 Borja Aldeguer-Riquelme1
Borja Aldeguer-Riquelme1 Esther Rubio-Portillo1
Esther Rubio-Portillo1 José Álvarez-Rogel2
José Álvarez-Rogel2 Francisca Giménez-Casalduero3
Francisca Giménez-Casalduero3 Xose Luis Otero4
Xose Luis Otero4 María-Dolores Belando5
María-Dolores Belando5 Jaime Bernardeau-Esteller5
Jaime Bernardeau-Esteller5 Rocío García-Muñoz5
Rocío García-Muñoz5 Aitor Forcada3
Aitor Forcada3 Juan M. Ruiz5
Juan M. Ruiz5 Fernando Santos1
Fernando Santos1 Josefa Antón1,6*
Josefa Antón1,6*- 1Department of Physiology, Genetics, and Microbiology, University of Alicante, Alicante, Spain
- 2Department of Agricultural Engineering of the Escuela Técnica Superior Ingeniería Agronómica (ETSIA) & Soil Ecology and Biotechnology Unit of the Institute of Plant Biotechnology, Technical University of Cartagena, Cartagena, Spain
- 3Department of Marine Science and Applied Biology, University of Alicante, Alicante, Spain
- 4Cross-Research in Environmental Technologies (CRETUS), Departamento de Edafoloxía e Química Agrícola, Facultade de Bioloxía, Universidade de Santiago de Compostela, Santiago de Compostela, Spain
- 5Seagrass Ecology Group, Spanish Oceanography Institute of the Spanish National Research Council, Oceanography Center of Murcia, Murcia, Spain
- 6Multidisciplinary Institute of Environmental Studies Ramón Margalef, University of Alicante, Alicante, Spain
Coastal marine lagoons are environments highly vulnerable to anthropogenic pressures such as agriculture nutrient loading or runoff from metalliferous mining. Sediment microorganisms, which are key components in the biogeochemical cycles, can help attenuate these impacts by accumulating nutrients and pollutants. The Mar Menor, located in the southeast of Spain, is an example of a coastal lagoon strongly altered by anthropic pressures, but the microbial community inhabiting its sediments remains unknown. Here, we describe the sediment prokaryotic communities along a wide range of environmental conditions in the lagoon, revealing that microbial communities were highly heterogeneous among stations, although a core microbiome was detected. The microbiota was dominated by Delta- and Gammaproteobacteria and members of the Bacteroidia class. Additionally, several uncultured groups such as Asgardarchaeota were detected in relatively high proportions. Sediment texture, the presence of Caulerpa or Cymodocea, depth, and geographic location were among the most important factors structuring microbial assemblages. Furthermore, microbial communities in the stations with the highest concentrations of potentially toxic elements (Fe, Pb, As, Zn, and Cd) were less stable than those in the non-contaminated stations. This finding suggests that bacteria colonizing heavily contaminated stations are specialists sensitive to change.
Introduction
Coastal lagoons are highly biodiverse and productive environments that provide critical ecosystem services with significant socioeconomic impacts such as food provision or climate regulation (Newton et al., 2018). They are shallow water bodies separated from the ocean by a barrier and connected to it by one or more restricted inlets (Kjerfve, 1994). Due to their proximity to land, small volumes, and long water renewal time, coastal lagoons are extremely sensitive to anthropic eutrophication and pollutant accumulation, such as agricultural nutrient loading, fishing, tourism, and mining (Barbier et al., 2011; Pérez-Ruzafa and Marcos, 2012), which may promote sudden and drastic changes in ecosystem structure, functions, and services (De Wit et al., 2001).
Sediments are a key compartment for the functioning and quality of coastal lagoons by acting as a sink for organic matter (Hewson and Fuhrman, 2008), potentially toxic elements (PTEs) (i.e., metals and metalloids), and inorganic nutrients such as N or P (Jiménez-Cárceles et al., 2006; Sharifuzzaman et al., 2016). Conversely, sediments can transfer nutrients and PTEs into the water column by diffusion or when sediment particles are suspended by winds and currents. Due to the presence of a high nutrient load in sediments, resident microbial communities are usually very dense, reaching up to 109 cells/g (Luna et al., 2002; Petro et al., 2019), which is 3-4 orders of magnitude higher than microbial densities in the water column (DeLong et al., 1999; Eilers et al., 2000). Furthermore, diffusion of O2 through the sediment is limited (Hewson and Fuhrman, 2008), which leads to redox zoning and changes in microbial metabolism. At the surface, organic matter is oxidized by aerobic microbes that deplete available O2; in deeper layers, anaerobes use alternative electron acceptors such as NO3–, MnO2, amorphous FeOOH, and SO42– (Sørensen et al., 1979). Due to their high abundances and metabolic diversity, microorganisms in marine sediments are a key factor in the remineralization of nutrients (Alongi, 1994, 1998); and thus, they have a strong influence on the biogeochemical cycles of coastal marine ecosystems.
The Mar Menor, located in the southeast of Spain (Figure 1), is the largest hypersaline (i.e., 42–47 psu) coastal lagoon in Europe and provides a paradigmatic example of a coastal marine ecosystem that is highly valuable yet strongly altered by several anthropogenic factors (Marín-Guirao et al., 2005; Conesa and Jiménez-Cárceles, 2007; León et al., 2013, 2017, 2018; Jiménez-Martinez et al., 2016; Romero et al., 2020). In the 1970s, one inlet (El Estacio channel) connecting the lagoon with the Mediterranean Sea was broadened (Mas, 1994), drastically changing the hydrodynamic regime and decreasing the lagoon temperature and salinity toward the values closer to those typically measured in the adjacent Mediterranean Sea (37 psu). This event, together with the massive loading of urban sewage into the lagoon, allowed the spread of the opportunistic chlorophyte Caulerpa prolifera (Ballester, 1985; Terrados and Ros, 1992). Furthermore, due to the past extractive activity in the Cartagena-La Unión mining district, located close to the southern basin (Conesa and Schulin, 2010), PTEs such as Pb, Zn, Cd, and As have been loaded into the lagoon by hydric and aeolian erosion and surface and subsurface runoffs. These PTEs accumulate in the sediment (Simonneau, 1973; García and Muñoz-Vera, 2015) or are taken up by vegetation and fauna (Serrano et al., 2019; Romero et al., 2020). In addition, the increase in tourism activity since the 1960s, and the effects of intensive agriculture since the 1970s, have deeply transformed the surrounding territory and poured huge amounts of sediments, nutrients, and contaminants into the lagoon (Jiménez-Martinez et al., 2016; Pérez-Ruzafa et al., 2019).
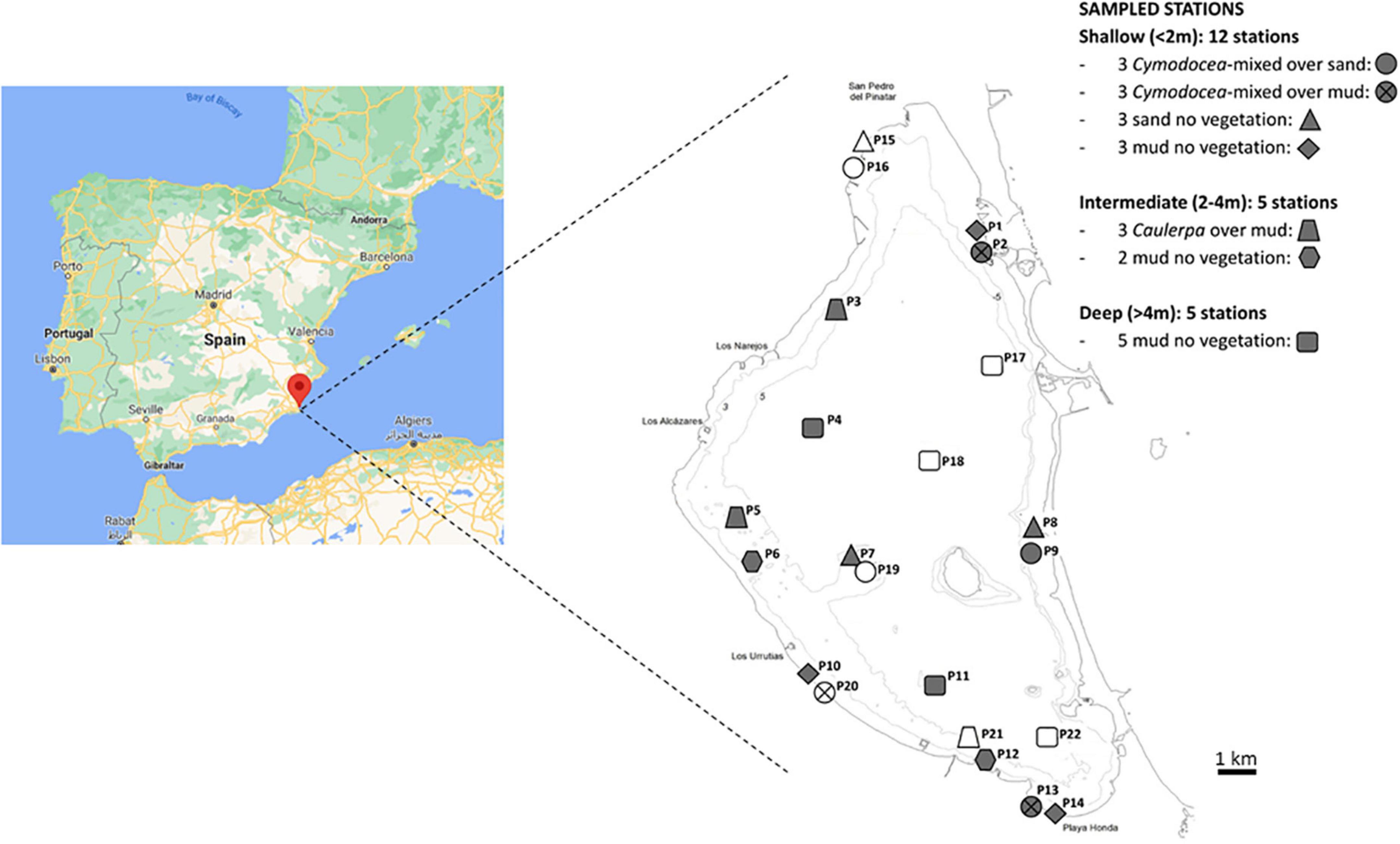
Figure 1. Geographic location and sampling stations in the Mar Menor lagoon. Levels of different factors (depth, vegetation, and texture) are indicated by symbols (see legend at the right) for each sampling site. The symbols colored in gray indicate stations selected for microbiological studies.
These impacts have shifted the lagoon from an oligotrophic system dominated by the seagrass Cymodocea nodosa (Ballester, 1985), to an eutrophic system that caused the macroalga C. prolifera to rapidly colonize, forming monospecific or mixed meadows with C. nodosa (Ballester, 1985; Terrados and Ros, 1991) (hereafter we will refer to these species as Caulerpa and Cymodocea, respectively). Contaminants of emerging concern, such as pesticides, polycyclic aromatic hydrocarbons (PAHs), surfactants, and pharmaceuticals, have also been detected in the lagoon-associated watercourses, water columns, sediments, and even in bivalves and fishes (Moreno-González et al., 2014, 2016; Jiménez-Martinez et al., 2016). Furthermore, the lagoon experienced a unique and severe event of the so-called “ecosystem disruptive algal bloom” as a consequence of a phytoplankton overgrowth initiated in 2016 by the cyanobacterium Synechoccocus (Aguilar-Escribano et al., 2016; Mercado et al., 2021). This caused the water turbidity to increase up to critical levels for macrophyte survival and led to the sudden loss of 82% of benthic vegetation, including Caulerpa and Cymodocea (Belando-Torrente et al., 2019; Marín-Guirao et al., 2019).
Some studies have addressed the effects of these anthropic impacts on the macro-organisms inhabiting the lagoon (Muñoz-Vera et al., 2015; Serrano et al., 2019; Romero et al., 2020) and on benthic food webs (Marín-Guirao et al., 2008). However, the information available on the microbial ecology of the system is very scarce and restricted to the water column (Gilabert, 2001; Ghai et al., 2012). Thus, despite the ecological relevance of sediment microbiota in coastal lagoons, to date, nothing is known about this component of the microbial assemblage in the Mar Menor. To fill this gap, we aimed at ascertaining whether (i) the taxonomic composition and inferred putative metabolic capabilities of the Mar Menor sediments were similar to that of other marine sediments, (ii) factors such as texture, depth, vegetation, and geographical location could be linked to the microbial community structure, and (iii) microbial communities in each station underwent temporal changes.
Our results showed that the microbial communities in the analyzed sediments were highly heterogeneous among stations, although a core microbiome could be detected. Sediment texture, vegetation type, depth, and geographical location were found to drive the structure of the microbial community, while time was only relevant in the most PTE contaminated stations. Mining activity-derived PTEs, which were present in extremely high concentrations in the south of the lagoon, also affected prokaryotic composition and stability, indicating the capacity of sustained human pressures to alter sediment microbiomes and, likely, their ecosystem functions.
Materials and methods
Study area
The Mar Menor is a hypersaline shallow coastal lagoon (maximum depth 7 m and average depth 4.5 m) located in the Region of Murcia, southeast of Spain, with an area of 135 km2. It is separated from the Mediterranean Sea by a sand bar of 22 km length (Figure 1) and connected to it by three small inlets through which water exchanges between both water bodies occur. The Mar Menor water mass is then relatively confined with a renewal time of about 1 year (Fraile-Nuez et al., 2018) and has a salinity range of 42–47 psu (Aravio-Torre and Arévalo, 1973), higher than that of the adjacent Mediterranean Sea in this area (37–38.5 psu; Pérez-Ruzafa et al., 2004). Water temperature varies from 10°C in winter to 31°C in summer, without a significant stratification in the water column (Fraile-Nuez et al., 2018). Nitrate concentrations were below 1 μM until the 1990s when agricultural and urban wastes increased the concentration above 8 μM (Ruiz et al., 2020). Curiously, chlorophyll concentrations did not significantly increase above 1–3 mg/m3 until 2016, when the macrophyte populations in the Mar Menor lagoon collapsed due to a massive phytoplankton bloom (Belando-Torrente et al., 2019; Pérez-Ruzafa et al., 2019; Ruiz et al., 2020).
Field sampling
A total of 22 sampling stations were distributed across the lagoon (Figure 1) based on depth (<2, 2–4, and >4 m), sediment texture (sandy and muddy), and vegetation (bared, Caulerpa and Cymodocea) according to the information from previous studies (bathymetry from MAGRAMA, 2008; sediment characteristics from Álvarez-Rogel et al., 2019; macrophytes distribution from Belando et al., 2021). At each sampling point, in March and September of 2018, cold and warm season, respectively, and both characterized to be dry seasons in this area, a lineal transect of 25 m was fixed, depth and presence of vegetation was validated in situ by divers, and sediment samples were collected using cores (7-cm diameter) along the transect area as follows. Three surface sediment samples (4.5 cm) were collected for microbial analysis using cores (7 cm diameter) that were distributed at the beginning, center, and end of each transect. For physicochemical determination, five replicates per sampling point, collected every 5 m of the line transect, were homogenized prior to the analyses. The samples were not split by the depth due to the heterogeneous vertical profiles of physicochemical parameters in the first 4 cm among the samples from different stations, which hampered the definition of a vertical common cutoff for the oxic–anoxic boundary. The samples were taken to the laboratory in a portable 4°C cooler.
For microbial analyses, the individual sediment samples were weighted and manually homogenized after adding 50 ml of 1X PBS containing 3.5% NaCl. Then, for cell density measurements, 1 ml of each sample was fixed with formaldehyde (4% final concentration, 4 h at 4°C) and the fixation stopped with 9 ml of 1X PBS. Cell counts in each station were determined by epifluorescence microscopy after DAPI staining (Gomariz et al., 2015). The remaining homogenized sediments were stored at –80°C until processed for nucleic acid extraction (see below).
Physicochemical measurements
Sediments for physicochemical analysis were processed immediately upon arrival at the laboratory. The redox potential (Eh) and pH values were measured three times per homogenized sample by inserting Eh and pH electrodes (Crison 50–55 and Crison 50–50, respectively). The Eh values were adjusted, according to Vepraskas and Faulkner (Vepraskas and Faulkner, 2001), by adding +200 mV to the measured voltage (the value of the Ag/AgCl reference electrode at 20°C). Then, sediment interstitial water was extracted by centrifugation at 11,300 g in a refrigerated centrifuge at 4°C under a nitrogen atmosphere. The dissolved sulfide was immediately measured (Cline, 1969). The rest of the interstitial water samples were stored at 4°C to measure electrical conductivity (EC), dissolved organic carbon (DOC), NO3– and NH4+ within less than 1 week after the sampling. The DOC was measured in a TOC-automatic analyzer (TOC-VCSH Shimadzu). The NO3– and NH4+ concentrations were measured with a V/UV spectrometer at λ = 220 nm and at λ = 670 nm, respectively, and the interference by organic matter was corrected by measuring the absorbance at λ = 275 nm (AOAC, 1975; Neiker, 2005). The solid material was dried at room temperature. Each dried sample was passed through a 2-mm mesh and a fraction larger than this size (mainly large mollusk shells) was discarded. The particle size was assessed with the 2-mm sieved aliquots, which were put in an ultrasonic bath for 10 min to promote particle dispersion. Then, they were sieved to 50 μm under a stream of water. Both fractions (sand, from 2 mm to 50 μm; silt + clay, <50 μm) were dried and weighed and the percentage of each one was calculated. Aliquots of the 2-mm sieved samples were crushed in an agate mortar for measuring total element concentrations (X-ray fluorescence). The amorphous or poorly crystalline Fe oxyhydroxides (FeA) were extracted by shaking 2-mm sieved samples for 6 h at 96°C with 30 ml 0.04 M hydroxylamine + 25% acetic acid (v/v) (Fortin et al., 1993). The degree of Fe pyritization (DOP) was calculated to establish the percentage of reactive iron incorporated into the pyrite fraction. The calculation was made using Eq. 1, assuming reactive-Fe to be extracted with dithionite (DOP-dithionite), according to the recommendations of Raiswell et al. (1994).
The concentration of acid volatile sulfides (AVS = ΣFeS, Fe3S4, H2S) was determined in triplicate using 0.5–1.0 g of wet sample, according to the method described by Kostka and Luther (1994). Sulfide from AVS was liberated with 20 ml 3N HCl previously deaerated for 40–50 min. The sample was digested in a gas-tight reaction flask for 40–50 min under a continuous flow of nitrogen, which was bubbled through the flask as slowly as possible. The evolved H2S was then received in a trap containing 25 ml of 3% Zn acetate, 1 ml of concentrated H2SO4, and 4 ml of diamine reagent, and precipitated as ZnS. Sulfide was then measured colorimetrically with a UV-VIS spectrophotometer at a wavelength of 670 nm using the methylene blue method of Cline (Cline, 1969). Metal(loid)s simultaneously extracted with AVS (SEM-AVS), which is an approach to assess the potential for metal ions found in sediment to cause toxic effects in sediment-dwelling organisms, were analyzed by ICP-OES (Perkin Elmer, Optima model 4300 DV, Sunny vale, CA, United States) in 3N HCl.
DNA extraction, 16S rRNA gene PCR amplification, and sequencing
Based on a 3D PCA with the physicochemical characteristics (Supplementary Figure 1), fourteen stations (corresponding to 82 samples, including triplicates per station and sampling time, except P8 with duplicates) were selected as representative of all the physicochemical and environmental conditions in the Mar Menor sediments and were analyzed microbiologically.
To extract DNA, the RNeasy Power Soil Total RNA Kit (Qiagen) and its complementary RNeasy Power Soil DNA Elution Kit (Qiagen) were used. Two grams of the homogenized sediment samples, stored at –80°C, were used according to the manufacturer’s instructions. Qubit™ dsDNA High Sensitivity (HS) or Broad Range (BR) (Invitrogen) kits were employed to measure DNA concentration.
The 82 extracted DNAs were used for PCR amplification of the 16S rRNA genes V3-V4 regions, using 25 cycles and primers 341F–805R (Takahashi et al., 2014). The PCR products from three independent reactions per sample were pooled and sequenced on an Illumina MiSeq using a 2 bp × 250 bp run (at Fundació per al Foment de la Investigació Sanitària i Biomédica, FISABIO, Valencia, Spain). Sequences were quality filtered removing sequences shorter than 50 bp or with a Phred quality below 30 by employing prinseq-lite (Schmieder and Edwards, 2011), and R1 and R2 reads were joined by FLASH (Magoč and Salzberg, 2011) using default parameters. Assembled sequences were analyzed using Qiime (Caporaso et al., 2010) for: (i) removing chimeras, using usearch6.1; (ii) clustering sequences in operational taxonomy units (OTUs) with 97% identity, using pick_open_reference_otus.py function; and (iii) assigning taxonomy of the longest sequence in each OTU, by BLAST against SILVA_132_release database at 97% identity. All samples were normalized at 47,461 sequences (the lowest number of sequences in a sample after post-assembly and filtering). To visualize the diversity and phylum or class abundances at each station and time of sampling, bubble plots were drawn in R with ggplot2 (Wickham, 2016).
Statistical analysis
Due to the limitation of analyzing microbiologically the total amount of samples collected, and with the aim of selecting a set of stations representative of all the physicochemical and environmental conditions in the Mar Menor sediments, a principal component analysis (PCA) of the environmental variables was carried out using “prcomp.” The environmental data were normalized using the function “decostand” of the vegan package (Oksanen, 2011) with the parameter “range.” Correlations among the environmental variables were assessed by the function “cor” and variables with correlations coefficients above 0.85 were considered correlated.
Rarefactions curves were generated in R with the rarecurve function of the “vegan” package. The α-diversity measures (Shannon index and observed OTUs) were calculated with the “estimate_richness” function of the “phyloseq” package (McMurdie and Holmes, 2013). In addition, unique and shared OTUs among samples were displayed with the “UpSet” (visualizing intersecting sets) diagram using the “R- bioconductor” package “UpSetR” (Conway et al., 2017). For this analysis, we only considered OTUs above 0.1% in each sample which could be considered an abundant biosphere given the high diversity of these samples.
An analysis of variance (ANOVA) was used to test for statistical differences in physicochemical variables, concentration of cells, observed OTUs, Shannon index, and relative abundance of detected microbial classes among the levels of each of the following factors considered: Texture (levels: mud and sand), time (levels: March and September), vegetation (levels: bared, Caulerpa and Cymodocea), depth (levels: shallow, intermediate, and deep), and zone (levels: north, center, and south). Prior to ANOVA, homogeneity of variance was confirmed with Levene’s test (Levene, 1960). All p-values below 0.05 were considered significant.
To identify the main environmental factors that explain the changes in the microbial communities among the sampled stations, Bray–Curtis distances were calculated using the “vegdist” function of the “vegan” package, and a NMDS analysis was carried out. SIMPER analysis (Clarke, 1993) was used to identify species potentially responsible for statistically significant differences among levels of each factor using “vegan” (Oksanen, 2011).
Differences in the multivariate structure of microbial communities of each station among sampling times were tested using permutational multivariate analysis of variance (PERMANOVA) (Anderson, 2001; McArdle and Anderson, 2001) considering two factors (time and station). Wherein each term in the analysis was tested using 4,999 random permutations of the appropriate units and similarities among samples were calculated using the Bray–Curtis similarity index. Prior to PERMANOVA, homogeneity of multivariate dispersions among groups was tested with “betadisper.” All p-values below 0.05 were considered significant.
To get an explanation concerning the different behavior of the microbial communities between the two sampling times in the contaminated and non-contaminated stations, observed with the above PERMANOVA test, several statistical analyses were performed. First, contaminated and non-contaminated stations were defined by clustering the stations by FeA, PbAVS, and ZnAVS concentrations (indicators of metal pollution) using the function “agnes” and the ward method of the “cluster” R package (Supplementary Figure 2) (Maechler et al., 2019). Then, PERMANOVA with two factors: Time (levels: March–September) and PTEs [levels: contaminated (stations 4, 10–14) and non-contaminated (stations 1–3, 5 and 6)] was employed to test for differences in physicochemical parameters, following the same procedure explained above. Finally, to identify the variables responsible for the differences reported by PERMANOVA, a two-way ANOVA with the same factors was tested on each environmental variable. Additionally, using SIMPER analysis, metallophilic, intermediate, and non-metallophilic OTUs were defined based on the fold change among the mean abundance in the contaminated and non-contaminated stations. Metallophilic OTUs were defined as those having a fold change equal to or higher than 2, intermediate OTUs as those with fold change values higher than 0.5 but lower than 2, and non-metallophilic OTUs included those with a fold change lower than 0.5.
Results and discussion
General characteristics of the sediments
Based on a 3D PCA constructed with the physicochemical variables, 14 stations (among the 22 sampled stations; Supplementary Figure 1) that covered the variability of physicochemical conditions included in the plot, were selected as representative and analyzed more deeply. In these stations, redox potential (Eh) ranged from 197 to −240 mV with significantly lower values in muddy than sandy sediments (Supplementary Table 1.1 and Supplementary Figure 3). The Eh was significantly more negative in September than in March and sulfide (S2–) presented significantly higher concentrations in September, probably reflecting a higher activity of sulfate-reducing bacteria in the summer as observed in other systems (Robador et al., 2009). The pH values varied from nearly neutral to alkaline (pH: 7.1–8.4), with a tendency to lower values at muddy stations closer to the coast. Soluble organic carbon (SOC) was significantly higher in vegetated than in bare sediments with the highest values found in sediments colonized by Cymodocea. On the other hand, TOC displayed significantly higher concentrations in muddy than in sandy sediments, mainly in the north. Regarding DOP, which is related to sulfate reduction (Raiswell et al., 1988), statistically significant differences were observed in depth, with the higher values found in the deepest sediments, while NH4+, presented statistically significant differences between zones, with the higher values found in the south.
Acid volatile sulfide (AVS) was detected in very high concentrations in muddy sediments (≈2.8–141 mg kg–1), especially in the south of the lagoon, displaying statistically significant differences for both factors (texture and zone). High AVS concentrations in the Mar Menor bed had been previously reported (Marín-Guirao et al., 2005) and might be related to a high sulfate-reducing rate in the sediment favored by the high content of organic matter and the fine texture (Thode-Andersen and Jørgensen, 1989; Llobet-Brossa et al., 2002). Similarly, high concentrations of PTEs evaluated by means of FeAVS, ZnAVS, PbAVS, CdAVS, and AsAVS were found in almost all the muddy stations in both samplings, with high spatial variability (Supplementary Table 1.1). Particularly, stations located in the south of the lagoon (stations 10–14, see Figure 1), showed significant and unusually high concentrations of PbAVS (≈1,000–8,600 mg kg–1) and ZnAVS (≈600–3,900 mg kg–1). These results indicated metal(loid) pollution, mainly in the southern part of the lagoon, due to the runoff of mine wastes from the old mining area of Sierra de Cartagena-La Unión, in agreement with previously reported data (Marín-Guirao et al., 2008; María-Cervantes et al., 2009). Since PbAVS was correlated with CdAVS and FeA with FeAVS and ZnAVS, hereinafter we will only analyze PbAVS and FeA although their individual effects cannot be distinguished.
Microbial abundances
Cell concentrations ranged from 6.6 × 107 to 130 × 107 cells/g of sediment (Figure 2), values in the ranges previously reported for other marine sediments (Luna et al., 2002; Sørensen et al., 2007; García-Martínez et al., 2009). Statistically significant differences in cell concentrations were detected for texture (p = 0.000121), with higher concentrations in muddy than in sandy sediments (42 × 107 to 130 × 107 cells/g vs. 6.6 × 107 to 32 × 107 cells/g) and for zone (p = 0.0131), but not for time (p = 0.115), depth (p = 0.244), or vegetation (p = 0.457) (Supplementary Figure 4). Note that since the sandy samples were only present in the center of the lagoon, this zone presented lower cell concentrations than the north and the south. Despite no statistical differences being found for the factor time, it is noteworthy that microbial abundances were higher in September than in March for most of the studied stations (Figure 2), which is in agreement with previous studies (Musat et al., 2006; Suárez-Suárez et al., 2011).
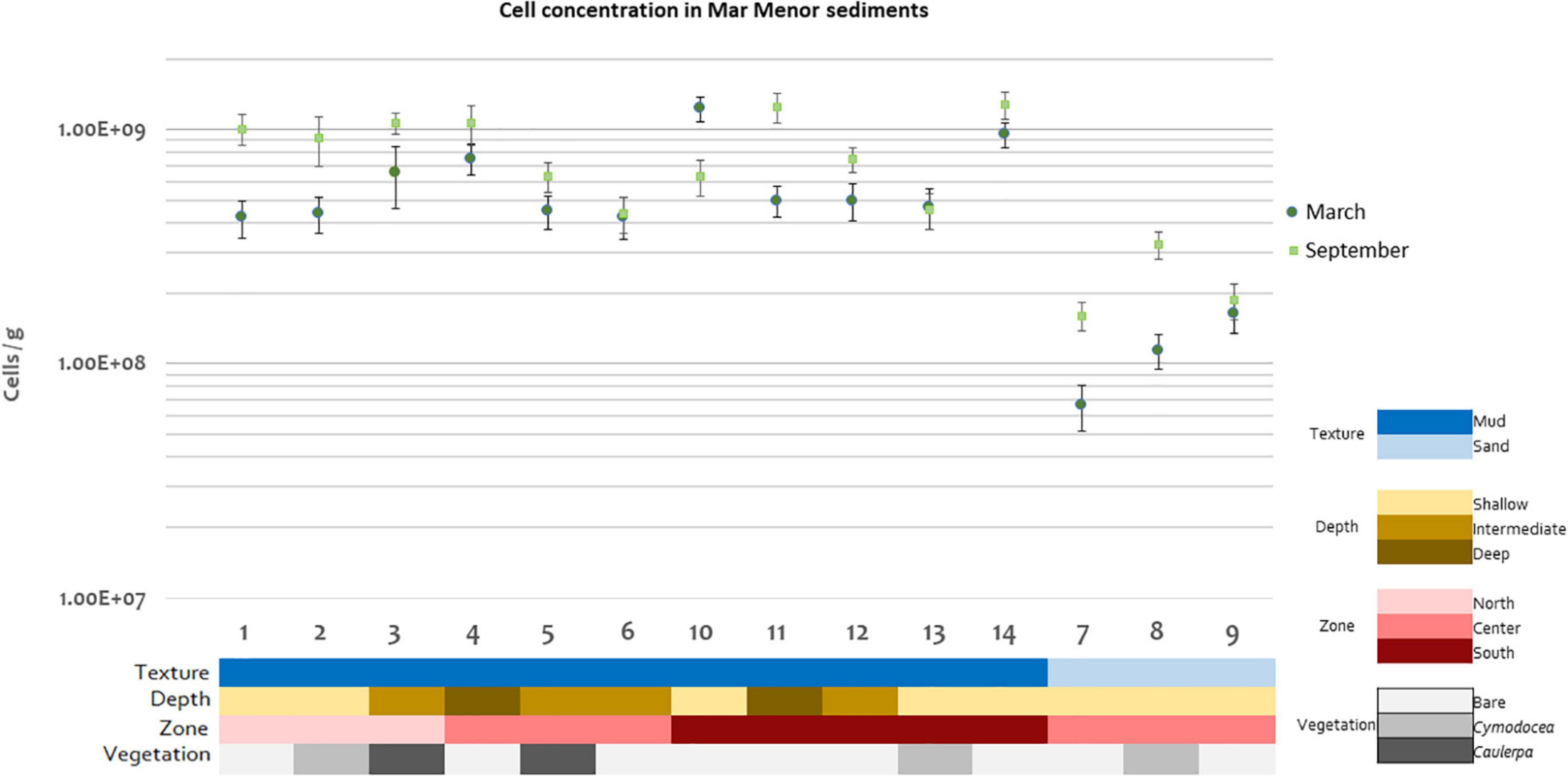
Figure 2. Microbial cell concentrations for each Mar Menor sediment station and time of sampling in logarithmic scale. One replicate per station and sampling time is shown. Error bars indicate standard deviation in DAPI count.
Overall microbial community description
The prokaryotic community of the Mar Menor sediments was analyzed by 16S rRNA gene metabarcoding. Although this is a powerful tool to investigate microbial community structure and dynamics, it has several limitations. For instance, biases due to different numbers of ribosomal RNA operons (Kembel et al., 2012), choice of amplification primers (Klindworth et al., 2013), and the amplification process (Suzuki et al., 1998; Sipos et al., 2007) have been reported. Furthermore, the bioinformatic pipeline is another source of bias, and Qiime1 is known to be very sensitive but usually overestimates diversity (Straub et al., 2020). Additionally, there is the possibility that some of these OTUs are derived from extracellular DNA from lysed cells (Torti et al., 2018). We are aware of these limitations and have tried to avoid overreaching conclusions.
For the microbiological analyses, DNA was extracted, amplified, and sequenced from 82 samples (including triplicates, except P8 with duplicates). A total of 3,891,802 reads were obtained and were grouped at 97% of similarity in 324,325 OTUs for the 82 samples after quality filtering, clustering, and normalizing (Supplementary Tables 1.2, 1.3). The rarefaction analysis indicated a good recovery of the microbial diversity inhabiting the sediments although the asymptote was not reached (Supplementary Figure 5). Regarding alpha diversity, the number of OTUs (richness) varied between 7,240 and 15,942, while the Shannon index values ranged from 6.7 to 8.5 (Figure 3A). The ANOVA test showed statistically significant differences in richness, and the Shannon index between textures (Shannon p = 0.009773; richness p = 0.001014) and zone (Shannon p = 2.695 × 10–6; richness p = 5.91 × 10–6), with higher values in muddy sediments and in the north area of the Mar Menor. However, similar values were found among times (Shannon p = 0.519; richness p = 0.122), depths (Shannon p = 0.567; richness p = 0.8468), or vegetations (Shannon p = 0.3696; richness p = 0.329) (Supplementary Figure 6).
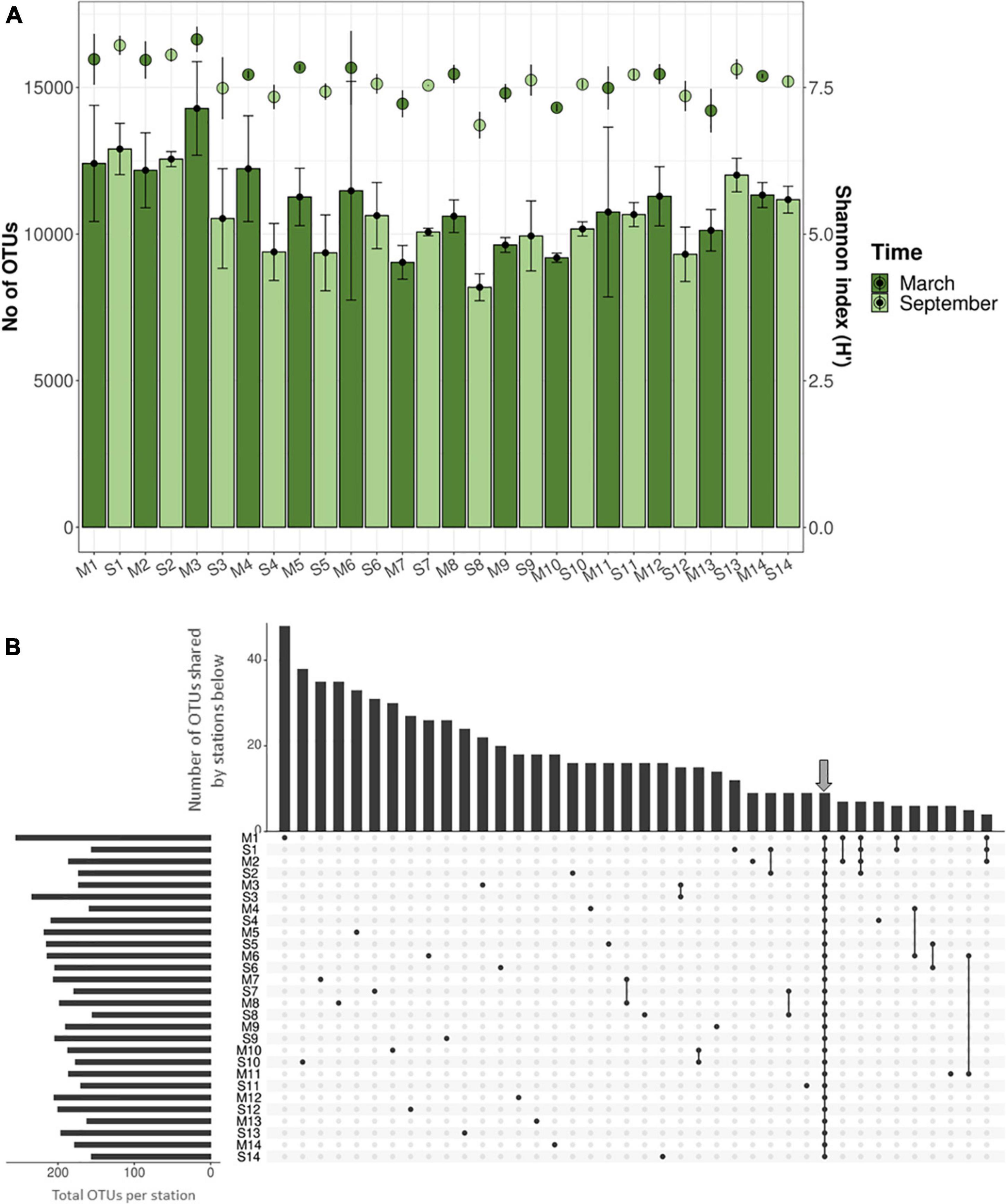
Figure 3. (A) Number of OTUs (columns) and Shannon index estimates (dots) for rarefied samples. Deviation bars indicate variability within replicates of each station and sampling time. (B) Upset plot showing the OTU distribution (for OTUs > 0.1% relative abundance) along the 14 stations during the cold and warm seasons (March and September, respectively). At the top, columns indicate the number of OTUs shared by stations, which are marked at the bottom with black dots. Arrow indicates the core microbiome.
The UpSet analysis, considering only OTUs with relative abundances above 0.1% [the “abundant” biosphere (Pedrós-Alió, 2012)], revealed that most of them were specific to individual stations with only a small proportion of shared OTUs (Figure 3B), highlighting the heterogeneity among stations. Nevertheless, a core microbiome of nine OTUs was detected in all stations and sampling times which represented a substantial fraction of the whole microbial community (from 2.4 up to 13.8% of total sequences) (Supplementary Table 1.4). The previous studies in intertidal sand and marine sediments (Sun et al., 2013; Boehm et al., 2014; Quero et al., 2015) have also detected both cosmopolitan (i.e., members of the core) and sample-specific microorganisms, likely representing generalists (i.e., members of the core) and specialists, respectively.
Microbial community structure
Proteobacteria was the phylum most frequently detected in all stations, accounting for 20.5–55.8% of the retrieved sequences (Supplementary Figure 7), followed by Bacteroidetes and Chloroflexi (4.3–26.4 and 1.9–17%, respectively). At the class level, Deltaproteobacteria (12.7–32.6%), now referred to as phylum Desulfobacterota, and Gammaproteobacteria (4.6–30.1%) dominated almost all the stations and times, with the exception of Bacteroidia dominating sample S7 and stations 1 and 2, both located on the channel connecting the lagoon with the Mediterranean Sea (Figure 4). The previous results on marine sediments from different locations also indicated the dominance of Proteobacteria and Bacteroidetes (Mahmoudi et al., 2015; Quero et al., 2015).
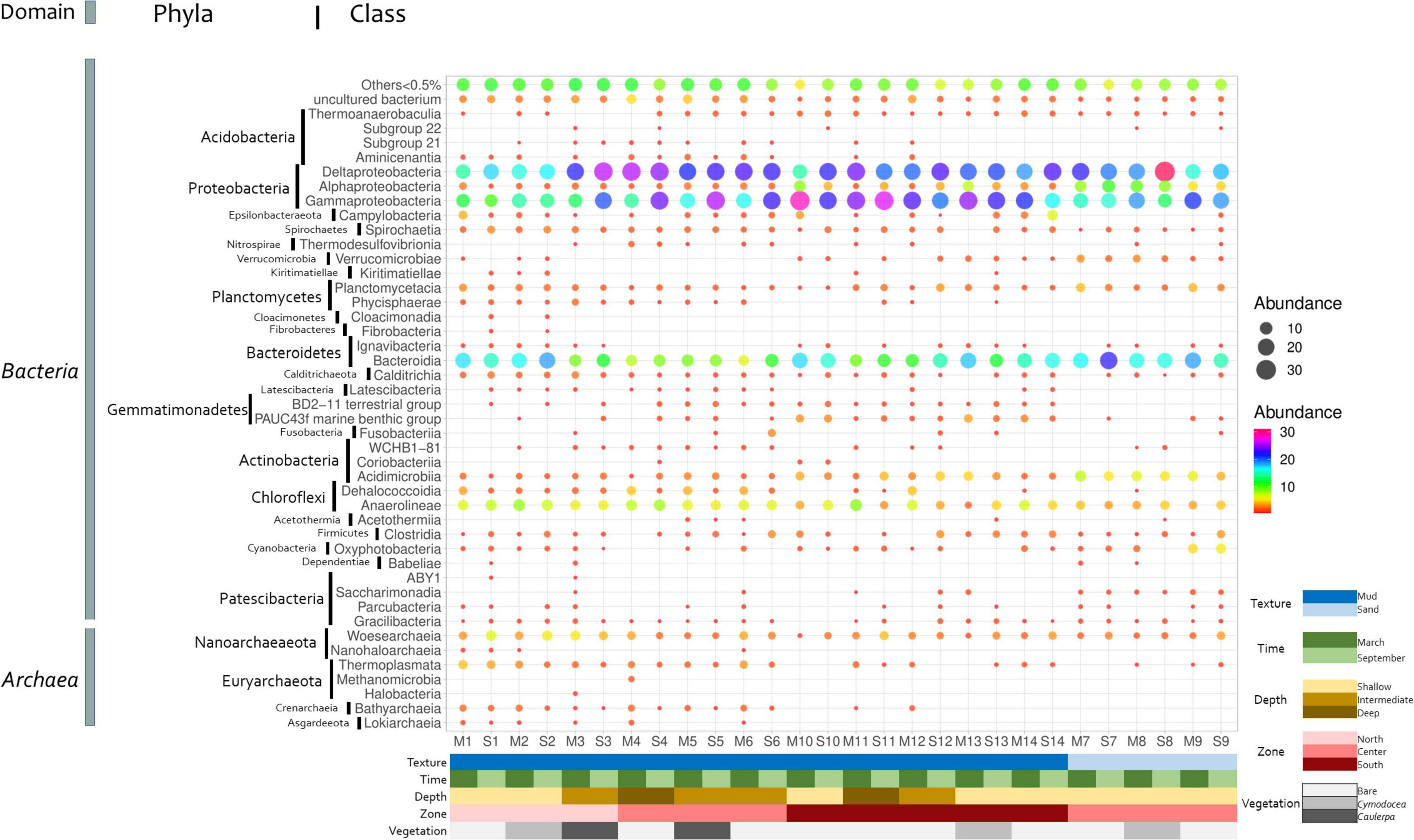
Figure 4. Distribution of bacterial and archaeal classes in Mar Menor sediments. Dot color and size indicate the relative abundance of each class. The main features of each station are shown at the bottom.
Although 16S rRNA gene metabarcoding provides little information regarding the metabolic capabilities of the community (Sun et al., 2020), some putative metabolisms can be hypothesized for some of the detected phyla in the Mar Menor sediments. For instance, the most frequently retrieved orders from the Deltaproteobacteria class were Desulfobacterales and Desulfuromonadales, which are typically composed of sulfate- and sulfur-reducing bacteria (Madigan et al., 2019). The order Myxococcales was also abundant in some samples, although very little is known about their physiology and metabolism in marine environments (Brinkhoff et al., 2012). This order has been associated with organic matter degradation in the seagrass rhizosphere (Ugarelli et al., 2017) and in Posidonia oceanica decaying material (Rubio-Portillo et al., 2021). In soil samples, members of Myxococcales present an aerobic metabolism, although some representatives are anaerobes that respire nitrate or Fe (III) (Treude et al., 2003) and produce a wide range of secondary metabolites (Reichenbach, 2015).
Within the Gammaproteobacteria class, the Steroidobacterales order was predominant in many stations, mainly due to the abundance of Woeseia, a genus with a versatile metabolism that goes from sulfur and hydrogen oxidation to chemorganoheterotrophy (Mußmann et al., 2017). The Chromatiales and Ectothiorhodospirales orders were also abundant. These groups grow autotrophically, acting as primary producers, using light and the energy provided by the oxidation of reduced sulfur compounds (Mori et al., 2015; Madigan et al., 2019; Sorokin et al., 2020).
The most frequently retrieved orders of the Bacteroidia class were Flavobacteriales and Bacteroidales, well-known degraders of polymeric organic matter (Fernández-Gómez et al., 2013; Bernardet, 2015; Krieg, 2015). Within this class, some OTUs could be assigned to genera within Flavobacteriales, which are aerobic or facultative aerobic bacteria with an obligate chemorganoheterotrophic metabolism (Cho and Giovannoni, 2004; Kim et al., 2008; Park et al., 2010; Choi et al., 2013; Bernardet, 2015). The OTUs assigned to Bacteroidales corresponded to uncultured genera, which hampers any metabolic inference, although most of the cultured members of this order are strict or facultative anaerobes (Krieg, 2015).
Considering only nine OTUs that formed the “abundant” core microbiome (Figure 3B and Supplementary Table 1.4), sulfate reduction and sulfur oxidation appeared also as potentially relevant and widespread metabolisms since three of these OTUs belonged to the Desulfobacterales and one to the Chromatiales. Furthermore, two OTUs related to Bacteroidetes were included in this core, highlighting the putative importance of polymeric organic matter degradation in the system.
In summary, based on the 16S rRNA gene diversity data, assuming all the limitations of this approach (Crosby and Criddle, 2003; Soergel et al., 2012; Ghyselinck et al., 2013; Rubio-Portillo et al., 2016) and the high proportion of OTUs related to uncultured microbes in the analyzed samples, the most frequently retrieved sequences corresponded to microorganisms related to S and C cycles, as expected for marine sediments (Plugge et al., 2011; Urich et al., 2014; Fike et al., 2015).
However, Mar Menor sediments also harbored a considerable degree of novelty since several uncultured groups were found in relatively high proportions. Sequences corresponding to the Asgardeota phylum accounted for up to 1.7% of the total sequences in some stations. Three classes of this phylum, Lokiarchaeia, Heimdallarchaeia, and Odinarchaeia, could be detected at relative abundances of 1.4, 0.3, and 0.2%, respectively. Asgardarchaeota, have been found in different aquatic sediments, and are known by their close phylogenetic relationship with Eukarya, thereby suggesting the emergence of eukaryotes within this phylum (Zaremba-Niedzwiedzka et al., 2017; Bulzu et al., 2019). The relative abundance of sequences related to Heimdallarchaeia, which seems to be the closest group to eukaryotes (Bulzu et al., 2019; Williams et al., 2020), is the highest reported so far.
Sequences corresponding to the Woesearchaeia class, within the phylum Nanoarchaeota and now classified as the order Woesearchaeales, were present in high proportions, accounting for up to 5.7% of the community. This group has been extensively detected around the world in many different ecosystems through 16S rRNA gene sequence studies, but little is known about their ecology and metabolism. The recent studies have shown the high diversity of this group and pointed to the oxic status as the main factor driving the distribution of Woesearchaeal members (Liu et al., 2018). A symbiotic or syntrophic lifestyle has been proposed although it has not been confirmed as a general trait of the group (Liu et al., 2018).
Finally, sequences from the Actinobacteria class WCHB1-81 accounted for up to 1.1% of the total. This group has been only described by 16S rRNA gene analyses so its metabolism remains completely unknown.
Environmental factors driving microbial composition of sediment communities
To identify the main environmental factors related with the changes in the microbial communities, a NMDS plot based on Bray–Curtis distances of the microbial composition and abundance was performed. As shown in Figure 5, the texture (mud vs. sand), the type of vegetation (Caulerpa vs. Cymodocea), the depth (shallow vs. intermediate/deep), and the zone (north vs south) clustered the samples while factor time did not.
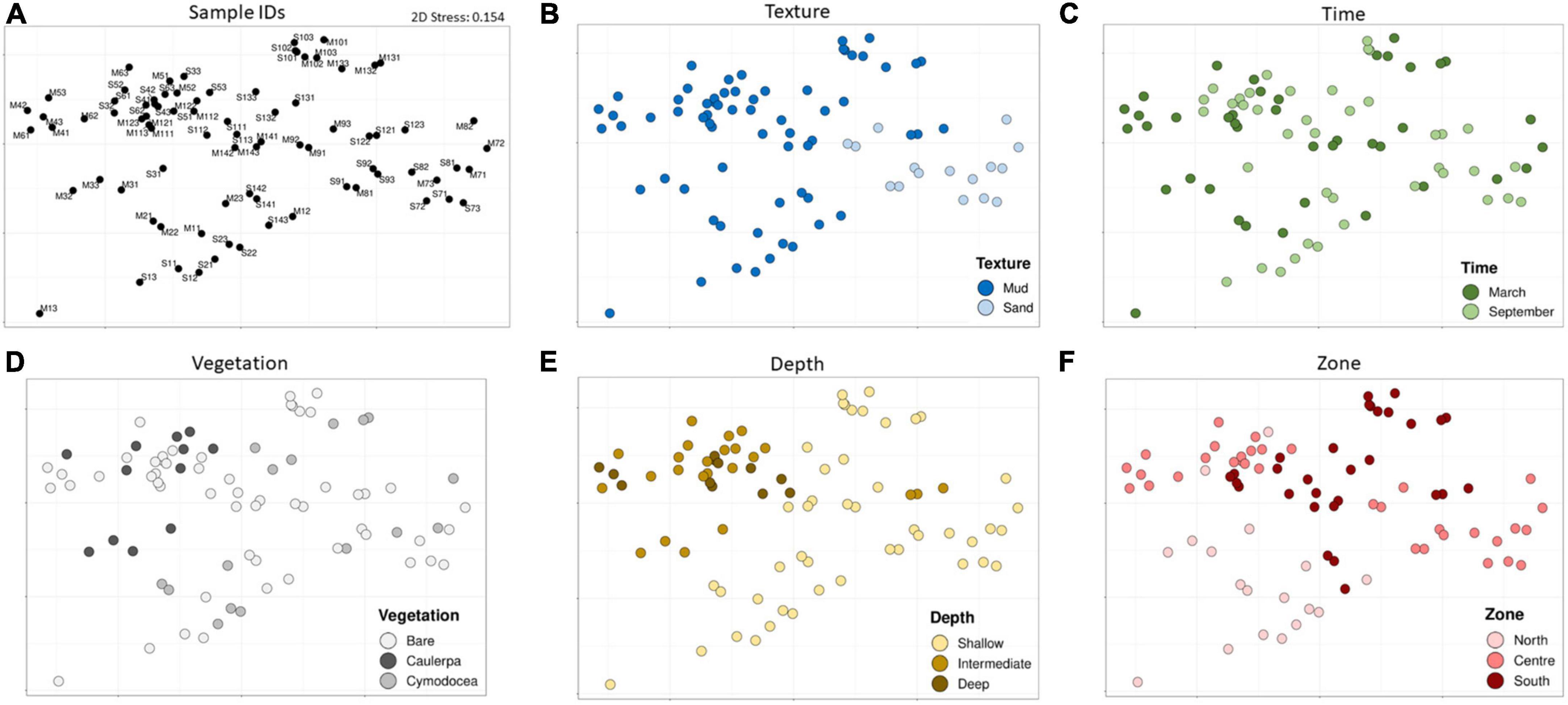
Figure 5. An NMDS plot of the Bray–Curtis distances among all samples (A), colored by factor texture (B), time (C), vegetation (D), depth (E), and zone (F).
Texture is a well-known factor influencing the microbial community composition (Llobet-Brossa et al., 1998; Sun et al., 2013) and variables such as Eh, AVS, TOC, AsAVS, and FeA, which were statistically significant between levels of this factor (Supplementary Figure 3), could be the responsible of this differences. Although the influence of texture was expected, information about the relationship between sediment texture and microbial community structure still remains scarce in the literature (Boey et al., 2022). Thus, to provide some light, a detailed analysis of the taxonomic differences was performed. At the class level, a significant higher relative abundance of Anaerolineae was observed in muddy sediments while sequences related to Bacteroidia, Alphaproteobacteria, and Acidimicrobiia were significantly more abundant in sandy ones (Figure 6A). The most relevant OTUs highlighting the differences between muddy and sandy sediments were identified by SIMPER (Supplementary Table 1.5). Since microorganisms within the class Anaerolineae are strict anaerobes (Yamada and Sekiguchi, 2015), their relatively higher abundances in muddy sediments with a low redox potential were not surprising. On the other hand, sandy sediments showed a more positive redox potential and a lower concentration of total organic carbon. Accordingly, they harbored higher abundances of putatively aerobic microorganisms, such as the genera Robiginitalea and Ulvibacter (from the Bacteroidia class) (Cho and Giovannoni, 2004; Choi et al., 2007; Kim et al., 2008), and members of Alphaproteobacteria and Acidimicrobiia, which have been detected in aerobic and oligotrophic environments (Oh et al., 2012, 2016; Mizuno et al., 2015). In summary, these results suggested that sandy sediments had higher abundances of putatively aerobic microorganisms whereas muddy sediments had higher abundances of putatively anaerobic microbes.
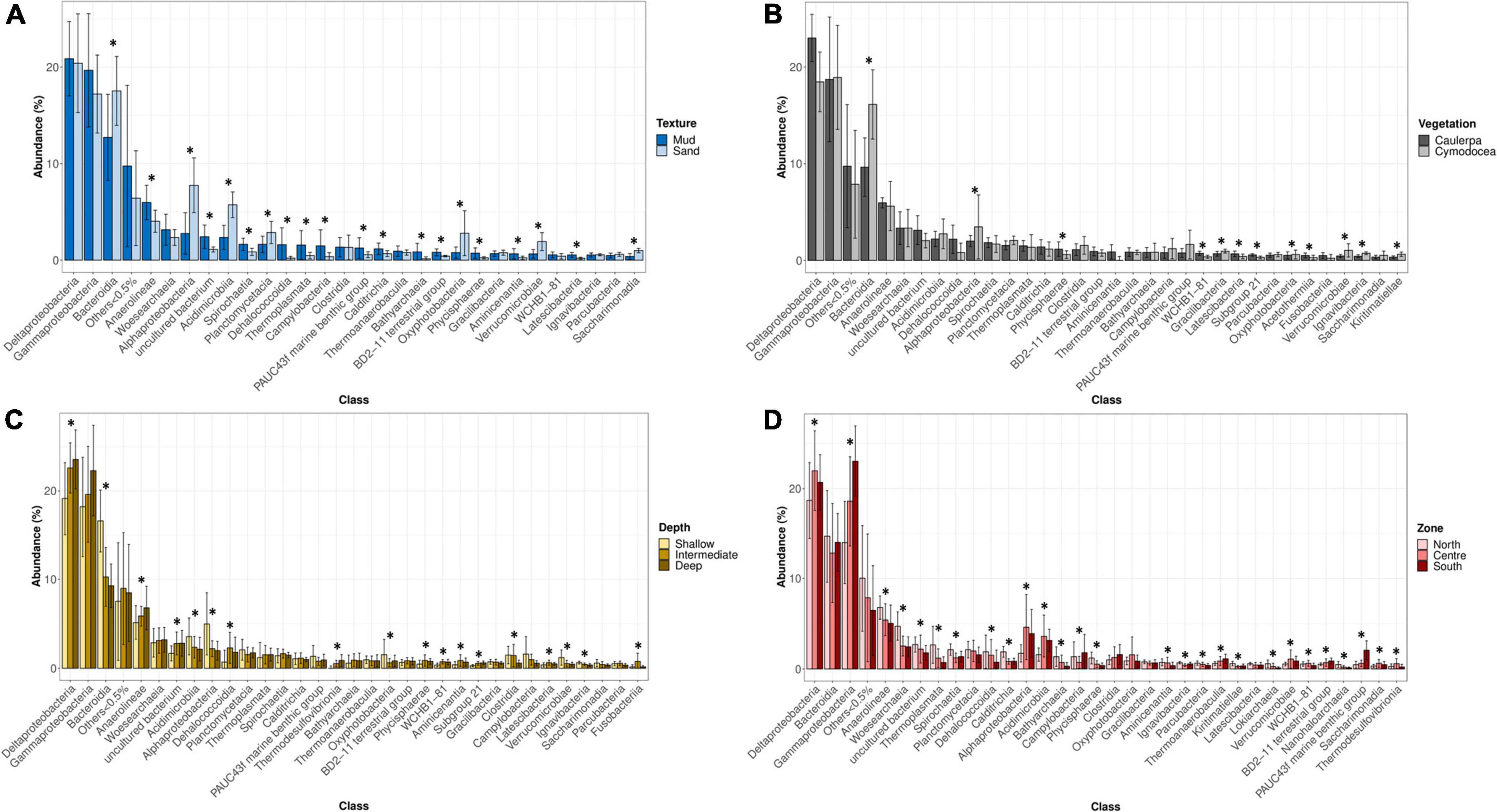
Figure 6. Relative abundance of detected microbial classes based on the levels of factor texture (A), vegetation (B), depth (C), and zone (D). Error bars indicate the range of abundances within each group of samples. Mud (n = 66); Sand (n = 16); Cymodocea nodosa (n = 16); C. prolifera (n = 16); Shallow (n = 46); Intermediate (n = 24); Deep (n = 12); North (n = 18); Centre (n = 34); and South (n = 30). Asterisk (*) indicates statistically significant differences tested by ANOVA (p < 0.05).
Regarding the type of benthic vegetation (i.e., Caulerpa and/or Cymodocea), our result is in good agreement with previous comparisons between seagrasses and macroalgae (Gribben et al., 2017). Curiously, only SOC was statistically significant for this factor (Supplementary Figure 3). Given that Caulerpa was able to spread in the Mar Menor due to human activity, the taxonomic differences and their corresponding metabolic potentials are of great importance to determine the anthropogenic influence on the lagoon. At the class level, the proportion of sequences related to Deltaproteobacteria was higher in Caulerpa-associated sediments despite not being statistically significant, while Bacteroidia sequences had a statistically significant higher abundance in Cymodocea-associated sediments (Figure 6B). The OTUs contributing to the differences between Caulerpa and Cymodocea communities are shown in Supplementary Table 1.6. Our data agree with previous observations of higher abundances of sulfate-reducing microorganisms (Deltaproteobacteria) in Caulerpa sediments compared to phanerogam-colonized ones (Holmer et al., 2009; Aires et al., 2013; Gribben et al., 2017). Besides the difference in their relative proportions, different families of sulfate reducers were also present in both sediments (Desulfobulbaceae and Desulfobacteraceae in Cymodocea and Caulerpa-colonized sediments, respectively). Desulfobacteraceae are considered complete oxidizers while Desulfobulbaceae are unable to oxidize acetate to CO2 (Xing et al., 2020). On the other hand, different sulfur-oxidizing bacteria were also found associated with both types of vegetation, with B2M28, Thioalkalivibrio and Thiohalophilus dominating Cymodocea sediments and Woeseia and Arenicellaceae prevailing in Caulerpa sediments. The class Bacteroidia, which was relatively more abundant in Cymodocea colonized sediments, has been found to be abundant in other seagrass species such as Zostera marina, Z. noltii, Thalassia testudinum, and Syringodium filliforme, with some phylotypes belonging to their core microbiome (Cúcio et al., 2016; Ugarelli et al., 2019). Furthermore, a study reported an enrichment in Bacteroidales inside a patch of the seagrass Zostera compared to the surrounding sediment (Ettinger et al., 2017). These observations suggest that some kind of stable association, direct or indirect, could exist between Bacteroidia and seagrasses, and is probably related to the higher content of labile organic matter inside seagrass patches compared to bare sediments (Honkoop et al., 2008; Ricart et al., 2015).
On the other hand, differences were also observed between the shallowest sediments (0–2 m) and those deeper than 2 m. This finding suggests that physicochemical conditions change at 2-m depth, shifting microbial communities, although only DOP, which was higher in deep sediments, presented statistically significant differences (Supplementary Figure 3). Intriguingly, a 2-m depth was also the limit for the survival of vegetation due to anoxia and light scarcity during the 2016 algal bloom (Belando et al., 2017). Regarding the taxonomic differences (Figure 6C), Deltaproteobacteria and Anaerolineae, both well-known anaerobic bacteria were significantly more abundant in intermediate/deep sediments while Bacteroidia, Acidimicrobiia, and Alphaproteobacteria were in shallow samples. The SIMPER analysis revealed the OTUs responsible for the differences between shallow and intermediate/deep samples (Supplementary Table 1.7). Remarkably, Desulfobacteraceae were enriched in intermediate/deep samples while Robiginitalea and Actinomarinales were in shallow samples. These differences are similar to those obtained with texture and based on the putative metabolic potential of these groups are probably due to oxygen availability.
Finally, we explored whether the difference between the north and the south microbial communities could be related to the extremely high concentrations of PTEs found in the southern basin (Supplementary Figure 3).
Relationships between potentially toxic elements concentrations and microbial communities
As explained above, clear differences in the microbial communities were observed between zones, mainly among the north and south microbial communities (Figure 5E), most likely due to the extremely high concentrations of PTEs present in the southern basin attributable to the presence of mining wastes carried from the old mining area (Marín-Guirao et al., 2008; María-Cervantes et al., 2009) (Supplementary Figure 3). Remarkably, when the temporal changes in the microbial communities of each station were tested by PERMANOVA (two factors: station and time) significant p-values were obtained (p = 0.0002), and a posteriori test indicated that the stations with the highest PTEs concentrations presented significant temporal changes while the less contaminated stations did not (Table 1). These differences were mainly due to changes in low abundance bacterial classes but some relatively abundant ones (up to 6.2% relative abundance) also displayed extensive changes. These differences between the most contaminated (P4, P10, P11, P12, P13, and P14) and the less contaminated stations (which we will refer to as contaminated and non-contaminated, respectively) can be explained by the following three hypotheses: (i) Contaminated and non-contaminated stations were exposed to different temporal environmental changes, (ii) there was an input of PTEs in the contaminated stations between March and September, and (iii) microbial communities inhabiting the sediments with high PTEs concentrations are more vulnerable to environmental changes. The PERMANOVA test of the environmental variables with two factors (time: March vs. September; PTEs: contaminated vs. non-contaminated), indicated that there were significant differences in the whole lagoon between March and September and between the contaminated and non-contaminated stations but not in the interaction between the two factors (Supplementary Table 1.8) which means that there was no evidence that temporal changes among the two groups of stations were different and thus, “hypothesis i” was rejected. Then, the two-way (time and PTEs) ANOVAs (Supplementary Table 1.9) identified redox potential and S2– as the variables responsible for temporal changes, and AVS and PTEs, for the differences between contaminated and non-contaminated stations. Thus, there was no evidence of input of PTEs between March and September and therefore “hypothesis ii” was also rejected. The only environmental variables that differed between contaminated and non-contaminated stations were those associated with PTEs. Therefore, based on our results, the most likely “hypothesis iii” is that the microbial communities that inhabit the sediments with high PTEs concentrations are more vulnerable to environmental changes. Further, in vitro experiments would be necessary to confirm the hypothesis.
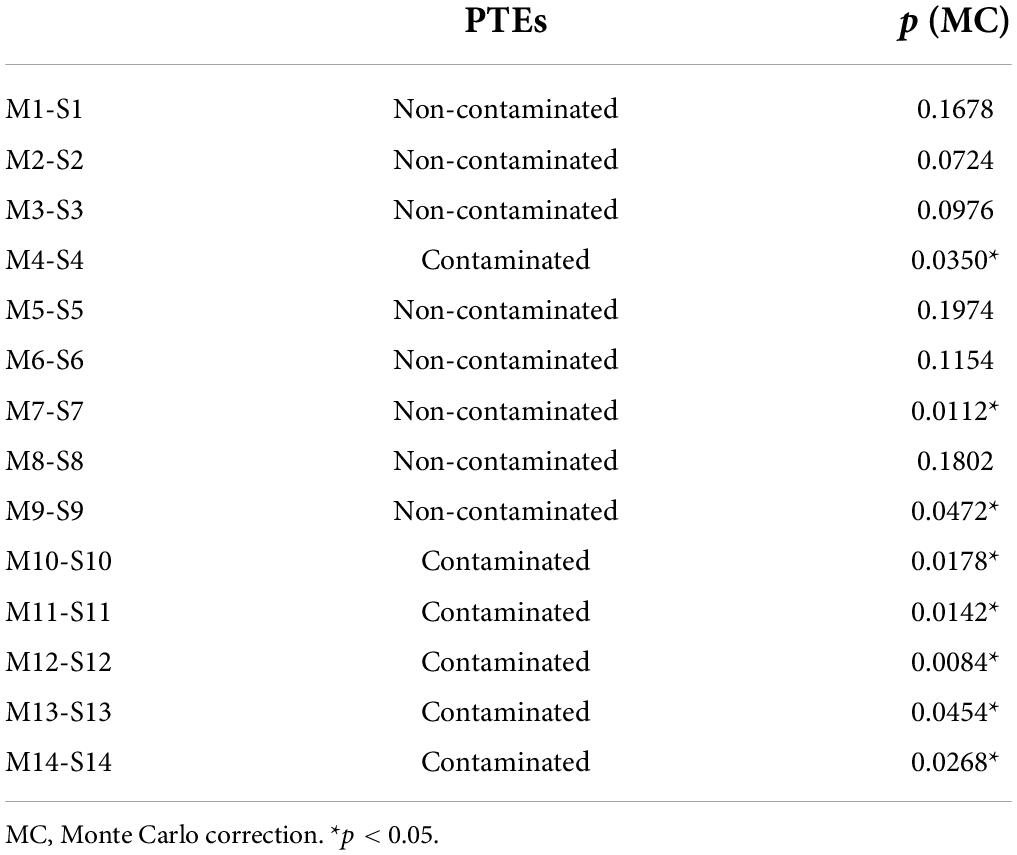
Table 1. The p-values of the PERMANOVA a posteriori test for the microbial communities of each station between March and September.
Microbes able to survive in the presence of high concentrations of PTEs (heavy metals and As), which can be considered an extreme condition, are known as metallophiles (Nies, 2000; Gupta et al., 2014). SIMPER analyses supported our hypothesis by revealing that temporal changes in PTE-contaminated samples were mainly due to metallophilic OTUs (defined as those OTUs statistically different or contributing to changes among contaminated and non-contaminated stations and whose mean relative abundance in contaminated stations was two-fold higher than in non-contaminated stations) (19.64–95.15%) (Table 2). Metallophilic OTUs could correspond to specialist bacteria harboring mechanisms to resist or use the PTEs which could be more susceptible to environmental perturbations. Indeed, it has been previously reported that such specialist microorganisms can be more vulnerable to fluctuations in the other environmental variables since the genetic adaptation to one environment may result in loss of fitness in another environment (Cooper and Lenski, 2000; Kassen, 2002; Zhong et al., 2004). Nevertheless, non-metallophilic OTUs also contributed to temporal changes but in a lower proportion (5.44–22.07%).
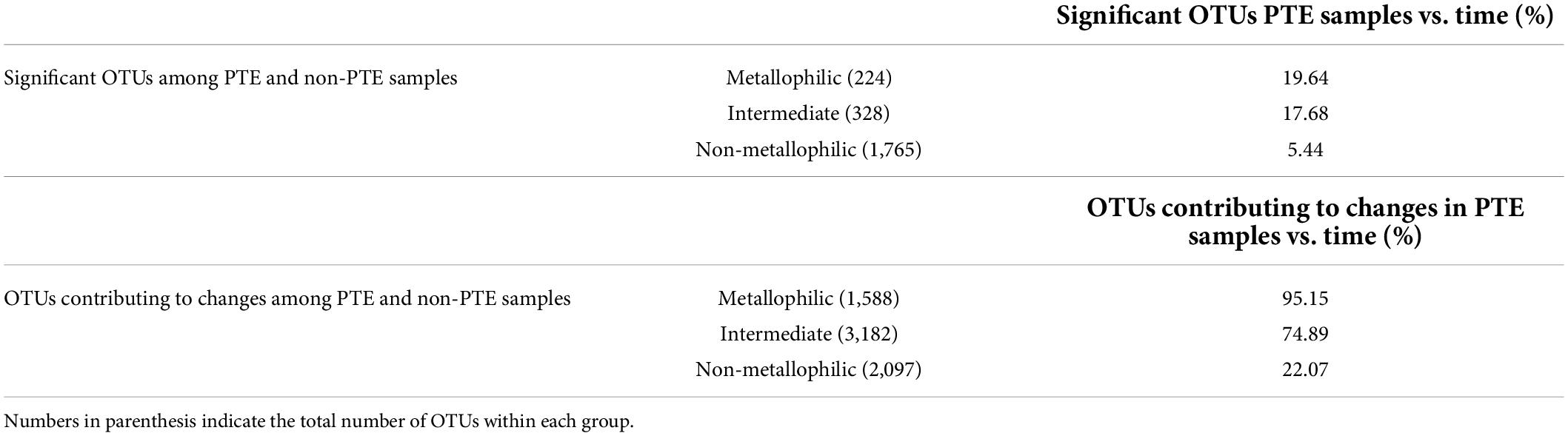
Table 2. Percentage of OTUs classified as metallophilic (fold change > 2), intermediate (0.5 < fold change < 2), and non-metallophilic (fold change < 0.5) that were also statistically different in the PTE contaminated samples between March and September.
Some of the extreme metallophilic OTUs found in the Mar Menor sediments have been previously associated with metal-rich environments. Metallophiles were identified by examining the Upset diagrams, which showed that P10, the most contaminated station, harbored a set of 15 specific OTUs that were present between March and September with abundances higher than 0.1% (Figure 3B). These OTUs were assigned to Sva1033, Woeseiaceae, Anaerolineaceae, OPB41, Desulfobulbaceae, Desulfuromusa, Flavobacteriaceae, and SAR324 families. Sva1033 metabolism has been associated with metal reduction (Sun et al., 2013; Buongiorno et al., 2019). Woeseia was the most abundant genus in a copper biofilm (Zhang et al., 2019) and metagenomic data suggested a broad range of metabolism for Woeseiaceae, including sulfur-based chemolithoautotrophy (Mußmann et al., 2017). OPB41 refers to a 16S rRNA gene amplicon sequence detected for the first time in a Yellowstone hot spring, an environment with high concentrations of reduced iron and sulfur (Hugenholtz et al., 1998). Desulfobulbaceae and SAR324 have been reported to negatively correlate with toxic elements (Baltar et al., 2018; Wu et al., 2019); however, the OTUs detected herein could be new representatives of these groups since their identities to sequences in the ARB-SILVA database were rather low (95.71 and 95.92%, respectively). Flavobacteriaceae was present in toxic metal-contaminated marine sediment (Gillan et al., 2005; Toes et al., 2008) and species of the genus Desulfuromusa are well-known Fe-reducing bacteria (Vandieken et al., 2006).
Remarkably, two out of the nine core OTUs (those shared between all stations and sampling times) with abundances >0.1% (NR_OTU1827 and AF286033.1.1521) were also defined as extreme metallophiles and belonged to the classes Gammaproteobacteria and PAUC43f (Gemmatimonadota phylum), respectively. As shown in Figure 6D, there were statistically significant higher abundances of Gammaprotebacteria and PAUC43f in the south of the lagoon, the most contaminated area. Sequences similar to this Gammaproteobacteria OTU have been detected in heavy metal-rich environments (Ben Said et al., 2010; Meyer-Dombard et al., 2012), a microbial mat at a shallow submarine hot spring (Hirayama et al., 2007), and marine sediments (Schöttner et al., 2011), as well as in the guts of fishes (NCBI accession number KJ197761). Regarding the PAUC43f 16S rRNA gene sequence (NCBI accession number AF286033), it was first detected in a saltmarsh contaminated with mercury and polychlorinated-biphenyls (PCBs). Closely related sequences were also detected in different types of sediments (Bowman and McCuaig, 2003; López-García et al., 2003; Genderjahn et al., 2018) and as a symbiont of marine invertebrates (Radwan et al., 2010; Griffiths et al., 2019; Bergo et al., 2021). Although it was deposited in the NCBI database more than 20 years ago and it has been found even in different ecosystems, there is no available information about the metabolism of this class. The high abundance and widespread presence of PAUC43f in the Mar Menor sediments make it an excellent scenario to study the characteristics of this uncultured class.
Conclusion
The Mar Menor lagoon is subjected to natural and intense anthropic pressures, which have left their imprints on the microbial community inhabiting the sediments. Here, we have shown that texture, type of vegetation, depth, and zone are strong structuring factors while the effect of time (season) is more limited and restricted to PTE contaminated stations. Our results provide a detailed description of the differences in microbial composition associated with these factors which are of great importance for the health of the lagoon. For instance, Caulerpa colonized sediments harbor a higher potential for sulfate reduction and thus a lower redox potential (as we have observed in aquaria experiments, data not shown). This human-introduced factor in the Mar Menor can be crucial in determining the response of the sediment microbiota to changes in the water column, which may favor the proliferation of sulfide-producing microbes that negatively affect the system. However, the sediment microbial community also harbors the potential for detoxification as well as microbes that are interesting both because of their novelty and resistance to PTEs. The history of mining impacts has most likely influenced the microbial communities, which have developed ways to tolerate, and even thrive, in this highly polluted surrounding. However, this specialization may have also increased the vulnerability of these communities to other environmental changes which add more instability to the whole ecosystem.
The sediment microbiota could be a key player in the fate of the Mar Menor since they could buffer the system from, and respond to, environmental changes. Here, we paved the way to understanding how this may occur.
Data availability statement
16S rRNA gene sequences generated in this work have been deposited in the NCBI Sequence Read Archive (SRA) database under BioProject PRJNA753791.
Author contributions
BA-R: conceptualization, data curation, formal analysis, investigation, methodology, validation, visualization, writing—original draft, and writing—review and editing. ER-P: data curation, formal analysis, investigation, methodology, supervision, validation, visualization, writing—original draft, and writing—review and editing. JÁ-R: conceptualization and writing—review and editing. FG-C: conceptualization, methodology, visualization, and writing—review and editing. XO: conceptualization, data curation, formal analysis, investigation, methodology, supervision, validation, visualization, writing—original draft, and writing—review and editing. M-DB: data curation and writing—review and editing. JB-E: conceptualization, data curation, funding acquisition, methodology, project administration, resources, and writing—review and editing. RG-M: data curation, project administration, and writing—review and editing. AF: data curation, formal analysis, methodology, supervision, writing—original draft, and writing—review and editing. JR: conceptualization, funding acquisition, methodology, project administration, resources, and writing—review and editing. FS: conceptualization, investigation, supervision, validation, visualization, writing—original draft, and writing—review and editing. JA: conceptualization, funding acquisition, investigation, project administration, supervision, validation, visualization, writing—original draft, and writing—review and editing. All authors contributed to the article and approved the submitted version.
Funding
The study of the microbial communities has been carried out within the project “Metafluidics” and it was funded by the European Union’s framework program Horizon 2020 (LEIT-BIO-2015-685474 to JA). Samplings were supported by the projects MEMM (financed by the Spanish Institute of Oceanography), 19-ESMARES2-ANG (financed by the Spanish Ministry of Ecological Transition and Demographic Challenge), and BIOFOM (Ref. TEC0004869 financed by the Regional Government of Murcia). M-DB and JB-E were supported by a contract within the Program Personal Técnico de Apoyo funded by the Ministerio de Economía y Competitividad. BA-R was also supported by the ACIF fellow of the Generalitat Valenciana government.
Acknowledgments
We thank Cristina López–Pascual for her help in the initial processing of the samples and Heather Maughan for her professional English editing and the critical reading of the manuscript. We thank Carles Pedrós–Alió for his critical reading of the manuscript and helpful suggestions. Sediment sampling and geochemical characterization was carried out by Universidad Politécnica de Cartagena, Universidad de Santiago de Compostela, and Centro Oceanográfico de Murcia (IEO) throughout a contract with TRAGSA, supported by Comunidad Autónoma de Murcia and FEDER funds. Thanks to M. C. Tercero, H. M. Conesa, A. Peñalver, I. Tunç, B. Straetemans, J. Santos, J. Patricio, J. Roberto, V. León, F. Torres, M. Santiso, and P. Guevara for helping in sample processing. We acknowledge logistical support provided by the President and Staff of the harbors Club Nautico Lo Pagán, Club Náutico La Puntica, Centro de Actividades Náuticas (San Pedro del Pinatar, Murcia, Spain), and Club Náutico Los Urrutias. We also thank the Regional Government of Murcia and national authorities (MITECO) for permission.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2022.937683/full#supplementary-material
References
Aguilar-Escribano, J., Mas-Hernández, J., and Ramos-Esplá, A. (2016). Evaluación del estado y composición de la comunidad fitoplanctónica de las agua del mar menor, Murcia (mayo de 2016). Alicante: University of Alicante.
Aires, T., Serrão, E. A., Kendrick, G., Duarte, C. M., and Arnaud-Haond, S. (2013). Invasion is a community affair: Clandestine followers in the bacterial community associated to green algae, Caulerpa racemosa, track the invasion source. PLoS One 8:e68429. doi: 10.1371/journal.pone.0068429
Alongi, D. M. (1994). The role of bacteria in nutrient recycling in tropical mangrove and other coastal benthic ecosystems. Hydrobiologia 285, 19–32. doi: 10.1007/BF00005650
Álvarez-Rogel, J., Santos, J., Belando, M. D., and Bernardeau, J. (2019). Proyecto BIOFOM. Estudio biogeoquímico de los fondos del mar menor- caracterización biológica de sedimentos (Ref. TEC0004869). Encomienda de Gestión de la CARM a través de Tecnologías y Servicios Agrarios, S.A., S.M.E., M.P. (TRAGSATEC).
Anderson, M. J. (2001). A new method for non-parametric multivariate analysis of variance. Austral Ecol. 26, 32–46. doi: 10.1046/j.1442-9993.2001.01070.x
Aravio-Torre, J., and Arévalo, A. (1973). La salinidad del Mar Menor, sus variaciones. Algunas consideraciones sobre el intercambio de aguas con el Mediterráneo. Boll. Inst. Esp. Ocean 146, 1–13.
Ballester, R. (1985). Biomasa, estacionalidad y distribución de tres macrófitos: Ruppia cirrhosa, Cymodocea nodosa y Caulerpa prolifera en el Mar Menor (Murcia, SE de España). An. Biol. 4, 31–36.
Baltar, F., Gutiérrez-Rodríguez, A., Meyer, M., Skudelny, I., Sander, S., Thomson, B., et al. (2018). Specific effect of trace metals on marine heterotrophic microbial activity and diversity: Key role of iron and zinc and hydrocarbon-degrading bacteria. Front. Microbiol. 9:3190. doi: 10.3389/fmicb.2018.03190
Barbier, E., Hacker, S., Kennedy, C., Koch, E., Stier, A., and Silliman, B. (2011). The value of estuarine and coastal ecosystem services. Ecol. Monogr. 81, 169–193.
Belando, M. D., Bernardeau-Esteller, J., García-Muñoz, R., Ramos-Segura, A., Santos-Echeandía, J., García-Moreno, P., et al. (2017). Evaluación del estado de conservación de las praderas de Cymodocea nodosa en la laguna costera del Mar Menor. 2014-2016. Murcia: Instituto Español de Oceanografía y Asociación de Naturalistas del Sureste.
Belando, M. D. Bernardeau-Esteller, J., and Paradinas, I. Ramos-Segura, A. García-Muñoz, R. García-Moreno, et al. (2021). Long-term coexistence between the macroalga Caulerpa prolifera and the seagrass Cymodocea nodosa in a Mediterranean lagoon. Aquat. Bot. 173:103415. doi: 10.1016/j.aquabot.2021.103415
Belando-Torrente, M. D., Ruiz, J. M., Muñoz, R., Segura, A., Esteller, J., Casero, J., et al. (2019). Collapse of macrophytic communities in a eutrophicated coastal lagoon. Front. Mar. Sci. 6:192. doi: 10.3389/conf.fmars.2019.08.00192
Ben Said, O., Goñi-Urriza, M., El Bour, M., Aissa, P., and Duran, R. (2010). Bacterial community structure of sediments of the bizerte lagoon (Tunisia), a southern mediterranean coastal anthropized lagoon. Microb. Ecol. 59, 445–456. doi: 10.1007/s00248-009-9585-x
Bergo, N. M., Bendia, A. G., Ferreira, J. C. N., Murton, B. J., Brandini, F. P., and Pellizari, V. H. (2021). Microbial diversity of deep-sea ferromanganese crust field in the Rio Grande rise, southwestern Atlantic ocean. Microb. Ecol. 82, 344–355. doi: 10.1007/s00248-020-01670-y
Bernardet, J.-F. (2015). “Flavobacteriales ord. nov,” in Bergey’s manual of systematics of archaea and bacteria, eds Trujillo, M. E., Dedysh, S., DeVos, P., Hedlund, B., P. Kämpfer, Rainey, F. A., et al. (New York, NY: Springer), 1–2. doi: 10.1002/9781118960608.obm00033
Boehm, A. B., Yamahara, K. M., and Sassoubre, L. M. (2014). Diversity and transport of microorganisms in intertidal sands of the california coast. Appl. Environ. Microbiol. 80, 3943–3951. doi: 10.1128/AEM.00513-14
Boey, J. S., Mortimer, R., Couturier, A., Worrallo, K., and Handley, K. M. (2022). Estuarine microbial diversity and nitrogen cycling increase along sand–mud gradients independent of salinity and distance. Environ. Microbiol. 24, 50–65. doi: 10.1111/1462-2920.15550
Bowman, J. P., and McCuaig, R. D. (2003). Biodiversity, community structural shifts, and biogeography of prokaryotes within Antarctic continental shelf sediment. Appl. Environ. Microbiol. 69, 2463–2483. doi: 10.1128/AEM.69.5.2463-2483.2003
Brinkhoff, T., Fischer, D., Vollmers, J., Voget, S., Beardsley, C., Thole, S., et al. (2012). Biogeography and phylogenetic diversity of a cluster of exclusively marine myxobacteria. ISME J. 6, 1260–1272. doi: 10.1038/ismej.2011.190
Bulzu, P. A., Andrei, A. Ş., Salcher, M. M., Mehrshad, M., Inoue, K., Kandori, H., et al. (2019). Casting light on Asgardarchaeota metabolism in a sunlit microoxic niche. Nat. Microbiol. 4, 1129–1137. doi: 10.1038/s41564-019-0404-y
Buongiorno, J., Herbert, L. C., Wehrmann, L. M., Michaud, A. B., Laufer, K., Røy, H., et al. (2019). Complex microbial communities drive iron and sulfur cycling in Arctic fjord sediments. Appl. Environ. Microbiol. 85:e00949-19. doi: 10.1128/AEM.00949-19
Caporaso, J. G., Kuczynski, J., Stombaugh, J., Bittinger, K., Bushman, F. D., Costello, E. K., et al. (2010). QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335–336. doi: 10.1038/nmeth.f.303
Cho, J. C., and Giovannoni, S. J. (2004). Robiginitalea biformata gen. nov., sp. nov., a novel marine bacterium in the family Flavobacteriaceae with a higher G + C content. Int. J. Syst. Evol. Microbiol. 54, 1101–1106. doi: 10.1099/ijs.0.03023-0
Choi, A., Yang, S. J., and Cho, J. C. (2013). Lutibacter flavus sp. nov., a marine bacterium isolated from a tidal flat sediment. Int. J. Syst. Evol. Microbiol. 63, 946–951. doi: 10.1099/ijs.0.043471-0
Choi, T. H., Lee, H. K., Lee, K., and Cho, J. C. (2007). Ulvibacter antarcticus sp. nov., isolated from Antarctic coastal seawater. Int. J. Syst. Evol. Microbiol. 64, 2041–2046. doi: 10.1099/ijs.0.65265-0
Clarke, K. R. (1993). Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 18, 117–143.
Cline, J. D. (1969). Spectrophotometric determination of hydrogen sulfide in natural waters. Limnol. Oceanogr. 14, 454–458. doi: 10.4319/lo.1969.14.3.0454
Conesa, H. M., and Jiménez-Cárceles, F. J. (2007). The Mar Menor lagoon (SE Spain): A singular natural ecosystem threatened by human activities. Mar. Pollut. Bull. 54, 839–849. doi: 10.1016/j.marpolbul.2007.05.007
Conesa, H. M., and Schulin, R. (2010). The Cartagena-La Unión mining district (SE spain): A review of environmental problems and emerging phytoremediation solutions after fifteen years research. J. Environ. Monit. 12, 1225–1233. doi: 10.1039/c000346h
Conway, J. R., Lex, A., and Gehlenborg, N. (2017). UpSetR: An R package for the visualization of intersecting sets and their properties. Bioinformatics 33, 2938–2940. doi: 10.1093/bioinformatics/btx364
Cooper, V. S., and Lenski, R. E. (2000). The population genetics of ecological specialization in evolving Escherichia coli populations. Nature 407, 736–739. doi: 10.1038/35037572
Crosby, L. D., and Criddle, C. S. (2003). Understanding bias in microbial community analysis techniques due to rrn operon copy number heterogeneity. Biotechniques 34, 790–802. doi: 10.2144/03344rr01
Cúcio, C., Engelen, A. H., Costa, R., and Muyzer, G. (2016). Rhizosphere microbiomes of European seagrasses are selected by the plant, but are not species specific. Front. Microbiol. 7:440. doi: 10.3389/fmicb.2016.00440
De Wit, R., Stal, L. J., Lomstein, B. A., Herbert, R. A., Van Gemerden, H., Viaroli, P., et al. (2001). Robust: The role of buffering capacities in stabilising coastal lagoon ecosystems. Cont. Shelf Res. 21, 2021–2041. doi: 10.1016/S0278-4343(01)00040-1
DeLong, E. F., Taylor, L. T., Marsh, T. L., and Preston, C. M. (1999). Visualization and enumeration of marine planktonic archaea and bacteria by using polyribonucleotide probes and fluorescent in situ hybridization. Appl. Environ. Microbiol. 65, 5554–5563. doi: 10.1128/aem.65.12.5554-5563.1999
Eilers, H., Pernthaler, J., Glöckner, F. O., and Amann, R. (2000). Culturability and in situ abundance of pelagic Bacteria from the North Sea. Appl. Environ. Microbiol. 66, 3044–3051. doi: 10.1128/AEM.66.7.3044-3051.2000
Ettinger, C. L., Voerman, S. E., Lang, J. M., Stachowicz, J. J., and Eisen, J. A. (2017). Microbial communities in sediment from Zostera marina patches, but not the Z. marina leaf or root microbiomes, vary in relation to distance from patch edge. PeerJ 5:e3246. doi: 10.7717/peerj.3246
Fernández-Gómez, B., Richter, M., Schüler, M., Pinhassi, J., Acinas, S. G., González, J. M., et al. (2013). Ecology of marine bacteroidetes: A comparative genomics approach. ISME J. 7, 1026–1037. doi: 10.1038/ismej.2012.169
Fike, D. A., Bradley, A. S., and Rose, C. V. (2015). Rethinking the ancient sulfur cycle. Annu. Rev. Earth Planet. Sci. 43, 593–622. doi: 10.1146/annurev-earth-060313-054802
Fortin, D., Leppard, G. G., and Tessier, A. (1993). Characteristics of lacustrine diagenetic iron oxyhydroxides. Geochim. Cosmochim. Acta 57, 4391–4404. doi: 10.1016/0016-7037(93)90490-N
Fraile-Nuez, E., Machín, F., Santana-Casiano, J. M., González-Dávila, M., Domínguez-Yanes, J. F., Mercado, J., et al. (2018). Estudio oceanográfico integral de alta resolución de la laguna costera de El Mar Menor. Murcia: Instituto Español de Oceanografía y Asociación de Naturalistas del Sureste.
García, G., and Muñoz-Vera, A. (2015). Characterization and evolution of the sediments of a Mediterranean coastal lagoon located next to a former mining area. Mar. Pollut. Bull. 100, 249–263. doi: 10.1016/j.marpolbul.2015.08.042
García-Martínez, M., López-López, A., Calleja, M. L., Marbà, N., and Duarte, C. M. (2009). Bacterial community dynamics in a seagrass (Posidonia oceanica) meadow sediment. Estuaries Coast. 32, 276–286. doi: 10.1007/s12237-008-9115-y
Genderjahn, S., Alawi, M., Mangelsdorf, K., Horn, F., and Wagner, D. (2018). Desiccation-and saline-tolerant bacteria and archaea in kalahari pan sediments. Front. Microbiol. 9:2082. doi: 10.3389/fmicb.2018.02082
Ghai, R., Hernandez, C. M., Picazo, A., Mizuno, C. M., Ininbergs, K., Díez, B., et al. (2012). Metagenomes of mediterranean coastal lagoons. Sci. Rep. 2:490. doi: 10.1038/srep00490
Ghyselinck, J., Pfeiffer, S., Heylen, K., Sessitsch, A., and De Vos, P. (2013). The effect of primer choice and short read sequences on the outcome of 16S rRNA gene based diversity studies. PLoS One 8:e71360. doi: 10.1371/journal.pone.0071360
Gilabert, J. (2001). Seasonal plankton dynamics in a Mediterranean hypersaline coastal lagoon: The Mar Menor. J. Plankton Res. 23, 207–218. doi: 10.1093/plankt/23.2.207
Gillan, D. C., Danis, B., Pernet, P., Joly, G., and Dubois, P. (2005). Structure of sediment-associated microbial communities along a heavy-metal contamination gradient in the marine environment. Appl. Environ. Microbiol. 71, 679–690. doi: 10.1128/AEM.71.2.679-690.2005
Gomariz, M., Martínez-García, M., Santos, F., Rodríguez, F., Capella-Gutiérrez, S., Gabaldón, T., et al. (2015). From community approaches to single-cell genomics: The discovery of ubiquitous hyperhalophilic Bacteroidetes generalists. ISME J. 9, 16–31. doi: 10.1038/ismej.2014.95
Gribben, P. E., Nielsen, S., Seymour, J. R., Bradley, D. J., West, M. N., and Thomas, T. (2017). Microbial communities in marine sediments modify success of an invasive macrophyte. Sci. Rep. 7:9845. doi: 10.1038/s41598-017-10231-2
Griffiths, S. M., Antwis, R. E., Lenzi, L., Lucaci, A., Behringer, D. C., Butler, M. J., et al. (2019). Host genetics and geography influence microbiome composition in the sponge Ircinia campana. J. Anim. Ecol. 88, 1684–1695. doi: 10.1111/1365-2656.13065
Gupta, G. N., Srivastava, S., Khare, S. K., and Prakash, V. (2014). Extremophiles: An overview of microorganism from extreme environment. Int. J. Agric. Environ. Biotechnol. 7, 371–380. doi: 10.5958/2230-732x.2014.00258.7
Hewson, I., and Fuhrman, J. A. (2008). “Viruses, bacteria, and the microbial loop,” in Nitrogen in the Marine Environment, eds D. Capone, D. Bronk, M. Mulholland, and E. Carpenter (Amsterdam: Elsevier), 1097–1134. doi: 10.1016/B978-0-12-372522-6.00025-6
Hirayama, H., Sunamura, M., Takai, K., Nunoura, T., Noguchi, T., Oida, H., et al. (2007). Culture-dependent and -independent characterization of microbial communities associated with a shallow submarine hydrothermal system occurring within a coral reef off Taketomi Island, Japan. Appl. Environ. Microbiol. 73, 7642–7656. doi: 10.1128/AEM.01258-07
Holmer, M., Marbà, N., Lamote, M., and Duarte, C. M. (2009). Deterioration of sediment quality in seagrass meadows (Posidonia oceanica) invaded by macroalgae (Caulerpa sp.). Estuaries Coast. 32, 456–466. doi: 10.1007/s12237-009-9133-4
Honkoop, P. J. C., Berghuis, E. M., Holthuijsen, S., Lavaleye, M. S. S., and Piersma, T. (2008). Molluscan assemblages of seagrass-covered and bare intertidal flats on the Banc d’Arguin, Mauritania, in relation to characteristics of sediment and organic matter. J. Sea Res. 60, 255–263. doi: 10.1016/j.seares.2008.10.001
Hugenholtz, P., Pitulle, C., Hershberger, K. L., and Pace, N. R. (1998). Novel division level bacterial diversity in a Yellowstone hot spring. J. Bacteriol. 180, 366–376. doi: 10.1128/jb.180.2.366-376.1998
Jiménez-Cárceles, F. J., Egea, C., Rodríguez-Caparrós, A. B., Barbosa, O. A., Delgado, M. J., Ortiz, R., et al. (2006). Contents of nitrogen, ammonium, phosphorus, pesticides and heavy metals, in a salt marsh in the coast of the Mar Menor lagoon (SE Spain). Fresenius Environ. Bull. 15, 370–378.
Jiménez-Martinez, J., Garcia-Aróstegui, J. L., Hunink, J. E., Contreras, S., Baudron, P., and Candela, L. (2016). The role of groundwater in highly human-modified hydrosystems: A review of impacts and mitigation options in the Campo de Cartagena-Mar Menor coastal plain (SE Spain). Environ. Rev. 24, 377–392.
Kassen, R. (2002). The experimental evolution of specialists, generalists, and the maintenance of diversity. J. Evol. Biol. 15, 173–190. doi: 10.1046/j.1420-9101.2002.00377.x
Kembel, S. W., Wu, M., Eisen, J. A., and Green, J. L. (2012). Incorporating 16S gene copy number information improves estimates of microbial diversity and abundance. PLoS Comput. Biol. 8:e1002743. doi: 10.1371/journal.pcbi.1002743
Kim, J. H., Kim, K. Y., Hahm, Y. T., Kim, B. S., Chun, J., and Cha, C. J. (2008). Actibacter sediminis gen. nov., sp. nov., a marine bacterium of the family Flavobacteriaceae isolated from tidal flat sediment. Int. J. Syst. Evol. Microbiol. 58, 139–143. doi: 10.1099/ijs.0.65346-0
Kjerfve, B. (1994). Chapter 1 coastal lagoons. Elsevier Oceanogr. Ser. 60, 1–8. doi: 10.1016/S0422-9894(08)70006-0
Klindworth, A., Pruesse, E., Schweer, T., Peplies, J., Quast, C., Horn, M., et al. (2013). Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 41:e1. doi: 10.1093/nar/gks808
Kostka, J. E., and Luther, G. W. (1994). Partitioning and speciation of solid phase iron in saltmarsh sediments. Geochim. Cosmochim. Acta 58, 1701–1710. doi: 10.1016/0016-7037(94)90531-2
Krieg, N. R. (ed.). (2015). “Bacteroidales ord. nov,” in Bergey’s manual of systematics of archaea and bacteria, (Hoboken, NJ: John Wiley & Sons, Inc). doi: 10.1002/9781118960608.obm00028
León, V. M., García, I., González, E., Samper, R., Fernández-González, V., and Muniategui-Lorenzo, S. (2018). Potential transfer of organic pollutants from littoral plastics debris to the marine environment. Environ. Pollut. 236, 442–453. doi: 10.1016/j.envpol.2018.01.114
León, V. M., Moreno-González, R., García, V., and Campillo, J. A. (2017). Impact of flash flood events on the distribution of organic pollutants in surface sediments from a Mediterranean coastal lagoon (Mar Menor, SE Spain). Environ. Sci. Pollut. Res. 24, 4284–4300. doi: 10.1007/s11356-015-4628-y
León, V. M., Moreno-González, R., González, E., Martínez, F., García, V., and Campillo, J. A. (2013). Interspecific comparison of polycyclic aromatic hydrocarbons and persistent organochlorines bioaccumulation in bivalves from a Mediterranean coastal lagoon. Sci. Total Environ. 463–464, 975–987. doi: 10.1016/j.scitotenv.2013.06.075
Levene, H. (1960). Robust tests for equality of variances. Redwood City, CA: Stanford University Press.
Liu, X., Li, M., Castelle, C. J., Probst, A. J., Zhou, Z., Pan, J., et al. (2018). Insights into the ecology, evolution, and metabolism of the widespread woesearchaeotal lineages. Microbiome 6:102. doi: 10.1186/s40168-018-0488-2
Llobet-Brossa, E., Rabus, R., Böttcher, M. E., Könneke, M., Finke, N., Schramm, A., et al. (2002). Community structure and activity of sulfate-reducing bacteria in an intertidal surface sediment: A multi-method approach. Aquat. Microb. Ecol. 29, 211–226. doi: 10.3354/ame029211
Llobet-Brossa, E., Rosselló-Mora, R., and Amann, R. (1998). Microbial community composition of wadden sea sediments as revealed by fluorescence in situ hybridization. Appl. Environ. Microbiol. 64, 2691–2696. doi: 10.1128/aem.64.7.2691-2696.1998
López-García, P., Duperron, S., Philippot, P., Foriel, J., Susini, J., and Moreira, D. (2003). Bacterial diversity in hydrothermal sediment and epsilonproteobacterial dominance in experimental microcolonizers at the Mid-Atlantic Ridge. Environ. Microbiol. 5, 961–976. doi: 10.1046/j.1462-2920.2003.00495.x
Luna, G. M., Manini, E., and Danovaro, R. (2002). Large fraction of dead and inactive bacteria in coastal marine sediments: Comparison of protocols for determination and ecological significance. Appl. Environ. Microbiol. 68, 3509–3513. doi: 10.1128/AEM.68.7.3509-3513.2002
Madigan, M., Bender, K., Buckley, D., Sattley, W., and Stahl, D. (2019). Brock biology of microorganisms, 15th Edn. London: Pearson.
Maechler, M., Rousseeuw, P., Struyf, A., Hubert, M., Hornik, K., Studer, M., et al. (2019). Package “cluster” for R: ‘Finding Groups in Data’: Cluster Analysis Extended.
Magoč, T., and Salzberg, S. L. (2011). FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 27, 2957–2963. doi: 10.1093/bioinformatics/btr507
MAGRAMA (2008). Ministerio de agricultura, alimentación y medio ambiente. Available online at: https://www.miteco.gob.es/es/costas/temas/proteccion-costa/ecocartografias/ecocartografia-murcia.aspx
Mahmoudi, N., Robeson, M. S. II, Castro, H. F., Fortney, J. L., Techtmann, S. M., Joyner, D. C., et al. (2015). Microbial community composition and diversity in Caspian Sea sediments. FEMS Microbiol. Ecol. 91, 1–11. doi: 10.1093/femsec/fiu013
María-Cervantes, A., Jiménez-Cárceles, F. J., and Álvarez-Rogel, J. (2009). As, Cd, Cu, Mn, Pb, and Zn contents in sediments and mollusks (Hexaplex trunculus and Tapes decussatus) from coastal zones of a mediterranean lagoon (mar menor, SE Spain) affected by mining wastes. Water Air Soil Pollut. 200, 289–304. doi: 10.1007/s11270-008-9913-7
Marín-Guirao, L., Atucha, A. M., Barba, J. L., López, E. M., and García Fernández, A. J. (2005). Effects of mining wastes on a seagrass ecosystem: Metal accumulation and bioavailability, seagrass dynamics and associated community structure. Mar. Environ. Res. 60, 317–337. doi: 10.1016/j.marenvres.2004.11.002
Marín-Guirao, L., Bernardeau-Esteller, J., Belando, M. D., Cerezo, I., Pérez, E., Ramos, A., et al. (2019). Physiological tipping points to light reduction underlie seagrass population collapse and abrupt shift in a shallow coastal lagoon. Front. Mar. Sci. 6:181. doi: 10.3389/conf.fmars.2019.08.00181
Marín-Guirao, L., Lloret, J., and Marin, A. (2008). Carbon and nitrogen stable isotopes and metal concentration in food webs from a mining-impacted coastal lagoon. Sci. Total Environ. 393, 118–130. doi: 10.1016/j.scitotenv.2007.12.023
Mas, J. (1994). El Mar menor: Relaciones, diferencias y afinidades entre la laguna costera y el Mar Mediterráneo adyacente. Ph.D. Thesis. Madrid: Autonomous University of Madrid.
McArdle, B. H., and Anderson, M. J. (2001). Fitting multivariate models to community data: A comment on distance-based redundancy analysis. Ecology 82, 290–297.
McMurdie, P. J., and Holmes, S. (2013). Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One 8:e61217. doi: 10.1371/journal.pone.0061217
Mercado, J. M., Cortés, D., Gómez-Jakobsen, F., García-Gómez, C., Ouaissa, S., Yebra, L., et al. (2021). Role of small-sized phytoplankton in triggering an ecosystem disruptive algal bloom in a Mediterranean hypersaline coastal lagoon. Mar. Pollut. Bull. 164:111989. doi: 10.1016/j.marpolbul.2021.111989
Meyer-Dombard, D. R., Price, R. E., Pichler, T., and Amend, J. P. (2012). Prokaryotic populations in arsenic-rich shallow-sea hydrothermal sediments of Ambitle island, Papua New Guinea. Geomicrobiol. J. 29, 1–17. doi: 10.1080/01490451.2010.520073
Mizuno, C. M., Rodriguez-Valera, F., and Ghai, R. (2015). Genomes of planktonic acidimicrobiales: Widening horizons for marine actinobacteria by metagenomics. mBio 6:02083-14. doi: 10.1128/mBio.02083-14
Moreno-González, R., Rodríguez-Mozaz, S., Gros, M., Pérez-Cánovas, E., Barceló, D., and León, V. M. (2014). Input of pharmaceuticals through coastal surface watercourses into a Mediterranean lagoon (Mar Menor, SE Spain): Sources and seasonal variations. Sci. Total Environ. 490, 59–72. doi: 10.1016/j.scitotenv.2014.04.097
Moreno-González, R., Rodríguez-Mozaz, S., Huerta, B., Barceló, D., and León, V. M. (2016). Do pharmaceuticals bioaccumulate in marine molluscs and fish from a coastal lagoon? Environ. Res. 146, 282–298. doi: 10.1016/j.envres.2016.01.001
Mori, K., Suzuki, K. I., Yamaguchi, K., Urabe, T., and Hanada, S. (2015). Thiogranum longum gen. nov., sp. nov., an obligately chemolithoautotrophic, sulfur-oxidizing bacterium of the family Ectothiorhodospiraceae isolated from a deep-sea hydrothermal field, and an emended description of the genus Thiohalomonas. Int. J. Syst. Evol. Microbiol. 65, 235–241. doi: 10.1099/ijs.0.070599-0
Muñoz-Vera, A., García, G., and García-Sánchez, A. (2015). Metal bioaccumulation pattern by Cotylorhiza tuberculata (Cnidaria, Scyphozoa) in the Mar Menor coastal lagoon (SE Spain). Environ. Sci. Pollut. Res. 22, 19157–19169. doi: 10.1007/s11356-015-5119-x
Musat, N., Werner, U., Knittel, K., Kolb, S., Dodenhof, T., van Beusekom, J. E. E., et al. (2006). Microbial community structure of sandy intertidal sediments in the North Sea, Sylt-Rømø Basin, Wadden Sea. Syst. Appl. Microbiol. 29, 333–348. doi: 10.1016/j.syapm.2005.12.006
Mußmann, M., Pjevac, P., Krüger, K., and Dyksma, S. (2017). Genomic repertoire of the Woeseiaceae/JTB255, cosmopolitan and abundant core members of microbial communities in marine sediments. ISME J. 11, 1276–1281. doi: 10.1038/ismej.2016.185
Neiker. (2005). Determinación del contenido de amonio en aguas, lixiviados y extractos. (PEC/EN/A-059).
Newton, A., Brito, A. C., Icely, J. D., Derolez, V., Clara, I., Angus, S., et al. (2018). Assessing, quantifying and valuing the ecosystem services of coastal lagoons. J. Nat. Conserv. 44, 50–65. doi: 10.1016/J.JNC.2018.02.009
Nies, D. H. (2000). Heavy metal-resistant bacteria as extremophiles: Molecular physiology and biotechnological use of Ralstonia sp. CH34. Extremophiles 4, 77–82. doi: 10.1007/s007920050140
Oh, K. H., Choi, W. C., Jung, Y. T., Kang, S. J., Oh, T. K., and Yoon, J. H. (2012). Tropicimonas aquimaris sp. nov., isolated from seawater, and emended description of the genus Tropicimonas Harwati, et al. 2009. Int. J. Syst. Evol. Microbiol. 62, 688–692. doi: 10.1099/ijs.0.029371-0
Oh, M., Kim, J. H., and Kim, W. (2016). Tropicimonas arenosa sp. nov., isolated from marine sand. Int. J. Syst. Evol. Microbiol. 66, 5514–5518. doi: 10.1099/ijsem.0.001549
Oksanen, J. (2011). Multivariate analysis of ecological communities in R: Vegan tutorial. Available Online at: https://cran.r-project.org/web/packages/vegan/index.html (accessed June 27, 2019).
Park, S., Kang, S. J., Oh, T. K., and Yoon, J. H. (2010). Lutibacter maritimus sp. nov., isolated from a tidal flat sediment. Int. J. Syst. Evol. Microbiol. 60, 610–614. doi: 10.1099/ijs.0.012401-0
Pedrós-Alió, C. (2012). The rare bacterial biosphere. Ann. Rev. Mar. Sci. 4, 449–466. doi: 10.1146/annurev-marine-120710-100948
Pérez-Ruzafa, A., Campillo, S., Fernández-Palacios, J. M., García-Lacunza, A., García-Oliva, M., Ibañez, H., et al. (2019). Long-term dynamic in nutrients, chlorophyll a, and water quality parameters in a coastal lagoon during a process of eutrophication for decades, a sudden break and a relatively rapid recovery. Front. Mar. Sci. 6:26. doi: 10.3389/fmars.2019.00026
Pérez-Ruzafa, A., and Marcos, C. (2012). Fisheries in coastal lagoons: An assumed but poorly researched aspect of the ecology and functioning of coastal lagoons. Estuar. Coast. Shelf Sci. 110, 15–31. doi: 10.1016/j.ecss.2012.05.025
Pérez-Ruzafa, A., Quispe-Becerra, J. I., García-Charton, J. A., and Marcos, C. (2004). Composition, structure and distribution of the ichthyoplankton in a Mediterranean coastal lagoon. J. Fish Biol. 64, 202–218. doi: 10.1111/j.1095-8649.2004.00301.x
Petro, C., Zäncker, B., Starnawski, P., Jochum, L. M., Ferdelman, T. G., Jørgensen, B. B., et al. (2019). Marine deep biosphere microbial communities assemble in near-surface sediments in Aarhus Bay. Front. Microbiol. 10:758. doi: 10.3389/fmicb.2019.00758
Plugge, C. M., Zhang, W., Scholten, J. C. M., and Stams, A. J. M. (2011). Metabolic flexibility of sulfate-reducing bacteria. Front. Microbiol. 2:81. doi: 10.3389/fmicb.2011.00081
Quero, G. M., Cassin, D., Botter, M., Perini, L., and Luna, G. M. (2015). Patterns of benthic bacterial diversity in coastal areas contaminated by heavy metals, polycyclic aromatic hydrocarbons (PAHs) and polychlorinated biphenyls (PCBs). Front. Microbiol. 6:1053. doi: 10.3389/fmicb.2015.01053
Radwan, M., Hanora, A., Zan, J., Mohamed, N. M., Abo-Elmatty, D. M., Abou-El-Ela, S. H., et al. (2010). Bacterial community analyses of two red sea sponges. Mar. Biotechnol. 12, 350–360. doi: 10.1007/s10126-009-9239-5
Raiswell, R., Buckley, F., Berner, R. A., and Anderson, T. F. (1988). Degree of pyritization of iron as a paleoenvironmental indicator of bottom-water oxygenation. J. Sediment. Res. 58, 812–819. doi: 10.1306/212F8E72-2B24-11D7-8648000102C1865D
Raiswell, R., Canfield, D. E., and Berner, R. A. (1994). A comparasion of iron extraction methods of degree of pyritisation and the recognition of iron-limited formation. Chem. Geol. 111, 101–110. doi: 10.1016/0009-2541(94)90084-1
Reichenbach, H. (2015). “Myxococcales,” in Bergey’s manual of systematics of archaea and bacteria, eds W. B. Whitman, F. Rainey, P. Kämpfer, M. Trujillo, J. Chun, P. DeVos, B. Hedlund, and S. Dedysh (Hoboken, NJ: John Wiley & Sons, Inc), 1–31. doi: 10.1002/9781118960608.obm00088
Ricart, A. M., York, P. H., Rasheed, M. A., Pérez, M., Romero, J., Bryant, C. V., et al. (2015). Variability of sedimentary organic carbon in patchy seagrass landscapes. Mar. Pollut. Bull. 100, 476–482. doi: 10.1016/j.marpolbul.2015.09.032
Robador, A., Brüchert, V., and Jørgensen, B. B. (2009). The impact of temperature change on the activity and community composition of sulfate-reducing bacteria in arctic versus temperate marine sediments. Environ. Microbiol. 11, 1692–1703. doi: 10.1111/J.1462-2920.2009.01896.X
Romero, D., Barcala, E., María-Dolores, E., and Muñoz, P. (2020). European eels and heavy metals from the Mar Menor lagoon (SE Spain). Mar. Pollut. Bull. 158:111368. doi: 10.1016/j.marpolbul.2020.111368
Rubio-Portillo, E., Martin-Cuadrado, A.-B., Ramos-Esplá, A. Á., and Antón, J. (2021). Metagenomics unveils Posidonia oceanica “banquettes” as a potential source of novel bioactive compounds and carbohydrate active enzymes (CAZymes). mSystems 6:e00866-21. doi: 10.1128/msystems.00866-21
Rubio-Portillo, E., Santos, F., Martínez-García, M., de los Ríos, A., Ascaso, C., Souza-Egipsy, V., et al. (2016). Structure and temporal dynamics of the bacterial communities associated to microhabitats of the coral Oculina patagonica. Environ. Microbiol. 18, 4564–4578. doi: 10.1111/1462-2920.13548
Ruiz, J. M., Albentosa, M., Aldeguer, B., Álvarez-Rogel, J., Antón, J., Belando, M. D., et al. (2020). Informe de evolución y estado actual del Mar Menor en relación al proceso de eutrofización y sus causas. Informe de asesoramiento técnico del Instituto Español de Oceanografía.
Schmieder, R., and Edwards, R. (2011). Quality control and preprocessing of metagenomic datasets. Bioinformatics 27, 863–864. doi: 10.1093/bioinformatics/btr026
Schöttner, S., Pfitzner, B., Grünke, S., Rasheed, M., Wild, C., and Ramette, A. (2011). Drivers of bacterial diversity dynamics in permeable carbonate and silicate coral reef sands from the Red Sea. Environ. Microbiol. 13, 1815–1826. doi: 10.1111/j.1462-2920.2011.02494.x
Serrano, R., Gras, L., Giménez-Casalduero, F., Del-Pilar-Ruso, Y., Grindlay, G., and Mora, J. (2019). The role of Cymodocea nodosa on the dynamics of trace elements in different marine environmental compartments at the Mar Menor Lagoon (Spain). Mar. Pollut. Bull. 141, 52–60. doi: 10.1016/j.marpolbul.2019.02.019
Sharifuzzaman, S. M., Rahman, H., Ashekuzzaman, S. M., Islam, M. M., Chowdhury, S. R., and Hossain, M. S. (2016). “Heavy metals accumulation in coastal sediments,” in Environmental Remediation technologies for metal-contaminated soils, eds H. Hasegawa I, M. M. Rahman, and M. A. Rahman (Tokyo: Springer), 21–42. doi: 10.1007/978-4-431-55759-3_2
Simonneau, J. (1973). Mar menor: Evolution sedimentologique et geoquimique recente du remplissage. Toulouse: Université Paul Sebatier of Tolouse.
Sipos, R., Székely, A. J., Palatinszky, M., Révész, S., Márialigeti, K., and Nikolausz, M. (2007). Effect of primer mismatch, annealing temperature and PCR cycle number on 16S rRNA gene-targetting bacterial community analysis. FEMS Microbiol. Ecol. 60, 341–350. doi: 10.1111/j.1574-6941.2007.00283.x
Soergel, D. A. W., Dey, N., Knight, R., and Brenner, S. E. (2012). Selection of primers for optimal taxonomic classification of environmental 16S rRNA gene sequences. ISME J. 6, 1440–1444. doi: 10.1038/ismej.2011.208
Sørensen, J., Jørgensen, B. B., and Revsbech, N. P. (1979). A comparison of oxygen, nitrate, and sulfate respiration in coastal marine sediments. Microb. Ecol. 5, 105–115. doi: 10.1007/BF02010501
Sørensen, K. B., Glazer, B., Hannides, A., and Gaidos, E. (2007). Spatial structure of the microbial community in sandy carbonate sediment. Mar. Ecol. Prog. Ser. 346, 61–74. doi: 10.3354/meps06996
Sorokin, D. Y., Merkel, A. Y., and Muyzer, G. (2020). “Thiohalophilus,” in Bergey’s manual of systematics of archaea and bacteria, ed. W. B. Whitman (Hoboken, NJ: John Wiley & Sons), 1–7. doi: 10.1002/9781118960608.gbm01693
Straub, D., Blackwell, N., Langarica-Fuentes, A., Peltzer, A., Nahnsen, S., and Kleindienst, S. (2020). Interpretations of environmental microbial community studies are biased by the selected 16S rRNA (gene) amplicon sequencing pipeline. Front. Microbiol. 11:550420. doi: 10.3389/fmicb.2020.550420
Suárez-Suárez, A., López-López, A., Tovar-Sánchez, A., Yarza, P., Orfila, A., Terrados, J., et al. (2011). Response of sulfate-reducing bacteria to an artificial oil-spill in a coastal marine sediment. Environ. Microbiol. 13, 1488–1499. doi: 10.1111/j.1462-2920.2011.02451.x
Sun, M. Y., Dafforn, K. A., Johnston, E. L., and Brown, M. V. (2013). Core sediment bacteria drive community response to anthropogenic contamination over multiple environmental gradients. Environ. Microbiol. 15, 2517–2531. doi: 10.1111/1462-2920.12133
Sun, S., Jones, R. B., and Fodor, A. A. (2020). Inference-based accuracy of metagenome prediction tools varies across sample types and functional categories. Microbiome 8, 1–9. doi: 10.1186/s40168-020-00815-y
Suzuki, M., Rappé, M. S., and Giovannoni, S. J. (1998). Kinetic bias in estimates of coastal picoplankton community structure obtained by measurements of small-subunit rRNA gene PCR amplicon length heterogeneity. Appl. Environ. Microbiol. 64, 4522–4529. doi: 10.1128/aem.64.11.4522-4529.1998
Takahashi, S., Tomita, J., Nishioka, K., Hisada, T., and Nishijima, M. (2014). Development of a prokaryotic universal primer for simultaneous analysis of Bacteria and Archaea using next-generation sequencing. PLoS One 9:e105592. doi: 10.1371/journal.pone.0105592
Terrados, J., and Ros, J. (1991). Production dynamics in a macrophyte-dominated ecosystem: The Mar Menor coastal lagoon (SE Spain). Oecologia Aquat. 10, 255–270.
Terrados, J., and Ros, J. D. (1992). Growth and primary production of Cymodocea nodosa (Ucria) Ascherson in a Mediterranean coastal lagoon: The Mar Menor (SE Spain). Aquat. Bot. 43, 63–74. doi: 10.1016/0304-3770(92)90014-A
Thode-Andersen, S., and Jørgensen, B. B. (1989). Sulfate reduction and the formation of 35S-labeled FeS, FeS2, and S0 in coastal marine sediments. Limnol. Oceanogr. 34, 793–806. doi: 10.4319/lo.1989.34.5.0793
Toes, A. C. M., Finke, N., Kuenen, J. G., and Muyzer, G. (2008). Effects of deposition of heavy-metal-polluted harbor mud on microbial diversity and metal resistance in sandy marine sediments. Arch. Environ. Contam. Toxicol. 55, 372–385. doi: 10.1007/s00244-008-9135-4
Torti, A., Jørgensen, B. B., and Lever, M. A. (2018). Preservation of microbial DNA in marine sediments: Insights from extracellular DNA pools. Environ. Microbiol. 20, 4526–4542. doi: 10.1111/1462-2920.14401
Treude, N., Rosencrantz, D., Liesack, W., and Schnell, S. (2003). Strain FAc12, a dissimilatory iron-reducing member of the Anaeromyxobacter subgroup of Myxococcales. FEMS Microbiol. Ecol. 44, 261–269. doi: 10.1016/S0168-6496(03)00048-5
Ugarelli, K., Chakrabarti, S., Laas, P., and Stingl, U. (2017). The seagrass holobiont and its microbiome. Microorganisms 5:81. doi: 10.3390/microorganisms5040081
Ugarelli, K., Laas, P., and Stingl, U. (2019). The microbial communities of leaves and roots associated with turtle grass (Thalassia testudinum) and manatee grass (Syringodium filliforme) are distinct from seawater and sediment communities, but are similar between species and sampling sites. Microorganisms 7:4. doi: 10.3390/microorganisms7010004
Urich, T., Lanzén, A., Stokke, R., Pedersen, R. B., Bayer, C., Thorseth, I. H., et al. (2014). Microbial community structure and functioning in marine sediments associated with diffuse hydrothermal venting assessed by integrated meta-omics. Environ. Microbiol. 16, 2699–2710. doi: 10.1111/1462-2920.12283
Vandieken, V., Mußmann, M., Niemann, H., and Jørgensen, B. B. (2006). Desulfuromonas svalbardensis sp. nov. and Desulfuromusa ferrireducens sp. nov., psychrophilic, Fe(III)-reducing bacteria isolated from Arctic sediments, Svalbard. Int. J. Syst. Evol. Microbiol. 56, 1133–1139. doi: 10.1099/ijs.0.63639-0
Vepraskas, M. J., and Faulkner, S. P. (2001). Redox chemistry of hydric soils. Boca Raton, FL: Lewis.
Williams, T. A., Cox, C. J., Foster, P. G., Szöllõsi, G. J., and Embley, T. M. (2020). Phylogenomics provides robust support for a two-domains tree of life. Nat. Ecol. Evol. 4, 138–147. doi: 10.1038/s41559-020-01347-2
Wu, S., Li, R., Xie, S., and Shi, C. (2019). Depth-related change of sulfate-reducing bacteria community in mangrove sediments: The influence of heavy metal contamination. Mar. Pollut. Bull. 140, 443–450. doi: 10.1016/j.marpolbul.2019.01.042
Xing, L., Zhang, W., Gu, M., Yin, Q., and Wu, G. (2020). Microbial interactions regulated by the dosage of ferroferric oxide in the co-metabolism of organic carbon and sulfate. Bioresour. Technol. 296:122317. doi: 10.1016/j.biortech.2019.122317
Yamada, T., and Sekiguchi, Y. (2015). “Anaerolineaceae,” in Bergey’s manual of systematics of archaea and bacteria, eds W. B. Whitman, F. Rainey, P. Kämpfer, M. Trujillo, J. Chun, P. DeVos, B. Hedlund, and S. Dedysh (Hoboken, NJ: John Wiley & Sons), 1–5. doi: 10.1002/9781118960608.fbm00301
Zaremba-Niedzwiedzka, K., Caceres, E. F., Saw, J. H., Bäckström, D., Juzokaite, L., Vancaester, E., et al. (2017). Asgard archaea illuminate the origin of eukaryotic cellular complexity. Nature 541, 353–358. doi: 10.1038/nature21031
Zhang, Y., Ma, Y., Zhang, R., Zhang, B., Zhai, X., Li, W., et al. (2019). Metagenomic resolution of functional diversity in copper surface-associated marine biofilms. Front. Microbiol. 10:2863. doi: 10.3389/fmicb.2019.02863
Keywords: 16S rRNA gene amplicon sequencing, coastal lagoon, Mar Menor, sediment, microbial community
Citation: Aldeguer-Riquelme B, Rubio-Portillo E, Álvarez-Rogel J, Giménez-Casalduero F, Otero XL, Belando M-D, Bernardeau-Esteller J, García-Muñoz R, Forcada A, Ruiz JM, Santos F and Antón J (2022) Factors structuring microbial communities in highly impacted coastal marine sediments (Mar Menor lagoon, SE Spain). Front. Microbiol. 13:937683. doi: 10.3389/fmicb.2022.937683
Received: 06 May 2022; Accepted: 27 July 2022;
Published: 07 September 2022.
Edited by:
Danny Ionescu, Leibniz-Institute of Freshwater Ecology and Inland Fisheries (IGB), GermanyReviewed by:
Gustavo Bueno Gregoracci, Federal University of São Paulo, BrazilCarmen Rizzo, Stazione Zoologica Anton Dohrn Napoli, Italy
Copyright © 2022 Aldeguer-Riquelme, Rubio-Portillo, Álvarez-Rogel, Giménez-Casalduero, Otero, Belando, Bernardeau-Esteller, García-Muñoz, Forcada, Ruiz, Santos and Antón. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Josefa Antón, YW50b25AdWEuZXM=