
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 25 May 2023
Sec. Infectious Agents and Disease
Volume 14 - 2023 | https://doi.org/10.3389/fmicb.2023.1181034
This article is part of the Research Topic Zoonoses- A Rising Threat to Healthcare System View all 12 articles
 Helena Pětrošová1,2*
Helena Pětrošová1,2* Abanoub Mikhael2
Abanoub Mikhael2 Sophie Culos1
Sophie Culos1 Alexandre Giraud-Gatineau3
Alexandre Giraud-Gatineau3 Alloysius M. Gomez1Matthew E. Sherman4
Alloysius M. Gomez1Matthew E. Sherman4 Robert K. Ernst4
Robert K. Ernst4 Caroline E. Cameron1,5
Caroline E. Cameron1,5 Mathieu Picardeau3
Mathieu Picardeau3 David R. Goodlett1,2*
David R. Goodlett1,2*Lipid A is the hydrophobic component of bacterial lipopolysaccharide and an activator of the host immune system. Bacteria modify their lipid A structure to adapt to the surrounding environment and, in some cases, to evade recognition by host immune cells. In this study, lipid A structural diversity within the Leptospira genus was explored. The individual Leptospira species have dramatically different pathogenic potential that ranges from non-infectious to life-threatening disease (leptospirosis). Ten distinct lipid A profiles, denoted L1-L10, were discovered across 31 Leptospira reference species, laying a foundation for lipid A-based molecular typing. Tandem MS analysis revealed structural features of Leptospira membrane lipids that might alter recognition of its lipid A by the host innate immune receptors. Results of this study will aid development of strategies to improve diagnosis and surveillance of leptospirosis, as well as guide functional studies on Leptospira lipid A activity.
Lipopolysaccharide (LPS) is one of the hallmark virulence factors of Gram-negative pathogens. It consists of three parts: O-antigen, core oligosaccharide antigen, and lipid A. The O-antigen is a polysaccharide exposed to the extracellular milieu, and its size and structural complexity delays the recognition of LPS by the host immune system and limits binding to host antibodies (Duerr et al., 2009; Domínguez-Medina et al., 2020). Core oligosaccharide consists of several different monosaccharide units, and connects O-antigen with lipid A. It contributes to stability of the outer membrane and it has antigenic properties (Silipo and Molinaro, 2010). Finally, lipid A, also known as endotoxin, anchors LPS to the outer leaflet of the outer membrane. Lipid A is the membrane anchor of LPS which attaches it to the outer leaflet of the outer membrane. It comprises two glucosamine sugars decorated with fatty acyl chains and terminal phosphate groups that can be further adorned with other functional moieties (Simpson and Trent, 2019).
The biological function of lipid A is dependent on its chemical structure. Bacteria modify their lipid A to adapt to changes in their surrounding environment (Needham and Trent, 2013; Simpson and Trent, 2019; Kawahara, 2021). These structural adaptations include, for example, modifications to the length and saturation of fatty acyl chains to overcome temperature shifts (Gunn and Ernst, 2007; Hassan et al., 2020) or addition of functional groups to gain resistance to antimicrobial peptides (Trent et al., 2001; Zhou et al., 2001), and are covered in great detail in recent reviews (Simpson and Trent, 2019; Kawahara, 2021). Lipid A is also a pathogen-associated molecular pattern. It interacts with the Toll-Like Receptor 4/Myeloid Differentiation protein 2 (TLR4/MD2) complex in a structure-dependent manner (Park et al., 2009; Ohto et al., 2012; Scott et al., 2017). The canonical hexa-acylated lipid A from Escherichia coli strongly activates TLR4/MD2, and is therefore highly endotoxic (Park et al., 2009). In contrast, tetra-acylated lipid A molecules are TLR4/MD2 antagonists (Baldridge and Crane, 1999; Deguchi et al., 2016). Similarly, lipid A molecules with two terminal phosphates are stronger TLR4/MD2 ligands than their monophosphorylated counterparts (Baldridge and Crane, 1999; Kong et al., 2012). Some pathogens, such as Yersinia and Salmonella, modify their lipid A structures accordingly to evade host inflammatory responses when establishing infection (Kawahara et al., 2002; Rebeil et al., 2004; Kong et al., 2011; Chandler et al., 2020).
Leptospira is a diverse group of bacteria comprising non-infectious free-living spirochetes, as well as pathogens that cause leptospirosis in a wide variety of hosts (Coburn et al., 2021). Unlike other spirochetes, all Leptospira possess LPS in their envelopes, and this molecule is central to the host immune responses to infection (Werts et al., 2001; Nahori et al., 2005; Viriyakosol et al., 2006; Murray et al., 2010; Srikram et al., 2011; Marcsisin et al., 2013). The lipid A structure has been established in serovars of the pathogenic Leptospira (Que-Gewirth et al., 2004; Eshghi et al., 2015; Novak et al., 2022). In contrast to the canonical di-glucosamine backbone of lipid A with amide- and ester-linked primary fatty acids (Simpson and Trent, 2019), the backbone of Leptospira lipid A comprises di-aminoglucose sugars, which results in linkage of all primary fatty acids through amide bonds (Que-Gewirth et al., 2004; Eshghi et al., 2015; Novak et al., 2022). In addition, the lipid A has only one terminal phosphate that is methylated; a structural feature that has not been described in any other bacterial species to date (Que-Gewirth et al., 2004; Simpson and Trent, 2019). These unique lipid A features are likely involved in the inability of Leptospira lipid A to bind to human TLR4/MD2 (Werts et al., 2001). Similar to other bacterial pathogens, L. interrogans modify their lipid A structure to adapt to temperature shifts (Gunn and Ernst, 2007; Eshghi et al., 2015).
Given the enormous diversity of the Leptospira genus (Vincent et al., 2019), the structural diversity of its lipid A is curiously understudied (Patra et al., 2015; Vanithamani et al., 2021; Novak et al., 2022). Leptospira are fastidious bacteria that grow slowly in rich and complex culturing media supplemented with host factors (Zuerner, 2005). The traditional protocols for lipid A extraction that require large volumes of bacterial culture are therefore likely the cause of this knowledge gap. To circumvent these limitations, we employed a rapid protocol for lipid A structural characterization, FLATn (Leung et al., 2017; Sorensen et al., 2020; Yang et al., 2022a), that allowed us to utilize an estimated equivalent of 107 Leptospira cells in 1 ml volume per assay. We examined lipid A mass spectral profiles, from which representative structures were proposed, in 31 Leptospira species from different phylogenetic groups. This work therefore represents the first comprehensive comparison of lipid A structure in virulent versus nonvirulent Leptospira species.
Leptospira species used in this study are listed in Table 1. Leptospira were grown in the Ellinghausen–McCullough–Johnson–Harris (EMJH) medium, as modified by Ellis and Thierman (EMJH T80/T40/LH); medium was prepared without the addition of rabbit serum and superoxide dismutase (Ellis and Thiermann, 1986; Zuerner, 2005). Cultures were kept at 30°C and shaking at 100 rpm. For all experiments, Leptospira species were grown in biological triplicates to mid-logarithmic phase (approximately 5× 10^8 cells/ml), as assessed by density and motility under a dark-field microscope (Zuerner, 2005).
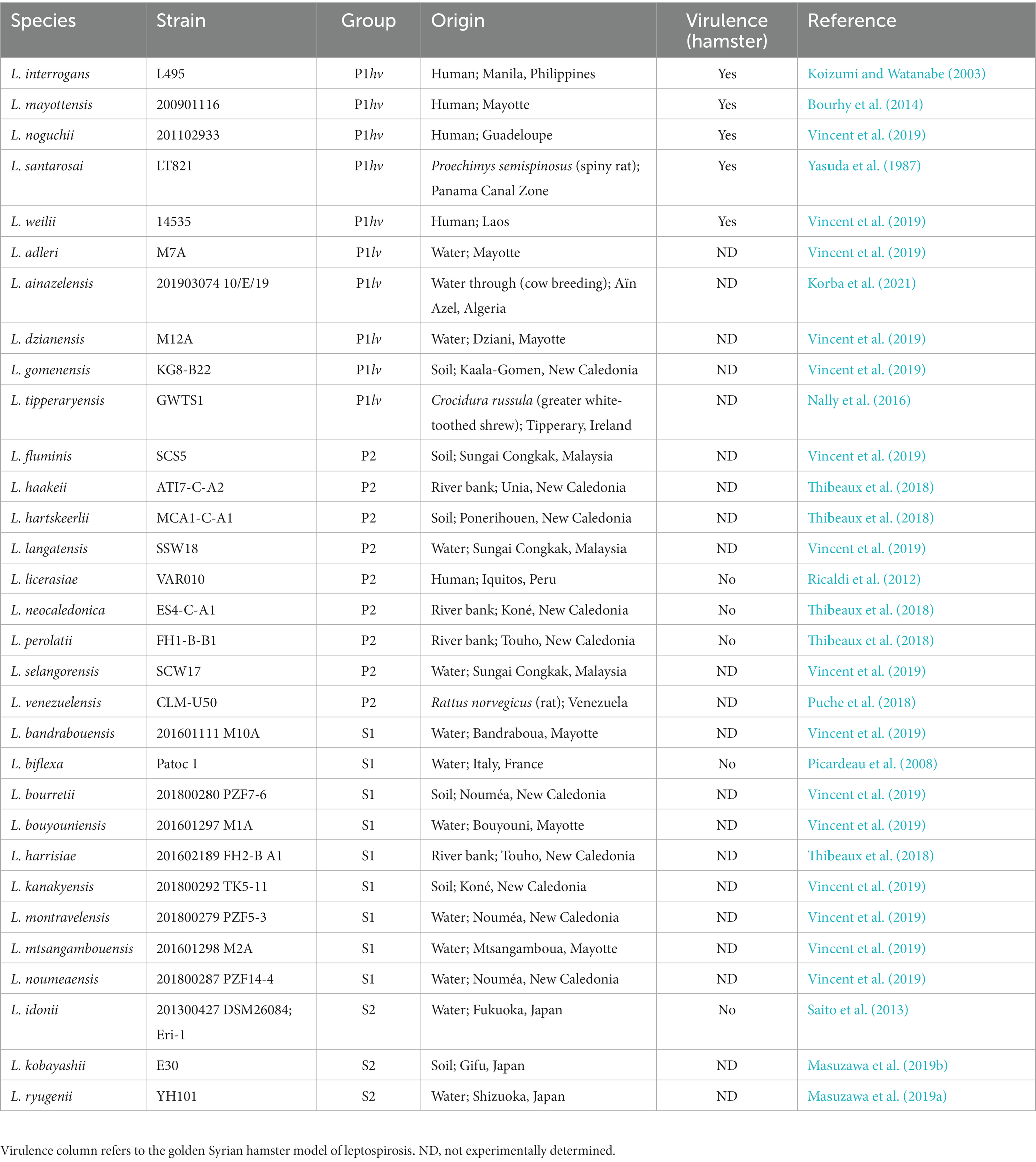
Table 1. Reference Leptospira species used in this study.
Lipid A structural analyses were performed using FLAT (Sorensen et al., 2020) and its tandem-MS version FLATn (Yang et al., 2022a). Five milliliter of logarithmic Leptospira culture was centrifuged at 4,000x g for 15 min. Resulting pellets were washed twice with 1 mL of phosphate buffered saline (Sigma Aldrich, St. Luis, MO, USA), and resuspended in 200 μL of MS-grade water (Fisher Chemical, Hampton, NH, USA). One microliter of the sample was spotted on a MALDI plate (MFX μFocus plate 12×8 c 2,400 μm 0.7 T; Hudson Surface Technology, Inc., South Korea) and air dried. One microliter of the FLAT extraction buffer (0.2 M citric acid, 0.1 M sodium citrate in MS-grade water; both from Fisher Chemical) was added on the top of each sample. MALDI plate was placed in an in-house made humidifier chamber and incubated at 110°C for 30 min. Plate was gently washed with MS-grade water for approximately 30s and let air dry. Finally, 1 μL of norharmane matrix (Sigma Aldrich) was spotted on the top of each sample and let dry. Norharmane matrix was prepared at 10 mg/mL in 2:1 v/v MS-grade chloroform and methanol (both from Fisher Chemical).
Mass spectra were obtained on a timsTOF flex MALDI-2 instrument (Bruker, Bremen, Germany) in the negative ion mode. Instrument was calibrated before each experiment in an electrospray mode by a direct infusion of the Agilent Calibration mix (Agilent Technologies, Santa Clara, CA, USA). Tandem MS analyses were performed with the following settings: 3,000 shots/spot on average, collision energy: 110–120 eV, isolation width: m/z 4, collision RF: 1,000 Vpp, transfer time: 110 μs and prepulse storage: 11 μs. To detect product ions in the low range m/z, the collision RF and transfer time were changed to 300 Vpp and 30 μs, respectively. Data were analyzed using mMass v5.5.0 (Strohalm et al., 2010) and Compass Data Analysis v 6.0 (Bruker). Fragmentation patterns of predicted lipid A structures were confirmed in ChemDraw v18.0 (PerkinElmer Informatics, Waltham, MA, USA). Theoretical isotopic distributions were predicted using Peak-by-Peak Metabolomics software v 2022.8.0 (Spectroswiss, Lausanne, Switzerland).
Recent advances in the field of Leptospira genomics led to identification of 68 reference Leptospira species, and their reclassification into four distinct phylogenetic subclades (Vincent et al., 2019; Korba et al., 2021). The P1 subclade encompasses species formerly known as “pathogens,” the P2 subclade comprises species formerly known as “intermediates,” and finally, the S1 and S2 subclades encompass non-infectious saprophytic species. The P1 subclade is further divided into P1 high virulence (P1hv) and P1 low virulence (P1lv) groups. Leptospira species most frequently involved in human disease, such as L. interrogans and L. noguchii, belong to the P1hv group, whereas species with no/unknown pathogenic potential cluster to the P1lv group (Vincent et al., 2019). Although the differences between the individual subclades are clear on the genome level, additional knowledge on phenotypic differences is warranted to fully understand the pathogenesis of leptospirosis. Here we examined clade-specific differences in the structure of lipid A, the hallmark virulence factor of bacterial pathogens.
The negative ion mass spectra of lipid A from 31 Leptospira species were examined by FLAT (Sorensen et al., 2020). This included five P1hv, five P1lv, nine P2, nine S1, and three S2 species (Table 1). In total, 10 different lipid A profiles denoted L1-L10 were detected across the examined species (Figure 1; Table 2). In the P1hv group, L. interrogans, L. noguchii, and L. weilii shared the L1 profile (Figure 1A), and L2 and L3 profiles were detected in L. mayottensis and L. santarosai, respectively (Figures 1B,C). The P1lv group was homogenous; all P1lv species shared the L4 profile (Figure 1D). In the P2 subclade, 7 out of the 9 species shared the L5 lipid A profile with L. licerasiae (Figure 1E). The L6 profile was detected in L. fluminis (Figure 1F) and the L7 profile in L. perolatii (Figure 1G). The lipid A profiles of S1 and S2 species were very similar to each other. Seven out of nine S1 species shared the L8 profile with the model saprophytic species L. biflexa (Figure 1H). L. noumeaensis and L. kanakyensis displayed the L9 phenotype (Figure 1I). Finally, the L10 profile was detected in all S2 species (Figure 1J). The individual lipid A profiles of all examined Leptospira species can be found in Supplementary material (Supplementary Figures S1–S3).
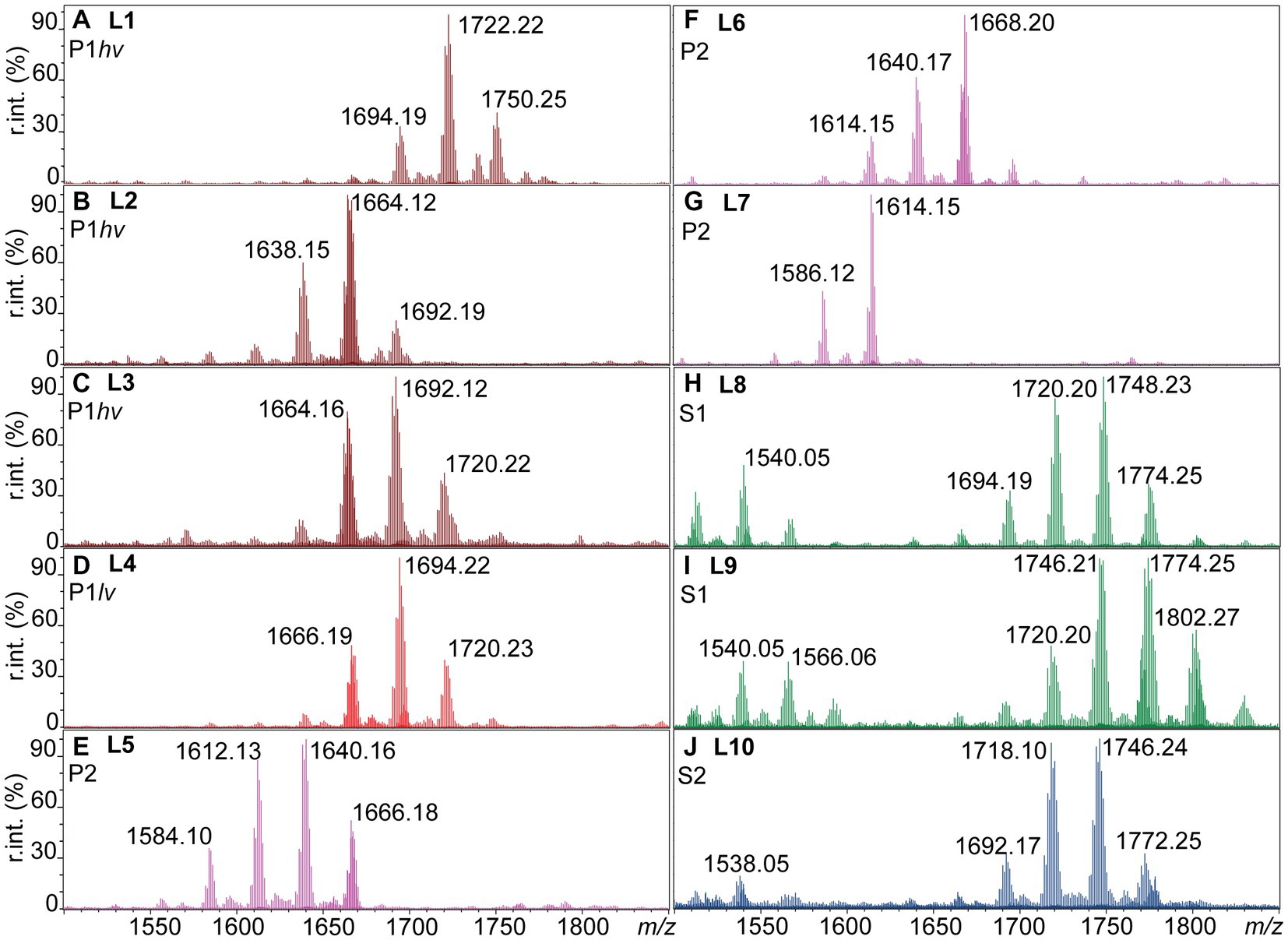
Figure 1. Ten lipid A profiles identified within the individual Leptospira subclades (L1-L10). One microliter of logarithmic cell suspensions in MS-grade water was spotted on a MALDI plate and subjected to FLAT (Sorensen et al., 2020). MS1 scans were acquired in the negative ion mode, relative intensities (r. int.) are shown. (A–C) P1hv group (dark red). (D) P1lv group (light red). (E–G) P2 subclade (purple). (H,I) S1 subclade (green). (J) S2 subclade (blue). Lipid A profiles of the individual Leptospira species can be found in Supplementary Figures S1–S3, and the information is summarized in Table 2.
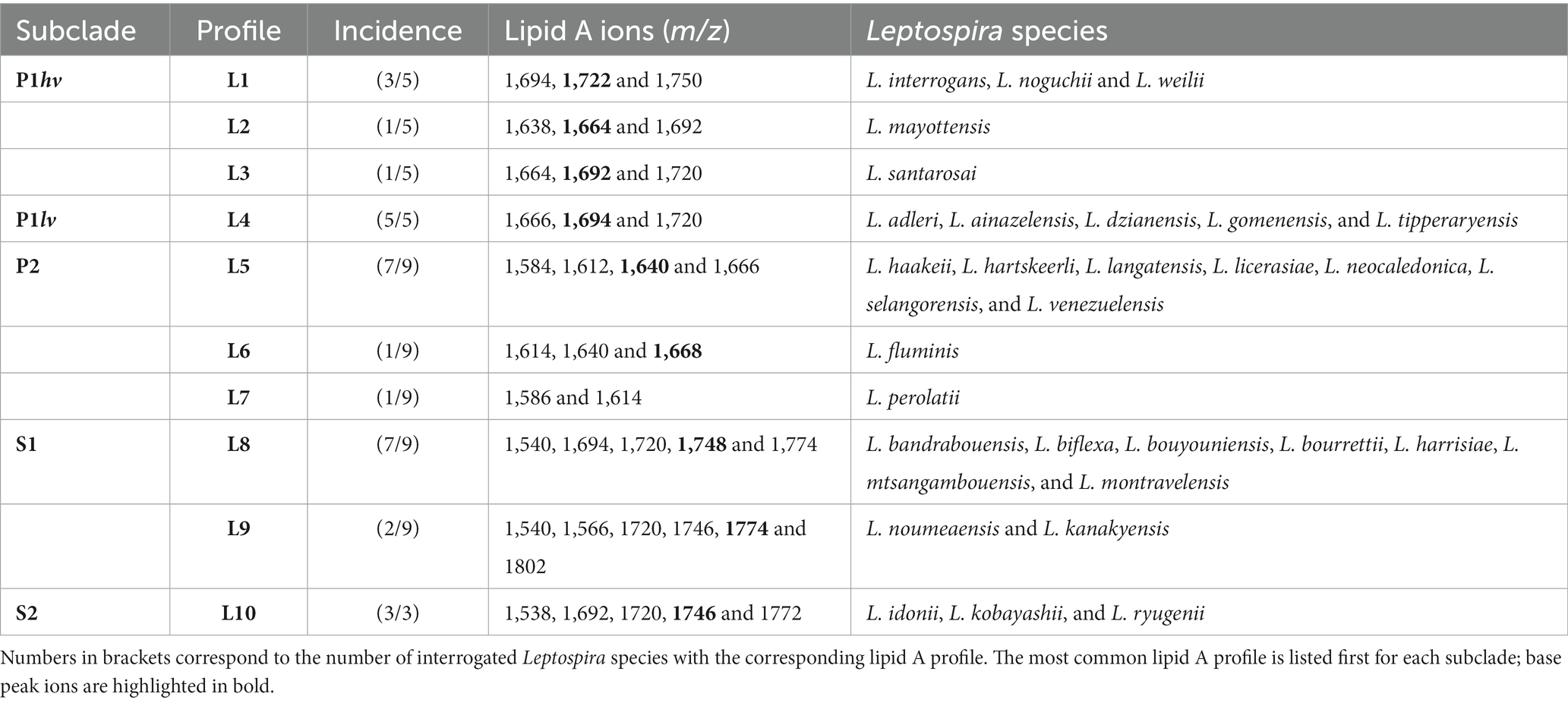
Table 2. Lipid A profiles identified in the individual subclades.
There was no obvious association between the origin of the examined Leptospira species and their lipid A profiles. For example, all but one examined P1lv species and all S1 species were isolated from water and soil environments (Table 1), yet their lipid A profiles were different (Figure 1). Presence of lipid A modifications that could aid survival of Leptospira in water and soil environments cannot be excluded. However, environment-induced lipid A modifications are often transient (Rebeil et al., 2004; Li et al., 2012) and unlikely to be carried over to bacteria grown under conditions where these modifications are not required. At the growth conditions used in this study (modified EMJH T80/T40/LH, 30°C, and shaking), the strongest association was observed between the lipid A profiles of the individual Leptospira species and their phylogenetic classification (Figure 1; Table 2).
The structures of the representative lipid A ions of each profile (L1–L10) were proposed based on tandem mass spectrometry analysis (FLATn) (Yang et al., 2022a). The lipid A structure of L. interrogans (L1) corresponded to the previously reported structure for this species (Que-Gewirth et al., 2004; Eshghi et al., 2015), validating our methodology (Figures 2A,B). Interpretation of lipid A profiles can be challenging. However, one main lipid A ion is usually surrounded by satellite molecules resulting from substantial modifications to this lipid A molecule (such as addition of a sugar moiety or a terminal phosphate group) (Leung et al., 2017; Liang et al., 2019). In contrast, all Leptospira lipid A profiles were complex with several clusters of lipid A ions separated by 26 or 28 Da (Figures 1, 2E). These mass differences corresponded to an addition of two carbons connected by a double bond or a single bond, respectively, and were previously described in L. interrogans and L. kirschneri (Novak et al., 2022) (Figure 2E). Each of these clusters was further predicted to consist of five individual lipid A ions separated by 2 Da (a double bond), revealing an unusual lipid A heterogeneity within a single bacterial species (Figure 2E). Briefly, if only a single lipid A ion was present, the isotopic distribution would look as depicted in Figure 2C. Instead, the measured isotopic distribution in each lipid A cluster (Figure 2E) closely corresponded to a mixed isotopic distribution consisting of five lipid A ions differentiated by a presence of a double bond (Figure 2D). Mass spectrometry-based strategies to locate positions of double bonds in unsaturated lipid molecules exist. They include chemical derivatization prior mass spectrometry analysis, and are yet to be tested on complex mixtures of lipid A molecules detected in Leptospira species (Figure 1E; Novak et al., 2022). Alternatively, proposed lipid A structures can be supported with other analytical techniques, such as nuclear magnetic resonance (NMR). However, dissolving lipid A in NMR-compatible solvents is challenging due to its amphipathic nature (Ribeiro et al., 1999; Zähringer et al., 2001; Silipo et al., 2002). The NMR approach is therefore more appropriate for characterization of the water-soluble components of LPS (core oligosaccharide and O-antigen). Both above-mentioned strategies require pure lipid A extracts from large volume of Leptospira culture, chemical derivatization reagents and rigorous method optimization for complex lipid A samples. Localization of double bonds was therefore not possible within the scope of this study. Like others (Eshghi et al., 2015; Novak et al., 2022) we therefore proposed structures of the representative lipid A for each of the lipid A profiles (L1–L10), and concluded that additional degrees of unsaturation were present (Figure 3). It is important to note that our approach allowed us to obtain valid structural information on Leptospira lipid A from an equivalent of 107 cells (approximately 100 μL of exponential culture). Experiments were therefore performed in a controlled manner, using biological triplicates on two independent experimental days. The low amount of starting material does not affect the results. Lipid A structures of Pseudomonas aeruginosa, Acinetobacter baumannii, and Klebsiella pneumoniae proposed by FLATn corresponded to those determined by other methodologies (Yang et al., 2022a). Here we also validated FLATn on lipid A of L. interrogans serovar Manilae strain L495 (Figure 1A; Eshghi et al., 2015).
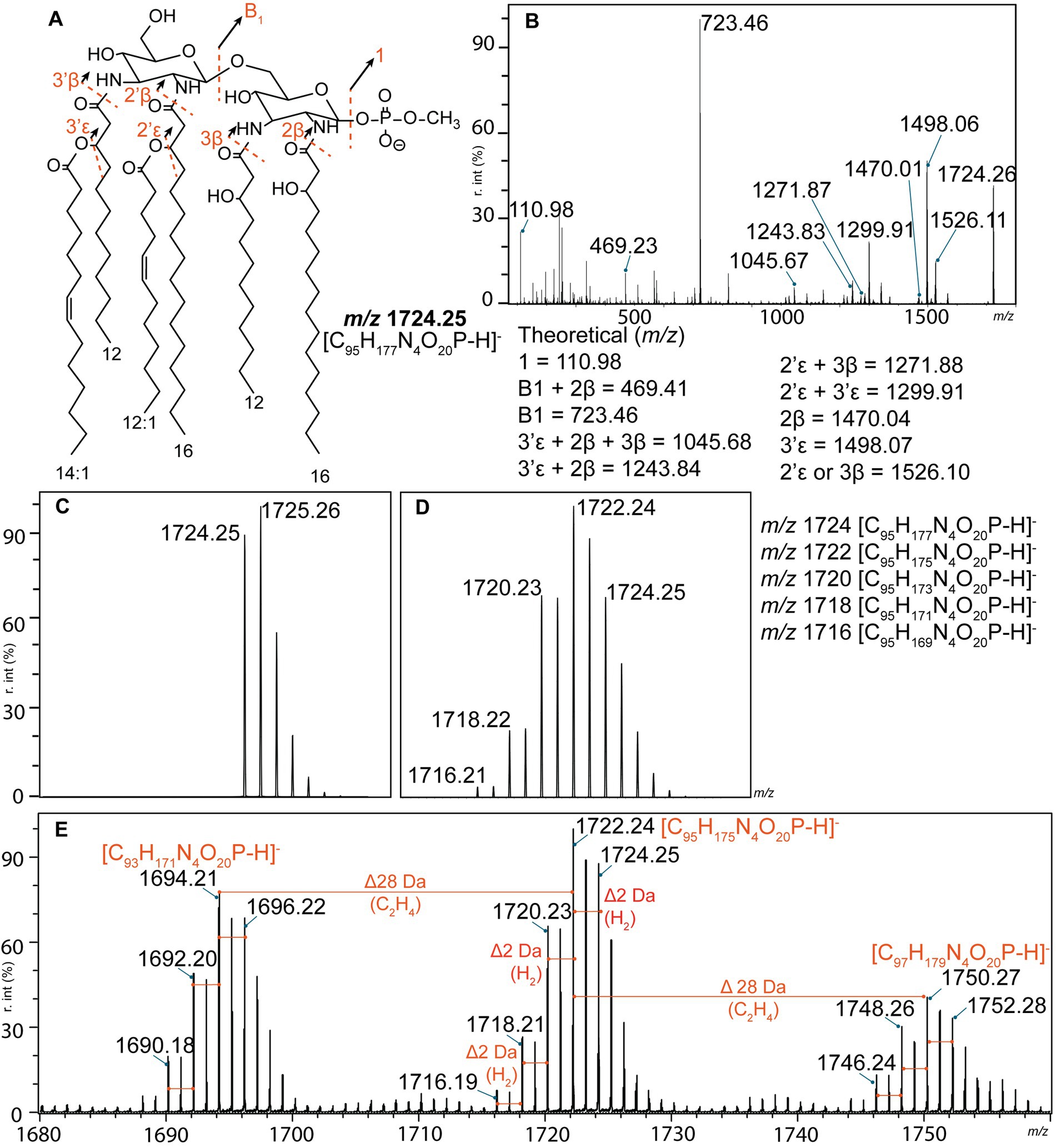
Figure 2. Structural determination of L. interrogans lipid A (L1) by tandem mass spectrometry. (A) Proposed structure of the m/z 1724.25 ion corresponds to the previously published structure for this species (Que-Gewirth et al., 2004; Eshghi et al., 2015). Fragmentation patterns are depicted as dashed red lines. (B) Product ion scan of the precursor ion m/z 1724.25. Calculated m/z of the product ions are listed at the bottom of the panel. (C,D) Theoretical isotopic distributions. (C) Isotopic distribution of a single lipid A ion (m/z 1724.25 corresponding to the structure in panel A). (D) Mixed isotopic distribution of five lipid A ions that differ from each other by a presence of a single double bond (2 Da). Please note that the abundances of each ion were not equal; ratio used for the simulation was 9:21:17:6:1 (m/z 1724:1722:1720:1718:1716). (E) Annotated mass spectrum of L. interrogans lipid A profile (L1). Three main clusters of lipid A ions separated by 28 Da were identified. Each individual cluster likely consisted of five lipid A ions that differ by a presence of a double bond (red lines). r. int. – relative intensity.
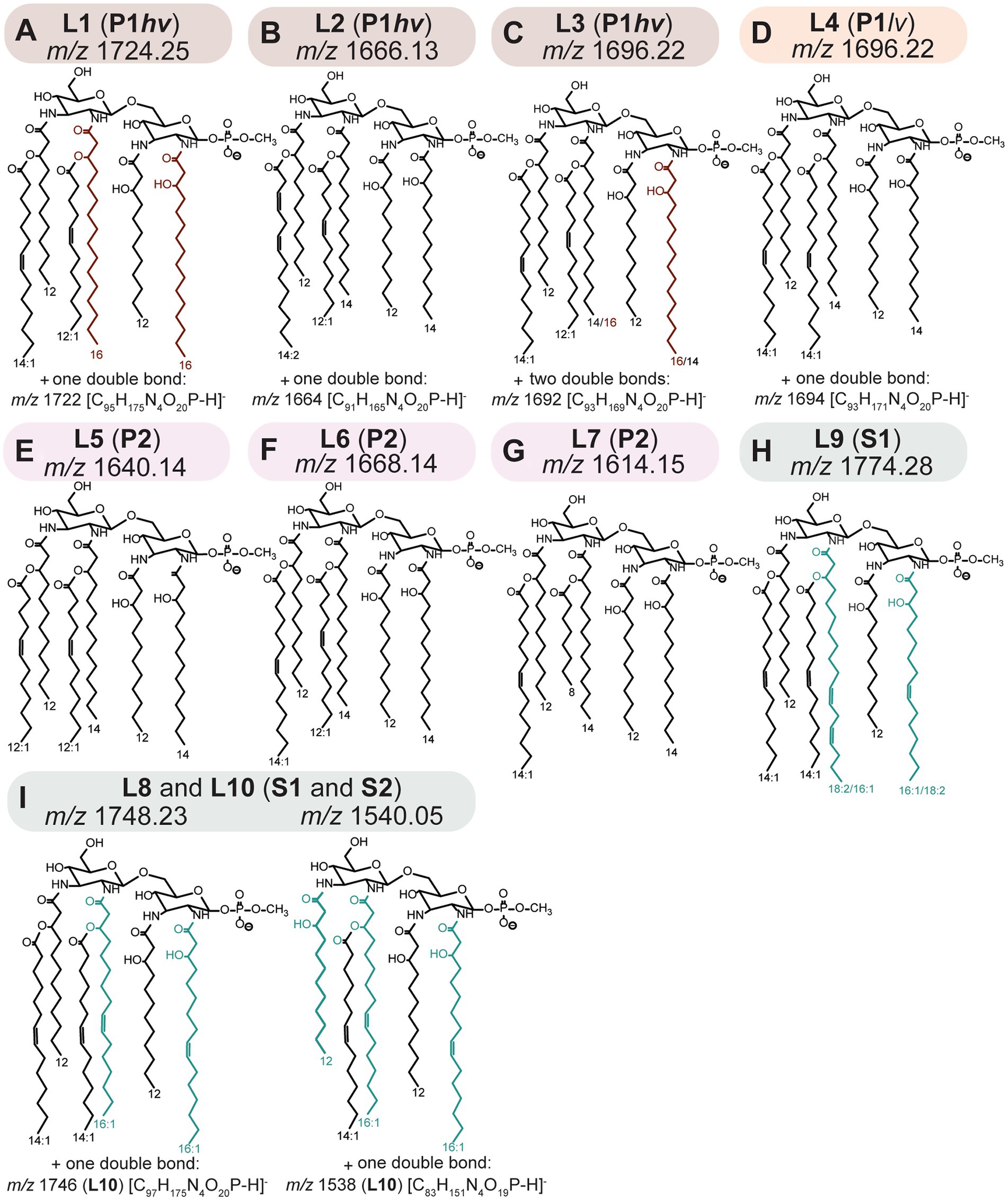
Figure 3. (A–C) L1-L3; P1hv group. (D) L4; P1lv group. (E–G) L5-L7; P2 subclade. (H) L9; S1 subclade. (I) L8 and L10; S1 and S2 subclades. Putative structures of lipid A ions representative of the L1–L10 profiles. Structures were proposed based on fragmentation patterns of the representative lipid A ions, as detected by FLATn (Yang et al., 2022a; Supplementary Figures S4–S6). Sugar and anomeric configurations were assigned in homology with previous data (Que-Gewirth et al., 2004; Eshghi et al., 2015; Novak et al., 2022) and the placement of double bonds is putative. Additional approaches discussed in Section 3.2 are needed to fully validate the proposed structures (out of the scope of the current study). Please refer to the Table 2 for information on distribution of the individual phenotype across Leptospira species.
Tandem MS analysis revealed that some previously described Leptospira lipid A features (Que-Gewirth et al., 2004; Eshghi et al., 2015) were conserved in all 31 examined species. Specifically, representative lipid A molecules were hexa-acylated and monophosphorylated, with all primary fatty acyl residues linked via amide bonds and a methylated terminal phosphate group present at 1 position of the di-aminoglucose backbone (Figure 3). A newly discovered conserved feature was the presence of C12:0 (OH) residues at the 3 and 3′ primary positions (Figure 3).
The structural variability of lipid A between the Leptospira species was determined by the length and saturation of 2 and 2′ primary acyl and 2′ and 3′ secondary acyl chains (Figure 3). The identity of 2 primary acyl chains could be deducted from the main product B1 ion that results from fragmentation of the bond connecting the glucosamine backbone (Figures 2A,B). The three main B1 ions identified in this study were m/z 695, m/z 721 and m/z 723 indicating acylation of the phosphorylated sugar unit (GlcN I) at position 2 with C14:0 (OH), C16:1 (OH) and C16:0 (OH), respectively (Supplementary Figures S4–S6). The 2 and 2′ primary acyl chains are usually identical, which stems from the mechanism of the lipid A biosynthesis (Raetz et al., 2009; Simpson and Trent, 2019). Lipid A is synthesized in a series of conserved reactions mediated by the family of Lpx enzymes; homologs of most Lpx enzymes were identified across Leptospira species (Hinckley et al., 2005; Eshghi et al., 2015; Nieves et al., 2023). In the early steps, LpxA, LpxC, and LpxD produce a molecule of uridine phosphate (UDP)-2,3-diacylglucosamine from UDP-N-acetylglucosamine and fatty acids bound to acyl carrier proteins. While LpxA is responsible for the addition of a fatty acyl to the 3 primary position, LpxD adds a fatty acyl to the 2 primary position of the glucosamine backbone. Both LpxA and LpxD have affinity toward specific fatty acyl chains, and this affinity differs across bacterial species (Simpson and Trent, 2019). Subsequently, a molecule of “lipid X” (2,3-diacylglucosamine-1-phoshate) is produced from some UDP-2,3-diacylglucosamine precursors via activity of LpxH or its homologs LpxI or LpxG. One UDP-2,3-diacylglucosamine and one “lipid X” molecule are then condensed together via the activity of LpxB, resulting in identical acyl chains in the 2 and 2′ and in the 3 and 3′ primary positions. As follows, each of the individual Leptospira subclades had a predominant primary acyl chain at the 2 and 2′ primary positions: C16:0 (OH) acyls were detected exclusively in P1hv species (Figures 3A,C), C14:0 (OH) in all P1lv and P2 species (Figures 3D–G) and C16:1 (OH) in all S1 and S2 species (Figures 3H,I). This was consistent with a previous study where C16 (OH) were detected exclusively in the pathogenic L. interrogans (Patra et al., 2015). Lipid A of L. mayottensis incorporated two C14:0 (OH) as the 2 and 2′ primary residues and its lipid A therefore resembled those of the P1lv and P2 species (Figure 3B, L2 profile).
Interestingly, fragmentation of the L3 and L9 representative lipid A ions resulted in two B1 ions instead of one (Supplementary Figures S4D, S6F). In L3 (L. santarosai), the two B1 ions m/z 695 and m/z 723 were detected, suggesting that the 2 and 2′ primary acyl chains were interchangeable, creating two possible isomers. A combination of C14:0 (OH)/C16:0 (OH) at the 2/2′ positions resulted in the m/z 695 B1 product ion, while the opposite configuration, C16:0 (OH)/C14:0 (OH) at the 2/2′ positions, resulted in the m/z 723 B1 ion (Figure 3C). The lipid A profile of this strain was also the most complex one with two extra double bonds in the base lipid A ion that could not be localized using the MS data alone (Figure 3C). In L9 (L. kanakyensis and L. noumeaensis), m/z 721 and m/z 747 were detected, likely resulting from combinations of C16:1 (OH)/C18:2 (OH) and C18:2 (OH)/C16:1 (OH) at the 2/2′ primary positions, respectively (Figure 3H). These unusual lipid A structures could be a result of simultaneous activity of two LpxD enzymes, as two copies of lpxD genes have been annotated in all Leptospira genomes except for those belonging to the P2 subclade (Supplementary Figure S7). To date, the function of two separate LpxD enzymes was studied only in two bacterial species (Simpson and Trent, 2019). In Francisella, the expression of LpxD1 and LpxD2 is temperature dependent. LpxD1 adds two C18:0 (OH) and LpxD2 adds two C16:0 (OH) to the 3 and 3′ primary positions of the lipid A when grown at 37°C and 25°C, respectively, aiding adaptation to temperature shifts (Gunn and Ernst, 2007; Scott et al., 2016). In L. interrogans, LpxD1 contributes to pathogenicity, adaptation to temperature changes and presence of toxic compounds (Eshghi et al., 2015). However, the conditions warranting expression of LpxD1/LpxD2 in L. interrogans remain elusive (Eshghi et al., 2015; Simpson and Trent, 2019). The representative structures of L3 and L9 phenotypes might provide a first hint to function of LpxD1 and LpxD2 in other Leptospira species. It occurs that Leptospira species with the L3 and the L9 phenotypes, co-expressed LpxD1 and LpxD2 enzymes might compete to add acyl residues to the 2 primary position of the early UDP-2,3-diacyl glucosamine product in lipid A biosynthesis. A similar phenomenon was described for the late acetyltransferases LpxL1 and LpxL2 in Klebsiella pneumoniae that compete to add either C12:0 or C14:0 at the 2′ secondary position (Li et al., 2016; Mills et al., 2017; Simpson and Trent, 2019). Annotated tandem mass spectra for all representative lipid A ions can be found in the supplementary material (Supplementary Figures S4–S6).
All Leptospira species incorporated short fatty acyl chains at the 2′ and 3′ secondary positions of their lipid A. In the representative lipid A molecules, these residues consisted of a combination of C12:1/C14:1 (L1, L3 and L6; Figures 3A,C,F), two C12:1 (L5; Figure 3E) or two C14:1 (L4, L8-10; Figures 3D,H,I). Representative lipid A molecules of L. mayottensis (L2; Figure 3B) and L. perolatii (L7; Figure 3G) contained a combination of C14:2/C12:1 and C14:1/C8:0, respectively (Figure 3). Secondary acyl residues are added to the lipid A by late acyltransferases (homologs of LpxL and LpxM from Escherichia coli) (Raetz et al., 2009). Each of these enzymes often adds an acyl chain of a specific length and degree of saturation (Simpson and Trent, 2019). To date, only one bi-functional acyltransferase capable of adding two different acyl chains to the 2′ and 3′ secondary positions was reported in Acinetobacter baumannii (Boll et al., 2015). Given the great variability of secondary acyl chains across Leptospira, its LpxL homolog is likely a multifunctional acyltransferase.
A novel structural feature of Leptospira lipid A was found in S1 and S2 subclades. In these species, additional clusters with lower m/z were identified (Figure 1; Supplementary Figure S3). Upon tandem mass spectrometry analysis, it was determined that these were penta-acylated lipid A molecules (Figure 3; Supplementary Figure S6). The mechanisms of synthesis of these penta-acylated lipid A species are unclear. In other bacteria, fatty acyl chains can be removed via activity of LpxR (Simpson and Trent, 2019). Although homologs of LpxR were identified in Leptospira, the LpxR usually removes two acyl chains, not one. PagL and PagP enzymes can remove a single acyl chain from the lipid A molecule (Ernst et al., 2006; Thaipisuttikul et al., 2014), however, homologs of these enzymes were not found in saprophytic Leptospira (Picardeau et al., 2008). Finally, in bacteria harboring two LpxL enzymes, such as Neisseria meningitidis, loss of one copy leads to synthesis of penta-acylated lipid A species (Fransen et al., 2009). This cannot be the case in Leptospira where only one LpxL homolog was annotated (Picardeau et al., 2008). Nonetheless, penta-acylated lipid A molecules are known to elicit reduced immune responses in the host (Fransen et al., 2010; Scott et al., 2017), and their presence in saprophytic species is intriguing.
Lipid A of pathogenic Leptospira has been previously shown to evade recognition by the human TLR4/MD2 (Werts et al., 2001). It has been speculated that this might be due to monophosphorylation of Leptospira lipid A that is associated with a reduced endotoxic activity in other bacteria (Baldridge and Crane, 1999; Wang et al., 2007). However, this is complicated by the unusual presence of a methyl group on the single terminal phosphate (Que-Gewirth et al., 2004). Here, we revealed other structural features that might contribute to this phenomenon.
Degree of TLR4/MD2 activation is also dependent on the length of fatty acyl residues. While C12 or C14 are optimal for TLR4/MD2 binding, C16 is not favorable (Rietschel et al., 1994; Park and Lee, 2013; Facchini et al., 2018). In deep-sea Moritella species, the lipid A either activates TLR4/MD2 or is “immune-silent,” not eliciting responses via TLR4/MD2 or other related host receptors (Gauthier et al., 2021). While the basic structural features between Moritella lipid As are conserved (hexa-acylated bis-phosphorylated molecules), the immune-silent Moritella lipid A has higher C16 content (Gauthier et al., 2021). In this study, C16 (OH) residues were found exclusively in pathogenic P1hv species (Figures 3A,C), which was consistent with previous findings (Patra et al., 2015). We therefore hypothesize that lipid A of P1lv and P2 species might be better binding partners of the innate immune receptors, contributing to faster clearance of these species and their lower pathogenic potential in humans. Future studies including assessing endotoxin activity of P1lv and P2 lipid A extracts using reporter assays are warranted to explore this hypothesis.
Finally, while the discussion to this point has centered around lipid A, other lipid molecules are known to confer immune evasion. Cardiolipins have been shown to suppress stimulatory activity of LPS (Khan et al., 2018). Cardiolipin species have been identified in pathogenic as well as non-pathogenic Leptospira species (Supplementary Figure S8). Since our lipid preparations for FLAT and FLATn consisted of whole cells, it is not possible to determine if the cardiolipins were located to the inner or the outer membrane and if they can attenuate LPS-mediated immune activation. However, their presence in the Leptospira membrane is intriguing and warrants further investigation.
Novel Leptospira species are isolated from various hosts or the environment on regular basis (Thibeaux et al., 2018; Masuzawa et al., 2019b; Korba et al., 2021). Extensive phenotype profiling including serotyping, assessing growth at 37°C, growth in presence of purine analog 8-azaguanine, and ultimately animal infection studies are needed to distinguish pathogens from saprophytes during characterization of novel species (Vincent et al., 2019). Here, we propose the use of L1-L10 lipid A profiles combined with FLAT for rapid classification of Leptospira isolates into the individual subclades (L1-L3 for P1hv, L4 for P1lv, L5-L7 for P2, L8-9 for S1 and L10 for S2 subclades). Lipid A-based MALDI-TOF assays allow for rapid (within an hour) identification of bacteria directly from a specimen using minimal input and hands-on-time (Leung et al., 2017; Liang et al., 2019; Sorensen et al., 2020). Lipid A-based assays allow for simultaneous identification and screening for antibiotic resistance markers and can be used directly from urine (Smith et al., 2021, 2022; Yang et al., 2022b). Thanks to minimal background in the m/z area where lipid A is detected, individual species can also be identified from multi-bacterial samples (Fondrie et al., 2018; Ryu et al., 2020). Protein-based profiling via MALDI-TOF is routinely used to characterize Leptospira species (Thibeaux et al., 2018; Sonthayanon et al., 2019; Girault et al., 2020; Korba et al., 2021), and the addition of lipid A phenotyping would provide valuable information while utilizing the existing infrastructure.
This is the first study focused on structural analysis of lipid A across the whole Leptospira genus. Ten distinct lipid A profiles were revealed that can be used for rapid molecular typing of novel clinical and environmental Leptospira isolates, aiding the leptospirosis surveillance. In addition, revealed structural differences between lipid A of individual species can lead to novel hypotheses on Leptospira pathogenicity.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding authors.
HP conceived and designed the experiments. HP, AM, SC, AG-G, and MS performed the experiments. HP, AM, and DG analyzed the data. AG, RE, CC, MP, and DG contributed reagents, materials, and analysis tools. HP and DG prepared the original draft. HP, AM, SC, AG-G, AG, MS, RE, CC, MP, and DG reviewed and edited the manuscript. All authors contributed to the article and approved the submitted version.
Funding for this study was provided to DRG from Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant (RGPIN-2022-04433). DG and RE received funding from National Institutes of Health (NIH) and National Institute of Allergy and Infectious Diseases (NIAID) #R01AI147314. DG and CC received support from the Canada Institutes of Health Research (CIHR), Catalyst Grant Sexually Transmitted and Blood Borne Infections Research in Canada: Beyond HIV/AIDS and Hepatitis (SC1–178732). DG and the work performed at the University of Victoria-Genome BC Proteomics Centre was also supported by funding to the Metabolomics Innovation Centre (TMIC) from Genome Canada and Genome British Columbia, through the Genomics Technology Platform (GTP) program for operations and technology development (265MET), as well operations support from the Canadian Foundation for Innovation Major Sciences Initiative (CFI-MSI) program (35456). Infrastructure and operations funding to support his project was provided to DRG from PacifiCan (Project No: 22591). MP was supported by Institut Pasteur.
We thank Darryl Hardie and Jun Han of the UVic Genome BC Proteomic Centre for their assistance with the LC–MS/MS assays. We thank Angela Jackson of the UVic Genome BC Proteomic Centre for her support and managing the laboratory’s day-to-day operations. We also thank Dr. Yuri O. Tsybin of Spectroswiss for assistance with isotopic distribution modeling.
RE and DG are co-founders and vice presidents of Patagain, a company that develops mass spectrometry-based microbiology tests to identify disease pathogens and determine antimicrobial resistance.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2023.1181034/full#supplementary-material
Baldridge, J. R., and Crane, R. T. (1999). Monophosphoryl lipid A (MPL) formulations for the next generation of vaccines. Methods 19, 103–107. doi: 10.1006/meth.1999.0834
Boll, J. M., Tucker, A. T., Klein, D. R., Beltran, A. M., Brodbelt, J. S., Davies, B. W., et al. (2015). Reinforcing lipid A acylation on the cell surface of Acinetobacter baumannii promotes cationic antimicrobial peptide resistance and desiccation. Survival 6, e00478–e00415. doi: 10.1128/mBio.00478-15
Bourhy, P., Collet, L., Brisse, S., and Picardeau, M. (2014). Leptospira mayottensis sp. nov., a pathogenic species of the genus Leptospira isolated from humans. Int. J. Syst. Evol. Microbiol. 64, 4061–4067. doi: 10.1099/ijs.0.066597-0
Chandler, C. E., Harberts, E. M., Pelletier, M. R., Thaipisuttikul, I., Jones, J. W., Hajjar, A. M., et al. (2020). Early evolutionary loss of the lipid A modifying enzyme PagP resulting in innate immune evasion in Yersinia pestis. Proc. Natl. Acad. Sci. U. S. A. 117, 22984–22991. doi: 10.1073/pnas.1917504117
Coburn, J., Picardeau, M., Woods, C. W., Veldman, T., and Haake, D. A. (2021). Pathogenesis insights from an ancient and ubiquitous spirochete. PLoS Pathog. 17:e1009836. doi: 10.1371/journal.ppat.1009836
Deguchi, A., Tomita, T., Ohto, U., Takemura, K., Kitao, A., Akashi-Takamura, S., et al. (2016). Eritoran inhibits S100A8-mediated TLR4/MD-2 activation and tumor growth by changing the immune microenvironment. Oncogene 35, 1445–1456. doi: 10.1038/onc.2015.211
Domínguez-Medina, C. C., Pérez-Toledo, M., Schager, A. E., Marshall, J. L., Cook, C. N., Bobat, S., et al. (2020). Outer membrane protein size and LPS O-antigen define protective antibody targeting to the Salmonella surface. Nat. Commun. 11:851. doi: 10.1038/s41467-020-14655-9
Duerr, C. U., Zenk, S. F., Chassin, C., Pott, J., Gütle, D., Hensel, M., et al. (2009). O-antigen delays lipopolysaccharide recognition and impairs antibacterial host defense in murine intestinal epithelial cells. PLoS Pathog. 5:e1000567. doi: 10.1371/journal.ppat.1000567
Ellis, W. A., and Thiermann, A. B. (1986). Isolation of Leptospira interrogans serovar Bratislava from sows in Iowa. Am. J. Vet. Res. 47, 1458–1460.
Ernst, R. K., Adams, K. N., Moskowitz, S. M., Kraig, G. M., Kawasaki, K., Stead, C. M., et al. (2006). The Pseudomonas aeruginosa lipid A deacylase: selection for expression and loss within the cystic fibrosis airway. J. Bacteriol. 188, 191–201. doi: 10.1128/JB.188.1.191-201.2006
Eshghi, A., Henderson, J., Trent, M. S., and Picardeau, M. (2015). Leptospira interrogans lpxD homologue is required for thermal acclimatization and virulence. Infect. Immun. 83, 4314–4321. doi: 10.1128/IAI.00897-15
Facchini, F. A., Zaffaroni, L., Minotti, A., Rapisarda, S., Calabrese, V., Forcella, M., et al. (2018). Structure-activity relationship in monosaccharide-based toll-like receptor 4 (TLR4) antagonists. J. Med. Chem. 61, 2895–2909. doi: 10.1021/acs.jmedchem.7b01803
Fondrie, W. E., Liang, T., Oyler, B. L., Leung, L. M., Ernst, R. K., Strickland, D. K., et al. (2018). Pathogen identification direct from Polymicrobial specimens using membrane glycolipids. Sci. Rep. 8:15857. doi: 10.1038/s41598-018-33681-8
Fransen, F., Hamstra, H. J., Boog, C. J., van Putten, J. P., van den Dobbelsteen, G. P. J. M., and van der Ley, P. (2010). The structure of Neisseria meningitidis lipid A determines outcome in experimental meningococcal disease. Infect. Immun. 78, 3177–3186. doi: 10.1128/IAI.01311-09
Fransen, F., Heckenberg, S. G. B., Hamstra, H. J., Feller, M., Boog, C. J. P., van Putten, J. P. M., et al. (2009). Naturally occurring lipid A mutants in Neisseria meningitidis from patients with invasive meningococcal disease are associated with reduced coagulopathy. PLoS Pathog. 5:e1000396. doi: 10.1371/journal.ppat.1000396
Gauthier, A. E., Chandler, C. E., Poli, V., Gardner, F. M., Tekiau, A., Smith, R., et al. (2021). Deep-sea microbes as tools to refine the rules of innate immune pattern recognition. Sci. Immunol. 6:eabe0531. doi: 10.1126/sciimmunol.abe0531
Girault, D., Kainiu, M., Barsac, E., Thibeaux, R., and Goarant, C. (2020). Use of MALDI-ToF mass spectrometry for identification of Leptospira. Methods Mol. Biol. 2134, 23–29. doi: 10.1007/978-1-0716-0459-5_3
Gunn, J. S., and Ernst, R. K. (2007). The structure and function of Francisella lipopolysaccharide. Ann. N. Y. Acad. Sci. 1105, 202–218. doi: 10.1196/annals.1409.006
Hassan, N., Anesio, A. M., Rafiq, M., Holtvoeth, J., Bull, I., Haleem, A., et al. (2020). Temperature driven membrane lipid adaptation in glacial psychrophilic Bacteria. Front. Microbiol. 11:824. doi: 10.3389/fmicb.2020.00824
Hinckley, M. B., Reynolds, C. M., Ribeiro, A. A., McGrath, S. C., Cotter, R. J., Lauw, F. N., et al. (2005). A Leptospira interrogans enzyme with similarity to yeast Ste14p that methylates the 1-phosphate group of lipid A. J. Biol. Chem. 280, 30214–30224. doi: 10.1074/jbc.M506103200
Kawahara, K. (2021). Variation, modification and engineering of lipid A in endotoxin of gram-negative Bacteria. Int. J. Mol. Sci. 22:2281. doi: 10.3390/ijms22052281
Kawahara, K., Tsukano, H., Watanabe, H., Lindner, B., and Matsuura, M. (2002). Modification of the structure and activity of lipid A in Yersinia pestis lipopolysaccharide by growth temperature. Infect. Immun. 70, 4092–4098. doi: 10.1128/IAI.70.8.4092-4098.2002
Khan, M. M., Ernst, O., Sun, J., Fraser, I. D. C., Ernst, R. K., Goodlett, D. R., et al. (2018). Mass spectrometry-based structural analysis and systems Immunoproteomics strategies for deciphering the host response to endotoxin. J. Mol. Biol. 430, 2641–2660. doi: 10.1016/j.jmb.2018.06.032
Koizumi, N., and Watanabe, H. (2003). Identification of a novel antigen of pathogenic Leptospira spp. that reacted with convalescent mice sera. J. Med. Microbiol. 52, 585–589. doi: 10.1099/jmm.0.05148-0
Kong, Q., Six, D. A., Liu, Q., Gu, L., Roland, K. L., Raetz, C. R. H., et al. (2011). Palmitoylation state impacts induction of innate and acquired immunity by the Salmonella enterica serovar typhimurium msbB mutant. Infect. Immun. 79, 5027–5038. doi: 10.1128/IAI.05524-11
Kong, Q., Six, D. A., Liu, Q., Gu, L., Wang, S., Alamuri, P., et al. (2012). Phosphate groups of lipid A are essential for Salmonella enterica serovar Typhimurium virulence and affect innate and adaptive immunity. Infect. Immun. 80, 3215–3224. doi: 10.1128/IAI.00123-12
Korba, A. A., Lounici, H., Kainiu, M., Vincent, A. T., Mariet, J.-F., Veyrier, F. J., et al. (2021). Leptospira ainlahdjerensis sp. nov., Leptospira ainazelensis sp. nov., Leptospira abararensis sp. nov. and Leptospira chreensis sp. nov., four new species isolated from water sources in Algeria. Int. J. Syst. Evol. Microbiol. 71:005148. doi: 10.1099/ijsem.0.005148
Leung, L. M., Fondrie, W. E., Doi, Y., Johnson, J. K., Strickland, D. K., Ernst, R. K., et al. (2017). Identification of the ESKAPE pathogens by mass spectrometric analysis of microbial membrane glycolipids. Sci. Rep. 7:6403. doi: 10.1038/s41598-017-04793-4
Li, Y., Powell, D. A., Shaffer, S. A., Rasko, D. A., Pelletier, M. R., Leszyk, J. D., et al. (2012). LPS remodeling is an evolved survival strategy for bacteria. Proc. Natl. Acad. Sci. U. S. A. 109, 8716–8721. doi: 10.1073/pnas.1202908109
Li, Y., Yun, J., Liu, L., Li, Y., and Wang, X. (2016). Identification of two genes encoding for the late acyltransferases of lipid A in Klebsiella pneumoniae. Curr. Microbiol. 73, 732–738. doi: 10.1007/s00284-016-1117-6
Liang, T., Leung, L. M., Opene, B., Fondrie, W. E., Lee, Y. I., Chandler, C. E., et al. (2019). Rapid microbial identification and antibiotic resistance detection by mass spectrometric analysis of membrane lipids. Anal. Chem. 91, 1286–1294. doi: 10.1021/acs.analchem.8b02611
Marcsisin, R. A., Bartpho, T., Bulach, D. M., Srikram, A., Sermswan, R. W., Adler, B., et al. (2013). Use of a high-throughput screen to identify Leptospira mutants unable to colonize the carrier host or cause disease in the acute model of infection. J. Med. Microbiol. 62, 1601–1608. doi: 10.1099/jmm.0.058586-0
Masuzawa, T., Saito, M., Nakao, R., Nikaido, Y., Matsumoto, M., Ogawa, M., et al. (2019a). Molecular and phenotypic characterization of Leptospira johnsonii sp. nov., Leptospira ellinghausenii sp. nov. and Leptospira ryugenii sp. nov. isolated from soil and water in Japan. Microbiol. Immunol. 63, 89–99. doi: 10.1111/1348-0421.12671
Masuzawa, T., Uno, R., Matsuhashi, N., Yamaguchi, M., Xu, J., and Nakamura, S. (2019b). Comparison of Leptospira species isolated from environmental water and soil in Japan. Microbiol. Immunol. 63, 469–473. doi: 10.1111/1348-0421.12741
Mills, G., Dumigan, A., Kidd, T., Hobley, L., and Bengoechea, J. A. (2017). Identification and characterization of two Klebsiella pneumoniae lpxL lipid A late Acyltransferases and their role in virulence. Infect. Immun. 85, e00068–e00017. doi: 10.1128/IAI.00068-17
Murray, G. L., Srikram, A., Henry, R., Hartskeerl, R. A., Sermswan, R. W., and Adler, B. (2010). Mutations affecting Leptospira interrogans lipopolysaccharide attenuate virulence. Mol. Microbiol. 78, 701–709. doi: 10.1111/j.1365-2958.2010.07360.x
Nahori, M.-A., Fournié-Amazouz, E., Que-Gewirth, N. S., Balloy, V., Chignard, M., Raetz, C. R. H., et al. (2005). Differential TLR recognition of leptospiral lipid A and lipopolysaccharide in murine and human cells. J. Immunol. 175, 6022–6031. doi: 10.4049/jimmunol.175.9.6022
Nally, J. E., Arent, Z., Bayles, D. O., Hornsby, R. L., Gilmore, C., Regan, S., et al. (2016). Emerging infectious disease implications of invasive mammalian species: the greater white-toothed shrew (Crocidura russula) is associated with a novel Serovar of pathogenic Leptospira in Ireland. PLoS Negl. Trop. Dis. 10:e0005174. doi: 10.1371/journal.pntd.0005174
Needham, B. D., and Trent, M. S. (2013). Fortifying the barrier: the impact of lipid A remodelling on bacterial pathogenesis. Nat. Rev. Microbiol. 11, 467–481. doi: 10.1038/nrmicro3047
Nieves, C., Vincent, A. T., Zarantonelli, L., Picardeau, M., Veyrier, F. J., and Buschiazzo, A. (2023). Horizontal transfer of the rfb cluster in Leptospira is a genetic determinant of serovar identity. Life Sci. Alliance 6:e202201480. doi: 10.26508/lsa.202201480
Novak, A., Pupo, E., Van't Veld, E., VPMG, R., Broere, F., and Sloots, A. (2022). Activation of canine, mouse and human TLR2 and TLR4 by inactivated Leptospira vaccine strains. Front. Immunol. 13:823058. doi: 10.3389/fimmu.2022.823058
Ohto, U., Fukase, K., Miyake, K., and Shimizu, T. (2012). Structural basis of species-specific endotoxin sensing by innate immune receptor TLR4/MD-2. Proc. Natl. Acad. Sci. U. S. A. 109, 7421–7426. doi: 10.1073/pnas.1201193109
Park, B. S., and Lee, J.-O. (2013). Recognition of lipopolysaccharide pattern by TLR4 complexes. Exp. Mol. Med. 45:e66. doi: 10.1038/emm.2013.97
Park, B. S., Song, D. H., Kim, H. M., Choi, B.-S., Lee, H., and Lee, J.-O. (2009). The structural basis of lipopolysaccharide recognition by the TLR4–MD-2 complex. Nature 458, 1191–1195. doi: 10.1038/nature07830
Patra, K. P., Choudhury, B., Matthias, M. M., Baga, S., Bandyopadhya, K., and Vinetz, J. M. (2015). Comparative analysis of lipopolysaccharides of pathogenic and intermediately pathogenic Leptospira species. BMC Microbiol. 15:244. doi: 10.1186/s12866-015-0581-7
Picardeau, M., Bulach, D. M., Bouchier, C., Zuerner, R. L., Zidane, N., Wilson, P. J., et al. (2008). Genome sequence of the saprophyte Leptospira biflexa provides insights into the evolution of Leptospira and the pathogenesis of leptospirosis. PLoS One 3:e1607. doi: 10.1371/journal.pone.0001607
Puche, R., Ferrés, I., Caraballo, L., Rangel, Y., Picardeau, M., Takiff, H., et al. (2018). Leptospira venezuelensis sp. nov., a new member of the intermediate group isolated from rodents, cattle and humans. Int. J. Syst. Evol. Microbiol. 68, 513–517. doi: 10.1099/ijsem.0.002528
Que-Gewirth, N. L. S., Ribeiro, A. A., Kalb, S. R., Cotter, R. J., Bulach, D. M., Adler, B., et al. (2004). A methylated phosphate group and four amide-linked acyl chains in Leptospira interrogans lipid A. the membrane anchor of an unusual lipopolysaccharide that activates TLR2. J. Biol. Chem. 279, 25420–25429. doi: 10.1074/jbc.M400598200
Raetz, C. R. H., Guan, Z., Ingram, B. O., Six, D. A., Song, F., Wang, X., et al. (2009). Discovery of new biosynthetic pathways: the lipid A story. J. Lipid Res. 50, S103–S108. doi: 10.1194/jlr.R800060-JLR200
Rebeil, R., Ernst, R. K., Gowen, B. B., Miller, S. I., and Hinnebusch, B. J. (2004). Variation in lipid A structure in the pathogenic yersiniae. Mol. Microbiol. 52, 1363–1373. doi: 10.1111/j.1365-2958.2004.04059.x
Ribeiro, A. A., Zhou, Z., and Raetz, C. R. H. (1999). Multi-dimensional NMR structural analyses of purified lipid X and lipid A (endotoxin). Magn. Reson. Chem. 37, 620–630. doi: 10.1002/(SICI)1097-458X(199909)37:9<620::AID-MRC517>3.0.CO;2-Q
Ricaldi, J. N., Fouts, D. E., Selengut, J. D., Harkins, D. M., Patra, K. P., Moreno, A., et al. (2012). Whole genome analysis of Leptospira licerasiae provides insight into leptospiral evolution and pathogenicity. PLoS Negl. Trop. Dis. 6:e1853. doi: 10.1371/journal.pntd.0001853
Rietschel, E. T., Kirikae, T., Schade, F. U., Mamat, U., Schmidt, G., Loppnow, H., et al. (1994). Bacterial endotoxin: molecular relationships of structure to activity and function. FASEB J. 8, 217–225. doi: 10.1096/fasebj.8.2.8119492
Ryu, S. Y., Wendt, G. A., Ernst, R. K., and Goodlett, D. R. (2020). MGMS2: membrane glycolipid mass spectrum simulator for polymicrobial samples. Rapid Commun. Mass Spectrom. 34:e8824. doi: 10.1002/rcm.8824
Saito, M., Villanueva, S. Y. A. M., Kawamura, Y., Iida, K.-I., Tomida, J., Kanemaru, T., et al. (2013). Leptospira idonii sp. nov., isolated from environmental water. Int. J. Syst. Evol. Microbiol. 63, 2457–2462. doi: 10.1099/ijs.0.047233-0
Scott, A. J., Flinders, B., Cappell, J., Liang, T., Pelc, R. S., Tran, B., et al. (2016). Norharmane matrix enhances detection of endotoxin by MALDI-MS for simultaneous profiling of pathogen, host, and vector systems. Pathog Dis. 74:ftw097. doi: 10.1093/femspd/ftw097
Scott, A. J., Oyler, B. L., Goodlett, D. R., and Ernst, R. K. (2017). Lipid A structural modifications in extreme conditions and identification of unique modifying enzymes to define the toll-like receptor 4 structure-activity relationship. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 1862, 1439–1450. doi: 10.1016/j.bbalip.2017.01.004
Silipo, A., Lanzetta, R., Garozzo, D., Lo Cantore, P., Iacobellis, N. S., Molinaro, A., et al. (2002). Structural determination of lipid A of the lipopolysaccharide from Pseudomonas reactans. A pathogen of cultivated mushrooms. Eur. J. Biochem. 269, 2498–2505. doi: 10.1046/j.1432-1033.2002.02914.x
Silipo, A., and Molinaro, A. (2010). “The diversity of the Core oligosaccharide in lipopolysaccharides” in Endotoxins: Structure, function and recognition subcellular biochemistry. eds. X. Wang and P. J. Quinn (Dordrecht: Springer Netherlands), 69–99.
Simpson, B. W., and Trent, M. S. (2019). Pushing the envelope: LPS modifications and their consequences. Nat. Rev. Microbiol. 17, 403–416. doi: 10.1038/s41579-019-0201-x
Smith, R. D., Izac, J. R., Ha, M., Yang, H., Johnson, J. K., and Ernst, R. K. (2021). Rapid identification of mcr-1-positive Escherichia coli from patient urine using a novel lipid-based MALDI-TOF-MS assay. Access Microbiol. 3:000309. doi: 10.1099/acmi.0.000309
Smith, R. D., McElheny, C. L., Izac, J. R., Gardner, F. M., Chandler, C. E., Goodlett, D. R., et al. (2022). A novel lipid-based MALDI-TOF assay for the rapid detection of Colistin-resistant Enterobacter species. Microbiol. Spectr. 10:e0144521. doi: 10.1128/spectrum.01445-21
Sonthayanon, P., Jaresitthikunchai, J., Mangmee, S., Thiangtrongjit, T., Wuthiekanun, V., Amornchai, P., et al. (2019). Whole cell matrix assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) for identification of Leptospira spp. in Thailand and Lao PDR. PLoS Negl. Trop. Dis. 13:e0007232. doi: 10.1371/journal.pntd.0007232
Sorensen, M., Chandler, C. E., Gardner, F. M., Ramadan, S., Khot, P. D., Leung, L. M., et al. (2020). Rapid microbial identification and colistin resistance detection via MALDI-TOF MS using a novel on-target extraction of membrane lipids. Sci. Rep. 10:21536. doi: 10.1038/s41598-020-78401-3
Srikram, A., Zhang, K., Bartpho, T., Lo, M., Hoke, D. E., Sermswan, R. W., et al. (2011). Cross-protective immunity against leptospirosis elicited by a live, attenuated lipopolysaccharide mutant. J. Infect. Dis. 203, 870–879. doi: 10.1093/infdis/jiq127
Strohalm, M., Kavan, D., Novák, P., Volný, M., and Havlícek, V. (2010). mMass 3: a cross-platform software environment for precise analysis of mass spectrometric data. Anal. Chem. 82, 4648–4651. doi: 10.1021/ac100818g
Thaipisuttikul, I., Hittle, L. E., Chandra, R., Zangari, D., Dixon, C. L., Garrett, T. A., et al. (2014). A divergent Pseudomonas aeruginosa palmitoyltransferase essential for cystic fibrosis-specific lipid A. Mol. Microbiol. 91, 158–174. doi: 10.1111/mmi.12451
Thibeaux, R., Iraola, G., Ferrés, I., Bierque, E., Girault, D., Soupé-Gilbert, M.-E., et al. (2018). Deciphering the unexplored Leptospira diversity from soils uncovers genomic evolution to virulence. Microb. Genom. 4:e000144. doi: 10.1099/mgen.0.000144
Trent, M. S., Ribeiro, A. A., Lin, S., Cotter, R. J., and Raetz, C. R. (2001). An inner membrane enzyme in Salmonella and Escherichia coli that transfers 4-amino-4-deoxy-L-arabinose to lipid A: induction on polymyxin-resistant mutants and role of a novel lipid-linked donor. J. Biol. Chem. 276, 43122–43131. doi: 10.1074/jbc.M106961200
Vanithamani, S., Akino Mercy, C. S., Kanagavel, M., Sumaiya, K., Bothammal, P., Saranya, P., et al. (2021). Biochemical analysis of leptospiral LPS explained the difference between pathogenic and non-pathogenic serogroups. Microb. Pathog. 152:104738. doi: 10.1016/j.micpath.2021.104738
Vincent, A. T., Schiettekatte, O., Goarant, C., Neela, V. K., Bernet, E., Thibeaux, R., et al. (2019). Revisiting the taxonomy and evolution of pathogenicity of the genus Leptospira through the prism of genomics. PLoS Negl. Trop. Dis. 13:e0007270. doi: 10.1371/journal.pntd.0007270
Viriyakosol, S., Matthias, M. A., Swancutt, M. A., Kirkland, T. N., and Vinetz, J. M. (2006). Toll-like receptor 4 protects against lethal Leptospira interrogans Serovar Icterohaemorrhagiae infection and contributes to in vivo control of Leptospiral burden. Infect. Immun. 74, 887–895. doi: 10.1128/IAI.74.2.887-895.2006
Wang, X., Ribeiro, A. A., Guan, Z., Abraham, S. N., and Raetz, C. R. H. (2007). Attenuated virulence of a Francisella mutant lacking the lipid A 4′-phosphatase. Proc. Natl. Acad. Sci. U. S. A. 104, 4136–4141. doi: 10.1073/pnas.0611606104
Werts, C., Tapping, R. I., Mathison, J. C., Chuang, T. H., Kravchenko, V., Saint Girons, I., et al. (2001). Leptospiral lipopolysaccharide activates cells through a TLR2-dependent mechanism. Nat. Immunol. 2, 346–352. doi: 10.1038/86354
Yang, H., Smith, R. D., Chandler, C. E., Johnson, J. K., Jackson, S. N., Woods, A. S., et al. (2022a). Lipid A structural determination from a single Colony. Anal. Chem. 94, 7460–7465. doi: 10.1021/acs.analchem.1c05394
Yang, H., Smith, R. D., Sumner, K. P., Goodlett, D. R., Johnson, J. K., and Ernst, R. K. (2022b). A matrix-assisted laser desorption ionization-time of flight mass spectrometry direct-from-urine-specimen diagnostic for gram-negative pathogens. Microbiol. Spectr. 10:e0373022. doi: 10.1128/spectrum.03730-22
Yasuda, P. H., Steigerwalt, A. G., Sulzer, K. R., Kaufmann, A. F., Rogers, F., and Brenner, D. J. (1987). Deoxyribonucleic acid relatedness between Serogroups and Serovars in the family Leptospiraceae with proposals for seven new Leptospira species. Int. J. Syst. Evol. Microbiol. 37, 407–415. doi: 10.1099/00207713-37-4-407
Zähringer, U., Salvetzki, R., Wagner, F., Lindner, B., and Ulmer, A. J. (2001). Structural and biological characterisation of a novel tetra-acyl lipid A from Escherichia coli F515 lipopolysaccharide acting as endotoxin antagonist in human monocytes. J. Endotoxin Res. 7, 133–146. doi: 10.1177/09680519010070020801
Zhou, Z., Ribeiro, A. A., Lin, S., Cotter, R. J., Miller, S. I., and Raetz, C. R. (2001). Lipid A modifications in polymyxin-resistant Salmonella typhimurium: PMRA-dependent 4-amino-4-deoxy-L-arabinose, and phosphoethanolamine incorporation. J. Biol. Chem. 276, 43111–43121. doi: 10.1074/jbc.M106960200
Keywords: lipid A, Leptospira, mass spectrometry, structure-activity relationship, molecular typing, lipopolysaccharide (LPS), fast lipid analysis technique, pathogenicity
Citation: Pětrošová H, Mikhael A, Culos S, Giraud-Gatineau A, Gomez AM, Sherman ME, Ernst RK, Cameron CE, Picardeau M and Goodlett DR (2023) Lipid A structural diversity among members of the genus Leptospira. Front. Microbiol. 14:1181034. doi: 10.3389/fmicb.2023.1181034
Edited by:
Laurent Rénia, Nanyang Technological University, SingaporeReviewed by:
Antonio Molinaro, University of Naples Federico II, ItalyCopyright © 2023 Pětrošová, Mikhael, Culos, Giraud-Gatineau, Gomez, Sherman, Ernst, Cameron, Picardeau and Goodlett. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Helena Pětrošová, aHBldHJvc0B1dmljLmNh; David R. Goodlett, Z29vZGxldHRAdXZpYy5jYQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.