 Marcos Martins
Marcos Martins Cássia Siqueira César
Cássia Siqueira César Rodrigo Cogni
Rodrigo Cogni- Department of Ecology, University of São Paulo, São Paulo, Brazil
Facultative inheritable endosymbionts are common and diverse in insects and are often found at intermediate frequencies in insect host populations. The literature assessing the relationship between environment and facultative endosymbiont frequency in natural host populations points to temperature as a major component shaping the interaction. However, a synthesis describing its patterns and mechanistic basis is lacking. This mini-review aims to bridge this gap by, following an evolutionary model, hypothesizing that temperature increases endosymbiont frequencies by modulating key phenotypes mediating the interaction. Field studies mainly present positive correlations between temperature and endosymbiont frequency at spatial and seasonal scales; and unexpectedly, temperature is predominantly negatively correlated with the key phenotypes. Higher temperatures generally reduce the efficiency of maternal transmission, reproductive parasitism, endosymbiont influence on host fitness and the ability to protect against natural enemies. From the endosymbiont perspective alone, higher temperatures reduce titer and both high and low temperatures modulate their ability to promote host physiological acclimation and behavior. It is necessary to promote research programs that integrate field and laboratory approaches to pinpoint which processes are responsible for the temperature correlated patterns of endosymbiont prevalence in natural populations.
1 Introduction
Nonessential or facultative inheritable endosymbionts are common and diverse in insects and are often found at intermediate frequencies in natural host populations (Oliver et al., 2014; Smith et al., 2021), prevailing as an intriguing and successful ecological division of the tree of life. Their long-term maintenance are associated with their contrasting acting as parasitic modulators of host’s reproductive system and/or by being mutualistic enhancers of host fitness (Zug and Hammerstein, 2015; Corbin et al., 2017; Renoz et al., 2019). Environmental factors directly affect the strength of the phenotypes induced by symbionts and the efficiency of maternal transmission (Oliver et al., 2014; Corbin et al., 2017; Smith et al., 2021). However, field studies exploring the missing link between the functional ecology of these inheritable microbes with environmental factors remains scarce (Oliver et al., 2014).
An evolutionary model states three key parameters regulating the dynamics and equilibria of facultative endosymbiont frequencies in their host populations: i. the proportion of uninfected individuals produced by infected ones (i.e., strength of imperfect maternal transmission); ii. the fitness of infected individuals relative to the uninfected ones (i.e., strength of mutualistic boost); iii. The costs associated with infection and its transmission phenotypes (i.e., strength of parasitism and parasitic reproductive manipulation; Hoffmann et al., 1990; Hague et al., 2020a). Environmental factors are important selective agents driving all those parameters in nature, hence spatial and temporal variation are expected to reflect different patterns of correlated endosymbiont prevalence in the field. A synthesis on the topic is complex and not yet established. Abiotic factors such as temperature (Hague et al., 2022), humidity and presence of inorganic toxicants (Lemoine et al., 2020), biotic factors such as parasitism and disease (Leclair et al., 2021), host resource (Hague et al., 2020a), competition (Gavotte et al., 2010), and symbiont multiple infections (Toju and Fukatsu, 2011; Zhu et al., 2018) assuredly play a role (Oliver et al., 2014; Heyworth and Ferrari, 2015). Among abiotic factors, temperature might be pivotal in modulating the conditioning for host physiology performance, resource availability and presence and abundance of natural enemies (Leclair et al., 2021), thus affecting endosymbiont titer and host-symbiont interactions (Hague et al., 2020a). Indeed, Corbin et al. (2017) reviews that thermal environments affect endosymbiotic association and its ability to alter host thermotolerance across many species beyond insects, but did not aim to link facultative endosymbiont prevalence patterns to the causative processes. Thus, temperature role remains elusive in shaping facultative endosymbiont success.
This mini-review aims to synthesize the current knowledge and bridge the gap by hypothesizing that temperature increases endosymbiont frequencies by modulating key phenotypes mediating the interaction. Field studies exploring the spatial and temporal variation in endosymbiont frequency related to temperature, as laboratory essays exploring the role of temperature on endosymbiont mediated interaction following the previous evolutionary model are essential sources for a comprehensive view of the phenomena.
2 Spatial and seasonal endosymbiont prevalence variation correlates with temperature
The prevalence of facultative inheritable bacteria symbionts in natural insect populations varies in spatial gradients, latitudinal clines and over seasons (Corbin et al., 2017; Carpenter et al., 2021; Smith et al., 2021). Studies on 41 different natural insect populations show spatial or temporal symbiont frequency variation. In 21 of these cases, endosymbiont frequency was positively correlated with temperatures (Table 1). Alternatively, in only 3 of those 41 populations, endosymbiont frequency was negatively correlated with temperature (Table 1). In 17 cases, spatial or temporal symbiont frequency variation was observed, but with no observed correlation with temperature (Table 1). Even though we observed a general pattern of more cases of positive correlations between temperature and endosymbiont prevalence, there was great variation among studies. Some studies report seasonal variation with increases in endosymbiont prevalence in warmer seasons (Carpenter et al., 2021), while other studies report spatial variation with increases in endosymbiont prevalence in warmer regions with lower latitudes and lower altitudes (Zhang et al., 2021). One study predicted higher endosymbiont prevalence from land surface temperature data (Morag et al., 2012). A negative correlation between temperature and endosymbiont prevalence was observed in one study conducted in one site that exhibits extremely high temperatures, showing higher endosymbiont prevalence at milder temperatures (Watts et al., 2009). It is important to note that some studies tested the effect of temperature on the entire endosymbiont community infecting a host and symbiont hitchhiking and gut-microbiota is likely affecting some of the observed patterns (Zhang et al., 2021). As an example, S. symbiotica exhibits different patterns of infection in aphids across Belgium that could be related with temperature. However, S. symbiotica is found both as an endosymbiont and as a gut-associated bacteria, with plant resources also containing the bacteria, serving as a permanent environmental source of contamination (Pons et al., 2022).
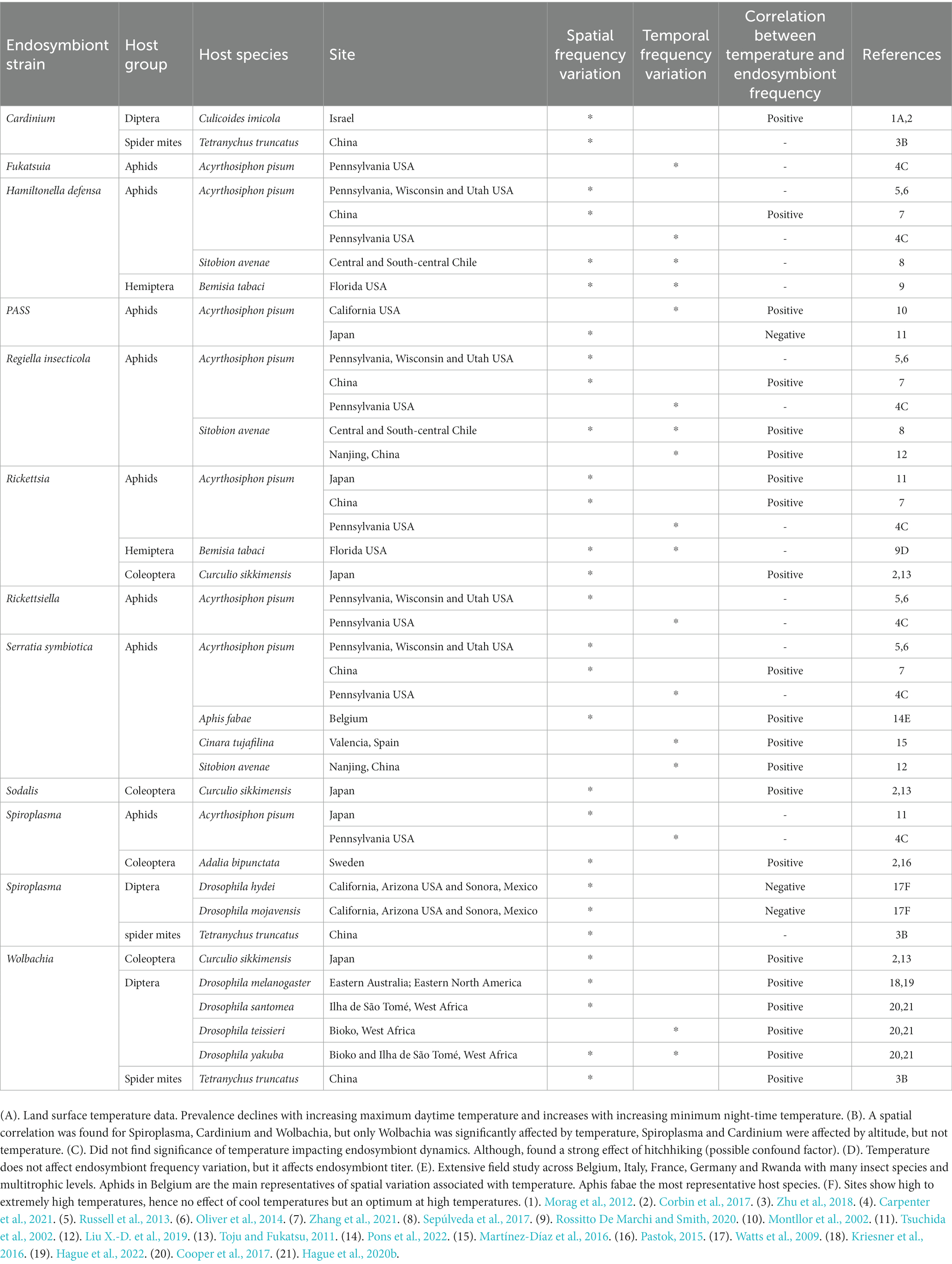
Table 1. Evidence of spatial and temporal frequency variation in facultative endosymbiont infections in different groups of insects and its correlation with temperature.
3 Temperature affects the efficiency of maternal transmission
Imperfect maternal transmission might be an important mechanism limiting the prevalence of inheritable symbionts (Oliver et al., 2014). We found 17 studies that showed that temperature affects maternal transmission, expanding studies found in Corbin et al. (2017). In eight studies transmission efficiency was negatively correlated with temperature (Table 2), in five studies transmission efficiency was positively correlated (Table 2), and in one study there was a non-linear correlation between temperature and transmission efficiency (Table 2). Hague et al. (2020b) point temperature as a fundamental determinant on the persistence of wMel-like Wolbachia on D. melanogaster showing that colder temperatures reduce maternal transmission with its effects on host oocytes providing a cellular basis for the wMel clines observed in Australia (Kriesner et al., 2016; Hague et al., 2022). For instance, both high and low temperature affects the vertical transmission of Spiroplasma on the native host Drosophila nebulosa (Anbutsu et al., 2008) and Drosophila hydei (Osaka et al., 2008). At lower temperatures the density of Spiroplasma was reduced as well. Low temperatures are known to cure Spiroplasma infections in D. melanogaster, whereas impeding higher temperatures might alter either their tissue tropism or motility, thereby resulting in less-efficient vertical transmission (Anbutsu et al., 2008). Liu X.-D. et al. (2019) showed failure in the rate of vertical transmission of R. insecticola in aphids at high temperatures. However, when high temperatures are maintained for several generations, the transmission rate of R. insecticola is increased by up to 100% (Liu X.-D. et al., 2019). Hence, temperature clearly affects the efficiency of maternal transmission, and this can explain some patterns of spatial variation in symbiont prevalence (Hague et al., 2020a).
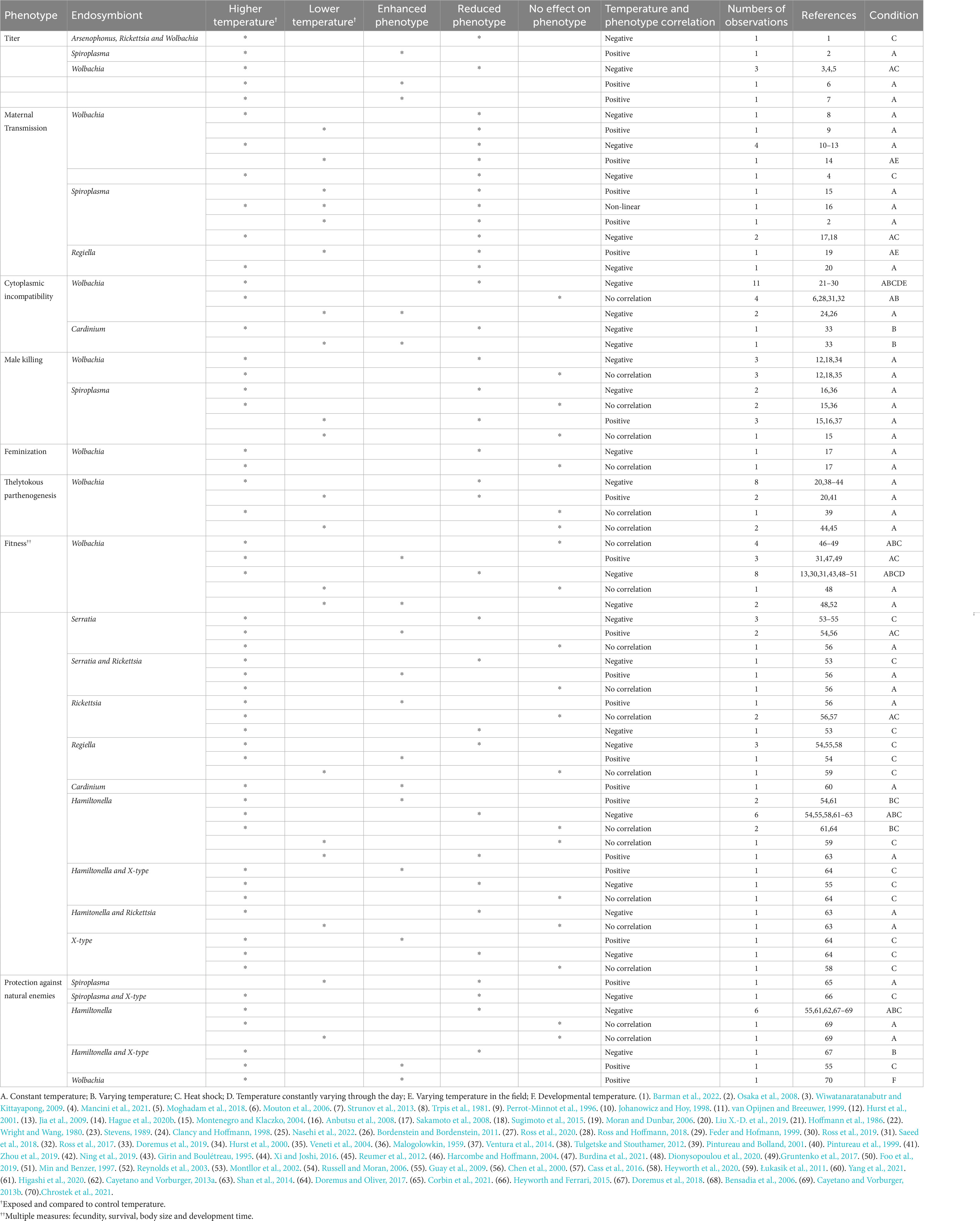
Table 2. Relationship between phenotype, temperature, and effects on fitness of common facultative endosymbionts.
4 Temperature affects reproductive parasitism phenotypes
Wolbachia, Cardinium and Spiroplasma can manipulate the reproduction of their hosts and temperature can affect the expression of these phenotypes (Corbin et al., 2017). Reproductive parasitism phenotypes include cytoplasmic incompatibility (CI), male killing (MK), feminization and thelytokous parthenogenesis (Werren et al., 2008). All these phenotypes increase the number of infected females in the population, either by modification of gametic compatibility and interference in sex determination as non fertilization and selective lethality to males (Hurst et al., 2000; Sakamoto et al., 2008; Sugimoto and Ishikawa, 2012; Liu Q.-Q. et al., 2019; Kaur et al., 2021). The effect of temperature on reproductive manipulation was reported in 46 cases (Table 2). In 28 cases the expression of the reproductive manipulation was stronger at lower temperatures (negative correlations in Table 2), in five, reproductive manipulation was stronger at higher temperature (positive correlations in Table 2) and 13 cases showed that the expression of reproductive manipulation was not affected by temperature.
Most studies show a pattern that at lower temperatures, Wolbachia and Cardinium can increase the expression of CI and at higher temperatures the expression of CI decreases or is completely suppressed (Table 2). As development time is slower under colder temperatures, endosymbionts might have enough time to induce full CI in the cells of their hosts, despite the effects on symbiont titer (Bordenstein and Bordenstein, 2011; Doremus et al., 2019). Regarding MK, feminization and thelytoky, at permissive temperatures the expression of these phenotypes is close to 100%. When hosts are exposed to both high or low temperatures, the expression of MK by Spiroplasma is greatly reduced (Malogolowkin, 1959; Montenegro and Klaczko, 2004; Anbutsu et al., 2008; Ventura et al., 2014) and exposure to extremely high temperatures reduces the expression of MK by Wolbachia (Hurst et al., 2000, 2001). Most studies show that both high and low temperatures reduce the expression of feminization and thelytoky by Wolbachia (Girin and Boulétreau, 1995; Pintureau et al., 1999; Ning et al., 2019; Zhou et al., 2019). Furthermore, the time and temperature required to affect phenotype penetrance varies among endosymbiont strains and host species (Pintureau et al., 1999; Corbin et al., 2017). This difference might occur because some strains have higher thermal stress tolerance that endosymbionts face according to their global distribution (Ross et al., 2017). Thus, temperature is an important environmental factor influencing the expression of reproductive parasitism phenotypes in nature.
5 Temperature affects endosymbiont influence on host fitness
The presence of symbionts usually affects many components of host fitness, such as fecundity, survival, body size, and development time. These effects are likely to be influenced by temperature. We compiled 57 studies testing the effect of temperature on the effects of symbionts on host fitness (Table 2). In 27 studies there was a negative correlation between temperature and host fitness, indicating that symbionts intensively decrease host fitness at higher temperatures. In 14 studies there was a positive correlation between host fitness and temperature and 16 studies showed no correlation (Table 2). Exposure to thermal stress can negatively affect fitness of insects (Harvey et al., 2020). Across studies that investigated fitness costs on insects associated with endosymbionts under thermal stress, there is a general pattern that higher temperatures causes lower fecundity, survival, body size, and development time (Table 2). Few studies investigated associated costs under low temperatures (Reynolds et al., 2003; Łukasik et al., 2011; Shan et al., 2014; Dionysopoulou et al., 2020). Shan et al. (2014) observed lower fitness associated with exposure to cold in aphids infected with Hamiltonella. Interestingly, in some cases, symbionts are capable of providing protection against heat stress, as observed in white flies infected with Cardinium (Yang et al., 2021), aphids infected with Fukatsuia (Heyworth et al., 2020), and flies infected with Wolbachia strain wMelPlus (Burdina et al., 2021).
6 Temperature affects endosymbiont protection against natural enemies
Several facultative endosymbionts protect its hosts against natural enemies and the outcome of the interactions can vary across different temperature ranges (Doremus and Oliver, 2017). Temperature effect on symbiont protection against natural enemies was reported in 11 studies (Table 2). In seven studies the protection phenotype was negatively correlated with temperature, in two, protection was positively correlated and the remaining two showed no correlation between protection and temperature (Table 2). Protection against parasitoids by Spiroplasma in flies ceases to happen at low temperatures (Corbin et al., 2021). Hamiltonella’s protection against parasitoid attacks in aphids is severely reduced under high temperature regimes (Bensadia et al., 2006; Guay et al., 2009; Cayetano and Vorburger, 2013a; Doremus et al., 2019; Higashi et al., 2020), but not under colder temperatures (Cayetano and Vorburger, 2013b). Interestingly, Fukatsuia (X-type) alone does not confer protection against natural enemies, but it is able to rescue Hamiltonella’s protection against parasitoids when aphids are exposed to high temperatures (Guay et al., 2009), but this rescue ability may vary between Fukatsuia strains (Doremus and Oliver, 2017). Spiroplasma titer is reduced under low temperatures, which can affect toxin production and lipid competition, both factors responsible for protection against parasitoids (Corbin et al., 2021). However, in Hamiltonella elevated temperatures do not reduce endosymbiont density in aphids. Wolbachia induced antiviral phenotype in Drosophila melanogaster depends on developmental temperature being strong at 25°C and abolished at 18°C (Chrostek et al., 2021). Therefore, although symbiont density can be a good indicator of protection against natural enemies for some host-endosymbiont-natural enemy interactions (Martinez et al., 2014; Corbin et al., 2021), its relevance in nature remains unknown (Cogni et al., 2021). The mechanisms behind microbe-mediated protection and its relation to changes in temperature range still need further investigation.
7 Temperature affects endosymbiont titer and modulates host physiological acclimation and behavior
From the endosymbiont perspective, temperature challenge may lead, in one extreme, to endosymbiont depletion and in the other to an overproliferation that harms its transmission route through host fitness. Since long-term endosymbiosis success depends on the interests of endosymbionts being aligned with those of their host (Werner et al., 2014; Fisher et al., 2017) titer regulation might be an important trait under selection (Duarte et al., 2021). Titer is often found positively correlated with temperature in experiments held at permissive and constant 18 to 29°C (Reynolds et al., 2003; Osaka et al., 2008; Strunov et al., 2013). However, contrasting patterns are found in experiments involving heat shocks. The white-fly Bemicia tabaci harbors several endosymbionts under the genera Rickettsia, Hamiltonella, and Wolbachia. All of them show reduced titer after exposition to higher temperatures, a pattern not displayed by the obligate endosymbiont Candidatus Portiera (Barman et al., 2022). Facultative symbionts were reported to mitigate the effect of heat shock on obligate symbionts, aiding the hosts’ survival (Lemoine et al., 2020) and thus ensuring endosymbiont route to offspring despite the reduced titer. In aphids, for example, heat shock increased the adaptive value of a symbiotic infection, implying that hosting a facultative symbiont was more beneficial under heat stress than under permissive temperatures (Tougeron and Iltis, 2022). This is represented by the endosymbionts’ ability to trigger stress-lowering compounds (e.g., heat shock proteins) that modify hosts thermal tolerance through diverse metabolic and physiological pathways (Burke et al., 2010; Brumin et al., 2011; Wernegreen, 2012; Barman et al., 2022; Iltis et al., 2022).
Deleterious effects of thermal stress on symbiont titers may be diminished via other mechanisms such as gene expression or modulation of the host behavior (Brumin et al., 2011; Truitt et al., 2018; Hague et al., 2020a; Barman et al., 2022). Divergent Wolbachia strains were reported to affect host preference to different microclimates. Drosophila melanogaster infected with A-group Wolbachia strains prefer significantly cooler temperatures relative to uninfected individuals (Hague et al., 2020a). On the other hand, Drosophila mauritiana infected with the B-group wMau causes hosts to prefer warmer temperatures. This experiment also shows that host temperature preference does not alter Wolbachia titer (Hague et al., 2020a). Therefore, coping with temperature challenge via the symbiont effect on host acclimation and behavior is an important feature evolving that can maintain facultative endosymbiont prevalence.
8 Discussion
Temperature promotes an everlasting influence on facultative endosymbionts. In spatial and temporal scales higher frequencies are correlated with higher temperatures, whereas, key phenotypes are found with more negative correlations with temperature. This leads to the following questions: How stable are the patterns of prevalence of facultative endosymbionts in field populations considering different temperature ranges? Are there divergent cost–benefit optima among phenotype intensity, fitness and titer at multiple temperatures? How does this shape and maintain the prevalence? To test the stability of the correlations presented here it is important to describe endosymbiont dynamics with long term resampling (Cogni et al., 2017; Rodrigues and Cogni, 2021) and sampling repeated patterns of correspondent temperature correlations such as clinal and seasonal correspondent variation (Cogni et al., 2015; Rodrigues et al., 2021).
Following the evolutionary model, temperature indeed affects the processes that might explain the prevalence patterns, by acting in the infection itself and directly regulating endosymbiont titer, or by affecting the cost–benefit associated with the infection to the host and regulating the intensity of many endosymbiont induced phenotypes. However, key phenotypes such as maternal transmission, reproductive parasitism, host fitness and protection against natural enemies all show more negative correlations with temperature opposed to the prevalence pattern found in spatial-seasonal scales. This is striking, once the evolutionary model points to intensity of the key phenotypes as an essential mechanism for endosymbionts to reach higher frequencies. This shows that the processes on endosymbiont prevalence and success in nature is indeed complex and need further investigation. Temperature is a strong contender but it did not enact in a predictive way. In some systems, such as maternal transmission intensity modulated by temperature explaining prevalence patterns (Hague et al., 2020a) can be bolstered. However, a generalization of the process in the diversity of endosymbiont insect systems is currently not possible. Thus, it is important to delineate studies that investigate titer and divergent cost–benefit optima among phenotype intensity at multiple temperatures (Reynolds et al., 2003; López-Madrigal and Duarte, 2019; Duarte et al., 2021); as investigate measurements of the reaction norm, rather than only a snapshot of the phenomena with just one set of conditions (Thomas and Blanford, 2003).
In an evolutionary context, it’s harder to demonstrate that laboratory-observed patterns can lead to processes such as frequency-dependent selection in the field (Thomas and Blanford, 2003), but we cannot discard this possibility. Therefore, some important caveats here is that almost all studies on the effect of temperature were set in the laboratory. Protection against natural enemies are frequently reported, but correlation with temperature and this phenotype in the field is currently unknown (Cogni et al., 2021). Another caveat is given to the origin of the populations considered in each study, as a lab-adapted insect population will certainly show different responses to the processes than a wild one. Although many studies reported the conditioning set in their experiments (described in Table 2), the depiction of the events of each considered lab-adapted population is uncertain. If populations were previously acclimated in higher or lower temperatures or were exposed to heat or cold shocks that will assuredly impact on the coevolution processes. An integrative approach combining field and laboratory studies, exploring each system’s field prevalence and laboratory studies including cage experiments exploring temperature variation and realistic upper and lower extremes, as well as constant temperatures is essential. To add another layer, the use of transcriptomics and genomics may help to elucidate possible endosymbiont molecular candidates that interact with hosts that are differentiable between different temperatures, hence directing towards a mechanistic understanding of the patterns.
Author contributions
MM: Writing – original draft, Writing – review & editing. CC: Writing – original draft, Writing – review & editing. RC: Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Funding for this work was provided by São Paulo Research Foundation (FAPESP) (2019/03997-2; 2021/13166-0; and 2021/06874-9), the National Council for Scientific and Technological Development (CNPq) (309468/2021-3), and the Foundation Coordination for the Improvement of Higher Education Personnel (CAPES) (88887.637427/2021-00).
Acknowledgments
We thank André Pimentel and Vitória Miranda for suggestions on the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Anbutsu, H., Goto, S., and Fukatsu, T. (2008). High and low temperatures differently affect infection density and vertical transmission of male-killing Spiroplasma symbionts in Drosophila hosts. Appl. Environ. Microbiol. 74, 6053–6059. doi: 10.1128/AEM.01503-08
Barman, M., Samanta, S., Ahmed, B., Dey, S., Chakraborty, S., Deeksha, M. G., et al. (2022). Transcription dynamics of heat-shock proteins (Hsps) and endosymbiont titres in response to thermal stress in whitefly, Bemisia tabaci (Asia-I). Front. Physiol. 13:1097459. doi: 10.3389/fphys.2022.1097459
Bensadia, F., Boudreault, S., Guay, J.-F., Michaud, D., and Cloutier, C. (2006). Aphid clonal resistance to a parasitoid fails under heat stress. J. Insect Physiol. 52, 146–157. doi: 10.1016/j.jinsphys.2005.09.011
Bordenstein, S. R., and Bordenstein, S. R. (2011). Temperature affects the tripartite interactions between bacteriophage WO, Wolbachia, and cytoplasmic incompatibility. PLoS One 6:e29106. doi: 10.1371/journal.pone.0029106
Brumin, M., Kontsedalov, S., and Ghanim, M. (2011). Rickettsia influences thermotolerance in the whitefly Bemisia tabaci B biotype. Insect Sci. 18, 57–66. doi: 10.1111/j.1744-7917.2010.01396.x
Burdina, E. V., Bykov, R. A., Menshanov, P. N., Ilinsky, Y. Y., and Gruntenko, N. Е. (2021). Unique Wolbachia strain wMelPlus increases heat stress resistance in Drosophila melanogaster. Arch. Insect Biochem. Physiol. 106:e21776. doi: 10.1002/arch.21776
Burke, G., Fiehn, O., and Moran, N. (2010). Effects of facultative symbionts and heat stress on the metabolome of pea aphids. ISME J. 4, 242–252. doi: 10.1038/ismej.2009.114
Carpenter, M., Peng, L., Smith, A. H., Joffe, J., O’Connor, M., Oliver, K. M., et al. (2021). Frequent drivers, occasional passengers: signals of symbiont-driven seasonal adaptation and hitchhiking in the pea aphid, Acyrthosiphon pisum. Insects 12:805. doi: 10.3390/insects12090805
Cass, B. N., Himler, A. G., Bondy, E. C., Bergen, J. E., Fung, S. K., Kelly, S. E., et al. (2016). Conditional fitness benefits of the Rickettsia bacterial symbiont in an insect pest. Oecologia 180, 169–179. doi: 10.1007/s00442-015-3436-x
Cayetano, L., and Vorburger, C. (2013a). Effects of heat shock on resistance to parasitoids and on life history traits in an aphid/endosymbiont system. PLoS One 8:e75966. doi: 10.1371/journal.pone.0075966
Cayetano, L., and Vorburger, C. (2013b). Genotype-by-genotype specificity remains robust to average temperature variation in an aphid/endosymbiont/parasitoid system. J. Evol. Biol. 26, 1603–1610. doi: 10.1111/jeb.12154
Chen, D.-Q., Montllor, C. B., and Purcell, A. H. (2000). Fitness effects of two facultative endosymbiotic bacteria on the pea aphid, Acyrthosiphon pisum, and the blue alfalfa aphid, A. kondoi. Entomol. Exp. Appl. 95, 315–323. doi: 10.1046/j.1570-7458.2000.00670.x
Chrostek, E., Martins, N., Marialva, M. S., and Teixeira, L. (2021). Wolbachia-conferred antiviral protection is determined by developmental temperature. MBio 12:e0292320. doi: 10.1128/mBio.02923-20
Clancy, D. J., and Hoffmann, A. A. (1998). Environmental effects on cytoplasmic incompatibility and bacterial load in Wolbachia-infected Drosophila simulans. Entomol. Exp. Appl. 86, 13–24. doi: 10.1046/j.1570-7458.1998.00261.x
Cogni, R., Ding, S. D., Pimentel, A. C., Day, J. P., and Jiggins, F. M. (2021). Wolbachia reduces virus infection in a natural population of Drosophila. Commun. Biol. 4:1327. doi: 10.1038/s42003-021-02838-z
Cogni, R., Kuczynski, K., Koury, S., Lavington, E., Behrman, E. L., O’Brien, K. R., et al. (2017). On the long-term stability of clines in some metabolic genes in Drosophila melanogaster. Sci. Rep. 7:42766. doi: 10.1038/srep42766
Cogni, R., Kuczynski, K., Lavington, E., Koury, S., Behrman, E. L., O’Brien, K. R., et al. (2015). Variation in Drosophila melanogaster central metabolic genes appears driven by natural selection both within and between populations. Proc. Biol. Sci. 282:20142688. doi: 10.1098/rspb.2014.2688
Cooper, B. S., Ginsberg, P. S., Turelli, M., and Matute, D. R. (2017). Wolbachia in the Drosophila yakuba complex: pervasive frequency variation and weak cytoplasmic incompatibility, but no apparent effect on reproductive isolation. Genetics 205, 333–351. doi: 10.1534/genetics.116.196238
Corbin, C., Heyworth, E. R., Ferrari, J., and Hurst, G. D. D. (2017). Heritable symbionts in a world of varying temperature. Heredity 118, 10–20. doi: 10.1038/hdy.2016.71
Corbin, C., Jones, J. E., Chrostek, E., Fenton, A., and Hurst, G. D. D. (2021). Thermal sensitivity of the Spiroplasma-Drosophila hydei protective symbiosis: the best of climes, the worst of climes. Mol. Ecol. 30, 1336–1344. doi: 10.1111/mec.15799
Dionysopoulou, N. K., Papanastasiou, S. A., Kyritsis, G. A., and Papadopoulos, N. T. (2020). Effect of host fruit, temperature and Wolbachia infection on survival and development of Ceratitis capitata immature stages. PLoS One 15:e0229727. doi: 10.1371/journal.pone.0229727
Doremus, M. R., Kelly, S. E., and Hunter, M. S. (2019). Exposure to opposing temperature extremes causes comparable effects on Cardinium density but contrasting effects on Cardinium-induced cytoplasmic incompatibility. PLoS Pathog. 15:e1008022. doi: 10.1371/journal.ppat.1008022
Doremus, M. R., and Oliver, K. M. (2017). Aphid heritable symbiont exploits defensive mutualism. Appl. Environ. Microbiol. 83. doi: 10.1128/AEM.03276-16
Doremus, M. R., Smith, A. H., Kim, K. L., Holder, A. J., Russell, J. A., and Oliver, K. M. (2018). Breakdown of a defensive symbiosis, but not endogenous defences, at elevated temperatures. Mol. Ecol. 27, 2138–2151. doi: 10.1111/mec.14399
Duarte, E. H., Carvalho, A., López-Madrigal, S., Costa, J., and Teixeira, L. (2021). Forward genetics in Wolbachia: regulation of Wolbachia proliferation by the amplification and deletion of an addictive genomic island. PLoS Genet. 17:e1009612. doi: 10.1371/journal.pgen.1009612
Feder, M. E., and Hofmann, G. E. (1999). Heat-shock proteins, molecular chaperones, and the stress response: evolutionary and ecological physiology. Annu. Rev. Physiol. 61, 243–282. doi: 10.1146/annurev.physiol.61.1.243
Fisher, R. M., Henry, L. M., Cornwallis, C. K., Kiers, E. T., and West, S. A. (2017). The evolution of host-symbiont dependence. Nat. Commun. 8:15973. doi: 10.1038/ncomms15973
Foo, I. J.-H., Hoffmann, A. A., and Ross, P. A. (2019). Cross-generational effects of heat stress on fitness and Wolbachia density in Aedes aegypti mosquitoes. Trop. Med. Infect. Dis. 4:13. doi: 10.3390/tropicalmed4010013
Gavotte, L., Mercer, D. R., Stoeckle, J. J., and Dobson, S. L. (2010). Costs and benefits of Wolbachia infection in immature Aedes albopictus depend upon sex and competition level. J. Invertebr. Pathol. 105, 341–346. doi: 10.1016/j.jip.2010.08.005
Girin, C., and Boulétreau, M. (1995). Microorganism-associated variation in host infestation efficiency in a parasitoid wasp, Trichogramma bourarachae (Hymenoptera: Trichogrammatidae). Experientia 51, 398–401. doi: 10.1007/BF01928904
Gruntenko, N. Е., Ilinsky, Y. Y., Adonyeva, N. V., Burdina, E. V., Bykov, R. A., Menshanov, P. N., et al. (2017). Various Wolbachia genotypes differently influence host Drosophila dopamine metabolism and survival under heat stress conditions. BMC Evol. Biol. 17:252. doi: 10.1186/s12862-017-1104-y
Guay, J.-F., Boudreault, S., Michaud, D., and Cloutier, C. (2009). Impact of environmental stress on aphid clonal resistance to parasitoids: role of Hamiltonella defensa bacterial symbiosis in association with a new facultative symbiont of the pea aphid. J. Insect Physiol. 55, 919–926. doi: 10.1016/j.jinsphys.2009.06.006
Hague, M. T. J., Caldwell, C. N., and Cooper, B. S. (2020a). Pervasive effects of Wolbachia on host temperature preference. MBio 11, 1–15. doi: 10.1128/mBio.01768-20
Hague, M. T. J., Mavengere, H., Matute, D. R., and Cooper, B. S. (2020b). Environmental and genetic contributions to imperfect wMel-like Wolbachia transmission and frequency variation. Genetics 215, 1117–1132. doi: 10.1534/genetics.120.303330
Hague, M. T. J., Shropshire, J. D., Caldwell, C. N., Statz, J. P., Stanek, K. A., Conner, W. R., et al. (2022). Temperature effects on cellular host-microbe interactions explain continent-wide endosymbiont prevalence. Curr. Biol. 32, 878–888.e8. doi: 10.1016/j.cub.2021.11.065
Harcombe, W., and Hoffmann, A. A. (2004). Wolbachia effects in Drosophila melanogaster: in search of fitness benefits. J. Invertebr. Pathol. 87, 45–50. doi: 10.1016/j.jip.2004.07.003
Harvey, J. A., Heinen, R., Gols, R., and Thakur, M. P. (2020). Climate change-mediated temperature extremes and insects: from outbreaks to breakdowns. Glob. Chang. Biol. 26, 6685–6701. doi: 10.1111/gcb.15377
Heyworth, E. R., and Ferrari, J. (2015). A facultative endosymbiont in aphids can provide diverse ecological benefits. J. Evol. Biol. 28, 1753–1760. doi: 10.1111/jeb.12705
Heyworth, E. R., Smee, M. R., and Ferrari, J. (2020). Aphid facultative symbionts aid recovery of their obligate symbiont and their host after heat stress. Front. Ecol. Evol. 8:56. doi: 10.3389/fevo.2020.00056
Higashi, C. H. V., Barton, B. T., and Oliver, K. M. (2020). Warmer nights offer no respite for a defensive mutualism. J. Anim. Ecol. 89, 1895–1905. doi: 10.1111/1365-2656.13238
Hoffmann, A. A., Turelli, M., and Harshman, L. G. (1990). Factors affecting the distribution of cytoplasmic incompatibility in Drosophila simulans. Genetics 126, 933–948. doi: 10.1093/genetics/126.4.933
Hoffmann, A. A., Turelli, M., and Simmons, G. M. (1986). Unidirectional incompatibility between populations of Drosophila simulans. Evolution 40, 692–701. doi: 10.2307/2408456
Hurst, G. D., Jiggins, F. M., and Robinson, S. J. (2001). What causes inefficient transmission of male-killing Wolbachia in Drosophila? Heredity 87, 220–226. doi: 10.1046/j.1365-2540.2001.00917.x
Hurst, G. D., Johnson, A. P., Schulenburg, J. H., and Fuyama, Y. (2000). Male-killing Wolbachia in Drosophila: a temperature-sensitive trait with a threshold bacterial density. Genetics 156, 699–709. doi: 10.1093/genetics/156.2.699
Iltis, C., Tougeron, K., Hance, T., Louâpre, P., and Foray, V. (2022). A perspective on insect-microbe holobionts facing thermal fluctuations in a climate-change context. Environ. Microbiol. 24, 18–29. doi: 10.1111/1462-2920.15826
Jia, F.-X., Yang, M.-S., Yang, W.-J., and Wang, J.-J. (2009). Influence of continuous high temperature conditions on Wolbachia infection frequency and the fitness of Liposcelis tricolor (Psocoptera: Liposcelididae). Environ. Entomol. 38, 1365–1372. doi: 10.1603/022.038.0503
Johanowicz, D. L., and Hoy, M. A. (1998). Experimental induction and termination of non-reciprocal reproductive incompatibilities in a parahaploid mite. Entomol. Exp. Appl. 87, 51–58. doi: 10.1046/j.1570-7458.1998.00303.x
Kaur, R., Shropshire, J. D., Cross, K. L., Leigh, B., Mansueto, A. J., Stewart, V., et al. (2021). Living in the endosymbiotic world of Wolbachia: a centennial review. Cell Host Microbe 29, 879–893. doi: 10.1016/j.chom.2021.03.006
Kriesner, P., Conner, W. R., Weeks, A. R., Turelli, M., and Hoffmann, A. A. (2016). Persistence of a Wolbachia infection frequency cline in Drosophila melanogaster and the possible role of reproductive dormancy. Evolution 70, 979–997. doi: 10.1111/evo.12923
Leclair, M., Buchard, C., Mahéo, F., Simon, J.-C., and Outreman, Y. (2021). A link between communities of protective endosymbionts and parasitoids of the pea aphid revealed in unmanipulated agricultural systems. Front. Ecol. Evol. 9:187. doi: 10.3389/fevo.2021.618331
Lemoine, M. M., Engl, T., and Kaltenpoth, M. (2020). Microbial symbionts expanding or constraining abiotic niche space in insects. Curr. Opin. Insect. Sci. 39, 14–20. doi: 10.1016/j.cois.2020.01.003
Liu, X.-D., Lei, H.-X., and Chen, F.-F. (2019). Infection pattern and negative effects of a facultative endosymbiont on its insect host are environment-dependent. Sci. Rep. 9:4013. doi: 10.1038/s41598-019-40607-5
Liu, Q.-Q., Zhou, J.-C., Zhang, C., Ning, S.-F., Duan, L.-J., and Dong, H. (2019). Co-occurrence of thelytokous and bisexual Trichogramma dendrolimi Matsumura (Hymenoptera: Trichogrammatidae) in a natural population. Sci. Rep. 9:17480. doi: 10.1038/s41598-019-53992-8
López-Madrigal, S., and Duarte, E. H. (2019). Titer regulation in arthropod-Wolbachia symbioses. FEMS Microbiol. Lett. 366. doi: 10.1093/femsle/fnz232
Łukasik, P., Hancock, E. L., Ferrari, J., and Godfray, H. C. J. (2011). Grain aphid clones vary in frost resistance, but this trait is not influenced by facultative endosymbionts. Ecol. Entomol. 36, 790–793. doi: 10.1111/j.1365-2311.2011.01321.x
Malogolowkin, C. (1959). Temperature effects on maternally inherited “sex-ratio” conditions in Drosophila willistoni and Drosophila equinoxialis. Am. Nat. 93, 365–368. doi: 10.1086/282096
Mancini, M. V., Ant, T. H., Herd, C. S., Martinez, J., Murdochy, S. M., Gingell, D. D., et al. (2021). High temperature cycles result in maternal transmission and dengue infection differences between Wolbachia strains in Aedes aegypti. MBio 12:e0025021. doi: 10.1128/mBio.00250-21
Martinez, J., Longdon, B., Bauer, S., Chan, Y.-S., Miller, W. J., Bourtzis, K., et al. (2014). Symbionts commonly provide broad spectrum resistance to viruses in insects: a comparative analysis of Wolbachia strains. PLoS Pathog. 10:e1004369. doi: 10.1371/journal.ppat.1004369
Martínez-Díaz, V., Latorre, A., and Gil, R. (2016). Seasonal changes in the endosymbiotic consortia of aphids from the genus Cinara. Microbes Environ. 31, 137–144. doi: 10.1264/jsme2.ME15118
Min, K. T., and Benzer, S. (1997). Wolbachia, normally a symbiont of Drosophila, can be virulent, causing degeneration and early death. Proc. Natl. Acad. Sci. U. S. A. 94, 10792–10796. doi: 10.1073/pnas.94.20.10792
Moghadam, N. N., Thorshauge, P. M., Kristensen, T. N., de Jonge, N., Bahrndorff, S., Kjeldal, H., et al. (2018). Strong responses of Drosophila melanogaster microbiota to developmental temperature. Fly 12, 1–12. doi: 10.1080/19336934.2017.1394558
Montenegro, H., and Klaczko, L. B. (2004). Low temperature cure of a male killing agent in Drosophila melanogaster. J. Invertebr. Pathol. 86, 50–51. doi: 10.1016/j.jip.2004.03.004
Montllor, C. B., Maxmen, A., and Purcell, A. H. (2002). Facultative bacterial endosymbionts benefit pea aphids Acyrthosiphon pisum under heat stress. Ecol. Entomol. 27, 189–195. doi: 10.1046/j.1365-2311.2002.00393.x
Morag, N., Klement, E., Saroya, Y., Lensky, I., and Gottlieb, Y. (2012). Prevalence of the symbiont Cardinium in Culicoides (Diptera: Ceratopogonidae) vector species is associated with land surface temperature. FASEB J. 26, 4025–4034. doi: 10.1096/fj.12-210419
Moran, N. A., and Dunbar, H. E. (2006). Sexual acquisition of beneficial symbionts in aphids. Proc. Natl. Acad. Sci. U. S. A. 103, 12803–12806. doi: 10.1073/pnas.0605772103
Mouton, L., Henri, H., Bouletreau, M., and Vavre, F. (2006). Effect of temperature on Wolbachia density and impact on cytoplasmic incompatibility. Parasitology 132, 49–56. doi: 10.1017/S0031182005008723
Nasehi, S. F., Fathipour, Y., Asgari, S., and Mehrabadi, M. (2022). Environmental temperature, but not male age, affects Wolbachia and prophage WO thereby modulating cytoplasmic incompatibility in the parasitoid wasp, Habrobracon Hebetor. Microb. Ecol. 83, 482–491. doi: 10.1007/s00248-021-01768-x
Ning, S.-F., Zhou, J.-C., Liu, Q.-Q., Zhao, Q., and Dong, H. (2019). Gradual, temperature-induced change of secondary sexual characteristics in Trichogramma pretiosum infected with parthenogenesis-inducing Wolbachia. PeerJ 7:e7567. doi: 10.7717/peerj.7567
Oliver, K. M., Smith, A. H., and Russell, J. A. (2014). Defensive symbiosis in the real world - advancing ecological studies of heritable, protective bacteria in aphids and beyond. Funct. Ecol. 28, 341–355. doi: 10.1111/1365-2435.12133
Osaka, R., Nomura, M., Watada, M., and Kageyama, D. (2008). Negative effects of low temperatures on the vertical transmission and infection density of a spiroplasma endosymbiont in Drosophila hydei. Curr. Microbiol. 57, 335–339. doi: 10.1007/s00284-008-9199-4
Perrot-Minnot, M. J., Guo, L. R., and Werren, J. H. (1996). Single and double infections with Wolbachia in the parasitic wasp Nasonia vitripennis: effects on compatibility. Genetics 143, 961–972. doi: 10.1093/genetics/143.2.961
Pintureau, B., and Bolland, P. (2001). A Trichogramma species showing a better adaptation to high temperature than its symbionts. Biocontrol Sci. Tech. 11, 13–20. doi: 10.1080/09583150020029691
Pintureau, B., Chapelle, L., and Delobel, B. (1999). Effects of repeated thermic and antibiotic treatments on a Trichogramma (Hym., Trichogrammatidae) symbiont. J. Appl. Entomol. 123, 473–483. doi: 10.1046/j.1439-0418.1999.00412.x
Pons, I., Scieur, N., Dhondt, L., Renard, M.-E., Renoz, F., and Hance, T. (2022). Pervasiveness of the symbiont Serratia symbiotica in the aphid natural environment: distribution, diversity and evolution at a multitrophic level. FEMS Microbiol. Ecol. 98. doi: 10.1093/femsec/fiac012
Renoz, F., Pons, I., and Hance, T. (2019). Evolutionary responses of mutualistic insect-bacterial symbioses in a world of fluctuating temperatures. Curr. Opin. Insect Sci. 35, 20–26. doi: 10.1016/j.cois.2019.06.006
Reumer, B. M., van Alphen, J. J. M., and Kraaijeveld, K. (2012). Occasional males in parthenogenetic populations of Asobara japonica (Hymenoptera: Braconidae): low Wolbachia titer or incomplete coadaptation? Heredity 108, 341–346. doi: 10.1038/hdy.2011.82
Reynolds, K. T., Thomson, L. J., and Hoffmann, A. A. (2003). The effects of host age, host nuclear background and temperature on phenotypic effects of the virulent Wolbachia strain popcorn in Drosophila melanogaster. Genetics 164, 1027–1034. doi: 10.1093/genetics/164.3.1027
Rodrigues, M. F., and Cogni, R. (2021). Genomic responses to climate change: making the Most of the Drosophila model. Front. Genet. 12:676218. doi: 10.3389/fgene.2021.676218
Rodrigues, M. F., Vibranovski, M. D., and Cogni, R. (2021). Clinal and seasonal changes are correlated in Drosophila melanogaster natural populations. Evolution 75, 2042–2054. doi: 10.1111/evo.14300
Ross, P. A., Axford, J. K., Yang, Q., Staunton, K. M., Ritchie, S. A., Richardson, K. M., et al. (2020). Heatwaves cause fluctuations in wMel Wolbachia densities and frequencies in Aedes aegypti. PLoS Negl. Trop. Dis. 14:e0007958. doi: 10.1371/journal.pntd.0007958
Ross, P. A., and Hoffmann, A. A. (2018). Continued susceptibility of the wMel Wolbachia infection in Aedes aegypti to heat stress following field deployment and selection. Insects 9:78. doi: 10.3390/insects9030078
Ross, P. A., Ritchie, S. A., Axford, J. K., and Hoffmann, A. A. (2019). Loss of cytoplasmic incompatibility in Wolbachia-infected Aedes aegypti under field conditions. PLoS Negl. Trop. Dis. 13:e0007357. doi: 10.1371/journal.pntd.0007357
Ross, P. A., Wiwatanaratanabutr, I., Axford, J. K., White, V. L., Endersby-Harshman, N. M., and Hoffmann, A. A. (2017). Wolbachia infections in Aedes aegypti differ markedly in their response to cyclical heat stress. PLoS Pathog. 13:e1006006. doi: 10.1371/journal.ppat.1006006
Rossitto De Marchi, B., and Smith, H. A. (2020). Bacterial endosymbiont diversity among Bemisia tabaci (Hemiptera: Aleyrodidae) populations in Florida. Insects 11:179. doi: 10.3390/insects11030179
Russell, J. A., and Moran, N. A. (2006). Costs and benefits of symbiont infection in aphids: variation among symbionts and across temperatures. Proc. Biol. Sci. 273, 603–610. doi: 10.1098/rspb.2005.3348
Russell, J. A., Weldon, S., Smith, A. H., Kim, K. L., Hu, Y., Łukasik, P., et al. (2013). Uncovering symbiont-driven genetic diversity across north American pea aphids. Mol. Ecol. 22, 2045–2059. doi: 10.1111/mec.12211
Saeed, N., Battisti, A., Martinez-Sanudo, I., and Mori, N. (2018). Combined effect of temperature and Wolbachia infection on the fitness of Drosophila suzukii. Bull. Insectology 71, 161–169.
Sakamoto, H., Kageyama, D., Hoshizaki, S., and Ishikawa, Y. (2008). Heat treatment of the adzuki bean borer, Ostrinia scapulalis infected with wolbachia gives rise to sexually mosaic offspring. J. Insect Sci. 8, 1–5. doi: 10.1673/031.008.6701
Sepúlveda, D. A., Zepeda-Paulo, F., Ramírez, C. C., Lavandero, B., and Figueroa, C. C. (2017). Diversity, frequency, and geographic distribution of facultative bacterial endosymbionts in introduced aphid pests. Insect Sci. 24, 511–521. doi: 10.1111/1744-7917.12313
Shan, H.-W., Lu, Y.-H., Bing, X.-L., Liu, S.-S., and Liu, Y.-Q. (2014). Differential responses of the whitefly Bemisia tabaci symbionts to unfavorable low and high temperatures. Microb. Ecol. 68, 472–482. doi: 10.1007/s00248-014-0424-3
Smith, A. H., O’Connor, M. P., Deal, B., Kotzer, C., Lee, A., Wagner, B., et al. (2021). Does getting defensive get you anywhere?-seasonal balancing selection, temperature, and parasitoids shape real-world, protective endosymbiont dynamics in the pea aphid. Mol. Ecol. 30, 2449–2472. doi: 10.1111/mec.15906
Stevens, L. (1989). Environmental factors affecting reproductive incompatibility in flour beetles, genus Tribolium. J. Invertebr. Pathol. 53, 78–84. doi: 10.1016/0022-2011(89)90076-1
Strunov, A., Kiseleva, E., and Gottlieb, Y. (2013). Spatial and temporal distribution of pathogenic Wolbachia strain wMelPop in Drosophila melanogaster central nervous system under different temperature conditions. J. Invertebr. Pathol. 114, 22–30. doi: 10.1016/j.jip.2013.05.001
Sugimoto, T. N., and Ishikawa, Y. (2012). A male-killing Wolbachia carries a feminizing factor and is associated with degradation of the sex-determining system of its host. Biol. Lett. 8, 412–415. doi: 10.1098/rsbl.2011.1114
Sugimoto, T. N., Kayukawa, T., Matsuo, T., Tsuchida, T., and Ishikawa, Y. (2015). A short, high-temperature treatment of host larvae to analyze Wolbachia-host interactions in the moth Ostrinia scapulalis. J. Insect Physiol. 81, 48–51. doi: 10.1016/j.jinsphys.2015.06.016
Thomas, M. B., and Blanford, S. (2003). Thermal biology in insect-parasite interactions. Trends Ecol. Evol. 18, 344–350. doi: 10.1016/S0169-5347(03)00069-7
Toju, H., and Fukatsu, T. (2011). Diversity and infection prevalence of endosymbionts in natural populations of the chestnut weevil: relevance of local climate and host plants. Mol. Ecol. 20, 853–868. doi: 10.1111/j.1365-294X.2010.04980.x
Tougeron, K., and Iltis, C. (2022). Impact of heat stress on the fitness outcomes of symbiotic infection in aphids: a meta-analysis. Proc. Biol. Sci. 289:20212660. doi: 10.1098/rspb.2021.2660
Trpis, M., Perrone, J. B., Reissig, M., and Parker, K. L. (1981). Control of cytoplasmic incompatibility in the Aedes scutellaris complexIncompatible crosses become compatible by treatment of larvae with heat or antibiotics. J. Hered. 72, 313–317. doi: 10.1093/oxfordjournals.jhered.a109513
Truitt, A. M., Kapun, M., Kaur, R., and Miller, W. J. (2018). Wolbachia modifies thermal preference in Drosophila melanogaster. Environ. Microbiol. 21, 3259–3268. doi: 10.1111/1462-2920.14347
Tsuchida, T., Koga, R., Shibao, H., Matsumoto, T., and Fukatsu, T. (2002). Diversity and geographic distribution of secondary endosymbiotic bacteria in natural populations of the pea aphid, Acyrthosiphon pisum. Mol. Ecol. 11, 2123–2135. doi: 10.1046/j.1365-294X.2002.01606.x
Tulgetske, G. M., and Stouthamer, R. (2012). Characterization of intersex production in Trichogramma kaykai infected with parthenogenesis-inducing Wolbachia. Naturwissenschaften 99, 143–152. doi: 10.1007/s00114-011-0880-2
van Opijnen, T., and Breeuwer, J. A. (1999). High temperatures eliminate Wolbachia, a cytoplasmic incompatibility inducing endosymbiont, from the two-spotted spider mite. Exp. Appl. Acarol. 23, 871–881. doi: 10.1023/A:1006363604916
Veneti, Z., Toda, M. J., and Hurst, G. D. D. (2004). Host resistance does not explain variation in incidence of male-killing bacteria in Drosophila bifasciata. BMC Evol. Biol. 4:52. doi: 10.1186/1471-2148-4-52
Ventura, I. M., Costa, T., and Klaczko, L. B. (2014). Low temperature reveals genetic variability against male-killing Spiroplasma in Drosophila melanogaster natural populations. Microb. Ecol. 67, 229–235. doi: 10.1007/s00248-013-0295-z
Watts, T., Haselkorn, T. S., Moran, N. A., and Markow, T. A. (2009). Variable incidence of Spiroplasma infections in natural populations of Drosophila species. PLoS One 4:e5703. doi: 10.1371/journal.pone.0005703
Wernegreen, J. J. (2012). Mutualism meltdown in insects: bacteria constrain thermal adaptation. Curr. Opin. Microbiol. 15, 255–262. doi: 10.1016/j.mib.2012.02.001
Werner, G. D. A., Strassmann, J. E., Ivens, A. B. F., Engelmoer, D. J. P., Verbruggen, E., Queller, D. C., et al. (2014). Evolution of microbial markets. Proc. Natl. Acad. Sci. U. S. A. 111, 1237–1244. doi: 10.1073/pnas.1315980111
Werren, J. H., Baldo, L., and Clark, M. E. (2008). Wolbachia: master manipulators of invertebrate biology. Nat. Rev. Microbiol. 6, 741–751. doi: 10.1038/nrmicro1969
Wiwatanaratanabutr, I., and Kittayapong, P. (2009). Effects of crowding and temperature on Wolbachia infection density among life cycle stages of Aedes albopictus. J. Invertebr. Pathol. 102, 220–224. doi: 10.1016/j.jip.2009.08.009
Wright, J. D., and Wang, B.-T. (1980). Observations on wolbachiae in mosquitoes. J. Invertebr. Pathol. 35, 200–208. doi: 10.1016/0022-2011(80)90185-8
Xi, Z., and Joshi, D. (2016). “Chapter 14 - genetic control of malaria and dengue using Wolbachia” in Genetic control of malaria and dengue. ed. Z. N. Adelman (Boston: Academic Press), 305–333.
Yang, K., Yuan, M.-Y., Liu, Y., Guo, C.-L., Liu, T.-X., Zhang, Y.-J., et al. (2021). First evidence for thermal tolerance benefits of the bacterial symbiont Cardinium in an invasive whitefly, Bemisia tabaci. Pest Manag. Sci. 77, 5021–5031. doi: 10.1002/ps.6543
Zhang, L., Cao, Y., Zhang, L.-J., Wang, M.-Y., Wang, X.-T., Yang, X.-Z., et al. (2021). Diversity and geographic variation of endosymbiotic bacteria in natural populations of the pea aphid (Acyrthosiphon pisum) in China. J. Asia Pac. Entomol. 24, 788–797. doi: 10.1016/j.aspen.2021.07.001
Zhou, J.-C., Li, Y.-Y., Liu, Q.-Q., Ning, S.-F., Che, W.-N., Cong, B., et al. (2019). Effects of temperature and superparasitism on quality and characteristics of thelytokous Wolbachia-infected Trichogramma dendrolimi Matsumura (Hymenoptera: Trichogrammatidae) during mass rearing. Sci. Rep. 9:18114. doi: 10.1038/s41598-019-54719-5
Zhu, Y.-X., Song, Y.-L., Zhang, Y.-K., Hoffmann, A. A., Zhou, J.-C., Sun, J.-T., et al. (2018). Incidence of facultative bacterial endosymbionts in spider mites associated with local environments and host plants. Appl. Environ. Microbiol. 84. doi: 10.1128/AEM.02546-17
Keywords: endosymbiont, temperature, insects, prevalence, induced phenotypes
Citation: Martins M, César CS and Cogni R (2023) The effects of temperature on prevalence of facultative insect heritable symbionts across spatial and seasonal scales. Front. Microbiol. 14:1321341. doi: 10.3389/fmicb.2023.1321341
Edited by:
George Tsiamis, University of Patras, GreeceReviewed by:
Katerina Nikolouli, International Atomic Energy Agency, AustriaMuhammad Zeeshan Ahmed, Bahauddin Zakariya University, Pakistan
Copyright © 2023 Martins, César and Cogni. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rodrigo Cogni, cmNvZ25pQHVzcC5icg==