 Xiaobing Guo
Xiaobing Guo Ruyan Chen
Ruyan Chen Qian Wang
Qian Wang Chenyu Li
Chenyu Li Haoyu Ge
Haoyu Ge Jie Qiao1
Jie Qiao1
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol., 06 September 2022
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 13 - 2022 | https://doi.org/10.3389/fmicb.2022.979558
This article is part of the Research TopicCommunity Series - Characterization of mobile genetic elements associated with acquired resistance mechanisms, Volume IIView all 6 articles
IncX3 plasmids are narrow host range plasmids mostly found in Enterobacteriaceae with great conjugation ability, high stability, no fitness cost, and the ability to improve biofilm formation in their bacterial hosts. IncX3 plasmids have spread swiftly, primarily in several nations and among different species over the last 10 years. blaNDM, blaKPC, and blaOXA-181 are the carbapenemase genes carried by IncX3 plasmids. Among them, blaNDM is often located on the IncX3 plasmid, which is deemed as the primary vehicle of blaNDM transmission. Isolates harboring IncX3 plasmids are found in nations all over the world from human, animal, and environmental sources. Cointegrate plasmids related to IncX3 have recently been discovered to increase the antibiotic resistance spectrum and potentially broaden the host range of plasmids, restricting the use of antibiotics in the clinic. There are, however, few reviews based on the physiological and epidemiological properties of IncX3 plasmid, as well as studies on the plasmid itself. Hence, we conducted a retrospective literature review to summarize the characteristics of IncX3 plasmids aiming to provide a theoretical basis for controlling the global prevalence of IncX3 plasmids and directions for further research on the functions of the related genes on the IncX3 plasmid.
Carbapenem antibiotics are often the last line of defense in the treatment of multidrug-resistant Gram-negative bacteria. However, with the widespread use of antibiotics, carbapenem-resistant Enterobacteriaceae (CRE) have become common and pose a great threat to human health. Phenotypic resistance to carbapenems is complex and mainly caused by four mechanisms, including (1) β-lactamase activity combined with structural mutations; (2) production of carbapenemases, enzymes that hydrolyze carbapenem antibiotics; (3) activation and massive expression of drug efflux pump system; and (4) alterations in penicillin-binding proteins. The most typical mechanism is carbapenemase production (KPC, NDM, VIM, IMP, and OXA-48; Brinkac et al., 2019).
Clonal spread and horizontal transmission are the two most common carbapenemase-producing Enterobacteriaceae (CPE) dissemination methods. Compared with chromosomes, genes encoding carbapenemases are generally found in incompatible plasmid groups (Poirel et al., 2011). Horizontal transfer of resistance genes via mobile plasmids can accelerate the spread of resistance genes in diverse strains and hosts.
Plasmid incompatibility refers to the inability of two plasmids to coexist in the same cell, resulting in the loss of one plasmid in daughter cells, which is determined by replication machinery. In Enterobacteriaceae, 27 main plasmid incompatibility groups have been linked to antibiotic resistance genes (ARGs). A number of plasmid replicon groups in CREs have been reported globally, such as IncF, N, X, A/C, L/M, R, P, H, I, and W. Compared with other Inc. groups, IncF, A/C, and X are the most commonly associated with carbapenemase production.
IncX plasmids appear to be a potential reservoir for various resistance combinations, decreasing susceptibility to clinically important antimicrobials (Juraschek et al., 2021). IncX plasmids can be divided into nine subgroups based on repA sequences and binding sites (IncX1α, IncX1β, and IncX2-IncX8) (Fang et al., 2018). Among the IncX plasmid family, the IncX3 subgroup is primarily responsible for the dissemination of ARGs for clinically relevant first-line and last-resort (carbapenems) antibiotics (Liakopoulos et al., 2018). The prevalence of IncX3 plasmids was underestimated due to a lack of sequences available for setting PCR-based replicon typing (PBRT) until the taxC gene was used as a target to successfully identify the first IncX3 plasmid that represents the prototype IncX3 plasmid (Johnson et al., 2012). Then, until 2011, the first relevant properties of IncX3 plasmid pNDM-HN380 carrying blaNDM were recorded, which was one of the early steps in the evolution and spread of blaNDM (Ho et al., 2012). IncX3 plasmids have been implicated in the distribution of a range of blaNDM in humans, animals, and the environment in the last 10 years, particularly in Southeast Asia (Mouftah et al., 2019). IncX3 plasmid is an important vector carrying blaNDM (Wu et al., 2019; Touati et al., 2021). Furthermore, IncX3 is the most common subgroup found to harbor blaKPC and blaNDM, the two most prevalent carbapenemase genes (Kopotsa et al., 2019). The IncX3 plasmid has high stability, low fitness cost, great conjugation ability, and strong biofilm formation ability (Ma et al., 2020), all of which can facilitate the rapid and dominant dissemination of ARGs.
However, current research on IncX3 plasmids mainly focus on the epidemiological and molecular characteristics, while the physiological features of these plasmids are still unclear. Therefore, the purpose of this review is to (1) summarize the global epidemiological and molecular characteristics of IncX3 over the last 10 years, providing a theoretical basis for controlling the spread of IncX3 plasmids; and (2) organize the literature that have studied the physiological characteristics of IncX3 plasmids, providing directions and ideas for future IncX3 plasmid research.
Publications chosen for this review were found on PubMed using the keywords “IncX3” as search criteria. As of May 21, 2022, a total of 273 articles were retrieved.
Isolates harboring IncX3 plasmids are found in many countries around the world (Figure 1), including China (CHN), South Korea (KOR), Switzerland (SWI), Czech (CZE), Italy (ITA), Australia (AUS), India (IND), Japan (JPN), Netherlands (NED), Brazil (BRA), Germany (GER), South Africa (ZAF), Myanmar (MYA), the United States (USA), France (FRA), Ghana (GHA), Spain (ESP), the United Arab Emirates (AE), Cambodia (CAM), Canada (CAN), Denmark (DEN), Egypt (EGY), Kuwait (KUW), Pakistan (PAK), Poland (POL), Portugal (POR), the United Kingdom (UK), Oman (OMA), Vietnam (VIE), Algeria (DZA), Angola (AGO), Burkina Faso (BFA), Chad (TCD), Nigeria (NGA), Tanzania (TZA; Supplementary material), Tunisia (TUN; Dziri et al., 2022), Lebanon (LBN; Dagher et al., 2018), Ethiopia (ETH; Legese et al., 2022), Croatia (HRV; Petrosillo et al., 2016), Turkey (TUR; Peirano et al., 2022), and Gabon (GAB; Moussounda et al., 2017). Among them, China is the country with the most reported IncX3 plasmids.
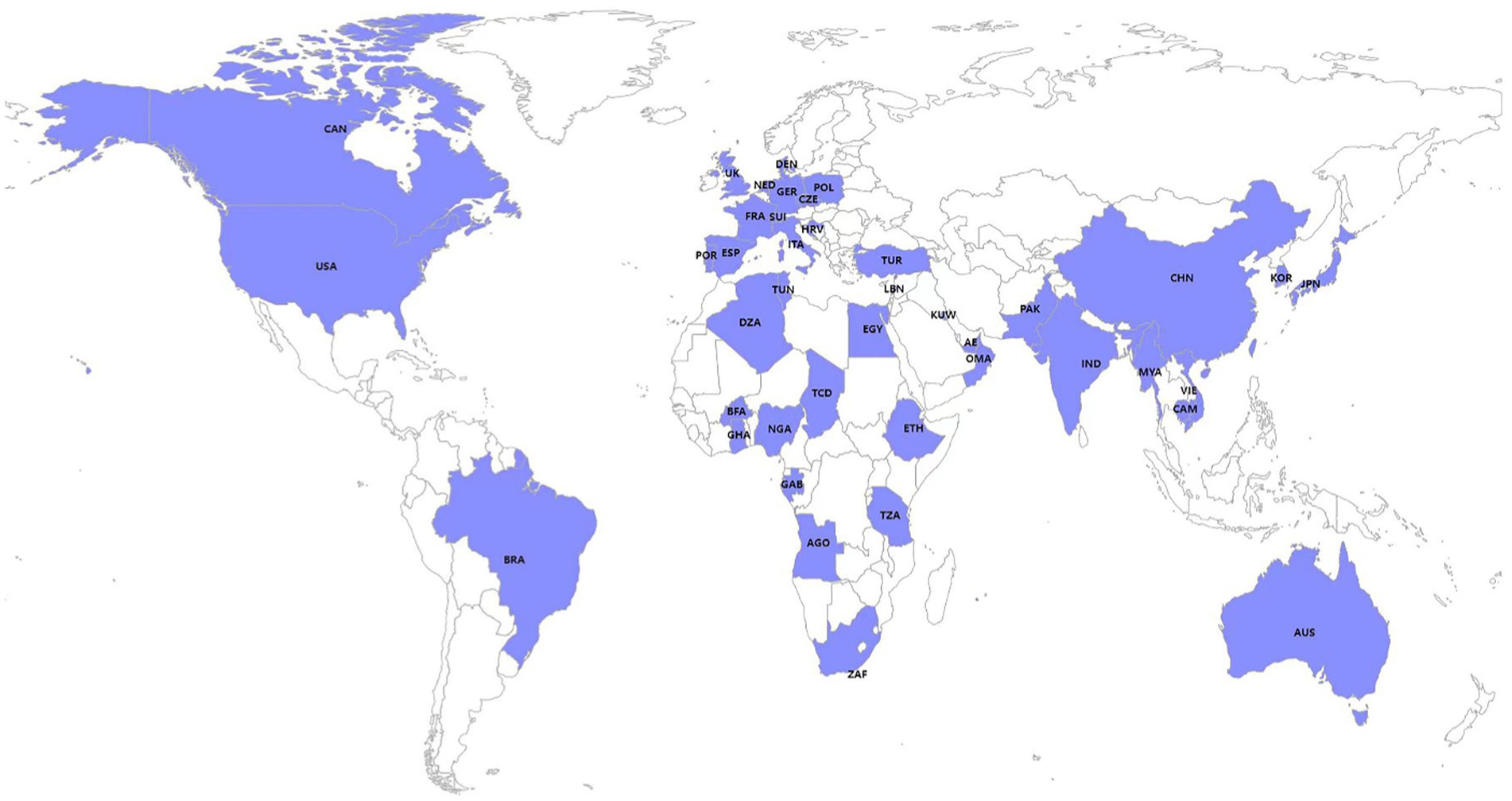
Figure 1. Global distribution of published strains harboring IncX3 plasmids included in this study.
The strains carrying IncX3 plasmids are from a rich source, mainly divided into clinical and non-clinical sources, with clinical sources accounting for two-thirds of the total (Supplementary material). Among them, urine ranked first, followed by blood, rectal swab, sputum, and stool specimens. In addition to the common specimens, clinical sources also include cerebrospinal fluid, bile, tracheal aspirate, drainage fluid, and wound. Non-clinical sources include livestock, environment (such as sewage treatment plants, rivers, and subway surfaces), companion animals, and food chains. Focusing on non-clinical specimens can help limit the threat of horizontal transfer to human health and take steps to regulate transmission routes and slow the global spread of antibiotic resistance genes.
One Health advocates that we should pay more attention to the interaction between humans, animals, and the shared environment, to achieve the best health outcomes, which is an important reason why we need to pay equal attention to the characteristics of IncX3 plasmids from non-clinical sources. Several investigations have shown interactions to mediate the spread of IncX3 plasmids in humans, animals, and the environment, resulting in the spread of resistance determinants. The blaNDM-5-producing Escherichia coli ST167 in both owners and companion animals suggests potential transmission between humans and dogs (Grönthal et al., 2018). IncX3 sequences in a blaNDM-5-positive strain isolated from a cat had areas with >99% nucleotide sequence identity to the human reference plasmid pNDM-MGR194 (Zhang et al., 2022). Furthermore, while carbapenems are not permitted for use in food-producing animals in China, the appearance of blaNDM-5-bearing IncX3 plasmids in swine-origin K. pneumoniae could be owing to environmental factors or human-to-livestock transmission (Zhao et al., 2021), but this hypothesis needs further confirmation. Meanwhile, the environment is also a potential reservoir for CRE as well as an essential source of transmission (Gu et al., 2022).
There are 23 species in the host of IncX3 plasmid (Supplementary material), including Escherichia coli, Klebsiella pneumonia, Citrobacter freundii, Enterobacter cloacae, Citrobacter braakii, Citrobacter portucalensis, Citrobacter sedlakii, Cronobacter sakazakii, Enterobacter asburiae, Enterobacter hormaechei, Klebsiella quasipneumoniae, Klebsiella aerogenes, Klebsiella oxytoca, Klebsiella variicola, Kluyvera cryocrescens, Kluyvera intermedia, Morganella morganii, Proteus mirabilis, Raoultella ornithinolytica, Salmonella Typhimurium, Raoultella planticola, and Serratia marcescens. Compared with other species, E. coli, K. pneumonia, C. freundii, and E. cloacae are the most commonly associated with IncX3 plasmid.
Among IncX3 plasmid-containing Escherichia coli and Klebsiella pneumoniae isolates, we discovered a surprising diversity of sequence types (STs). The most popular E. coli with distinct sequence types are ST410, ST167, ST48, and ST101. In Chinese clinical settings, ST167 E. coli strains had close ties to blaNDM-5, and high-level tigecycline resistance Escherichia coli carrying blaNDM-5 also belonged to the ST167 clonal lineage (Li et al., 2018). This suggests that the ST167 is a major blaNDM-5 reservoir in China. It is worth noting that ST48 only involves livestock sources, while ST101 only involves clinical sources. ST410 also includes clinical, animal, and environmental sources. The ST410 Escherichia coli carries diverse ARGs, indicating ST410 Escherichia coli is widely distributed and has a robust transmission and pathogenic potential. In Klebsiella pneumoniae, the most popular types are ST307, ST11, and ST15.
IncX3 plasmids are narrow host range plasmids of Enterobacteriaceae (Zhu et al., 2020). The backbone and accessory module are the two main parts of the structure of the IncX3 plasmid. The core genes of the backbone include plasmid replication-related genes, conjugation-related genes, entry exclusion system-related genes, and partitioning system-related genes. The core genes are shared among the three typical completely sequenced IncX3 plasmids (pEC13_35, pIncX-SHV, and pNDM-HN380): replication (replication initiation protein, pir; replication accessory protein, bis), partitioning (parA and parB), plasmid maintenance (a putative DNA-binding protein, hns; a putative type III topoisomerase, topB), conjugation/type IV secretion system (T4SS, with 11 genes, pilX1 to pilX11), transcriptional activator (actX), and putative DNA transfer proteins (taxA and taxC) (Ho et al., 2012). A retrospective analysis of IncX3 in China shows that the IncX3 plasmid backbone has been highly conserved over the past decade (Liu et al., 2021a). The accessory module mainly includes transposase genes, metabolic functions related genes, and antimicrobial resistance encoding genes. These are identified as acquired DNA sections coupled with and bordered by mobile elements and inserted at various backbone sites (Shi et al., 2018). In a study of IncX3 plasmids harboring blaNDM-5 in retail beef in China (Zhang et al., 2019), a total of seven IncX3 genetic backgrounds were discovered. The variation is primarily due to mobile element insertions, truncation, and/or deletions. The conserved sequence of type I is IS3000-ΔISAba125-IS5-blaNDM-bleMBL-trpF-tat-Δdct-IS26-ΔumuD, which is the most common structure in Enterobacteriaceae of human, pig, and chicken sources in China and other countries. This shows that distinct mobile components are primarily responsible for the variety of the genomic environment in the IncX3 plasmid.
IncX3 plasmids have lower fitness costs. A study using the typical IncX3 plasmid pNDM-HN380 suggests that the IncX3 plasmid affected the motility of the host in addition to conferring multiple antibiotic resistance. Furthermore, the plasmid-encoded new sRNA IGR plas2 can influence fucose metabolism and biofilm formation to increase strain survival in a competitive environment (Huang et al., 2020). To our understanding, plasmids carrying resistance genes in strains impose fitness costs on their hosts in the absence of antibiotic selection pressure (Andersson and Hughes, 2010). However, in Ma’s study, 75.9% of wild-type strains receiving IncX3 plasmid demonstrated no fitness cost as a result of plasmid acquisition, and 6.9% of strains showed improved growth above their wild-type counterparts (Ma et al., 2020). The low fitness cost of the IncX3 plasmid may contribute to the stability of host bacteria and facilitate the horizontal transfer.
IncX3 plasmids are highly stable. In serial passage tests, the IncX3 plasmid is stable over time and confers a fitness benefit in selective antibiotic pressure (Flerlage et al., 2020). After 100 serial generations, IncX3 plasmids demonstrate remarkable stability in both clinical isolates and transconjugants, with no apparent plasmid loss (Tian et al., 2020). The IncX3 blaNDM-5-carrying plasmids in one K. pneumoniae clinical isolate showed high stability in clinical isolates, with no apparent plasmid loss after repeated cultures for 5 days (Zhu et al., 2020). Additionally, two blaNDM-5 were found in Escherichia coli and were shown to be localized to the IncX3 and IncFII plasmids, respectively, in a study. Transconjugants carrying the IncX3 plasmid had higher levels of carbapenem resistance and blaNDM-5 expression than those carrying both plasmids or only carrying the IncFII plasmid (Yang et al., 2020).
IncX3 plasmids are conjugative and could enhance biofilm formation. Research suggests that IncX3 plasmid can transfer blaNDM between different enterobacterial species over a wide temperature range (Liu et al., 2019). And this is attributed to the type IV secretion system encoded by the IncX3 plasmid, which, in addition to providing transfer functions for the plasmid, can also provide auxiliary functions such as resistance gene acquisition and biofilm formation to its host strains (Johnson et al., 2012; Wang et al., 2018). Biofilms can improve bacteria’s antibiotic resistance and raise their tolerance to harsh environmental conditions (Rabin et al., 2015). The acquired resistance to β-lactam antibiotics and heightened biofilm-forming potential of the wild-type strain after receiving the IncX3 plasmid may impair the drug’s efficacy (Ma et al., 2020). Transferring this plasmid to extremely virulent bacteria may improve pathogenicity and multidrug resistance, potentially increasing treatment failure rates (Ma et al., 2020). However, some studies have demonstrated that IncX3 plasmids do not promote bacterial pathogenicity (Liakopoulos et al., 2018; Liu et al., 2020). This could be related to the virulence and pathogenicity of the recipient strain itself, which requires further research.
The conjugation capacity of the IncX3 plasmid is inextricably linked to its high stability, and inhibiting conjugation transfer can lead to the elimination of the plasmid from the strain (Zhu et al., 2020). Plasmid stability can be improved further via IncX3 plasmid conjugation transfer. Linoleic acid is a conjugative transfer inhibitor. It can limit the frequency of plasmid conjugation and induce the loss of IncX3 plasmids in strains (Zhu et al., 2020). This conclusion reveals that conjugative transfer inhibitors can lessen the two key benefits of IncX3 plasmids, which is an excellent strategy to keep plasmids from spreading.
The majority of IncX3 plasmids are between 30 kb and 60 kb in size, with a few between 60 kb and 70 kb. The smallest plasmid in this collection, pKB11 (accession number: MK264769), is 12,757 kb in size (Fuga et al., 2020). Only sequences encoding replication, resistance (blaKPC-2 on NTEKPC-Ic), and auxiliary genes were found in the basic structure of pKB11. Although this novel tiny plasmid lacks conjugation genes, the host of pKB11 contained additional plasmids from various incompatibility groups, which may aid the transfer of this novel IncX3 plasmid to a new host. Following that was an unnamed3 plasmid (accession number: MN847777) with 18,178 bp but no conjugation genes (Cao et al., 2020). This is nearly identical to a non-conjugative 17-kb IncX3 plasmid harboring blaNDM. It eliminated large-scale regions compared to the typical IncX3 plasmid, suggesting a low fitness cost dissemination strategy for the IncX3 plasmid (Li et al., 2022). The 75,415-bp plasmid pWLK-NDM (accession number: CP038280) is the largest in this collection. In China, this plasmid was isolated from urban river debris. Despite having a conjugation gene, it is unable to conjugate due to the disruption of the pilX3-4 gene. This plasmid is also significantly larger than other IncX3 plasmids, which could be the result of genetic exchange between different bacterial species from various environments (Dang et al., 2020).
All IncX3 plasmids cited in this review carry ARGs, the majority of which are connected to carbapenemase genes. blaNDM had the highest prevalence, followed by blaOXA-181 and its variants, blaKPC, and blaSHV. Interestingly, the majority of gene types carried on plasmids in China are blaNDM and its variations. Except for blaNDM-4, all blaNDM variants located on IncX3 plasmids can be found in China. And no IncX3 plasmid carrying blaKPC was recorded in China. The frequency distribution of carbapenemase genes reported in different nations and species covered by this review are shown in Figures 2, 3.
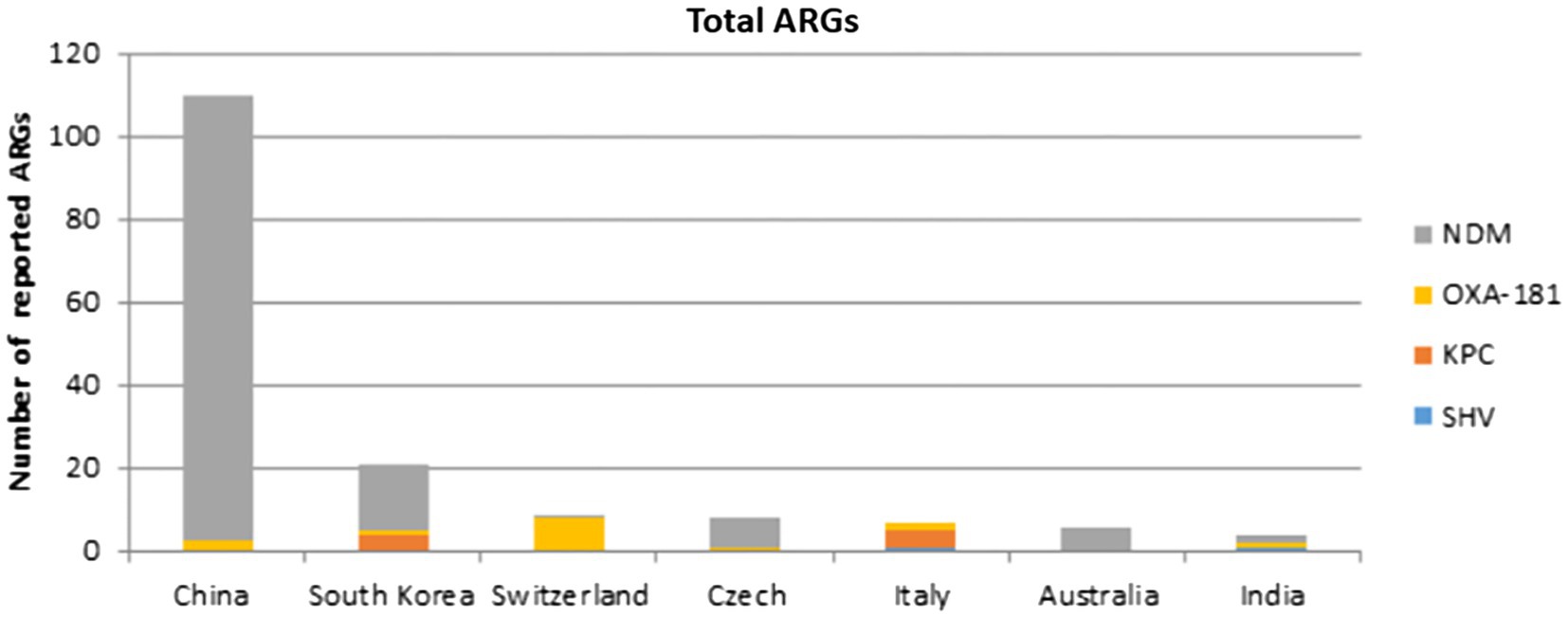
Figure 2. Number of carbapenemase genes reported in different countries represented by this review.
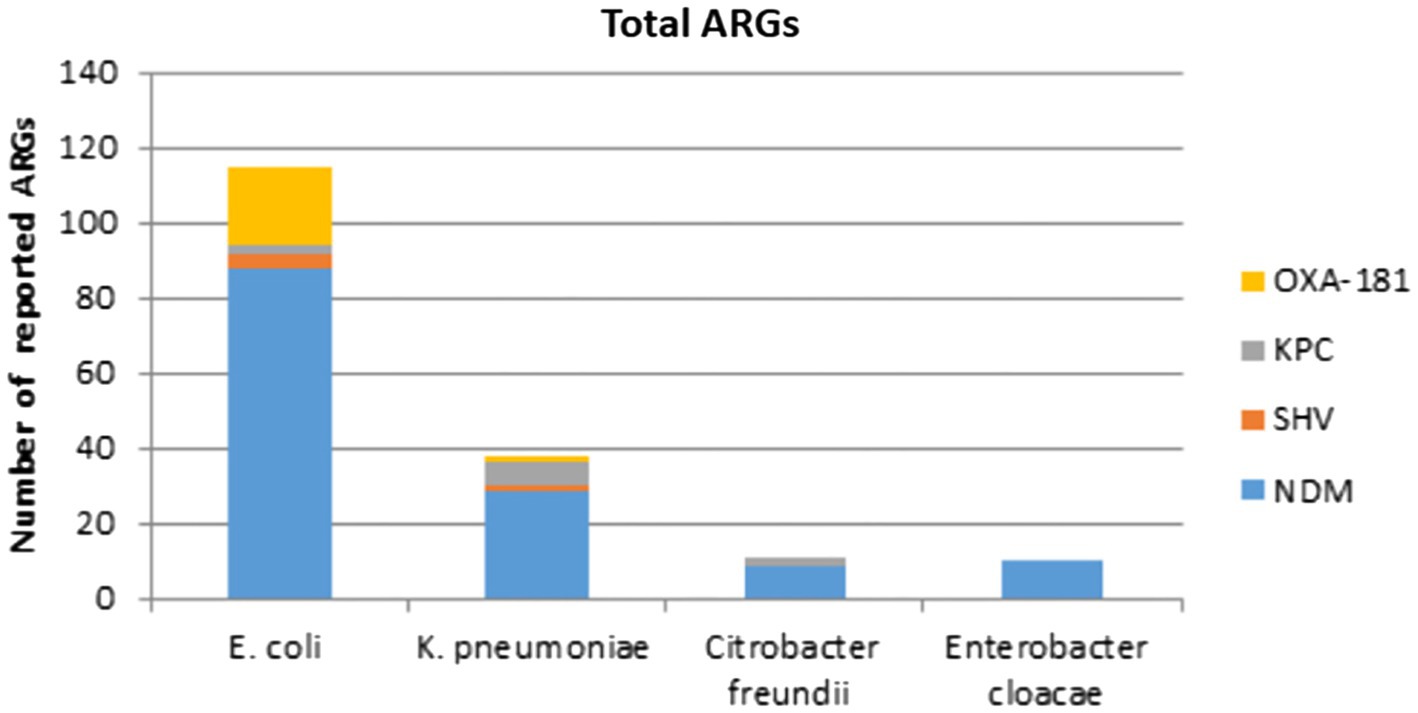
Figure 3. Number of carbapenemase genes reported in different species represented by this review.
The IncX3 plasmids carry various NDM variants, including NDM-5, NDM-1, NDM-7, NDM-4, NDM-11, NDM-13, NDM-17, NDM-19, NDM-20, NDM-21, and NDM-33 (Supplementary material). This suggests that the blaNDM-bearing IncX3 plasmids descended from a common ancestor plasmid through a series of mutations. The easy propagation of the IncX3 plasmid could be responsible for the spread of various NDM variants in Enterobacteriaceae isolates (Tian et al., 2020). NDM-positive strains can carry multiple types of plasmids, with IncX3 plasmids accounting for roughly one-third of the plasmids in the gene bank (Ma et al., 2020). The IncX3 plasmid is not only the most common vehicle for harboring blaNDM but may also be one of the most important platforms for blaNDM evolution as novel NDM variants emerge (Wu et al., 2019). Notably, blaNDM-5-carrying IncX3 plasmid was the most common type in blaNDM, followed by blaNDM-1-carrying IncX3 plasmid, which is consistent with the findings of Kopotsa et al. (2019). NDM-1 and NDM-5 were found to be more frequently connected with IncX3 plasmids than other NDM variations, according to the researchers. NDM-1 and NDM-5 differ in amino acid sequence at positions 88 (ValLeu) and 154 (MetLeu) and NDM-5 has a greater carbapenem hydrolysis ability (Sun et al., 2019).
Notably, we found conserved sequences in the genetic environment of most blaNDM and its variants. The majority of the strains have the conserved structure “blaNDM-bleMBL-trpF-dsbC,” but some simply have the “blaNDM-bleMBL-trpF” structure, or the dsbC was changed into the dcbD “blaNDM-bleMBL -trpF-dcbD,” and only two strains carrying blaNDM-5 had the conserved sequence” blaNDM-bleMBL -trpT-dcbD.” This conserved sequence may play a key role in stability and retention, increasing the transfer of ARGs and improving enzyme activity. However, the specific functions of related genes need to be investigated further (Brinkac et al., 2019).In addition, the environment surrounding blaNDM contains not only conserved sequences but also mobile elements such as ISAba125, IS3000, IS5, and IS26, indicating that blaNDM acts as an external gene to recombine into the IncX3 plasmid by insertion or transposition depending on IS elements (Liu et al., 2021b). Meanwhile, IS26 is linked to blaSHV, blaKPC to ISKpn6 and ISKpn7, blaNDM to ISAba125 (Sun et al., 2019), blaOXA-181 to IS26 and ISEcp1 (Supplementary material). This is consistent with Li et al. (2020). They claim that the IncX3 plasmid contains a changeable area that primarily encodes resistance to clinically significant antimicrobial agents and encodes genes required for its maintenance and dissemination. The findings confirm the potential of IncX3 plasmid for accumulating resistance genes via IS-mediated transposition, with the likely consequence of limiting effective treatment options for possible human infections. In blaNDM-carrying IncX3 plasmids, deletion events with a 500-bp deletion in ISAba125 and a 180-bp deletion in dsbC were observed, which is significant and powerful evidence that ISAba125 and dsbC were unstable, implying that blaNDM is evolving to reduce the burden to facilitate rapid and stable horizontal transfer (Liu et al., 2021b).
We also observed IncX3 plasmids to examine if they also carried other ARGs. Besides harboring blaNDM, blaKPC, and blaSHV, most IncX3 plasmids carry no additional antibiotic resistance genes, especially IncX3 blaNDM-5-carrying plasmids (García-Fernández et al., 2012; Chakraborty et al., 2021; Lu et al., 2022). The size of the plasmid likely limits the number of antibiotic resistance genes that the plasmid can carry. IncX3 plasmid carrying only blaNDM is similar to the original plasmid pNDM-MGR194 (accession number: KF220657) (Krishnaraju et al., 2015), which is an early prototype in the dissemination of blaNDM. This is the first report of blaNDM being inserted into a plasmid without the presence of other resistance gene determinants. Interestingly, all plasmids carrying blaOXA-181 also carry the qnrS1, while plasmids carrying blaNDM-1 often also carry blaSHV-12. This is consistent with the study by Xiang et al. (2021). IncX3 plasmids can disseminate blaOXA-181 and qnrS via an IS26-flanked composite transposon, or blaNDM and blaSHV-12 via a Tn125-like transposon. And these are the two main epidemic types of IncX3 plasmids.
The IncX3 plasmids with conjugation ability accounted for the majority (Kim et al., 2020; Ma et al., 2020; Zhu et al., 2020). The deletion or destruction of conjugation genes may be the reason for IncX3 plasmids losing conjugation ability (Pál et al., 2017; Di Pilato et al., 2020). However, some plasmids still failed to transfer in conjugation studies despite having entire backbone structures (Wang et al., 2017; Dang et al., 2020). The cause for this remains unknown. The IncX3 plasmids have a conjugation frequency of between 10−9 and 10−2 (Liu et al., 2017; Lu et al., 2022). The short size of the IncX3 plasmid may also play a part in the high transfer frequency, and such a high frequency and stable conjugation ability have played a key role in the rapid and widespread dissemination of IncX3 over the last decade.
The conjugation frequency of IncX3 plasmid varies in different strains, according to current research on the properties of IncX3 plasmid and gene function. Varying ST strains have different conjugation frequencies, even within the same strain. The IncX3 plasmid in the donor strain is inaccessible to Acinetobacter baumannii and Pseudomonas aeruginosa as receivers for conjugative transfer (Wang et al., 2018). The frequency of conjugation varies among strains from different sources. The conjugation frequency of animal-derived plasmids is lower at 37°, whereas the conjugation frequency of human-derived plasmids is higher (Liakopoulos et al., 2018). In Baomo’s study, four different temperatures were used to investigate the role of IncX3 plasmids in frequency, fitness cost, and stability at different temperatures: environment temperature in Asia and the Middle East (25 and 30°), human body temperature (37°), and chicken body temperature (42°; Sugawara et al., 2019). The data demonstrate that at 37°, the IncX3 plasmid has a higher transfer frequency, more stability, and a lower fitness cost, indicating that the plasmid has the highest transfer in human and other mammalian intestines and has been adapted to exist in the mammalian gut. This could explain why the IncX3 plasmid can be found in humans and other mammals, as well as in bacteria with different antibiotic resistance genes. The IncX3 plasmids had conjugation frequencies equivalent to or higher than the reference IncFII plasmid at 30 and 37° (Wang et al., 2018). IncX4 conjugal transfer to recipient E. coli at 30° is also higher than IncFII, demonstrating that this trait is common to all IncX plasmid subtypes (Wang et al., 2018).
Ho et al. hypothesized that the IncX3 plasmid backbone contains a histone-like nucleoid structuring (H-NS)–like protein that stabilizes plasmid DNA and regulates plasmid transfer at different temperatures (Ho et al., 2012). Furthermore, Sugawara et al. recently discovered that hns genes are ubiquitous in IncX3 plasmids, implying that H-NS-like proteins may influence the propagation of IncX3 plasmids (Sugawara et al., 2019). The role of the hns gene in plasmid dispersion, stability, fitness cost, and temperature regulation was fully clarified by Baomo’s study (Liu et al., 2020; Baomo et al., 2021). The author claims that the IncX3 plasmid, which lacks the plasmid-encoded hns gene, can increase conjugation frequency by upregulating conjugation-related genes and that the hns gene regulates conjugation transfer temperature-dependently. By boosting the expression of the parB gene, H-NS-like proteins can also improve plasmid stability in the host. H-NS-like protein also impacts the virulence of the plasmid by regulating the virulence gene encoded by the chromosome to create fimbriae for the adhesion and invasion of the strain.
Strains carrying IncX3 plasmids tend to carry more than one plasmid. Inc. typing is a method of classifying plasmids based on their ability to coexist stably with other plasmids in the same bacterial strain, determined by their replication machinery. When co-resident plasmids have the same replication mechanisms, they are incompatible (Johnson and Nolan, 2009). The number of plasmids carried by the strains ranged from 1 to 8 plasmids (Supplementary material), with 2, 3, and 4 being the most common. Multiple plasmids in the same strain are uncommon (e.g., 7–8), and the coexistence of a large number of plasmids reflects that active horizontal transfer events may have occurred before.
In most cases, a plasmid will only contain one type of replicon gene. Recent research has found that large MDR conjugative plasmids can harbor numerous replicon genes, expanding the host range of such plasmids. Furthermore, mobile elements are the main contributors to plasmid replicon fusion. Plasmid evolution by combining two or more MDR plasmids would increase the resistance profile of the resulting plasmid. Moreover, it would broaden the host spectrum of such resistance-encoding mobile elements, limiting antibiotic selection and application in clinical practice. Increasing the fitness cost could, in general, limit the spread of cointegrate plasmids. In high-risk strains, however, cointegrate plasmids remained stable in the presence of antibiotics, implying that drug residue is an essential driving force in the evolution of MDR pathogens (Liu et al., 2021c). Cointegrate plasmids have more ARGs and insertion sequences with high plasticity than regular plasmids because of the integration of genetic information from original plasmids. In response to a complicated environment, the cointegrate plasmid could dynamically discard some areas and acquire new ARGs, exhibiting its great dispersion and evolution potential (Liu et al., 2021c). The explosive proliferation of cointegrate plasmids in Klebsiella pneumoniae was observed by the HIN Hospital in the United States, which could be a means for plasmids to seek stable forms in evolution (Liu et al., 2021c). Transferring this plasmid to clinic-associated strains could improve biofilm formation capabilities, resulting in higher treatment failure rates (Hargreaves et al., 2015). Cointegrate plasmids should therefore be given special attention. A better understanding of the molecular mechanisms underlying the formation and evolution of cointegrate MDR plasmids will pave the way for preventing the spread of resistance elements among bacterial pathogens via IncX3 plasmid.
The IncX3 plasmid, as a typical self-transfer plasmid, can behave as a helper plasmid, assisting other non-conjugative plasmids in their transfer to other bacterial hosts and spreading antibiotic resistance genes further (Conlan et al., 2014). IncA/C, FII, FIA, FIB, IncR, IncY, and ColE plasmids are among the plasmids that cointegrate with IncX3 plasmids. The IS26 element plays a significant role in creating cointegrate plasmids, and its insertion promotes the dissemination of antibiotic resistance genes and the formation of cointegrate plasmids (Hao et al., 2021). Plasmid stability experiments on Salmonella containing IncX3 and IncA/C cointegrate plasmids revealed that cointegrate plasmids had high stability but were easily decomposed into single plasmids during conjugation and transfer, and the decomposed IncX3 plasmid could still transfer to the recipient strain (Li et al., 2020). K. aerogenes was found to have the IncC-IncX3 cointegrate plasmid pNUITM-VK5_mdr (accession number: LC633285), which was resistant to practically all antibiotics, including tigecycline, tetracyclines, carbapenems, cephalosporins, fluoroquinolones, and aminoglycosides (except amikacin) (Hirabayashi et al., 2021). The fusion is thought to have an increased risk of carrying more ARGs and spreading more stably and efficiently among bacteria in humans, animals, and the environment. Hence, IncX3 plasmids with high stability, low fitness cost, and self-transferability can assist broad-host-range plasmids that carry multiple antibiotic resistance genes but do not have the conjugation ability to achieve their transfer and dissemination. In return, broad-host-range plasmids may also promote the stabilization of co-resident IncX3 plasmid (Bitar et al., 2019).
Carbapenem resistance is a clinical problem to be solved urgently. The ability of antibiotic-resistant strains to produce carbapenemase is an important mechanism of carbapenem resistance, while plasmids play an important role in spreading and carrying antibiotic resistance genes. Among these, the IncX3 plasmid has low fitness costs, high stability, high transfer frequency, and improved strain biofilm-forming capabilities. The IncX3 plasmid is crucial for transporting and spreading carbapenemase genes (blaNDM, blaKPC, and blaOXA), particularly blaNDM. IncX3 plasmid is not only a vehicle for spreading blaNDM but also a platform for evolution. Many NDM variations, particularly blaNDM-5, can be found in IncX3. IncX3 plasmids have been isolated from different human, animal, and environmental strains worldwide, especially in China. The plasmids are primarily found in E. coli, K. pneumoniae, C. freundii, and Enterobacter cloacae. There are usually 1–3 co-resident plasmids in the IncX3 plasmid, with the transfer frequency ranging from 10−9 to 10−2 and the size of the plasmid usually ranging from 30 to 60 kb. It can be used as a co-resident plasmid to form a fusion with a non-self-transfer plasmid, and IncX3 plasmids could help expand the resistance profile and disseminate resistance genes. The structure of the IncX3 plasmid is divided into two parts: the backbone and the accessory module. The backbone is highly conserved, and its related genes play an important role in the stability and spread of the plasmid. More research into the physiological aspects is still needed. Based on this, future research should focus on the physiological role of the plasmid structure to provide a theoretical basis for controlling the global prevalence of IncX3.
XG contributed to the conceptualization. RC, CL, and HG searched articles and collected data. JQ and YL analyzed the data. RC and QW contributed to the writing and final approval. All authors contributed to the article and approved the submitted version.
This work was supported by grants from the Henan Province Medical Science and Technology Research Project Joint Construction Project (no. LHGJ20190232) and the Youth Innovation Fund project of the First Affiliated Hospital of Zhengzhou University (no. YNQN2017168).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2022.979558/full#supplementary-material
Andersson, D. I., and Hughes, D. (2010). Antibiotic resistance and its cost: is it possible to reverse resistance? Nat. Rev. Microbiol. 8, 260–271. doi: 10.1038/nrmicro2319
Baomo, L., Lili, S., Moran, R. A., van Schaik, W., and Chao, Z. (2021). Temperature-regulated Inc X3 plasmid characteristics and the role of plasmid-encoded H-NS in thermoregulation. Front. Microbiol. 12:765492. doi: 10.3389/fmicb.2021.765492
Bitar, I., Caltagirone, M., Villa, L., Mattioni Marchetti, V., Nucleo, E., Sarti, M., et al. (2019). Interplay among IncA and Bla (KPC)-carrying plasmids in Citrobacter freundii. Antimicrob. Agents Chemother. 63:02609-18. doi: 10.1128/aac.02609-18
Brinkac, L. M., White, R., D'Souza, R., Nguyen, K., Obaro, S. K., and Fouts, D. E. (2019). Emergence of New Delhi Metallo-β-lactamase (NDM-5) in Klebsiella quasipneumoniae from neonates in a Nigerian hospital. mSphere 4:e00685-18. doi: 10.1128/mSphere.00685-18
Cao, T., Liu, Y., Li, Y., Wang, Y., Shen, Z., Shao, B., et al. (2020). A public health concern: emergence of carbapenem-resistant Klebsiella pneumoniae in a public transportation environment. J. Antimicrob. Chemother. 75, 2769–2772. doi: 10.1093/jac/dkaa260
Chakraborty, T., Sadek, M., Yao, Y., Imirzalioglu, C., Stephan, R., Poirel, L., et al. (2021). Cross-border emergence of Escherichia coli producing the Carbapenemase NDM-5 in Switzerland and Germany. J. Clin. Microbiol. 59:e02238-20. doi: 10.1128/jcm.02238-20
Conlan, S., Thomas, P. J., Deming, C., Park, M., Lau, A. F., Dekker, J. P., et al. (2014). Single-molecule sequencing to track plasmid diversity of hospital-associated carbapenemase-producing Enterobacteriaceae. Sci. Transl. Med. 6, 254ra126. doi: 10.1126/scitranslmed.3009845
Dagher, C., Salloum, T., Alousi, S., Arabaghian, H., Araj, G. F., and Tokajian, S. (2018). Molecular characterization of Carbapenem resistant Escherichia coli recovered from a tertiary hospital in Lebanon. PLoS One 13:e0203323. doi: 10.1371/journal.pone.0203323
Dang, B., Zhang, H., Li, Z., Ma, S., and Xu, Z. (2020). Coexistence of the Bla(NDM-1)-carrying plasmid pWLK-NDM and the Bla(KPC-2)-carrying plasmid pWLK-KPC in a Raoultella ornithinolytica isolate. Sci. Rep. 10, 2360. doi: 10.1038/s41598-020-59341-4
Di Pilato, V., Ceccherini, F., Sennati, S., D'Agostino, F., Arena, F., D'Atanasio, N., et al. (2020). In vitro time-kill kinetics of dalbavancin against staphylococcus spp. biofilms over prolonged exposure times. Diagn. Microbiol. Infect. Dis. 96:114901. doi: 10.1016/j.diagmicrobio.2019.114901
Dziri, R., Kuşkucu, M. A., Arfaoui, A., Fethi, M., Ifaoui, S., Bellaaj, R., et al. (2022). Whole genome sequencing of a Citrobacter freundii strain isolated from the hospital environment: an extremely multiresistant NDM-1 and VIM-48 coproducing isolate. Microb. Drug Resist. 28, 18–22. doi: 10.1089/mdr.2020.0417
Fang, H., Feng, J., Xu, Y., Zhang, Y., Zhan, Z., Yin, Z., et al. (2018). Sequencing of pT5282-CTXM, p13190-KPC and p30860-NR, and comparative genomics analysis of IncX8 plasmids. Int. J. Antimicrob. Agents 52, 210–217. doi: 10.1016/j.ijantimicag.2018.04.012
Flerlage, T., Brazelton de Cardenas, J. N., Garner, C. D., Hasan, N. A., Karathia, H., Qudeimat, A., et al. (2020). Multiple NDM-5-expressing Escherichia coli isolates from an immunocompromised pediatric host. Open Forum Infect. Dis. 7, ofaa018. doi: 10.1093/ofid/ofaa018
Fuga, B., Ferreira, M. L., Cerdeira, L. T., de Campos, P. A., Dias, V. L., Rossi, I., et al. (2020). Novel small IncX3 plasmid carrying the Bla(KPC-2) gene in high-risk Klebsiella pneumoniae ST11/CG258. Diagn. Microbiol. Infect. Dis. 96:114900. doi: 10.1016/j.diagmicrobio.2019.114900
García-Fernández, A., Villa, L., Carta, C., Venditti, C., Giordano, A., Venditti, M., et al. (2012). Klebsiella pneumoniae ST258 producing KPC-3 identified in Italy carries novel plasmids and OmpK36/OmpK35 porin variants. Antimicrob. Agents Chemother. 56, 2143–2145. doi: 10.1128/aac.05308-11
Grönthal, T., Österblad, M., Eklund, M., Jalava, J., Nykäsenoja, S., Pekkanen, K., et al. (2018). Sharing more than friendship–transmission of NDM-5 ST167 and CTX-M-9 ST69 Escherichia coli between dogs and humans in a family, Finland, 2015. Euro Surveill. 23:1700497. doi: 10.2807/1560-7917.Es.2018.23.27.1700497
Gu, C., Li, X., Zou, H., Zhao, L., Meng, C., Yang, C., et al. (2022). Clonal and plasmid-mediated dissemination of environmental carbapenem-resistant Enterobacteriaceae in large animal breeding areas in northern China. Environ. Pollut. 297:118800. doi: 10.1016/j.envpol.2022.118800
Hao, M., Schuyler, J., Zhang, H., Shashkina, E., Du, H., Fouts, D. E., et al. (2021). Apramycin resistance in epidemic carbapenem-resistant Klebsiella pneumoniae ST258 strains. J. Antimicrob. Chemother. 76, 2017–2023. doi: 10.1093/jac/dkab131
Hargreaves, M. L., Shaw, K. M., Dobbins, G., Snippes Vagnone, P. M., Harper, J. E., Boxrud, D., et al. (2015). Clonal dissemination of Enterobacter cloacae harboring blaKPC-3 in the upper Midwestern United States. Antimicrob. Agents Chemother. 59, 7723–7734. doi: 10.1128/aac.01291-15
Hirabayashi, A., Dao, T. D., Takemura, T., Hasebe, F., Trang, L. T., Thanh, N. H., et al. (2021). A transferable IncC-IncX3 hybrid plasmid Cocarrying Bla(NDM-4), tet(X), and tmexCD3-toprJ3 confers resistance to Carbapenem and Tigecycline. mSphere 6:e0059221. doi: 10.1128/mSphere.00592-21
Ho, P. L., Li, Z., Lo, W. U., Cheung, Y. Y., Lin, C. H., Sham, P. C., et al. (2012). Identification and characterization of a novel incompatibility group X3 plasmid carrying Bla NDM-1 in Enterobacteriaceae isolates with epidemiological links to multiple geographical areas in China. Emerg. Microbes. Infect. 1:e39. doi: 10.1038/emi.2012.37
Huang, C., Liu, L. Z., Kong, H. K., Law, C. O. K., Hoa, P. Q., Ho, P. L., et al. (2020). A novel incompatibility group X3 plasmid carrying Bla (NDM-1) encodes a small RNA that regulates host fucose metabolism and biofilm formation. RNA Biol. 17, 1767–1776. doi: 10.1080/15476286.2020.1780040
Johnson, T. J., Bielak, E. M., Fortini, D., Hansen, L. H., Hasman, H., Debroy, C., et al. (2012). Expansion of the IncX plasmid family for improved identification and typing of novel plasmids in drug-resistant Enterobacteriaceae. Plasmid 68, 43–50. doi: 10.1016/j.plasmid.2012.03.001
Johnson, T. J., and Nolan, L. K. (2009). Plasmid replicon typing. Methods Mol. Biol. 551, 27–35. doi: 10.1007/978-1-60327-999-4_3
Juraschek, K., Käsbohrer, A., Malorny, B., Schwarz, S., Meemken, D., and Hammerl, J. A. (2021). Dissection of highly prevalent qnrS1-carrying IncX plasmid types in commensal Escherichia coli from German food and livestock. Antibiotics (Basel) 10:1236. doi: 10.3390/antibiotics10101236
Kim, J. S., Jin, Y. H., Park, S. H., Han, S., Kim, H. S., Yu, J. K., et al. (2020). Horizontal transfer of Bla(NDM-1)-carrying IncX3 plasmid between carbapenem-resistant Enterobacteriaceae in a single patient. J. Infect. 81, 816–846. doi: 10.1016/j.jinf.2020.07.013
Kopotsa, K., Osei Sekyere, J., and Mbelle, N. M. (2019). Plasmid evolution in carbapenemase-producing Enterobacteriaceae: A review. Ann. N. Y. Acad. Sci. 1457, 61–91. doi: 10.1111/nyas.14223
Krishnaraju, M., Kamatchi, C., Jha, A. K., Devasena, N., Vennila, R., Sumathi, G., et al. (2015). Complete sequencing of an IncX3 plasmid carrying blaNDM-5 allele reveals an early stage in the dissemination of the blaNDM gene. Indian J. Med. Microbiol. 33, 30–38. doi: 10.4103/0255-0857.148373
Legese, M. H., Asrat, D., Mihret, A., Hasan, B., Mekasha, A., Aseffa, A., et al. (2022). Genomic epidemiology of Carbapenemase-producing and Colistin-resistant Enterobacteriaceae among sepsis patients in Ethiopia: A whole-genome analysis. Antimicrob. Agents Chemother. 66:e0053422. doi: 10.1128/aac.00534-22
Li, X., Fu, Y., Shen, M., Huang, D., Du, X., Hu, Q., et al. (2018). Dissemination of Bla(NDM-5) gene via an IncX3-type plasmid among non-clonal Escherichia coli in China. Antimicrob. Resist. Infect. Control 7, 59. doi: 10.1186/s13756-018-0349-6
Li, R., Liu, Z., Li, Y., Xiao, X., and Wang, Z. (2022). Characterization of Bla(NDM)-positive Enterobacteriaceae reveals the clonal dissemination of Enterobacter hormaechei coharboring Bla(NDM) and tet(X4) along the pork production chain. Int. J. Food Microbiol. 372:109692. doi: 10.1016/j.ijfoodmicro.2022.109692
Li, R., Xie, M., Liu, L., Huang, Y., Wu, X., Wang, Z., et al. (2020). Characterisation of a cointegrate plasmid harbouring Bla(NDM-1) in a clinical salmonella Lomita strain. Int. J. Antimicrob. Agents 55:105817. doi: 10.1016/j.ijantimicag.2019.09.021
Liakopoulos, A., van der Goot, J., Bossers, A., Betts, J., Brouwer, M. S. M., Kant, A., et al. (2018). Genomic and functional characterisation of IncX3 plasmids encoding Bla(SHV-12) in Escherichia coli from human and animal origin. Sci. Rep. 8, 7674. doi: 10.1038/s41598-018-26073-5
Liu, B., Guo, Y., Liu, N., Wang, J., Li, F., Yao, L., et al. (2021a). In silico evolution and comparative genomic analysis of IncX3 plasmids isolated From China Over ten years. Front. Microbiol. 12:725391. doi: 10.3389/fmicb.2021.725391
Liu, B., Shui, L., Zhou, K., Jiang, Y., Li, X., Guan, J., et al. (2020). Impact of plasmid-encoded H-NS-like protein on blaNDM-1-bearing IncX3 plasmid in Escherichia coli. J. Infect. Dis. 221, S229–s236. doi: 10.1093/infdis/jiz567
Liu, Z., Wang, Z., Lu, X., Peng, K., Chen, S., He, S., et al. (2021c). Structural diversity, fitness cost, and stability of a Bla(NDM-1)-bearing Cointegrate plasmid in Klebsiella pneumoniae and Escherichia coli. Microorganisms 9:2435. doi: 10.3390/microorganisms9122435
Liu, Z., Wang, Y., Walsh, T. R., Liu, D., Shen, Z., Zhang, R., et al. (2017). Plasmid-mediated novel Bla(NDM-17) gene encoding a Carbapenemase with enhanced activity in a sequence type 48 Escherichia coli strain. Antimicrob. Agents Chemother. 61:e02233-16. doi: 10.1128/aac.02233-16
Liu, Z., Wang, K., Zhang, Y., Xia, L., Zhao, L., Guo, C., et al. (2021b). High prevalence and diversity characteristics of Bla (NDM), mcr, and Bla (ESBLs) harboring multidrug-resistant Escherichia coli from chicken, pig, and cattle in China. Front. Cell. Infect. Microbiol. 11:755545. doi: 10.3389/fcimb.2021.755545
Liu, Z., Xiao, X., Li, Y., Liu, Y., Li, R., and Wang, Z. (2019). Emergence of IncX3 plasmid-harboring Bla (NDM-) (5) dominated by Escherichia coli ST48 in a goose farm in Jiangsu. China Front. Microbiol. 10, 2002. doi: 10.3389/fmicb.2019.02002
Lu, X., Du, Y., Peng, K., Zhang, W., Li, J., Wang, Z., et al. (2022). Coexistence of tet(X4), mcr-1, and Bla(NDM-5) in ST6775 Escherichia coli isolates of animal origin in China. Microbiol Spectr 10:e0019622. doi: 10.1128/spectrum.00196-22
Ma, T., Fu, J., Xie, N., Ma, S., Lei, L., Zhai, W., et al. (2020). Fitness cost of Bla(NDM-5)-carrying p3R-IncX3 plasmids in wild-type NDM-free Enterobacteriaceae. Microorganisms 8:377. doi: 10.3390/microorganisms8030377
Mouftah, S. F., Pál, T., Darwish, D., Ghazawi, A., Villa, L., Carattoli, A., et al. (2019). Epidemic IncX3 plasmids spreading carbapenemase genes in the United Arab Emirates and worldwide. Infect. Drug Resist. 12, 1729–1742. doi: 10.2147/idr.S210554
Moussounda, M., Diene, S. M., Dos Santos, S., Goudeau, A., François, P., and van der Mee-Marquet, N. (2017). Emergence of Bla(NDM-7)-producing Enterobacteriaceae in Gabon, 2016. Emerg. Infect. Dis. 23, 356–358. doi: 10.3201/eid2302.161182
Pál, T., Ghazawi, A., Darwish, D., Villa, L., Carattoli, A., Hashmey, R., et al. (2017). Characterization of NDM-7 Carbapenemase-producing Escherichia coli isolates in the Arabian peninsula. Microb. Drug Resist. 23, 871–878. doi: 10.1089/mdr.2016.0216
Peirano, G., Chen, L., Nobrega, D., Finn, T. J., Kreiswirth, B. N., DeVinney, R., et al. (2022). Genomic epidemiology of global carbapenemase-producing Escherichia coli, 2015-2017. Emerg. Infect. Dis. 28, 924–931. doi: 10.3201/eid2805.212535
Petrosillo, N., Vranić-Ladavac, M., Feudi, C., Villa, L., Fortini, D., Barišić, N., et al. (2016). Spread of Enterobacter cloacae carrying blaNDM-1, blaCTX-M-15, blaSHV-12 and plasmid-mediated quinolone resistance genes in a surgical intensive care unit in Croatia. J. Glob. Antimicrob. Resist. 4, 44–48. doi: 10.1016/j.jgar.2015.09.008
Poirel, L., Dortet, L., Bernabeu, S., and Nordmann, P. (2011). Genetic features of blaNDM-1-positive Enterobacteriaceae. Antimicrob. Agents Chemother. 55, 5403–5407. doi: 10.1128/aac.00585-11
Rabin, N., Zheng, Y., Opoku-Temeng, C., Du, Y., Bonsu, E., and Sintim, H. O. (2015). Biofilm formation mechanisms and targets for developing antibiofilm agents. Future Med. Chem. 7, 493–512. doi: 10.4155/fmc.15.6
Shi, L., Liang, Q., Zhan, Z., Feng, J., Zhao, Y., Chen, Y., et al. (2018). Co-occurrence of 3 different resistance plasmids in a multi-drug resistant Cronobacter sakazakii isolate causing neonatal infections. Virulence 9, 110–120. doi: 10.1080/21505594.2017.1356537
Sugawara, Y., Akeda, Y., Hagiya, H., Sakamoto, N., Takeuchi, D., Shanmugakani, R. K., et al. (2019). Spreading patterns of NDM-producing Enterobacteriaceae in clinical and environmental settings in Yangon, Myanmar. Antimicrob. Agents Chemother. 63:e01924-18. doi: 10.1128/aac.01924-18
Sun, P., Xia, W., Liu, G., Huang, X., Tang, C., Liu, C., et al. (2019). Characterization Of Bla (NDM-5)-positive Escherichia coli prevalent in a university hospital in eastern China. Infect. Drug Resist. 12, 3029–3038. doi: 10.2147/idr.S225546
Tian, D., Wang, B., Zhang, H., Pan, F., Wang, C., Shi, Y., et al. (2020). Dissemination of the Bla (NDM-5) gene via IncX3-type plasmid among Enterobacteriaceae in children. mSphere 5:e00699-19. doi: 10.1128/mSphere.00699-19
Touati, A., Manseur, L., Mehidi, I., and Mairi, A. (2021). Epidemiological and genetic features of plasmids carrying Bla(NDM) genes: an in silico analysis with emphasis on replicon types, and resistome. Microb. Drug Resist. 27, 1232–1242. doi: 10.1089/mdr.2020.0427
Wang, Y., Tong, M. K., Chow, K. H., Cheng, V. C., Tse, C. W., Wu, A. K., et al. (2018). Occurrence of highly conjugative IncX3 epidemic plasmid carrying Bla (NDM) in Enterobacteriaceae isolates in geographically widespread areas. Front. Microbiol. 9:2272. doi: 10.3389/fmicb.2018.02272
Wang, J., Yuan, M., Chen, H., Chen, X., Jia, Y., Zhu, X., et al. (2017). First report of Klebsiella oxytoca strain simultaneously producing NDM-1, IMP-4, and KPC-2 Carbapenemases. Antimicrob. Agents Chemother. 61:e00877-17. doi: 10.1128/aac.00877-17
Wu, W., Feng, Y., Tang, G., Qiao, F., McNally, A., and Zong, Z. (2019). NDM Metallo-β-lactamases and their bacterial producers in health care settings. Clin. Microbiol. Rev. 32:e00115-18. doi: 10.1128/cmr.00115-18
Xiang, G., Lan, K., Cai, Y., Liao, K., Zhao, M., Tao, J., et al. (2021). Clinical molecular and genomic epidemiology of Morganella morganii in China. Front. Microbiol. 12:744291. doi: 10.3389/fmicb.2021.744291
Yang, L., Lin, Y., Lu, L., Xue, M., Ma, H., Guo, X., et al. (2020). Coexistence of two Bla (NDM-) (5) genes carried on IncX3 and IncFII plasmids in an Escherichia coli isolate revealed by illumina and nanopore sequencing. Front. Microbiol. 11, 195. doi: 10.3389/fmicb.2020.00195
Zhang, Q., Lv, L., Huang, X., Huang, Y., Zhuang, Z., Lu, J., et al. (2019). Rapid increase in Carbapenemase-producing Enterobacteriaceae in retail meat driven by the spread of the Bla (NDM-5)-carrying IncX3 plasmid in China from 2016 to 2018. Antimicrob. Agents Chemother. 63:e00573-19. doi: 10.1128/aac.00573-19
Zhang, Z., Zhang, L., Dai, H., Zhang, H., Song, Y., An, Q., et al. (2022). Multidrug-resistant Klebsiella pneumoniae complex from clinical dogs and cats in China: molecular characteristics, phylogroups, and hypervirulence-associated determinants. Front Vet Sci 9:816415. doi: 10.3389/fvets.2022.816415
Zhao, W., Li, S., Schwarz, S., Li, A., Yao, H., and Du, X. D. (2021). Detection of a NDM-5-producing Klebsiella pneumoniae sequence type 340 (CG258) high-risk clone in swine. Vet. Microbiol. 262:109218. doi: 10.1016/j.vetmic.2021.109218
Keywords: IncX3, plasmid, epidemiology, carbapenemases, drug resistance
Citation: Guo X, Chen R, Wang Q, Li C, Ge H, Qiao J and Li Y (2022) Global prevalence, characteristics, and future prospects of IncX3 plasmids: A review. Front. Microbiol. 13:979558. doi: 10.3389/fmicb.2022.979558
Edited by:
John Osei Sekyere, University of Pretoria, South AfricaReviewed by:
Jayalaxmi Wangkheimayum, Assam University, IndiaCopyright © 2022 Guo, Chen, Wang, Li, Ge, Qiao and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiaobing Guo, Z3hiaW5nOTI4QHp6dS5lZHUuY24=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.