
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 21 September 2022
Sec. Evolutionary and Genomic Microbiology
Volume 13 - 2022 | https://doi.org/10.3389/fmicb.2022.942603
This article is part of the Research Topic Basidiomycete Fungi: From Biosystematics and Biodiversity to Biotechnology View all 21 articles
 Yuan Yuan1†
Yuan Yuan1† Ying-Da Wu1,2†Ya-Rong Wang1
Ying-Da Wu1,2†Ya-Rong Wang1 Meng Zhou1
Meng Zhou1 Jun-Zhi Qiu3De-Wei Li4
Jun-Zhi Qiu3De-Wei Li4 Josef Vlasák5
Josef Vlasák5 Hong-Gao Liu6Yu-Cheng Dai1*
Hong-Gao Liu6Yu-Cheng Dai1*Phaeolus schweinitzii (Fr.) Pat. was originally described in Europe and is considered a common forest pathogen on conifers in the Northern Hemisphere. Our molecular phylogeny based on samples from China, Europe, and North America confirms that P. schweinitzii is a species complex, including six taxa. P. schweinitzii sensu stricto has a distribution in Eurasia; the samples from Northeast and Southwest China are distantly related to P. schweinitzii sensu stricto, and two new species are described after morphological, phylogenetic, and geographical analyses. The species growing on Larix, Picea, and Pinus in Northeast China is described as Phaeolus asiae-orientalis. Another species mostly occurring on Pinus yunnanensis in Southwest China is Phaeolus yunnanensis. In addition, three taxa distributed in North America differ from P. schweinitzii sensu stricto. Phaeolus tabulaeformis (Berk.) Pat. is in Southeast North America, “P. schweinitzii-1” in Northeast North America, and “P. schweinitzii-2” in western North America.
Phaeolus (Pat.) Pat. is a well-known polypore genus because of its type species; Phaeolus schweinitzii (Fr.) Pat. has conspicuous and colorful basidiocarps and is the major cause of butt rotting in many commercial timber species (Gilbertson and Ryvarden, 1987; Sinclair et al., 1987; Núñez and Ryvarden, 2001; Ryvarden and Melo, 2017). The genus is characterized by annual, pileate to stipitate, orange to brown basidiocarps with a fibrous to spongy context, a monomitic hyphal system with simple septate hyphae, the presence of gloeoplerous hyphae and cystidia, ellipsoid to cylindric, hyaline, thin-walled, acyanophilous, neither amyloid nor dextrinoid basidiospores, and causing brown rot. According to current taxonomy, Phaeolus is closely related to Wolfiporia Ryvarden & Gilb. and belongs to the family Laetiporaceae in the order Polyporales (Justo et al., 2017; Wu et al., 2020).
Although many epithets are listed in Phaeolus, most were treated as synonyms of P. schweinitzii (Donk, 1974; Index Fungorum: http://www.indexfungorum.org/names/Names.asp, MycoBank: https://www.mycobank.org/page/Simple%20names%20search). Previously, five species only, viz., Phaeolus amazonicus M.A. De Jesus & Ryvarden, P. subbulbipes (Henn.) O. Fidalgo & M. Fidalgo, P. manihotis R. Heim, P. tabulaeformis (Berk.) Pat., and P. schweinitzii (Fr.) Pat., were accepted in the genus. The former three species were described from Brazil and Madagascar, respectively, with a limited distribution (Heim, 1931; Fidalgo and Fidalgo, 1957; Jesus and Ryvarden, 2010), and P. tabulaeformis was described from Georgia, USA (Patouillard, 1900). P. schweinitzii was described from Europe and reported to be widely distributed in The Northern Hemisphere (Donk, 1974; Gilbertson and Ryvarden, 1987; Núñez and Ryvarden, 2001; Ryvarden and Melo, 2017). P. schweinitzii was also reported in Australia and New Zealand (Buchanan and Ryvarden, 2000; Simpson and May, 2002).
During investigations on brown-rot fungi in China, specimens morphologically similar to P. schweinitzii were collected from Northeast (NE) and Southwest (SW) China, but they are different from the European P. schweinitzii based on our preliminary phylogenetic study. Thus, it appears that P. schweinitzii is a species complex with more independent species that exist in the Northern Hemisphere. Then, more samples from North America and Europe were added for the multigene phylogeny, and six independent lineages were formed. After morphological examinations and phylogenetic and geographical analyses, six taxa are recognized in the P. schweinitzii complex; two taxa are confirmed as new members of Phaeolus, and they are pathogens on coniferous trees in NE and WS China. The present study aims to clarify the Chinese species of Phaeolus and outline the phylogeny of Phaeolus based on available data in the Northern Hemisphere.
The examined specimens are deposited at the herbaria of the Institute of Microbiology, Beijing Forestry University (BJFC), and the Institute of Applied Ecology, Chinese Academy of Sciences (IFP). Macro-morphological descriptions were based on field notes and measurements of voucher herbarium specimens. Microscopic measurements and drawings were obtained from the slides prepared from voucher specimens and stained with Cotton Blue and Melzer's reagent following Wu et al. (2022) using a Nikon Eclipse 80i microscope. The following abbreviations were used in the description: CB = Cotton Blue, CB– = acyanophilous, IKI = Melzer's reagent, IKI– = neither amyloid nor dextrinoid, L = mean spore length (arithmetic average of spores), W = mean spore width (arithmetic average of spores), Q = variation in the L/W ratios between specimens studied, n (a/b) = number of spores (a) measured from a given number of specimens (b). In presenting spore size variation, mean ± SD (standard deviation at 95% confidence) was reported as the range; 5% of measurements were excluded from each end of the range, and the values are given in parentheses. Special color terms follow Anonymous (1969) and Petersen (1996). Herbarium abbreviations follow Thiers (2022).
The Rapid Plant Genome kit based on acetyl trimethylammonium bromide extraction (Aidlab Biotechnologies Co., Ltd, Beijing, China) was used to extract total genomic DNA from dried specimens and for polymerase chain reaction (PCR), according to the manufacturer's instructions with some modifications (Song and Cui, 2017; Xing et al., 2018). The ITS region was amplified with the primer pairs ITS5 and ITS4 (White et al., 1990). The nLSU region was amplified with primer pairs LR0R and LR7 (http://www.biology.duke.edu/fungi/mycolab/primers.htm). The PCR procedure was followed by Yuan et al. (2021). The PCR products were purified and sequenced at the Beijing Genomics Institute, China, with the same primers as in the original PCR amplifications.
The phylogenetic tree was constructed using sequences obtained in this study and additional sequences downloaded from GenBank (Table 1). The sequences were aligned within MAFFT version 7 (Katoh et al., 2019) and ClustalX (Thompson et al., 1997), then manual proofreading was performed in BioEdit (Hall, 1999). The downloaded sequences were chosen to cover Laetiporaceae Jülich and related clades, including Fomitopsidaceae Jülich and Sparassidaceae Herter (Justo et al., 2017; Song and Cui, 2017; Song et al., 2018). Ambiguous regions were deleted, and gaps were manually adjusted to optimize alignment before phylogenetic analyses. Sparassis latifolia Y. C. Dai and Zheng Wang was used as an outgroup in the phylogeny of Phaeolus (Zhao et al., 2013; Figure 1). The data matrix was edited in Mesquite version 3.04 software. Phylogenetic analyses were performed with maximum parsimony (MP), maximum likelihood (ML), and Bayesian Inference (BI) based on ITS + nLSU aligned datasets.
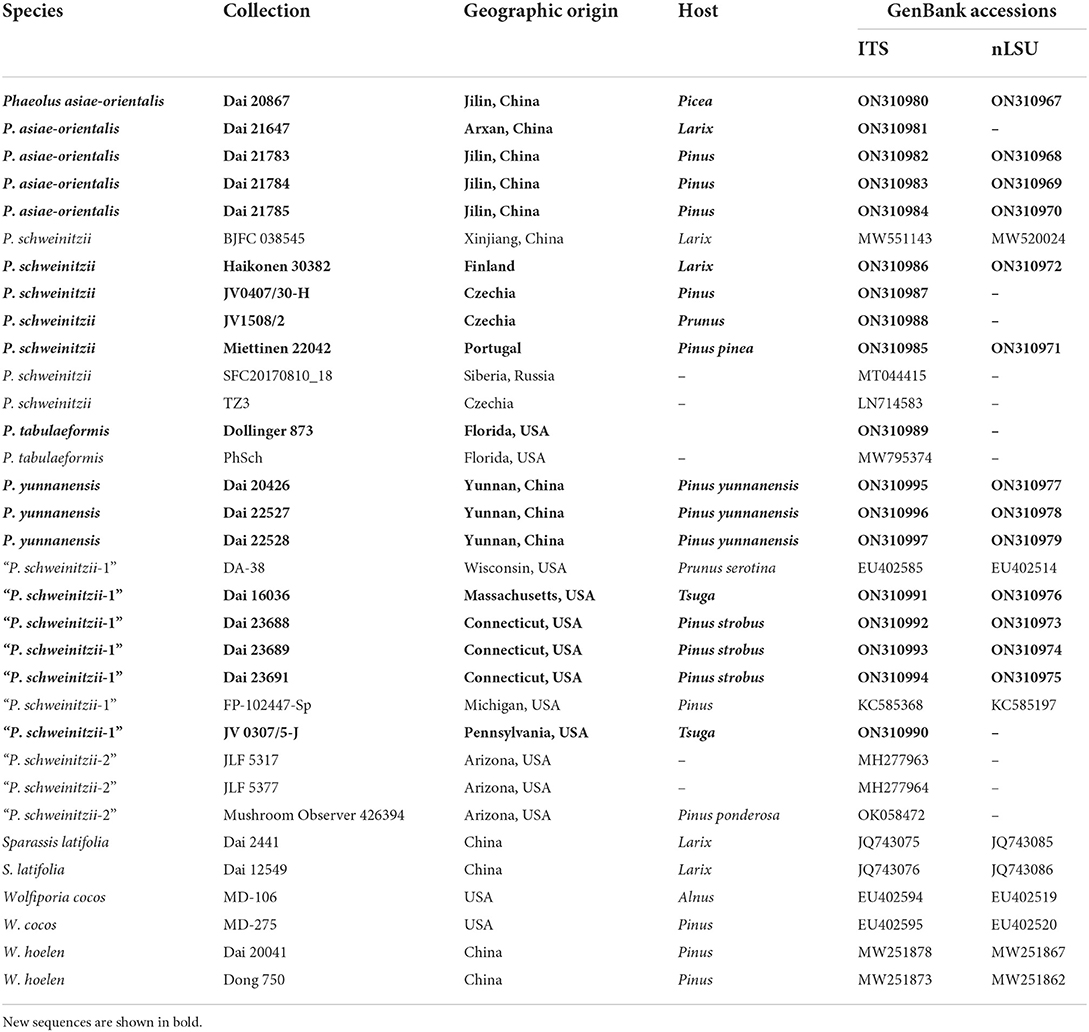
Table 1. Taxa information and GenBank accession numbers of the sequences used in this study.
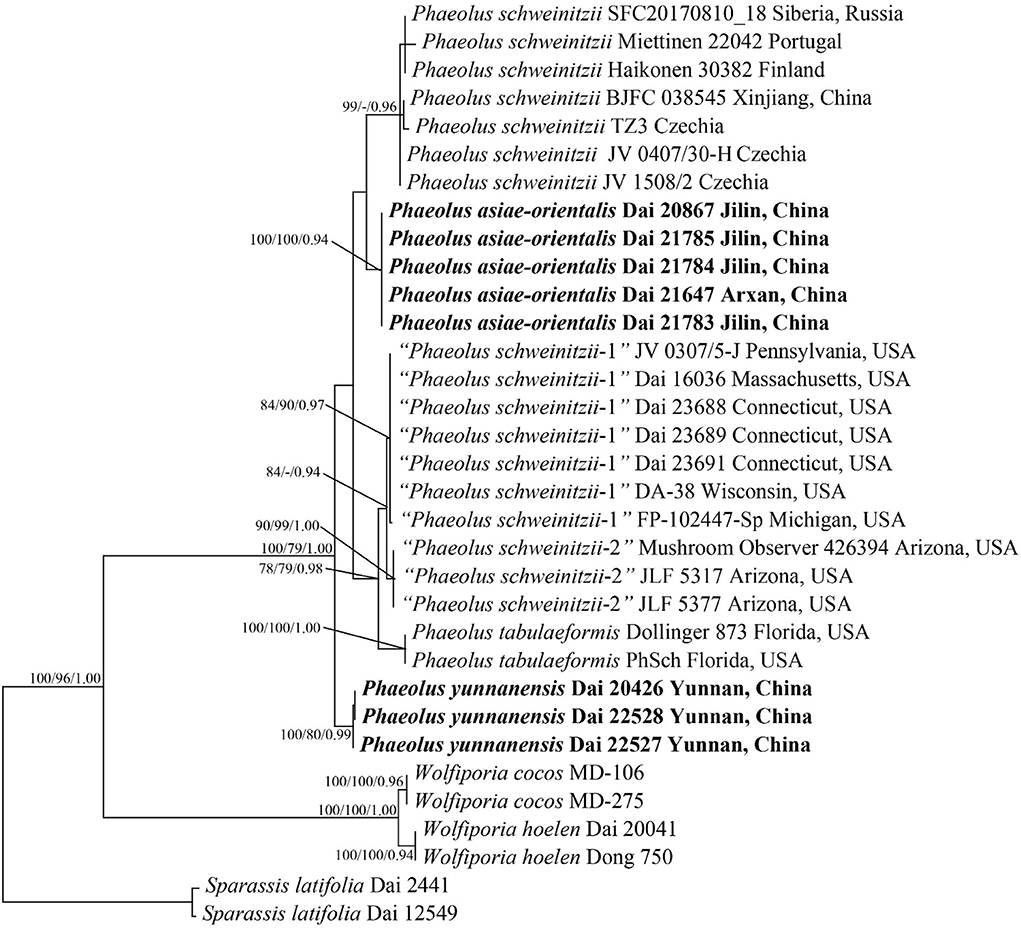
Figure 1. Phylogeny of Phaeolus generated by maximum parsimony based on a dataset of ITS + nLSU. Branches are labeled with bootstrap values (MP/ML) higher than 75% and posterior probabilities (BI) more than 0.90, respectively. New taxa are in bold.
MP analysis was applied to the dataset containing the ITS + nLSU sequences. The tree construction procedure was performed using PAUP* version 4.0b10 (Swofford, 2002). All characters were equally weighted, and gaps were treated as missing data. Trees were inferred using the heuristic search option with TBR branch swapping and 1,000 random sequence additions. Max-trees were set to 5,000, branches of zero length were collapsed, and all parsimonious trees were saved. Clade robustness was assessed using a bootstrap analysis with 1,000 replicates (Felsenstein, 1985). Descriptive tree statistics: tree length (TL), consistency index (CI), retention index (RI), rescaled consistency index (RCI), and homoplasy index (HI) were calculated for each maximum parsimonious tree generated.
jModeltest v.2.17 (Darriba et al., 2012) was used to determine the best-fit evolution model of the combined dataset for ML and BI. Four unique partitions were established; GTR + I + G was the selected substitution model for each partition. RAxML version 8.2.12 (Stamatakis, 2014) was used for ML analysis with default parameters. Only the best maximum likelihood tree from all searches was kept.
The BI was calculated with MrBayes version 3.2.6 (Ronquist et al., 2012) in two independent runs, each of which had four chains for 10 million generations and started from random trees. Trees were sampled every 100 generations. The first 25% of sampled trees were discarded as burn-in, whereas other trees were used to construct a 50% majority consensus tree and for calculating Bayesian posterior probabilities (BPPs).
Phylogenetic trees were visualized using TreeView (Page, 1996). Branches that received bootstrap support for Maximum likelihood (BS), Maximum parsimony (BP), and Bayesian posterior probabilities (BPP) ≥75% (BS and BP) and 0.90 (BPP) were considered as significantly supported, respectively.
The ITS + nLSU sequences from 33 fungal collections represent nine species. Among them, 31 new sequences were generated in this study. The dataset had an aligned length of 1,944 characters, of which 1,473 characters are constant, six are variable and parsimony-uninformative, and 465 are parsimony-informative. MP analysis yielded a tree (TL = 626, CI = 0.925, RI = 0.972, RC = 0.899, HI = 0.075). The best model for the ITS + nLSU sequences dataset estimated and applied in the Bayesian analysis was GTR + I + G with an equal frequency of nucleotides, lset nst = 6 rates = invgamma; prset statefreqpr = dirichlet (1,1,1,1). The Bayesian analysis resulted in a similar topology to MP, with an average standard deviation of split frequencies = 0.004684; thus, only the MP tree is displayed (Figure 1). Two new species, Phaeolus asiae-orientalis (100% MP, 100% ML, 0.94 BI) and P. yunnanensis (100% MP, 80% ML, 1.00 BI), formed well-supported phylogenetic lineages, respectively.
According to the present phylogenetic analyses, the P. schweinitzii complex consists of six taxa: P. schweinitzii sensu stricto seems to be distributed in Eurasia; P. asiae-orientalis and P. yunnanensis are so far known in Northeast and Southwest China; P. tabulaeformis (Berk.) Pat., “P. schweinitzii-1,” and “P. schweinitzii-2” have distributions in North America.
Phaeolus asiae-orientalis Y.C. Dai & Yuan Yuan, sp. nov. Figures 2, 3.

Figure 2. Basidiocarps of Phaeolus asiae-orientalis. (a,b) Dai 20867 (holotype); (c) Dai 789; (d) Dai 7089; (e) Dai 21784; (f) Dai 14566. Scale bars = 1 cm.
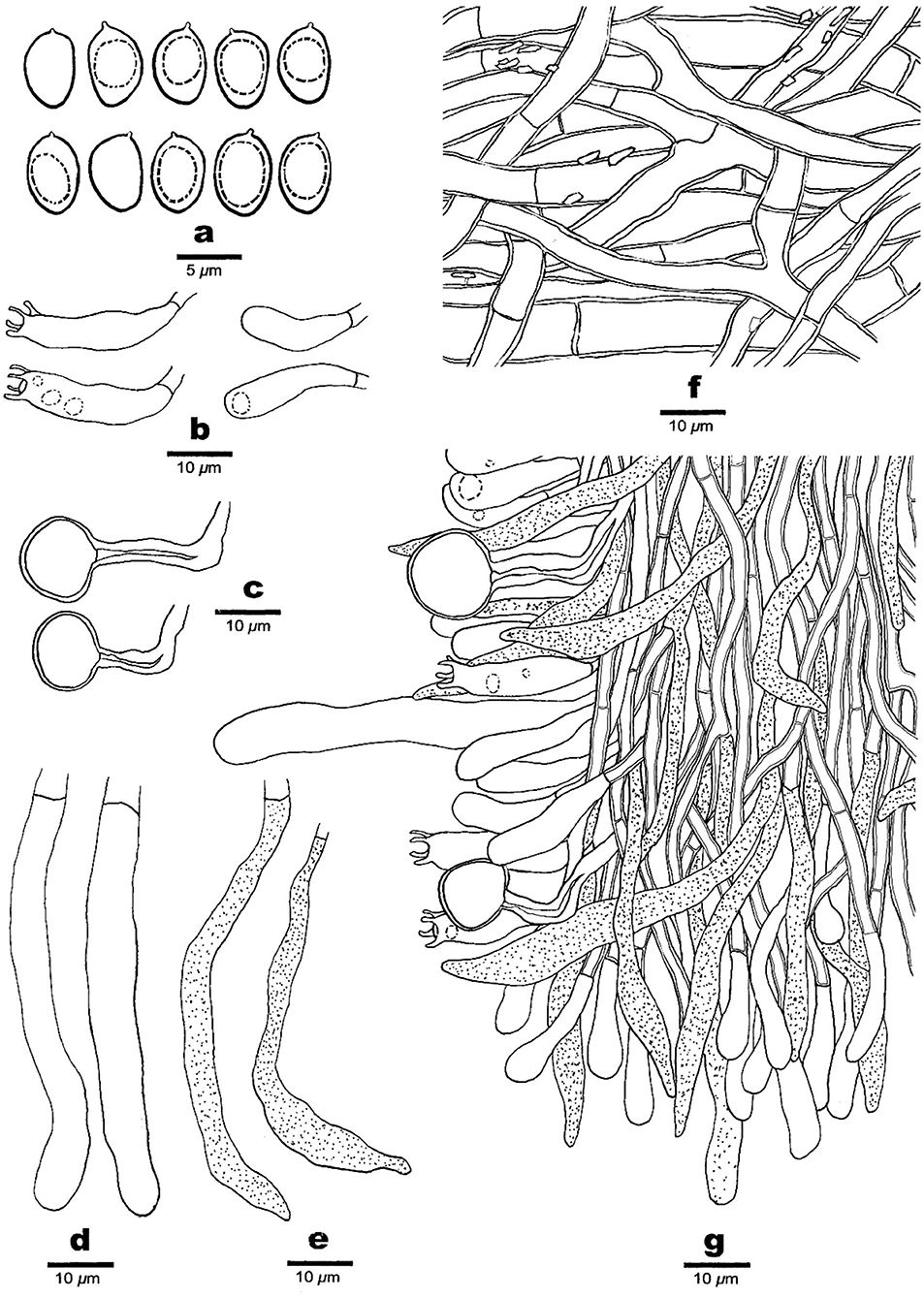
Figure 3. Microscopic structures of Phaeolus asiae-orientalis (drawn from the holotype, Dai 20867). (a) Basidiospores. (b) Basidia and basidioles. (c) Vascular elements. (d) Cystidia. (e) Gloeocystidia. (f) Hyphae from context. (g) A section from tube trama.
MycoBank number: MB 845327.
Diagnosis: Phaeolus asiae-orientalis is characterized by pileate to laterally or centrally stipitate basidiocarps, irregular pores 1–2 per mm, abundant vascular elements, mango-shaped basidiospores 6–7 × 3.5–4.2 μm, L = 6.34 μm, W = 3.82 μm, Q = 1.48–1.63 and distribution in Northeast China.
Type: China, Jilin Province, Antu County, the Changbaishan Nature Reserve, dead tree of Picea, 21 Sep. 2019, Y.C. Dai, Dai 20867 (holotype, BJFC032536!).
Etymology: Asiae-orientalis (Lat.): referring to East Asia, where the species was found.
Basidiocarps: Basidiocarps annual, pileate to laterally or centrally stipitate on substrate or ground from roots, soft and watery when fresh, and fragile and light in weight when dry. Pilei imbricate, sometimes developing a large number of imbricate, petaloid, or flabelliform and often confluent pilei, circular or semicircular to fan-shaped with irregularly lobed margin, projecting up to 15, 26 cm wide, up to 2 cm thick at the base or center; pileal surface cream, buff, orange when juvenile, pinkish buff, reddish brown to fuscous with age, concentrically zonate with various shades of cinnamon, brown, reddish brown to fuscous colors when fresh, become tomentose to hirsute, umber, rusty tawny to dark brown, indistinctly concentrically zonate when dry; margin acute, incurved when dry. Pore surface greenish yellow, citrine to sulfur yellow, become dark brown when bruised, rusty brown, bay, date brown to purplish chestnut when dry, sterile margin distinct, up to 3 mm wide; pores irregular and labyrinthine, 1–2 per mm; dissepiments thick, entire to lacerate. Context concolorous with a pileal surface, soft corky to fibrous, azonate, up to 10 mm thick. Tubes are pinkish buff to curry yellow, paler than the context, fragile when dry, decurrent, and up to 10 mm long. Stipe central or lateral, sometimes branched, concolorous with a pileal surface, and up to 2 cm long and 1 cm in diameter.
Hyphal structure: Hyphal system monomitic; generative hyphae simple septate, thin- to slightly thick-walled, IKI–, CB–; tissue darkening otherwise unchanged in KOH.
Context: Generative hyphae are hyaline to yellowish, thin- to slightly thick-walled with a wide lumen, occasionally branched, loosely interwoven, encrusted by crystals, and 4.2–15 μm in diameter; gloeoplerous hyphae are present, thin-walled, dark blue in CB.
Tubes: Generative hyphae are hyaline to yellowish, thin- to thick-walled, occasionally branched, subparallel among the tubes, some encrusted by crystals, 3–4 μm in diameter, gloeoplerous hyphae present, thin-walled, and dark blue in CB. Cystidia present, subulate and ventricose or clavate, hyaline, thin-walled, smooth, 65–80 × 10–14 μm, gloeocystidia present, long fusoid, hyaline, thin-walled, dark blue in CB, and 22.5–32.5 × 6.5–8.2 μm. Basidia clavate, bearing four sterigmata and a simple basal septum, and 24–30 × 7.5–9 μm; basidioles dominant, similar to basidia in shape, but smaller. Vascular elements are frequently present in the hymenium.
Spores: Basidiospores are mostly mango-shaped, some ellipsoid, hyaline, thin-walled, smooth, mostly mono-guttulate, IKI–, CB–, (5.5–)6–7(−7.1) × (3.2–)3.5–4.2(−4.5) μm, L = 6.34 μm, W = 3.82 μm, Q = 1.48–1.63 (n = 90/3).
Additional materials (paratypes) examined: China, Heilongjiang Province, Tangyuan County, The Daliangzihe National Park, on Pinus, August 25, 2014, B.K. Cui, and Cui 11481 (BJFC016723); Wuma County, The Nanwenghe Nature Reserve, on root of Larix, August 27, 2014, Y.C. Dai, and Dai 14566 (BJFC017811); Yichun, The Fenglin Nature Reserve, on fallen gymnosperm trunk, 2.VIII.2011, Cui 9894 (BJFC010787). Inner Mongolia, Arxan, Arxan Forest Park, on the living tree of Larix gmelinii, August 24, 2020, Y.C. Dai, Dai 21647 (BJFC035548). Jilin Prov., Antu County, The Changbaishan Nature Reserve, on the fallen trunk of Picea, August 28, 2005, Y.C. Dai, and Dai 7089 (BJFC001617); on rotten Pinus, July 28, 1993, Y.C. Dai, Dai 789 (IFP004023), August 18, 2020, Y.C. Dai, Dai 21783 (BJFC035684), Dai 21784 (BJFC035685), and Dai 21785 (BJFC035686).
Phaeolus yunnanensis Y.C. Dai & Yuan Yuan, sp. nov. Figures 4, 5.

Figure 4. Basidiocarps of Phaeolus yunnanensis. (a,b) Dai 20426 (holotype); (c,d) Dai 22528. Scale bars = 1 cm.
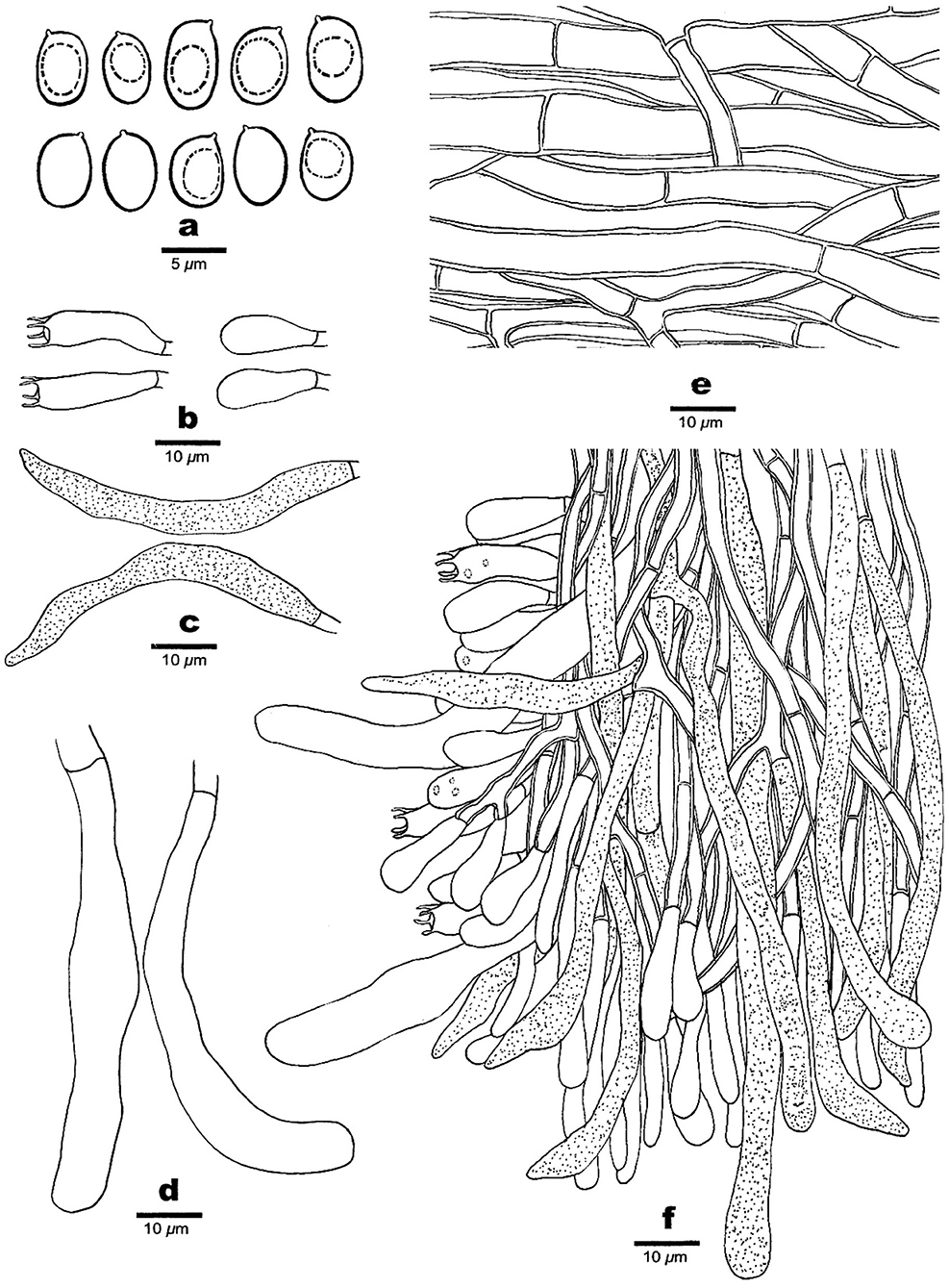
Figure 5. Microscopic structures of Phaeolus yunnanensis (drawn from the holotype, Dai 20426). (a) Basidiospores. (b) Basidia and basidioles. (c) Gloeocystidia. (d) Cystidia. (e) Hyphae from context. (f) A section from tube trama.
MycoBank number: MB 845328.
Diagnosis: Phaeolus yunnanensis is characterized by pileate to laterally stipitate basidiocarps, irregular pores of 0.5–1 per mm, absence of vascular elements, ellipsoid to oblong ellipsoid basidiospores 5.5–6.2 × 3.6–4 μm, L = 5.85 μm, W = 3.83 μm, Q = 1.45–1.65 and distribution in Southwest China.
Type: China, Yunnan Province, Yuxi, Xinping County, Longquan Park, the root of Pinus yunnanensis, 16 Aug. 2019, Y.C. Dai, Dai 20426 (holotype, BJFC032094!).
Etymology: Yunnanensis (Lat.) refers to the species found on Pinus yunnanensis in Yunnan province.
Basidiocarps: Basidiocarps annual, pileate to laterally stipitate, always on wood, soft when fresh, corky to fragile, and light in weight when dry. Pilei applanate, semicircular to fan-shaped with lobed margin, projecting up to 9, 14 cm wide, and 13 mm thick at the base or center. Upper surface buff, fawn to vinaceous brown, concentrically zonate with various shades of buff, cinnamon, vinaceous brown to bay colors when fresh, tomentose to hirsute, indistinctly concentrically zonate when dry; margin acute or blunt, curved inwards when dry. Pore surface cream, greenish yellow to sulfur yellow, slightly glistening, become dark brown when bruised, cinnamon buff, cinnamon, rusty brown, bay to purplish chestnut when dry; sterile margin indistinct to almost lacking; pores irregular and labyrinthine, 0.5–1 per mm; dissepiments thin, entire in juvenile to lacerate to dentate with age. Context concolorous with a pileal surface, azonate, and corky when dry, up to 10 mm thick. Tubes are buff to orange-yellow, paler than the context, fragile when dry, decurrent, and up to 3 mm long. Stipe central or lateral, unbranched, concolorous with a pileal surface, up to 1 cm long and 1 cm in diameter.
Hyphal structure: Hyphal system monomitic; generative hyphae simple septate, thin- to slightly thick-walled, IKI–, CB–; tissue darkening otherwise unchanged in KOH.
Context: Generative hyphae are hyaline to yellowish, thin- to slightly thick-walled with a wide lumen, occasionally branched, loosely interwoven, and 4.5–12 μm in diameter; gloeoplerous hyphae present, thin-walled, and dark blue in CB.
Tubes: Generative hyphae are hyaline to yellowish, thin- to slightly thick-walled, occasionally branched, subparallel among the tubes, 3–4.2 μm in diameter; gloeoplerous hyphae are present, thin-walled, and dark blue in CB. Cystidia present, subulate and ventricose or clavate, hyaline, thin-walled, smooth, and 76–90 × 10–20 μm; gloeocystidia occasionally present, fusoid, thin-walled, dark blue in CB, and 24–31 × 6–7 μm. Basidia clavate, bearing four sterigmata and a simple basal septum, and 25–30 × 6.5–7.5 μm; basidioles dominant, similar to basidia in shape, but smaller. Vascular elements are absent.
Spores: Basidiospores ellipsoid to oblong ellipsoid, hyaline, thin-walled, smooth, mostly mono-guttulate, IKI–, CB–, (5.2–) 5.5–6.2 (−6.5) × (3.3–) 3.6–4 (−4.2) μm, L = 5.85 μm, W = 3.83 μm, Q = 1.45–1.65 (n = 90/3).
Additional materials (paratypes) examined: China, Yunnan Province, Chuxiong, The Zixishan National Forest Park, on the rotten root of Pinus yunnanensis, 02 July 2021; Y.C. Dai, Dai 22527 (BJFC037106), Dai 22528 (BJFC037107). Nanhua County, The Dazhongshan Nature Reserve, on the root of Pinus yunnanensis, 15 July 2013, Y.C. Dai, Dai 13279 (BJFC014768). Yuxi, Xinping County, Longquan Park, on the stump of Pinus yunnanensis, July 5, 2021; Y.C. Dai, Dai 22529 (BJFC037108).
Other materials studied.—P. schweinitzii. China. Xinjiang Autonomous Region, Altay, Burqin County, on the stump of Larix, August 19, 2019, J.Z. Qiu, M542 (BJFC038545). Belarus. Brestskaya Voblasts, Belavezhskaya Pushcha National Park, on Picea, 19 Oct. 2019, Y.C. Dai, Dai 21061 (BJFC032720). The Czech Republic. Ceské Budějovice, Hluboka, on Pinus, Jul. 2004, J. Vlasák, JV0407/30-H (dupl. in BJFC038546). Finland. Helsinki Botanical Garden, on the root of Larix, July 5. 1997, Y.C. Dai, Dai 2267 (IFP004025); Etela-Hame, Heinola, Kirkonkyla, Papilla, on Larix, 11 Apr. 2013, Haikonen 29485 (H); Tammmela, Mustiala, Maatalousoppilaitos, on Larix, April 17, 2016, Haikonen 30632 (H). Portugal. Algarve, Vila do Bispo, Budens, on Pinus pinea, 26 Jul. 2018, Miettinen (dupl. in BJFC033041).—“P. schweinitzii-1.” The USA. Connecticut, Griswold, Hopeville Pond State Park, on Pinus strobus, 07 Jun. 2021, D.W. Li, dupl. Y.C. Dai, Dai 23689 (BJFC038260), D.W. Li, dupli. Y.C. Dai, Dai 23690 (BJFC038262), D.W. Li, dupli. Dai 23691 (BJFC038263); New Haven, on the ground, October 29, 2021, D.W. Li, dupl. Dai 23688 (BJFC038260). Massachusetts, Boston, Forestry Hill, on Tsuga, July 27, 2015, Y.C. Dai, Dai 16036 (BJFC020137). Pennsylvania, Wilkes-Barre, Rickettes Glen St. Park, on Tsuga roots, July 2003, J. Vlasák Jr., JV0307/5-J (dupl. BJFC033026).—Phaeolus tabulaeformis. The USA. Florida, Sarasota, Water Tower Park, Frisbee golf course, August 13, 2016, Dollinger 873 (dupli. JV and PRM).
Six taxa are detected in the P. schweinitzii complex: P. schweinitzii sensu stricto seems to be distributed in Eurasia; P. asiae-orientalis and P. yunnanensis are so far known in Northeast and Southwest China; P. tabulaeformis (Berk.) Pat., “P. schweinitzii-1” and “P. schweinitzii-2” have distributions in North America. P. schweinitzii sensu stricto is most probably not distributed in North America, and its previous records in North America (Gilbertson and Ryvarden, 1987) should be analyzed by molecular data.
Twelve names were listed as synonyms of P. schweinitzii (Donk, 1974; Index Fungorum Database 2021, http://www.speciesfungorum.org/GSD/GSDspecies.asp?RecordID=121352; MycoBank Database 2021, https://www.mycobank.org/page/Simple%20names%20search); most of these taxa were described from Europe (Donk, 1974), and four were outside of Europe, viz. Phaeolus amazonicus, P. subbulbipes, P. manihotis, and P. tabulaeformis.
Inonotus sulphureopulverulentus P. Karst. was described from Baikal, Russia (Karsten, 1904). Pilát (1936–1942) considered it a species of uncertain identity; Lowe (1956) treated it as a synonym of P. schweinitzii. The senior author studied the type (in H), and it is a sterile fragment of basidiocarp. To confirm its identity, new samples from the type locality are needed. However, one sample, SFC2017081018 from Siberia, is P. schweinitzii, and our sample BJFC 038545 from the Altay area is also P. schweinitzii (Figure 1); the Russian Baikal is closer to Siberia and Altay, and the type of Inonotus sulphureopulverulentus is most probably a representation of P. schweinitzii.
Polyporus tabulaeformis Berk. (1845, type from Augusta, Georgia), Polyporus spectabilis Fr. (1851, type from South Carolina, illegitimate) and Polyporus hispidoides Peck (1880, type from New York) were described from North America; Overholts (1953) and Donk (1974) treated them as synonyms of P. schweinitzii. The former taxon was combined as Phaeolus tabulaeformis (Berk.) Pat. by Patouillard (1900). Ryvarden (1977) considered P. tabulaeformis to be a synonym of P. schweinitzii. However, our phylogenetic analyses show that three taxa of Phaeolus exist in North America (Figure 1). Samples from Florida (Dollinger 873 and PhSch) fit the description of Polyporus tabulaeformis well (Berkeley, 1845), and they are treated as Phaeolus tabulaeformis. Samples from Northeast North America (JV0307/5-J, Dai 16036, Dai 23688, Dai 23689, Dai 23691, DA-38, and FP-102447) most probably represent Polyporus hispidoides (type from New York). We treated them as “P. schweinitzii-1” because we did not study the type of P. hispidoides. Samples from western North America (JLF 5317, JLF 5377, and Mushroom Observer 426394) are treated as “P. schweinitzii-2.” We studied two collections of JV0108/104 (from California) and JV0308/64 (from Washington) but failed to extract their DNA, and they were not analyzed in our phylogeny. Samples of JLF 5317, JLF 5377, and Mushroom Observer 426394 from Arizona formed an independent lineage in our phylogeny (Figure 1); they may represent an undescribed taxon. Because our present paper focuses on the Chinese taxa of Phaeolus, we are not going to comment much on American taxa.
Phaeolus schweinitzii sensu stricto is easily distinguished from the two new species by its bigger basidiospores [6–9 × 4.5–5 μm in Ryvarden and Melo (2017) vs. 6–7 × 3.5–4.2 μm in P. asiae-orientalis and 5.5–6.2 × 3.6–4 μm in P. yunnanensis].
Phaeolus tabulaeformis has pores of 2–3 per mm and basidiospores of 6–7 × 4–5 μm (from Dollinger 873), so its pores are smaller than those in P. asiae-orientalis and P. yunnanensis, and its basidiospores are wider than those in P. asiae-orientalis and P. yunnanensis.
“Phaeolus schweinitzii-1” resembles P. asiae-orientalis and P. yunnanensis, but it has mostly oblong ellipsoid basidiospores and scanty vascular elements, while P. asiae-orientalis has mostly mango-shaped basidiospores and abundant vascular elements. “P. schweinitzii-1” differs from P. yunnanensis by longer basidiospores [(6.5–) 6.6–7.5 (−7.7) × (3.5–) 3.7–4.1 (−4.3) μm, L = 6.98 μm, W = 3.90 μm, Q = 1.79 (n = 90/3) vs. (5.2–) 5.5–6.2 (−6.5) × (3.3–) 3.6–4 (−4.2) μm, L = 5.85 μm, W = 3.83 μm, Q = 1.45–1.65 (n = 90/3)].
Phaeolus asiae-orientalis is different from P. yunnanensis by longer basidiospores (6–7 × 3.5–4.2 μm vs. 5.5–6.2 × 3.6–4 μm). In addition, vascular elements are abundant in P. asiae-orientalis, while they are absent in P. yunnanensis.
Phaeolus asiae-orientalis and P. yunnanensis are common species in both natural and planted coniferous forests in China; they were previously considered P. schweinitzii, a forest pathogen in the Chinese forests (Dai et al., 2007; Dai, 2012). The present results demonstrate that the three pathogenetic species of Phaeolus exist in China: viz. P. asiae-orientalis mostly on Larix gmelinii, L. olgensis, Picea jezoensis, and Pinus koraiensis in Northeast China, Phaeolus yunnanensis usually on Pinus yunnanensis in Southwest China, and P. schweinitzii on Larix in Northwest China. P. asiae-orientalis is the cause of butt rot in natural coniferous forests in Northeast China, while P. yunnanensis causes butt rot in planted coniferous forests in Southwest China.
Phaeolus schweinitzii was also reported in New Zealand and Australia on species of Araucariaceae and Myrtaceae (Cunningham, 1965; Buchanan and Ryvarden, 2000) and in the tropical pacific areas of Hawaii Island on Acacia (the Bega, 1979). The taxon differs from P. schweinitzii sensu stricto and will be published by another team (personal communication).
Phaeolus amazonicus M.A. De Jesus & Ryvarden was described from Brazil, and it differs from the members of the genus in the Northern Hemisphere by its dimitic hyphal structure (Jesus and Ryvarden, 2010). Phaeolus manihotis (Heim, 1931) was described from Madagascar. It is different from our two new species by the presence of cuticle or crust at the upper surface and the absence of cystidia (Heim, 1931). Phaeolus subbulbipes (Henn.) O. Fidalgo & M. Fidalgo was also originally described from Brazil, and it differs from our new species by its smaller and globose basidiospores measuring 3.5–4.1 μm (Fidalgo and Fidalgo, 1957).
Phaeolus rigidus (Lév.) Pat. was described in Java, Indonesia, but according to Ryvarden (1981), it is a synonym of Trichaptum durum (Jungh.) Corner.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/supplementary material.
YY and Y-CD: design of the research. YY and Y-DW: performance of the research. YY, Y-DW, Y-RW, and MZ: data analysis and interpretation. Y-CD, JV, J-ZQ, D-WL, and H-GL: collection of the materials. YY, Y-DW, Y-CD, JV, and D-WL: writing and revising the manuscript. All authors contributed to the article and approved the submitted version.
The research was supported by the National Natural Science Foundation of China (Project Nos. 32161143013, U1802231) and the Second Tibetan Plateau Scientific Expedition and Research Program (STEP, Grant No. 2019QZKK0503).
The authors are grateful to Dr. Otto Miettinen (Helsinki) for arranging specimens on loan for our study. Y-CD is grateful to Dr. Tuomo Niemelä for his friendly support in studying specimens in the Botanical Museum of the University of Helsinki, Finland.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Anonymous (1969). Flora of British Fungi. Colour Identification Chart. London: Her Majesty's Stationery Office.
Bega, R. V. (1979). Heart and root tot fungi associated with deterioration of Acacia koa on the Island of Hawaii. Plant Dis. Rep. 63, 682–684.
Berkeley, M. J. (1845). Decades of fungi. Decades VIII-X. Australian and North American fungi. Lond. J. Bot. 4, 298–315.
Buchanan, P. K., and Ryvarden, L. (2000). An annotated checklist of polypore and polypore-like fungi recorded from New Zealand. N. Zeal. J. Bot. 38, 265–323. doi: 10.1080/0028825X.2000.9512683
Cunningham, G. H. (1965). Polyporaceae of New Zealand. N. Zeal. Depart. Sci. Ind. Res. Bull. 64, 1–304.
Dai, Y. C., Cui, B. K., Yuan, H. S., and Li, B. D. (2007). Pathogenic wood-decaying fungi in China. For. Pathol. 37, 105–120. doi: 10.1111/j.1439-0329.2007.00485.x
Darriba, D., Taboada, G. L., Doallo, R., and Posada, D. (2012). jModelTest 2: more models, new heuristics and parallel computing. Nat. Methods 9, 772. doi: 10.1038/nmeth.2109
Donk, M. A. (1974). Check list of European polypores. Verh Koninkl Nederl Akad Wet Natuurk 62, 1–469.
Felsenstein, J. (1985). Confidence intervals on phylogenetics: an approach using bootstrap. Evolution 39, 783–791. doi: 10.1111/j.1558-5646.1985.tb00420.x
Fidalgo, O., and Fidalgo, M. E. P. K. (1957). Revisão de Fungi São Paulensis. Arquivos do Museu Nacional Rio de Janeiro 43, 157–188.
Gilbertson, R. L., and Ryvarden, L. (1987). North American polypores 2. Megasporoporia - Wrightoporia. Oslo: Fungiflora, pp. 434–885.
Hall, T. A. (1999). Bioedit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 41, 95–98. doi: 10.1021/bk-1999-0734.ch008
Heim, R. (1931). Les Phaeolus manihotis sp. nov., parasite du manioc a Madagascar et considération sur le genre Phaeolus. Pat. Ann. Crypt. Exotiq. 4, 175–189.
Jesus, M. A. D., and Ryvarden, L. (2010). Studies in neotropical polypores 28. Two new species from Amazonas, Brazil. Synop. Fung. 27, 73–77.
Justo, A., Miettinen, O., Floudas, D., Ortiz-Santana, B., Sjökvist, E., Lindner, D. L., et al. (2017). A revised family-level classification of the Polyporales (Basidiomycota). Fungal Biol. 121, 798–824. doi: 10.1016/j.funbio.2017.05.010
Karsten, P. A. (1904). Fungi novi, paucis exceptis, in Sibiria a clarissimo O.A.F. Loennbohm collecti. Öfversigt af Finska Vetenskaps-Societetens Förhandlingar 46, 1–9.
Katoh, K., Rozewicki, J., and Yamada, K. D. (2019). MAFFT online service: multiple sequence alignment, interactive sequence choice, and visualization. Brief. Bioinform. 20, 1160–1166. doi: 10.1093/bib/bbx108
Lowe, J. L. (1956). Type studies of the polypores described by Karsten. Mycologia 48, 99–125. doi: 10.1080/00275514.1956.12024520
Núñez, M., and Ryvarden, L. (2001). East Asian polypores 2. Polyporaceae s. lato. Synop. Fung. 14, 170–522.
Overholts, L. D. (1953). The Polyporaceae of the United States, Alaska, and Canada. Ann Arbor: University of Michigan Press.
Page, R. D. M. (1996). TreeView: application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 12, 357–358. doi: 10.1093/bioinformatics/12.4.357
Patouillard, N. T. (1900). Essai taxonomique sur les familles et les genres des Hym?nomycètes. Lucien Declume: Lons-le-Saunier. p. 1–184.
Petersen, J. H. (1996). Farvekort. Foreningen Til Svampekundskabens Fremme. Greve: Foreningen til Svampekundskabens Fremme.
Pilát, A. (1936–1942). “Polyporaceae,” in: Atlas des champignons de l' Europe, eds C. Kavina, and A. Pilát (Praha: Privately published), pp. 374.
Ronquist, F., Teslenko, M., Mark, P. V. D., Ayres, D., Darling, A., Höhna, S., et al. (2012). MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542. doi: 10.1093/sysbio/sys029
Ryvarden, L. (1977). Type studies in the Polyporaceae 10. Species described by J.M. Berkeley, either alone or with other authors, from 1844 to 1855. Norwegian J. Bot. 24, 213–230.
Ryvarden, L. (1981). Type studies in the Polyporaceae 13. Species described by J.H. Léveillé. Mycotaxon 13, 175–186.
Simpson, J. A., and May, T. W. (2002). Phaeolus schweinitzii in Australia. Australas. Plant Path. 31, 99–100. doi: 10.1071/AP01076
Sinclair, W. A., Lyon, H. H., and Johnson, W. T. (1987). Diseases of trees and shrubs. Ithaca: Cornell University Press, pp. 1–574.
Song, J., and Cui, B. K. (2017). Phylogeny, divergence time, and historical biogeography of Laetiporus (Basidiomycota, Polyporales). BMC Evol. Biol. 17, 102. doi: 10.1186/s12862-017-0948-5
Song, J., Sun, Y. F., Ji, X., Dai, Y. C., and Cui, B. K. (2018). Phylogeny and taxonomy of Laetiporus (Basidiomycota, Polyporales) with descriptions of two new species from western China. MycoKeys 37, 57–71. doi: 10.3897/mycokeys.37.26016
Stamatakis, A. (2014). RAxML Version 8: a tool for phylogenetic analyses and post analyses of large phylogenies. Bioinformatics 30, 1312–1313. doi: 10.1093/bioinformatics/btu033
Swofford, D. L. (2002). PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4.0 Beta. Sunderland, MA: Sinauer.
Thiers, B. (2022). Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. New York: New York Botanical Garden's Virtual Herbarium.
Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F., and Higgins, D. G. (1997). The Clustal_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucl. Acids Res. 25, 4876–4882. doi: 10.1093/nar/25.24.4876
White, T. J., Bruns, T., Lee, S., and Taylor, J. W. (1990). “Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics,” in PCR Protocols: A Guide to Methods and Applications, eds M. A. Innis, D. H. Gelfand, J. J. Sninsky, T. J. White (San Diego: Academic Press), 315–322. doi: 10.1016/B978-0-12-372180-8.50042-1
Wu, F., Li, S. J., Dong, C. H., Dai, Y. C., and Papp, V. (2020). The genus Pachyma (syn. Wolfiporia) reinstated and species clarification of the cultivated medicinal mushroom “Fuling” in China. Front. Microbiol. 11, 59078. doi: 10.3389/fmicb.2020.590788
Wu, F., Zhou, L. W., Vlasák, J., and Dai, Y. C. (2022). Global diversity and systematics of Hymenochaetaceae with poroid hymenophore. Fungal Divers. 113, 1–192. doi: 10.1007/s13225-021-00496-4
Xing, J. H., Sun, Y. F., Han, Y. L., Cui, B. K., and Dai, Y. C. (2018). Morphological and molecular identification of two new Ganoderma species on Casuarina equisetifolia from China. MycoKeys 34, 93–108. doi: 10.3897/mycokeys.34.22593
Yuan, Y., Chen, J. J., Korhonen, K., Martin, F., and Dai, Y. C. (2021). An updated global species diversity and phylogeny in the forest pathogenic genus Heterobasidion (Basidiomycota, Russulales). Front. Microbiol. 11, 596393. doi: 10.3389/fmicb.2020.596393
Keywords: brown rot, pathogenetic fungi, phylogeny, polypore, wood-decaying fungi
Citation: Yuan Y, Wu Y-D, Wang Y-R, Zhou M, Qiu J-Z, Li D-W, Vlasák J, Liu H-G and Dai Y-C (2022) Two new forest pathogens in Phaeolus (Polyporales, Basidiomycota) on Chinese coniferous trees were confirmed by molecular phylogeny. Front. Microbiol. 13:942603. doi: 10.3389/fmicb.2022.942603
Received: 12 May 2022; Accepted: 26 August 2022;
Published: 21 September 2022.
Edited by:
Bálint Dima, Eötvös Loránd University, HungaryReviewed by:
Peter Pristas, Pavol Jozef Šafárik University in Košice, SlovakiaCopyright © 2022 Yuan, Wu, Wang, Zhou, Qiu, Li, Vlasák, Liu and Dai. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yu-Cheng Dai, eXVjaGVuZ2RhaUBiamZ1LmVkdS5jbg==
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.