 Yingjie Shen
Yingjie Shen Hao Liu2†
Hao Liu2† Xiangyi Meng
Xiangyi Meng Fulan Hu
Fulan Hu
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 22 December 2023
Sec. Systems Microbiology
Volume 14 - 2023 | https://doi.org/10.3389/fmicb.2023.1290909
This article is part of the Research Topic Untangle the Broad Connections and Tight Interactions Between Human Microbiota and Complex Diseases Through Data-Driven Approaches, Volume II View all 32 articles
Background: Recent studies have suggested that the composition of gut microbiota (GM) may change after intracerebral hemorrhage. However, the causal inference of GM and hemorrhagic stroke is unknown. Mendelian Randomization (MR) is an effective research method that removes confounding factors and investigates the causal relationship between exposure and outcome. This study intends to explore the causal relationship between GM and hemorrhagic stroke with the help of MR.
Methods: Univariable and multivariable MR analyses were performed using summary statistics of the GM (n = 18,340) in the MiBioGen consortium vs. the FinnGen consortium R9 summary statistics (intracerebral hemorrhage, subarachnoid hemorrhage, and nontraumatic intracranial hemorrhage). Causal associations between gut microbiota and hemorrhagic stroke were analyzed using inverse variance weighted, MR-Egger regression, weighted median, weighted mode, simple mode, and MR-PRESSO. Cochran’s Q statistic, MR-Egger regression, and leave-one-out analysis were used to test for multiplicity and heterogeneity of instrumental variables. Separate reverse MR analyses were performed for microbiota found to be causally associated with hemorrhagic stroke in the forward MR analysis. Also, multivariate MR analyses were conducted after incorporating common confounders.
Results: Based on the results of univariable and multivariate MR analyses, Actinobacteria (phylum) (OR, 0.80; 95%CI, 0.66–0.97; p = 0.025) had a protective effect against hemorrhagic stroke, while Rikenellaceae RC9 gut group (genus) (OR, 0.81; 95%CI, 0.67–0.99; p = 0.039) had a potential protective effect. Furthermore, Dorea (genus) (OR, 1.77; 95%CI, 1.27–2.46; p = 0.001), Eisenbergiella (genus) (OR, 1.24; 95%CI, 1.05–1.48; p = 0.013) and Lachnospiraceae UCG008 (genus) (OR, 1.28; 95%CI, 1.01–1.62; p = 0.041) acted as potential risk factors for hemorrhagic stroke. The abundance of Dorea (genus) (β, 0.05; 95%CI, 0.002 ~ 0.101; p = 0.041) may increase, and that of Eisenbergiella (genus) (β, −0.072; 95%CI, −0.137 ~ −0.007; p = 0.030) decreased after hemorrhagic stroke according to the results of reverse MR analysis. No significant pleiotropy or heterogeneity was detected in any of the MR analyses.
Conclusion: There is a significant causal relationship between GM and hemorrhagic stroke. The prevention, monitoring, and treatment of hemorrhagic stroke through GM represent a promising avenue and contribute to a deeper understanding of the mechanisms underlying hemorrhagic stroke.
Hemorrhagic stroke is a sudden condition caused by the rupture of a specific blood vessel in the brain (Di Biase et al., 2023). It is classified based on two main types of bleeding sites: intracerebral hemorrhage (ICH) within the brain parenchyma and subarachnoid hemorrhage (SAH) within the subarachnoid space (Ikram et al., 2012; Hoh et al., 2023). In the year 2019, the global burden of stroke exhibited notable figures: 12.2 million incident cases (95% UI 11.0–13.6), 101 million prevalent cases (95% UI 93.2–111), 143 million disability-adjusted life-years attributed to stroke (95% UI 133–153), and 6.55 million deaths stemming from this condition (95% UI 6.00–7.02). Within this spectrum, intracerebral hemorrhage accounted for 27.9% of all incident strokes, while subarachnoid hemorrhage constituted 9.7% (Collaborators, 2021). Such statistics shed light on the significant global health impact of stroke in terms of morbidity and mortality, necessitating sustained efforts in research and intervention to mitigate its consequences. The hemorrhagic stroke seriously affects the prognosis of the patients and, at the same time, creates a heavy financial burden for the patients. Therefore, early identification of risk factors for hemorrhagic stroke is necessary. So far, hypertension, diabetes mellitus, alcohol, obesity, smoking, and genetic factors are risk factors for hemorrhagic stroke, and the irrational development of these factors promotes cerebrovascular instability and thus contributes to the occurrence of hemorrhagic events (Magid-Bernstein et al., 2022). In addition, the link between dietary and intestinal factors and nerves is gradually being emphasized, although there is not sufficient evidence yet (Li et al., 2018).
The human gut is characterized by a microbiota in which bacteria are the dominant species (Lozupone et al., 2012). Recently, there has been increasing evidence that the gut microbiota (GM) is closely related to host health and is involved in the development of the etiology of a wide range of human diseases (Clemente et al., 2012). GM may communicate with the central nervous system through the microbiota-gut-brain (MGB) axis, including immune responses, neural connections, and so on (Cryan et al., 2019). Recent studies have demonstrated that gut microbial alterations after cerebral hemorrhage are a key factor in promoting secondary brain injury and prognosis (Zou et al., 2022). In addition, several studies have demonstrated that dysregulation of GM and their metabolites may have an impact on a variety of phenotypes, including vascular inflammation, blood pressure, lipid metabolism, and tissue injury and repair, which may suggest that GM may indirectly contribute to the development of cerebrovascular disease (Witkowski et al., 2020). However, GM is a large class of populations, and it is unclear whether one or more bacterial traits are involved in the development of hemorrhagic stroke. Potential shortcomings of clinical studies and preclinical trials, such as limited sample size and retrospective design, have hindered our determination of a causal relationship between GM and hemorrhagic stroke.
Mendelian randomization (MR) is a technique that employs single nucleotide polymorphisms (SNPs) as instrumental variables (IVs) obtained from a genome-wide association study (GWAS) to ascertain the causal relationship between an exposure and an outcome (Emdin et al., 2017). By Mendel’s principles of inheritance, at conception, alleles are transmitted randomly from parent to child, akin to the random allocation principle in randomized controlled trials. Therefore, MR can circumvent the limitations found in observational studies, such as prejudice and reverse causation, and enable the inference of causal relationships between complex diseases (Bowden and Holmes, 2019). MR has been widely used to explore the relationship between GM and a variety of disease phenotypes, such as immunologic disorders (Xu et al., 2021), psychiatric disorders (Ni et al., 2021), and cardiovascular diseases (Meng et al., 2023). In this study, we will discuss the causal effect of GM on hemorrhagic stroke and the corresponding inverse relationship using MR analysis based on GWAS summary statistics from the MiBioGen and FinnGen consortiums.
This study did not require additional ethical approval since we used publicly available data, which had already received approval from the appropriate ethical and institutional review boards. We selected SNPs significantly associated with exposure as IVs based on strict inclusion and exclusion criteria and included relevant IVs for MR analysis and sensitivity analyses through hypothetical criteria for MR.
We used GM and hemorrhagic stroke as exposures to observe their effects on outcomes separately. All MR analysis of this study was executed under three basic assumptions (Davies et al., 2018). Firstly, we need to ensure that genetic variants are independently and significantly associated with the exposure, a requirement that we fulfilled by utilizing p-values and F-statistics from correlation tests. Secondly, we must establish that there is no association between the genetic variants related to the exposure and the outcomes. Lastly, the third premise assumes these variants should not correlate with any confounding variables (Figure 1).
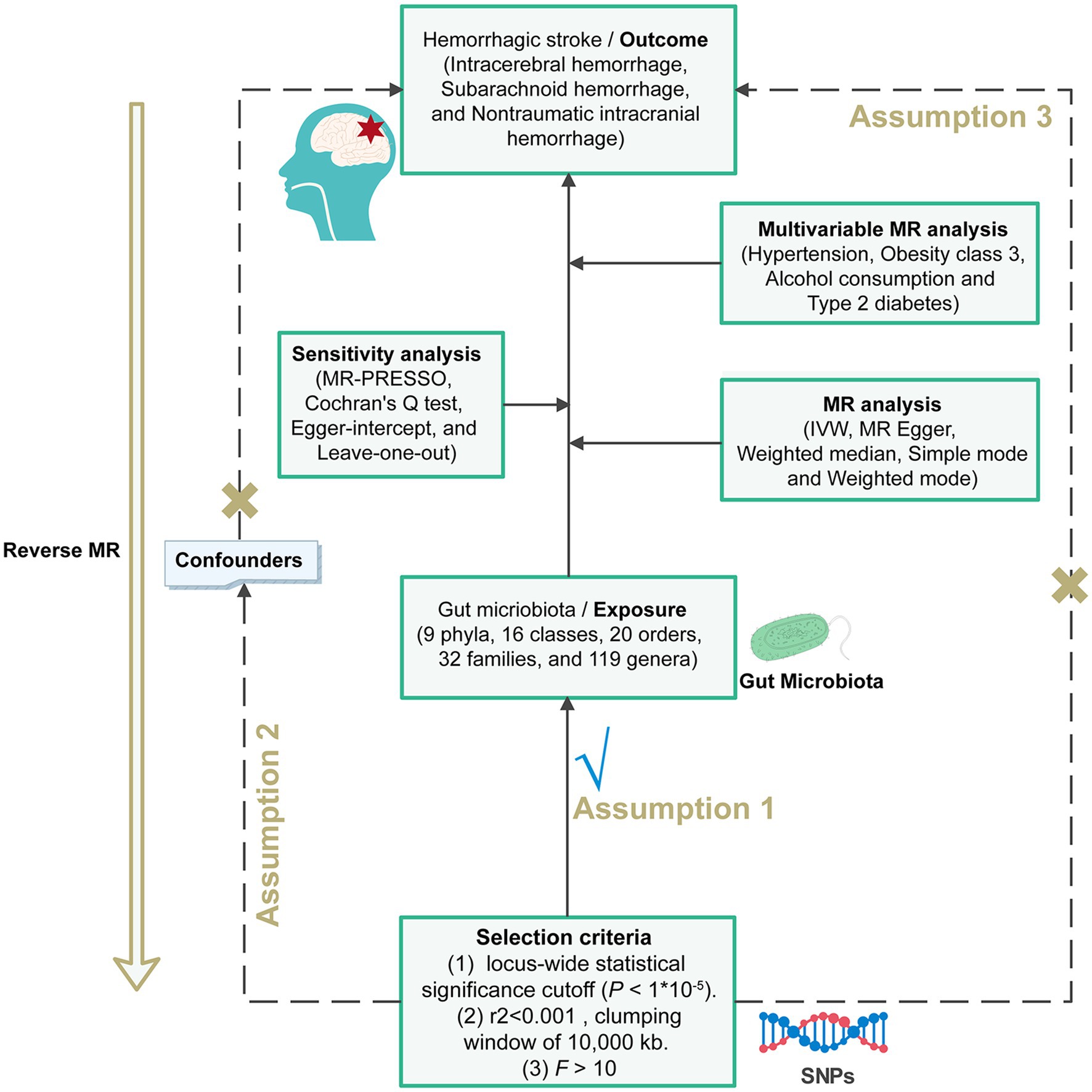
Figure 1. Flowchart of the MR randomization study and key assumptions. Three key assumptions for a valid Mendelian randomization analysis. (I) the instrumental variable selected from the dataset was significantly associated with exposure; (II) the instrumental variable was not associated with any unknown confounders of exposure; and (III) the instrumental variable could only be associated with the outcome through exposure. MR, Mendelian randomization; SNPs, single nucleotide polymorphisms; IVW, inverse-variance weighted; MR-PRESSO, MR pleiotropy residual sum and outlier.
This study is reported in accordance with the Strengthening the Reporting of Observational Studies in Epidemiology Using Mendelian Randomization guidelines (STROBE-MR) (Skrivankova et al., 2021) (Supplementary Table S1).
We acquired the most recent GWAS dataset of the human gut microbiome from the MiBioGen consortium. This dataset is the largest, multi-ethnic, genome-wide analysis of human autosomal genetic variation and the gut microbiome, specifically designed to investigate host-genetics-microbiome associations (Wang et al., 2018; Kurilshikov et al., 2021). The study involved 18,340 participants from diverse countries and regions, predominantly representing European groups. The consortium utilized standardized analytical pipelines for both microbiota phenotype and genotype, ensuring uniform data processing methods. As a result of this comprehensive study, we successfully identified 211 bacterial taxa, including 9 phyla, 16 classes, 20 orders, 35 families (of which 3 were unknown), and 131 genera (of which 12 were unknown) (Kurilshikov et al., 2021). For our study, we excluded 15 microbial taxa with missing information regarding families and genera, ultimately leaving us with 196 bacteria presenting in our initial analysis. The GWAS details are shown in Supplementary Table S2.
Hemorrhagic stroke (ICH and SAH) was defined as non-traumatic subarachnoid hemorrhage or intracerebral hemorrhage (Ohashi et al., 2023). Therefore, we obtained GWAS summary data for ICH, SAH, and nontraumatic intracranial hemorrhage (nITH) from the FinnGen Consortium’s R9 data. Specifically, the dataset for ICH consisted of 3,749 cases and 339,914 controls; the dataset for SAH included 3,289 cases and 339,922 controls; and the dataset for nITH included 6,530 cases and 342,673 controls. All the populations included in these datasets belonged to European groups (details shown in Supplementary Table S2).
The GWAS summary data for the risk factors of hemorrhagic stroke, including hypertension, obesity, alcohol drinking, and diabetes, were obtained from the corresponding consortia of the Integrative Epidemiology Unit (IEU) Open GWAS Project (Supplementary Table S2).
We used SNPs with p < 1 × 10−5 as the initial genetic instrumental variable for MR analyses. This value was determined to be the optimal threshold for many intestinal flora-associated MR studies to maximize the amount of genetic variance explained by the genetic predictors and to increase the number of eligible SNPs for sensitivity analyses (Sanna et al., 2019). Meanwhile, the F-statistic serves as a reliable measure to evaluate the robustness of instruments. It is commonly computed using the formula:
For Univariable MR analysis (UVMR), we used a variety of MR-related statistical methods, including inverse variance weighted (IVW), MR-Egger regression, weighted median, weighted mode, simple mode, and MR-PRESSO to examine whether there is a causal relationship between GM and hemorrhagic stroke. The IVW method allows us to estimate the causal effect of exposure on the outcome by incorporating ratio estimates for each SNP (Choi et al., 2019). Essentially, it transforms MR estimates into a weighted regression of SNP-outcome effects on SNP-exposure effects. While the weighted median method can produce unbiased estimates even when up to 50% of the data derives from invalid IVs (Bowden et al., 2016), the weighted mode approach is reliable when most individual instrument causal effect estimates are derived from valid instruments (Hartwig et al., 2017), even if some IVs are considered invalid. The MR-Egger method is a valuable tool for estimating causal effects through the slope coefficient from Egger regression, which helps identify and address potential small study bias (Bowden et al., 2015). Additionally, the simple mode represents an unweighted empirical density function for causal estimation (Hemani et al., 2018). The IVW provides the most precise estimates among these methods, assuming that all SNPs are valid instruments. If the IVW method yields a significant result (p < 0.05), even when other methods do not, and there is no evidence of pleiotropy or heterogeneity, the outcome can be considered positive as long as the beta values of the other methods point in the same direction (Chen et al., 2020). Meanwhile, the p-value of MR-PRESSO was less than 0.05, reinforcing the robustness of the positive results.
For the primary MR results, we corrected the p-values for False discovery rate (FDR) based on different taxa of GM and type of hemorrhagic stroke, resulting in corresponding PFDR value with a false discovery rate of PFDR < 0.1 (Storey and Tibshirani, 2003). A potential significant causality between the exposure and the outcome was considered to be present when p < 0.05 but PFDR ≥ 0.1.
For both significant and potential significant causalities, we employed MR-Egger regression to assess whether the genetic instruments had a pleiotropic effect on the outcome (Bowden et al., 2015). Additionally, MR-PRESSO was utilized to identify horizontal pleiotropy and heterogeneity and mitigate its effect by removing outliers. The Cochran’s Q test was also applied to detect heterogeneity. If heterogeneity was detected by Cochran’s Q test, along with horizontal pleiotropy of selected SNPs by MR-Egger (egger-intercept) or MR-PRESSO (Global test), the analysis was repeated after excluding these pleiotropic SNPs. An insignificant p-value (p > 0.05) indicated the absence of heterogeneity or pleiotropy. Additionally, to ensure unbiased causal estimates, we performed leave-one-out analyses by iteratively excluding each instrumental SNP and re-running the IVW analyses, enabling the identification of any influential SNP (Li et al., 2022).
In order to assess the complete correlation between GM and hemorrhagic stroke, we conducted a reverse MR analysis using GM, which had been previously identified by forward MR analysis to have a causal relationship with hemorrhagic stroke. The settings and methodologies employed were consistent with those used in the forward MR analysis. In addition, some confounding factors (hypertension, obesity, alcohol drinking, and diabetes) may influence the causal relationship between GM and hemorrhagic stroke. In order to fully adjust for confounding factors, we included the above confounders (hypertension, obesity class 3, alcohol consumption, and type 2 diabetes) further using multivariate MR analysis (MVMR) in the forward MR analysis. Similarly, IVW, weighted median, and MR-Egger regression were used for MVMR analysis. The Egger-intercept test and Cochran’s Q test were also used to assess the stability of the results.
All statistical analyses were conducted with R (version 4.3.1), and MR analyses were executed using the TwosampleMR (Hemani et al., 2018), MR-PRESSO (Verbanck et al., 2018), and Mendelian Randomization R packages.
Strictly based on the selection criteria of IVs, we excluded four intestinal bacterial taxa that did not make it into the MR analysis. Finally, 192 intestinal bacterial taxa were included in the MR analysis. In total, we performed MR analysis after selecting SNPs that met the criteria for ICH (2030), SAH (2030) and nITH (2032).
The results of all MR analyses are shown in Figures 2–4, which record the beta values of all MR analysis methods and the p-values of the IVW method. And then, we identified the positive results of the MR analysis based on IVW’s significant p-values. In addition, the direction of the beta results for all five methods is consistent, increasing the credibility of the true causal relationships (Table 1, Figures 5, 6). The F-statistics of the IVs included in the analysis significantly associated with the GM were all greater than 10, indicating that the estimates are unlikely to suffer from weak instrumental bias (Supplementary Table S1).
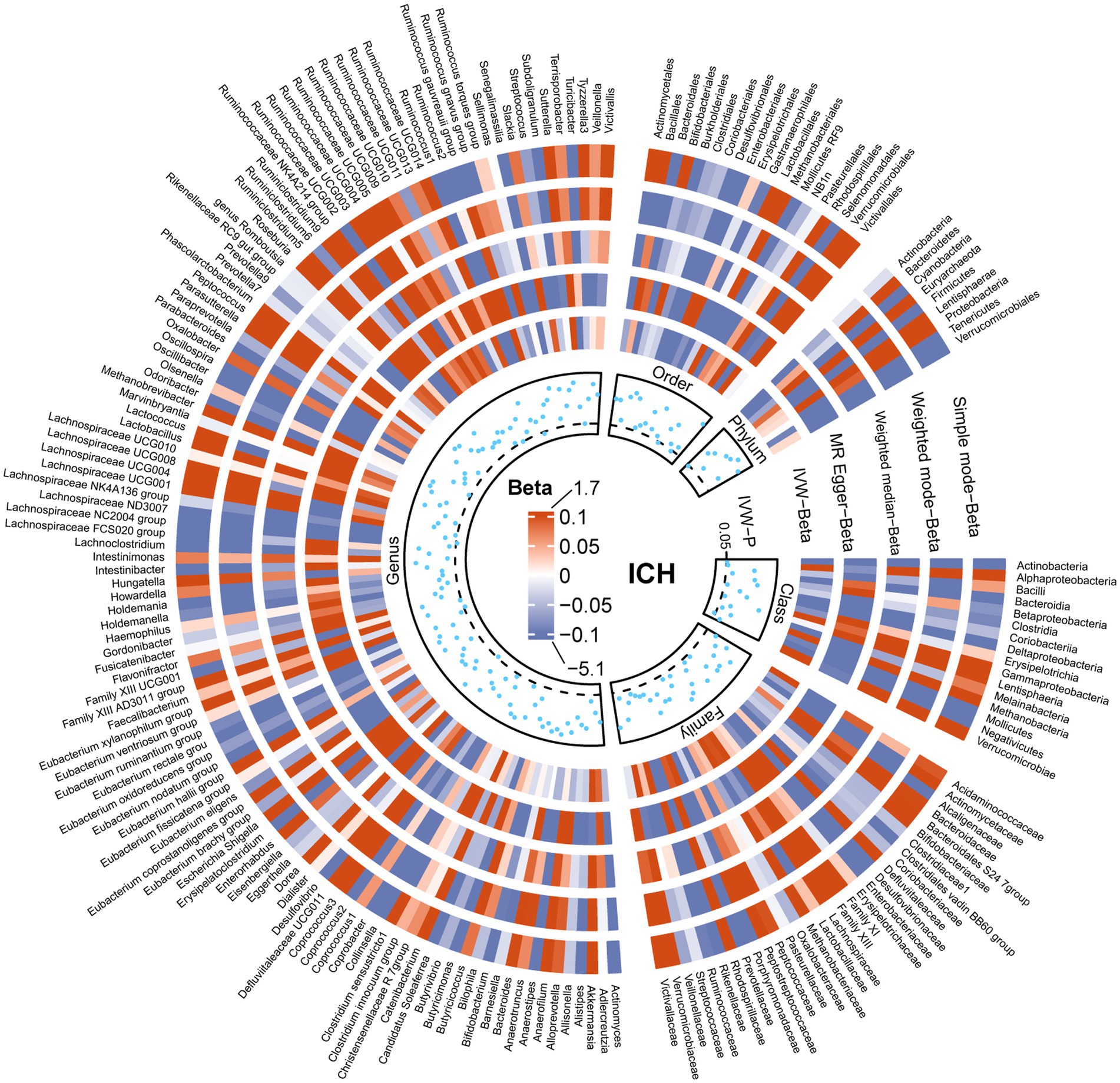
Figure 2. Causal effect of the gut microbiota on intracerebral hemorrhage based on MR analyses. From outside to inside, the beta values of simple mode, weighted mode, weighted median, MR-Egger, inverse variance weighted, and p-value inverse variance weighted are represented, respectively. MR, Mendelian randomization; IVW, Inverse variance weighted.
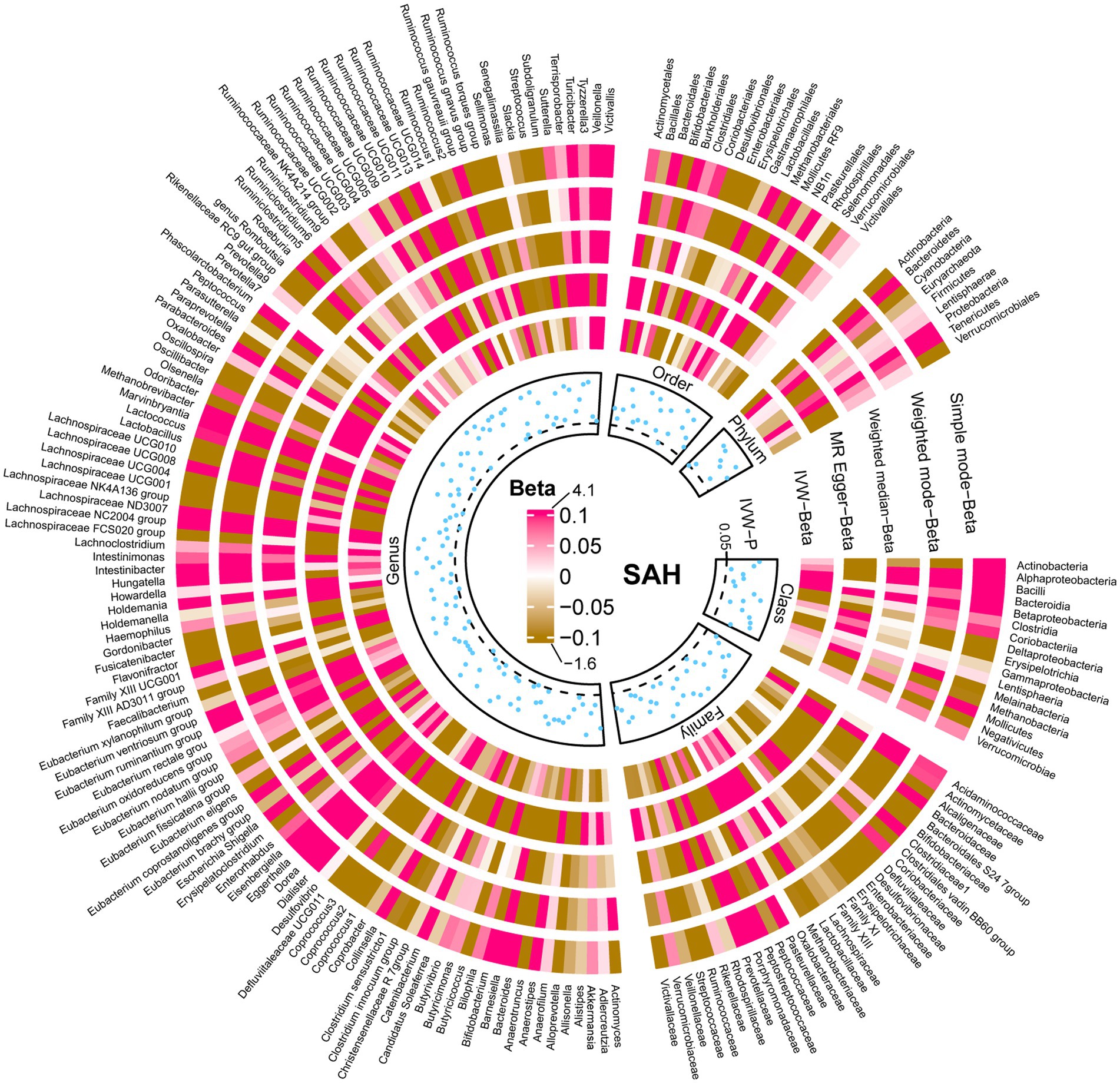
Figure 3. Causal effect of the gut microbiota on subarachnoid hemorrhage based on MR analyses. From outside to inside, the beta values of simple mode, weighted mode, weighted median, MR-Egger, inverse variance weighted, and p-value inverse variance weighted are represented, respectively. MR, Mendelian randomization; IVW, Inverse variance weighted.
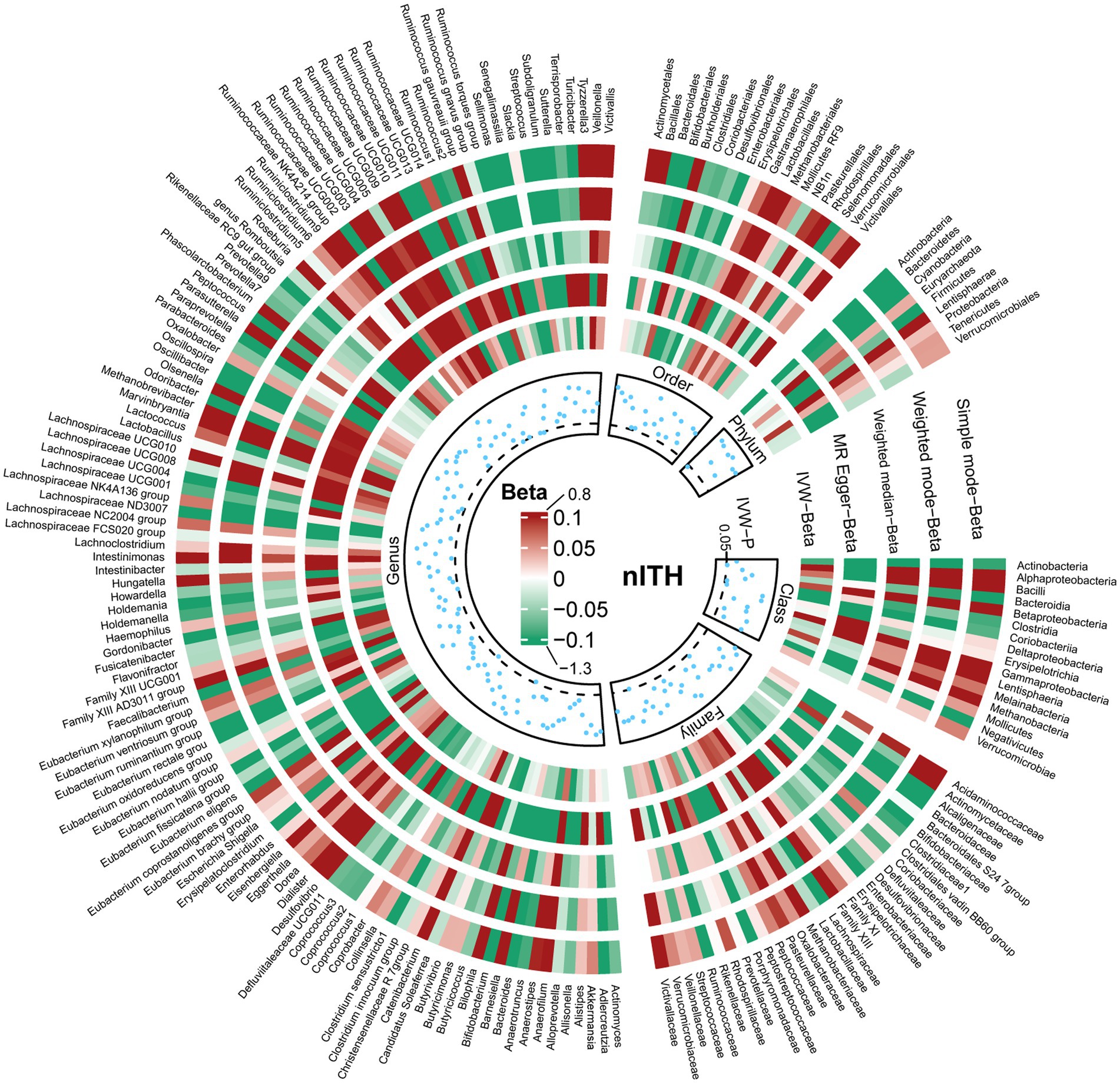
Figure 4. Causal effect of the gut microbiota on nontraumatic intracranial hemorrhage based on MR analyses. From outside to inside, the beta values of simple mode, weighted mode, weighted median, MR-Egger, inverse variance weighted, and p-value inverse variance weighted are represented, respectively. MR, Mendelian randomization; IVW, Inverse variance weighted.
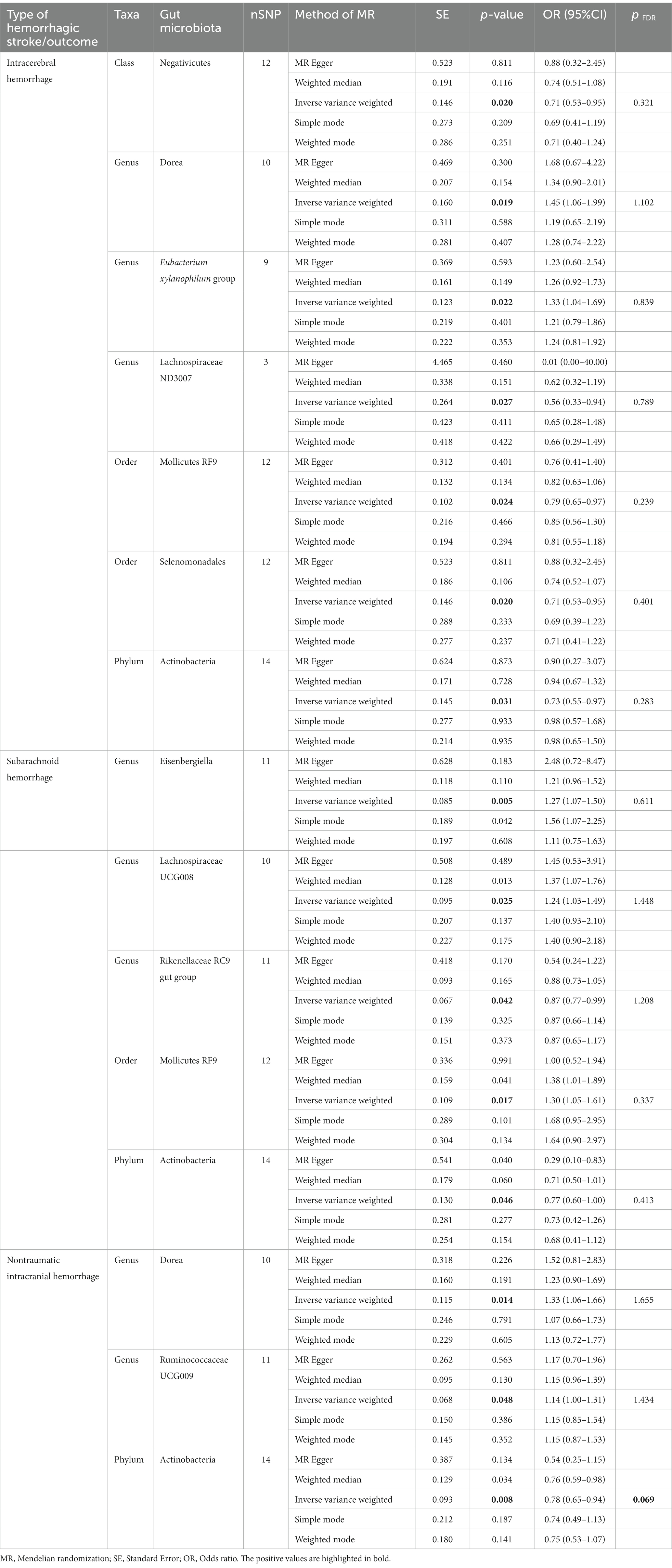
Table 1. Mendelian randomization analysis results between Gut microbiota and hemorrhagic stroke.
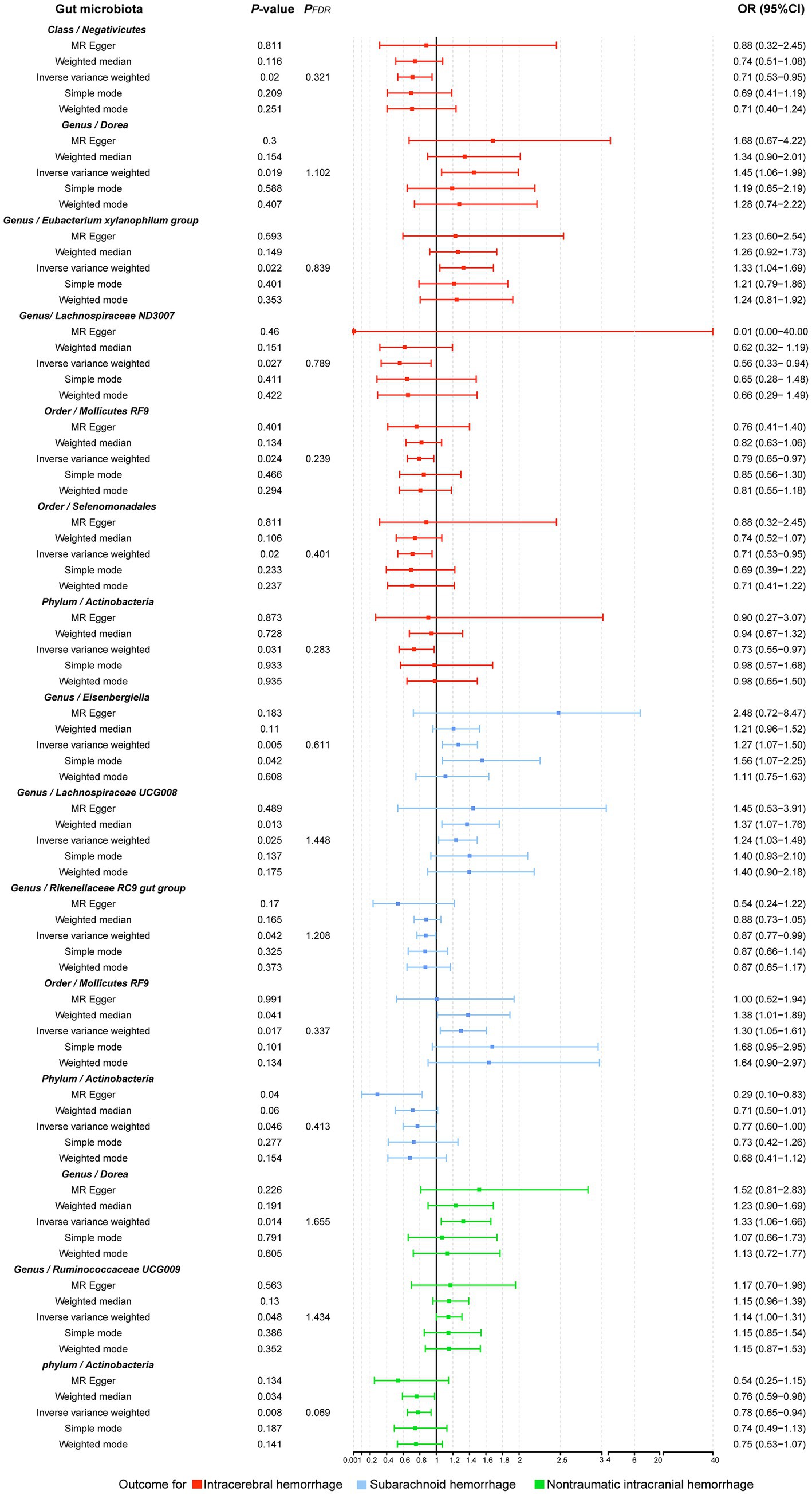
Figure 5. Forest plot illustrating the causal effect of the gut microbiota on hemorrhagic stroke using five methods of MR analyses.
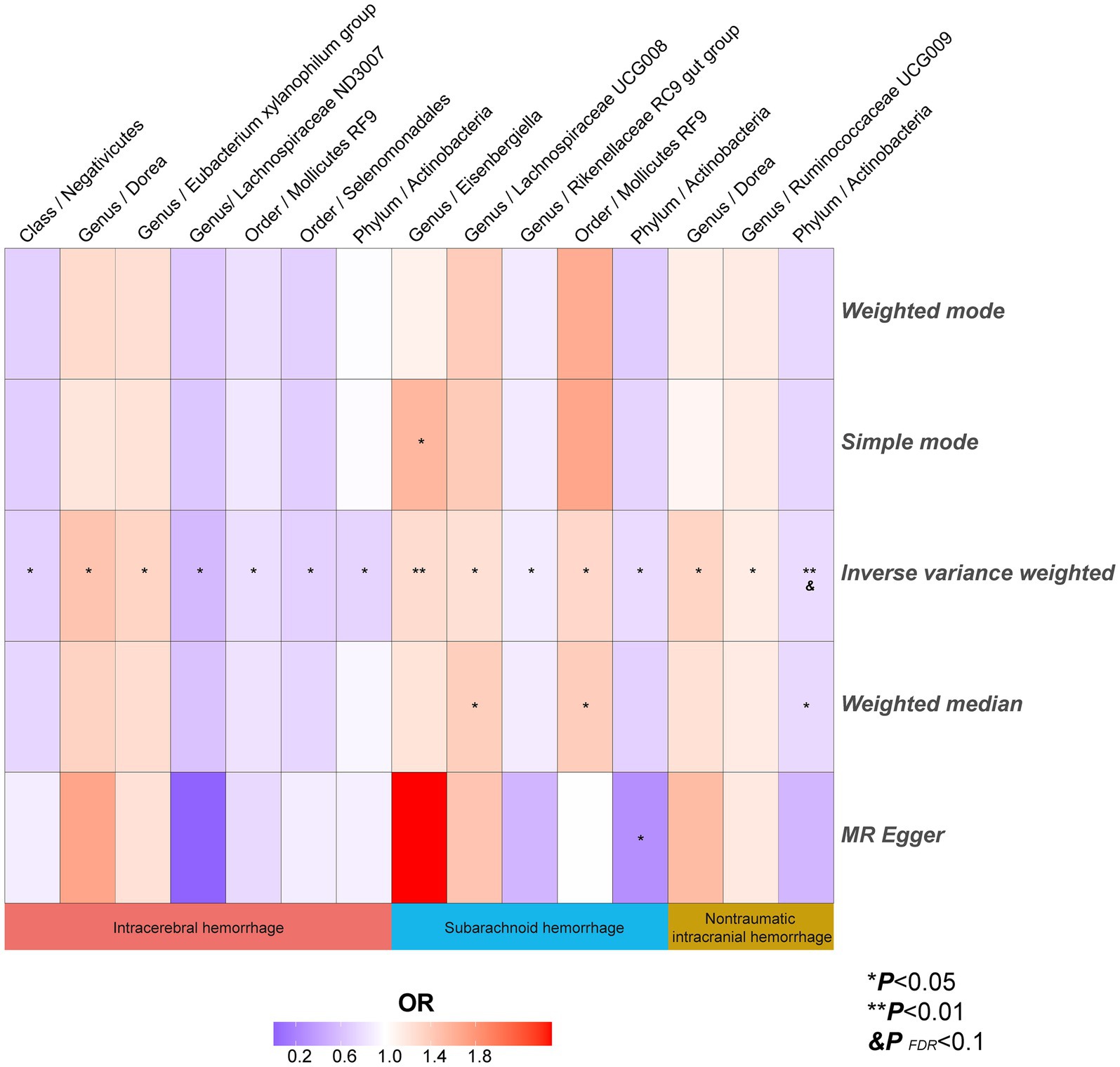
Figure 6. The heatmaps of five MR analysis methods. Different color blocks represent different odds ratio values. MR, Mendelian randomization.
Analysis of the MR showed that the relative abundance of the genetically predicted 1 class (Negativicutes), 3 genera (Dorea, Eubacterium xylanophilum group, Lachnospiraceae ND3007),2 order (Mollicutes RF9, Selenomonadales), 1 phylum (Actinobacteria) were potentially associated with the risk of ICH. Specifically, Negativicutes (class) (OR, 0.71; 95%CI, 0.53–0.95; p = 0.020), Lachnospiraceae ND3007 (genus), (OR, 0.56; 95%CI, 0.33–0.94; p = 0.027), Mollicutes RF9 (order) (OR, 0.79; 95%CI, 0.65–0.97; p = 0.024), Selenomonadales (order) (OR, 0.71; 95%CI, 0.53–0.95; p = 0.020), Actinobacteria (phylum) (OR, 0.73; 95%CI, 0.55–0.97; p = 0.031) were potentially protective against ICH, i.e., higher abundance of them, the lower the risk of ICH may be. The relative abundance of Dorea (genus) (OR, 1.45; 95%CI, 1.06–1.99; p = 0.019) and Eubacterium xylanophilum group (genus) (OR, 1.33; 95%CI, 1.04–1.69; p = 0.022) may contribute to the development of ICH, and the higher the abundance of these two genera, the higher the potential risk of ICH occurrence (Table 1, Figures 5, 6).
Similarly, the relative abundance of genetically predicted taxa showed potential significant associations with the risk of developing SAH. Specifically, the Eisenbergiella (genus) (OR, 1.27; 95%CI, 1.07–1.50; p = 0.005), Lachnospiraceae UCG008 (genus) (OR, 1.24; 95%CI, 1.03–1.49; p = 0.025) and Mollicutes RF9 (order) (OR, 1.30; 95%CI, 1.05–1.61; p = 0.017) were potential positive association with SAH, with their higher relative abundance associated with a higher potential risk of disease. Conversely, for the Rikenellaceae RC9 gut group (genus) (OR, 0.87; 95%CI, 0.77–0.99; p = 0.042) and Actinobacteria (phylum) (OR, 0.77; 95%CI, 0.60–1.00; p = 0.046), the higher the relative abundance, the lower the potential risk of developing SAH (Table 1, Figures 5, 6).
In order to include the completeness of the population, we also selected the nITH population for MR analysis. The results showed that 2 genera and 1 phylum had a significant causal relationship with nITH. Specifically, Actinobacteria (phylum) (OR, 0.78; 95%CI, 0.65–0.94; p = 0.008, PFDR = 0.069) was negatively associated with the risk of nITH. In other words, it was a protective factor against nITH. In contrast, Dorea (genus) (OR, 1.33; 95%CI, 1.06–1.66; p = 0.014), Ruminococcaceae UCG009 (genus) (OR, 1.14; 95%CI, 1.00–1.31; p = 0.048) were potentially positively associated with the risk of nITH (Table 1, Figures 5, 6).
In the sensitivity analyses, we performed pleiotropy, heterogeneity test, and leave-one-out analysis, respectively. All MR-Egger regression intercepts did not significantly deviate from zero (all intercepts p > 0.05). Cochran’s Q test and MR-PRESSO had p-values greater than 0.05; all of the above suggested that there was no significant heterogeneity and pleiotropy (shown in Supplementary Tables S4, S5). In addition, The leave-one-out analysis revealed no specific SNPs driving the association between GM and hemorrhagic stroke (Supplementary Table S6).
By reverse MR analysis, we found the abundance of Dorea (genus) (β, 0.05; 95%CI, 0.002 ~ 0.101; p = 0.041) may be up-regulated after ICH. Furthermore, the abundance of Eisenbergiella (genus) (β, −0.072; 95%CI, −0.137 ~ −0.007; p = 0.030, PFDR = 0.090) was down-regulated when SAH acted as an exposure (shown in Supplementary Tables S7, S8). In the reverse MR analysis, we did not find heterogeneity and pleiotropy, and the leave-one-out analysis suggested that no SNP was significantly associated with the results (Supplementary Tables S9, S10).
To determine whether the above-selected microbiota exerted an impact on hemorrhagic stroke risk directly or through common hemorrhagic stroke confounders, we further conducted an MVMR analysis. In the MR analysis with different types of hemorrhagic stroke as the outcome, we considered confounding variables such as hypertension, obesity class 3, alcohol consumption, and type 2 diabetes separately, and the final results are shown in Figure 7 and Supplementary Table S11. After we subsequently considered all confounding variables simultaneously, the results are shown in Figure 8 and Table 2. Specifically, when ICH was the outcome, only Dorea (genus) remained as a potential risk factor for ICH. When SAH was used as the outcome, potential risk factors for SAH remained for Eisenbergiella (genus) and Lachnospiraceae UCG008 (genus), and potential protective factors for SAH remained for Rikenellaceae RC9 gut group (genus). When nITH was the outcome, potential risk factors for nITH were present for Dorea (genus), and protective factors for nITH were present for Actinobacteria (phylum). The causal inference was further supported by consistent direction from distinct MR models. Besides, Cochran’s Q test and intercept term derived from multivariable IVW and multivariable MR-Egger did not detect potential heterogeneity and pleiotropy (Supplementary Table S12).
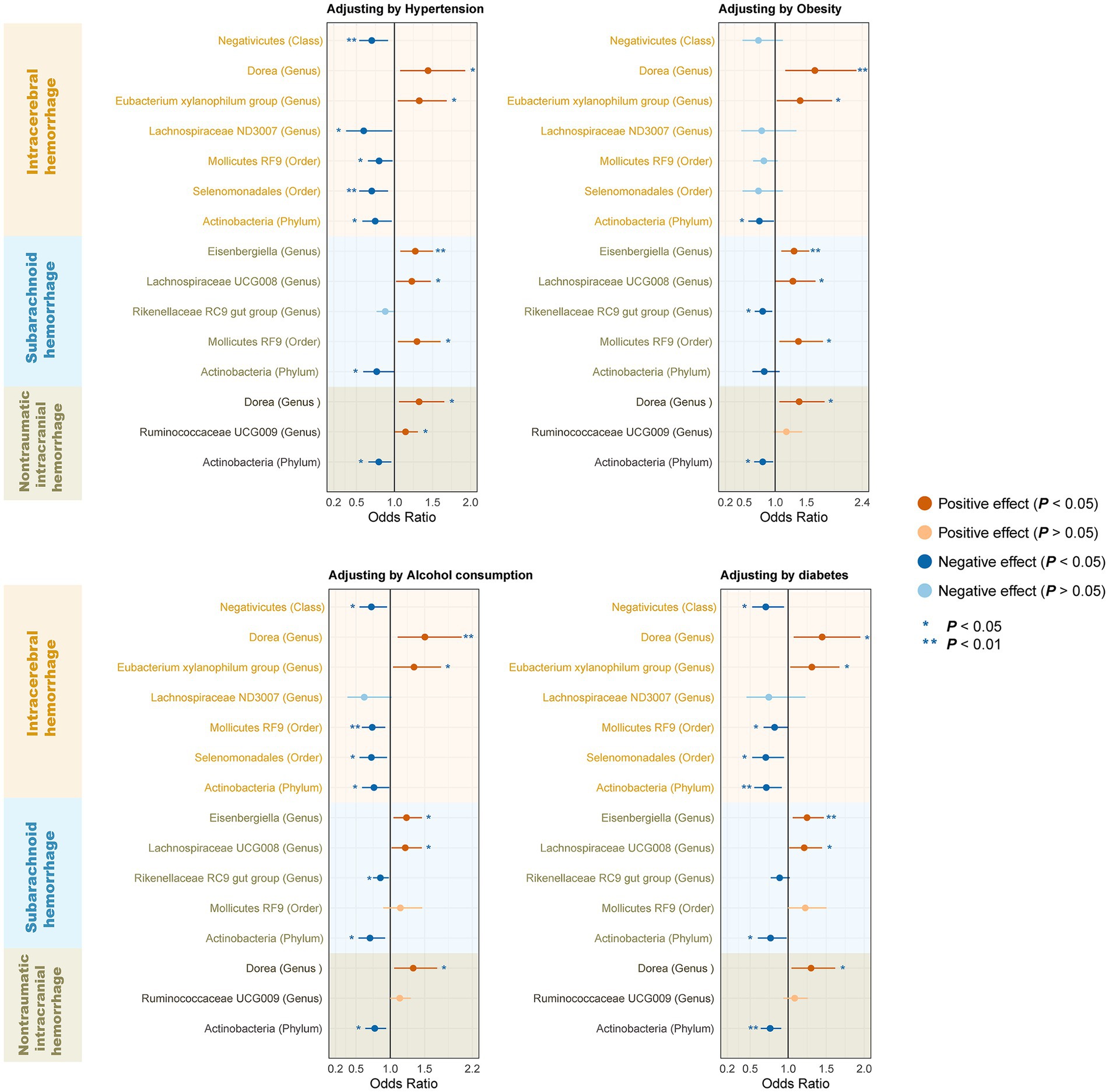
Figure 7. Forest plot illustrating the causal effect of gut microbiota on hemorrhagic stroke using the IVW method to adjust for each of the four confounders. IVW, Inverse variance weighted.
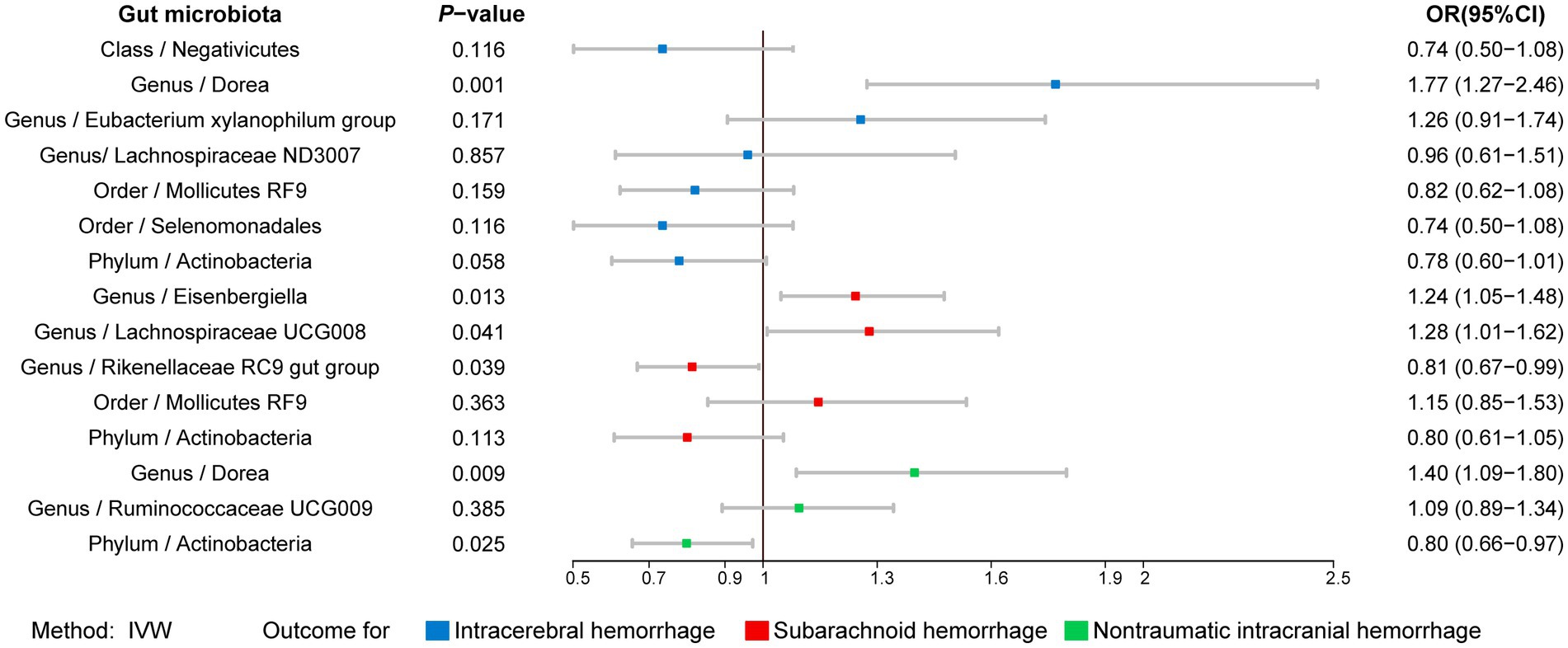
Figure 8. Forest plot illustrating the causal effect of gut microbiota on hemorrhagic stroke using IVW methods with simultaneous adjustment for four confounders. IVW, Inverse variance weighted.
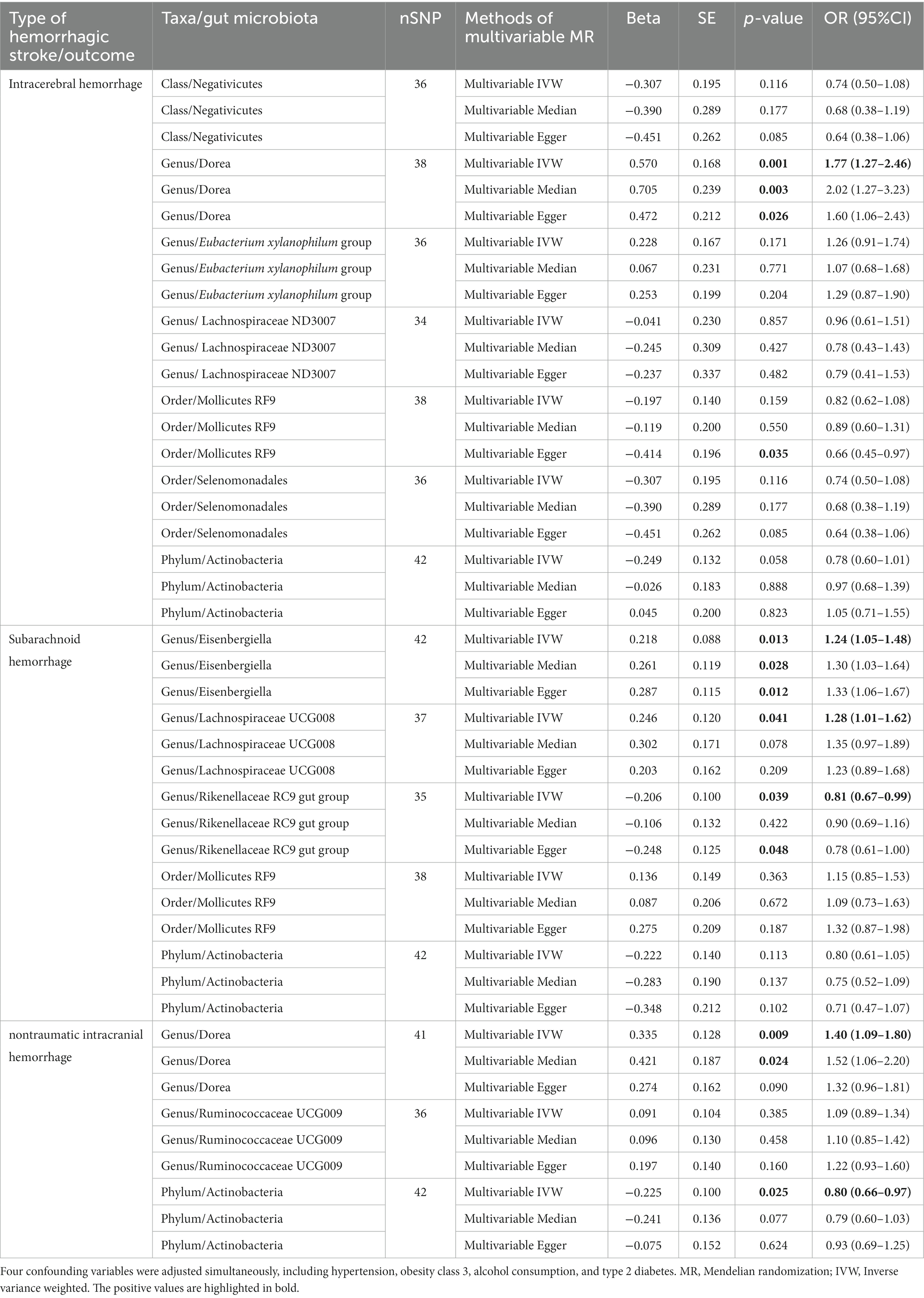
Table 2. Multivariable mendelian randomization analysis results between gut microbiota and hemorrhagic stroke (adjusting four variables).
To our best knowledge, this is the first MR analysis of GM for hemorrhagic stroke. We conducted bidirectional, two-sample, and MVMR analysis to assess the causal relationship between GM and hemorrhagic stroke. Our findings revealed that after adjusting for a range of confounders, Actinobacteria (phylum) had a protective effect against hemorrhagic stroke, while Rikenellaceae RC9 gut group (genus) had a potential protective effect. On the other hand, Dorea (genus), Eisenbergiella (genus), and Lachnospiraceae UCG008 (genus) acted as potential risk factors for hemorrhagic stroke. The occurrence of hemorrhagic stroke also alters the composition of the GM, as evidenced primarily by a potential increase in the relative abundance of Dorea (genus) and a decrease in the relative abundance of Eisenbergiella (genus). These findings suggest the potential value of GM in the identification, prevention, and management of hemorrhagic stroke.
GM plays a crucial role in human growth, development, immune responses, metabolism, and various pathophysiological processes. It exerts a direct or indirect influence on numerous aspects, including host cell proliferation, angiogenesis, intestinal endocrine function, nerve signaling, and the synthesis of essential substances (Kåhrström et al., 2016; Fan and Pedersen, 2021). Moreover, GM affects remote organs, such as the liver, kidneys, and even the central nervous system. MGB axis introduction leads to a better understanding of the onset and progression of neurological disorders (Mulak and Bonaz, 2015).
Drawing from the MGB axis, connections have been identified linking GM to an array of neurological disorders, including Parkinson’s disease, Alzheimer’s disease, and ischemic stroke. After the commencement of ischemic stroke, ischemia induces gastrointestinal paralysis and an excessive generation of nitrates, leading to a modified flora composition (Xu et al., 2021). The Aerococcaceae (family) and Flavobacterium (genus) demonstrated a substantial increase. Conversely, the Clostridiales (order), as well as the genera Lactobacillus and Stenotrophomonas, exhibited a marked decrease within the intestinal tracts of ischemic stroke patients when contrasted with the control group. Furthermore, the flora’s composition proved capable of effectively discerning between patients with differing prognoses (Chang et al., 2021). Additionally, a diminished presence of GM responsible for butyrate production is an autonomous predictor for post-stroke infection development (Haak et al., 2021). Substantial strides have been achieved in recent years concerning alterations in the abundance and composition of GM within patients afflicted by ICH. In a cohort study investigating GM in ICH patients, significant decreases in the abundance of the genera Brucella, Bacteroidota, Fusobacterium, E. faecalis, Bifidobacterium, Romboutsia, and Agathobacteria were observed. Conversely, significant increases in the abundance of the genera Ackermannia, Escherichia, Shigella, Clostridium, and Lactobacillus were observed after the onset of ICH (Xiong et al., 2022). A separate study identified increased levels of Enterococcus, Parabacteroides, Lachnoclostridium, Acidaminococcus, and Streptococcus after ICH onset, accompanied by a notable reduction in the abundance of Prevotella and Faecalibacterium (Luo et al., 2022). Therefore, changes in the GM following hemorrhagic stroke have been continuously validated. Within our investigation, we examined the causal impact of hemorrhagic stroke on select members of the GM through reversed MR. Additionally, we supplemented our analysis with Dorea (genus) and Eisenbergiella (genus), taxa that exhibited changes in response to hemorrhagic stroke.
The disruption of GM and metabolite composition further amplifies secondary damage after hemorrhagic stroke. Within preclinical investigations, mice within the ICH model displayed disrupted GM alongside an inflammatory reaction in the hematoma’s periphery, coupled with the presence of intestinal lymphocytes. Notably, mice that received a transplantation of regular GM exhibited a marked reduction in neuroinflammation (Yu et al., 2021). To put it differently, the modified GM following cerebral hemorrhage intensifies the harm caused by neuroinflammation—an element recognized as pivotal in secondary brain injury and unfavorable post-hemorrhagic stroke prognoses (Candelario-Jalil et al., 2022). Furthermore, ongoing inflammatory cell infiltration progressively harms vascular endothelial cells, resulting in heightened fragility of vessel walls along with both structural and functional decline. Consequently, this elevates the vulnerability to hemorrhagic stroke (Low et al., 2019). More specifically, in pathological conditions such as hemorrhagic stroke, the GM changes as described above. For example, there is a reduction in the populations of beneficial GM, while the presence of GM that produces harmful compounds increases. The subsequent gradual accumulation of harmful metabolites from GM induces changes in gut permeability. Histomorphological and ultrastructural examinations after ICH revealed deterioration of the intestinal epithelium. This was manifested as thickening, shortening, and fusion of the villi and fragmentation of epithelial cells on the surface of the villi (Cheng et al., 2016). Inflammation initiates a cascade of events resulting in harm to the intestinal epithelium, ongoing impairment of the intestinal barrier function, and eventual liberation of metabolites from intestinal bacteria into the circulatory system. Sequential stimulation of inflammatory pathway molecules in the bloodstream, including NOD-like receptor thermal protein domain associated protein-3 and Nuclear factor-κB, the inflammatory mediator Matrix metalloprotein-9, specific members of the interleukin family, and reactive oxygen species, induces the disruption of tight junctions within the blood–brain barrier’s architecture. Concurrently, vascular endothelial cell apoptosis occurs, further heightening susceptibility to hemorrhagic stroke and secondary cerebral edema (Guo et al., 2015; Zhang et al., 2022). Within a population-based study, an increase in Enterococcus abundance and a reduction in Prevotella abundance within the intestines of patients from the ICH group was observed to stimulate the progression of stroke-associated pneumonia, potentially via linked inflammatory factors (Luo et al., 2022), this outcome additionally bolsters the firmly interconnected and distinguishable correlation between GM and inflammation. Dorea (genus) is believed to be closely linked to inflammation and may worsen the severity of the disease when present in increased abundance, as seen in irritable bowel syndrome (Xu et al., 2021). Furthermore, the abundance of Dorea (genus) was significantly higher in an obese rat model, suggesting that it could contribute to the development of chronic inflammation in the organism (Jiao et al., 2018). In our study by MVMR analysis, Dorea (genus) was identified as a potential risk factor for stroke, suggesting that it may influence the development of hemorrhagic stroke through inflammation. Eisenbergiella (genus), however, yielded inconsistent results. We found that Eisenbergiella could promote the development of hemorrhagic stroke, and it is believed that its abundance significantly rises in patients following a high-fat/low-fiber diet (Bailén et al., 2020). However, the abundance of Eisenbergiella was reduced after hemorrhagic stroke. We speculate that (1) differences may be due to population samples, and (2) Eisenbergiella (genus) can also contribute to the production of short-chain fatty acids (SCFAs) (Tian et al., 2022), which are thought to be closely associated with the development of secondary brain injury (Sadler et al., 2020). Thus, it may be low in abundance in the hemorrhagic stroke population.
We found that Actinobacteria (phylum) reduces the risk of hemorrhagic stroke. A current study found an elevated abundance of Actinobacteria in a rat model of hemorrhagic transformation compared to non-hemorrhagic transformation rats (Huang et al., 2022). Bifidobacterium belongs to Actinobacteria (phylum) and is thought to improve blood glucose and lipids, as well as act as an antioxidant (Tonucci et al., 2017). Meanwhile, Actinobacteria (phylum), one of the major producers of antibiotics and butyrate, may reduce the risk of cerebrovascular lesions by controlling inflammation (Barka et al., 2016; Zhao et al., 2018). Butyrate is an essential SCFA that strengthens the intestinal barrier and exerts anti-inflammatory properties (Mills et al., 2019). Butyrate depletion is associated with risk factors for hemorrhagic stroke, such as diabetes, obesity, and neuroinflammation (Huang et al., 2022). Therefore, all of the factors mentioned above may help Actinomycetes (phylum) to reduce the risk of hemorrhagic stroke. It has also been shown that the Rikenellaceae RC9 gut group (genus) is significantly associated with resistance to oxidative stress and regulation of lipid metabolism, which explains its role as a protective bacterium against hemorrhagic stroke (Qin et al., 2022). On the other hand, Lachnospiraceae UCG008 (genus) is closely related to T cells and NK cells, suggesting that it may also have contributed to hemorrhagic stroke through an inflammatory response (Pang et al., 2022). Thus, inflammation-related factors may link GM and hemorrhagic stroke.
Given the complexities of conducting randomized controlled trials, we have used MR methods to delineate further the genetic-level determinants associated with hemorrhagic stroke, thereby improving the precision of early hemorrhagic stroke prevention and secondary injury mitigation. Nevertheless, large randomized controlled trials remain essential.
Our study has several limitations. First, the GWAS statistics are derived from a European population, which may limit the generalizability of the results. Second, due to the strict threshold filtering (p < 1 × 10−5) applied to IVs, there is a possibility of some potential spurious associations. Third, since we could not obtain individual-level data from the GWAS data, we were unable to analyze the detailed stratification of patient data, and thus were unable to explore the interactions between GM and different strata of hemorrhagic stroke patients. Despite this, as in this case, we were able to establish a causal relationship between GM and hemorrhagic stroke through a series of rigorous MR analyses, which set the foundation for our further studies.
We established a causal relationship between GM and the occurrence of hemorrhagic stroke through univariate and multivariate MR analyses. Dorea (genus), Eisenbergiella (genus), and Lachnospiraceae UCG008 (genus) may be associated with an increased risk of hemorrhagic stroke, whereas Rikenellaceae RC9 gut group (genus) and Actinobacteria (phylum) may be associated with a decreased risk of hemorrhagic stroke. Specifically, a higher abundance of Actinobacteria (phylum) is linked to a reduced risk of nITH. Inflammation likely plays a crucial role in connecting GM and hemorrhagic stroke. Future research should include more comprehensive investigations to verify the causal relationship between GM and hemorrhagic stroke, as well as to explore the underlying mechanisms.
Publicly available datasets were analyzed in this study. This data can be found here: the MiBioGen, https://mibiogen.gcc.rug.nl/menu/main/home; the FinnGen, https://r9.finngen.fi/; and the IEU Open GWAS, https://gwas.mrcieu.ac.uk/.
Ethical approval was not required for the studies involving humans because This study did not require additional ethical approval since we used publicly available data, which had already received approval from the appropriate ethical and institutional review boards. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent for participation was not required from the participants or the participants’ legal guardians/next of kin in accordance with the national legislation and institutional requirements because this study did not require additional ethical approval since we used publicly available data, which had already received approval from the appropriate ethical and institutional review boards.
YS: Conceptualization, Formal Analysis, Methodology, Visualization, Writing – original draft. HLiu: Data curation, Formal Analysis, Software, Writing – review & editing. XM: Methodology, Software, Validation, Writing – review & editing. AG: Investigation, Methodology, Supervision, Writing – review & editing. YL: Data curation, Investigation, Writing – review & editing. WM: Software, Visualization, Writing – review & editing. HLi: Conceptualization, Funding acquisition, Resources, Supervision, Writing – review & editing. FH: Conceptualization, Funding acquisition, Project administration, Resources, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the National Natural Science Foundations of China (No. 82071368, 82271340) and the Outstanding Youth Funding of Harbin Medical University (No. HYD2020JQ0016).
We thank all the volunteers who participated in this study and expressed our gratitude to the participants and researchers in the MiBioGen and FinnGen study groups.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2023.1290909/full#supplementary-material
Bailén, M., Bressa, C., Martínez-López, S., González-Soltero, R., Montalvo Lominchar, M. G., San Juan, C., et al. (2020). Microbiota features associated with a high-fat/Low-Fiber diet in healthy adults. Front. Nutr. 7:583608. doi: 10.3389/fnut.2020.583608
Barka, E. A., Vatsa, P., Sanchez, L., Gaveau-Vaillant, N., Jacquard, C., Meier-Kolthoff, J. P., et al. (2016). Taxonomy, physiology, and natural products of Actinobacteria. Microbiol. Mol. Biol. Rev. 80, 1–43. doi: 10.1128/MMBR.00019-15
Bowden, J., Davey Smith, G., and Burgess, S. (2015). Mendelian randomization with invalid instruments: effect estimation and bias detection through Egger regression. Int. J. Epidemiol. 44, 512–525. doi: 10.1093/ije/dyv080
Bowden, J., Davey Smith, G., Haycock, P. C., and Burgess, S. (2016). Consistent estimation in Mendelian randomization with some invalid instruments using a weighted median estimator. Genet. Epidemiol. 40, 304–314. doi: 10.1002/gepi.21965
Bowden, J., and Holmes, M. V. (2019). Meta-analysis and Mendelian randomization: a review. Res. Synth. Methods 10, 486–496. doi: 10.1002/jrsm.1346
Candelario-Jalil, E., Dijkhuizen, R. M., and Magnus, T. (2022). Neuroinflammation, stroke, blood-brain barrier dysfunction, and imaging modalities. Stroke 53, 1473–1486. doi: 10.1161/STROKEAHA.122.036946
Chang, Y., Woo, H. G., Jeong, J. H., Kim, G. H., Park, K. D., and Song, T. J. (2021). Microbiota dysbiosis and functional outcome in acute ischemic stroke patients. Sci. Rep. 11:10977. doi: 10.1038/s41598-021-90463-5
Chen, X., Kong, J., Diao, X., Cai, J., Zheng, J., Xie, W., et al. (2020). Depression and prostate cancer risk: a Mendelian randomization study. Cancer Med. 9, 9160–9167. doi: 10.1002/cam4.3493
Cheng, Y., Wei, Y., Yang, W., Cai, Y., Chen, B., Yang, G., et al. (2016). Ghrelin attenuates intestinal barrier dysfunction following intracerebral hemorrhage in mice. Int. J. Mol. Sci. 17:2032. doi: 10.3390/ijms17122032
Choi, K. W., Chen, C. Y., Stein, M. B., Klimentidis, Y. C., Wang, M. J., Koenen, K. C., et al. (2019). Assessment of bidirectional relationships between physical activity and depression among adults: a 2-sample Mendelian randomization study. JAMA Psychiatry 76, 399–408. doi: 10.1001/jamapsychiatry.2018.4175
Clemente, J. C., Ursell, L. K., Parfrey, L. W., and Knight, R. (2012). The impact of the gut microbiota on human health: an integrative view. Cells 148, 1258–1270. doi: 10.1016/j.cell.2012.01.035
Collaborators, G. S. (2021). Global, regional, and national burden of stroke and its risk factors, 1990-2019: a systematic analysis for the global burden of disease study 2019. Lancet Neurol. 20, 795–820. doi: 10.1016/S1474-4422(21)00252-0
Cryan, J. F., O'riordan, K. J., Cowan, C. S. M., Sandhu, K. V., Bastiaanssen, T. F. S., Boehme, M., et al. (2019). The microbiota-gut-brain Axis. Physiol. Rev. 99, 1877–2013. doi: 10.1152/physrev.00018.2018
Davies, N. M., Holmes, M. V., and Davey Smith, G. (2018). Reading Mendelian randomisation studies: a guide, glossary, and checklist for clinicians. BMJ 362:k601. doi: 10.1136/bmj.k601
Di Biase, L., Bonura, A., Pecoraro, P. M., Carbone, S. P., and Di Lazzaro, V. (2023). Unlocking the potential of stroke blood biomarkers: early diagnosis, ischemic vs.: Haemorrhagic differentiation and Haemorrhagic transformation risk a comprehensive review. Int. J. Mol. Sci. 24:24. doi: 10.3390/ijms241411545
Emdin, C. A., Khera, A. V., and Kathiresan, S. (2017). Mendelian Randomization. JAMA 318, 1925–1926. doi: 10.1001/jama.2017.17219
Fan, Y., and Pedersen, O. (2021). Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 19, 55–71. doi: 10.1038/s41579-020-0433-9
Guo, Z., Xu, L., Wang, X., and Sun, X. (2015). MMP-9 expression and activity is concurrent with endothelial cell apoptosis in the basilar artery after subarachnoid hemorrhaging in rats. Neurol. Sci. 36, 1241–1245. doi: 10.1007/s10072-015-2092-6
Haak, B. W., Westendorp, W. F., Van Engelen, T. S. R., Brands, X., Brouwer, M. C., Vermeij, J. D., et al. (2021). Disruptions of anaerobic gut Bacteria are associated with stroke and post-stroke infection: a prospective case-control study. Transl. Stroke Res. 12, 581–592. doi: 10.1007/s12975-020-00863-4
Hartwig, F. P., Davey Smith, G., and Bowden, J. (2017). Robust inference in summary data Mendelian randomization via the zero modal pleiotropy assumption. Int. J. Epidemiol. 46, 1985–1998. doi: 10.1093/ije/dyx102
Hemani, G., Zheng, J., Elsworth, B., Wade, K. H., Haberland, V., Baird, D., et al. (2018). The MR-base platform supports systematic causal inference across the human phenome. elife 7:e34408. doi: 10.7554/eLife.34408
Hoh, B. L., Ko, N. U., Amin-Hanjani, S., Hsiang-Yi Chou, S., Cruz-Flores, S., Dangayach, N. S., et al. (2023). 2023 guideline for the Management of Patients with Aneurysmal Subarachnoid Hemorrhage: a guideline from the American Heart Association/American Stroke Association. Stroke 54, e314–e370. doi: 10.1161/STR.0000000000000436
Huang, Q., Di, L., Yu, F., Feng, X., Liu, Z., Wei, M., et al. (2022). Alterations in the gut microbiome with hemorrhagic transformation in experimental stroke. CNS Neurosci. Ther. 28, 77–91. doi: 10.1111/cns.13736
Ikram, M. A., Wieberdink, R. G., and Koudstaal, P. J. (2012). International epidemiology of intracerebral hemorrhage. Curr. Atheroscler. Rep. 14, 300–306. doi: 10.1007/s11883-012-0252-1
Jiao, N., Baker, S. S., Nugent, C. A., Tsompana, M., Cai, L., Wang, Y., et al. (2018). Gut microbiome may contribute to insulin resistance and systemic inflammation in obese rodents: a meta-analysis. Physiol. Genomics 50, 244–254. doi: 10.1152/physiolgenomics.00114.2017
Kåhrström, C. T., Pariente, N., and Weiss, U. (2016). Intestinal microbiota in health and disease. Nature 535:47. doi: 10.1038/535047a
Kurilshikov, A., Medina-Gomez, C., Bacigalupe, R., Radjabzadeh, D., Wang, J., Demirkan, A., et al. (2021). Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat. Genet. 53, 156–165. doi: 10.1038/s41588-020-00763-1
Li, Y., Huang, Z., Jin, C., Xing, A., Liu, Y., Huangfu, C., et al. (2018). Longitudinal change of perceived salt intake and stroke risk in a Chinese population. Stroke 49, 1332–1339. doi: 10.1161/STROKEAHA.117.020277
Li, P., Wang, H., Guo, L., Gou, X., Chen, G., Lin, D., et al. (2022). Association between gut microbiota and preeclampsia-eclampsia: a two-sample Mendelian randomization study. BMC Med. 20:443. doi: 10.1186/s12916-022-02657-x
Low, A., Mak, E., Rowe, J. B., Markus, H. S., and O'brien, J. T. (2019). Inflammation and cerebral small vessel disease: a systematic review. Ageing Res. Rev. 53:100916. doi: 10.1016/j.arr.2019.100916
Lozupone, C. A., Stombaugh, J. I., Gordon, J. I., Jansson, J. K., and Knight, R. (2012). Diversity, stability and resilience of the human gut microbiota. Nature 489, 220–230. doi: 10.1038/nature11550
Luo, J., Chen, Y., Tang, G., Li, Z., Yang, X., Shang, X., et al. (2022). Gut microbiota composition reflects disease progression, severity and outcome, and dysfunctional immune responses in patients with hypertensive intracerebral hemorrhage. Front. Immunol. 13:869846. doi: 10.3389/fimmu.2022.869846
Magid-Bernstein, J., Girard, R., Polster, S., Srinath, A., Romanos, S., Awad, I. A., et al. (2022). Cerebral hemorrhage: pathophysiology, treatment, and future directions. Circ. Res. 130, 1204–1229. doi: 10.1161/CIRCRESAHA.121.319949
Meng, C., Deng, P., Miao, R., Tang, H., Li, Y., Wang, J., et al. (2023). Gut microbiome and risk of ischaemic stroke: a comprehensive Mendelian randomization study. Eur. J. Prev. Cardiol. 30, 613–620. doi: 10.1093/eurjpc/zwad052
Mills, S., Stanton, C., Lane, J. A., Smith, G. J., and Ross, R. P. (2019). Precision nutrition and the microbiome, Part I: current state of the science. Nutrients 11:923. doi: 10.3390/nu11040923
Mulak, A., and Bonaz, B. (2015). Brain-gut-microbiota axis in Parkinson's disease. World J. Gastroenterol. 21, 10609–10620. doi: 10.3748/wjg.v21.i37.10609
Ni, J. J., Xu, Q., Yan, S. S., Han, B. X., Zhang, H., Wei, X. T., et al. (2021). Gut microbiota and psychiatric disorders: a two-sample Mendelian randomization study. Front. Microbiol. 12:737197. doi: 10.3389/fmicb.2021.737197
Ohashi, S. N., Delong, J. H., Kozberg, M. G., Mazur-Hart, D. J., Van Veluw, S. J., Alkayed, N. J., et al. (2023). Role of inflammatory processes in hemorrhagic stroke. Stroke 54, 605–619. doi: 10.1161/STROKEAHA.122.037155
Pang, R., Wang, J., Xiong, Y., Liu, J., Ma, X., Gou, X., et al. (2022). Relationship between gut microbiota and lymphocyte subsets in Chinese Han patients with spinal cord injury. Front. Microbiol. 13:986480. doi: 10.3389/fmicb.2022.986480
Qin, S., He, Z., Wu, Y., Zeng, C., Zheng, Z., Zhang, H., et al. (2022). Instant dark tea alleviates Hyperlipidaemia in high-fat diet-fed rat: from molecular evidence to redox balance and beyond. Front. Nutr. 9:819980. doi: 10.3389/fnut.2022.819980
Sadler, R., Cramer, J. V., Heindl, S., Kostidis, S., Betz, D., Zuurbier, K. R., et al. (2020). Short-chain fatty acids improve Poststroke recovery via immunological mechanisms. J. Neurosci. 40, 1162–1173. doi: 10.1523/JNEUROSCI.1359-19.2019
Sanna, S., Van Zuydam, N. R., Mahajan, A., Kurilshikov, A., Vich Vila, A., Võsa, U., et al. (2019). Causal relationships among the gut microbiome, short-chain fatty acids and metabolic diseases. Nat. Genet. 51, 600–605. doi: 10.1038/s41588-019-0350-x
Skrivankova, V. W., Richmond, R. C., Woolf, B. A. R., Yarmolinsky, J., Davies, N. M., Swanson, S. A., et al. (2021). Strengthening the reporting of observational studies in epidemiology using Mendelian randomization: the STROBE-MR statement. JAMA 326, 1614–1621. doi: 10.1001/jama.2021.18236
Staiger, D. O., and Stock, J. H. (1994). Instrumental variables regression with weak instruments. National Bureau of Economic Research, Cambridge, MA.
Storey, J. D., and Tibshirani, R. (2003). Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. U. S. A. 100, 9440–9445. doi: 10.1073/pnas.1530509100
Tian, B., Geng, Y., Wang, P., Cai, M., Neng, J., Hu, J., et al. (2022). Ferulic acid improves intestinal barrier function through altering gut microbiota composition in high-fat diet-induced mice. Eur. J. Nutr. 61, 3767–3783. doi: 10.1007/s00394-022-02927-7
Tonucci, L. B., Olbrich Dos Santos, K. M., Licursi De Oliveira, L., Rocha Ribeiro, S. M., and Duarte Martino, H. S. (2017). Clinical application of probiotics in type 2 diabetes mellitus: a randomized, double-blind, placebo-controlled study. Clin. Nutr. 36, 85–92. doi: 10.1016/j.clnu.2015.11.011
Verbanck, M., Chen, C. Y., Neale, B., and Do, R. (2018). Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat. Genet. 50, 693–698. doi: 10.1038/s41588-018-0099-7
Wang, J., Kurilshikov, A., Radjabzadeh, D., Turpin, W., Croitoru, K., Bonder, M. J., et al. (2018). Meta-analysis of human genome-microbiome association studies: the MiBioGen consortium initiative. Microbiome 6:101. doi: 10.1186/s40168-018-0479-3
Witkowski, M., Weeks, T. L., and Hazen, S. L. (2020). Gut microbiota and cardiovascular disease. Circ. Res. 127, 553–570. doi: 10.1161/CIRCRESAHA.120.316242
Xiong, Z., Peng, K., Song, S., Zhu, Y., Gu, J., Huang, C., et al. (2022). Cerebral Intraparenchymal hemorrhage changes Patients' gut Bacteria composition and function. Front. Cell. Infect. Microbiol. 12:829491. doi: 10.3389/fcimb.2022.829491
Xu, K., Gao, X., Xia, G., Chen, M., Zeng, N., Wang, S., et al. (2021). Rapid gut dysbiosis induced by stroke exacerbates brain infarction in turn. Gut 70, 1486–1494. doi: 10.1136/gutjnl-2020-323263
Xu, H., Ma, C., Zhao, F., Chen, P., Liu, Y., Sun, Z., et al. (2021). Adjunctive treatment with probiotics partially alleviates symptoms and reduces inflammation in patients with irritable bowel syndrome. Eur. J. Nutr. 60, 2553–2565. doi: 10.1007/s00394-020-02437-4
Xu, Q., Ni, J. J., Han, B. X., Yan, S. S., Wei, X. T., Feng, G. J., et al. (2021). Causal relationship between gut microbiota and autoimmune diseases: a two-sample Mendelian randomization study. Front. Immunol. 12:746998. doi: 10.3389/fimmu.2021.746998
Yu, X., Zhou, G., Shao, B., Zhou, H., Xu, C., Yan, F., et al. (2021). Gut microbiota Dysbiosis induced by intracerebral hemorrhage aggravates Neuroinflammation in mice. Front. Microbiol. 12:647304. doi: 10.3389/fmicb.2021.647304
Zhang, Y., Yu, W., Flynn, C., Chang, W., Zhang, L., Wang, M., et al. (2022). Interplay between gut microbiota and NLRP3 Inflammasome in intracerebral hemorrhage. Nutrients 14:5251. doi: 10.3390/nu14245251
Zhao, J., Liu, P., Wu, Y., Guo, P., Liu, L., Ma, N., et al. (2018). Dietary Fiber increases butyrate-producing Bacteria and improves the growth performance of weaned piglets. J. Agric. Food Chem. 66, 7995–8004. doi: 10.1021/acs.jafc.8b02545
Keywords: causal inference, gut microbiota, hemorrhagic stroke, Mendelian randomization, odds ratio
Citation: Shen Y, Liu H, Meng X, Gao A, Liu Y, Ma W, Liang H and Hu F (2023) The causal effects between gut microbiota and hemorrhagic stroke: a bidirectional two-sample Mendelian randomization study. Front. Microbiol. 14:1290909. doi: 10.3389/fmicb.2023.1290909
Edited by:
Qi Zhao, University of Science and Technology Liaoning, ChinaReviewed by:
Georgia Damoraki, National and Kapodistrian University of Athens, GreeceCopyright © 2023 Shen, Liu, Meng, Gao, Liu, Ma, Liang and Hu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hongsheng Liang, bGlhbmdob25nc2hlbmdAaHJibXUuZWR1LmNu; Fulan Hu, aHVmdTE1MjVAMTYzLmNvbQ==
†These authors have contributed equally to this work and share first authorship
‡These authors have contributed equally to this work and share last authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.