 Nunziatina Russo1,2†
Nunziatina Russo1,2† Viviana Floridia3†
Viviana Floridia3† Enrico D’Alessandro3
Enrico D’Alessandro3 Vincenzo Lopreiato3
Vincenzo Lopreiato3 Alessandra Pino1,2,4*
Alessandra Pino1,2,4* Vincenzo Chiofalo3,5
Vincenzo Chiofalo3,5 Cinzia Caggia1,2,4
Cinzia Caggia1,2,4 Luigi Liotta3
Luigi Liotta3 Cinzia Lucia Randazzo1,2,4
Cinzia Lucia Randazzo1,2,4- 1Department of Agriculture, Food and Environment, University of Catania, Catania, Italy
- 2ProBioEtna SRL, Spin-Off of University of Catania, Catania, Italy
- 3Animal Production Unit, Department of Veterinary Sciences, University of Messina, Messina, Italy
- 4CERNUT, Interdepartmental Research Centre in Nutraceuticals and Health Products, University of Catania, Catania, Italy
- 5Consortium Research of Meat and Agribusiness Chain, Messina, Italy
Olive by-products represent a valuable low-price feed supplement for animal nutrition. In the present study, the effect of the dietary destoned olive cake supplementation, on both composition and dynamics of the fecal bacterial biota of cow, was assessed by Illumina MiSeq analysis of the 16S rRNA gene. In addition, metabolic pathways were predicted by using the PICRUSt2 bioinformatic tool. Eighteen lactating cows, according to the body condition score, the days from calving, and the daily milk production were homogeneously allocated into two groups, control or experimental, and subjected to different dietary treatments. In detail, the experimental diet contained, along with the components of the control one, 8% of destoned olive cake. Metagenomics data revealed significant differences in abundance rather than in richness between the two groups. Results showed that Bacteroidota and Firmicutes were identified as the dominant phyla, accounting for over 90% of the total bacterial population. The Desulfobacterota phylum, able to reduce sulfur compounds, was detected only in fecal samples of cows allocated to the experimental diet whereas the Elusimicrobia phylum, a common endosymbiont or ectosymbiont of various flagellated protists, was detected only in cows subjected to the control diet. In addition, both Oscillospiraceae and Ruminococcaceae families were mainly found in the experimental group whereas fecal samples of control cows showed the presence of Rikenellaceae and Bacteroidaceae families, usually associated with the high roughage or low concentrate diet. Based on the PICRUSt2 bioinformatic tool, pathways related to carbohydrate, fatty acid, lipid, and amino acids biosynthesis were mainly up regulated in the experimental group. On the contrary, in the control group, the metabolic pathways detected with the highest occurrence were associated with amino acids biosynthesis and degradation, aromatic compounds degradation, nucleosides and nucleotides biosynthesis. Hence, the present study confirms that the destoned olive cake is a valuable feed supplement able to modulate the fecal microbiota of cows. Further studies will be conducted in order to deepen the inter-relationships between the GIT microbiota and the host.
1. Introduction
Recently, the use of agro-industrial by-products as feed supplements in animal nutrition represents an interesting and successfully adopted strategy to reduce feeding costs and satisfy the nutritional needs of livestock (Chiofalo B. et al., 2020; Chiofalo V. et al., 2020; El Otmani et al., 2021; Bionda et al., 2022). In this context, the use of by-products of the olive oil industry, such as destoned olive cake, became widespread as a feed supplement since they are rich in nutraceutical molecules with antioxidant and antimicrobial features, including polyphenols (flavonoids, anthocyans, cyanidins, and phenolic acids), tyrosol, hydroxytyrosol, and oleuropein (Mannelli et al., 2018; Foti et al., 2022). Recent evidence suggests that the use of olive by-products did not negatively impact nutrients utilization, microbiota composition, and rumen fermentation variables, such as pH, ammonia and volatile fatty acids (VFA) concentrations (Ruiz et al., 2004; Estaún et al., 2014; Pallara et al., 2014; Tzamaloukas et al., 2021). Differently by influencing the ruminal fermentation and the rumen pH, the increase of the content of linoleic acid, in both milk and meat, as well as change in the aromatic and microbiological profiles of milk are reported (Chiofalo V. et al., 2020; Liotta et al., 2020; Foti et al., 2021; Tzamaloukas et al., 2021; Rabee et al., 2022). In fact, the high polyphenol content could modulate the rumen microbiota biodiversity and, consequently, affect the rumen metabolism, decreasing dietary protein degradation and fatty acid biohydrogenation by means of targeting specific groups of microorganisms (Mannelli et al., 2018; Biondi et al., 2019; Milani et al., 2020).
To date, modern sequencing technologies, based on culture-independent methods, represent the most powerful tools available for elucidating the diversity of animal microbiomes, positioning the microbial ecology of cattle for its renaissance (Hagey et al., 2019; Vaccalluzzo et al., 2020). In addition, the significant advances in sequence data analysis allow deep insights into this ecosystem (Hagey et al., 2019). The vast majority of available data, related to the microbial community identification in cattle, is focused on the rumen microbiota and less attention was paid to the study of fecal samples (Dowd et al., 2008; Shanks et al., 2011; Tang et al., 2017). Nevertheless, fecal sampling represents a non-invasive way suitable to link changes in bacterial abundance and diversity, along with related functional traits, to bovine rumen microbiome, with a high level of reproducibility and repeatability (Mott et al., 2022). In fact, the deepening of the fecal microbiota can provide valuable insights into the effect of feeding strategies on the nutritional status and wellbeing of livestock along with helping in manure management mitigating the environmental impact of pollution (Hagey et al., 2019). Recently published data, using 16S rRNA gene sequencing-based approaches, suggest that by studying the fecal microbiota of cattle is possible to elucidate the effects of a variety of feeds (Dowd et al., 2008; Callaway et al., 2010; Rice et al., 2012; Li et al., 2017; Kotz et al., 2021).
According to that, the main goal of the present study was to investigate the effect of the dietary destoned olive cake supplementation on fecal microbiota of cows using 16S rRNA gene amplicon sequencing. In addition, metabolic pathways, at KEGG level 3, were predicted by using the PICRUSt2 bioinformatic tools.
2. Materials and methods
2.1. Animal welfare
The Ethical Committee of the Department of Veterinary Science of the University of Messina approved all procedures (code 041/2020), conducted according to the European guidelines for the care and use of animals in research (Directive 2010/63/EU).
2.2. Animal management and diet
This experiment was conducted for 100 days (from February to June 2022) in a commercial dairy farm located 520 m above sea level in the province of Ragusa (Sicily, Italy). A total of 18 lactating cows, homogenous for the Body Condition Score (3 ± 0.5), distance from calving (90–120 days), and daily milk production (25 ± 3 kg/day) were randomly allocated in two groups, control (CTRL; 9 cows) or experimental (TRT; 9 cows). In detail, all cows were fed as total mixed ration (TMR) once daily at 0700 h where diet was composed by concentrate and meadow hay. The CTRL group received a concentrate without any olive cake integration, whereas the TRT group received a concentrate integrated with the enriched olive cake at the inclusion of 8%. The chemical composition of concentrates is reported in Table 1. The enriched olive cake, used as supplement, was obtained by mechanical pressing of olives carried out through a two-stage process, applied to produce extra virgin olive oil, by adding about 5% of a concentrate of vegetation waters, subsequently pitted by centrifugation and dried in the open air. The chemical composition (on DM basis) of the olive cake used in the present experiment was as follow: 95.6% of dry matter, 8.6% of crude protein, 30.3% of ether extract, 49.4% of neutral detergent fiber, 39.4% of acid detergent fiber, 23.1% of acid detergent lignin, 4.1% of ash, 1.5% of starch, and 9.360 mg/kg of polyphenols. A flowchart of olive cake production is reported in Figure 1.
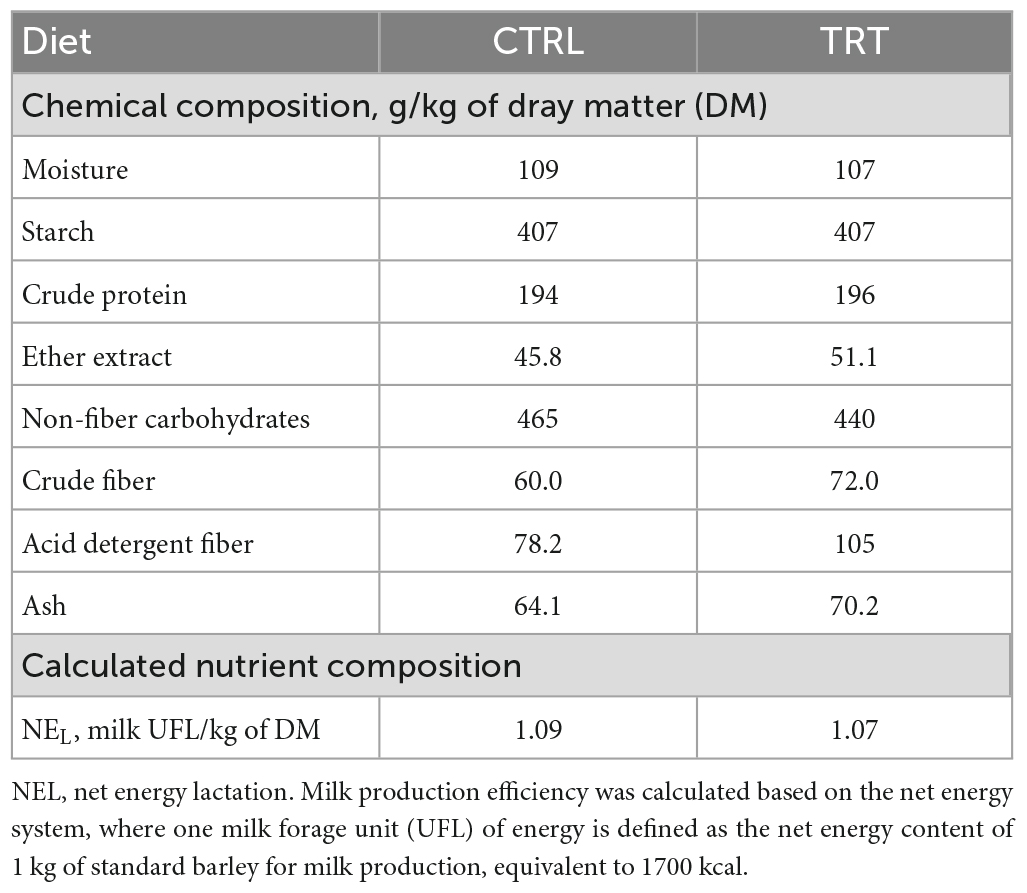
Table 1. Nutritional characteristics of concentrates used in the experiment.
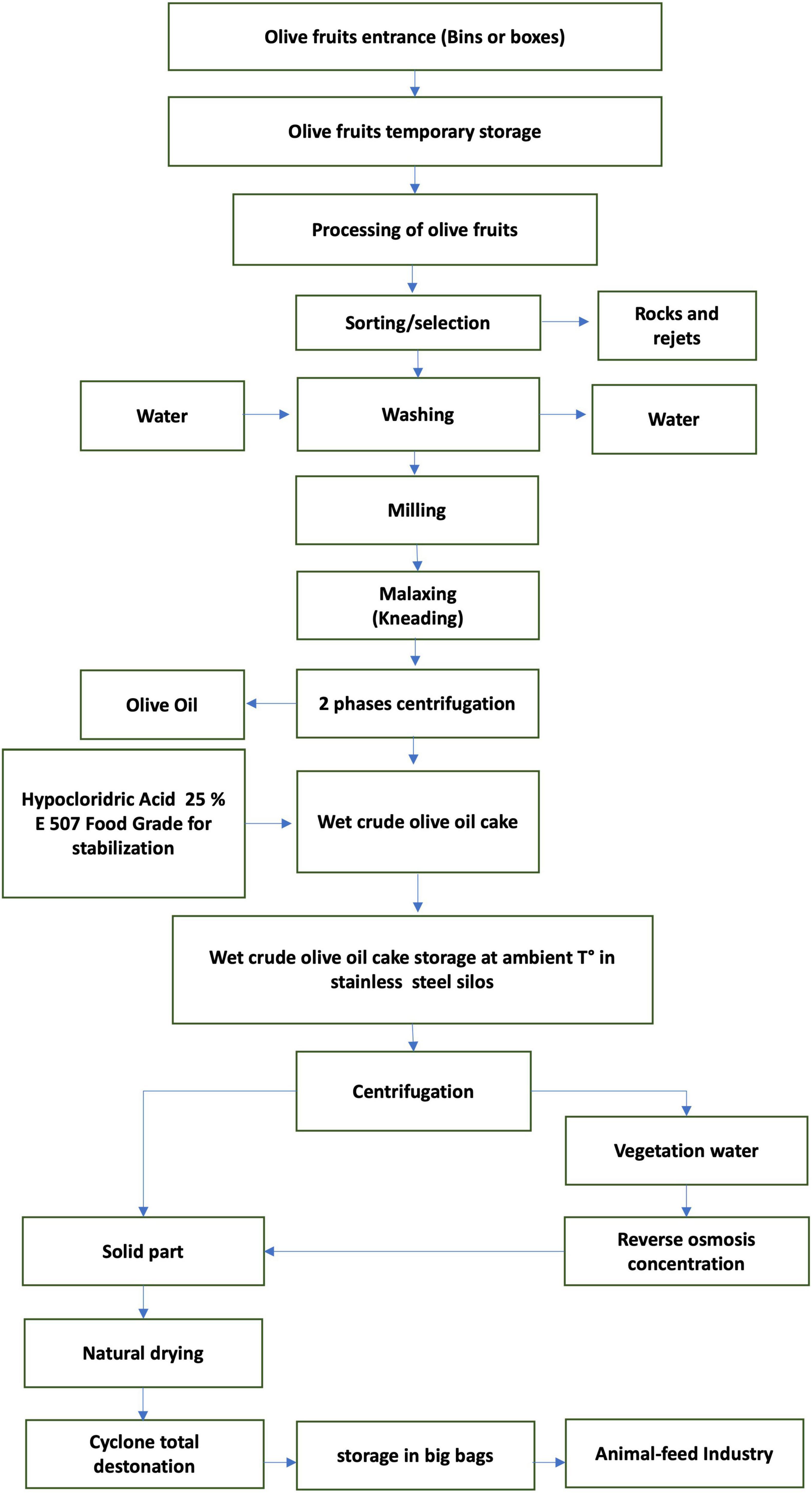
Figure 1. Flowchart of the destoned olive cake production.
2.3. Fecal samples collection and DNA extraction
Fecal samples were collected, from each cow, 100 days after the start of the control or experimental diet administration. Samples were aseptically collected from the rectal ampoule by using sterile gloves then placed in sterile containers and transferred under refrigerated conditions to the Laboratory of Microbiology of the Department of Agricultural Food and Environment (University of Catania, Italy) and immediately frozen at −80°C until analysis. Total genomic DNA was extracted using the commercial QIAamp® DNA Stool Mini Kit (QIAgen, Hilden, Germany) following the manufacturer instructions with a slight modification consisting of a repeated bead beating (RBB) pre-treatment step (Randazzo et al., 2015). In detail, after melted on ice, 0.25 g of feces was weighted under sterile conditions and transferred into a 2 ml screw-cap tube containing four glass-beads (2.7 mm, Biospec Products, Inc., USA) and 0.5 g of zirconia beads (0.1 mm, Biospec Products, Inc., USA). After the addition of 1 ml of lysis buffer (50 mM Tris–HCl, 500 mM NaCl, 50 mM EDTA, 4% w/v sodium dodecyl sulfate), the sample was subjected to mechanical cell lysis by performing three rounds of bead-beating with Precellys 24 Tissue Homogenizer (Bertin Technologies, Montigny-le-Bretonneux, France). Treatment time was 3 min for each bead-beating round and samples were cool on ice in between. After the RBB treatment the samples were centrifuged at + 4°C for 5 min at full speed and the collected supernatant was used for the DNA extraction following the kit manufacturer’s instructions. DNA concentration was evaluated using the fluorimeter Qubit 4.0 (Invitrogen, Carlsbad, CA, United States) before storing at −20°C until use.
2.4. 16S rRNA gene library construction and sequencing
The fecal microbiome composition was determined by 16S rRNA gene sequencing as previously described (Milani et al., 2013; Vaccalluzzo et al., 2022). Briefly, the V3 region of the 16S rRNA gene was amplified using PCR to build qualified libraries, which were subjected to MiSeq (Illumina) sequencing at the facilities of GenProbio Srl.1 The obtained 16S rRNA raw data were deposited at NCBI Sequence Read Archive (SRA)2 under accession code PRJNA909483.
2.5. Bioinformatic analysis
The raw reads were processed using Quantitative Insights Into Microbial Ecology (QIIME2) version 2022.2 (Bolyen et al., 2019). The sequences were quality filtered, trimmed, and denoised using Divisive Amplicon Denoising Algorithms 2 (DADA2). The high-quality sequences were then used to construct the amplicon sequence variant (ASV) feature table. Taxonomic classification was made through the SILVA reference database (v138) (Robeson et al., 2021) with a percentage of identity of 75, 87, and 95% for phylum, family, and genus levels, respectively (Henderson et al., 2019). ASVs with relative abundance lower than 0.1% were grouped as “others.”
2.6. Alpha and beta diversity, differential analysis, and enzymatic prediction
The determination of diversity and differences in the abundance of the fecal bacterial community was performed using RStudio software (version 4.1.2). The phyloseq packing of R (McMurdie and Holmes, 2013) was used for alpha and beta diversity. The alpha diversity, based on genus level, was evaluated considering three measures: observed richness, Chao1 index, and Shannon index. The box plot of alpha diversity was generated using the ggplot2 package (Wickham, 2016). Beta diversity, based on genus level, was evaluated with the Bray-Curtis distance and plotted with the Principal Coordinate Analysis (PCoA). Differential analysis, based on genus level, was performed using the DESeq2 package in R (Love et al., 2014) and the differences between CTRL and TRT groups were evaluated.
The Phylogenetic Investigation of Communities by Reconstruction of Unobserved States (PICRUSt2) v2.5.0 (Douglas et al., 2020) was used to predict the functional abundances at KEGG level 3 (i.e., Environmental Information Processing) based on 16S rRNA gene sequencing data obtained during the denoise step by QIIME2. Statistical analysis of taxonomic and functional profiles (STAMP) (v2.1.3) (Parks et al., 2014) was used to illustrate, based on the PICRUSt2 outputs, the difference in the predicting microbial functions associated with the diet.
2.7. Statistical analysis
The alpha diversity, according to observed richness, Chao1 and Shannon indices, between CTRL and TRT groups was compared with one-way non-parametric Wilcoxon test. Beta diversity, according to Bray Curtis distance, was evaluated through permutational multivariate analysis of variance (PERMANOVA) with 999 permutations with the vegan package.
Differential abundance (DA) was considered with a false discovery rate (FDR) cut-off of 0.05 and a fold-change (FC) higher than 1.5 or lower than −1.5 (i.e., |log2FC| > 0.59). The difference in the prediction of microbial functions associated with the diet (CTRL vs. TRT) was detected with Welch’s t-test with a confidence interval of 95% and the data were corrected with Benjamini–Hochberg FDR. Significance was determined at P ≤ 0.05.
3. Results
3.1. Taxonomy classification
After trimming, denoising, chimera-removal and merging with QIIME2, the obtained high-quality sequences were used for the taxonomic classification. Sample metadata, denoising statistics, taxonomy classification and the relative abundances for each taxonomic level are reported in Supplementary Data 1. As reported in Table 2, the total ASVs were assigned to 10 phyla, 51 families and 78 genera in CTRL group, whereas 10 phyla, 44 families and 78 genera were detected in TRT group. The relative abundance of bacteria, detected in both CTRL and TRT groups, at phylum, family, and genus levels is shown in Figures 2A–C. In detail, Firmicutes, Bacteroidota, Actinobacteriota, Spirochaetota, Proteobacteria, Verrucomicrobiota, Patescibacteria, Cyanobacteria, and Fibrobacterota phyla were detected in both CTRL and TRT groups with relative abundance higher than 0.1%. Although Firmicutes and Bacteroidota were the most abundant phyla in both CTRL and TRT groups, the Firmicutes phylum showed higher relative abundance in TRT samples compared with CTRL. Moreover, the Desulfobacterota and Elusimicrobia phyla were not detected in CTRL and TRT groups, respectively (Figure 2A).
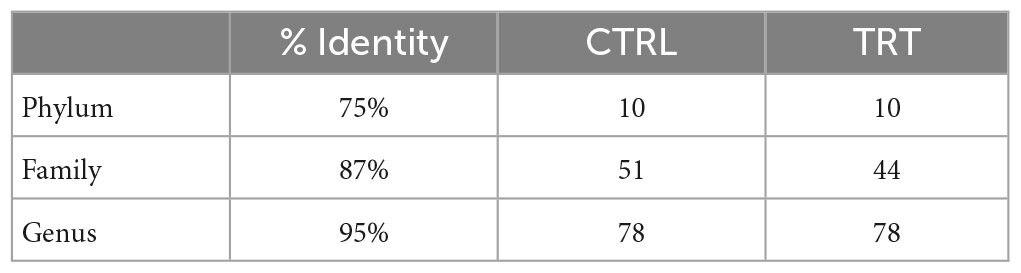
Table 2. Overall microbiota composition of fecal samples collected from cows subjected to control (CTRL) and experimental (TRT) diet.
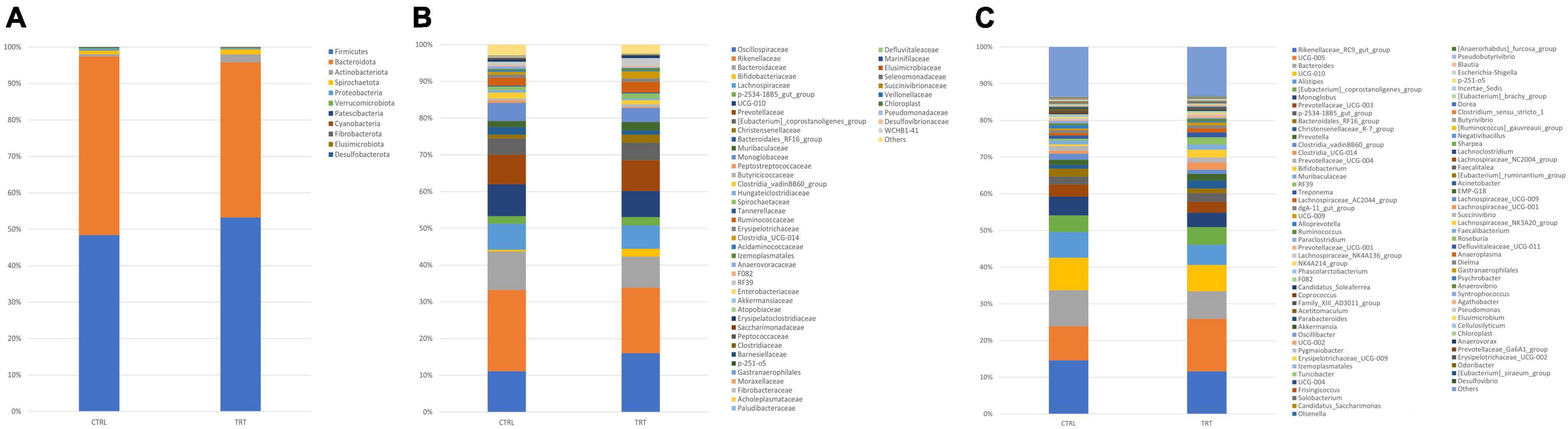
Figure 2. Relative abundance of phyla (A), families (B), and genera (C) detected in fecal samples from cows subjected to control (CTRL) and experimental (TRT) diet.
Overall, at family level, the ASVs of CTRL group were classified into 51 families, whereas the ASVs of TRT group were classified into 44 families (Figure 2B). In fact, compared with CTRL group, the Peptococcaceae, Defluviitaleaceae, Elusimicrobiaceae, Succinivibrionaceae, Veillonellaceae, Pseudomonadaceae, and WCHB1-41 families where not detected in TRT group. As reported in Figure 3, showing the ten most abundant families, Oscillospiraceae and Ruminococcaceae were dominant in in TRT group, whereas Rikenellaceae and Bacteroidaceae were most abundant in CTRL group.
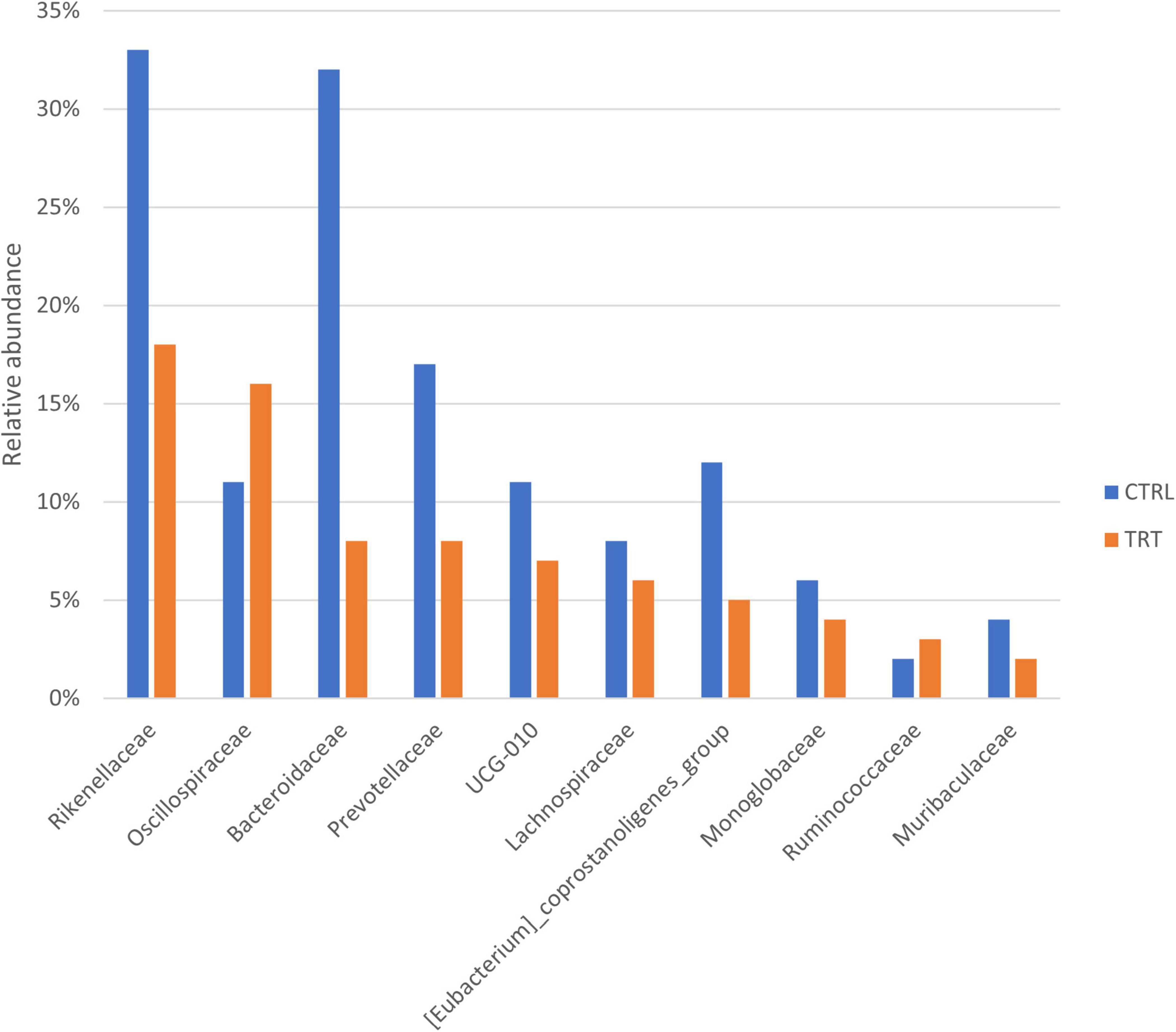
Figure 3. Top 10 most abundant families detected in control (CTRL) and experimental (TRT) groups.
At genus level, 78 genera were overall identified in both CTRL and TRT groups (Figure 2C) and the most abundant are reported in Figure 4. In particular, Rikenellaceae_RC9_gut_group, UCG-010 and Monoglobus showed higher relative abundance in CTRL group, while UCG-005, Prevotellaceae_UCG-003 and p-2534-18B5_gut_group were prevalent in TRT group (Figure 4).
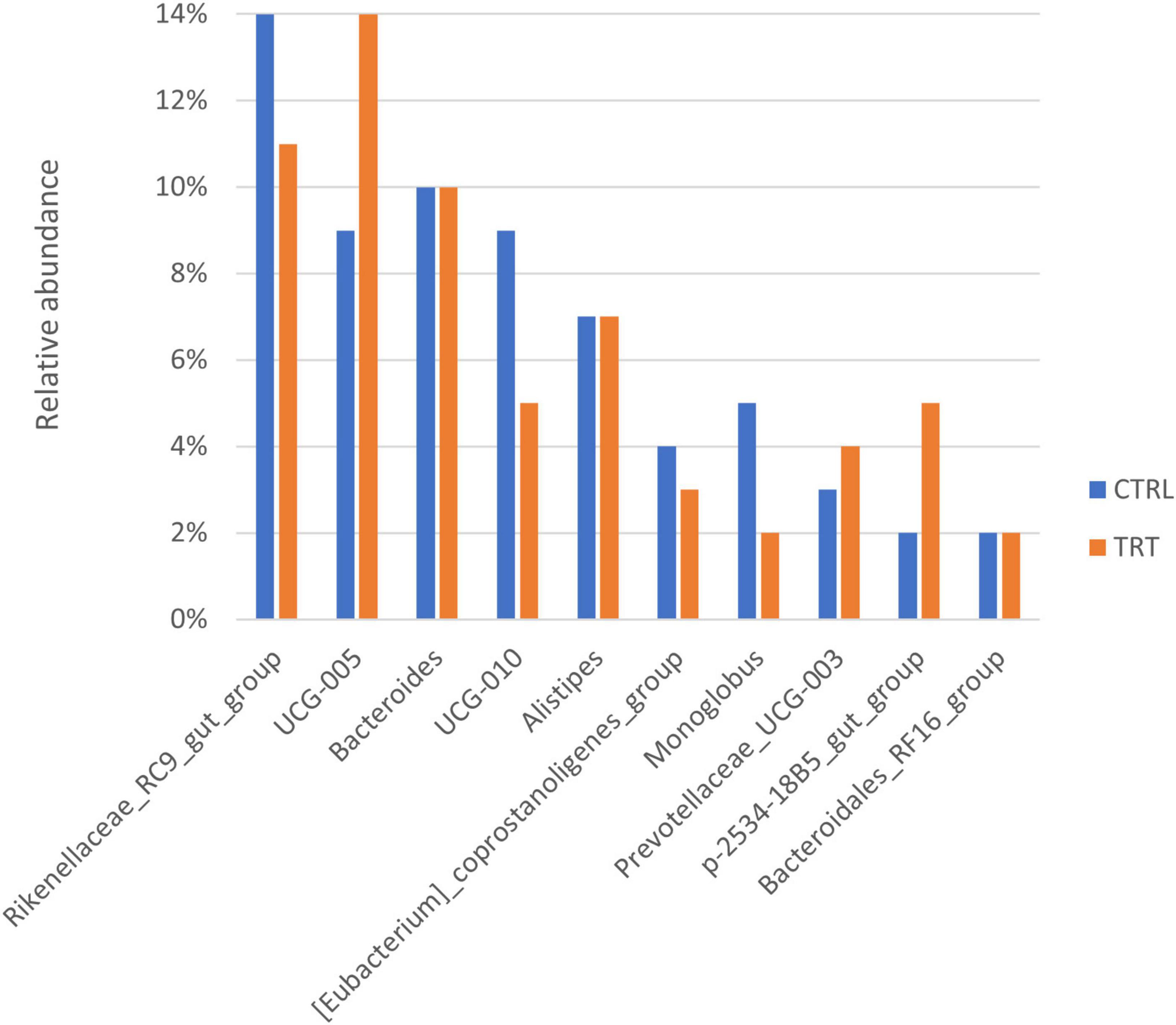
Figure 4. Top 10 most abundant genera detected in control (CTRL) and experimental (TRT) groups.
Moreover, Roseburia, Prevotellaceae_Ga6A1_g -roup, Cellulosilyticum, Gastranaerophilales, [Eubacterium]_siraeum_group, Lachnospiraceae_UCG-009, Dielma, Odoribacter, Anaeroplasma, Faecalibacterium, Chloroplast and Desulfovibrio were not found in CTRL group whereas Negativibacillus, Acinetobacter, Agathobacter, Erysipelotrichaceae_UCG-002, Defluviitaleaceae_UCG-011, Anaerovorax, Elusimicrobium, Anaerovibrio, Succinivibrio, Syntrophococcus, Psychrobacter, and Pseudomonas were not found in TRT group (Figure 2C).
3.2. Alpha and beta diversity, differential analysis, and enzymatic prediction
Observed richness, Chao1, and Shannon indices are plotted in Supplementary Figures 1A–C. Based on the Wilcoxon test, no difference in richness was detected between CTRL and TRT groups.
The beta diversity (Figure 5), based on the Bray-Curtis distance method, allowed to group the analyzed samples in relation to the dietary treatment (CTRL or TRT). Based on PERMANOVA results, the diet significantly affected the fecal microbial communities (p < 0.001).
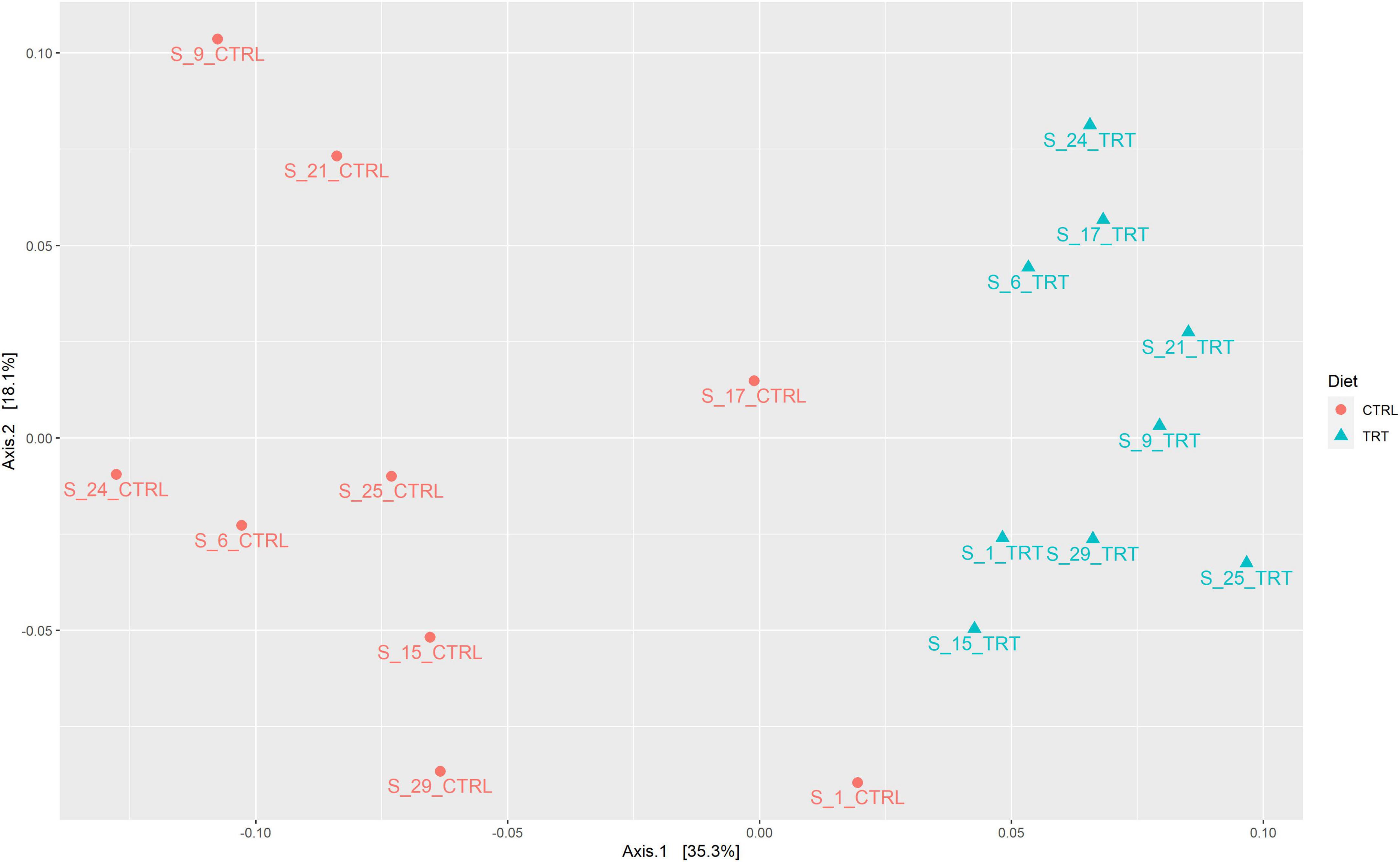
Figure 5. Principal coordinate analysis (PCoA) depicting the distribution of samples according to Bray—Curtis distance.
The model used in the differential analysis allowed to detect the genera, occurring in both CTRL and TRT groups, with significant differences in terms of percentage of occurrence (Table 3).
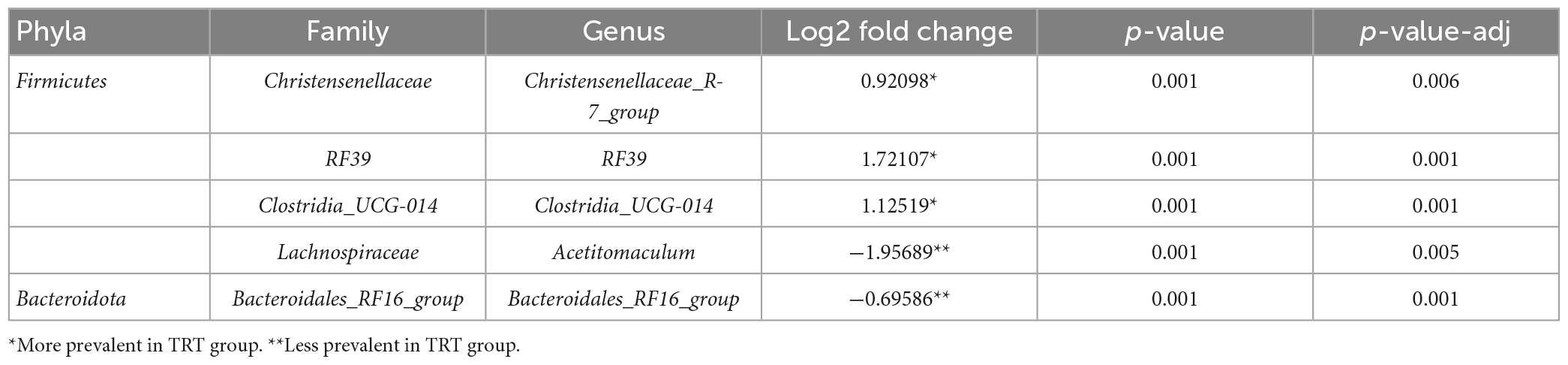
Table 3. Different taxa identified by comparing the fecal microbiota of cows allocated to control (CTRL) and experimental (TRT) groups.
Overall, 366 pathways were detected based on KEGG prediction (Supplementary Data 2) and 58 of them, with different abundance between CTRL and TRT groups, are shown in Figure 6. In detail, in TRT group, the metabolic pathways mainly identified were involved in the biosynthesis of carbohydrates (Glycogen biosynthesis I, UDP-N-acetyl-D-glucosamine biosynthesis I, O-antigen building blocks biosynthesis), fatty acids and lipids (Phosphatidylglycerol biosynthesis I and II), and amino acids (Superpathway of aromatic amino acid biosynthesis, Superpathway of L-threonine biosynthesis, L-histidine biosynthesis, Superpathway of L-isoleucine biosynthesis I and L-lysine biosynthesis III). In the CTRL group, the identified pathways were mainly involved in amino acids degradation (L-histidine degradation I and III) and biosynthesis (L-arginine biosynthesis III and Superpathway of L-methionine biosynthesis) as well as in aromatic compounds degradation and nucleosides and nucleotides biosynthesis (Figure 6).
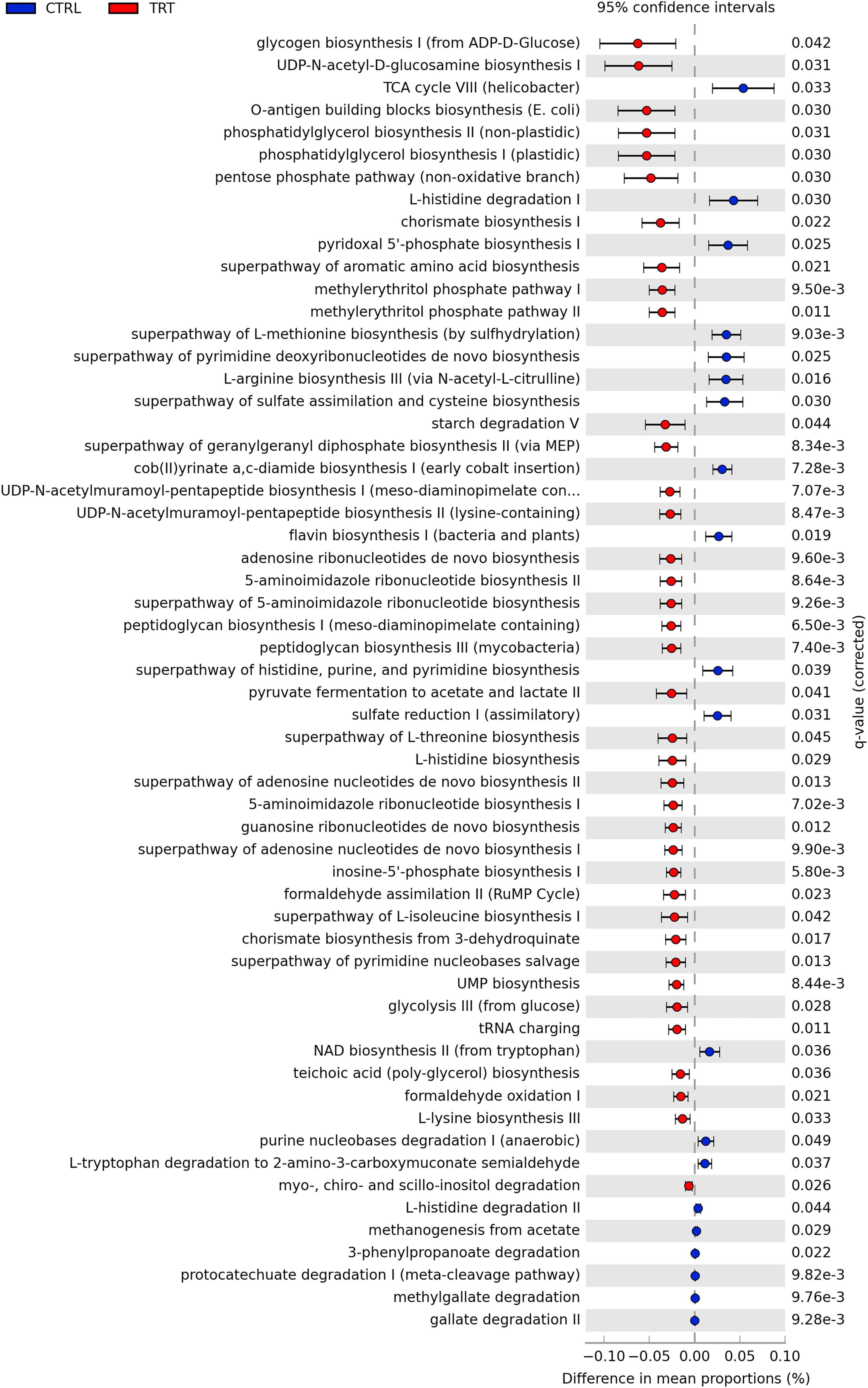
Figure 6. Differentially metabolic pathways between control (CTRL) and experimental (TRT) groups.
4. Discussion
The development of alternative feedstuffs and the use of by-products, as feed supplements, represent a challenge for animal nutrition researchers to boost farm livestock wellbeing as well as increase both the production and quality of animal-derived products. The diet becomes a key factor contributing to changes in the composition of the gastrointestinal tract (GIT) microbiota (Zhang et al., 2021; Welch et al., 2022). According to that, the present study, by applying a metagenomic approach, aimed to deepen the effect of the dietary destoned olive cake supplementation on both composition and dynamics of the fecal bacterial biota of cow as well as on the KEGGs functional profile.
Consistent with previously reported data, our study confirmed that polyphenols rich diet is able to modulate the microbial community affecting, in turn, the fecal microbiota composition (Mao et al., 2013; Plaizier et al., 2017; Hagey et al., 2019). In fact, although no significant difference in richness was detected, beta diversity allowed to discriminate the analyzed samples based on the diet regime. Metagenomics approaches revealed that the fecal microbiota of cattle is dominated by Firmicutes and Bacteroidetes phyla, commonly recognized as health-promoting (Henderson et al., 2015; Mannelli et al., 2019; Conte et al., 2022; Welch et al., 2022). More in depth, Firmicutes are involved in the degradation of oligosaccharide, fiber, and starch, helping the host intestinal tract in the absorption of energy from food. In addition, members of the Firmicutes phylum are able to produce volatile fatty acids such as butyrate, which is linked with gut health (Kim et al., 2011). Similarly, members of the Bacteroidetes phylum have many functions in the gut, including degradation of carbohydrates, such as complex plant cell walls, as well as production of butyrate, a significant player in energy metabolism in the rumen (Thomas et al., 2011; Miguel et al., 2019). It is well known that the Firmicutes phylum acts by increasing the nutrients availability whereas the Bacteroidetes one is energetically less favorable to the host (Xu et al., 2021).
According to previous studies, suggesting that dietary changes significantly affect the Firmicutes:Bacteroidota ratio (Mao et al., 2013; Plaizier et al., 2017, 2018), our data revealed increased abundance of Firmicutes and decreased occurrence of Bacteroidota in fecal samples of cows subjected to destoned olive cake supplementation. In addition, the Desulfobacterota phylum was detected only in fecal samples of treated cows. Noteworthy, the aforementioned phylum includes microorganisms able to reduce sulfur compounds via the sulfite reductase pathway, followed by butyrate degradation, playing a significant role in energy metabolism (Miguel et al., 2019). Moreover, Elusimicrobia, a recently defined animal-associated phylum, occurring as endosymbiont or ectosymbiont of various flagellated protists (Méheust et al., 2020), was detected only in cows subjected to control diet. The presence of both Oscillospiraceae and Ruminococcaceae families mainly in experimental group can be associate to the diet regime. In fact, as recently reported by Yang et al. (2021), polyphenols can increase the abundance of the families mentioned above which are able to ferment complex plant carbohydrates and to produce short chain fatty acids competing with activation of energy metabolism. Conversely, fecal samples of control cows showed the presence of Rikenellaceae and Bacteroidaceae families, usually associated with the high roughage or low concentrate diet (Mpanza et al., 2022), playing a key role in carbohydrates degradation and in the production of VFAs, including succinate, acetate and propionate (Wang et al., 2020). Consistent with metagenetic data, in the present study, most of the predicted metabolic pathways are involved in biosynthetic processes. Among these, pathways related to carbohydrate, fatty acid, lipid and amino acids biosynthesis were mainly present in experimental group than in control one. This finding is in accordance with the observed dominance of both Bacteroidota and Firmicutes phyla in fecal microbiota. In fact, it is well known that Firmicutes are involved in the degradation of complex polysaccharides, with subsequent synthesis of VFA, whereas Bacteroidetes mainly degrades carbohydrates, fats, and proteins (Jami et al., 2013; Yildirim et al., 2021). On the contrary, in control group, the metabolic pathways detected with highest occurrence were associated to amino acids biosynthesis and degradation, aromatic compounds degradation, nucleosides and nucleotides biosynthesis. It is well known that bacteria require the synthesis and/or acquisition of purines and pyrimidines, which form the basis of nucleotides, to survive, even having strong links with the virulence factors of opportunistic and bacterial pathogens (Goncheva et al., 2022). In fact, in most bacteria, the nucleotides are synthesized de novo and the products are used in many cell functions, including DNA replication, energy storage, and as signaling molecules (Goncheva et al., 2022). Our results confirm that the feeding regime significantly affect the composition and dynamics of the fecal microbiota as well as the microbial metabolism. Further studies will be conducted in order to in depth investigate the molecular functions of microbiota by using insightful methods, such as metabolomics and metatranscriptomics.
5. Conclusion
The present study confirms that, among olive by-products, destoned olive cake is a valuable feed supplement for cow’s nutrition. The destoned olive cake supplementation was able to modulate the fecal microbiota determining the increase of Firmicutes phylum, associated to growing nutrients availability, and the reduction of Bacteroidetes, energetically less favorable to the host. The prediction of metabolic pathways revealed a significant effect of the regime diet on carbohydrate, fatty acid, lipid and amino acids biosynthesis. Further studies will be conducted in order to deepen the inter-relationships between the GIT microbiota and the host.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
Ethics statement
The study was reviewed and approved by the Ethical Committee of the Department of Veterinary Science of the University of Messina (code 041/2020). All procedures were conducted according to the European guidelines for the care and use of animals in research (Directive 2010/63/EU). Written informed consent was obtained from the owners for the participation of their animals in this study.
Author contributions
LL, CR, and VL: conceptualization. AP, VC, and CR: methodology. VF and AP: software. VL, AP, LL, and CR: validation. NR and VF: formal analysis and writing—original draft preparation. VL and LL: investigation. VF, ED’A, AP, and LL: data curation. VL, AP, CC, LL, and CR: writing—review and editing. LL and CR: visualization and supervision. LL: project administration and funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by P.O. FESR SICILIA 2014/2020. Obiettivo Tematico 1—Ricerca, Sviluppo Tecnologico e Innovazione Obiettivo specifico 1.1–Incremento dell’attività di Innovazione delle Imprese Azione 1.1.5–Sostegno all’avanzamento Tecnologico delle Imprese Attraverso il Finanziamento di linee pilota e azioni di validazione precoce dei prodotti e di dimostrazione su larga scala. Project BioTrak. Grant number 08SR1091000150 -CUP G69J18001000007 (Principal Investigator LL).
Acknowledgments
We thank the Feed Manufacturing Industry Dipasquale srl (Avola, SR, Italy) for the production of feed, the Oil Mill Industry Consoli (Adrano, CT, Italy) for the production of destoned olive cake, and the Lissandrello dairy farm for guesting the infield trials.
Conflict of interest
NR, AP, CC, and CR declare that they are members of ProBioEtna, a spinoff of the University of Catania, Italy.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2023.1137452/full#supplementary-material
Supplementary Figure 1 | Alpha diversity based on (A) Observed, (B) Chao1, and (C) Shannon index. Statistical analysis provides the difference of richness at genus level in the two groups (CTRL vs. TRT).
Supplementary Data 1 | 16S rRNA raw data.
Supplementary Data 2 | Abundance of each pathway detected by prediction analysis.
Footnotes
References
Bionda, A., Lopreiato, V., Crepaldi, P., Chiofalo, V., Fazio, E., Oteri, M., et al. (2022). Diet supplemented with olive cake as a model of circular economy: metabolic and endocrine responses of beef cattle. Front. Sustain. Food Syst. 6:1077363. doi: 10.3389/fsufs.2022.1077363
Biondi, L., Randazzo, C. L., Russo, N., Pino, A., Natalello, A., Van Hoorde, K., et al. (2019). Dietary supplementation of tannin-extracts to lambs: effects on meat fatty acids composition and stability and on microbial characteristics. Foods 8:469. doi: 10.3390/foods8100469
Bolyen, E., Rideout, J. R., Dillon, M. R., Bokulich, N. A., Abnet, C. C., Al-Ghalith, G. A., et al. (2019). Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857. doi: 10.1038/s41587-019-0209-9
Callaway, T. R., Dowd, S. E., Edrington, T. S., Anderson, R. C., Krueger, N., Bauer, N., et al. (2010). Evaluation of bacterial diversity in the rumen and feces of cattle fed different levels of dried distillers grains plus solubles using bacterial tag-encoded FLX amplicon pyrosequencing. J. Anim. Sci. 88, 3977–3983. doi: 10.2527/jas.2010-2900
Chiofalo, B., Di Rosa, A. R., Lo Presti, V., Chiofalo, V., and Liotta, L. (2020). Effect of supplementation of herd diet with olive pâté on the composition profile of milk and on the composition, quality and sensory profile of cheeses made therefrom. Animal 10:977. doi: 10.3390/ani10060977
Chiofalo, V., Liotta, L., Lo Presti, V., Gresta, F., Di Rosa, A., and Chiofalo, B. (2020). Effect of dietary olive cake supplementation on performance, carcass characteristics, and meat quality of beef cattle. Animals 10:1176. doi: 10.3390/ani10071176
Conte, G., Dimauro, C., Daghio, M., Serra, A., Mannelli, F., McAmmond, B. M., et al. (2022). Exploring the relationship between bacterial genera and lipid metabolism in bovine rumen. Animal 16:100520. doi: 10.1016/j.animal.2022.100520
Douglas, G. M., Maffei, V. J., Zaneveld, J. R., Yurgel, S. N., Brown, J. R., Taylor, C. M., et al. (2020). PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 38, 685–688. doi: 10.1038/s41587-020-0548-6
Dowd, S. E., Callaway, T. R., Wolcott, R. D., Sun, Y., McKeehan, T., Hagevoort, R. G., et al. (2008). Evaluation of the bacterial diversity in the feces of cattle using 16S rDNA bacterial tag-encoded FLX amplicon pyrosequencing (bTEFAP). BMC Microbiol. 8:125. doi: 10.1186/1471-2180-8-125
El Otmani, S., Chebli, Y., Taminiau, B., Chentouf, M., Hornick, J. L., and Cabaraux, J. (2021). Effect of olive cake and cactus cladodes incorporation in goat kids’ diet on the rumen microbial community profile and meat fatty acid composition. Biology 10:1237. doi: 10.3390/biology10121237
Estaún, J., Dosil, J., Al Alami, A., Gimeno, A., and De Vega, A. (2014). Effects of including olive cake in the diet on performance and rumen function of beef cattle. Anim. Prod. Sci. 54, 1817–1821. doi: 10.1071/AN14352
Foti, P., Pino, A., Romeo, F. V., Vaccalluzzo, A., Caggia, C., and Randazzo, C. L. (2022). Olive pomace and pâté olive cake as suitable ingredients for food and feed. Microorganisms 10:237. doi: 10.3390/microorganisms10020237
Foti, P., Romeo, F. V., Russo, N., Pino, A., Vaccalluzzo, A., Caggia, C., et al. (2021). Olive mill wastewater as renewable raw materials to generate high added-value ingredients for agro-food industries. Appl. Sci. 11:7511. doi: 10.3390/app11167511
Goncheva, M. I., Chin, D., and Heinrichs, D. E. (2022). Nucleotide biosynthesis: the base of bacterial pathogenesis. Trends Microbiol. 30, 793–804. doi: 10.1016/j.tim.2021.12.007
Hagey, J. V., Bhatnagar, S., Heguy, J. M., Karle, B. M., Price, P. L., Meyer, D., et al. (2019). Fecal microbial communities in a large representative cohort of California dairy cows. Front. Microbiol. 10:1093. doi: 10.3389/fmicb.2019.01093
Henderson, G., Cox, F., Ganesh, S., Jonker, A., Young, W., Collaborators, G. R. C., et al. (2015). Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 5:14567. doi: 10.1038/srep14567
Henderson, G., Yilmaz, P., Kumar, S., Forster, R. J., Kelly, W. J., Leahy, S. C., et al. (2019). Improved Taxonomic Assignment of Rumen Bacterial 16S RRNA Sequences Using a Revised SILVA Taxonomic Framework. PeerJ 7:e6496. doi: 10.7717/peerj.6496
Jami, E., Israel, A., Kotser, A., and Mizrahi, I. (2013). Exploring the bovine rumen bacterial community from birth to adulthood. ISME J. 7, 1069–1079. doi: 10.1038/ismej.2013.2
Kim, M., Morrison, M., and Yu, Z. (2011). Status of the phylogenetic diversity census of ruminal microbiomes. FEMS Microbiol. Ecol. 76, 49–63. doi: 10.1111/j.1574-6941.2010.01029.x
Kotz, A., Azevedo, P. A., Khafipour, E., and Plaizier, J. C. (2021). Effects of the dietary grain content on rumen and fecal microbiota of dairy cows. Can. J. Anim. Sci. 101, 274–286. doi: 10.1139/cjas-2020-0122
Li, Y., Meng, Q., Zhou, B., and Zhou, Z. (2017). Effect of ensiled mulberry leaves and sun-dried mulberry fruit pomace on the fecal bacterial community composition in finishing steers. BMC Microbiol. 17:9. doi: 10.1186/s12866-017-1011-9
Liotta, L., Chiofalo, V., Lo Presti, V., and Chiofalo, B. (2020). Effect of production system on growth performances and meat traits of suckling Messinese goat kids. Ital. J. Anim. Sci. 19, 245–252. doi: 10.1080/1828051X.2020.1726832
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15:550. doi: 10.1186/s13059-014-0550-8
Mannelli, F., Cappucci, A., Pini, F., Pastorelli, R., Decorosi, F., Giovannetti, L., et al. (2018). Effect of different types of olive oil pomace dietary supplementation on the rumen microbial community profile in Comisana ewes. Sci. Rep. 8:8455. doi: 10.1038/s41598-018-26713-w
Mannelli, F., Daghio, M., Alves, S. P., Bessa, R. J. B., Minieri, S., Giovannetti, L., et al. (2019). Effects of chestnut tannin extract, vescalagin and gallic acid on the dimethyl acetals profile and microbial community composition in rumen liquor: an in vitro study. Microrganisms 7:202. doi: 10.3390/microorganisms7070202
Mao, S. Y., Zhang, R. Y., Wang, D. S., and Zhu, W. Y. (2013). Impact of subacute ruminal acidosis (SARA) adaptation on rumen microbiota in dairy cattle using pyrosequencing. Anaerobe 24, 12–19. doi: 10.1016/j.anaerobe.2013.08.003
McMurdie, P. J., and Holmes, S. (2013). Phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One 8:e61217. doi: 10.1371/journal.pone.0061217
Méheust, R., Castelle, C. J., Matheus Carnevali, P. B., Farag, I. F., He, C., Chen, L. X., et al. (2020). Groundwater Elusimicrobia are metabolically diverse compared to gut microbiome Elusimicrobia and some have a novel nitrogenase paralog. ISME J. 14, 2907–2922. doi: 10.1038/s41396-020-0716-1
Miguel, M. A., Lee, S. S., Mamuad, L. L., Choi, Y. J., Jeong, C. D., Son, A., et al. (2019). Enhancing butyrate production, ruminal fermentation and microbial population through supplementation with Clostridium saccharobutylicum. J. Microbiol. Biotechnol. 29, 1083–1095. doi: 10.4014/jmb.1905.05016
Milani, C., Hevia, A., Foroni, E., Duranti, S., Turroni, F., Lugli, G. A., et al. (2013). Assessing the fecal microbiota: an optimized ion torrent 16S rRNA gene-based analysis protocol. PLoS One. 8:68739. doi: 10.1371/journal.pone.0068739
Milani, M., Consoli, S., Marzo, A., Pino, A., Randazzo, C., Barbagallo, S., et al. (2020). Treatment of winery wastewater with a multistage constructed wetland system for irrigation reuse. Water 12:1260. doi: 10.3390/w12051260
Mott, A. C., Schneider, D., Hünerberg, M., Hummel, J., and Tetens, J. (2022). Bovine rumen microbiome: impact of dna extraction methods and comparison of non-invasive sampling sites. Ruminan 2, 112–132. doi: 10.3390/ruminants2010007
Mpanza, T. D. E., Dhlamini, T. C., Pierneef, R. E., and Mbatha, K. R. (2022). Enteric methane emission, rumen fermentation and microbial profiles of meat-master lambs supplemented with barley fodder sprouts. Ferment 8:434. doi: 10.3390/fermentation8090434
Pallara, G., Buccioni, A., Pastorelli, R., Minieri, S., Mele, M., Rapaccini, S., et al. (2014). Effect of stoned olive pomace on rumen microbial communities and polyunsaturated fatty acid biohydrogenation: an in vitro study. BMC Veter. Res. 10:271. doi: 10.1186/s12917-014-0271-y
Parks, D. H., Tyson, G. W., Hugenholtz, P., and Beiko, R. G. (2014). STAMP: statistical analysis of taxonomic and functional profiles. Bioinform 30, 3123–3124. doi: 10.1093/bioinformatics/btu494
Plaizier, J. C., Danesh Mesgaran, M., Derakhshani, H., Golder, H., Khafipour, E., Kleen, J. L., et al. (2018). Review: enhancing gastrointestinal health in dairy cows. Animal 12, S399–S418. doi: 10.1017/S1751731118001921
Plaizier, J. C., Li, S., Tun, H. M., and Khafipour, E. (2017). Nutritional models of experimentally-induced subacute ruminal acidosis (SARA) differ in their impact on rumen and hindgut bacterial communities in dairy cows. Front. Microbiol. 7:2128. doi: 10.3389/fmicb.2016.02128
Rabee, A. E., Kewan, K. Z., El Shaer, H. M., Lamara, M., and Sabra, E. A. (2022). Effect of olive and date palm by-products on rumen methanogenic community in Barki sheep. AIMS Microbiol. 27, 26–41. doi: 10.3934/microbiol.2022003
Randazzo, C. L., Pino, A., Ricciardi, L., Romano, C., Comito, D., Arena, E., et al. (2015). Probiotic supplementation in systemic nickel allergy syndrome patients: study of its effects on lactic acid bacteria population and on clinical symptoms. J. Appl. Microbiol. 118, 202–211. doi: 10.1111/jam.12685
Rice, W. C., Galyean, M. L., Cox, S. B., Dowd, S. E., and Cole, N. A. (2012). Influence of wet distillers grains diets on beef cattle fecal bacterial community structure. BMC Microbiol. 12:25. doi: 10.1186/1471-2180-12-25
Robeson, M. S., O’Rourke, D. R., Kaehler, B. D., Ziemski, M., Dillon, M. R., Foster, J. T., et al. (2021). RESCRIPt: reproducible sequence taxonomy reference database management. PLoS Comput. Biol. 17:e1009581. doi: 10.1371/journal.pcbi.1009581
Ruiz, D. R. Y., Moumen, A., García, A. I. M., and Alcaide, E. M. (2004). Ruminal fermentation and degradation patterns, protozoa population, and urinary purine derivatives excretion in goats and wethers fed diets based on two-stage olive cake: effect of peg supply1. J. Anim. Sci. 82, 2023–2032. doi: 10.2527/2004.8272023x
Shanks, O. C., Kelty, C. A., Archibeque, S., Jenkins, M., Newton, R. J., Mclellan, S. L., et al. (2011). Community structures of fecal bacteria in cattle from different animal feeding operations. Appl. Environ. Microbiol. 77, 2992–3001. doi: 10.1128/AEM.02988-10
Tang, M. T., Han, H., Yu, Z., Tsuruta, T., and Nishino, N. (2017). Variability, stability, and resilience of fecal microbiota in dairy cows fed whole crop corn silage. Appl. Microbiol. Biotechnol. 101, 6355–6364. doi: 10.1007/s00253-017-8348-8
Thomas, F., Hehemann, J.-H., Rebuffet, E., Czjzek, M., and Michel, G. (2011). Environmental and gut Bacteroidetes: the food connection. Front. Microbiol. 2:93. doi: 10.3389/fmicb.2011.00093
Tzamaloukas, O., Neofytou, M. C., and Simitzis, P. E. (2021). Application of olive by-products in livestock with emphasis on small ruminants: implications on rumen function, growth performance, milk and meat quality. Animal 11:531. doi: 10.3390/ani11020531
Vaccalluzzo, A., Celano, G., Pino, A., Calabrese, F. M., Foti, P., Caggia, C., et al. (2022). Metagenetic and volatilomic approaches to elucidate the effect of Lactiplantibacillus plantarum starter cultures on Sicilian table olives. Front. Microbiol. 25:771636. doi: 10.3389/fmicb.2021.771636
Vaccalluzzo, A., Pino, A., Russo, N., De Angelis, M., Caggia, C., and Randazzo, C. L. (2020). FoodOmics as a new frontier to reveal microbial community and metabolic processes occurring on table olives fermentation. Food Microbiol. 92:103606. doi: 10.1016/j.fm.2020.103606
Wang, Q., Wang, Y., Wang, X., Dai, C., Tang, W., Li, J., et al. (2020). Effects of dietary energy levels on rumen fermentation, microbiota, and gastrointestinal morphology in growing ewes. Food Sci. Nutr. 8, 6621–6632. doi: 10.1002/fsn3.1955
Welch, C. B., Ryman, V. E., Pringle, T. D., and Lourenco, J. M. (2022). Utilizing the gastrointestinal microbiota to modulate cattle health through the microbiome-gut-organ axes. Microorganisms 10:1391. doi: 10.3390/microorganisms10071391
Xu, Q., Qiao, Q., Gao, Y., Hou, J., Hu, M., Du, Y., et al. (2021). Gut Microbiota and their role in health and metabolic disease of dairy cow. Front. Nutr. 8:701511. doi: 10.3389/fnut.2021.701511
Yang, J., Li, Y., Wen, Z., Liu, W., Meng, L., and Huang, H. (2021). Oscillospira - a candidate for the next-generation probiotics. Gut Microbes 13:1987783. doi: 10.1080/19490976.2021.1987783
Yildirim, E., Ilina, L., Laptev, G., Filippova, V., Brazhnik, E., Dunyashev, T., et al. (2021). The structure and functional profile of ruminal microbiota in young and adult reindeers (Rangifer tarandus) consuming natural winter-spring and summer-autumn seasonal diets. PeerJ 9:e12389. doi: 10.7717/peerj.12389
Keywords: olive by-products, stool samples, metagenomics, microbiota, enzymatic pathways
Citation: Russo N, Floridia V, D’Alessandro E, Lopreiato V, Pino A, Chiofalo V, Caggia C, Liotta L and Randazzo CL (2023) Influence of olive cake dietary supplementation on fecal microbiota of dairy cows. Front. Microbiol. 14:1137452. doi: 10.3389/fmicb.2023.1137452
Received: 04 January 2023; Accepted: 17 April 2023;
Published: 03 May 2023.
Edited by:
Antonio Bevilacqua, University of Foggia, ItalyReviewed by:
Gabriella Siesto, University of Basilicata, ItalyAngela Racioppo, University of Foggia, Italy
Copyright © 2023 Russo, Floridia, D’Alessandro, Lopreiato, Pino, Chiofalo, Caggia, Liotta and Randazzo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alessandra Pino, YWxlc3NhbmRyYS5waW5vQHVuaWN0Lml0
†These authors have contributed equally to this work and share first authorship