolates HSE, ELB, DSL, and COM and the novel Acetoanaerobium isolate DSL2.
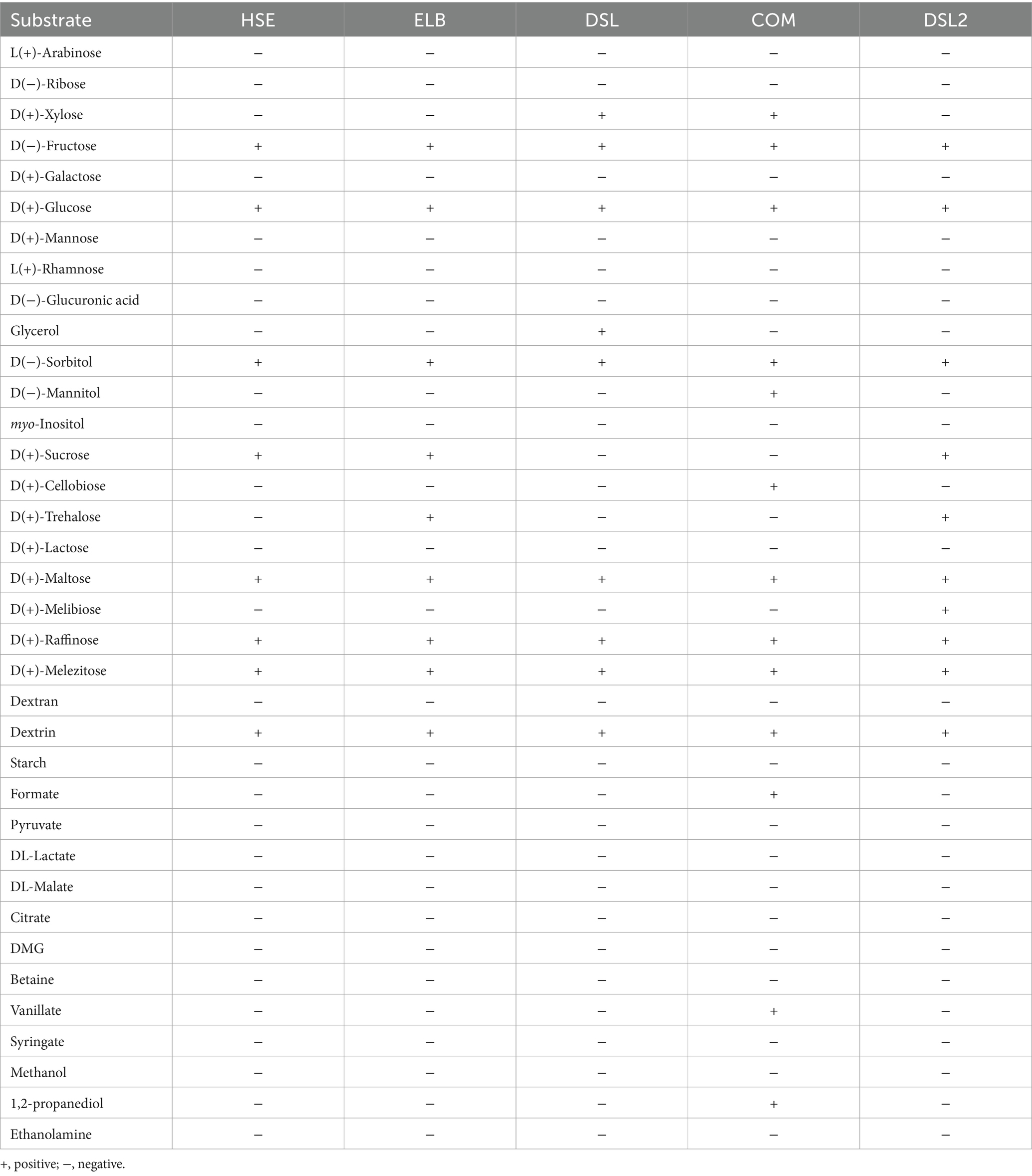
Table 4. Substrate utilization of the novel Terrisporobacter isolates HSE, ELB, DSL, and COM and the novel Acetoanaerobium isolate DSL2.
The Terrisporobacter isolates HSE, DSL and COM as well as the Acetoanaerobium isolate DSL2 grew in the initial 17 h of incubation faster in the N2 + CO2 control cultures than under an H2 + CO2 atmosphere. Only the Terrisporobacter isolate ELB achieved a higher OD600 after 17 h with H2 + CO2 than in the control. Nevertheless, all isolates achieved their maximal OD600 within 48 h of incubation under H2 + CO2 and the N2 + CO2 control (Figure 4; Supplementary Figure S1). The Terrisporobacter isolates HSE, ELB, DSL and COM achieved higher maximal growth in H2 + CO2 cultures (OD600: 0.401, 0.345, 0.442 and 0.37) than in control cultures with an atmosphere of N2 + CO2 (OD600: 0.343, 0.327, 0,431 and 0.31). Furthermore, the Terrisporobacter isolates HSE, ELB, DSL and COM produced considerably higher amounts of acetate after 96 h of incubation in cultures with a H2 + CO2 atmosphere (31.60 mM, 33.38 mM, 45.69 mM and 54.58 mM,) than in control cultures with an atmosphere of N2 + CO2 (17.44 mM, 19.07 mM, 23.42 mM and 15.64 mM). The Terrisporobacter isolate COM was the only isolate showing small amounts of ethanol (up to 2.92 mM) besides the main product acetate under a H2 + CO2 atmosphere. Ethanol production was not observed in N2 + CO2 control cultures and occurred in H2 + CO2 cultures mainly with the onset of the stationary growth phase, simultaneous to the production of acetate (Figure 4). The Acetoanaerobium isolate DSL2 showed considerably lower maximal growth values in H2 + CO2 cultures (OD600: 0.231) than in the N2 + CO2 control cultures (OD600: 0.379). DSL2 produced similar amounts of acetate after 96 h of incubation under a H2 + CO2 atmosphere (21.54 mM) than under the N2 + CO2 control atmosphere (24.36 mM) (Supplementary Figure S1). Growth of DSL2 in the N2 + CO2 control cultures stagnated after the initial 17 h of incubation, while H2 + CO2 cultures of DSL2 continued to grow until 48 h of incubation. The GC-measurements of all isolates under a H2 + CO2 and a N2 + CO2 atmosphere showed besides the main products acetate and ethanol trace amounts of other products. The concentration of these trace amounts did not considerably differ between cultures of both atmospheres and did not change over the course of incubation. These products were not further investigated.
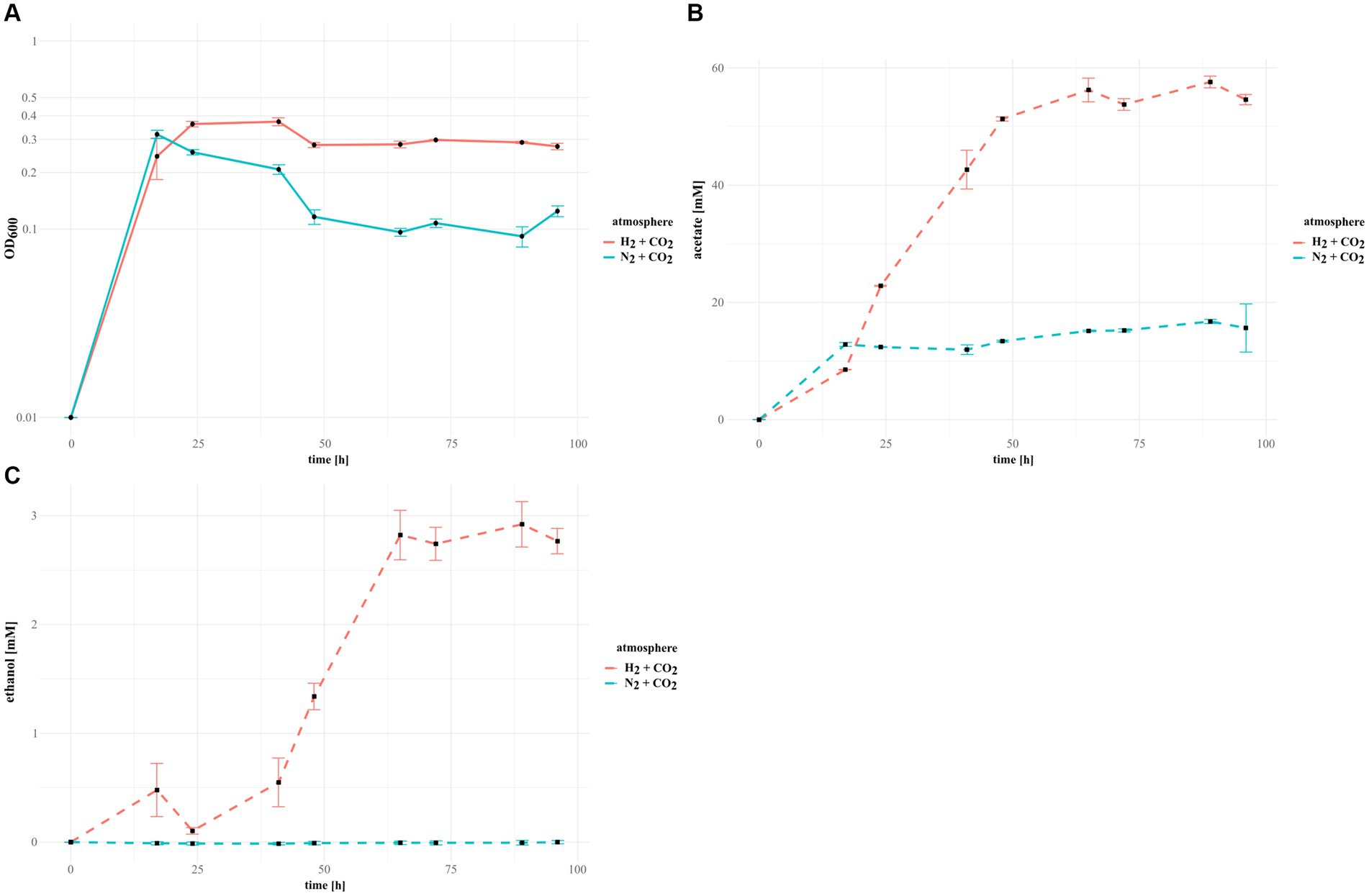
Figure 4. Growth in H2-supplemented cultures and control cultures of the novel Terrisporobacter isolate COM. Measured OD600 growth values for COM cultures grown with an atmosphere of H2 + CO2 (solid red line) and control cultures with an atmosphere of N2 + CO2 (solid blue line) (A). GC-measurements detecting the amount of produced acetate (B) and ethanol (C) in H2 + CO2 (dotted red line) and N2 + CO2 (dotted blue line) cultures.
We performed a prophage activity screening for all of our novel isolates, in order to identify potential growth inhibitions by prophage activity. Isolate DSL showed two active prophage regions (Supplementary Figure S2). The first active prophage region (TPDSL_ph1) was 55 Kb in size and matched to the prediction of a complete prophage by PHASTEST and a genomic island predicted by IslandViewer4, while the second active prophage region (TPDSL_ph2) was 37 Kb in size and matched to another complete prophage region predicted by PHASTEST. Isolate ELB showed one active prophage region (TPELB_ph1) with a size of 36 Kb matching one of the two predicted prophage regions by PHASTEST (Supplementary Figure S3). A proportion of this active region was also predicted as a genomic island by IslandViewer4. Isolate COM showed one active prophage region (TVCOM_ph1) with a size of 42 Kb. The position aligned with two adjacent regions predicted as incomplete and complete prophages by PHASTEST (Supplementary Figure S4). Isolate HSE did not yield phage DNA in our activity screening. Isolate DSL2 showed an active prophage region with a size of 36 Kb (ANDSL2_ph1) matching the only predicted region of a complete prophage by PHASTEST (Supplementary Figure S5). All prophage genomes were annotated to encode genes assigned to the function classes of transcription regulation, lysis, integration and excision, head and packaging, connector and tail. Furthermore, every prophage region encoded genes of the class DNA/RNA and nucleotide metabolism. However, TPDSL_ph1 was the only prophage region encoding a DNA invertase flanked on both sides by an integrase. The TPDSL_ph1 prophage region was also the only region encoding a γ-glutamyl cyclotransferase assigned to the class moron, auxiliary metabolic gene and host takeover. The γ-glutamyl cyclotransferase was preceded by two genes annotated as amidoligase, which were also only identified in the TPDSL_ph1 prophage. We did not detect significant hits to reference genomes of phage isolates in the viral databases of NCBI Virus or PhageScope for our novel phage genomes. BLASTp search of the sequence of the large terminase subunit of the novel phage genomes identified several hits to metagenomic assembled phage genomes (Table 5). PhageScope predicted the novel phage genomes and metagenomic assembled phage genomes to be affiliated to the class of Caudoviricetes. Interestingly, Clostridioides difficile was the only host predicted to also belong to the Peptostreptococcaceae family, like Terrisporobacter or Acetoanaerobium. The phages TPELB_ph1, TVCOM_ph1, TPDSL_ph2 and ANDSL2_ph1 were all predicted to employ a temperate lifestyle, only TPDSL_ph1 was predicted to be a virulent bacteriophage. The whole genome-based phylogram created by VICTOR placed the potential relatives into the same phylogenomic group in all cases, with the only exception of the TVCOM_ph1 genome not clustering with the BK017189 phage genome (Figure 5). The phylogenomic groups of TPELB_ph1, TVCOM_ph1, TPDSLph2 and ANDSL2_ph1 were assigned to the same phage family of different phage genera, while the phylogenomic group of TPDSL_ph1 was assigned to another phage family.
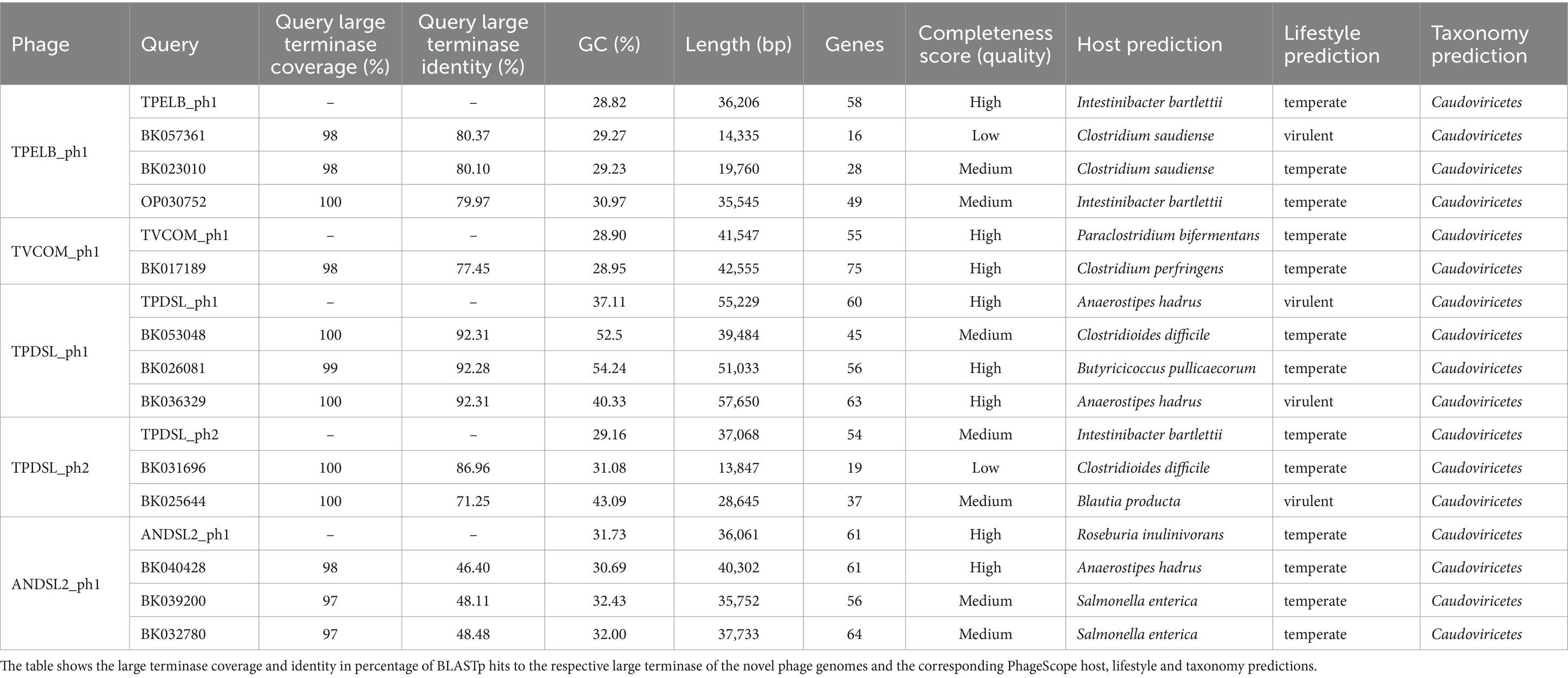
Table 5. Related prophage genomes of the novel phage genomes derived from the prophage activity screening identified by BLASTp search of the protein sequence of the large terminase subunit.
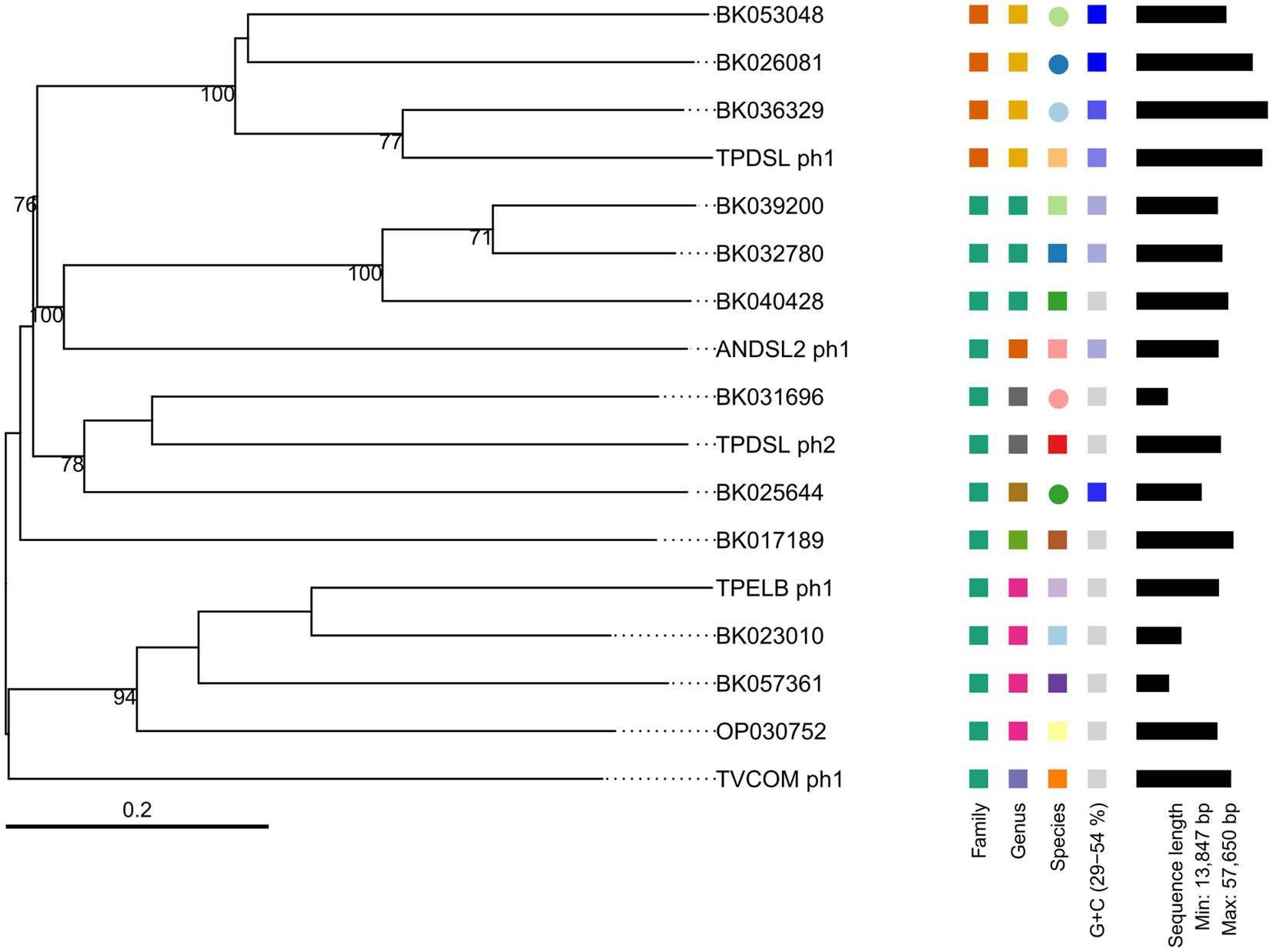
Figure 5. Phylogram of novel Terrisporobacter and Acetoanaerobium phage genomes and related phages. Branch lengths are scaled in terms of GBDP distance formula d0 and the black numbers below branches are confidence scores yielding an average branch support of 71%. The tree was rooted at the midpoint.
The Terrisporobacter isolates HSE, ELB, DSL, and COM showed genome sizes of 4 to 4.1 Mb and a GC-content of 29%, which is in line with the available genomes of other Terrisporobacter members. Plasmids were reported for the circular high-quality genomes of T. hibernicus NCTC14625T, T, glycolicus DSM 1288T, T. mayombei DSM 6539T and T. petrolearius JCM 19845T (Böer et al., 2023; Mitchell et al., 2023). The plasmids pTPHSE2 (21.696 Kb), pTPELB1 (21.684 Kb), pTPDSL1 (21.645 Kb) and pTVTCOM1 (21.519 Kb) showed the genetic structure of the Terrisporobacter-specific plasmids encoding genes for the proline-dependent reductive branch of the Stickland reaction (Böer et al., 2023). The pTVTCOM2 plasmid of T. vanillatitrophus encoded uviA/B genes, which were shown to be involved in the UV-inducible production of bacteriocin Bcn5 in Clostridium perfringens or the production of toxins in Clostridioides difficile, Clostridium tetani or Clostridium botulinum (Dupuy et al., 2005). Other plasmids detected in the Terrisporobacter isolates were unique for the respective isolate and only hypothetical proteins were encoded. ANIm analysis showed a phylogenomic cluster applying a species threshold of 95% (Goris et al., 2007) of the Terrisporobacter strains HSE, JCM 19845T, NCTC 14625T, DSL, KPPR-9, WW3900, UN03-225, FS03 and ELB with ANIm values ranging from 96.0 to 99.0%. The dDDH (d4) comparisons between those Terrisporobacter strains confirmed the phylogenomic cluster with values ranging from 84.0 to 91.2%, with the exception of the strains HSE and JCM 19845T where multiple comparisons fell below the dDDH species threshold of 70%. In such cases it is recommended to consider the confidence intervals of the in silico dDDH (d4) predictions (Meier-Kolthoff et al., 2013). These ranged across the species threshold of 70% with confidence scores ranging from 63.6 to 74.1% for the dDDH (d4) values of strains HSE and JCM 19845T (Supplementary Table S3). Furthermore, as we are comparing circular high-quality genomes, we can also consult the dDDH values calculated by the d0 and d6 formula considering the genome length, and not only the d4 formula, which performs most robust for draft genomes as it is independent on genome lengths (Meier-Kolthoff et al., 2013). The dDDH values of strains HSE and JCM 19845T compared to the other strains of the phylogenomic cluster ranged from 70.4 to 85.1% for formula d0 and 72.0 to 85.1% for formula d6 supporting the phylogenomic clustering of the ANIm analysis (Supplementary Table S3). Therefore, we propose to reclassify and summarize the Terrisporobacter isolates HSE, JCM 19845T, NCTC 14625T, DSL, KPPR-9, WW3900, UN03-225, FS03 and ELB under one species name. As the species T. petrolearius was described prior to T. hibernicus, we propose to classify all genomes and the corresponding strains of this phylogenomic cluster as T. petrolearius. The closest relative of the COM isolate was T. mayombei DSM 6539T but the ANIm and dDDH values remained under the corresponding species thresholds in all comparisons. Hence, the COM isolate represented a novel Terrisporobacter species, which was further supported by the 599 unique OGs detected in this isolate and the whole genome-based phylogram created by TYGS. The name Terrisporobacter vanillatitrophus is proposed for the COM isolate. Several unique OGs of the COM isolate allowed to infer unique physiological characteristics of the novel Terrisporobacter species. For example, the unique OGs of ArgBCDFGHJ indicate a functional ornithine-based synthesis of arginine in T. vanillatitrophus, which was identified as incomplete in the genome-based metabolic analysis of the type strains of T. glycolicus, T. petrolearius and T. mayombei (Böer et al., 2023). The PFOR links catabolism and anabolism during autotrophic growth in acetogens and is encoded by one fused gene in every other Terrisporobacter genome (Furdui and Ragsdale, 2000; Katsyv et al., 2021; Böer et al., 2023). In contrast, besides the fused PFOR (TVTCOM_10920), the COM isolate encodes an additional PFOR encoded by four separate genes, which was hitherto reported to be characteristically present in methanogenic archaea (Blamey and Adams, 1993; Ma et al., 1997) or hyperthermophilic bacteria (Blamey and Adams, 1994; Eram et al., 2015). Besides the additional PFOR identified for the COM isolate, every Terrisporobacter isolate fulfilled all prerequisites for H2/CO2-dependent acetogenesis in line with the metabolic model described for the Terrisporobacter genus based on the type strains of T. mayombei, T. petrolearius and T. glycolicus (Böer et al., 2023). The model comprises a complete WLP-gene cluster, a potential HDCR-complex, an electron-bifurcating hydrogenase (HydABC), an electron-bifurcating Sporomusa-type Nfn transhydrogenase (StnABC) and a Rnf-complex (RnfABCDEG).
The Acetoanaerobium isolate DSL2, with a genome size of 2.6 Mb, was smaller than the type strains of A. noterae (2.8 Mb), A. sticklandii (2.7 Mb) and A. pronyense (3 Mb). The GC-content of 34% was slightly higher than that of other Acetoanaerobium genomes (33%). The closest relative of DSL2 was identified by ANIm and dDDH analyses as the type strain of A. noterae. Both strains, together with the type strain of A. sticklandii, formed a phylogenomic cluster with ANIm values ranging from 96.5 to 97.7% and dDDH (d4) values ranging from 70 to 78.6%. As these values fall into the respective species threshold of 95 and 70%, respectively, this phylogenomic cluster was identified to correspond to the same bacterial species. As A. sticklandii was described before A. noterae, we propose to reclassify all strains of this phylogenomic cluster to the species A. sticklandii. The only available Acetoanaerobium genome analyzed falling under the species thresholds of ANIm and dDDH was A. pronyense DSM 27512T, forming its own phylogenomic cluster. The structure of the WLP-gene cluster in all strains of the phylogenomic cluster of A. sticklandii resembled the structure described for different Clostridium or Terrisporobacter species (Poehlein et al., 2015; Böer et al., 2023), with the only difference that the gene encoding the bifunctional cyclohydrolase/dehydrogenase (FolD) was not encoded as part of the WLP-cluster, but located in another genomic region.
Members of the genus Terrisporobacter were described as anaerobic and Gram-positive bacteria, forming rod-shaped and motile cells producing terminal endospores (Gerritsen et al., 2014). These characteristics were met by all four obtained isolates assigned to the genus Terrisporobacter. Growth dependence on the addition of yeast extract or peptone has been described for the type strains of T. mayombei and T. glycolicus (Kane et al., 1991; Chamkha et al., 2001). A. noterae was described to form rod-shaped Gram-negative cells without endospore formation (Sleat et al., 1985), which was in line with the observed morphology of the DSL2 isolate and the isolation from a non-pasteurized enrichment culture. Although A. noterae was reported to stain Gram-negative, cells were described to form a two-layered cell wall structure atypical for Gram-negative bacteria (Sleat et al., 1985). A. sticklandii and A. pronyense were both described as Gram-positive bacteria (Stadtman and McClung, 1957; Bes et al., 2015).
The major cellular fatty acids detected in T. vanillatitrophus COM showed considerable differences to the composition of other Terrisporobacter type strains. With 19% of C16:0, the COM isolate contained lower proportions than T. mayombei (26.3%), T. petrolearius (23.6%) and T. hibernicus (22.8%). T. glycolicus contained similar proportions of C16:0 with 18.5%. The other major cellular fatty acids C16:1 ∆9 (7.3%) and C16:1 ∆7 (6%) detected for the COM isolate were identified in similar abundances for the other four Terrisporobacter type strains. DMA fatty acids were not reported by Deng et al. (2015) and Mitchell et al. (2023) for the type strains of T. glycolicus, T. mayombei, T. petrolearius and T. hibernicus (Deng et al., 2015). The proposal of the genus Terrisporobacter in 2014 by Gerritsen et al. reported lower values for C16:0 DMA for the type strains of T. glycolicus (9.3%) and T. mayombei (8.3%) than we identified in the COM isolate (Gerritsen et al., 2014). In summary, the major cellular fatty acid analysis of the COM isolate supported the classification as a novel Terrisporobacter species.
The physiological optima and substrates utilized by the novel Terrisporobacter isolates were generally consistent with the descriptions of the four type strains of the Terrisporobacter genus (Table 6). However, some unique physiological features of the novel isolates were identified. The isolates HSE and ELB showed lower temperatures and higher pH values for optimal growth than other Terrisporobacter isolates or reference strains, although the physiological optima may not be directly comparable due to methodological differences. ELB was the first Terrisporobacter strain identified to utilize trehalose for growth. The COM isolate utilized vanillate, mannitol and 1,2-propanediol for growth, which was not described for any of the type strains of the four Terrisporobacter species. The fermentation of mannitol and 1,2-propanediol by the COM isolate matched the respective unique OGs detected in the pan/core genome analysis. The degradation of glucose to acetate and ethanol by the COM isolate was in line with the detected products of sugar or ethylene glycol fermentation reported for the T. glycolicus DSM 1288T type strain (Gaston and Stadtman, 1963) or the acetogenic T. glycolicus RD-1 strain (Küsel et al., 2001). The determined optimal growth conditions of the A. noterae DSL2 isolate were similar to the data reported for the type strains of A. noterae (Sleat et al., 1985) and A. sticklandii (Bes et al., 2015) (Table 7). Furthermore, DSL2 utilized glucose and maltose, which matched with the description of A. noterae ATCC 35199T. The Acetoanaerobium isolate DSL2 additionally fermented fructose, sorbitol, sucrose, trehalose, melibiose, raffinose, melezitose and dextrin. Growth with fructose, sorbitol, sucrose, trehalose and melezitose deviates from the description of the A. noterae type strain, utilization of the other substrates was not assessed for other type strains. Surprisingly, the Acetoanaerobium isolate DSL2 did not grow with DMG, although it was isolated from an enrichment culture containing DMG as substrate. As no other energy source was supplemented than yeast extract, the isolate was likely growing by pairwise fermentation of amino acids via the Stickland reaction in the enrichment culture (Stickland, 1935). Substrates identified to be used for growth by the isolates of this study were only identified to support growth when yeast extract and peptone were also supplemented. This was required due to the dependency on the addition of complex substrates for bacteria of the genera Terrisporobacter and Acetoanaerobium. However, as complex substrates were also supplemented during the characterization of T. mayombei (Kane et al., 1991), T. glycolicus (Chamkha et al., 2001), T. petrolearius (Deng et al., 2015), A. noterae (Sleat et al., 1985), and A. pronyense (Bes et al., 2015) the substrate utilization data of this study is comparable to the reported data of the respective reference strains.
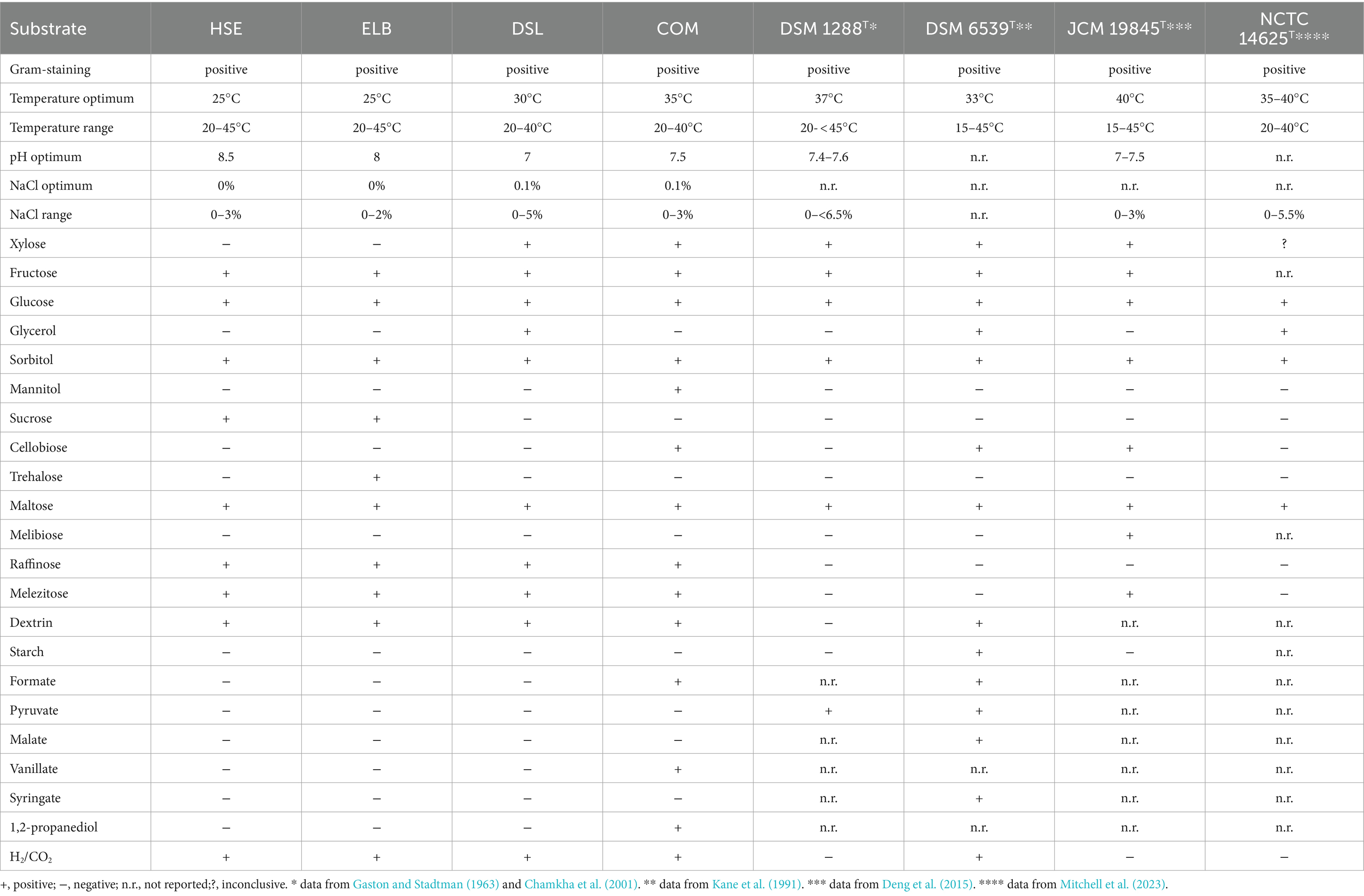
Table 6. Results of the physiological characterization of the T. petrolearius isolates HSE, ELB, DSL, and the T. vanillatitrophus isolate COM in comparison to the type strain descriptions of the Terrisporobacter species T. glycolicus DSM 1288T, T. mayombei DSM 6539T, T. petrolearius JCM 19845T and T. hibernicus NCTC 14625T.
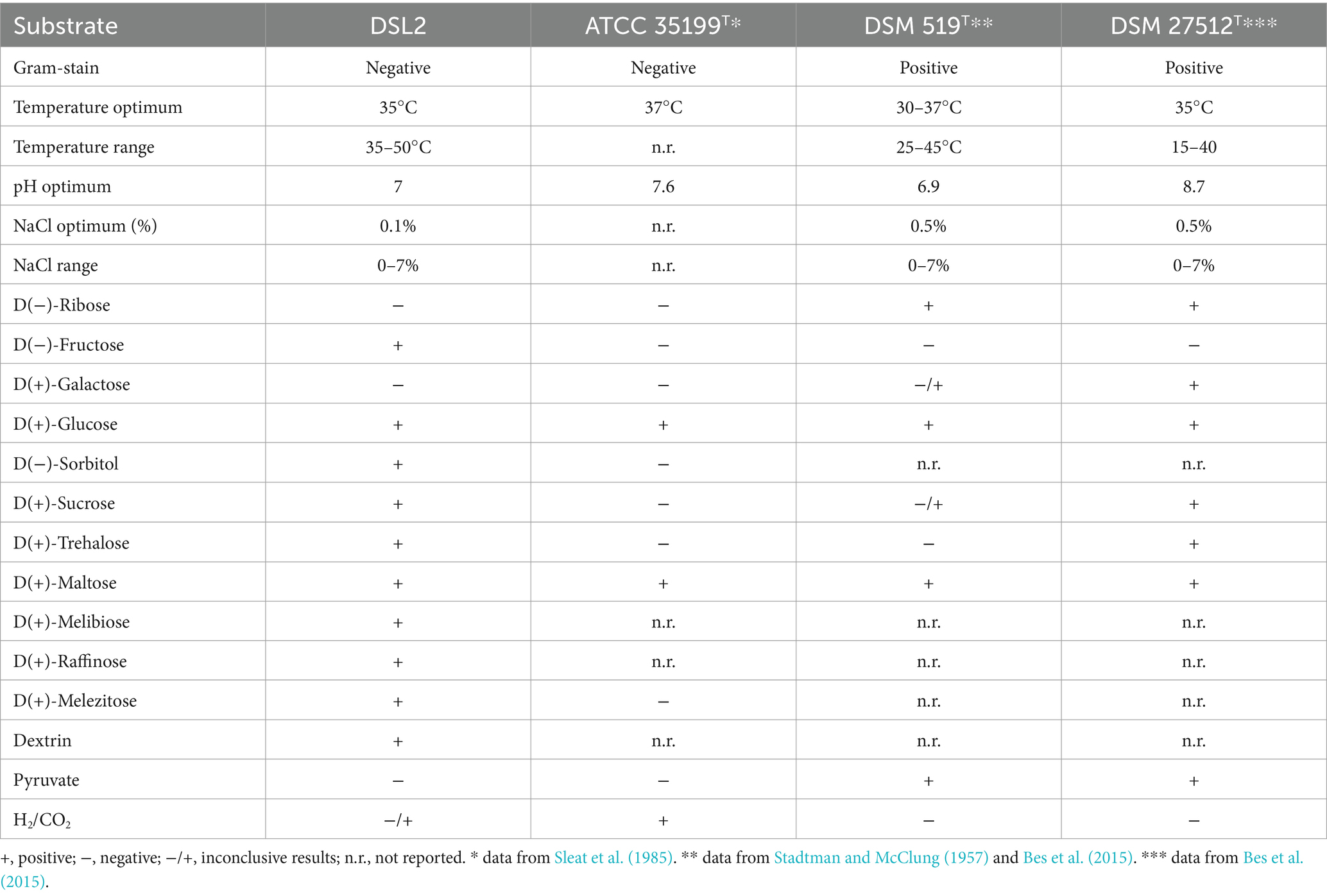
Table 7. Results of the physiological characterization of the A. sticklandii isolate DSL2 in comparison to the type strain descriptions of the Acetoanaerobium species A. noterae ATCC 35199T, A. sticklandii DSM 519T, A. pronyense DSM 27512T.
All Terrisporobacter isolates shared a common growth pattern in H2 + CO2-supplemented cultures. Initial growth took place by consuming nutrients provided by the complex substrates peptone and yeast extract in the medium, likely by performing the acetate-producing pairwise fermentation of amino acids via the Stickland reaction (Stickland, 1935; Sangavai and Chellapandi, 2019). The trace amounts of other products besides acetate and ethanol detected by the GC-measurements likely originated from these Stickland reactions. T. mayombei was reported to produce trace amounts of isovalerate from yeast extract (Kane et al., 1991) and T. glycolicus was reported to produce trace amounts of isovalerate from yeast extract, casamino acids, trypticase and peptone (Chamkha et al., 2001). We hypothesize that the H2-producing Stickland reaction is inhibited by high partial pressures of H2, leading to lower initial growth of some isolates supplied with H2 + CO2 than in control cultures. The initial heterotrophic growth was then followed by the stationary growth phase, in which continuous acetate production was observed in H2-supplemented cultures of the Terrisporobacter isolates indicative of acetogenesis. This growth pattern on H2 + CO2 has been reported for the acetogenic type strain of T. mayombei and CO2-fixation via the WLP into acetate was verified by 14C isotopic labelling of CO2 (Kane et al., 1991). T. vanillatitrophus COM was the only Terrisporobacter isolate found to produce besides acetate also ethanol during the stationary growth phase in H2-supplemented cultures. From our experiments, we could not exclude the possibility that this ethanol was produced from the provided complex substrates via the Stickland reaction, however, the ethanol production coincided with the onset of the stationary growth phase where these substrates were already consumed. Furthermore, there was no ethanol production observed in the control cultures. There are two hypothetical routes for the production of ethanol by acetogenesis with H2 + CO2 in T. vanillatitrophus. The ethanol precursor acetaldehyde could be produced from reabsorbed acetate by an acetaldehyde ferredoxin oxidoreductase (Aor) or directly from acetyl-CoA using an acetaldehyde dehydrogenase (Aldh) (Daniell et al., 2012). Synthesis of ethanol could then occur via alcohol dehydrogenases (Adh). We detected a total of three possible aor genes (TVTCOM_11530, TVTCOM_02900 and TVTCOM_02970) and three bifunctional acetaldehyde/alcohol dehydrogenase genes (adhE; TVTCOM_05440, TVTCOM_30030 and TVTCOM_30090) encoded by the T. vanillatitrophus genome. Other putative alcohol dehydrogenases identified comprised genes encoding a long-chain-alcohol dehydrogenase 1 (adh1; TVTCOM_02640), an alcohol dehydrogenase 2 (adhB; TVTCOM_24670) and an aldehyde dehydrogenase (aldH1; TVTCOM_18480).
In summary, the physiological characterizations of the COM isolate showed considerable differences in comparison to the data of other Terrisporobacter type strains and thereby further supported the classification of the COM isolate from the genomic characterization as a novel Terrisporobacter species.
In contrast to the Terrisporobacter isolates, the A. sticklandii isolate DSL2 did not achieve higher maximal growth values in H2-supplemented cultures than in the control cultures. This was also reported for the acetogenic type strain of A. noterae. However, A. noterae was found to produce more than 4 times the amount of acetate in H2-supplemented cultures compared to control cultures (Sleat et al., 1985). Hence, we could not confirm acetogenesis of the DSL2 isolate in our experiments, as we observed equal amounts of acetate being formed in H2-supplemented cultures compared to control cultures. It has to be considered that the acetate measurements of A. noterae were also conducted after longer incubation times (376 h) than in our experiments (96 h). Some H2-supplemented cultures of the A. noterae type strain were reported to continue utilizing H2 for more than 100 days after entering the stationary growth phase (Sleat et al., 1985). Therefore, the DSL2 isolate requires follow-up experiments with prolonged incubation times to verify the continuous production of acetate in H2-supplemented cultures. These experiments should also include the A. sticklandii type strain.
Active prophage regions were identified, under the tested conditions in every novel isolate screened with the exception of the HSE isolate. The size of the inferred phage genomes (36–55 kb) and the identification of multiple tail-associated genes indicated an affiliation to the class of Caudoviricetes (Zrelovs et al., 2020), which was also supported by the PhageScope taxonomy prediction. The DNA invertase uniquely identified in TPDSL_ph1 may represent a tool acting as a genetic switch for the alternate expression of gene sets as described for the Gin DNA invertase of the Escherichia coli K12 phage Mu controlling host specificity (van de Putte et al., 1980). Furthermore, TPDSL_ph1 contained two amidoligases in proximity to a γ-glutamyl cyclotransferase, which were predicted to potentially modify cell walls by linking novel peptides in the enterobacteriophage phiEco32 and thereby preventing host accession by other phages (Iyer et al., 2009). To the best of our knowledge this is the first report of active prophages of bacteria from the genus Terrisporobacter and Acetoanaerobium. The absence of comparable phage genomes in viral databases underlines the novelty of these bacteriophages. Related phage genomes could be identified by comparisons of the sequence of the terminase large subunit and a whole genome-based phylogram indicated the presence of two different novel phage families and a total of eight different phage genera.
Terrisporobacter vanillatitrophus (va.nil.la.ti.tro’phus. N.L. masc. n. vanillas, vanillate; Gr. masc. Adj. trophos, feeder; N.L. masc. Adj. vanillatitrophus, growing with vanillate). Cells are Gram-positive rods and produce terminal endospores. Growth occurs only under strict anaerobic conditions and is dependent on the addition of yeast extract. Growth ranges from the temperatures 20 to 40°C (optimum 35°C) and the NaCl concentrations 0 to 3% (optimum 0.1%). Optimal growth occurs at pH 7.5. The best growth under the tested conditions occurs with glucose as substrate, which is fermented to acetate and ethanol as fermentation products. Other substrates utilized for growth comprise xylose, fructose, sorbitol, mannitol, cellobiose, maltose, raffinose, melezitose, dextrin, formate, vanillate, 1,2-propanediol and H2 + CO2. Growth on H2 + CO2 occurs by initial heterotrophic growth followed by the stationary growth phase, where continuous acetate production is indicative of H2-dependent acetogenesis. H2-supplemented cultures also show the production of small amounts of ethanol in the stationary growth phase. Does not grow with arabinose, ribose, galactose, mannose, rhamnose, glucuronic acid, glycerol, inositol, sucrose, trehalose, lactose, melibiose, dextran, starch, pyruvate, lactate, malate, citrate, DMG, betaine, syringate, methanol, ethanolamine and CO under the tested conditions. The predominant cellular fatty acids are C16:0 (19%), C16:0 DMA (12.4%), C16:1 ∆9 (7.3%) and C16:1 ∆7 (6%). The type strain genome comprises a chromosome with the size of 4 Mb and three plasmids with the sizes of 21.5 Kb, 19.5 Kb and 5.6 Kb. The GC-content averages to 29%. The type strain COM was isolated from a compost sample taken from the Königsbühl composting plant near Göttingen, Germany. The Genbank/EMBL/DDBJ accession number of the 16S rRNA gene is PP196406. The type strain of T. vanillatitrophus is COMT and was deposited under the identifiers DSM 116160T and CCOS 2104T.
The novel acetogenic Terrisporobacter isolates expand the pool of available acetogens for biotechnological applications, in particular the novel species T. vanillatitrophus producing in addition to acetate also ethanol in H2-supplemented cultures. We propose to classify all isolates of the T. petrolearius/T. hibernicus phylogenomic cluster to the species T. petrolearius and all isolates of the A. noterae/A. sticklandii to the species A. sticklandii. Our results indicate that acetogenesis could be a more frequent trait of Terrisporobacter and Acetoanaerobium strains than hitherto described. However, these require follow-up experiments including all isolated strains of the respective genera. We could show that acetogenesis characteristically occurs in acetogenic strains from the genus Terrisporobacter by an initial heterotrophic growth phase on complex substrates followed up by a stationary growth phase with continuous acetate production. Furthermore, we provide first insights into active prophages in acetogenic bacteria from the genera of Terrisporobacter and Acetoanaerobium.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
TB: Conceptualization, Funding acquisition, Methodology, Writing – review & editing, Data curation, Formal analysis, Investigation, Visualization, Writing – original draft. MS: Investigation, Resources, Writing – review & editing. AL: Investigation, Writing – review & editing. LeE: Investigation, Writing – review & editing. JD: Investigation, Writing – review & editing. MH: Investigation, Writing – review & editing. LiE: Data curation, Investigation, Writing – review & editing. MB: Funding acquisition, Investigation, Writing – review & editing. RD: Conceptualization, Funding acquisition, Project administration, Supervision, Writing – review & editing. AP: Conceptualization, Funding acquisition, Methodology, Supervision, Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. We acknowledge the support of Tim Böer by the “Deutsche Bundesstiftung Umwelt” (DBU; 20020/640). Rolf Daniel and Anja Poehlein are grateful for support from the Bundesministerium für Bildung und Forschung (BMBF) for the project “Mikrobielle Biofabriken: THERMOSYNCON—Entwicklung thermophiler Mikroorganismen als Biokatalysatoren für die Umwandlung von Synthesegas zu Biobrennstoffen und Chemikalien“(grant number 031B0857C). Mirko Basen is grateful for support from the Bundesministerium für Bildung und Forschung (BMBF) for the project “Mikrobielle Biofabriken: THERMOSYNCON—Entwicklung thermophiler Mikroorganismen als Biokatalysatoren für die Umwandlung von Synthesegas zu Biobrennstoffen und Chemikalien” (grant number 031B0857A). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. This publication has been partially financed with the help of the publication fund NiedersachenOPEN, supported by zukunft.niedersachsen.
We thank Mechthild Bömeke for technical assistance. Furthermore, we thank Wolfgang R. Streit and Luise Göbbels for providing the Elbe marsh sediment sample. We thank the Open Access Publication Funds of the University of Göttingen and the “Ministerium für Wissenschaft und Kultur” of Lower Saxony (Germany) for supporting publishing open access.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2024.1426882/full#supplementary-material
Alikhan, N.-F., Petty, N. K., Ben Zakour, N. L., and Beatson, S. A. (2011). BLAST ring image generator (BRIG): simple prokaryote genome comparisons. BMC Genomics 12:402. doi: 10.1186/1471-2164-12-402
Aryal, N., Tremblay, P.-L., Lizak, D. M., and Zhang, T. (2017). Performance of different Sporomusa species for the microbial electrosynthesis of acetate from carbon dioxide. Bioresour. Technol. 233, 184–190. doi: 10.1016/j.biortech.2017.02.128
Aspinall, S. T., and Hutchinson, D. N. (1992). New selective medium for isolating Clostridium difficile from faeces. J. Clin. Pathol. 45, 812–814. doi: 10.1136/jcp.45.9.812
Baum, C., Zeldes, B., Poehlein, A., Daniel, R., Müller, V., and Basen, M. (2024). The energy-converting hydrogenase Ech2 is important for the growth of the thermophilic acetogen Thermoanaerobacter kivui on ferredoxin-dependent substrates. Microbiol Spectr 12:e0338023. doi: 10.1128/spectrum.03380-23
Bertelli, C., Laird, M. R., Williams, K. P., Lau, B. Y., Hoad, G., Winsor, G. L., et al. (2017). IslandViewer 4: expanded prediction of genomic islands for larger-scale datasets. Nucleic Acids Res. 45, W30–W35. doi: 10.1093/nar/gkx343
Bes, M., Merrouch, M., Joseph, M., Quéméneur, M., Payri, C., Pelletier, B., et al. (2015). Acetoanaerobium pronyense sp. nov., an anaerobic alkaliphilic bacterium isolated from a carbonate chimney of the Prony hydrothermal field (New Caledonia). Int. J. Syst. Evol. Microbiol. 65, 2574–2580. doi: 10.1099/ijs.0.000307
Blamey, J. M., and Adams, M. W. W. (1993). Purification and characterization of pyruvate ferredoxin oxidoreductase from the hyperthermophilic archaeon Pyrococcus furiosus. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1161, 19–27. doi: 10.1016/0167-4838(93)90190-3
Blamey, J. M., and Adams, M. W. W. (1994). Characterization of an ancestral type of pyruvate ferredoxin oxidoreductase from the hyperthermophilic bacterium, Thermotoga maritima. Biochemistry 33, 1000–1007. doi: 10.1021/bi00170a019
Böer, T., Bengelsdorf, F. R., Bömeke, M., Daniel, R., and Poehlein, A. (2023). Genome-based metabolic and phylogenomic analysis of three Terrisporobacter species. PLoS One 18:e0290128. doi: 10.1371/journal.pone.0290128
Bolger, A. M., Lohse, M., and Usadel, B. (2014). Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
Bouras, G., Nepal, R., Houtak, G., Psaltis, A. J., Wormald, P.-J., and Vreugde, S. (2023). Pharokka: a fast scalable bacteriophage annotation tool. Bioinformatics 39, 1–4. doi: 10.1093/bioinformatics/btac776
Budianto, Z. O., and Yasin, R. E. (2024). The role of bacterial and archaea in determining the metabolic pathway of biogas fermentation at low temperatures. Acta Biol Colomb 29, 99–111. doi: 10.15446/abc.v29n1.106266
Chamkha, M., Labat, M., Patel, B. K., and Garcia, J. L. (2001). Isolation of a cinnamic acid-metabolizing Clostridium glycolicum strain from oil mill wastewaters and emendation of the species description. Int. J. Syst. Evol. Microbiol. 51, 2049–2054. doi: 10.1099/00207713-51-6-2049
Chen, T., Xie, G., Mi, J., Wen, X., Cao, Z., Ma, B., et al. (2022). Recovery of the structure and function of the pig manure bacterial community after enrofloxacin exposure. Microbiol Spectr 10, e02004–e02021. doi: 10.1128/spectrum.02004-21
Chen, S., Zhou, Y., Chen, Y., and Gu, J. (2018). fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 34, i884–i890. doi: 10.1093/bioinformatics/bty560
Claus, D. (1992). A standardized gram staining procedure. World J. Microbiol. Biotechnol. 8, 451–452. doi: 10.1007/BF01198764
Daniell, J., Köpke, M., and Simpson, S. (2012). Commercial biomass syngas fermentation. Energies 5, 5372–5417. doi: 10.3390/en5125372
Deng, Y., Guo, X., Wang, Y., He, M., Ma, K., Wang, H., et al. (2015). Terrisporobacter petrolearius sp. nov., isolated from an oilfield petroleum reservoir. Int. J. Syst. Evol. Microbiol. 65, 3522–3526. doi: 10.1099/ijsem.0.000450
Dupuy, B., Mani, N., Katayama, S., and Sonenshein, A. L. (2005). Transcription activation of a UV-inducible Clostridium perfringens bacteriocin gene by a novel σ factor. Mol. Microbiol. 55, 1196–1206. doi: 10.1111/j.1365-2958.2004.04456.x
Eram, M. S., Wong, A., Oduaran, E., and Ma, K. (2015). Molecular and biochemical characterization of bifunctional pyruvate decarboxylases and pyruvate ferredoxin oxidoreductases from Thermotoga maritima and Thermotoga hypogea. J. Biochem. 158, mvv058–mvv066. doi: 10.1093/jb/mvv058
Furdui, C., and Ragsdale, S. W. (2000). The role of pyruvate ferredoxin oxidoreductase in pyruvate synthesis during autotrophic growth by the wood-Ljungdahl pathway. J. Biol. Chem. 275, 28494–28499. doi: 10.1074/jbc.M003291200
Galperin, M. Y., Brover, V., Tolstoy, I., and Yutin, N. (2016). Phylogenomic analysis of the family Peptostreptococcaceae (Clostridium cluster XI) and proposal for reclassification of Clostridium litorale (Fendrich et al. 1991) and Eubacterium acidaminophilum (Zindel et al. 1989) as Peptoclostridium litorale gen. Nov. comb. nov. and Peptoclostridium acidaminophilum comb. nov. Int. J. Syst. Evol. Microbiol. 66, 5506–5513. doi: 10.1099/ijsem.0.001548
Gaston, L. W., and Stadtman, E. R. (1963). Fermentation of ethylene glycol by Clostridium glycolicum, sp. n. J. Bacteriol. 85, 356–362. doi: 10.1128/jb.85.2.356-362.1963
Gencic, S., and Grahame, D. A. (2020). Diverse energy-conserving pathways in Clostridium difficile: growth in the absence of amino acid stickland acceptors and the role of the wood-Ljungdahl pathway. J. Bacteriol. 202, e00233–e00220. doi: 10.1128/JB.00233-20
Gerritsen, J., Fuentes, S., Grievink, W., van Niftrik, L., Tindall, B. J., Timmerman, H. M., et al. (2014). Characterization of Romboutsia ilealis gen. Nov., sp. nov., isolated from the gastro-intestinal tract of a rat, and proposal for the reclassification of five closely related members of the genus Clostridium into the genera Romboutsia gen. Nov., Intestinibacter gen. Nov., Terrisporobacter gen. Nov., Asaccharospora gen. Nov. Int. J. Syst. Evol. Microbiol. 64, 1600–1616. doi: 10.1099/ijs.0.059543-0
Goris, J., Konstantinidis, K. T., Klappenbach, J. A., Coenye, T., Vandamme, P., and Tiedje, J. M. (2007). DNA–DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 57, 81–91. doi: 10.1099/ijs.0.64483-0
Hertel, R., Rodríguez, D. P., Hollensteiner, J., Dietrich, S., Leimbach, A., Hoppert, M., et al. (2015). Genome-based identification of active prophage regions by next generation sequencing in Bacillus licheniformis DSM13. PLoS One 10:e0120759. doi: 10.1371/journal.pone.0120759
Hunt, M., De, S. N., Otto, T. D., Parkhill, J., Keane, J. A., and Harris, S. R. (2015). Circlator: automated circularization of genome assemblies using long sequencing reads. Genome Biol. 16:294. doi: 10.1186/s13059-015-0849-0
Iyer, L. M., Abhiman, S., Maxwell Burroughs, A., and Aravind, L. (2009). Amidoligases with ATP-grasp, glutamine synthetase-like and acetyltransferase-like domains: synthesis of novel metabolites and peptide modifications of proteins. Mol. BioSyst. 5:1636. doi: 10.1039/b917682a
Kane, M. D., Brauman, A., and Breznak, J. A. (1991). Clostridium mayombei sp. nov., an H2/CO2 acetogenic bacterium from the gut of the African soil-feeding termite, Cubitermes speciosus. Arch. Microbiol. 156, 99–104. doi: 10.1007/BF00290980
Katsyv, A., Schoelmerich, M. C., Basen, M., and Müller, V. (2021). The pyruvate:ferredoxin oxidoreductase of the thermophilic acetogen Thermoanaerobacter kivui. FEBS Open Bio 11, 1332–1342. doi: 10.1002/2211-5463.13136
Kolmogorov, M., Yuan, J., Lin, Y., and Pevzner, P. A. (2019). Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 37, 540–546. doi: 10.1038/s41587-019-0072-8
Köpke, M., Straub, M., and Dürre, P. (2013). Clostridium difficile is an autotrophic bacterial pathogen. PLoS One 8:e62157. doi: 10.1371/journal.pone.0062157
Koren, S., Walenz, B. P., Berlin, K., Miller, J. R., Bergman, N. H., and Phillippy, A. M. (2017). Canu: scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 27, 722–736. doi: 10.1101/gr.215087.116
Küsel, K., Karnholz, A., Trinkwalter, T., Devereux, R., Acker, G., and Drake, H. L. (2001). Physiological ecology of Clostridium glycolicum RD-1, an aerotolerant acetogen isolated from sea grass roots. Appl. Environ. Microbiol. 67, 4734–4741. doi: 10.1128/AEM.67.10.4734-4741.2001
Langmead, B., and Salzberg, S. L. (2012). Fast gapped-read alignment with bowtie 2. Nat. Methods 9, 357–359. doi: 10.1038/nmeth.1923
Lechner, M., Findeiß, S., Steiner, L., Marz, M., Stadler, P. F., and Prohaska, S. J. (2011). Proteinortho: detection of (co-)orthologs in large-scale analysis. BMC Bioinformatics 12:124. doi: 10.1186/1471-2105-12-124
Li, Y., Li, X., Sun, Y., Zhao, X., and Li, Y. (2018). Cathodic microbial community adaptation to the removal of chlorinated herbicide in soil microbial fuel cells. Environ. Sci. Pollut. Res. 25, 16900–16912. doi: 10.1007/s11356-018-1871-z
Ma, K., Hutchins, A., Sung, S.-J. S., and Adams, M. W. W. (1997). Pyruvate ferredoxin oxidoreductase from the hyperthermophilic archaeon, Pyrococcus furiosus, functions as a CoA-dependent pyruvate decarboxylase. Proc. Natl. Acad. Sci. 94, 9608–9613. doi: 10.1073/pnas.94.18.9608
Madjarov, J., Soares, R., Paquete, C. M., and Louro, R. O. (2022). Sporomusa ovata as catalyst for bioelectrochemical carbon dioxide reduction: a review across disciplines from microbiology to process engineering. Front. Microbiol. 13:913311. doi: 10.3389/fmicb.2022.913311
Manni, M., Berkeley, M. R., Seppey, M., Simão, F. A., and Zdobnov, E. M. (2021). BUSCO update: novel and streamlined workflows along with broader and deeper phylogenetic coverage for scoring of eukaryotic, prokaryotic, and viral genomes. Mol. Biol. Evol. 38, 4647–4654. doi: 10.1093/molbev/msab199
Marçais, G., Delcher, A. L., Phillippy, A. M., Coston, R., Salzberg, S. L., and Zimin, A. (2018). MUMmer4: A fast and versatile genome alignment system. PLoS Comput. Biol. 14:e1005944. doi: 10.1371/journal.pcbi.1005944
Meier-Kolthoff, J. P., Auch, A. F., Klenk, H.-P., and Göker, M. (2013). Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinformatics 14:60. doi: 10.1186/1471-2105-14-60
Meier-Kolthoff, J. P., Carbasse, J. S., Peinado-Olarte, R. L., and Göker, M. (2022). TYGS and LPSN: a database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acids Res. 50, D801–D807. doi: 10.1093/nar/gkab902
Meier-Kolthoff, J. P., and Göker, M. (2017). VICTOR: genome-based phylogeny and classification of prokaryotic viruses. Bioinformatics 33, 3396–3404. doi: 10.1093/bioinformatics/btx440
Meier-Kolthoff, J. P., and Göker, M. (2019). TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 10:2182. doi: 10.1038/s41467-019-10210-3
Mitchell, M., Nguyen, S. V., Connor, M., Fairley, D. J., Donoghue, O., Marshall, H., et al. (2023). Terrisporobacter hibernicus sp. nov., isolated from bovine faeces in Northern Ireland. Int. J. Syst. Evol. Microbiol. 73:005667. doi: 10.1099/ijsem.0.005667
Ohwaki, K., and Hungate, R. E. (1977). Hydrogen utilization by Clostridia in sewage sludge. Appl. Environ. Microbiol. 33, 1270–1274. doi: 10.1128/aem.33.6.1270-1274.1977
Poehlein, A., Cebulla, M., Ilg, M. M., Bengelsdorf, F. R., Schiel-Bengelsdorf, B., Whited, G., et al. (2015). The complete genome sequence of Clostridium aceticum: a missing link between Rnf- and cytochrome-containing autotrophic acetogens. MBio 6, e01168–e01115. doi: 10.1128/mBio.01168-15
Pritchard, L., Glover, R. H., Humphris, S., Elphinstone, J. G., and Toth, I. K. (2016). Genomics and taxonomy in diagnostics for food security: soft-rotting enterobacterial plant pathogens. Anal. Methods 8, 12–24. doi: 10.1039/C5AY02550H
Sangavai, C., and Chellapandi, P. (2019). A metabolic study to decipher amino acid catabolism-directed biofuel synthesis in Acetoanaerobium sticklandii DSM 519. Amino Acids 51, 1397–1407. doi: 10.1007/s00726-019-02777-4
Schüler, M. A., Daniel, R., and Poehlein, A. (2024). Novel insights into phage biology of the pathogen Clostridioides difficile based on the active virome. Front. Microbiol. 15:15. doi: 10.3389/fmicb.2024.1374708
Seemann, T. (2014). Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069. doi: 10.1093/bioinformatics/btu153
Sleat, R., Mah, R. A., and Robinson, R. (1985). Acetoanaerobium noterae gen. Nov., sp. nov.: an anaerobic bacterium that forms acetate from H2 and CO2. Int. J. Syst. Bacteriol. 35, 10–15. doi: 10.1099/00207713-35-1-10
Stadtman, T. C., and McClung, L. S. (1957). Clostridium sticklandii nov. spec. J. Bacteriol. 73, 218–219. doi: 10.1128/jb.73.2.218-219.1957
Stickland, L. H. (1935). Studies in the metabolism of the strict anaerobes (genus Clostridium). Biochem. J. 29, 889–898. doi: 10.1042/bj0290889
Thulluru, L. P., Ghangrekar, M. M., and Chowdhury, S. (2023). Progress and perspectives on microbial electrosynthesis for valorisation of CO2 into value-added products. J. Environ. Manag. 332:117323. doi: 10.1016/j.jenvman.2023.117323
van de Putte, P., Cramer, S., and Giphart-Gassler, M. (1980). Invertible DNA determines host specificity of bacteriophage mu. Nature 286, 218–222. doi: 10.1038/286218a0
Vaser, R., and Šikić, M. (2021). Time- and memory-efficient genome assembly with raven. Nat Comput Sci 1, 332–336. doi: 10.1038/s43588-021-00073-4
Wang, D., Liang, Q., Chu, N., Zeng, R. J., and Jiang, Y. (2023). Deciphering mixotrophic microbial electrosynthesis with shifting product spectrum by genome-centric metagenomics. Chem. Eng. J. 451:139010. doi: 10.1016/j.cej.2022.139010
Wang, R. H., Yang, S., Liu, Z., Zhang, Y., Wang, X., Xu, Z., et al. (2024). PhageScope: a well-annotated bacteriophage database with automatic analyses and visualizations. Nucleic Acids Res. 52, D756–D761. doi: 10.1093/nar/gkad979
Wick, R. R., and Holt, K. E. (2022). Polypolish: short-read polishing of long-read bacterial genome assemblies. PLoS Comput. Biol. 18:e1009802. doi: 10.1371/journal.pcbi.1009802
Wick, R. R., Judd, L. M., Cerdeira, L. T., Hawkey, J., Méric, G., Vezina, B., et al. (2021). Trycycler: consensus long-read assemblies for bacterial genomes. Genome Biol. 22:266. doi: 10.1186/s13059-021-02483-z
Wickham, H. (2016). ggplot2: Elegant graphics for data analysis. New York: Springer International Publishing.
Wishart, D. S., Han, S., Saha, S., Oler, E., Peters, H., Grant, J. R., et al. (2023). PHASTEST: faster than PHASTER, better than PHAST. Nucleic Acids Res. 51, W443–W450. doi: 10.1093/nar/gkad382
Keywords: acetogen, Terrisporobacter , Acetoanaerobium , ethanol, prophage, Wood-Ljungdahl pathway
Citation: Böer T, Schüler MA, Lüschen A, Eysell L, Dröge J, Heinemann M, Engelhardt L, Basen M, Daniel R and Poehlein A (2024) Isolation and characterization of novel acetogenic strains of the genera Terrisporobacter and Acetoanaerobium. Front. Microbiol. 15:1426882. doi: 10.3389/fmicb.2024.1426882
Edited by:
John A. Fuerst, The University of Queensland, AustraliaReviewed by:
Armin Ehrenreich, Technical University of Munich, GermanyCopyright © 2024 Böer, Schüler, Lüschen, Eysell, Dröge, Heinemann, Engelhardt, Basen, Daniel and Poehlein. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Anja Poehlein, YW5qYS5wb2VobGVpbkBiaW9sb2dpZS51bmktZ29ldHRpbmdlbi5kZQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.