 Peng Zhou
Peng Zhou Yu-Xin Bu1
Yu-Xin Bu1 Lin Xu
Lin Xu Xue-Wei Xu
Xue-Wei Xu Hong-Bin Shen
Hong-Bin Shen
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
PERSPECTIVE article
Front. Microbiol. , 13 March 2023
Sec. Extreme Microbiology
Volume 14 - 2023 | https://doi.org/10.3389/fmicb.2023.1111472
This article is part of the Research Topic Adaptation of Halophilic/Halotolerant Microorganisms and Their Applications View all 12 articles
Halotolerant microorganisms have developed versatile mechanisms for coping with saline stress. With the increasing number of isolated halotolerant strains and their genomes being sequenced, comparative genome analysis would help understand the mechanisms of salt tolerance. Six type strains of Pontixanthobacter and Allopontixanthobacter, two phylogenetically close genera, were isolated from diverse salty environments and showed different NaCl tolerances, from 3 to 10% (w/v). Based on the co-occurrence greater than 0.8 between halotolerance and open reading frame (ORF) among the six strains, possible explanations for halotolerance were discussed regarding osmolyte, membrane permeability, transportation, intracellular signaling, polysaccharide biosynthesis, and SOS response, which provided hypotheses for further investigations. The strategy of analyzing genome-wide co-occurrence between genetic diversity and physiological characteristics sheds light on how microorganisms adapt to the environment.
Halotolerance is a relative term that refers to the ability to tolerate salt concentrations higher than those necessary for growth, and microorganisms are considered halotolerant if they survive at high salt concentrations but do not require these conditions for growth (Anton, 2014). With advances in technology, halotolerance mechanisms have been investigated using omics approaches. For instance, comparative transcriptomic and physiological analysis revealed that the halotolerant bacterium Egicoccus halophilus EGI 80432T increased inorganic ions uptake and accumulated trehalose and glutamate in response to moderate salinity condition, while the high salt condition led to up-regulated transcription of genes required for the synthesis of compatible solutes, such as glutamate, histidine, threonine, proline, and ectoine (Chen et al., 2021). The role of glutamate as a key compatible solute for halotolerance was also reported in a halotolerant strain of Staphylococcus saprophyticus based on transcriptome comparison of cells cultivated in media containing different concentrations of NaCl (0, 10, and 20%; Jo et al., 2022). In the exoproteome of the halotolerant bacterium Tistlia consotensis grown at high salinity, proteins associated with osmosensing, exclusion of Na+ and transport of compatible solutes, such as glycine betaine or proline are abundant (Rubiano-Labrador et al., 2015). Similarly, the proteomic analysis of halotolerant nodule endophytes, Rahnella aquatilis strain Ra4 and Serratia plymuthica strain Sp2 identified that different trans-membrane ABC transporters (ATP-binding cassettes) were the most represented among the up-regulated proteins in response to salt stress (Novello et al., 2022). Moreover, the proteome comparison of halotolerant bacterium Staphylococcus aureus under different osmotic stress conditions revealed the differentially expressed proteins (DEPs) involved in fatty acid synthesis, proline/glycine betaine biosynthesis and transportation, stress tolerance, cell wall biosynthesis, and the TCA cycle, which may contribute to the osmotic stress tolerance of S. aureus (Ming et al., 2019). These findings shed light on halotolerance mechanisms. However, halotolerance-related genes may be ignored in transcriptomic and proteomic comparison if there is no significant change in their expression under the experimental conditions.
Genomic comparisons of closely related halotolerant microorganisms can identify genes conserved among species as well as genes that may give an organism its unique characteristics, which helps to understand the mechanisms of salt tolerance. For example, through comparative genome analysis it was uncovered that the members of Acidihalobacter genus contained similar genes for the synthesis and transport of ectoine, as well as genes encoding low affinity potassium pumps. Variations were observed in genes encoding high affinity potassium pumps and proteins involved in the synthesis and/or transport of periplasmic glucans, sucrose, proline, taurine, and glycine betaine (Khaleque et al., 2019). To elucidate salt adaptation strategies in Nitriliruptoria, the genomes of five members from group Nitriliruptoria were analyzed. The results showed that Nitriliruptoria harbor similar synthesis systems of solutes, such as trehalose, glutamine, glutamate, and proline, and on the other hand each member of Nitriliruptoria species possesses specific mechanisms, K+ influx and efflux, betaine and ectoine synthesis, and compatible solutes transport (Chen et al., 2020). Using whole-genome analysis, the halotolerant strains of Martelella soudanensis, NC18T and NC20, were predicted to harbor various halotolerant-associated genes, including K+ uptake protein, K+ transport system, ectoine transport system, glycine betaine transport system, and glycine betaine uptake protein, indicating that strains NC18T and NC20 might tolerate high salinity through the accumulation of potassium ions, ectoine, glycine betaine (Lee and Kim, 2022). Although these findings help to understand the versatile mechanisms of halotolerance existing in halotolerant microbes, genomic comparisons are usually based on genome-wide searches for homologs of known halotolerance-related genes, such as those involved in K+ and Na+ influx and efflux and the synthesis and transport of compatible solutes.
The aim of this perspective is to provide new insights into the development of novel hypotheses and promote further studies on the halotolerance mechanisms. Therefore, co-occurrence analysis between halotolerance and open reading frames (ORFs) was performed to provide intuitive information on halotolerance.
Microorganisms develop abilities that enable them to deal with evolutionary pressure from the environment, such as salinity, temperature, and the power of hydrogen (pH). The phylogenetically closely related strains, which showed similar growth temperature and pH range but different halotolerance, would simplify the analysis. Furthermore, considering the ionic strength of different media may affect the cell growth, the tolerance to NaCl used for co-occurrence analysis should be determined by using same medium. Herein six type strains from two phylogenetically close genera, Pontixanthobacter and Allopontixanthobacter, were chosen for this study. Because of their close phylogenetic relationship, Allopontixanthobacter sediminis and Allopontixanthobacter confluentis have been previously classified as Pontixanthobacter species (Xu et al., 2020; Liu et al., 2021b), and later were reclassified as Allopontixanthobacter species (Xu et al., 2020; Liu et al., 2021a,b). Notably, all the type strains belonging to the two genera were isolated from the Yellow Sea and surrounding areas, but from diverse salty environments, such as Pontixanthobacter aestiaquae KCTC 42006T and Pontixanthobacter rizhaonensis KCTC 62828T from seawater (Jung et al., 2014; Liu et al., 2021b), Pontixanthobacter gangjinensis JCM 17802T and Pontixanthobacter luteolus KCTC 12311T from tidal flat (Yoon et al., 2005; Jeong et al., 2013), Pontixanthobacter aquaemixtae KCTC 52763T from the junction between ocean and fresh spring (Park et al., 2017), A. sediminis KCTC 42453T from lagoon sediments (Kim et al., 2016), and A. confluentis KCTC 52259T from water of estuary environment (Park et al., 2017). These strains showed similar optimum NaCl concentrations for growth (1–3%, w/v), but displayed different halotolerances, from 3 to 10% (w/v; Table 1), indicating that these strains adapt to their diverse habitats, including lagoon, junction between ocean and fresh spring, tidal flat, and seawater. The availability of their genomes provides remarkable opportunity to understand their different halotolerances by comparative genome analysis. Here, co-occurrence between halotolerance and the open reading frames (ORFs) was calculated among six strains of Pontixanthobacter and Allopontixanthobacter, and the ORFs showing high co-occurrence were discussed for possible contribution to halotolerance.
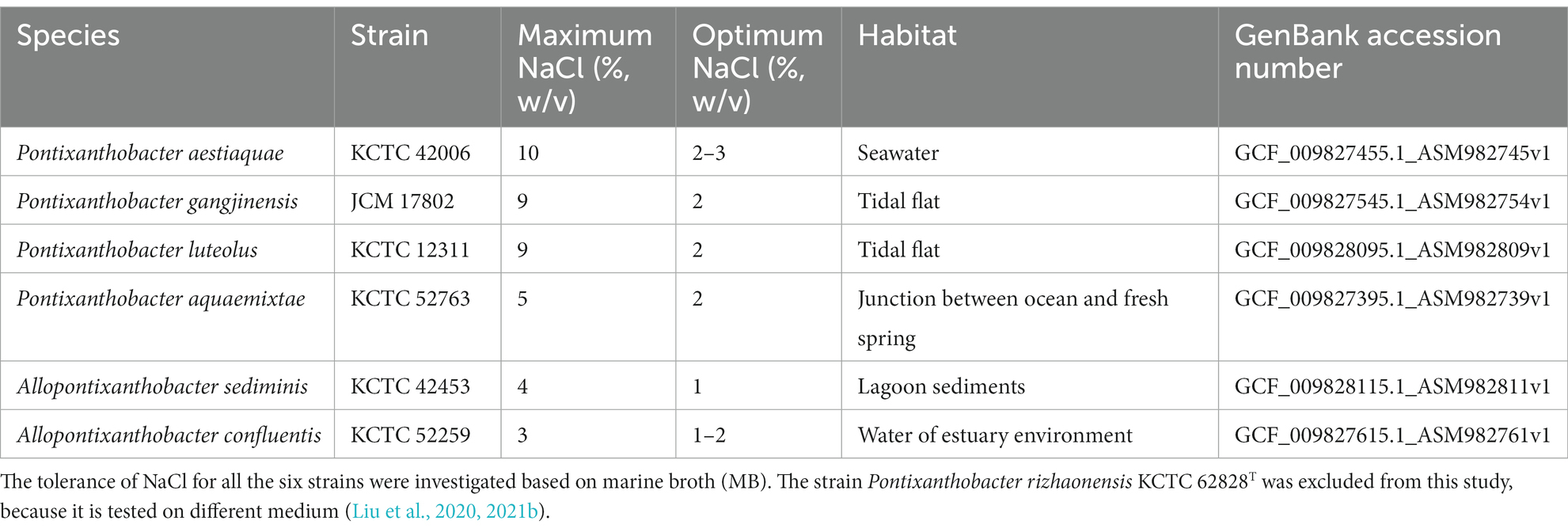
Table 1. Strains used for analysis in this study.
Open reading frames in the six genomes were predicted and clustered based on similarity using R package micropan (Snipen and Liland, 2015). Analysis of co-occurrence between ORFs and the maximum NaCl concentration tolerated among the six strains was conducted, and 113 clusters of ORFs were identified with co-occurrence greater than 0.8 (Table 2). The co-occurrence for the remaining clusters is listed in Supplementary material, as well as ORFs predicted in the six genomes and the index for clusters and ORFs. ORFs were annotated by searching standard database using protein–protein BLAST.1
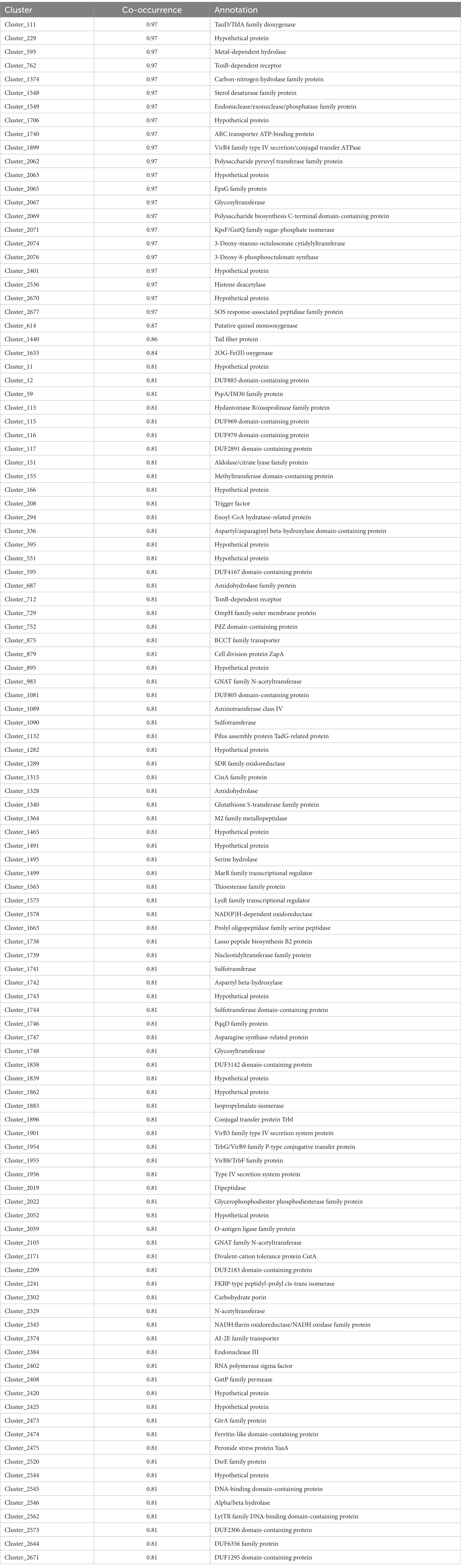
Table 2. Clusters highly co-occurred with halotolerance.
The ORFs of Cluster_111 (co-occurrence of 0.97, Table 2) were annotated as TauD/TfdA family dioxygenase. TauD is involved in the utilization of taurine (VanderPloeg et al., 1996), an organic osmolyte involved in cell volume regulation (Harris and Wen, 2012). Taurine is used as an osmoprotectant, such as in Escherichia coli at high osmolarity (McLaggan and Epstein, 1991) and in microbial communities from biofilms in metal-rich environment (Mosier et al., 2013). The ORFs of Cluster_111 only exist in three halotolerant strains, suggesting that taurine may be accumulated as an osmoprotectant. Interestingly, halotolerant strains harbor genes involved in various pathways related to glutamate generation. For instance, according to annotation, ORFs of Cluster_113 (co-occurrence of 0.81, Table 2) belong to the hydantoinase B/oxoprolinase family, which includes 5-oxoprolinase, catalyzing the formation of L-glutamate from 5-oxo-L-proline (Niehaus et al., 2017). Besides, ORFs of Cluster_1328 (co-occurrence of 0.81, Table 2) possess similarity to p-aminobenzoyl-glutamate (PABA-GLU) hydrolase subunit from Altererythrobacter insulae (GenBank Accession Number: RGP41665.1). PABA-GLU is a folate catabolite found in bacteria, and the enzyme PABA-GLU hydrolase breaks down PABA-GLU by cleaving glutamate (Larimer et al., 2014). Additionally, ORFs of Cluster_1747 (co-occurrence of 0.81 Table 2) showed similarity to asparagine synthase from Salinigranum halophilum (GenBank Accession Number: WP_136601134.1). Asparagine synthetase catalyzes an ATP-dependent amidotransferase reaction between aspartate and glutamine, which produces asparagine and glutamate (Richards and Kilberg, 2006).
To ensure a physiologically acceptable level of cellular hydration and turgor at high osmolarity, many bacteria accumulate compatible solutes as osmoprotectants (Ziegler et al., 2010). ORFs of Cluster_875 (co-occurrence of 0.81, Table 2) were annotated as proteins of Betaine/Carnitine/Choline Transporter (BCCT) family. The BCCT family includes transporters for carnitine, choline and glycine betaine, and some of which exhibit osmosensory and osmoregulatory properties (Ziegler et al., 2010). Furthermore, the ORFs of Cluster_1740, annotated as ABC transporter ATP-binding proteins, were present only in these three halotolerant strains. The salt-induced ABC transporter Ota from Methanosarcina mazei Gö1 acts as a glycine betaine transporter (Schmidt et al., 2007). Another ABC transporter in Listeria, OpuC, is shown to be necessary for glycine betaine and choline chloride uptake (Verheul et al., 1997). Compared to the wild type of S. aureus, mutating OpuC did reduce their ability to grow under osmotic stress (10% NaCl; Kiran et al., 2009). The function of ORFs of Cluster_1740 and their contribution to halotolerance can be further characterized. Additionally, previous studies have shown that water permeability is clearly affected by the number of double bonds in the fatty acid conjugates of lipids, the higher the degree of unsaturation, the greater the water permeability (Graziani and Livne, 1972), and sterol type is one of the determining factors in the permeability of membranes to small solutes (Frallicciardi et al., 2022). The genomes of three halotolerant strains contain ORFs of Cluster_1548, annotated as sterol desaturase family proteins, indicating that sterols might be used to change permeability.
Cluster_1549 also consists of three ORFs present in the three halotolerant strains, which showed similarity to the domain superfamily found in a large number of proteins including magnesium dependent endonucleases and phosphatases involved in intracellular signaling (Dlakic, 2000). Its role in the regulation of gene expression, such as triggering the salt-stress response, is worth of further study.
It has been reported that extracellular polysaccharides (EPS) may influence the salt tolerance of certain rhizobial strains (Samir and Kanak, 1997) and the lipopolysaccharide pattern could alter according to different salinities in a salt-tolerant strain of Mesorhizobium cicero (Soussi et al., 2001). All three halotolerant strains harbor ORFs annotated with polysaccharide/lipopolysaccharide biosynthesis (Cluster_2062, 2065, 2067, 2069, 2071, 2074, and 2076 in Table 2), such as 3-deoxy-d-manno-octulosonate cytidylyltransferase, a key enzyme in the biosynthesis of lipopolysaccharide (LPS) in Gram-negative organisms (Yi et al., 2011). Furthermore, ORFs of Cluster_2473 (co-occurrence as 0.81 Table 2) were annotated to encode proteins of the GtrA family, whose members are often involved in the synthesis of cell surface polysaccharides (Kolly et al., 2015).
Open reading frames of Cluster_2677 are annotated encoding SOS response-associated peptidase family protein. The bacterial SOS response induced under stress conditions is recruited to DNA repair and adaptive mutagenesis (Shinagawa, 1996; Aravind et al., 2013). Hence, ORFs of Cluster_2677 could be further investigated for its importance to halotolerance.
Salinity is one of the most important environmental factors for aquatic microorganisms and varies among habitats. Therefore, halotolerant microorganisms have developed versatile strategies to cope with saline stress. Based on the findings of co-occurrence analysis, possible explanations for mechanisms resulting in different salt tolerances among six strains are discussed above, which provided hypotheses for further investigations. Moreover, among the highly co-occurred clusters, there are several uncharacterized or hypothetical proteins (Table 2), which may contribute to halotolerance. It should be noted that the genes related to resistance to salts other than sodium chloride could also be discovered by co-occurrence analysis, since various salts co-exist in high ionic environments. For instance, ORFs of Cluster_2171 (co-occurrence as 0.81, Table 2) were annotated as divalent-cation tolerance protein CutA, which is required for copper tolerance in E. coli and affects tolerance levels to zinc, nickel, cobalt, and cadmium salts (Fong et al., 1995). This study sheds light on the mechanisms through which microorganisms cope with environmental stress. With the increasing number of isolated halotolerant strains and their genomes being sequenced, analyzing genome-wide co-occurrence between genetic diversity and physiological characteristics would expand the knowledge of the salinity adaptation strategies and provide comprehensive information on how microorganisms adapt to the environment, together with findings at the transcriptomic and proteomic levels.
Publicly available datasets were analyzed in this study. This data can be found here: https://www.ncbi.nlm.nih.gov. Accession Numbers are as follows: GCF_009827455.1_ASM982745v1, GCF_009827545.1_ASM982754v1, GCF_009828095.1_ASM982809v1, GCF_009827395.1_ASM982739v1, GCF_009828115.1_ASM982811v1, and GCF_009827615.1_ASM982761v1.
PZ contributed to study concept and design and performed data acquisition, analysis and visualization, and interpretation of results. PZ and Y-XB drafted the manuscript. LX, X-WX, and H-BS revised the manuscript. All authors contributed to the article and approved the submitted version.
This work was supported by grants from the National Science and Technology Fundamental Resources Investigation Program of China (2021FY100900), the Scientific Research Fund of the Second Institute of Oceanography, MNR (No. JZ1901 and JB2003), Natural Science Foundation of China (32000001), and the Oceanic Interdisciplinary Program of Shanghai Jiao Tong University (SL2022ZD108).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2023.1111472/full#supplementary-material
Anton, J. (2014). “Halotolerance” in Encyclopedia of Astrobiology. eds. R. Amils, M. Gargaud, J. C. Quintanilla, H. J. Cleaves, W. M. Irvine, and D. Pinti, et al. (Berlin, Heidelberg: Springer Berlin Heidelberg), 1–2.
Aravind, L., Anand, S., and Iyer, L. M. (2013). Novel autoproteolytic and DNA-damage sensing components in the bacterial SOS response and oxidized methylcytosine-induced eukaryotic DNA demethylation systems. Biol. Direct 8:20. doi: 10.1186/1745-6150-8-20
Chen, D. D., Fang, B. Z., Manzoor, A., Liu, Y. H., Li, L., Mohamad, O. A. A., et al. (2021). Revealing the salinity adaptation mechanism in halotolerant bacterium Egicoccus halophilus EGI 80432(T) by physiological analysis and comparative transcriptomics. Appl. Microbiol. Biotechnol. 105, 2497–2511. doi: 10.1007/s00253-021-11190-5
Chen, D. D., Tian, Y., Jiao, J. Y., Zhang, X. T., Zhang, Y. G., Dong, Z. Y., et al. (2020). Comparative genomics analysis of Nitriliruptoria reveals the genomic differences and salt adaptation strategies. Extremophiles 24, 249–264. doi: 10.1007/s00792-019-01150-3
Dlakic, M. (2000). Functionally unrelated signalling proteins contain a fold similar to Mg2+−dependent endonucleases. Trends Biochem. Sci. 25, 272–273. doi: 10.1016/s0968-0004(00)01582-6
Fong, S.-T., Camakaris, J., and Lee, B. T. O. (1995). Molecular genetics of a chromosomal locus involved in copper tolerance in Escherichia coli K-12. Mol. Microbiol. 15, 1127–1137. doi: 10.1111/j.1365-2958.1995.tb02286.x
Frallicciardi, J., Melcr, J., Siginou, P., Marrink, S. J., and Poolman, B. (2022). Membrane thickness, lipid phase and sterol type are determining factors in the permeability of membranes to small solutes. Nat. Commun. 13:1605. doi: 10.1038/s41467-022-29272-x
Graziani, Y., and Livne, A. (1972). Water permeability of bilayer lipid membranes: sterol-lipid interaction. J. Membr. Biol. 7, 275–284. doi: 10.1007/BF01867920
Harris, R., and Wen, S. (2012). Review: Taurine: A “very essential” amino acid. Mol Vis. 18, 2673–2686.
Jeong, S. H., Jin, H. M., Lee, H. J., and Jeon, C. O. (2013). Altererythrobacter gangjinensis sp. nov., a marine bacterium isolated from a tidal flat. Int. J. Syst. Evol. Microbiol. 63, 971–976. doi: 10.1099/ijs.0.039024-0
Jo, E., Hwang, S., and Cha, J. (2022). Transcriptome analysis of Halotolerant Staphylococcus saprophyticus isolated from Korean fermented shrimp. Foods 11:524. doi: 10.3390/foods11040524
Jung, Y. T., Park, S., Lee, J. S., and Yoon, J. H. (2014). Altererythrobacter aestiaquae sp. nov., isolated from seawater. Int. J. Syst. Evol. Microbiol. 64, 3943–3949. doi: 10.1099/ijs.0.066639-0
Khaleque, H. N., Gonzalez, C., Shafique, R., Kaksonen, A. H., Holmes, D. S., and Watkin, E. L. J. (2019). Uncovering the mechanisms of Halotolerance in the extremely acidophilic members of the Acidihalobacter genus through comparative genome analysis. Front. Microbiol. 10:155. doi: 10.3389/fmicb.2019.00155
Kim, J. H., Yoon, J. H., and Kim, W. (2016). Altererythrobacter sediminis sp. nov., isolated from lagoon sediments. Int. J. Syst. Evol. Microbiol. 66, 5424–5429. doi: 10.1099/ijsem.0.001535
Kiran, M. D., Akiyoshi, D. E., Giacometti, A., Cirioni, O., Scalise, G., and Balaban, N. (2009). OpuC—an ABC transporter that is associated with Staphylococcus aureus pathogenesis. Int. J. Artif. Organs 32, 600–610. doi: 10.1177/039139880903200909
Kolly, G. S., Mukherjee, R., Kilacskova, E., Abriata, L. A., Raccaud, M., Blasko, J., et al. (2015). GtrA protein Rv3789 is required for Arabinosylation of Arabinogalactan in Mycobacterium tuberculosis. J. Bacteriol. 197, 3686–3697. doi: 10.1128/JB.00628-15
Larimer, C. M., Slavnic, D., Pitstick, L. D., and Green, J. M. (2014). Comparison of substrate specificity of Escherichia coli p-Aminobenzoyl-glutamate hydrolase with Pseudomonas Carboxypeptidase G. Adv. Enzyme Res. 2, 39–48. doi: 10.4236/aer.2014.21004
Lee, J. Y., and Kim, D. H. (2022). Genomic analysis of Halotolerant bacterial strains Martelella soudanensis NC18(T) and NC20. J. Microbiol. Biotechnol. 32, 1427–1434. doi: 10.4014/jmb.2208.08011
Liu, A., Xue, Q. J., Li, S. G., and Zhang, Y. J. (2021a). Corrigendum: Pontixanthobacter rizhaonensis sp. nov., a marine bacterium isolated from surface seawater of the Yellow Sea, and the proposal of Pseudopontixanthobacter gen. Nov., Pseudopontixanthobacter confluentis comb. nov. and Pseudopontixanthobacter sediminis comb. nov. Int. J. Syst. Evol. Microbiol. 71:004931. doi: 10.1099/ijsem.0.004931
Liu, A., Xue, Q. J., Li, S. G., and Zhang, Y. J. (2021b). Pontixanthobacter rizhaonensis sp. nov., a marine bacterium isolated from surface seawater of the Yellow Sea, and proposal of Pseudopontixanthobacter gen. Nov., Pseudopontixanthobacter confluentis comb. nov. and Pseudopontixanthobacter sediminis comb. nov. Int. J. Syst. Evol. Microbiol. 71:004780. doi: 10.1099/ijsem.0.004780
Liu, A., Zhang, Y. J., Xue, Q. J., Wang, H., Yang, Y. Y., Du, F., et al. (2020). Litorilituus lipolyticus sp. nov., isolated from intertidal sand of the Yellow Sea in China, and emended description of Colwellia asteriadis. Antonie Van Leeuwenhoek 113, 449–458. doi: 10.1007/s10482-019-01355-8
McLaggan, D., and Epstein, W. (1991). Escherichia coli accumulates the eukaryotic osmolyte taurine at high osmolarity. FEMS Microbiol. Lett. 81, 209–213. doi: 10.1016/0378-1097(91)90304-s
Ming, T., Geng, L., Feng, Y., Lu, C., Zhou, J., Li, Y., et al. (2019). iTRAQ-based quantitative proteomic profiling of Staphylococcus aureus under different osmotic stress conditions. Front. Microbiol. 10:1082. doi: 10.3389/fmicb.2019.01082
Mosier, A. C., Justice, N. B., Bowen, B. P., Baran, R., Thomas, B. C., Northen, T. R., et al. (2013). Metabolites associated with adaptation of microorganisms to an acidophilic, metal-rich environment identified by stable-isotope-enabled metabolomics. MBio 4, e00484–e00412. doi: 10.1128/mBio.00484-12
Niehaus, T. D., Elbadawi-Sidhu, M., de Crecy-Lagard, V., Fiehn, O., and Hanson, A. D. (2017). Discovery of a widespread prokaryotic 5-oxoprolinase that was hiding in plain sight. J. Biol. Chem. 292, 16360–16367. doi: 10.1074/jbc.M117.805028
Novello, G., Gamalero, E., Massa, N., Cesaro, P., Lingua, G., Todeschini, V., et al. (2022). Proteome and physiological characterization of Halotolerant nodule Endophytes: the case of Rahnella aquatilis and Serratia plymuthica. Microorganisms 10:890. doi: 10.3390/microorganisms10050890
Park, S., Jung, Y. T., Choi, S. J., and Yoon, J. H. (2017). Altererythrobacter aquaemixtae sp. nov., isolated from the junction between the ocean and a freshwater spring. Int. J. Syst. Evol. Microbiol. 67, 3446–3451. doi: 10.1099/ijsem.0.002136
Richards, N. G., and Kilberg, M. S. (2006). Asparagine synthetase chemotherapy. Annu. Rev. Biochem. 75, 629–654. doi: 10.1146/annurev.biochem.75.103004.142520
Rubiano-Labrador, C., Bland, C., Miotello, G., Armengaud, J., and Baena, S. (2015). Salt stress induced changes in the exoproteome of the Halotolerant bacterium Tistlia consotensis deciphered by Proteogenomics. PLoS One 10:e0135065. doi: 10.1371/journal.pone.0135065
Samir, K. M., and Kanak, R. S. (1997). The correlation between salt tolerance and extracellular Polysaccharid. Microbes Environ. 12, 9–13.
Schmidt, S., Pfluger, K., Kogl, S., Spanheimer, R., and Muller, V. (2007). The salt-induced ABC transporter Ota of the methanogenic archaeon Methanosarcina mazei Go1 is a glycine betaine transporter. FEMS Microbiol. Lett. 277, 44–49. doi: 10.1111/j.1574-6968.2007.00938.x
Shinagawa, H. (1996). SOS response as an adaptive response to DNA damage in prokaryotes. EXS 77, 221–235. doi: 10.1007/978-3-0348-9088-5_14
Snipen, L., and Liland, K. H. (2015). Micropan: an R-package for microbial pan-genomics. BMC Bioinf. 16:79. doi: 10.1186/s12859-015-0517-0
Soussi, M., Santamaria, M., Ocana, A., and Lluch, C. (2001). Effects of salinity on protein and lipopolysaccharide pattern in a salt-tolerant strain of Mesorhizobium ciceri. J. Appl. Microbiol. 90, 476–481. doi: 10.1046/j.1365-2672.2001.01269.x
VanderPloeg, J. R., Weiss, M. A., Saller, E., Nashimoto, H., Saito, N., Kertesz, M. A., et al. (1996). Identification of sulfate starvation-regulated genes in Escherichia coli: a gene cluster involved in the utilization of taurine as a sulfur source. J. Bacteriol. 178, 5438–5446. doi: 10.1128/jb.178.18.5438-5446.1996
Verheul, A., Glaasker, E., Poolman, B., and Abee, T. (1997). Betaine and L-carnitine transport by Listeria monocytogenes Scott a in response to osmotic signals. J. Bacteriol. 179, 6979–6985. doi: 10.1128/jb.179.22.6979-6985.1997
Xu, L., Sun, C., Fang, C., Oren, A., and Xu, X. W. (2020). Genomic-based taxonomic classification of the family Erythrobacteraceae. Int. J. Syst. Evol. Microbiol. 70, 4470–4495. doi: 10.1099/ijsem.0.004293
Yi, L., Velasquez, M. S., Holler, T. P., and Woodard, R. W. (2011). A simple assay for 3-deoxy-d-manno-octulosonate cytidylyltransferase and its use as a pathway screen. Anal. Biochem. 416, 152–158. doi: 10.1016/j.ab.2011.05.022
Yoon, J. H., Kang, K. H., Yeo, S. H., and Oh, T. K. (2005). Erythrobacter luteolus sp. nov., isolated from a tidal flat of the Yellow Sea in Korea. Int. J. Syst. Evol. Microbiol. 55, 1167–1170. doi: 10.1099/ijs.0.63522-0
Keywords: halotolerance, co-occurrence, comparative genomics, Erythrobacteraceae, adaptation
Citation: Zhou P, Bu Y-X, Xu L, Xu X-W and Shen H-B (2023) Understanding the mechanisms of halotolerance in members of Pontixanthobacter and Allopontixanthobacter by comparative genome analysis. Front. Microbiol. 14:1111472. doi: 10.3389/fmicb.2023.1111472
Edited by:
Rosa María Martínez-Espinosa, University of Alicante, SpainReviewed by:
Kesava Priyan Ramasamy, Nanyang Technological University, SingaporeCopyright © 2023 Zhou, Bu, Xu, Xu and Shen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Peng Zhou, emhvdXBlbmdAc2lvLm9yZy5jbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.