 Sihai Zhao
Sihai Zhao Jianglin Fan
Jianglin Fan Enqi Liu
Enqi Liu
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
OPINION article
Front. Microbiol., 24 June 2022
Sec. Virology
Volume 13 - 2022 | https://doi.org/10.3389/fmicb.2022.907406
This article is part of the Research TopicInsights of Important Mammalian Viruses: Infection, Pathogenesis and DrugsView all 10 articles
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) causes coronavirus disease 2019 (COVID-19), which has spread worldwide and reached a pandemic level within several months (Bedford et al., 2020; Li et al., 2020; Hu et al., 2021). Globally, scientists are trying to understand and defeat COVID-19 using animal studies to explore potential therapeutics. Vaccination remains the best choice to protect susceptible populations; therefore, many institutions and companies have developed vaccines against SARS-CoV-2 infection and many therapeutics are under clinical investigations. For these undertakings, it is essential to use suitable COVID-19 animal models (Caldera-Crespo et al., 2021; Munoz-Fontela et al., 2022).
Animal models are usually used in medical experimentation as an alternative to avoid potential hazards in humans (Liu and Fan, 2017). Appropriate COVID-19 animal models should be a very powerful tool in the study of the onset, development and culmination of this disease and provide helpful knowledge for its management. Making and choosing good models for COVID-19 requires taking full consideration of the following: (1) the animals should be susceptible to SARS-CoV-2, especially easily infected by the virus via the respiratory route; (2) the characteristics of the induced phenotypes or manifestations are similar to those of human COVID-19 patients, such as fever, coughing and myalgia or fatigue, pneumonia detected by computed tomography (CT) examination, along with sex- or age-related differences in severity (Huang et al., 2020); (3) immune responses should be similar to those in human coronavirus infections; and (4) the animals should have an appropriate cost. When testing vaccine protection effects, recovered infected animals should gain immunity to resist reinfection. These principles should be used for creating new models or evaluating candidate models for the development of COVID-19 therapeutics.
Due to the similarity of SARS-CoV-2 to other two beta coronaviruses, SARS-CoV-1 and Middle Eastern respiratory syndrome coronavirus (MERS-CoV), lessons from SARS-CoV-1 vaccine development can be helpful for accelerating SARS-CoV-2 vaccine development (Amanat and Krammer, 2020). Scientists have designed several SARS-CoV-2 vaccines to prevent COVID-19. Although the SARS-CoV-2 vaccine did not catch up with the first wave of the pandemic, the application of the vaccine has reduced the severe incidence of COVID-19 patients. Several faithful animal models for studying the pathogenesis of SARS-CoV-2 and its vaccine development have recently been created by scientists and have shed light on COVID-19 research. Here, we provide a brief review of these models, summarize what we have learned, and discuss their usefulness for testing the immunological response of vaccine injection and post-challenge safety.
Non-human primates (the rhesus macaque or Macaca mulatta, cynomolgus monkey, or Macaca fascicularis, common marmoset or Callithrix jacchus and African green monkeys or Chlorocebus aethiops) have been used to explore the pathogenesis and test the potential therapies of SARS-CoV-2 (Hartman et al., 2020; Lu et al., 2020; Rockx et al., 2020; Yu et al., 2020; Woolsey et al., 2021). All the above mentioned primates can be infected by experimental inoculation with SARS-CoV-2. The rhesus macaque is the most susceptible to SARS-CoV2 infection, followed by the cynomolgus monkey and the common marmoset (Lu et al., 2020). The viral replication state of nasopharyngeal swabs, anal swabs and lungs in old rhesus macaques was more active than that in young monkeys for 14 days after SARS-CoV-2 challenge (Yu et al., 2020). In SARS-CoV-2-infected macaques, viruses were discharged from the nose and throat in the absence of clinical symptoms and could be detected in both type I and II pneumocytes in the foci of diffuse alveolar damage and ciliated epithelial cells of the nasal, bronchial, and bronchiolar mucosae (Rockx et al., 2020). SARS-CoV-2 also causes pulmonary infiltrates in these monkeys, a hallmark of human disease shown in CT scans, which can be detected by chest radiographs in all infected rhesus macaques (Munster et al., 2020). Another important finding is that monkeys that have recovered from SARS-CoV-2 infection cannot be infected again in the near future (Yu et al., 2020). This finding also provided an important clue for antibody-mediated protection, suggesting that COVID-19 can be prevented through either natural infection or vaccination. Autopsy examinations revealed that the lungs, throat, bronchi, and spleen were infected in both rhesus monkeys and cynomolgus monkeys (Lu et al., 2020; Yu et al., 2020). These observations suggested that the rhesus monkey and cynomolgus monkey models can be used to evaluate vaccines and drugs to treat or prevent COVID-19. Gao et al. developed a purified inactivated SARS-CoV-2 virus vaccine candidate (PiCoVacc) and then conducted a challenge experiment in rhesus monkeys. They reported that vaccine immunization significantly reduced the pathological changes in the lungs of rhesus monkeys and the viral titers were also significantly decreased whereas no antibody-dependent enhancement was observed (Gao et al., 2020).
A recent report showed that African green monkey (AGM), another non-human primate, had robust SARS-CoV-2 replication and developed pronounced respiratory disease (Hartman et al., 2020; Woolsey et al., 2021). Shedding of SARS-CoV-2 from both respiratory and gastrointestinal tracts was also documented in AGMs, which may mimic human COVID-19 cases (Hartman et al., 2020). Therefore, AGM may provide another option for COVID-19 therapeutics and vaccine evaluation. These data suggest that monkey models are useful to evaluate vaccines and therapeutics for treating or preventing COVID-19. The advantages and disadvantages of non-human primates compared with those of other animal models are summarized in Table 1. However, high costs and ethical issues may strictly limit the widespread use of non-human primate models in most institutions.
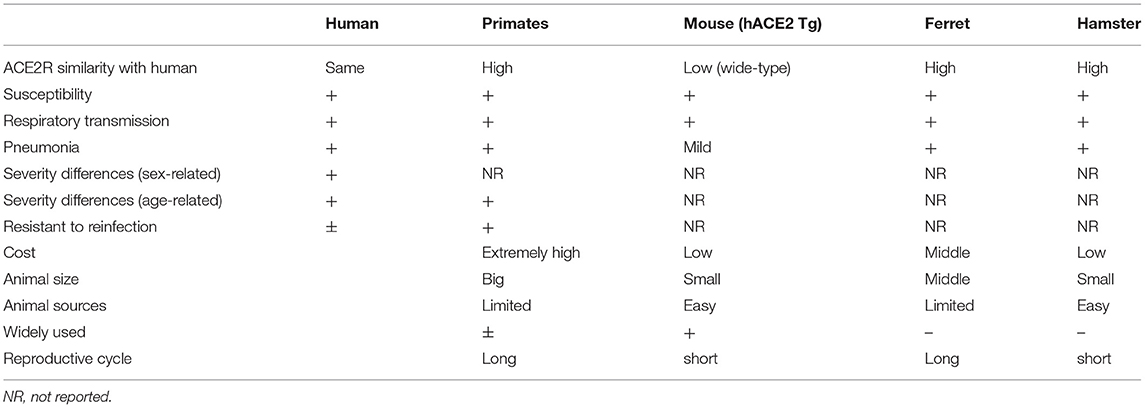
Table 1. Comparison of animal models and human COVID-19 features.
The genetically manipulated mouse model is relatively cheap and easy to use (Korner et al., 2020). It has been reported that SARS-CoV-2 infection can occur through the angiotensin converting enzyme II receptor (ACE2R) for cellular entry (Zhou et al., 2020). Since 11 of the 29 amino acids in the critical region of the human ACE2R receptor are different from those of the mouse, SARS-CoV-2 is unable to naturally infect mice. Therefore, human ACE2R transgenic mice were developed and analyzed for potential application in the study of COVID-19 (Netland et al., 2008; Bao et al., 2020). Although hACE2R transgenic mice showed signs of weight loss and interstitial pneumonia after infection, the symptoms were mild and much different from those of humans (Bao et al., 2020). The hACE2R transgenic mice developed by Netland et al. also exhibited brain infection of SARS-CoV-2, which is rare in humans (Netland et al., 2008). These drawbacks may puzzle or mislead researchers for interpretation of their studies. The development of lung cell-specific expression of the hACE2R gene in transgenic or knock-in models without endogenous mouse ACE2R gene may help to broaden the use of mouse models in COVID-19 studies in the future. Except for modifying the mouse ACE2R for SARS-CoV-2 infecting host cells, another reverse genetics strategy was to design a recombinant mouse-adapted SARS-CoV-2, designated as SARS-CoV-2 MA, which can use mouse ACE2R for entry into cells (Dinnon et al., 2020; Leist et al., 2020). SARS-CoV-2 MA was reported to successfully infect and replicate in the upper and lower airways in both adult and aged BALB/c mice (Dinnon et al., 2020). SARS-CoV-2 MA caused more severe disease in aged mice, which resembles the human features of COVID-19, and exhibited more clinically relevant phenotypes than those seen in Hfh4-ACE2R transgenic mice (Dinnon et al., 2020). This model provided a powerful tool for the study of SARS-CoV-2 pathogenesis and has been used to evaluate vaccine and antiviral therapeutics performance (Leist et al., 2020).
As rats are unsusceptible to SARS-CoV-2, they are not used to study of COVID-19. However, Syrian hamster ACE2R proteins are reported to be highly similar to human ACE2R with 3–4 mutations at the interface (Chan et al., 2020). After being infected with SARS-CoV-2, hamsters showed weight loss, drowsiness, hairiness, hunched back, and shortness of breath. Olfactory and taste dysfunctions are common in mildly symptomatic COVID-19 patients. The food-searching behavioral test showed that anosmia may happen in SARS-CoV-2-infected hamsters, and the level of anosmia was associated with olfactory epithelium damage (Reyna et al., 2022). In addition, high titers of SARS-CoV-2 were found in the hamster lungs and intestines. The phenotypes of human upper and lower respiratory tract infections are very similar to those of hamsters, so this model benefits scientists who like to use small animals to perform COVID-19 research or therapeutic screening (Sia et al., 2020).
Ferrets, another small animal, have special value in the research of virus-related diseases. After being infected with respiratory viruses, ferrets can show symptoms similar to those of humans. For example, ferrets infected with influenza virus can even sneeze like humans, and can easily transmit the virus to other ferrets (Herfst et al., 2012). SARS-CoV-2 can infect ferrets through the respiratory tract and can replicate in infected cells (Shi et al., 2020). The temperature of the infected ferrets rises slightly and infection does not cause serious illness or death. Although fatalities were not observed, SARS-CoV-2-infected ferrets shed the virus in nasal washes, saliva, urine, and feces up to 8 days post-infection (Kim et al., 2020). Ferrets give birth once per year, with 6–8 l, so they do not breed as rapidly as mice. To a certain extent, the slow reproduction speed may limit the wide application of ferrets. Similar to ferrets, farmed minks are also susceptible to coronaviruses. On farms, minks are infected with SARS-CoV-2 and can transmit it to humans (Enserink, 2020; Oreshkova et al., 2020; Sharun et al., 2021). Laboratory evidence also showed SARS-CoV-2 can be transmitted among minks and resulted severe respiratory syndrome (Shuai et al., 2021). Among middle- or large-sized animals, it was reported that SARS-CoV-2 can replicate efficiently in cats, with younger cats being more permissive and perhaps more importantly, the virus can transmit between cats via the airborne route (Shi et al., 2020). Dogs exhibit low susceptibility, whereas livestock, including pigs, chickens, and ducks, are not susceptible to SARS-CoV-2 (Shi et al., 2020). Previous reviews also have discussed the potentials of these animal species in COVID-19 studies (Munoz-Fontela et al., 2020; Sharun et al., 2020; Shou et al., 2021).
The currently reported COVID-19 animal models were summarized in Table 1. The advantages and disadvantages of the COVID-19 animal models that are currently available was discussed in this and previous study (Shou et al., 2021). To date, it seems that no animal model is perfect when evaluating potential drugs and vaccines (Munoz-Fontela et al., 2022). The most useful animal model is the rhesus macaque, but because of the substantial cost and specific facility requirements, the monkey model is unlikely to be widely used in common laboratories. The ferret model has special applicability in the study of respiratory viruses, but the low breeding rate and the lack of suitable facilities make it difficult for wide use in COVID-19 studies. The limitations of COVID-19 animal models are also apparent (Sharun et al., 2020). First, no model can fully mimic human COVID-19, and the findings of each model are needed to confirm with other animal models and to increase its credibility. Second, although the non-human primate model can better represent human COVID-19, it faces stricter ethical review and is not easy to be widely used. Mice are cheap and there are many genetically modified models available, but their immune system responses differ from those of humans. Humanized mouse models may become a feasible study tool for COVID-19 research in the future. At present, various genetically engineered mouse models are currently on the way, and hopefully, these new models will provide a unique research tool to fight against COVID-19 in the future.
JF and EL designed this project. SZ wrote the paper. All authors contributed to the article and approved the submitted version.
This study was supported by grants from the Natural Science Foundation of Shaanxi Province (Grant Nos. 2020PT-004 and 2020PT-001).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Amanat, F., and Krammer, F. (2020). SARS-CoV-2 vaccines: status report. Immunity 52, 583–589. doi: 10.1016/j.immuni.2020.03.007
Bao, L., Deng, W., Huang, B., Gao, H., Liu, J., Ren, L., et al. (2020). The pathogenicity of SARS-CoV-2 in hACE2 transgenic mice. Nature 583, 830–833. doi: 10.1038/s41586-020-2312-y
Bedford, J., Enria, D., Giesecke, J., Heymann, D. L., Ihekweazu, C., Kobinger, G., et al. (2020). COVID-19: towards controlling of a pandemic. Lancet 395, 1015–1018. doi: 10.1016/S0140-6736(20)30673-5
Caldera-Crespo, L. A., Paidas, M. J., Roy, S., Schulman, C. I., Kenyon, N. S., Daunert, S., et al. (2021). Experimental models of COVID-19. Front. Cell. Infect. Microbiol. 11, 792584. doi: 10.3389/fcimb.2021.792584
Chan, J. F., Zhang, A. J., Yuan, S., Poon, V. K., Chan, C. C., Lee, A. C., et al. (2020). Simulation of the clinical and pathological manifestations of Coronavirus disease 2019 (COVID-19) in a golden syrian hamster model: implications for disease pathogenesis and transmissibility. Clin. Infect. Dis. 71, 2428–2446. doi: 10.1093/cid/ciaa644
Dinnon, K. H. III., Leist, S. R., Schafer, A., Edwards, C. E., Martinez, D. R., Montgomery, S. A., et al. (2020). A mouse-adapted model of SARS-CoV-2 to test COVID-19 countermeasures. Nature 586, 560–566. doi: 10.1038/s41586-020-2708-8
Enserink, M. (2020). Coronavirus rips through Dutch mink farms, triggering culls. Science 368, 1169. doi: 10.1126/science.368.6496.1169
Gao, Q., Bao, L., Mao, H., Wang, L., Xu, K., Yang, M., et al. (2020). Development of an inactivated vaccine candidate for SARS-CoV-2. Science 369, 77–81. doi: 10.1126/science.abc1932
Hartman, A. L., Nambulli, S., McMillen, C. M., White, A. G., Tilston-Lunel, N. L., Albe, J. R., et al. (2020). SARS-CoV-2 infection of African green monkeys results in mild respiratory disease discernible by PET/CT imaging and shedding of infectious virus from both respiratory and gastrointestinal tracts. PLoS Pathog. 16, e1008903. doi: 10.1371/journal.ppat.1008903
Herfst, S., Schrauwen, E. J., Linster, M., Chutinimitkul, S., de Wit, E., Munster, V. J., et al. (2012). Airborne transmission of influenza A/H5N1 virus between ferrets. Science 336, 1534–1541. doi: 10.1126/science.1213362
Hu, B., Guo, H., Zhou, P., and Shi, Z. L. (2021). Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 19, 141–154. doi: 10.1038/s41579-020-00459-7
Huang, C., Wang, Y., Li, X., Ren, L., Zhao, J., Hu, Y., et al. (2020). Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 395, 497–506. doi: 10.1016/S0140-6736(20)30183-5
Kim, Y. I., Kim, S. G., Kim, S. M., Kim, E. H., Park, S. J., Yu, K. M., et al. (2020). Infection and rapid transmission of SARS-CoV-2 in ferrets. Cell Host Microbe 27, 704–709.e2. doi: 10.1016/j.chom.2020.03.023
Korner, R. W., Majjouti, M., Alcazar, M. A. A., and Mahabir, E. (2020). Of mice and men: the Coronavirus MHV and mouse models as a translational approach to understand SARS-CoV-2. Viruses 12, 880. doi: 10.3390/v12080880
Leist, S. R., Dinnon, K. H. III., Schafer, A., Tse, L. V., Okuda, K., Hou, Y. J., et al. (2020). A mouse-adapted SARS-CoV-2 induces acute lung injury and mortality in standard laboratory mice. Cell 183, 1070–1085.e12. doi: 10.1016/j.cell.2020.09.050
Li, Q., Guan, X., Wu, P., Wang, X., Zhou, L., Tong, Y., et al. (2020). Early transmission dynamics in Wuhan, China, of novel coronavirus-infected pneumonia. N. Engl. J. Med. 382, 1199–1207. doi: 10.1056/NEJMoa2001316
Liu, E., and Fan, J. (2017). Fundamentals of Laboratory Animal Science, 1st Edn. New York, NY: CRC Press. doi: 10.1201/9781315368993
Lu, S., Zhao, Y., Yu, W., Yang, Y., Gao, J., Wang, J., et al. (2020). Comparison of nonhuman primates identified the suitable model for COVID-19. Signal Transduct. Target Ther. 5, 157. doi: 10.1038/s41392-020-00269-6
Munoz-Fontela, C., Dowling, W. E., Funnell, S. G. P., Gsell, P. S., Riveros-Balta, A. X., Albrecht, R. A., et al. (2020). Animal models for COVID-19. Nature 586, 509–515. doi: 10.1038/s41586-020-2787-6
Munoz-Fontela, C., Widerspick, L., Albrecht, R. A., Beer, M., Carroll, M. W., de Wit, E., et al. (2022). Advances and gaps in SARS-CoV-2 infection models. PLoS Pathog. 18, e1010161. doi: 10.1371/journal.ppat.1010161
Munster, V. J., Feldmann, F., Williamson, B. N., van Doremalen, N., Perez-Perez, L., Schulz, J., et al. (2020). Respiratory disease in rhesus macaques inoculated with SARS-CoV-2. Nature 585, 268–272. doi: 10.1038/s41586-020-2324-7
Netland, J., Meyerholz, D. K., Moore, S., Cassell, M., and Perlman, S. (2008). Severe acute respiratory syndrome coronavirus infection causes neuronal death in the absence of encephalitis in mice transgenic for human ACE2. J. Virol. 82, 7264–7275. doi: 10.1128/JVI.00737-08
Oreshkova, N., Molenaar, R. J., Vreman, S., Harders, F., Oude Munnink, B. B., Hakze-van der Honing, R. W., et al. (2020). SARS-CoV-2 infection in farmed minks, the Netherlands, April and May 2020. Euro Surveill. 25, 2001005. doi: 10.2807/1560-7917.ES.2020.25.23.2001005
Reyna, R. A., Kishimoto-Urata, M., Urata, S., Makishima, T., Paessler, S., and Maruyama, J. (2022). Recovery of anosmia in hamsters infected with SARS-CoV-2 is correlated with repair of the olfactory epithelium. Sci. Rep. 12, 628. doi: 10.1038/s41598-021-04622-9
Rockx, B., Kuiken, T., Herfst, S., Bestebroer, T., Lamers, M. M., Oude Munnink, B. B., et al. (2020). Comparative pathogenesis of COVID-19, MERS, and SARS in a nonhuman primate model. Science 368, 1012–1015. doi: 10.1126/science.abb7314
Sharun, K., Tiwari, R., Natesan, S., and Dhama, K. (2021). SARS-CoV-2 infection in farmed minks, associated zoonotic concerns, and importance of the one health approach during the ongoing COVID-19 pandemic. Vet. Q. 41, 50–60. doi: 10.1080/01652176.2020.1867776
Sharun, K., Tiwari, R., Patel, S. K., Karthik, K., Iqbal Yatoo, M., Malik, Y. S., et al. (2020). Coronavirus disease 2019 (COVID-19) in domestic animals and wildlife: advances and prospects in the development of animal models for vaccine and therapeutic research. Hum. Vaccin. Immunother. 16, 3043–3054. doi: 10.1080/21645515.2020.1807802
Shi, J., Wen, Z., Zhong, G., Yang, H., Wang, C., Huang, B., et al. (2020). Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science 368, 1016–1020. doi: 10.1126/science.abb7015
Shou, S., Liu, M., Yang, Y., Kang, N., Song, Y., Tan, D., et al. (2021). Animal models for COVID-19: hamsters, mouse, ferret, mink, tree shrew, non-human primates. Front. Microbiol. 12, 626553. doi: 10.3389/fmicb.2021.626553
Shuai, L., Zhong, G., Yuan, Q., Wen, Z., Wang, C., He, X., et al. (2021). Replication, pathogenicity, and transmission of SARS-CoV-2 in minks. Natl. Sci. Rev. 8, nwaa291. doi: 10.1093/nsr/nwaa291
Sia, S. F., Yan, L. M., Chin, A. W. H., Fung, K., Choy, K. T., Wong, A. Y. L., et al. (2020). Pathogenesis and transmission of SARS-CoV-2 in golden hamsters. Nature 583, 834–838. doi: 10.1038/s41586-020-2342-5
Woolsey, C., Borisevich, V., Prasad, A. N., Agans, K. N., Deer, D. J., Dobias, N. S., et al. (2021). Establishment of an African green monkey model for COVID-19 and protection against re-infection. Nat. Immunol. 22, 86–98. doi: 10.1038/s41590-020-00835-8
Yu, P., Qi, F., Xu, Y., Li, F., Liu, P., Liu, J., et al. (2020). Age-related rhesus macaque models of COVID-19. Anim. Model Exp. Med. 3, 93–97. doi: 10.1002/ame2.12108
Keywords: animal models, COVID-19, therapeutics, vaccines, humanized mouse model
Citation: Zhao S, Fan J and Liu E (2022) Animal Models for COVID-19 Therapeutic Development: Where We Are and Where We Need to Go. Front. Microbiol. 13:907406. doi: 10.3389/fmicb.2022.907406
Received: 29 March 2022; Accepted: 30 May 2022;
Published: 24 June 2022.
Edited by:
Anna Kramvis, University of the Witwatersrand, South AfricaReviewed by:
Junki Maruyama, University of Texas Medical Branch at Galveston, United StatesCopyright © 2022 Zhao, Fan and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Enqi Liu, bGl1ZW5xaUB4anR1LmVkdS5jbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.