
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 23 October 2024
Sec. Microorganisms in Vertebrate Digestive Systems
Volume 15 - 2024 | https://doi.org/10.3389/fmicb.2024.1425218
This article is part of the Research TopicUnravelling the Unknown of the Rumen Microbiome: Implications for Animal Health, Productivity, and BeyondView all 23 articles
 Mu-Long Lu1
†
Mu-Long Lu1
† Guo-Hong Yuan1
†
Guo-Hong Yuan1
† Halidai Rehemujiang1,2Chang-Chang Li1Li-Hong Hu1Ping-Ping Duan1Li-Dong Zhang1
Halidai Rehemujiang1,2Chang-Chang Li1Li-Hong Hu1Ping-Ping Duan1Li-Dong Zhang1 Qi-Yu Diao3Kai-Dong Deng4Gui-Shan Xu1,2*
Qi-Yu Diao3Kai-Dong Deng4Gui-Shan Xu1,2*Introduction: The study aimed to evaluate the effects of Pleurotus Spent Mushroom Substrate (P.SMS) on the rumen microbiota, encompassing bacteria and fungi, as well as their interactions in Hu sheep.
Methods: A total of forty-five 3-month-old Hu sheep were randomly assigned to five groups. Each group was fed diets in which whole-plant corn silage (WPCS) was substituted with P.SMS at varying levels: 0% (CON), 5% (PSMS5), 10% (PSMS10), 15% (PSMS15), or 20% (PSMS20).
Results: The results indicated that higher proportions of P.SMS during the experimental period might have a detrimental effect on feed utilization efficiency, kidney function, and blood oxygen-carrying capacity. Notably, moderate levels of P.SMS, specifically below 15%, were associated with improvements in rumen NH3-N levels and absorption capacity. The results indicated that (1) PSMS20 exhibited a significantly higher feed-to-gain ratio compared to CON (P < 0.05); (2) PSMS15 showed a significantly higher NH3-N content than CON, PSMS5, and PSMS20. Additionally, PSMS10 and PSMS20 had elevated concentrations of NH3-N compared to CON and PSMS5 (P < 0.05); (3) The length and width of rumen papillae were significantly greater in PSMS20 compared to CON and PSMS5 (P < 0.05); (4) Creatinine levels were significantly higher in PSMS20 than in CON, PSMS5, and PSMS10 (P < 0.05); (5) By the conclusion of the experiment, hemoglobin concentration in PSMS20 showed a significant increase compared to CON (P < 0.05). Furthermore, the addition of P.SMS influenced microorganisms at both the phylum and genus levels: (1) At the phylum level, the prevalence of Patescibacteria was significantly lower in PSMS20 compared to the other groups; (2) PSMS15 exhibited significantly higher relative abundances of Basidiomycota compared to CON and PSMS10, while PSMS20 also demonstrated significantly higher relative abundances compared to CON (P < 0.05); (3) At the genus level, the prevalence of Candidatus_Saccharimonas in PSMS20 was significantly lower than in PSMS5, PSMS10, and PSMS15. Conversely, the prevalence of Phanerochaete in PSMS15 was notably higher than in CON and PSMS10, and it was also significantly elevated in PSMS20 compared to CON (P < 0.05); (4) Correlation analysis indicated no significant correlation between changes in the structure of bacterial and fungal communities.
Discussion: Considering these findings, a high percentage of P.SMS negatively impacted feed utilization efficiency, blood oxygen carrying capacity, and kidney function, while a moderate percentage of P.SMS promotes rumen absorption capacity, indicating that feeding 10% P.SMS is optimal.
Mushrooms have traditionally been utilized in Asian countries, particularly in China, for their medicinal properties and as functional foods due to their high-quality proteins, carbohydrates, minerals, vitamins, and low-fat content (Martin et al., 2023). Spent mushroom substrate (SMS) typically consists of the plant biomass that is discarded after mushroom harvesting cycles (Lucas De Jesus et al., 2023). With the growing demand for mushrooms, approximately five times the amount of SMS is generated with each kilogram of mushrooms produced, resulting in significant waste. A previous study highlighted the nutraceutical potential of SMS, particularly for its prebiotic properties (Baptista et al., 2023). A review noted that the mycelium and fruiting bodies exhibit different compositions based on their environments, but this does not compromise their safety or quality when cultivated under controlled conditions (Berger et al., 2022). While research on the feeding of mycelium remains limited, existing studies suggest that mycelium consumption can enhance the body’s immunity and possess anti-inflammatory properties (Shin et al., 2021; Yang et al., 2022). Nonetheless, the majority of SMS has been either incinerated or discarded as waste, leading to the loss of valuable biological resources and contributing to severe environmental pollution (Huang et al., 2023). Therefore, the full utilization of SMS shows promise, but investigation is needed to explore the impact of SMS feed utilization on ruminant animals.
Pleurotus mushrooms are recognized for their safety and edibility, along with diverse biotechnological and environmental applications. This versatility is largely attributed to their non-specific oxidative enzymatic systems and ligninolytic peroxidases, which play a crucial role in various biodegradation processes, including the fungal pretreatment of lignocellulosic biomass for sustainable waste management (Kainthola et al., 2021; Scholtmeijer et al., 2023). Pleurotus spp. exhibit an exceptional ability to selectively mineralize lignin through the production of various extracellular oxidizing enzymes, thereby establishing an effective lignin degradation system (Knop et al., 2015; van Kuijk et al., 2015). This process enhances the accessibility of cellulose and hemicellulose for the rumen microbe. A recent study by Agustinho et al. (2021) identified three lignin-degrading enzymes—lignin peroxidase, and manganese peroxidase—in the spent mushroom substrate of Pleurotus ostreatus (P.SMS). When incorporated into whole corn plant ensiling fermentation, these enzymes contributed to a reduction in lignin concentration. Additionally, research by Xu et al. (2010) indicated that increasing the concentration of SMS in total mixed ration silage tended to elevate the acetate to propionate ratio and pH in the rumen. Song et al. (2007) demonstrated that feeding finishing Berkshire pigs with fermented oyster mushroom by-products influenced multiple indicators during the fattening process. Furthermore, a study conducted by Huang et al. (2023) revealed that the addition of fermented SMS significantly improved the diversity and abundance of the rumen bacterial community. Our previous study pointed out that the inclusion of a high proportion (20%) of P.SMS exhibited a favorable feeding effect during the initial 2 months of the fattening period; however, as the feeding duration was extended, the effectiveness of the feeding began to decline (Guohong et al., 2023). Despite these findings, there remains a notable lack of studies directly investigating the effects of feeding SMS to animals, as well as limited research on its impact on rumen fungi.
The microbial composition of an animal’s digestive tract plays a crucial role in various physiological responses of the organism. The complex relationship between the host and its gut microbiota influences various aspects, including nutrient digestion and absorption, immune system modulation, overall health, and even behaviors (de Vos et al., 2022; Lamontagne et al., 2023; Ma Y. et al., 2022; Martinez-Guryn et al., 2018; Zhang X. et al., 2022). The degradation of plant cell wall polysaccharides in the rumen is known to be associated with ruminal bacteria, protozoa, and fungi (Rehemujiang et al., 2023).
Given that SMS incorporates mycelium, a unique type of fungus, and that Pleurotus-treated substrates enhance microbial access to cellulose and hemicellulose, we hypothesized that the addition of P.SMS would create additional utilization sites for rumen microbes or alter the rumen flora in sheep. Our objective was to investigate the impact of direct SMS feeding on Hu sheep by examining growth performance, rumen, and blood indices, as well as the rumen microbial community. This study focused on assessing the effects of incorporating SMS at various supplementation levels, replacing whole-plant corn silage (WPCS), on the microbial community structure in the rumen of sheep. We anticipated that this research would provide a novel perspective on SMS feeding, offering valuable guidance and insights for related research and practical applications.
The animal study was reviewed and approved by the animal ethics committee of Tarim University (2,024,049 and 2,024,067), written informed consent was obtained from the owners for the participation of their animals in this study.
P.SMS was cultivated using a substrate consisting of cultured from 70% cottonseed hulls, 20% sawdust, 7% wheat bran, and 3% lime sourced from a mushroom plantation located on the outskirts of Alar City, Xinjiang. The chemical composition of P.SMS and WPCS used in the experiment is described in Table 1.
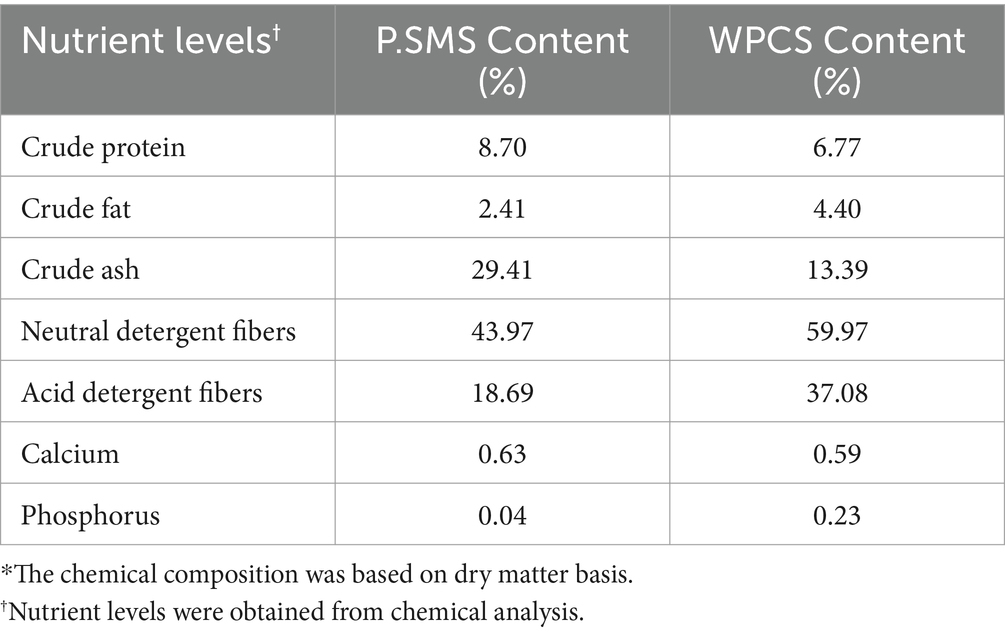
Table 1. The chemical composition* of P.SMS and WPCS used in the experiment.
A completely randomized experimental design was employed in this study. Forty-five 3-month-old male Hu sheep with an average weight of 30.0 ± 1.2 kg were selected and randomly assigned to 5 groups. These groups received varying levels of spent mushroom substrate (SMS) as treatments: CON (0%), PSMS5 (5%), PSMS10 (10%), PSMS15 (15%), and PSMS20 (20%). Each treatment group consisted of 9 sheep. The basic diet, formulated to meet the requirements of a 30-kg sheep with a daily gain of 200 g, is detailed in Table 2. The diet was administered twice daily, once in the morning at 10:00 and again at 19:00. An adaptation period of 10 days preceded a 90-day trial period. The feed ingredients in the basal diet were sourced from the farm, and their percentages were adjusted to meet the energy requirements for growth. The formulation remained unchanged throughout the feeding process; only the quantity fed was adjusted based on leftovers. During the trial, all sheep were housed individually in a partially open enclosure, provided with free access to salt blocks and clean water, and exposed to natural light and a cool environmental temperature.
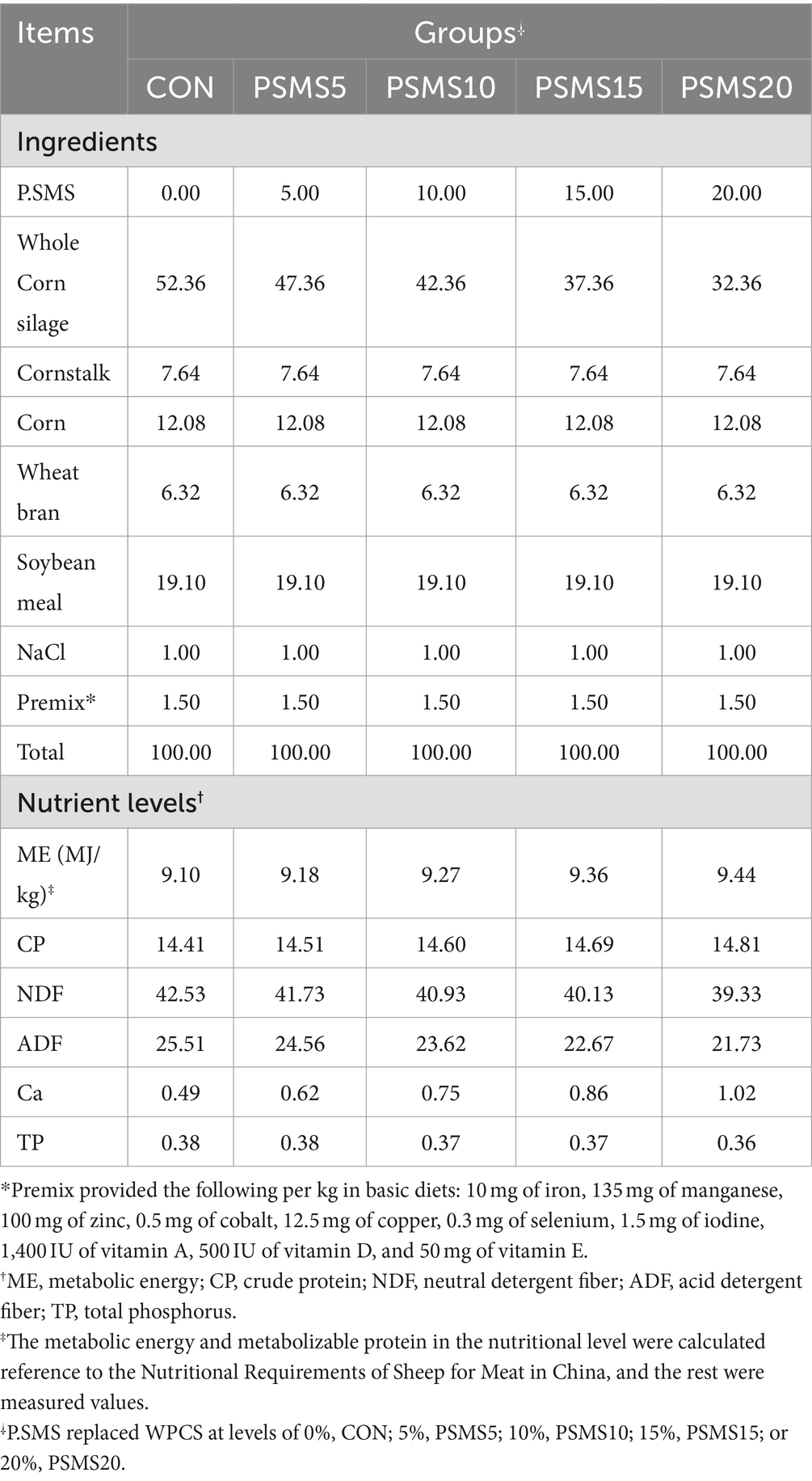
Table 2. Test diet composition and nutrition level (dry matter basis) %.
The feed intake was monitored daily, with leftovers from the previous day collected before the morning feed, dried, and weighed to determine the dry matter intake (DMI) and feed-to-gain ratio (F/G). Sheep were weighed before morning feeding on the 1st and 90th days to compute the average daily weight gain (ADG) over the feeding period.
One day prior to slaughter, a veterinarian examined the sheep to confirm the absence of gastrointestinal diseases. At the conclusion of the experiment, 25 sheep were randomly chosen from each group (5 sheep per group) and subjected to a 24-h fast, and 12 h without water before slaughter. All sheep were slaughtered, and their rumen was isolated. All the rumen fluid from each sheep was collected by filtering the digest contents through sterile 4-layer gauze. We then froze and stored 150 mL of rumen fluid from each sheep. The pH of the rumen fluid was promptly measured using a pH detector (FE28-standard, Mettler Toledo Co., China), while the NH3-N content was determined using a modified colorimetric technique (Feng and Gao, 2010). Subsequently, the rumen fluid was promptly transferred into liquid nitrogen and stored at −80°C for the analysis of rumen microbiota.
The rumen tissue was washed with saline and subsequently immersed in 4% paraformaldehyde. Subsequently, paraffin sections were prepared by removing the samples from the paraformaldehyde and rinsing them under running water for 24 h. The paraffin sections were observed under an electron microscope (ECLIPSE E200, Nikon Instruments Inc., Japan) and then photographed using a microscope image processing system (TL-507). The length and width of the rumen papilla, along with the thickness of the rumen muscular wall, were measured using ImageJ software.
Blood routine and serum biochemical indices were collected from the sheep on the 1st and 90th days of the experimental period, prior to morning feeding using conventional blood collection methods. For the blood routine, samples were drawn from blood tubes containing EDTA with 5 mL of blood. The samples were mixed and the total leukocyte count, hemoglobin concentration, and platelet count were determined using a fully automated blood cell analyzer for animals (BC-2800Vet, Mindray Co., China). Serum biochemical indices were collected from standard blood tubes containing 5 mL of blood. The tubes were allowed to stand for 40 min before being centrifuged for 15 min. Following centrifugation, the supernatant was transferred to 1.5 mL tubes for the determination of urea nitrogen and creatinine concentration using an automatic biochemical analyzer (Hitachi 7020 automatic analyzer, Hitachi Ltd., Japan).
The rumen samples underwent total genomic DNA extraction using the TGuide S96 Magnetic Soil/Stool DNA Kit (TIANGEN Biotech, Beijing Co., Ltd.), following the manufacturer’s protocol. The quality and quantity of the extracted DNA were assessed via electrophoresis on a 1.8% agarose gel and quantified using a NanoDrop 2000 UV–Vis spectrophotometer (Thermo Scientific, Wilmington, USA). For amplification of the full-length 16S rRNA gene, primer pairs 27F: AGRGTTTGATYNTGGCTCAG and 1492R: TASGGHTACCTTGTTASGACTT were utilized. The Internal Transcribed Spacer Identification (ITS) region was amplified using primer pairs ITS1-F: CTTGGTCATTTAGAGGAAGTAA and ITS4: TCCTCCGCTTATTGATATGC. Following quantification, amplicons with equimolar concentrations were pooled and sequenced on the PacBio Sequel II platform (Beijing Biomarker Technologies Co., Ltd., Beijing, China).
The raw reads obtained from sequencing underwent filtering and demultiplexing using SMRT Link software (v8.0), with parameters set to minPasses ≥5 and minPredictedAccuracy ≥0.9, to obtain circular consensus sequencing (CCS) reads. Subsequently, Lima software (v1.7.0)1 was employed to assign the CCS sequences to the corresponding samples based on their barcodes. CCS reads devoid of primers and those falling outside the length range of 1,200–1,650 bp were discarded, utilizing forward and reverse primers identification and quality filtering via Cutadapt (v2.7, Martin, 2011). The UCHIME algorithm (v8.1)2 was utilized to detect and eliminate chimera sequences, resulting in obtaining clean reads. Sequences with a similarity of >97% were clustered into the same operational taxonomic unit (OTU) using USEARCH (v10.0, Edgar, 2013), and OTUs with counts less than 2 in all samples were filtered out.
The data regarding growth performance, rumen fermentation, and blood parameters, represented as mean values, were gathered and analyzed using one-way ANOVA in SPSS (v26.0). Multiple comparisons were carried out utilizing the LSD method subsequent to identifying significant differences, with a significance level set at p < 0.05.
Taxonomy annotation of the OTUs was performed based on the Naive Bayes classifier in QIIME2 (Bolyen et al., 2019) using the SILVA database (Quast et al., 2013) with a confidence threshold of 70%. Alpha diversity was assessed to determine the species diversity complexity of each sample using QIIME2 software. Beta diversity calculations were conducted via Non-Metric Multi-Dimensional Scaling (NMDS) to evaluate the diversity in samples for species complexity. To investigate the influence on microbial communities, metastats analysis (White et al., 2009) was performed with R (v3.1.1). The R package of psych (v2.1.9), igraph (v1.2.5) and visNetwork (v2.1.0) we used for drafting the correlation network diagram.
ADG, DMI, and F/G of five groups during the experimental period are presented in Table 3. The results indicated that there were no significant differences in ADG and DMI among the Hu sheep across the five groups. The F/G ratios for the groups were 7.94, 8.08, 8.00, 8.89, and 9.16, respectively; notably, the F/G ratio in the PSMS20 group was a significantly higher value than that in the control group (p < 0.05; Table 3).
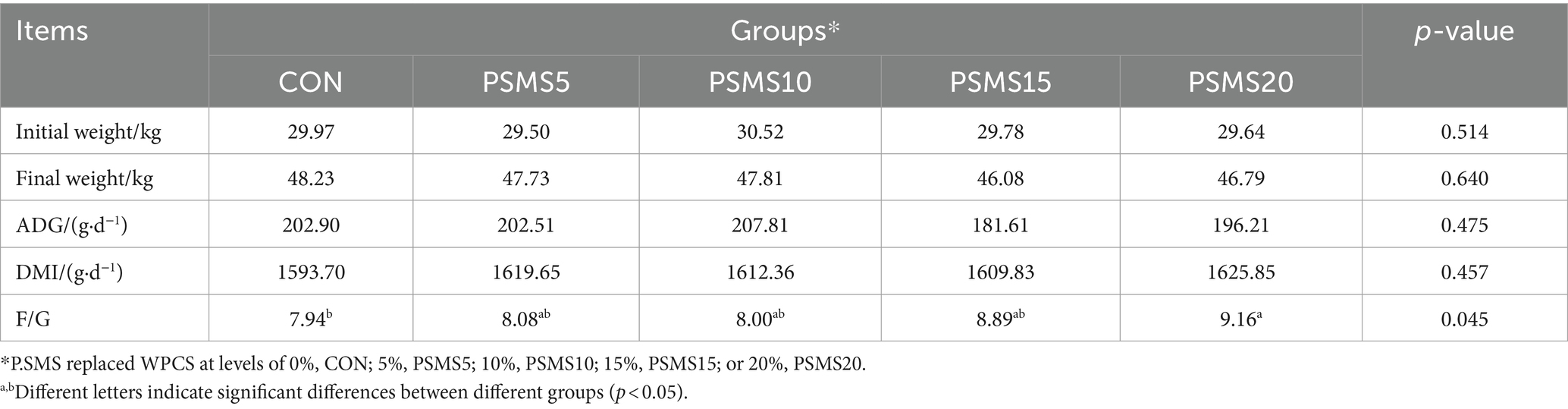
Table 3. Effects of dietary P.SMS supplementation on the growth performance of Hu sheep throughout the experimental period.
As illustrated in Figure 1, the NH3-N concentration in PSMS15 was significantly higher than that in CON, PSMS5, and PSMS20 (p < 0.05). Additionally, the NH3-N concentration in PSMS10 and PSMS20 was significantly elevated compared to CON and PSMS5 (p < 0.05).
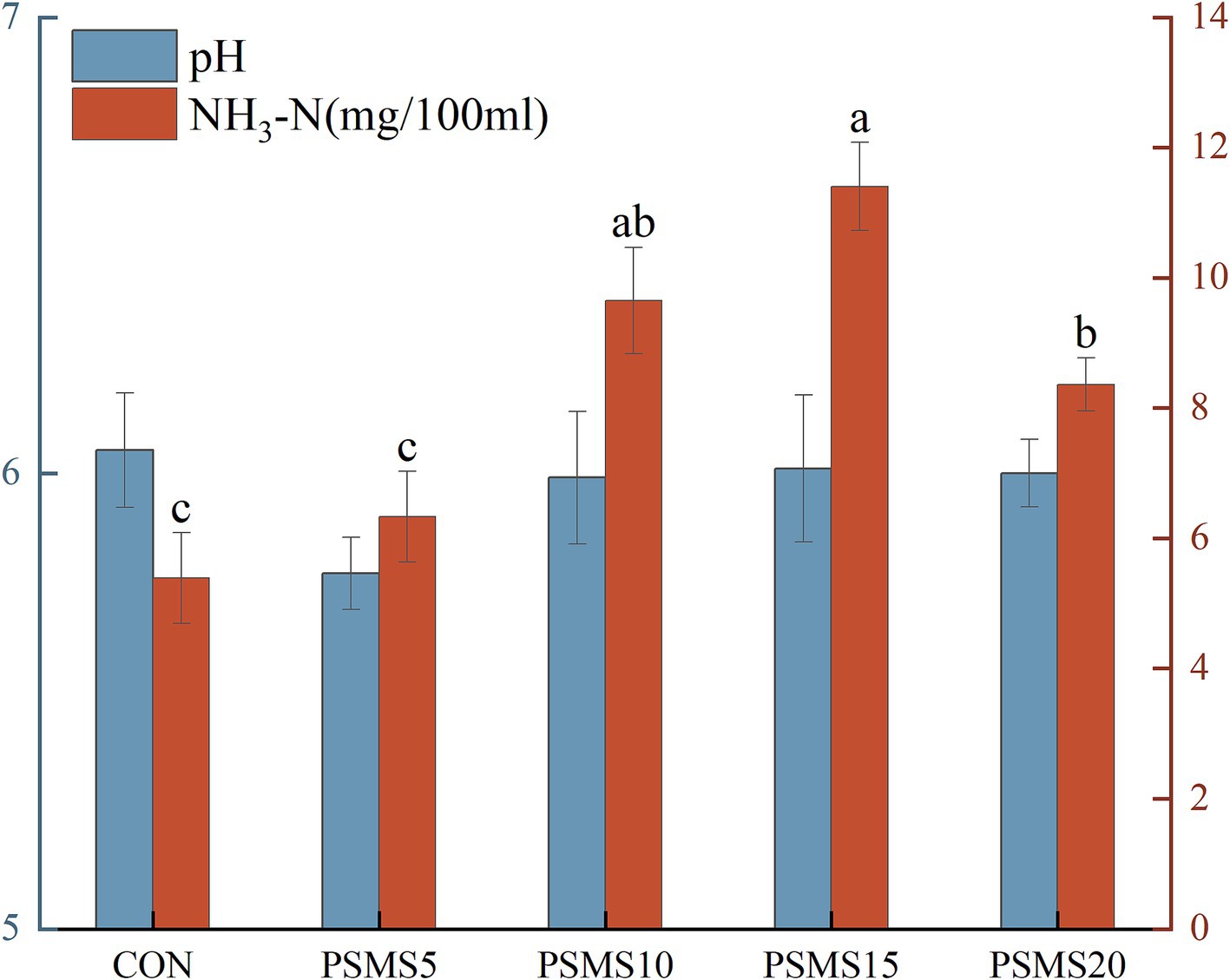
Figure 1. Effects of different proportions of P.SMS on rumen pH and NH3-N in sheep. The labels ‘a,’ ‘b,’ and ‘c’ above the bars indicate significant differences between different groups (p < 0.05). The absence of labels indicates no significant difference. P.SMS replaced WPCS at levels of 0%, CON; 5%, PSMS5; 10%, PSMS10; 15%, PSMS15; or 20%, PSMS20.
Table 4 presents the measurements of rumen papilla length and width, as well as rumen muscular thickness. The results indicate an increase in both the length and width of the rumen papilla with increasing proportions of P.SMS. Specifically, PSMS15 exhibited a significantly greater rumen papilla length compared to both CON and PSMS5 (p < 0.05). Moreover, PSMS15 and PSMS20 demonstrated significantly wider rumen papillae compared to CON and PSMS5 (p < 0.05).
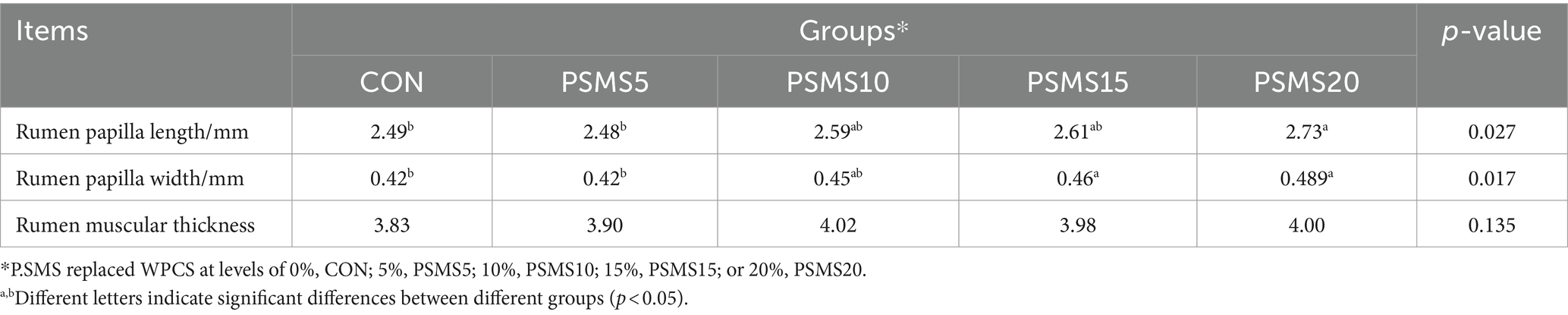
Table 4. Effects of P.SMS on rumen tissue morphology in Hu sheep.
Figure 2 presents the hematological and serological parameters before and after feeding throughout the experiment, highlighting significant differences in white blood cell count, hemoglobin concentration, and creatinine levels following P.SMS feeding. Notably, white blood cell counts were higher in PSMS5 and PSMS20 compared to PSMS10, with a significant increase in PSMS5 relative to both CON and PSMS15 (p < 0.05). Additionally, hemoglobin concentration exhibited a significant increase in PSMS20 compared to CON (p < 0.05). Furthermore, creatinine levels were significantly elevated in PSMS20 compared to CON, PSMS5, and PSMS10 (p < 0.05).
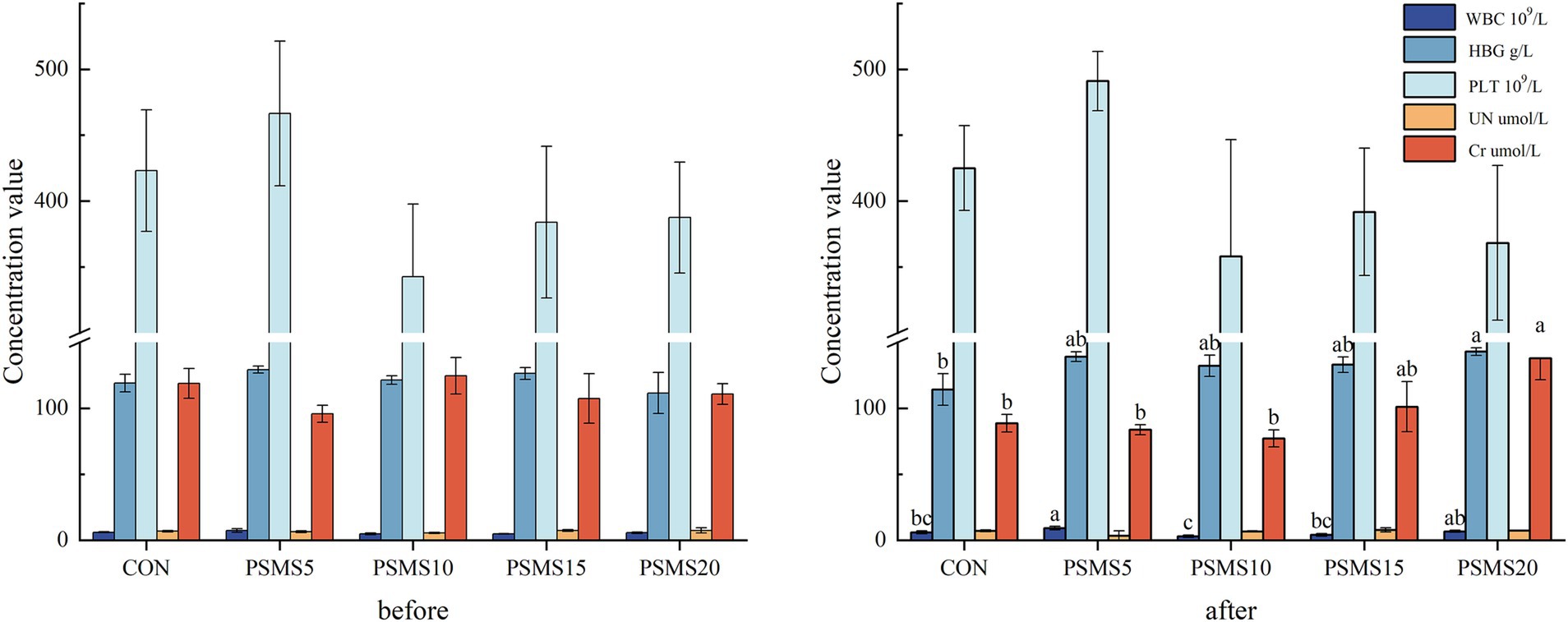
Figure 2. Selected blood parameters before and after feeding P.SMS. The labels ‘a’, ‘b’, and ‘c’ above the bars indicate significant differences between different groups (p < 0.05). The absence of labels indicates no significant difference. WBC, white blood cell; HGB, hemoglobin concentration; PLT, Platelet count; UN, urea nitrogen; Cr, creatinine. P.SMS replaced WPCS at levels of 0%, CON; 5%, PSMS5; 10%, PSMS10; 15%, PSMS15; or 20%, PSMS20.
The raw data were processed according to the aforementioned method, and data quality was assessed by evaluating factors such as read length and read counts at each stage, as detailed in Supplementary Table 1. The sequencing results for bacterial and fungal samples revealed the following ranges for clean circular consensus sequences (CCS), sequence effectiveness, and sequencing lengths: 6713–8,451 and 4,078–8,417; 94.84–98.98% and 99.13–99.95%; 1,454–1,459 bp and 593–653 bp, respectively.
Based on the analysis of 97% of nucleotide sequences from a total of 90,133 bacterial reads, 862 Operational Taxonomic Units (OTUs) were identified (Figure 3a). Among these, 454 OTUs were common to all five groups, representing 52.67% of the total OTUs. The number of unique OTUs in CON, PSMS5, PSMS10, PSMS15, and PSMS20 groups were 3 (0.35%), 5 (0.58%), 1 (0.12%), 13 (1.51%), and 3 (0.35%), respectively. PSMS15 exhibited the highest number of OTUs, whereas PSMS20 showed the lowest.
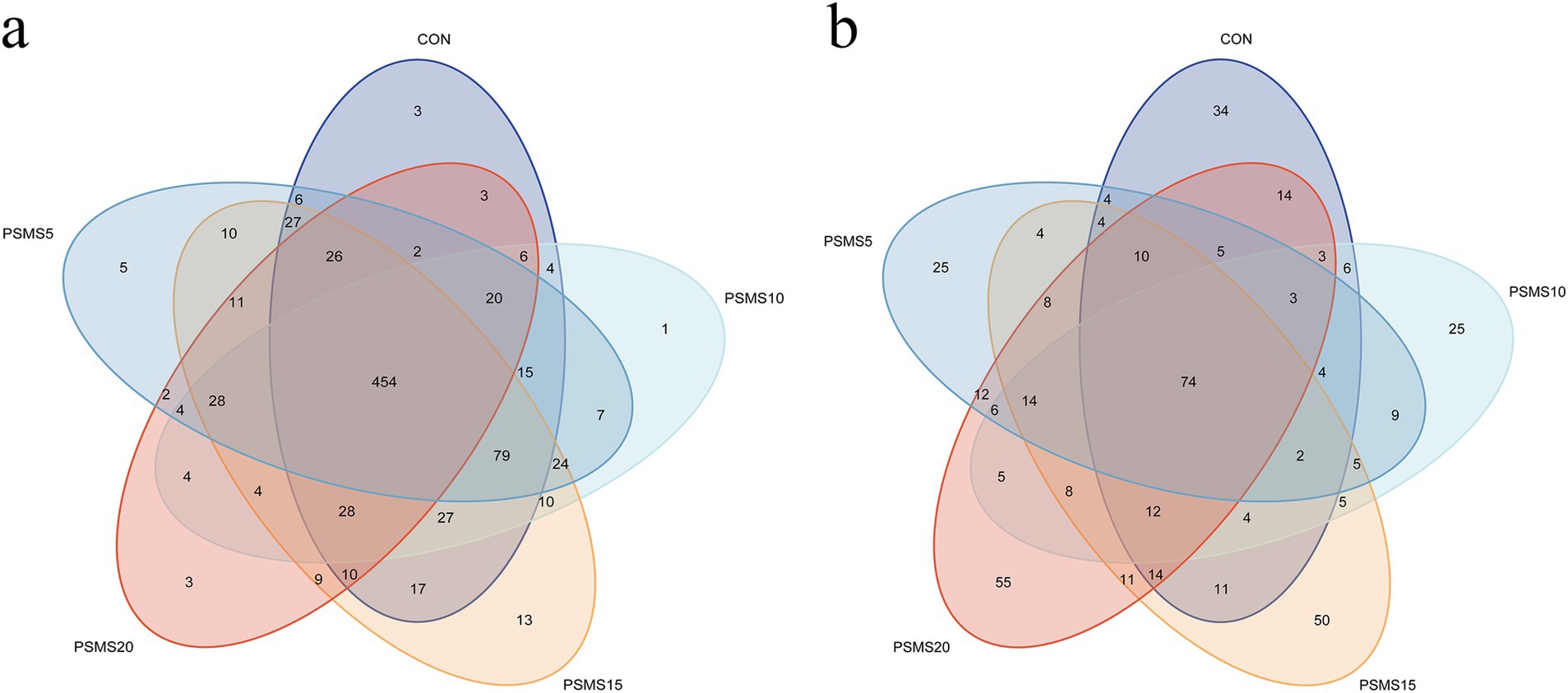
Figure 3. Venn diagram of OTUs in (a) rumen bacteria and (b) fungi of Hu sheep. The different colors of the ellipses in the Venn diagram represent different groups, and the overlapping regions indicate the number of shared features among the groups. The non-overlapping regions represent the number of features specific to each group. P.SMS replaced WPCS at levels of 0%, CON; 5%, PSMS5; 10%, PSMS10; 15%, PSMS15; or 20%, PSMS20.
For fungal sequencing, a total of 178,688 reads identified 446 OTUs (Figure 3b). Among these, 74 OTUs were shared among all five groups, accounting for 16.59% of the total OTUs. The unique OTU counts in CON, PSMS5, PSMS10, PSMS15, and PSMS20 were 34 (7.62%), 25 (5.61%), 25 (5.61%), 50 (11.21%), and 55 (12.33%), respectively. PSMS20 displayed the highest number of OTUs, while CON had the lowest.
Table 5 indicates that the bacterial and fungal indices did not exhibit significant differences among the groups. The Fungal ACE index and Chao1 index demonstrated a pattern of initially decreasing and then increasing (p > 0.05), with the fungal group in PSMS10 displaying the lowest abundance of the fungal community. A closer inspection of the table, reveals that fungal indices showed a decreasing trend in both the abundance and diversity of the fungal community from CON to PSMS5. However, as community abundance decreased, community diversity increased from PSMS5 to PSMS10. The increase in community abundance from PSMS5 to PSMS10 and PSMS15 was accompanied by a trend toward flattening community diversity. Conversely, the trend of bacterial diversity in the rumen was opposite to that of fungi. Notably, the decrease in colony abundance from CON to PSMS5 was associated with an increase in colony diversity. The trends in bacterial colony abundance and colony diversity were consistent across the other groups.
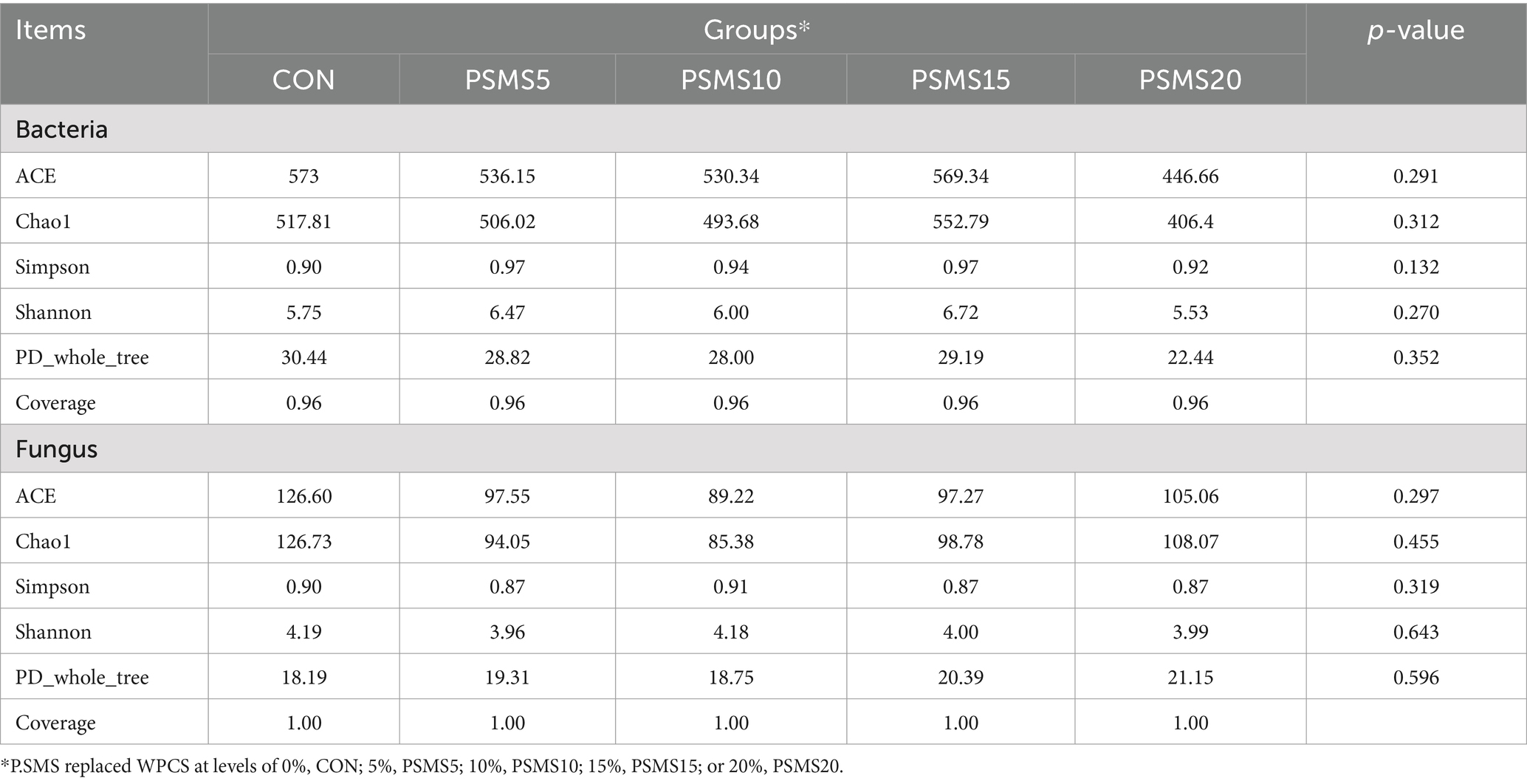
Table 5. Effects of P.SMS on Alpha diversity of rumen bacteria and fungi in Hu sheep.
As depicted in Figure 4, both stress values in the NMDS analyses are below 0.2, indicating a relatively reliable NMDS analysis. Both analyses revealed no significant differences in the distribution of bacterial and fungal microbiota among the groups. However, there appeared to be a tendency for the distribution of fungal communities to cluster, particularly between PSMS15 and PSMS20.
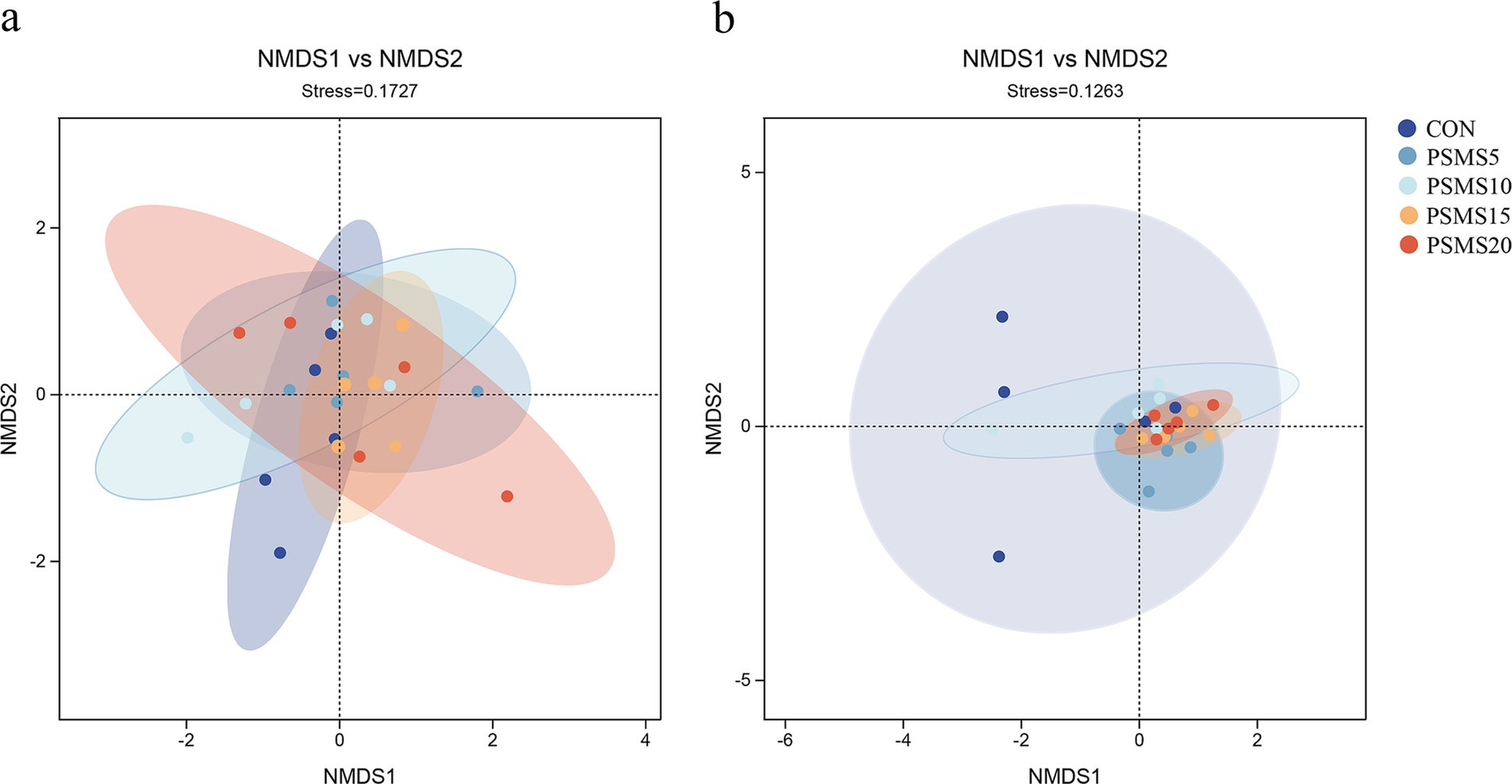
Figure 4. NMDS analysis of (a) rumen bacteria and (b) fungi of Hu sheep. Each point in the figure represents a sample; different colors represent different subgroups; the oval circle indicates that it was a 95% confidence ellipse. When Stress is less than 0.2, it indicates that the NMDS analysis has some reliability. The closer the samples are to each other on the coordinate graph, the higher the similarity. P.SMS replaced WPCS at levels of 0%, CON; 5%, PSMS5; 10%, PSMS10; 15%, PSMS15; or 20%, PSMS20.
The dominant organisms at the phylum level of rumen microbiota in all five groups of Hu sheep were Firmicutes and Bacteroidetes, with relative abundances ranging from 63.10 to 72.28% (Supplementary Table 3). Notably, as illustrated in Figure 5a and Supplementary Table 3 Firmicutes displayed an increasing trend followed by a decrease across the groups. PSMS5 had the highest abundance, while PSMS20 exhibited the lowest (p > 0.05). In contrast, Bacteroidetes showed a decreasing trend followed by an increase across the groups, with PSMS20 demonstrating the highest abundance and PSMS10 the lowest (p > 0.05). The ratio of Firmicutes to Bacteroidetes across the five groups was 0.94, 1.37, 1.53, 1, and 0.81, indicating an upward trend followed by a downward trend, as shown in Supplementary Table 3. At the genus level, as depicted in Figure 5b and Supplementary Table 4, Prevotella and uncultured_rumen_bacterium were the dominant genera, with relative abundances ranging from 36.61 to 44.47%. Prevotella exhibited a decreasing trend followed by an increase, with the highest abundance found in PSMS10 and the lowest in CON (p > 0.05). The Christensenellaceae_R_7_group also displayed an increasing and then decreasing trend, with the highest abundance in PSMS10 and the lowest in CON (p > 0.05).
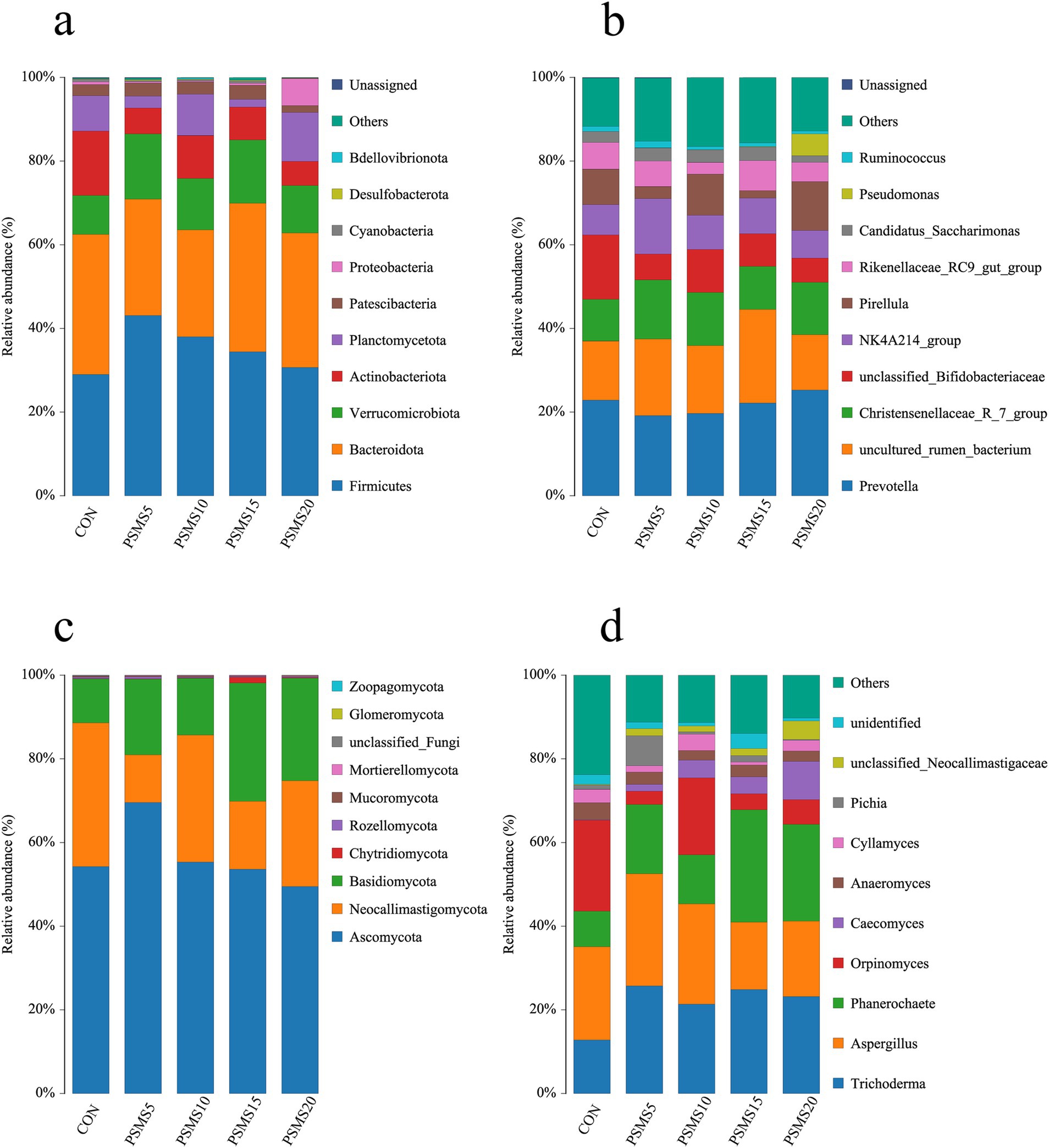
Figure 5. The relative abundance of ruminal bacteria at the level of (a) phylum and (b) genus. The relative abundance of ruminal fungi at the level of (c) phylum and (d) genus. The horizontal axis in the figure represents group names, and the vertical axis represents relative abundance percentages. Different colors represent different species, and stacked bars indicate the top 10 taxonomic groups of relative abundance at each classification level. P.SMS replaced WPCS at levels of 0%, CON; 5%, PSMS5; 10%, PSMS10; 15%, PSMS15; or 20%, PSMS20.
The predominant organisms at the phylum level of rumen fungi in all five groups of Hu sheep were Ascomycota and Neocallimastigomycota, with relative abundances ranging from 69.41 to 89.00%. Notably, Ascomycota displayed a trend of initially increasing and then decreasing with the addition of P.SMS, peaking in PSMS5 and reaching its lowest point in PSMS20 (p > 0.05). (Supplementary Table 3 and Figure 5c). Furthermore, Supplementary Table 3 indicates that the percentage of Basidiomycota was significantly higher in PSMS15 and PSMS20 compared to CON, and significantly higher in PSMS15 than in PSMS10 (p < 0.05). Additionally, Mucoromycota showed an initial increase followed by a subsequent decrease, with PSMS10 exhibiting the highest percentage and CON the lowest (p > 0.05). At the genus level, as illustrated in Figure 5d and Supplementary Table 4, Trichoderma and Aspergillus emerged as the dominant genera, with relative abundances ranging from 33.35 to 52.84%. According to Supplementary Table 4, the percentage of Phanerochaete was significantly higher in PSMS15 compared to CON and PSMS10, while both PSMS15 and PSMS20 were significantly higher than CON, PSMS5, and PSMS10 (p < 0.05).
LEfSe analysis of the rumen microbiota revealed the dominance of specific bacterial and fungal groups, as illustrated in Figure 6. Figures 6a,c highlighted the presence of 7 bacterial and 11 fungal differences observed at various taxonomic levels between the CON and PSMS15 groups. At the phylum level, Cyanobacteria and Basidiomycota were identified as biomarkers distinguishing CON from PSMS15. Figures 6b,d provide a comparison of the distribution of these two phyla across different groups. The Cyanobacteria phylum was found to be relatively abundant in the CON group, whereas the Basidiomycota phylum was more prevalent in PSMS15.
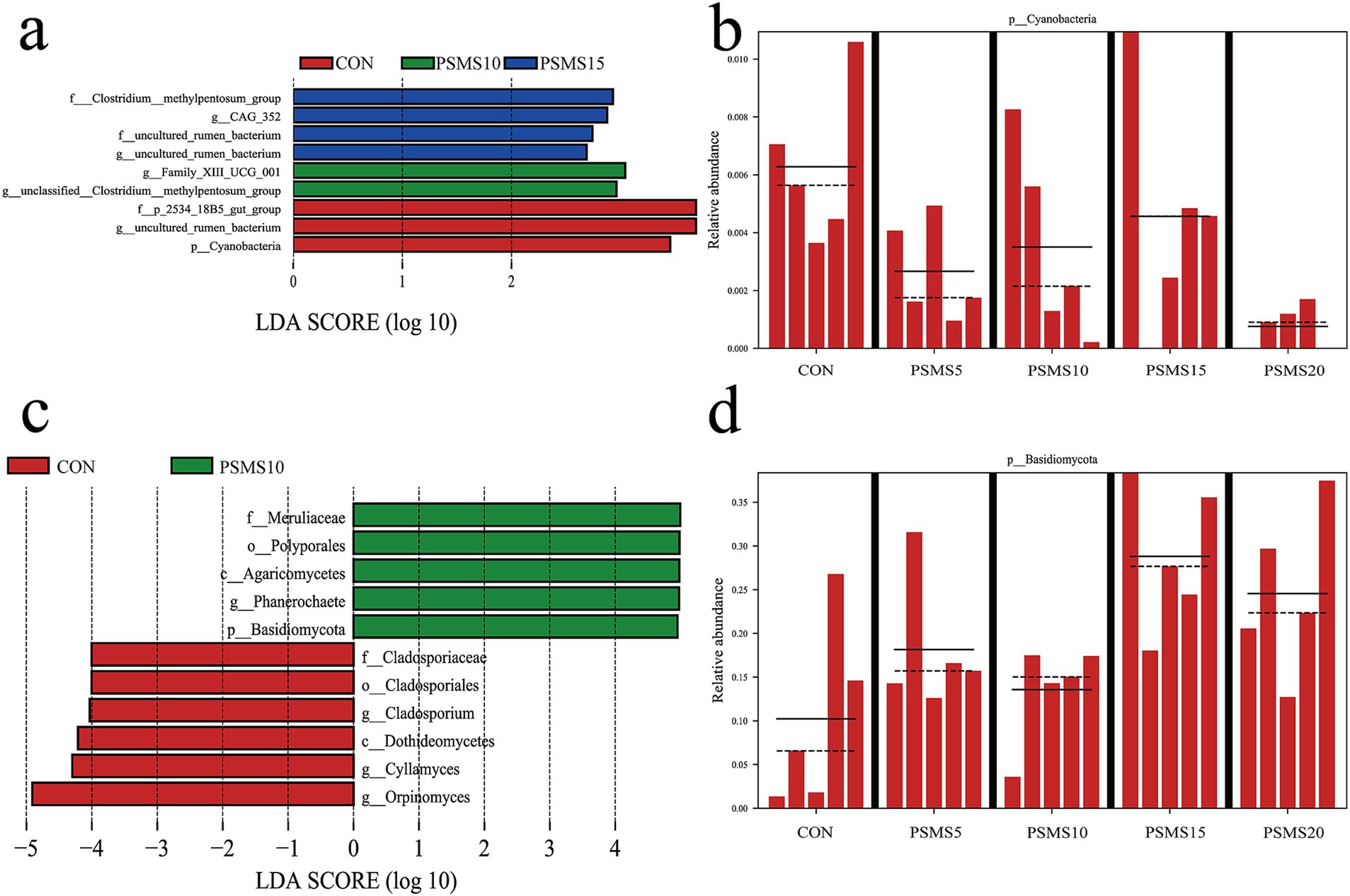
Figure 6. LEfSe analysis of rumen bacteria and fungi. Linear discriminant analysis effect size cladogram based on classification of (a) rumen bacteria and (c) fungi. Comparison of the relative abundance of (b) bacterial and (d) fungal biomarkers at the phylum level between five groups. P.SMS replaced WPCS at levels of 0%, CON; 5%, PSMS5; 10%, PSMS10; 15%, PSMS15; or 20%, PSMS20.
In Figure 7a, the majority of bacterial genera at the center of the association are classified within the Firmicutes phylum, while the Desulfovibrio genus (NO. 7) from the Desulfobacterota phylum also occupies a central position. The figure primarily illustrates a positive correlation. After excluding undefined and uncultured bacteria, we observed the presence of two pairs of weaker negative correlations: one pair consists of Candidatus Omnitrophus (NO. 1) and Saccharofermentans (NO. 25), while the other pair includes Prevotella (NO. 42) and Christensenellaceae_R_7_group (NO. 30).
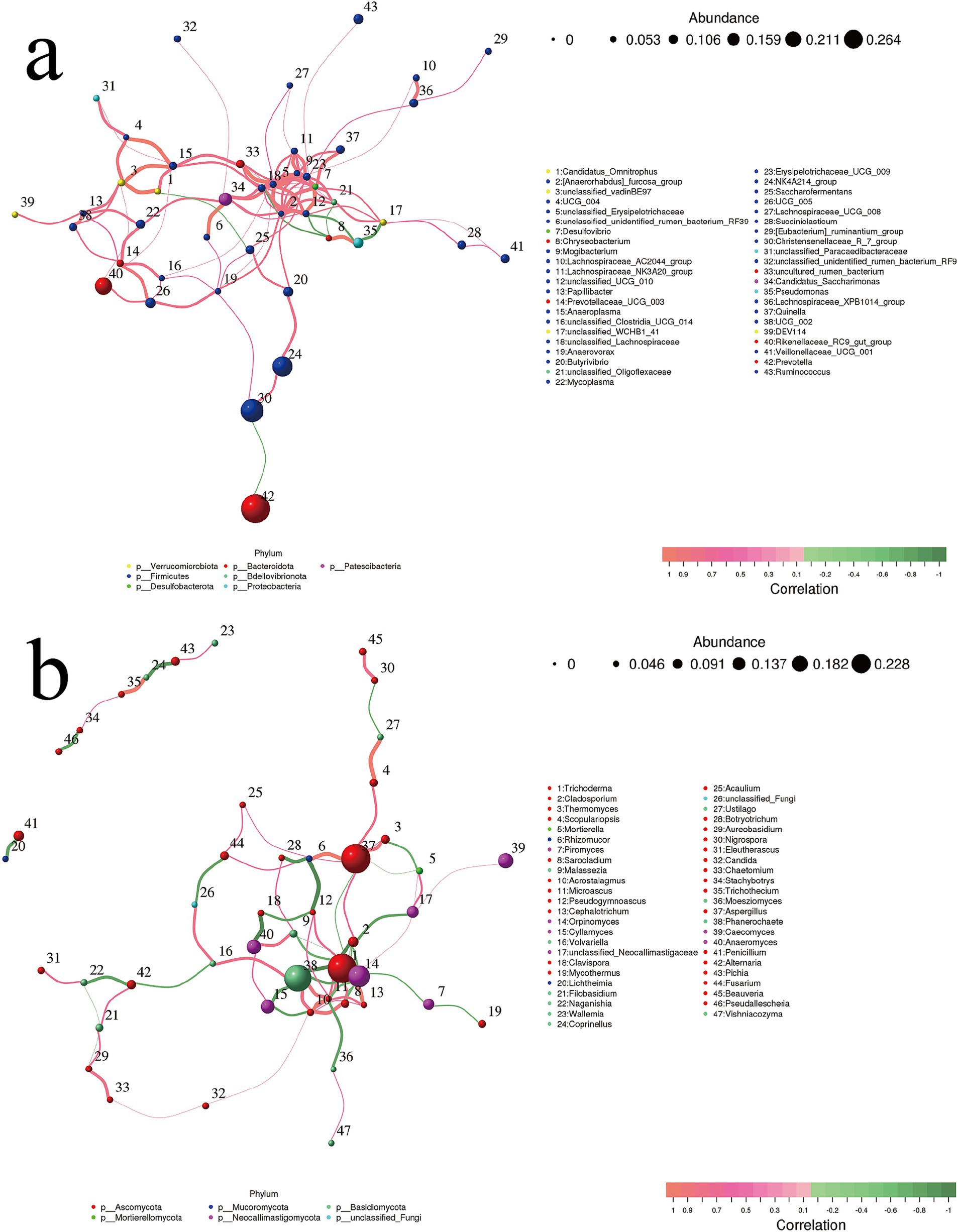
Figure 7. Correlation network of top 50 genera for (a) rumen bacteria and (b) fungi. Species are represented by circles, where the size of each circle indicates the abundance of the species. Furthermore, different colors are used to denote distinct phylum classifications. Edges represent the correlation between two species, with line thickness indicating strength and line color indicating correlation direction: orange for positive correlation and green for negative correlation. P.SMS replaced WPCS at levels of 0%, CON; 5%, PSMS5; 10%, PSMS10; 15%, PSMS15; or 20%, PSMS20.
In Figure 7b and Supplementary Figure 1, two distinct layout arrangements of the correlation analysis network are presented. The main interactions are concentrated among three phyla: Ascomycota, Basidiomycota, and Neoxallimastigomycota. Specifically, Trichoderma (NO. 1) and Aspergillus (NO. 37) are members of Ascomycota, Phanerochaete (NO. 38) belongs to Basidiomycota, and Orpincomycota (NO. 14) is part of Neoxallimastigomycota, all of which contribute to the core interactions. Two clusters of fungi show independent correlations: one between Ascomycota and Basidiomycota, and the other between Ascomycota and Mucoromycota (Figure 7b).
The correlation network of the top 20 genera between rumen bacteria and fungi is illustrated in Figure 8, which reveals a statistically significant negative correlation between Cladosporium and Prevotellaceae_UCG_001, with an absolute correlation coefficient exceeding 0.65.
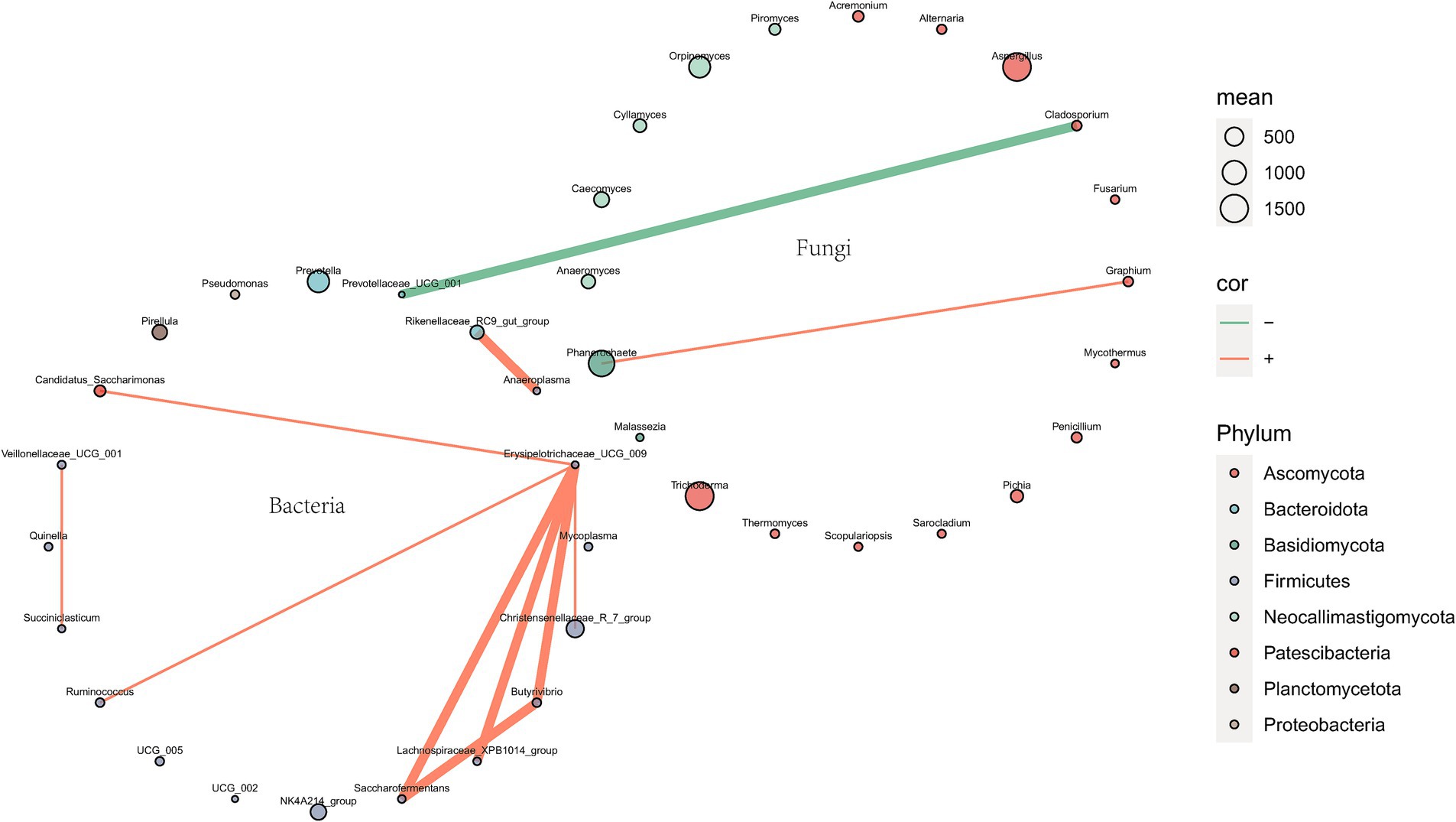
Figure 8. Correlation network of top 20 genera between rumen bacteria and fungi. Species are represented by circles, where the size of each circle indicates the abundance of the species. Furthermore, edges represent the correlation between two species, with a width of 2 when the absolute value of correlation >0.65 and a width of 0.5 otherwise. The line color indicates correlation direction: orange for positive correlation and green for negative correlation. P.SMS replaced WPCS at levels of 0%, CON; 5%, PSMS5; 10%, PSMS10; 15%, PSMS15; or 20%, PSMS20.
Each kilogram of mushroom production yields approximately five kilograms of SMS (Lucas De Jesus et al., 2023). SMS is classified as agricultural waste and is often either landfilled, incinerated, or disposed of improperly, which prevents the full utilization of its untapped nutrients and mycelium (Huang et al., 2023; Yang et al., 2022). However, the implementation of reuse or recycling strategies could significantly reduce this solid waste, generating new economic opportunities and promoting positive environmental outcomes (Huang et al., 2023). In addition to the residual nutrients in SMS, mushroom polysaccharides present in the fermentation liquid of edible fungi, mycelium, or fruit bodies contribute to regulating the gut microbiome, enhancing the intestinal mucosal barrier, modulating lipid metabolism, and increasing the production of short-chain fatty acids (Zhao J. et al., 2023). While some studies have explored the utilization of SMS, limited attention has been paid to its effects on rumen bacterial composition and the relationships between rumen bacteria and fungi. In our study, varied results were observed as the percentage of substitutions processed SMS (P.SMS) changed.
In this experiment, the F/G was significantly higher in the PSMS20 group compared to the CON group, while ADG and DMI did not differ among the five groups. This finding indicates a lower feed utilization efficiency when a high percentage of P.SMS was fed. Hanlon et al. (2023) noted that nutrient synchronization of protein and carbohydrates is an effective strategy for enhancing rumen nutrient utilization. The F/G ratio of PSMS20 was significantly greater than that of the CON group. This may be attributed to the fact that, although the protein content increased with the proportion of P.SMS, the efficiency of nutrient utilization decreased due to the lower soluble carbohydrate content of P.SMS compared to whole-plant corn silage. PSMS15 exhibited a significantly higher NH3-N concentration compared to CON, PSMS5, and PSMS20, while PSMS10 and PSMS20 also demonstrated significantly higher NH3-N concentrations than CON and PSMS5. The NH3-N content is influenced by the rate of microbial degradation of nitrogenous substances in the ration, as well as the rate of NH3-N utilization by microorganisms and rumen epithelial cells (Liang et al., 2024). Rehemujiang et al. (2023) indicated that the normal physiological range of rumen NH3-N is 6-30 mg/dL; all groups were within this normal range except for the CON group, which had NH3-N levels exceeding the minimum concentration required for the development of rumen microorganisms, which is 5 mg/dL. Despite the consistent nitrogen content in diets formulated according to isoenergetics and isonitrogen principles, variations were observed in the microbial and rumen epithelial utilization rates, as well as in the composition of bacteria and fungi at both the phylum and genus levels. These variations may contribute to differences in the NH3-N concentrations. Shen et al. (2023) noted a positive correlation between rumen fluid pH and its free ammonia nitrogen (FAN), along with a strong negative correlation between FAN and microbial population. However, there were no significant differences in rumen fluid pH among the five groups, suggesting that there may also be no significant difference in FAN among these groups. Rumen papillae length and width, along with rumen musculature thickness, are key indicators of rumen development (Lesmeister et al., 2004). In this experiment, with the exception of PSMS5, both the length and width of the rumen papillae increased with higher proportions of P.SMS added, indicating that the inclusion of P.SMS can promote rumen development in sheep. Gadelha et al. (2014) reported that monogastric animals, including birds, fish, pigs, and rodents, exhibit greater susceptibility to gossypol toxicity compared to ruminants; these effects tend to accumulate over a period of 1 to 3 months of ingestion. This was further corroborated by blood parameters observed in this experiment, where Cr content was significantly higher in PSMS20 compared to CON, PSMS5, and PSMS10, and the concentration of HGB was significantly higher in PSMS20 than in CON. Serum Cr levels reflect the animal’s utilization of proteins and amino acids, and a high serum Cr level may indicate impairment of kidney function (Wong Vega et al., 2020). The presence of gossypol may also affect the utilization of Fe2+ ions in the blood, potentially impacting the function of HGB as an oxygen transport carrier, leading to alterations in oxygen-carrying capacity (Gadelha et al., 2014).
In the analysis of bacterial composition in this experiment, changes were observed in the phyla Bacteroidetes and Firmicutes, indicating shifts in substrate utilization patterns. This microbiome investigation revealed that Bacteroidetes and Firmicutes were the dominant bacterial phyla, consistent with previous studies (Chen et al., 2022; Cui Y. et al., 2022; Han et al., 2019; Pinnell et al., 2022). These two phyla serve as indicators for assessing the energy needs of ruminants (Xue et al., 2017), with Bacteroidetes being responsible for acetate and propionate production, while Firmicutes are responsible for butyrate production (Cui Y. et al., 2022). The Firmicutes/Bacteroidetes ratio across the five groups demonstrated a clear trend of change, indicating significant structural alterations related to substrate utilization. Gharechahi and Salekdeh (2018) found that Bacteroidetes-associated species exhibited a high abundance of genes encoding debranching and oligosaccharide-degrading enzymes, whereas Firmicutes-associated species showed a richness in cellulases and hemicellulases. White et al. (2014) pointed that Firmicutes utilize cell surface-associated cellulose-degrading complexes, while Bacteroidetes employ outer membrane binding proteins encoded by gene clusters to degrade soluble polysaccharides primarily within the periplasmic or intracellular space. The transition from Bacteroidetes being the dominant phylum to Firmicutes, as observed in the experiment conducted by Xue et al. (2022), was attributed to a decreased level of dietary physically effective neutral detergent fiber (peNDF). P.SMS was provided in a crushed state while WPCS was fed in its original form, the crushed P.SMS exhibited a lower NDF compared to the original WPCS (Tables 1, 2). The experiment conducted by Palangi et al. (2022) also demonstrated a significant reduction in the acid detergent fiber and crude fiber contents of the substrate following mushroom production. This difference may contribute to the observed transition of the dominant phylum from Bacteroidetes to Firmicutes as the proportion of P.SMS from 0 to 10%, aligns with the findings of Xue et al. (2022). Ma G. et al. (2022) reported that the polysaccharide from Pleurotus eryngii significantly promoted the relative abundance of Firmicutes while inhibiting that of Bacteroidetes, suggesting a similar effect may occur with the polysaccharide in Pleurotus ostreatus. Furthermore, as the proportion of P.SMS continued to increase, the dominant phylum transitioned from Firmicutes to Bacteroidetes in PSMS15 and PSMS20. Although the level of NDF decreased with the increasing proportion of P.SMS in the diet, there was a gradual increase in the content of gossypol in cottonseed hulls, which may contribute to this transition. Wang et al. (2021) noted that low concentrations of gossypol enhanced the abundance of rumen bacteria involved in fiber and starch metabolism; however, gossypol at high concentrations (>0.5 mg/g) exhibited an inhibitory effect on the activity of rumen microbes, including six genera within the Firmicutes phylum. This conclusion, consistent with the current experimental findings, reflects both the significantly higher NH3-N content in PSMS15 compared to CON, PSMS5, and PSMS20, and the observed transition of the major phyla from Firmicutes to Bacteroidetes. Given that the two phyla share common characteristics for the degradation of phytopolysaccharides, they may be particularly efficient in utilizing nitrogenous material at a close ratio. Xue et al. (2022) discovered in their experiment that Prevotella and Christensenellaceae_R_7_group were the dominant genera, exhibiting a positive quadratic correlation with the level of peNDF. Zhang X. et al. (2022) confirmed that Prevotella was indeed the dominant genus across all samples, while the relative abundance of Christensenellaceae_R_7_group varied significantly among the treatment groups. Other studies have also identified the prevailing status of these genera within the microbiota (Cui et al., 2021; Zhao W. et al., 2023). Numerous reports have highlighted the strong capacity of Prevotella for the degradation of hemicellulose (Cui Y. et al., 2022; Li et al., 2015), as it plays a crucial role in the decomposition of starch, xylan, crude proteins, and pectin (Han et al., 2019; Huang et al., 2023; Qian et al., 2019; Xue et al., 2022). Christensenellaceae is a prevalent bacterial family in both human and animal guts, potentially capable of metabolizing non-fiber carbohydrates (Zhang et al., 2023). Previous studies have suggested possible associations between Christensenellaceae, lipid metabolism, and ether extract digestibility (Shang et al., 2021; Waters and Ley, 2019; Zhang et al., 2023). The observed negative correlation between the abundance changes of Christensenellaceae_R_7_group and Prevotella may be attributed to their competition for similar nutrient resources within the gut environment. While both Christensenellaceae_R_7_group and Prevotella play significant roles in feed digestion, they exhibit different preferences for specific types of carbohydrates. Consequently, fluctuations in the abundance of one genus may influence the growth and survival of the other, leading to a negative correlation between their respective abundances.
In addition to the two phyla that account for a relatively large proportion, Patescibacteria and Cyanobacteria were also affected by the addition of P.SMS. Patescibacteria, also known as the candidate phyla radiation (CPR), constitute a diverse and disproportionately large fraction of microbial dark matter, which remains largely unidentified, unclassified, and understudied (Wang et al., 2023). Members of the CPR exhibit a variety of unique traits, including their small width, compact genomes typically less than 1 Mb, and limited metabolic capabilities (Ji et al., 2022; Wang et al., 2023). This has led to the hypothesis of their universal dependence on host organisms for growth, which may explain the positive correlation observed between Erysipelotrichaceae_UCG_009 and Candidatus_Saccharimonas. Experimental findings support this notion, indicating that the majority of cultivated Patescibacteria adhere to and thrive on the surfaces of other bacteria, functioning as obligate epibionts (Batinovic et al., 2021; Kuroda et al., 2022). Erysipelotrichaceae are likely related to host metabolism or cholesterol phenotypes in humans and mice, while in ruminants, they may aid in the degradation of plant fibers in the rumen (Kaakoush, 2015; Qian et al., 2019; Zhang et al., 2023). The genera Candidatus_Saccharimonas and Christensenellaceae_R_7_group, as previously mentioned, demonstrated a positive correlation with metabolites involved in energy substrate metabolism and amino acid biosynthesis (Ogunade et al., 2019). Zhen et al. (2023) observed that the addition of both sodium acetate and sodium propionate resulted in increased abundances of specific rumen bacteria, including Candidatus Saccharimonas and Christensenellaceae R-7. The molar proportion of acetate significantly increased with gossypol supplementation (Wang et al., 2020), subsequently elevating the proportion of Candidatus Saccharimonas in PSMS5, PSMS10, and PSMS15. However, as the proportion of gossypol increased, the abundance of Candidatus_Saccharimonas decreased, potentially due to a negative effect of gossypol on its growth or the promotion of other bacteria that inhibited the growth of Candidatus_Saccharimonas in PSMS20. Furthermore, the positive correlation between Candidatus_Saccharimonas and the Christensenellaceae_R_7_group suggests a potential interaction within the gut microbiota, indicating their joint involvement in energy metabolism and nutrient synthesis. Cyanobacteria, commonly referred to as blue-green algae, were significantly less abundant in PSMS20 compared to other groups. The Cyanobacteria present in the rumen are recognized as polysaccharolytic organisms, capable of breaking down complex polysaccharides (McGovern et al., 2018). As algal populations proliferate, algal blooms can lead to adverse consequences including changes in water color, elevated biotoxin levels in aquatic organisms, and harm to both humans and other organisms (Hallegraeff et al., 2021). Previous studies have demonstrated that the enzyme systems of white-rot fungi exhibit an inhibitory effect on algae and that these fungi can utilize algae as a carbon and nitrogen source for growth and utilization (Yu et al., 2023). It is evident that a large proportion of P.SMS results in a significant reduction in Cyanobacteria abundance; this may be attributed to either the inhibitory effect of gossypol or the utilization of algae by the enzyme systems of white-rot fungi. The precise mechanisms underlying this phenomenon warrant further investigation.
In the realm of rumen microbiota research, a noticeable disparity exists between the number of studies conducted on rumen fungi and those on rumen bacteria. Anaerobic fungi have been established as a significant component of the population of ruminal microorganisms responsible for fiber digestion (Hartinger and Zebeli, 2021; Zhang et al., 2007). Among the fungal kingdoms, Basidiomycota and Ascomycota, are the dominant phyla with the latter comprising over 1.5 million species. Basidiomycota is distinguished from Ascomycota by the presence of specialized cellular structures called basidia, where haploid sexual spores (basidiospores) are formed (Schmidt-Dannert, 2016). Despite the increasing proportions of P.SMS, a significant increase in Basidiomycota abundance was observed (p < 0.05). However, at the genus level, Phanerochaete exhibited consistent changes in abundance alongside Basidiomycota. This suggests that the observed variations in Basidiomycota are more likely attributable to fluctuations in Phanerochaete rather than to residuals of Pleurotus ostreatus in the diet. Furthermore, these results may also stem from the potential misidentification of Pleurotus as Phanerochaete during the amplicon analysis, possibly due to similarities in their genetic sequences targeted by the primers used. Additionally, we speculated that Phanerochaete may have also been present in the basal diet, which could explain its detection in the CON group. According to Datsomor et al. (2022), the co-culture of P. chrysosporium and P. ostreatus did not demonstrate a significant advantage over axenic cultures in achieving a substantial reduction in lignin content or enriching the biomass with holo-cellulose. This finding indicates a prevailing mutual intermingling and inhibitory relationship between P. chrysosporium and P. ostreatus, despite their ability to grow together on the same medium. The inhibition relationship between P. chrysosporium and P. ostreatus is likely responsible for the changes in Basidiomycota abundance.
In this experiment, a negative correlation was observed between Prevotellaceae_UCG_001 (belonging to Bacteroidota) and Cladosporium (belonging to Ascomycota) among bacteria and fungi. Prevotellaceae_UCG_001, a strain of Prevotella, plays a crucial role in the degradation of various substrates, including hemicellulose, proteins, starches, xylan, and pectin (Zhang H. et al., 2022). This strain exhibited a strong correlation with the enrichment score of the AMPK signaling pathway, which is recognized as a vital sensor in regulating glycolipid metabolism (Song et al., 2019). Cladosporium is primarily known as a ubiquitous environmental saprophytic fungus or plant endophyte (Bensch et al., 2015). Although a few species have been documented as etiologic agents in vertebrate hosts, the number of such species identified to date remains limited (Sandoval-Denis et al., 2016). A significant positive correlation was found between Cladosporium abundance and isobutyrate concentration, while a significant negative correlation was observed between Cladosporium abundance and the levels of β-alanine and xanthine (Cui X. et al., 2022). Ascomycota, the most abundant fungal phylum across all groups in this study, comprises species that predominantly produce mycelia, which may contribute to the breakdown of plant cell walls (Wallace et al., 2019; Han et al., 2019). The pricise cause of this negative correlation remains unclear, necessitating further investigations to explore additional perspectives, such as examining the correlation between metabolites, to confirm and illuminate this relationship.
SMS shows promise as a feed resource due to its availability and potential to reduce the environmental impact of mushroom production residues. However, its nutritional consistency varies based on substrate types and cultivation practices, necessitating a thorough evaluation before integration into livestock diets. Processing methods aimed at enhancing digestibility and mitigating anti-nutritional factors are crucial considerations. Given that the substrate exhibits lower NDF and ADF after mushroom treatment, it can provide more fiber to microorganisms. However, the reduced carbohydrate content may limit its role in protein synthesis by these microorganisms. Therefore, proper formulation and feeding strategies are essential to realizing the benefits of P.SMS in livestock nutrition and sustainable farming practices.
In this experiment, we aimed to utilize P.SMS to determine the appropriate dosage and evaluate its effects on the rumen flora of Hu sheep. Alongside growth performance, rumen indices, and blood indices, our findings indicate that feeding a high percentage of P.SMS over an extended period adversely affects feed utilization efficiency, blood oxygen-carrying capacity, and kidney function, likely due to the influence of cottonseed hulls used in this study. Conversely, a moderate percentage of P.SMS appears to enhance rumen absorption capacity. Consistent with established expectations regarding rumen microbes, the addition of P.SMS impacted individual phylum-level and genus-level microorganisms; however, there is no evident rationale to implicate mushroom polysaccharides or mycelium. Furthermore, within the parameters of this experiment, no correlation was observed between changes in the microbial community structures of bacteria and fungi. In conclusion, substituting 10% of P.SMS for whole-plant corn silage proved to be the most effective approach in this study, demonstrating the feasibility of P.SMS feeding. However, the addition ratio is constrained by the P.SMS substrate, which varies by region. Notably, the substrate treated with white-rot fungi exhibited an enhancement in digestion and absorption in Hu sheep. Further experimental studies are needed to determine whether mushroom polysaccharides or mycelium affect rumen microbes.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: https://www.ncbi.nlm.nih.gov/, PRJNA1085887.
The animal study was approved by the Tarim University Science and Technology Ethics Committee. The study was conducted in accordance with the local legislation and institutional requirements.
M-LL: Conceptualization, Methodology, Writing – original draft, Writing – review & editing. G-HY: Methodology, Writing – review & editing. HR: Conceptualization, Formal analysis, Methodology, Writing – review & editing. C-CL: Data curation, Methodology, Writing – review & editing. L-HH: Methodology, Writing – review & editing. P-PD: Methodology, Writing – review & editing. L-dZ: Methodology, Writing – review & editing. Q-YD: Formal analysis, Methodology, Writing – review & editing. K-DD: Formal analysis, Methodology, Writing – review & editing. G-SX: Formal analysis, Funding acquisition, Methodology, Project administration, Resources, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the Corps Science and Technology Development Special Project “Meat Sheep Industry Science and Technology Support Project in Bostan Village, Kavak Township, Moyu County” (2018DA001-1). This work was also supported by the presidential research fund of Tarim University “Optimization of bacterial and enzyme treatment parameters and evaluation of feeding value of spent substrate of oyster mushroom (Pleurotus ostreatus)” (TDZKYB202303).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2024.1425218/full#supplementary-material
1. ^ github.com
2. ^ drive5.com
Agustinho, B. C., Daniel, J. L. P., Zeoula, L. M., Alcalde, C. R., Machado, E., Bragatto, J. M., et al. (2021). Enzymatic effects of Pleurotus ostreatus spent substrate on whole-plant corn silage and performance of lactating goats. J. Dairy Sci. 104, 11660–11672. doi: 10.3168/jds.2021-20775
Baptista, F., Campos, J., Costa-Silva, V., Pinto, A. R., Saavedra, M. J., Ferreira, L. M., et al. (2023). Nutraceutical potential of Lentinula edodes' spent mushroom substrate: a comprehensive study on phenolic composition, antioxidant activity, and antibacterial effects. J. Fungi 9:1200. doi: 10.3390/jof9121200
Batinovic, S., Rose, J. J. A., Ratcliffe, J., Seviour, R. J., and Petrovski, S. (2021). Cocultivation of an ultrasmall environmental parasitic bacterium with lytic ability against bacteria associated with wastewater foams. Nat. Microbiol. 6, 703–711. doi: 10.1038/s41564-021-00892-1
Bensch, K., Groenewald, J. Z., Braun, U., Dijksterhuis, J., de Jesús Yáñez-Morales, M., and Crous, P. W. (2015). Common but different: the expanding realm of Cladosporium. Stud. Mycol. 82, 23–74. doi: 10.1016/j.simyco.2015.10.001
Berger, R. G., Bordewick, S., Krahe, N. K., and Ersoy, F. (2022). Mycelium vs. fruiting bodies of edible Fungi-a comparison of metabolites. Microorganisms 10:1379. doi: 10.3390/microorganisms10071379
Bolyen, E., Rideout, J. R., Dillon, M. R., Bokulich, N. A., Abnet, C. C., al-Ghalith, G. A., et al. (2019). Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857. doi: 10.1038/s41587-019-0209-9
Chen, X., An, M., Zhang, W., Li, K., Kulyar, M. F., Duan, K., et al. (2022). Integrated Bacteria-Fungi diversity analysis reveals the gut microbial changes in Buffalo with mastitis. Front. Vet. Sci. 9:918541. doi: 10.3389/fvets.2022.918541
Cui, Y., Liu, H., Gao, Z., Xu, J., Liu, B., Guo, M., et al. (2022). Whole-plant corn silage improves rumen fermentation and growth performance of beef cattle by altering rumen microbiota. Appl. Microbiol. Biotechnol. 106, 4187–4198. doi: 10.1007/s00253-022-11956-5
Cui, X., Liu, Y., Wu, H., Meng, Q., Liu, S., Chai, S., et al. (2022). Dynamic changes in the yak rumen eukaryotic community and metabolome characteristics in response to feed type. Front. Vet. Sci. 9:1027967. doi: 10.3389/fvets.2022.1027967
Cui, X., Wang, Z., Tan, Y., Chang, S., Zheng, H., Wang, H., et al. (2021). Selenium yeast dietary supplement affects rumen bacterial population dynamics and fermentation parameters of Tibetan sheep (Ovis aries) in alpine meadow. Front. Microbiol. 12:663945. doi: 10.3389/fmicb.2021.663945
Datsomor, O., Gou-Qi, Z., and Miao, L. (2022). Effect of ligninolytic axenic and coculture white-rot fungi on rice straw chemical composition and in vitro fermentation characteristics. Sci. Rep. 12:1129. doi: 10.1038/s41598-022-05107-z
de Vos, W. M., Tilg, H., Van Hul, M., and Cani, P. D. (2022). Gut microbiome and health: mechanistic insights. Gut 71, 1020–1032. doi: 10.1136/gutjnl-2021-326789
Edgar, R. C. (2013). UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 10, 996–998. doi: 10.1038/nmeth.2604
Feng, Z. C., and Gao, M. (2010). Improvement of the method for determining the ammonia-nitrogen content of rumen liquor by colourimetry. Anim. Husb. Feed Sci. 31:37. doi: 10.16003/j.cnki.issn1672-5190.2010.z1.027
Gadelha, I. C. N., Fonseca, N. B. S., Oloris, S. C. S., Melo, M. M., Soto-Blanco, B., and Chase, C. C. (2014). Gossypol toxicity from cottonseed products. ScientificWorldJournal 2014, 231611–231635. doi: 10.1155/2014/231635
Gharechahi, J., and Salekdeh, G. H. (2018). A metagenomic analysis of the camel rumen's microbiome identifies the major microbes responsible for lignocellulose degradation and fermentation. Biotechnol. Biofuels 11:216. doi: 10.1186/s13068-018-1214-9
Guohong, Y., Lihong, H. U., Changchang, L. I., Lidong, Z., Pingping, D., Mulong, L. U., et al. (2023). 22Effects of dietary Pleurotus ostreatus substrate supplementation on growth performance and nutrient apparent digestibility of fattening Hu sheep. Chin. Anim. Hus. Vet. Med. 50, 3985–3993. doi: 10.16431/j.cnki.1671-7236.2023.10.012
Hallegraeff, G., Enevoldsen, H., and Zingone, A. (2021). Global harmful algal bloom status reporting. Harmful Algae 102:101992. doi: 10.1016/j.hal.2021.101992
Han, X., Li, B., Wang, X., Chen, Y., and Yang, Y. (2019). Effect of dietary concentrate to forage ratios on ruminal bacterial and anaerobic fungal populations of cashmere goats. Anaerobe 59, 118–125. doi: 10.1016/j.anaerobe.2019.06.010
Hanlon, M. E., Simoni, M., Moorby, J. M., Righi, F., Tsiplakou, E., Kantas, D., et al. (2023). Effects of the addition of non-fibre carbohydrates with different rumen degradation rates in dairy cow high-forage diets using the rumen simulation technique. Animal 17:100732. doi: 10.1016/j.animal.2023.100732
Hartinger, T., and Zebeli, Q. (2021). The present role and new potentials of anaerobic Fungi in ruminant nutrition. J. Fungi 7:200. doi: 10.3390/jof7030200
Huang, X., Zhou, L., You, X., Han, H., Chen, X., and Huang, X. (2023). Production performance and rumen bacterial community structure of Hu sheep fed fermented spent mushroom substrate from Pleurotus eryngii. Sci. Rep. 13:8696. doi: 10.1038/s41598-023-35828-8
Ji, Y., Zhang, P., Zhou, S., Gao, P., Wang, B., and Jiang, J. (2022). Widespread but poorly understood Bacteria: candidate Phyla radiation. Microorganisms 10:2232. doi: 10.3390/microorganisms10112232
Kaakoush, N. O. (2015). Insights into the role of Erysipelotrichaceae in the human host. Front. Cell. Infect. Microbiol. 5:84. doi: 10.3389/fcimb.2015.00084
Kainthola, J., Podder, A., Fechner, M., and Goel, R. (2021). An overview of fungal pretreatment processes for anaerobic digestion: applications, bottlenecks and future needs. Bioresour. Technol. 321:124397. doi: 10.1016/j.biortech.2020.124397
Knop, D., Yarden, O., and Hadar, Y. (2015). The ligninolytic peroxidases in the genus Pleurotus: divergence in activities, expression, and potential applications. Appl. Microbiol. Biotechnol. 99, 1025–1038. doi: 10.1007/s00253-014-6256-8
Kuroda, K., Yamamoto, K., Nakai, R., Hirakata, Y., Kubota, K., Nobu, M. K., et al. (2022). Symbiosis betweenCandidatus Patescibacteria and Archaea discovered in wastewater-treating bioreactors. MBio 13:e0171122. doi: 10.1128/mbio.01711-22
Lamontagne, J., Rico, D. E., Perdomo, C. M., Ronholm, J., Gervais, R., and Chouinard, P. Y. (2023). Effects of direct-fed Bacillus subtilis and Bacillus licheniformis on production performance and milk fatty acid profile in dairy cows. J. Dairy Sci. 106, 1815–1825. doi: 10.3168/jds.2022-22564
Lesmeister, K. E., Tozer, P. R., and Heinrichs, A. J. (2004). Development and analysis of a rumen tissue sampling procedure. J. Dairy Sci. 87, 1336–1344. doi: 10.3168/jds.S0022-0302(04)73283-X
Li, Z., Wright, A. D., Liu, H., Fan, Z., Yang, F., Zhang, Z., et al. (2015). Response of the rumen microbiota of sika deer (Cervus nippon) fed different concentrations of tannin rich plants. PLoS One 10:e0123481. doi: 10.1371/journal.pone.0123481
Liang, J., Zhang, R., Chang, J., Chen, L., Nabi, M., Zhang, H., et al. (2024). Rumen microbes, enzymes, metabolisms, and application in lignocellulosic waste conversion - a comprehensive review. Biotechnol. Adv. 71:108308. doi: 10.1016/j.biotechadv.2024.108308
Lucas De Jesus, G., José Lavoranti, O., Schafer, G., Dias De Oliveira, G., De Andrade, S., Silva, R., et al. (2023). Nutrient uptake in supplemented substrate by oyster mushroom. World J. Microbiol. Biotechnol. 39:175. doi: 10.1007/s11274-023-03622-3
Ma, Y., Han, X., Fang, J., and Jang, H. (2022). Role of dietary amino acids and microbial metabolites in the regulation of pig intestinal health. Anim. Nutr. 9, 1–6. doi: 10.1016/j.aninu.2021.10.004
Ma, G., Xu, Q., Du, H., Muinde Kimatu, B., Su, A., Yang, W., et al. (2022). Characterization of polysaccharide from Pleurotus eryngii during simulated gastrointestinal digestion and fermentation. Food Chem. 370:131303. doi: 10.1016/j.foodchem.2021.131303
Martin, M. (2011). Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 17, 10–12. doi: 10.14806/ej.17.1.200
Martin, C., Zervakis, G. I., Xiong, S., Koutrotsios, G., and Straetkvern, K. O. (2023). Spent substrate from mushroom cultivation: exploitation potential toward various applications and value-added products. Bioengineered 14:2252138. doi: 10.1080/21655979.2023.2252138
Martinez-Guryn, K., Hubert, N., Frazier, K., Urlass, S., Musch, M. W., Ojeda, P., et al. (2018). Small intestine microbiota regulate host digestive and absorptive adaptive responses to dietary lipids. Cell Host Microbe 23, 458–469.e5. doi: 10.1016/j.chom.2018.03.011
McGovern, E., Kenny, D. A., McCabe, M. S., Fitzsimons, C., McGee, M., Kelly, A. K., et al. (2018). 16S rRNA sequencing reveals relationship between potent cellulolytic genera and feed efficiency in the rumen of bulls. Front. Microbiol. 9:1842. doi: 10.3389/fmicb.2018.01842
Ogunade, I., Schweickart, H., McCoun, M., Cannon, K., and McManus, C. (2019). Integrating 16S rRNA sequencing and LC−MS-based metabolomics to evaluate the effects of live yeast on rumen function in beef cattle. Animals 9:28. doi: 10.3390/ani9010028
Palangi, V., Kaya, A., Kaya, A., and Giannenas, I. (2022). Ecofriendly usability of mushroom cultivation substrate as a ruminant feed: anaerobic digestion using gas production techniques. Animals 12:1583. doi: 10.3390/ani12121583
Pinnell, L. J., Reyes, A. A., Wolfe, C. A., Weinroth, M. D., Metcalf, J. L., Delmore, R. J., et al. (2022). Bacteroidetes and Firmicutes drive differing microbial diversity and community composition among Micro-environments in the bovine rumen. Front. Vet. Sci. 9:897996. doi: 10.3389/fvets.2022.897996
Qian, W., Ao, W., Jia, C., and Li, Z. (2019). Bacterial colonisation of reeds and cottonseed hulls in the rumen of Tarim red deer (Cervus elaphus yarkandensis). Antonie Van Leeuwenhoek 112, 1283–1296. doi: 10.1007/s10482-019-01260-0
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., et al. (2013). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41, D590–D596. doi: 10.1093/nar/gks1219
Rehemujiang, H., Yusuf, H. A., Ma, T., Diao, Q., Kong, L., Kang, L., et al. (2023). Fermented cottonseed and rapeseed meals outperform soybean meal in improving performance, rumen fermentation, and bacterial composition in Hu sheep. Front. Microbiol. 14:1119887. doi: 10.3389/fmicb.2023.1119887
Sandoval-Denis, M., Gené, J., Sutton, D. A., Wiederhold, N. P., Cano-Lira, J. F., and Guarro, J. (2016). New species of Cladosporium associated with human and animal infections. Persoonia 36, 281–298. doi: 10.3767/003158516X691951
Schmidt-Dannert, C. (2016). Biocatalytic portfolio of Basidiomycota. Curr. Opin. Chem. Biol. 31, 40–49. doi: 10.1016/j.cbpa.2016.01.002
Scholtmeijer, K., van den Broek, L. A. M., Fischer, A. R. H., and van Peer, A. (2023). Potential protein production from lignocellulosic materials using edible mushroom forming Fungi. J. Agric. Food Chem. 71, 4450–4457. doi: 10.1021/acs.jafc.2c08828
Shang, Q., Liu, S., Liu, H., Mahfuz, S., and Piao, X. (2021). Impact of sugar beet pulp and wheat bran on serum biochemical profile, inflammatory responses and gut microbiota in sows during late gestation and lactation. J. Anim. Sci. Biotechnol. 12:54. doi: 10.1186/s40104-021-00573-3
Shen, J., Zheng, W., Xu, Y., and Yu, Z. (2023). The inhibition of high ammonia to in vitro rumen fermentation is pH dependent. Front. Vet. Sci. 10:1163021. doi: 10.3389/fvets.2023.1163021
Shin, M., Lee, J. H., Lee, J. A., Kim, M. J., Park, H., Park, B. W., et al. (2021). Immunomodulatory and anti-inflammatory effects of Phellinus linteus mycelium. BMC Complement. Med. Ther. 21:269. doi: 10.1186/s12906-021-03441-9
Song, Y. M., Lee, S. D., Chowdappa, R., Kim, H. Y., Jin, S. K., and Kim, I. S. (2007). Effects of fermented oyster mushroom (Pleurotus ostreats) by-product supplementation on growth performance, blood parameters and meat quality in finishing Berkshire pigs. Animal 1, 301–307. doi: 10.1017/S1751731107683785
Song, X., Zhong, L., Lyu, N., Liu, F., Li, B., Hao, Y., et al. (2019). Inulin can alleviate metabolism disorders in Ob/Ob mice by partially restoring leptin-related pathways mediated by gut microbiota. Genomics Proteomics Bioinformatics 17, 64–75. doi: 10.1016/j.gpb.2019.03.001
van Kuijk, S. J. A., Sonnenberg, A. S. M., Baars, J. J. P., Hendriks, W. H., and Cone, J. W. (2015). Fungal treated lignocellulosic biomass as ruminant feed ingredient: a review. Biotechnol. Adv. 33, 191–202. doi: 10.1016/j.biotechadv.2014.10.014
Wallace, R. J., Sasson, G., Garnsworthy, P. C., Tapio, I., Gregson, E., Bani, P., et al. (2019). A heritable subset of the core rumen microbiome dictates dairy cow productivity and emissions. Sci. Adv. 5:eaav8391. doi: 10.1126/sciadv.aav8391
Wang, Y., Gallagher, L. A., Andrade, P. A., Liu, A., Humphreys, I. R., Turkarslan, S., et al. (2023). Genetic manipulation of Patescibacteria provides mechanistic insights into microbial dark matter and the epibiotic lifestyle. Cell 186, 4803–4817.e13. doi: 10.1016/j.cell.2023.08.017
Wang, C., Li, Y., MaiTiSaiYiDi, T., Yang, H., and Yang, K. (2020). Effect of dietary gossypol supplement on fermentation characteristics and bacterial diversity in the rumen of sheep. PLoS One 15:e0234378. doi: 10.1371/journal.pone.0234378
Wang, W., Wang, Y., Li, W., Wu, Q., Li, S., and Yang, H. (2021). Gossypol exhibited higher detrimental effect on ruminal fermentation characteristics of low-forage in comparison with high-forage mixed feeds. Toxics 9:51. doi: 10.3390/toxics9030051
Waters, J. L., and Ley, R. E. (2019). The human gut bacteria Christensenellaceae are widespread, heritable, and associated with health. BMC Biol. 17:83. doi: 10.1186/s12915-019-0699-4
White, B. A., Lamed, R., Bayer, E. A., and Flint, H. J. (2014). Biomass utilization by gut microbiomes. Ann. Rev. Microbiol. 68, 279–296. doi: 10.1146/annurev-micro-092412-155618
White, J. R., Nagarajan, N., and Pop, M. (2009). Statistical methods for detecting differentially abundant features in clinical metagenomic samples. PLoS Comput. Biol. 5:e1000352. doi: 10.1371/journal.pcbi.1000352
Wong Vega, M., Swartz, S. J., Devaraj, S., and Poyyapakkam, S. (2020). Elevated serum creatinine: but is it renal failure? Pediatrics 146:e20192828. doi: 10.1542/peds.2019-2828
Xu, C., Yimin, C., Jianguo, Z., and Hiroki, M. (2010). Feeding value of total mixed ration silage with spent mushroom substrate. Anim. Sci. J. 81, 194–198. doi: 10.1111/j.1740-0929.2009.00728.x
Xue, D., Chen, H., Zhao, X., Xu, S., Hu, L., Xu, T., et al. (2017). Rumen prokaryotic communities of ruminants under different feeding paradigms on the Qinghai-Tibetan plateau. Syst. Appl. Microbiol. 40, 227–236. doi: 10.1016/j.syapm.2017.03.006
Xue, B., Wu, M., Yue, S., Hu, A., Li, X., Hong, Q., et al. (2022). Changes in rumen bacterial community induced by the dietary physically effective neutral detergent Fiber levels in goat diets. Front. Microbiol. 13:820509. doi: 10.3389/fmicb.2022.820509
Yang, S., Fang, C., Chen, Y., Chen, W., Lee, L., Chen, C., et al. (2022). Hericium erinaceus mycelium ameliorates in vivo progression of osteoarthritis. Nutrients 14:2605. doi: 10.3390/nu14132605
Yu, H., Lei, P., Ma, J., Jin, J., Ma, Y., Fang, Y., et al. (2023). The potential of white-rot fungi for algal control: mechanisms, strategies, and challenges. Environ. Res. 236:116738. doi: 10.1016/j.envres.2023.116738
Zhang, Y., Gao, W., and Meng, Q. (2007). Fermentation of plant cell walls by ruminal bacteria, protozoa and fungi and their interaction with fibre particle size. Arch. Anim. Nutr. 61, 114–125. doi: 10.1080/17450390701204020
Zhang, H., Lang, X., Zhang, Y., and Wang, C. (2022). Distribution of bacteria in different regions of the small intestine with Zanthoxylum bungeanum essential oil supplement in small-tailed Han sheep. Front. Microbiol. 13:1062077. doi: 10.3389/fmicb.2022.1062077
Zhang, X., Liu, X., Chang, S., Zhang, C., Du, W., and Hou, F. (2022). Effect of Cistanche deserticola on rumen microbiota and rumen function in grazing sheep. Front. Microbiol. 13:840725. doi: 10.3389/fmicb.2022.840725
Zhang, J., Shang, J., Hao, Y., Wang, Y., Cao, Z., Yang, H., et al. (2023). Growth performance, blood metabolites, ruminal fermentation, and bacterial community in preweaning dairy calves fed corn silage-included starter and total mixed ration. J. Dairy Sci. 106, 4545–4558. doi: 10.3168/jds.2022-22476
Zhao, W., Abdelsattar, M. M., Wang, X., Zhang, N., Chai, J., and Fu, Y. (2023). In vitro modulation of rumen fermentation by microbiota from the recombination of rumen fluid and solid phases. Microbiol. Spectr. 11:e0338722. doi: 10.1128/spectrum.03387-22
Zhao, J., Hu, Y., Qian, C., Hussain, M., Liu, S., Zhang, A., et al. (2023). The interaction between mushroom polysaccharides and gut microbiota and their effect on human health: a review. Biology 12:122. doi: 10.3390/biology12010122
Zhen, Y., Xi, Z., Nasr, S. M., He, F., Han, M., Yin, J., et al. (2023). Multi-omics reveals the impact of exogenous short-chain fatty acid infusion on rumen homeostasis: insights into crosstalk between the microbiome and the epithelium in a goat model. Microbiol. Spectr. 11:e0534322. doi: 10.1128/spectrum.05343-22
Keywords: white-rot fungi, fungal pretreatment, gossypol, rumen microorganisms, spent mushroom substrate
Citation: Lu M-L, Yuan G-H, Rehemujiang H, Li C-C, Hu L-H, Duan P-P, Zhang L-D, Diao Q-Y, Deng K-D and Xu G-S (2024) Effects of spent substrate of oyster mushroom (Pleurotus ostreatus) on ruminal fermentation, microbial community and growth performance in Hu sheep. Front. Microbiol. 15:1425218. doi: 10.3389/fmicb.2024.1425218
Edited by:
Gonzalo Martinez Fernandez, Commonwealth Scientific and Industrial Research Organisation (CSIRO), AustraliaReviewed by:
Houqiang Luo, Wenzhou Vocational College of Science and Technology, ChinaCopyright © 2024 Lu, Yuan, Rehemujiang, Li, Hu, Duan, Zhang, Diao, Deng and Xu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gui-Shan Xu, Z3Vpc2hhbnh1QDEyNi5jb20=
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.