 Simon Houston1
Simon Houston1 Ethan Schovanek1
Ethan Schovanek1 Kate M. E. Conway1
Kate M. E. Conway1 Sarah Mustafa1
Sarah Mustafa1 Alloysius Gomez1Raghavendran Ramaswamy1Ayman Haimour1
Alloysius Gomez1Raghavendran Ramaswamy1Ayman Haimour1 Martin J. Boulanger1
Martin J. Boulanger1 Lisa A. Reynolds1
Lisa A. Reynolds1 Caroline E. Cameron1,2*
Caroline E. Cameron1,2*
- 1Department of Biochemistry and Microbiology, University of Victoria, Victoria, BC, Canada
- 2Division of Allergy and Infectious Diseases, Department of Medicine, University of Washington, Seattle, WA, United States
The etiological agent of syphilis, Treponema pallidum ssp. pallidum, is a highly invasive “stealth” pathogen that can evade the host immune response and persist within the host for decades. This obligate human pathogen is adept at establishing infection and surviving at sites within the host that have a multitude of competing microbes, sometimes including pathogens. One survival strategy employed by bacteria found at polymicrobial sites is elimination of competing microorganisms by production of antimicrobial peptides (AMPs). Antimicrobial peptides are low molecular weight proteins (miniproteins) that function directly via inhibition and killing of microbes and/or indirectly via modulation of the host immune response, which can facilitate immune evasion. In the current study, we used bioinformatics to show that approximately 7% of the T. pallidum proteome is comprised of miniproteins of 150 amino acids or less with unknown functions. To investigate the possibility that AMP production is an unrecognized defense strategy used by T. pallidum during infection, we developed a bioinformatics pipeline to analyze the complement of T. pallidum miniproteins of unknown function for the identification of potential AMPs. This analysis identified 45 T. pallidum AMP candidates; of these, Tp0451a and Tp0749 were subjected to further bioinformatic analyses to identify AMP critical core regions (AMPCCRs). Four potential AMPCCRs from the two predicted AMPs were identified and peptides corresponding to these AMPCCRs were experimentally confirmed to exhibit bacteriostatic and bactericidal activity against a panel of biologically relevant Gram-positive and Gram-negative bacteria. Immunomodulation assays performed under inflammatory conditions demonstrated that one of the AMPCCRs was also capable of differentially regulating expression of two pro-inflammatory chemokines [monocyte chemoattractant protein-1 (MCP-1) and interleukin-8 (IL-8)]. These findings demonstrate proof-of-concept for our developed AMP identification pipeline and are consistent with the novel concept that T. pallidum expresses AMPs to defend against competing microbes and modulate the host immune response.
Introduction
The spirochete bacterium, Treponema pallidum ssp. pallidum (hereafter T. pallidum), is the causative agent of syphilis, a chronic, multistage infection that is transmitted sexually or in utero. Following infection, T. pallidum traverses endothelial barriers and undergoes rapid and widespread dissemination via the circulatory system to infect every organ and tissue, including immunologically privileged sites such as the eyes (Marra et al., 1991; Muller et al., 2007), testes (Sell et al., 1980), and central nervous system (Collart et al., 1971; Lukehart et al., 1988). Despite host-initiated innate and adaptive immune responses, T. pallidum is able to persist within the host for decades (Lafond and Lukehart, 2006). The remarkable ability of T. pallidum to evade the immune system and establish and maintain persistent infection has earned it the designation of the “stealth” pathogen (Radolf, 1994).
During infection of a host, T. pallidum, which has a slow generation time of 30–33 h (Magnuson and Eagle, 1948; Cumberland and Turner, 1949), are introduced into anatomical sites that are abundant in species of microbiota, including the genital tract, skin, rectum, and oral cavity (Lafond and Lukehart, 2006), features that may put T. pallidum at a growth disadvantage in a polymicrobial environment. In addition, T. pallidum can present as a co-infection with other viral, fungal, parasitic, and bacterial pathogens, including the sexually transmitted pathogen, Neisseria gonorrhoeae (Bala et al., 2011; World Health Organization, 2019; Pinho-Bandeira et al., 2020; Coelho et al., 2021). Inhibition and elimination of competing microorganisms via production of antimicrobial peptides (AMPs) allows the microbiota and pathogenic bacteria found at polymicrobial sites to gain a competitive advantage (Meade et al., 2020). The ability of T. pallidum to establish an infection and survive in anatomical locations with a complex polymicrobial profile raised the question of whether this bacterium could use AMP production to eliminate microbial competition.
AMPs are a structurally and functionally diverse class of low molecular weight proteins produced by all branches of life (Kumar et al., 2018). Typically comprised of 10–150 amino acids, they often form amphipathic alpha helices facilitated by their net positive charge and high hydrophobic content (Kumar et al., 2018). AMPs have a direct mechanism of action that occurs via electrostatic interactions between positively-charged AMPs and negatively-charged microbial surfaces (Kumar et al., 2018). The amphipathic secondary structure of AMPs promotes membrane integration and pore formation, resulting in membrane destabilization and cell lysis as well as inhibition of essential intracellular functions such as DNA and protein synthesis (Kumar et al., 2018; Mishra et al., 2018). A second, indirect effect of AMPs can be alteration of the host immune response, including modulation of inflammatory cytokine production, immune cell recruitment and activation (Hilchie et al., 2013). The immunomodulatory effects of eukaryotic AMPs have been well documented, and recent studies have shown bacterial AMPs can have similar immunomodulatory activities (Kindrachuk et al., 2013; Malaczewska et al., 2019) that can promote bacterial survival and host infection via subversion and evasion of the host immune response (Li et al., 2014).
An important characteristic of AMPs is the presence of functionally essential regions that correspond to the shortest stretch of amino acids (often ∼10–20 residues) that retain antimicrobial effects (Chang et al., 2015). Identification of these key regions, defined as antimicrobial peptide critical core regions (AMPCCRs), allows for the design and development of discrete peptides with antimicrobial activity that are derived from, and more tractable than, their larger precursor proteins (Torrent et al., 2009, 2012).
More than two decades have passed since the first T. pallidum whole genome sequence was published (Fraser et al., 1998). Since then, many laboratory and clinical strains of T. pallidum ssp. pallidum have been sequenced, yet only three genes have been annotated as homologs of known AMP-related genes in other bacteria. These are tp0688 [Bacillus anthracis mccF, encoding the Microcin C7 self-immunity protein (Gonzalez-Pastor et al., 1995)], tp0522 [Escherichia coli cvpA, encoding the Colicin V Production protein that is required for production and secretion of the AMP Colicin V (Gilson et al., 1990)], and tp0405 [E. coli mcbG, encoding the Microcin B17 self-immunity protein (San Millan et al., 1985)]. Recent T. pallidum genome sequencing also identified a novel 91-amino acid miniprotein (TPANIC_RS05485) which has been annotated as a putative CPBP (CAAX Protease and Bacteriocin-Processing) family intramembrane metalloprotease. Evidence suggests some members of this family may be involved in bacterial AMP processing (Pei et al., 2011). One reason that may partially account for the low number of AMP-related genes detected within T. pallidum to date is that the bacterium is phylogenetically distinct, with approximately 300 genes/30% of the genome predicted to encode proteins with no known orthologs or assigned functions (Fraser et al., 1998; Petrosova et al., 2013).
The present study shows that approximately one quarter of T. pallidum genes of unknown function are predicted to encode miniproteins of 150 amino acids or less. Bioinformatic analyses show a portion of these miniproteins possess characteristics consistent with known AMPs. These findings, when considered in the context of the success of this bacterium at establishing infection at polymicrobial anatomical sites, prompted us to investigate whether AMP production is an unexplored pathogenic mechanism used by T. pallidum to defend against competing microbes and the host. Herein, we investigated this potential treponemal defense strategy using a combination of bioinformatics, structure modeling, antimicrobial susceptibility testing, and immunomodulation assays. Our investigations have provided experimental confirmation of AMP activity within two T. pallidum miniproteins, consistent with the novel concept that T. pallidum expresses AMPs to establish and maintain infection at polymicrobial sites in the human host.
Materials and Methods
Bacterial Strains and Culture
Bacterial strains used in this study were: E. coli ATCC 9723H, Pseudomonas aeruginosa ATCC 10148, Staphylococcus aureus ATCC 6538P (penicillin resistant), Streptococcus pyogenes (hospital isolate, strain unknown), Mycobacterium smegmatis MC2155, N. gonorrhoeae ATCC 700825 (streptomycin resistant), and Salmonella enterica subsp. enterica serovar Typhimurium SL1344 (streptomycin resistant). E. coli, P. aeruginosa, S. aureus, M. smegmatis, and S. enterica were cultured aerobically at 37°C in Mueller Hinton broth (MHB) (Sigma-Aldrich, MO, United States) and on nonselective Mueller Hinton agar (MHA) plates. S. pyogenes was cultured in 5% carbon dioxide at 37°C in MHB supplemented with 5% lysed horse blood (Quad Five, MT, United States) (MHB + 5% HB) and on nonselective MHA plates supplemented with 5% defibrinated sheep blood (Quad Five, MT, United States) (MHA + 5% SB). N. gonorrhoeae was cultured in 5% carbon dioxide at 37°C in gonococcal (GC) chocolate broth medium and on nonselective GC agar plates [GC medium base (BD Difco, MD, United States) supplemented with 1% BBL™ hemoglobin (BD Biosciences, MD, United States) and 1% IsoVitaleX (BD Biosciences, MD, United States)]. Prior to antimicrobial susceptibility assays (as described below), N. gonorrhoeae cultures were subcultured on nonselective GC agar plates (as described above), to ensure bacterial viability. All bacterial stocks were stored in 20% glycerol at –80°C.
Treponema pallidum Propagation and in vitro Culture
Treponema pallidum subsp. pallidum (Nichols strain) was propagated in, and extracted from, New Zealand White rabbits as described elsewhere (Lukehart and Marra, 2007), and stored in liquid nitrogen. Frozen treponemal stocks were then used for in vitro culture and sub-culture of T. pallidum in the presence of Sf1Ep (NBL-11) cottontail rabbit epithelial cells (ATCC CCL-68) [American Type Culture Collection (ATCC), Rockville, MD, United States]. Continuous axenic culture of T. pallidum in the absence of mammalian cells has not been achieved, and it is believed that the direct adherence of T. pallidum to Sf1Ep cells is required for the long term replication of T. pallidum in vitro (Edmondson et al., 2018). Dissociation of T. pallidum from Sf1Ep cells was accomplished using trypsin-free dissociation buffer [2 mL: 64% cell culture grade water (Sigma Aldrich), 10% modified EBSS (Earle’s Balanced salt solution, 10×), 1% non-essential amino acids (Thermo Fisher Scientific), 0.15% sodium bicarbonate (Sigma Aldrich), 0.728% 100 mM sodium pyruvate (Sigma Aldrich), 0.136% 0.5M EDTA (Thermo Fisher Scientific), 0.16 mg dithiothreitol (DTT) (Sigma Aldrich)] followed by a low speed centrifugation step (220 × g) to separate T. pallidum from the rabbit cells, as previously described (Edmondson et al., 2018; Edmondson and Norris, 2021).
Bioinformatics Pipeline: In silico Analysis of T. pallidum Whole Proteomes
The flow diagram shown in Figure 1 outlines all major steps that comprised our bioinformatics pipeline for the identification of potential AMPs and AMPCCRs in the T. pallidum proteome. As the first step in this approach, the whole proteome of T. pallidum (Nichols strain NC_021490) was obtained from the National Center for Biotechnology Information (NCBI) Genome database1 and manually searched in order to identify all functionally-unannotated miniproteins containing 150 amino acids or less (Figure 1A). All in silico proteomic analyses performed on the T. pallidum strain reported in the current study were based on the NCBI whole proteome annotation from July 2021.
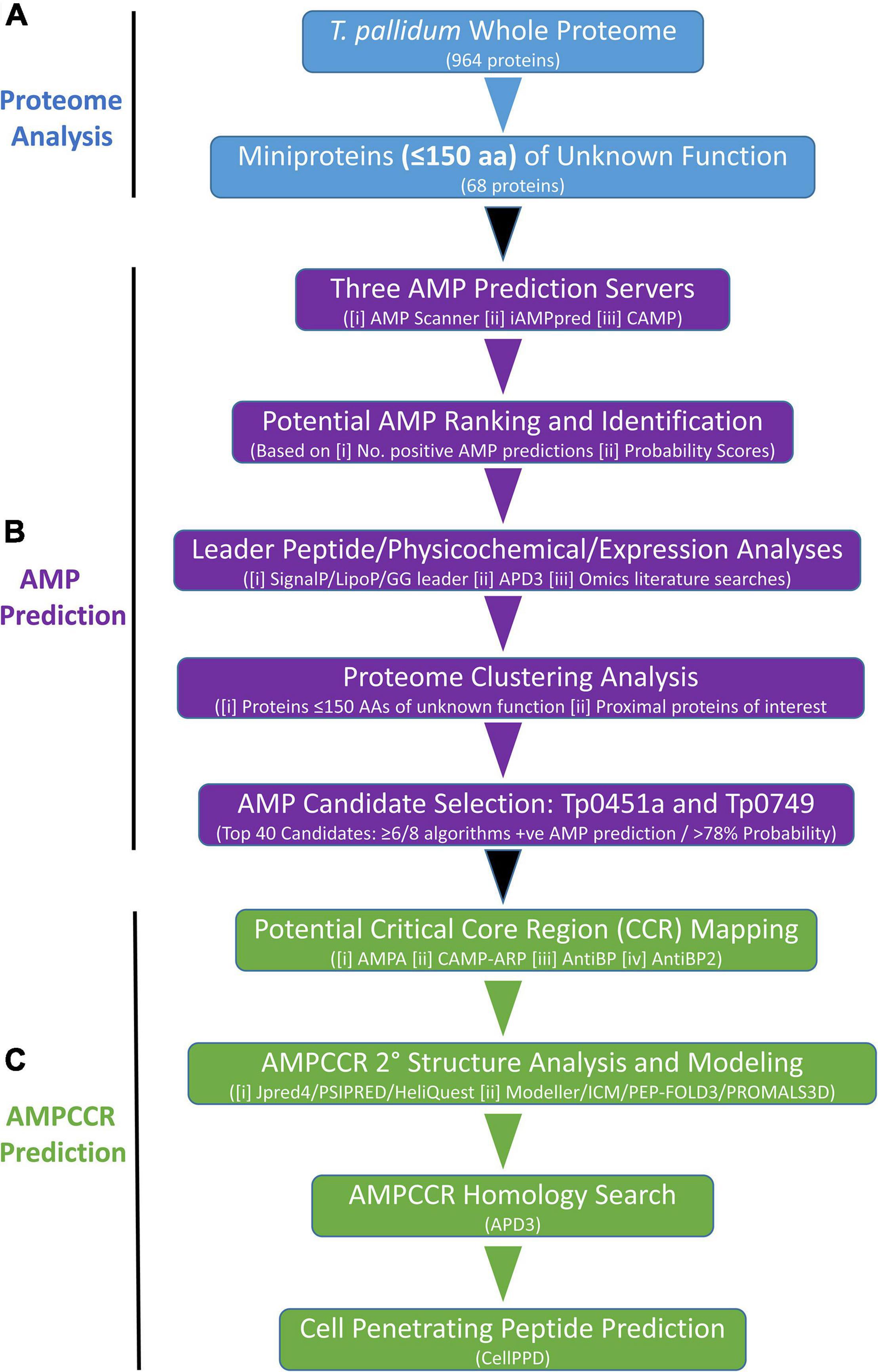
Figure 1. Bioinformatics pipeline for the identification of potential AMPs and AMPCCRs in the proteome of T. pallidum. (A) Proteome Analysis: The whole proteome (from NCBI genome database, July 2021 annotation) of T. pallidum was searched for proteins with no assigned function and 150 amino acids or less. (B) AMP Prediction: Complete amino acid sequences of 68 T. pallidum proteins (≤150 amino acids) of unknown function were submitted to three AMP prediction servers which allowed for ranking from most likely AMP to least likely AMP. Additional leader peptide, physicochemical, expression, and proteome clustering analyses facilitated the identification of two potential AMPs (Tp0451a and Tp0749) for further analyses. (C) AMPCCR prediction: A combination of critical core region (CCR) mapping, structural, homology, and cell penetrating peptide analyses were then performed to further confirm the initial AMP predictions generated by the three prediction servers and to identify putative antimicrobial peptide critical core regions (AMPCCRs) within the identified predicted AMPs, Tp0451a and Tp0749.
Bioinformatics Pipeline: AMP Prediction
The full-length amino acid sequences of all functionally-unannotated miniproteins identified in the above analyses were submitted to three AMP prediction servers: (i) AMP Scanner Version 2 (Deep Neural Learning Method for predicting antibacterial activity only)2 (Veltri et al., 2018), (ii) iAMPpred [Support Vector Machine (SVM) algorithms for predicting antibacterial, antiviral, and antifungal activities; machine learning method based on amino acid composition, physicochemical, and structural features]3 (Meher et al., 2017), and (iii) CAMP [SVM, Random Forest (RF), Artificial Neural Network (ANN), and Discriminant Analysis (DA) algorithms for predicting antimicrobial activity; machine learning method based on different physicochemical properties of proteins]4 (Waghu et al., 2016; Figure 1B). All T. pallidum miniproteins of unknown function were ranked from most likely AMP to least likely AMP based on the number of different server algorithms (out of eight total) that produced positive AMP predictions. Corresponding mean probability scores (1 [most likely prediction score]-0 [least likely prediction score]) were then used to rank all miniproteins within each positive AMP prediction class (Figure 1B).
Bioinformatics Pipeline: Leader Peptide, Physicochemical, and Proteome Clustering Analyses of Putative T. pallidum AMPs
To determine if any of the miniproteins of unknown function from T. pallidum contain potential Sec-dependent leader peptides (Sec/SP1 peptides), we used the signal peptide prediction servers, SignalP 5.05 (Almagro Armenteros et al., 2019) and LipoP 1.06 (Juncker et al., 2003). Manual searches of the T. pallidum miniproteins for the conserved double-glycine/glycine-alanine leader peptide motif (M[R/K]ELX3E[I/L]X2[I/V]XG[G/A]) that has been observed in AMPs from Gram-negative bacteria (Michiels et al., 2001; Dirix et al., 2004) were performed to identify proteins that contain Glycine-Glycine and/or Glycine–Alanine pairs within the first 31 residues of the N-terminus (Figure 1B). A multiple sequence alignment of the N-termini of each of the Glycine-Glycine and/or Glycine-Alanine-containing proteins was generated using Clustal Omega7 (Madeira et al., 2019). WebLogo8 (Crooks et al., 2004) was then used to generate a sequence logo for the identification of sites within the N-terminal residues of these proteins that exhibit homology to the conserved double-glycine/glycine-alanine leader peptide motif found in AMPs from Gram-negative bacteria (Michiels et al., 2001; Dirix et al., 2004). Physicochemical properties, including hydrophobicity, net charge, and presence/absence of cysteine residues, of the T. pallidum miniproteins of unknown function were analyzed using the APD3 database Antimicrobial Peptide Calculator and Predictor9 (Wang et al., 2016; Figure 1B). Tertiary structure modeling of functionally-unannotated proteins of interest (>150 amino acids) found to be located in close proximity to T. pallidum miniproteins identified above, was performed using the protein structure modeling server, Phyre210 (Kelley et al., 2015).
Bioinformatics Pipeline: Tp0451a and Tp0749 AMPCCR Prediction
Two identified AMP candidates [Tp0451a (accession number WP_014342798) and Tp0749 (accession number WP_010882194)] were further analyzed using our in-house bioinformatics pipeline (Figure 1C). This analysis was performed to help confirm the initial AMP predictions generated by the three prediction servers and to identify potential antimicrobial peptide critical core regions (AMPCCRs) within Tp0451a and Tp0749. This pipeline was comprised of the following four stages:
(i) Multi-server AMPCCR mapping
Full-length amino acid sequences of Tp0451a and Tp0749 were submitted to four servers: AMPA11 (Torrent et al., 2012), CAMP antimicrobial region prediction server (CAMP-ARP)12 (Waghu et al., 2016), AntiBP13 (Lata et al., 2007), and AntiBP214 (Lata et al., 2010). Potential AMPCCRs were identified in each protein based on high probability scoring regions (15–23 amino acid stretches) that were predicted by three or more servers.
(ii) Structure analyses and modeling
Secondary structure analyses of potential AMPCCRs were performed using the structure prediction servers Jpred 415 (Drozdetskiy et al., 2015) and PSIPRED 4.016 (Jones, 1999) and the alpha helix screening and physicochemical characterization server, HeliQuest17 (Gautier et al., 2008). Structure modeling of potential AMPCCRs based on PSIPRED secondary structure predictions were performed using the de novo peptide structure prediction server, PEP-FOLD 318 (Thevenet et al., 2012; Shen et al., 2014) and Swiss Model19 (Bienert et al., 2017; Waterhouse et al., 2018). All models were then vetted through the comparative protein structure modelling server, Modeller20 (Webb and Sali, 2016) and the homology modeling program, ICM (Molsoft L.L.C., CA, United States)21 (Cardozo et al., 1995), for their lowest normalized discrete optimized protein energy value (zDOPE) and GA341 score closest to 1. Comparative homology modeling using structure-based alignment was performed using PROMALS3D22 (Pei et al., 2008). Together, these structure prediction analyses were used to support the multi-server AMPCCR predictions through the identification of structural folds known to be important for AMP function and for facilitating subsequent peptide design via the prediction of intact secondary structure elements.
(iii) AMPCCR homology analyses
Amino acid homology searches using the APD3 database (see text footnote 9) (Wang et al., 2016) were employed to determine similarity and identity of predicted T. pallidum AMPCCRs with known and experimentally-validated AMPs and for the identification of short orthologous AMP sequences that would be otherwise missed using the NCBI BLAST tools23 (Altschul et al., 1990);
(iv) AMPCCR cell penetration prediction
AMPCCR cell penetrating abilities, a key functional feature of AMPs, were predicted using the CellPPD Protein Scanning Tool24 (Gautam et al., 2013) (peptide fragment length = 10; prediction method = SVM based with scores ranging from –1.0 to +1.0). To increase prediction stringency, the SVM threshold for positive cell penetrating peptide predictions was increased from the default threshold (0) to ≥0.1.
RNA Extraction and RT-PCR
RNA was isolated and purified from in vivo-harvested T. pallidum subsp. pallidum (Nichols strain) using Invitrogen TRIzol™ reagent (Thermo Fisher Scientific, MA United States) and the RNeasy mini kit (Qiagen, ON, Canada), according to the manufacturer’s instructions. RT-PCR was performed (after genomic DNA digestion/removal) using the orientation-specific RT-PCR sense (5’-aatgtcggctaccatcgctc) and antisense (5’-acgtgctctgccaattactgc) primers for tp0451a and the Invitrogen SuperScript™ IV First-Strand Synthesis System (Thermo Fisher Scientific), according to the manufacturer’s instructions. The negative control RT-PCR reaction did not include reverse transcriptase. PCR products were electrophoresed on agarose gels and visualized with ethidium bromide staining.
Peptide Synthesis
For the experimental validation of antimicrobial and immunomodulatory activity, four putative AMPCCR peptides (Table 1) from two T. pallidum miniproteins that were identified using our AMP bioinformatics prediction pipeline (Tp0451a_N, Tp0451a_C, Tp0749_N, and Tp0749_C), a cysteine-to-serine substituted version of Tp0749_C (Tp0749_C_C61S), and a cysteine-to-serine substituted version of Tp0451a_C (Tp0451a_C_C85S), were synthesized without chemical modifications using the PepPower™ solid state peptide synthesis (SSPS) platform at GenScript (NJ, United States). The known AMP, human cathelicidin LL-37 (Turner et al., 1998), the known bullfrog (Rana [Lithobates] catesbeiana) AMP, RaCa-2 (Li et al., 2022), a scrambled version of LL-37 (sLL-37), and a peptide (Tp0751_p5) from the T. pallidum adhesin Tp0751 (Cameron et al., 2005) (Table 1) were also synthesized via the same SSPS platform at GenScript, and used as positive (LL-37 and RaCa-2) and negative (sLL-37 and Tp0751_p5) controls in antimicrobial susceptibility and immunomodulation assays, as described below.

Table 1. Chemically synthesized peptides used in the current study.
Antimicrobial Susceptibility Assay—Broth Microdilution
The broth microdilution technique (Wiegand et al., 2008) was used to determine if the T. pallidum peptides are capable of exhibiting bacteriostatic [minimal inhibitory concentration (MIC) measurements] and/or bactericidal [minimal bactericidal concentration (MBC) measurements] activity. Bacterial suspensions were prepared by transferring bacterial colonies into MHB and resuspending using a vortex mixer to ensure complete suspension of any bacterial aggregates. Turbidity of the colony suspensions was adjusted spectrophotometrically to the required optical densities to achieve a turbidity equivalent to that of a 0.5 McFarland standard (1–2 × 108 CFU/mL) followed by dilution in MHB to achieve the standardized microbial inoculum of approximately 5 × 105 CFU/mL. Total viable counts (TVC) were routinely performed on all inoculum suspensions to ensure correct bacterial cell densities. The standardized bacterial suspensions were then incubated with two-fold serial dilutions of each peptide (dissolved in 11 μL of ultrapure sterile water; final peptide concentration range of 256 μg/mL–0.5 μg/mL) in Greiner polypropylene round bottom 96-well microtiter plates (Sigma-Aldrich, MO, United States). Each peptide was tested once per experiment, with a range of 3–9 independent experiments performed per peptide. Negative growth/sterility control wells contained bacterial growth media (100 μL) and the peptide solvent (11 μL of ultrapure sterile water). Positive growth control wells contained the standardized number of bacterial cells (100 μL of ∼ 5.0 × 105 CFU/mL) and peptide solvent (11 μL of ultrapure sterile water). Plates were incubated at 37°C for 16–24 h and MICs were determined using the unaided eye to identify the lowest concentration of AMP that inhibited visible growth of the tested bacterial species. If the sterility control well was turbid, the test was not considered valid. MBCs were determined by plating the entire content of the wells containing the peptide/bacteria mixture representing the MIC and the entire contents of the preceding wells containing two-fold and four-fold more concentrated AMP dilutions onto nonselective agar plates. Plates were incubated for 24 h at 37°C and MBCs were calculated as the percentage of bacteria killed at the different AMP concentrations tested (decrease in TVCs from the MBC plates compared to the initial bacterial suspension of ∼5 × 105 CFU/mL).
Antimicrobial Susceptibility Assay—Neisseria Modified Agar Dilution Method
The potential antimicrobial activity of the T. pallidum peptides against N. gonorrhoeae was determined using a modified agar dilution method. In this assay, N. gonorrhoeae colonies from GC chocolate agar plates were resuspended in MHB and the turbidity of the suspension was adjusted, as described above, to achieve the standardized microbial inoculum of approximately 5 × 105 CFU/mL. A two-fold serial dilution of the peptides (11 μL in sterile ultrapure water) was then prepared in wells 1–10 of a 96-well sterile polypropylene plate to obtain a dilution series corresponding to 10 times the required testing concentrations (2,560, 1,280, 640, 320, 160, 80, 40, 20, 10, 5 μg/mL). The bacterial suspension (100 μL) was dispensed into the wells containing the peptides. A negative growth/sterility control (well 12) contained bacterial growth media (100 μL) and the peptide solvent (11 μL of ultrapure sterile water). A positive growth control (well 11) contained the standardized number of bacterial cells (100 μL of ∼ 5.0 × 105 CFU/mL) and peptide solvent (11 μL of ultrapure sterile water). The 96-well plate was then incubated at 37°C in an atmosphere of 5% CO2 for 3 h to allow for peptide binding and antimicrobial activity. After the incubation period, an aliquot (20 μL) from wells 1–12 was removed and spotted onto the surface of a GC chocolate agar plate. N. gonorrhoeae spotted plates and TVC plates were incubated at 37°C in an atmosphere of 5% CO2 for 18–24 h. Following the incubation period, MICs were determined by identifying the lowest concentration of peptide that completely inhibited visible growth on the agar plate. To determine the bactericidal activity of peptides, total viable counts (TVCs) were also prepared on GC chocolate agar plates for the 3 h-incubated peptide/bacteria mixtures. These counts were compared with TVCs from the corresponding positive growth wells to give the percentage of bacteria killed by each of the peptides following the 3-h incubation.
Antimicrobial Susceptibility Assay – T. pallidum
An antimicrobial susceptibility assay was developed to assess the activity of the four treponemal peptides (Tp0451a_N, Tp0451a_C, Tp0749_N, and Tp0749_C), an equimolar mix of Tp0451a_N and Tp0451a_C, and the negative (Tp0751_p5) and positive (LL-37) control peptides, against T. pallidum. In vitro-grown T. pallidum (100μL; 1.0–1.2 × 106 Tp/mL), prepared as described above, were incubated with each peptide at three concentrations (4, 16, 64 μg/mL) or the Tp0451a_N/Tp0451a_C mix (21.6 μM; ∼85 μg/mL Tp0451a_N and ∼64 μg/mL Tp0451a_C) at 34°C in an atmosphere of 93.5% nitrogen, 5% carbon dioxide, and 1.5% oxygen. Darkfield microscopy was used to monitor T. pallidum viability by counting motile treponemes at 0, 1, 2, and 4 h post co-incubation. For each viability measurement, at least 50 treponemes were observed for each sample at each time point.
THP-1 Monocyte Culture and Macrophage-Like Differentiation
Human THP-1 (ATCC TIB-202) monocytes (American Type Culture Collection, VA, United States) were cultured and maintained in 5% CO2 at 37°C in RPMI-1640 medium (Gibco, Life Technologies, ON, Canada) supplemented with 10% (v/v) fetal bovine serum (FBS) (Fisher Scientific, ON, Canada), 0.05 mM 2-mercaptoethanol (BME) (Sigma-Aldrich, ON, Canada), penicillin (100 units), and streptomycin (0.1 mg/mL) (Sigma-Aldrich, ON, Canada) (hereafter referred to as “complete RPMI-1640 medium”). Cells were passaged at a density of 8 × 105 cells/mL to a maximum of 15 passages and re-seeded at 3 × 105 cells/mL for maintenance. For differentiation into plastic-adherent macrophage-like cells, THP-1 monocytes were seeded at a density of 3 × 105 cells/mL in T75 tissue culture flasks (Fisher Scientific, Ottawa, ON, Canada) and stimulated with a low concentration (25 ng/mL) of phorbol-12-myristate-13-acetate (PMA) (Sigma-Aldrich, ON, Canada) in complete RPMI-1640 medium for 48 h in 5% CO2 at 37°C. The PMA-mediated differentiation method results in the generation of cells with phenotypic characteristics that are similar to human peripheral blood mononuclear cell (PBMC) monocyte-derived macrophages; they are adherent, larger, more phagocytic, are less proliferative, and exhibit cell surface markers that are characteristic of macrophages (Chanput et al., 2014).
Following incubation with PMA, light microscopy was used to ensure the differentiated cells were adherent, and exhibited morphological changes consistent with PBMC monocyte-derived macrophages. Non-adherent cells were then removed by washing with sterile, calcium- and magnesium-free phosphate-buffered saline (PBS) (ThermoFisher Scientific, MA United States). Plastic-adherent macrophage-like cells were detached by a three- to five-minute treatment with trypsin-EDTA (0.05%) (ThermoFisher Scientific, MA, United States) and physical agitation. The macrophage-like cells were then centrifuged at 1,500 rpm using a Sorvall Model STR04 centrifuge (ThermoFisher Scientific, MA United States) for 5 min at room temperature and seeded in PMA-free complete RPMI-1640 media, as described below.
AMP Stimulatory and AMP/IL-32γ Co-stimulatory Immunomodulation Assays
THP-1 monocytes and THP-1 macrophage-like cells were seeded into the wells of 12-well plates (Thermo Fisher Scientific, ON, Canada) in complete RPMI-1640 medium (1 mL) at a density of 0.5 × 106 cells/mL. Following seeding and prior to stimulation, macrophage-like cells were rested overnight. For AMP alone stimulation, monocytes or macrophages were exposed for 24 h at 37°C in 5% CO2 to either (i) no stimulation (negative control for baseline cytokine production), (ii) lipopolysaccharide [LPS; from S. enterica serovar Typhimurium (Sigma-Aldrich, ON, Canada)] (positive control for cytokine production; final concentration 1.0 μg/mL), or (iii) the test peptides (control peptides and potential T. pallidum AMPs listed in Table 1; final concentration 25 μg/mL). For AMP/IL-32γ co-stimulation, 20 ng/mL IL-32γ (R&D Systems, MN, United States) in fresh complete RPMI-1640 medium was added to the rested macrophages. IL-32γ stimulation was immediately followed by co-stimulation by the addition of the test peptides at a final concentration of 25 μg/mL. Macrophage cells left unstimulated or stimulated with IL-32γ alone were used as negative and positive controls, respectively. Cells were stimulated for 24 h at 37°C in 5% CO2. Following stimulation, monocytes and macrophage cells were centrifuged at 1,500 rpm using a Sorvall Model STR04 centrifuge (ThermoFisher Scientific, MA, United States) for 5 min at room temperature and the cell-free supernatant was stored at -80°C prior to quantification of cytokine levels, as described below.
THP-1 Monocyte and Macrophage Cytokine Expression Analyses
The BD™ Cytometric Bead Array (CBA) system (BD Biosciences, CA, United States) was used to quantify the expression of tumor necrosis factor (TNF), MCP-1, IL-6, IL-8, IL-10, and IL-1β according to manufacturer’s instructions. For statistical analyses, data were analyzed for normality using a D’Agostino-Pearson omnibus normality test and a Shapiro–Wilk normality test. An ordinary one-way ANOVA followed by Dunnett’s multiple comparisons test was used to assess differences between three or more groups of normally distributed data. A Kruskal–Wallis test followed by Dunn’s multiple comparisons test was used to assess differences between three or more groups of data that were not normally distributed.
Results
Identification of T. pallidum Miniproteins of Unknown Function
Although the size of AMPs can vary greatly, ranging from approximately five amino acids to several hundred amino acids, a search of the AMP database, APD325 (Wang et al., 2016), indicated that 97% of the 3324 AMPs listed at the time of analysis fall within the 10–150 amino acid size range. With this knowledge, we sought to identify potential T. pallidum AMPs by manually searching the whole proteome of T. pallidum and filtering for proteins containing 150 amino acids or less. This resulted in the identification of 151 miniproteins (≤150 amino acids) (Supplementary Table 1), representing ∼16% of the T. pallidum proteome. We then filtered for miniproteins with no assigned function or weak/incomplete annotated functions and for miniproteins with a potential AMP-related function, resulting in the identification of 68 proteins (Supplementary Table 2), representing 7% of the T. pallidum proteome. Genes corresponding to four of these 68 proteins (Tp0039, Tp0130, Tp0451a, and Tp0867) were not included in the latest annotation of the T. pallidum proteome (Nichols strain NC_021490), however, all four genes have been shown to be expressed at the transcript level (Smajs et al., 2005 and current study) justifying their inclusion in this study (Supplementary Table 2). Sixty-seven of the 68 miniproteins of unknown function that were identified in T. pallidum were annotated in the published proteome from July 2021 as either “hypothetical proteins,” “DUF (Domains of Unknown Function) domain-containing proteins,” or as proteins with motifs/domains that do not provide enough insight to confidently indicate potential protein functions (e.g., helix-turn-helix domain-containing proteins, DNA- or RNA-binding proteins, zinc ribbon domain-containing protein) (Supplementary Table 2). One of the 68 miniproteins was annotated as a putative CPBP family intramembrane metalloprotease (TPANIC_RS05485), some of which may be involved in AMP processing (Pei et al., 2011). However, this 91-amino acid treponemal protein is at least two-four-fold smaller than other bacterial CPBP proteins, is only predicted to contain one transmembrane segment (unlike the four or more present in known CPBP proteins), and does not contain the four conserved sequence motifs required for proteolytic activity that are found in other CPBP proteins (Pei et al., 2011). In light of these findings, Tp_RS05485 was included in the list of 68 miniproteins for further bioinformatics analyses (Supplementary Table 2).
Prediction Analyses for the Identification and Ranking of Putative T. pallidum AMPs
Full-length amino acid sequences of all 68 miniproteins identified in the analyses described above were submitted to three AMP prediction servers. Prediction data were then used to rank the 68 treponemal proteins from most likely AMP (ranking 1/68) to least likely AMP (68/68) depending on (i) how many of the eight server algorithms produced positive AMP predictions and (ii) the mean probability scores from each of the AMP predictions for each protein. In summary, 45 high-priority T. pallidum AMP candidates predicted by at least four of the eight algorithms were assigned mean probability scores of at least 0.505 (50.5% probability) (Figure 2 and Supplementary Table 3). AMP prediction results for all 68 T. pallidum miniproteins and corresponding probability scores from each of the AMP prediction servers are listed in Supplementary Table 3.
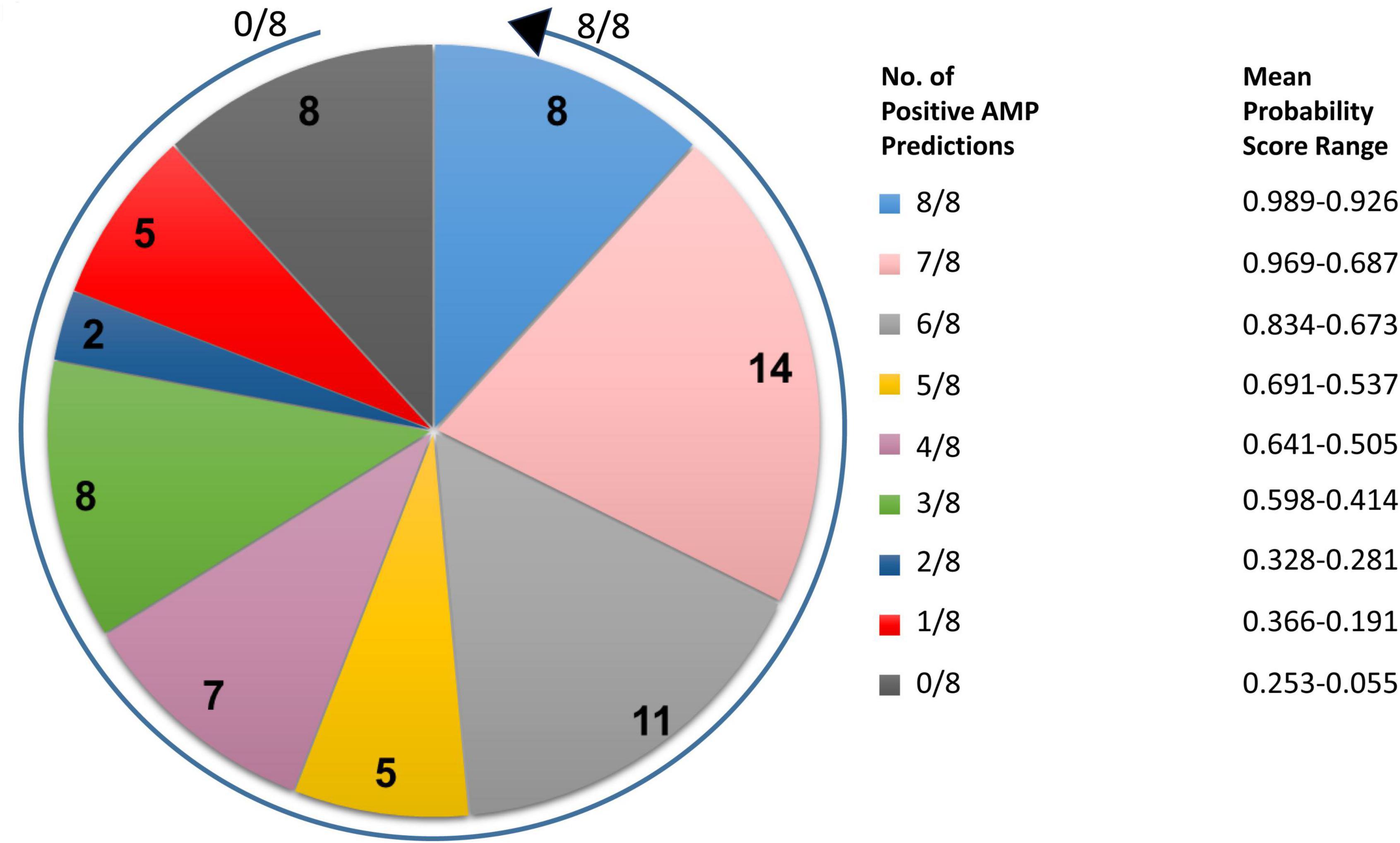
Figure 2. Distribution of the number of positive AMP predictions and corresponding mean probability score ranges for T. pallidum miniproteins of unknown function. Pie chart indicating the number of positive AMP predictions following submission of the full-length amino acid sequences of 68 T. pallidum miniproteins (≤150 amino acids) to three AMP prediction servers (eight AMP prediction algorithms total). The arrow begins at the 0/8 positive AMP prediction class and finishes at the 8/8 positive AMP prediction class. The corresponding mean probability score range for all proteins from each positive AMP prediction class is also shown.
Leader Peptide Analyses of T. pallidum Miniproteins
Many bacterial AMPs are synthesized as inactive preproteins with N-terminal leader/signal peptides whose presence and cleavage are required for export and activation, respectively. These include signal peptides recognized by the general secretory (Sec) pathway [Sec-dependent signal peptides (Sec/SP1 peptides)] (Leer et al., 1995; Chiorean et al., 2018) and Sec-independent double-glycine/glycine-alanine (GG/GA) leader peptides that have been documented in AMPs from both Gram-positive and Gram-negative bacteria (Havarstein et al., 1994; Oman and van der Donk, 2010). The signal peptide prediction servers, SignalP and LipoP, were used to search for the presence of Sec-dependent SP1 signal peptides, which predicted the presence of Sec/SP1 signal peptides in only four of the 68 miniproteins (Supplementary Table 4). Manual searches of the 68 T. pallidum AMP candidates for the conserved double-glycine/glycine-alanine leader peptide motif identified 24 proteins that contain Glycine-Glycine and/or Glycine-Alanine pairs within the first 31 residues of the N-terminus (Supplementary Figure 1). WebLogo analysis of the 24 proteins identified an N-terminal region with similarity to the double-glycine/glycine-alanine leader peptide motif from AMPs from Gram-negative bacteria, suggesting the presence of a similar secretion/activation recognition signal in T. pallidum candidate AMPs (Supplementary Figure 2).
Physicochemical Analyses of T. pallidum Miniproteins
Physicochemical properties known to be important for AMP function were calculated using the APD3 online AMP calculator and predictor tool. Consistent with the high content of arginine and lysine residues in AMPs, the top 22-ranking potential T. pallidum AMPs were found to have mean net charges at pH 7.0 of 5.56 (8 proteins with 8/8 positive AMP predictions) and 7.91 (14 proteins with 7/8 positive AMP predictions) (Figure 3A and Supplementary Table 4). Although no trend was observed between AMP likelihood rankings and the percentage of hydrophobic amino acids found within this group of proteins, the mean hydrophobic residue content of all 68 miniproteins was high (43.1%), with 66/68 proteins comprised of more than 30% hydrophobic residues (Figure 3B and Supplementary Table 4). Also observed was a trend in AMP likelihood rankings and the number / percentage of cysteine residues per protein. In general, cysteines were found to be more common in higher-ranking predicted AMPs (Figure 3C and Supplementary Table 4). In comparison to an average cysteine content of ∼1.9% found in all T. pallidum proteins [calculated from the Nichols strain (NC_021490) proteome], the top eight-ranking predicted AMPs (8/8 positive AMP predictions) contained on average almost three-fold more cysteines (mean cysteine residue content = 5.31%). Interestingly, the cysteine-rich nature of these T. pallidum miniproteins is shared with distinct classes of eukaryotic (Simmaco et al., 1994; Fahrner et al., 1996; Shafee et al., 2016) and prokaryotic (Baindara et al., 2017; Sugrue et al., 2020) AMPs, and thus may represent an important physicochemical property for protein structure and/or function.
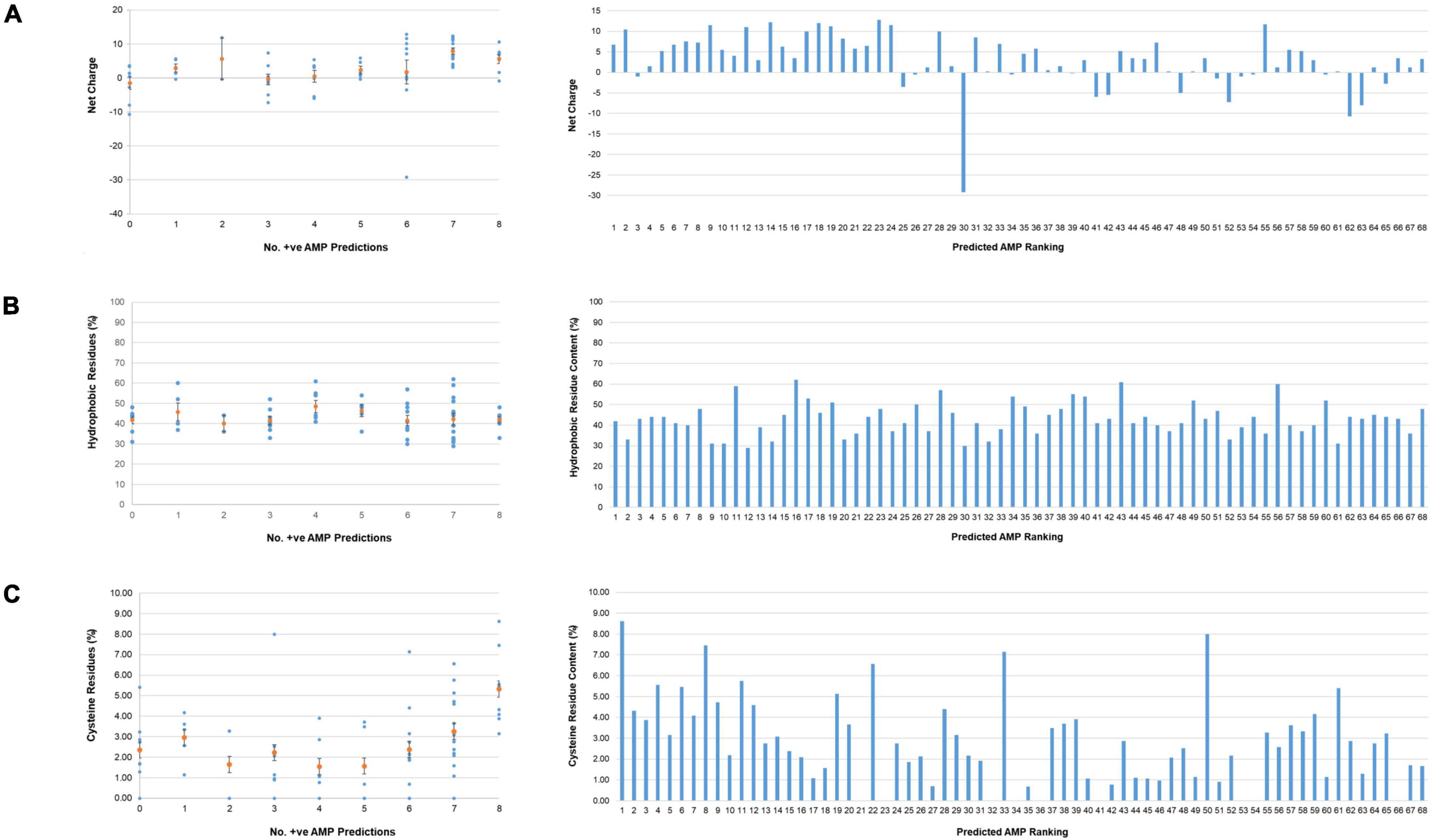
Figure 3. Physicochemical properties of T. pallidum miniproteins of unknown function. Physicochemical properties known to be important for AMP function were calculated for the 68 T. pallidum proteins (≤150 amino acids) of unknown function using the APD3 online AMP calculator and predictor tool. Left: Scatter plots depicting (A) net charges, (B) hydrophobic residue content, and (C) cysteine residue content (orange circles represent the mean values, +/- standard error) of all proteins from each positive AMP prediction class. Right: Bar graphs showing (A) net charge, (B) hydrophobic amino acid content, and (C) cysteine residue content of all 68 miniproteins (≤150 amino acids) from predicted AMP ranking 1–68. No. +ve AMP Predictions equates to the number of AMP prediction programs that assigned the miniprotein as an AMP.
Expression Analyses of T. pallidum Miniproteins
Most of the 68 T. pallidum miniproteins from the current study are annotated in the published T. pallidum proteome as “hypothetical” proteins. However, DNA microarray-based analysis of the T. pallidum transcriptome following experimental rabbit infection (Smajs et al., 2005) demonstrated that 56/68 genes encoding these functionally uncharacterized proteins are expressed at the transcriptional level (Supplementary Table 5). Transcripts from most of the other 12 genes were not searched for in the study (Smajs et al., 2005) as they were not annotated in the T. pallidum genome at the time the study was performed. In addition, peptides from 15/68 miniproteins were detected in mass spectrometry-based proteomics studies of rabbit infections (McGill et al., 2010; Osbak et al., 2016), including the protein with the highest level of expression in the Osbak and colleagues study, Tp0214 (Supplementary Table 5). The use of trypsin for T. pallidum protein digestion in the two mass spectrometry studies may have contributed to the low number of miniproteins detected in these experiments. Given that miniproteins are small and contain high numbers of lysine and arginine residues, trypsin treatment, which results in cleavage after lysine and arginine residues, would be expected to cleave the miniproteins into many small peptides. Many of these peptides would be below the size detection limit, a major limiting factor for protein identification in mass spectrometry studies.
Proteome Clustering of T. pallidum Miniproteins
Bacterial genomic analyses show genes with related functions tend to form gene clusters (Tamames et al., 1997). To determine the spatial arrangement of all 68 miniproteins (≤150 amino acids) of unknown function within the T. pallidum proteome, each protein was arranged from the lowest (Tp0004) to the highest (Tp1032) locus tag number and clusters comprised of at least two miniproteins separated by five or less intervening proteins were identified. Forty-three of the 68 miniproteins (63%) were found to be located within one of 17 clusters, with 23 of the 43 proteins located in clusters comprised of at least three miniproteins of unknown function (Supplementary Table 6 and Figure 4A). Twenty of the top 30-ranking predicted AMPs (67%) were found to be located within 13 miniprotein clusters, with 11 of the 20 proteins located in clusters containing at least three miniproteins of unknown function (Supplementary Table 6 and Figure 4B). The top-ranking predicted AMP (Tp_RS02215) was found in a three-miniprotein cluster including the 8th-ranking predicted AMP (Tp0451a). Interestingly, analysis of the surrounding proteome identified two annotated proteins of note including the outer-membrane inner-leaflet-associated lipoprotein, Tp0453, which contains multiple outer membrane-inserting amphipathic alpha helices that result in membrane bilayer destabilization and enhanced permeability (Hazlett et al., 2005; Luthra et al., 2011). Also included in this region is Tp0454; structure modeling of Tp0454 using Phyre2 predicted tertiary structure similarity to several response regulators (Supplementary Table 6), including the DNA-binding response regulator, PhoP, from the PhoP-PhoQ two-component system that is a central regulator for AMP resistance in Gram-negative bacteria (Brodsky and Gunn, 2005). Proteome functional annotation analyses and Phyre2 structure modeling of open reading frames located close to other putative AMPs identified several additional potential homologs and structural orthologs with potential functions that are consistent with AMP secretion, activation, transport, and self-immunity (Supplementary Table 6), including the ORFs Tp0405 and Tp0688 that have been previously annotated as self-immunity proteins (Fraser et al., 1998). The close spatial arrangement of the miniproteins, in particular the high-ranking predicted AMPs, together with the observed proximity of potential AMP accessory proteins in the proteome of T. pallidum is consistent with the concept that functionally-related genes have a tendency to form clusters within bacterial genomes (Tamames et al., 1997).
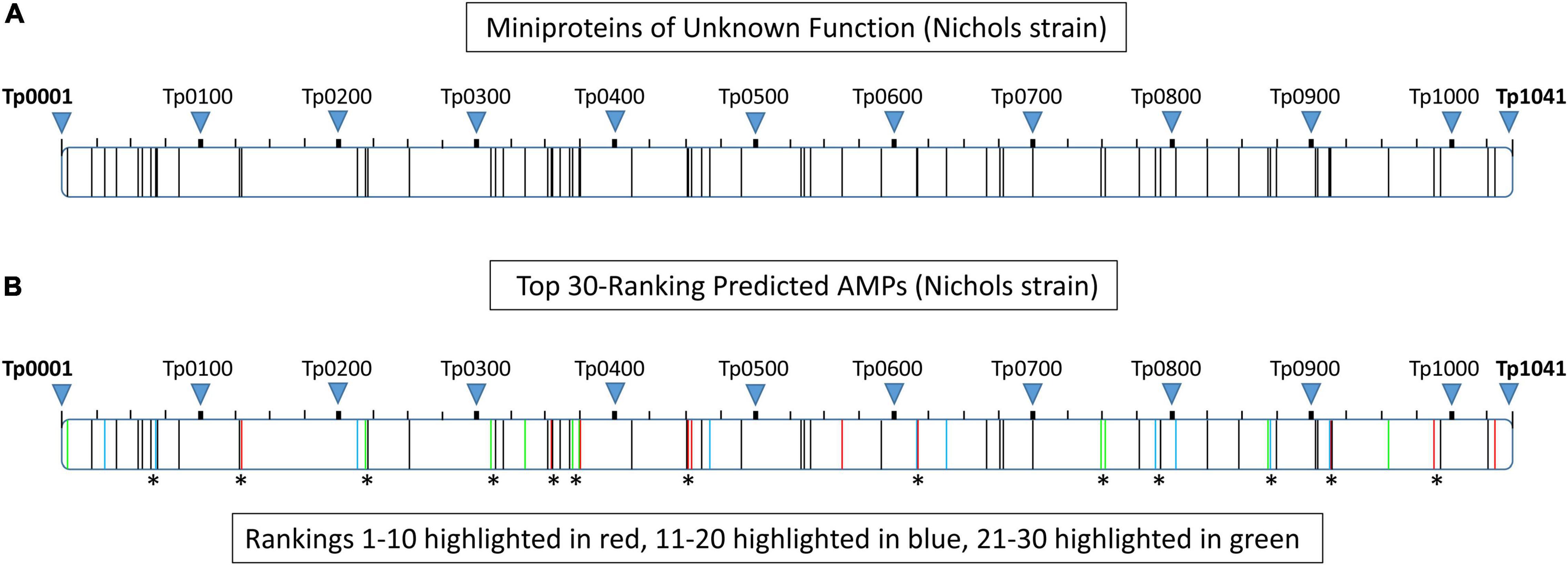
Figure 4. Proteome clustering of T. pallidum (Nichols strain) miniproteins of unknown function. Each of the T. pallidum (Nichols strain) proteins (≤150 amino acids) of unknown function were arranged from the lowest (Tp0004) to the highest (Tp1032) locus tag number within the proteome (Tp0001–Tp1041) and clusters comprised of at least two miniproteins separated by five or less intervening proteins were identified. (A) Schematic depicting the spatial arrangement of all miniproteins (≤50 amino acids) of unknown function within the T. pallidum (Nichols strain) proteome. The location of each protein within the proteome is represented by a vertical black line. (B) Schematic showing the spatial arrangement of the 30 top-ranking predicted AMPs within the T. pallidum (Nichols strain) proteome. The location of proteins corresponding to rankings 1–10, 11–20, and 21–30 are shown in red, blue, and green, respectively. Asterisks denote the location of 20 of the top 30-ranking predicted AMPs that are found to be located within 13 miniprotein clusters.
AMP Candidate Selection: Tp0451a and Tp0749
Two AMP candidates, Tp0451a (accession number WP_014342798) and Tp0749 (accession number WP_010882194), were selected for the identification of potential AMPCCRs, the important minimalistic functional regions of AMPs. Tp0451a was selected as it (i) is one of the top-ranking predicted AMPs (8/8 positive AMP predictions, mean probability score of 92.6%) (Supplementary Table 3), (ii) possesses classical AMP properties (high content of positively-charged and hydrophobic amino acid residues) (Supplementary Table 4), (iii) is clustered in the proteome with several other potential AMPs/related proteins, as described above, and (iv) tp0451a is expressed at the transcript level, as described below. Although Tp0749 is a lower-ranking predicted AMP (ranked 24/68, 6/8 positive AMP predictions, mean probability score of 78.4%) (Supplementary Table 3), it was selected for further analyses as it (i) is highly positively-charged and contains high hydrophobic content, consistent with pore-forming AMPs (Supplementary Table 4), (ii) was identified in preliminary bioinformatics analyses as having clearly defined potential critical core regions, indicative of future success in AMPCCR design and synthesis, (iii) is known to be expressed at the transcript level (Smajs et al., 2005), unlike several of the higher-ranking predicted AMPs (Supplementary Table 5), (iv) is the second highest expressed ORF at the transcript level in the top-30 ranking T. pallidum miniproteins (Smajs et al., 2005) (Supplementary Table 5), and (v) of particular importance, it is one of only six minproteins within the top-30 ranking predicted AMPs whose expression has been detected at the protein level in experimental rabbit infections (Osbak et al., 2016) (Supplementary Table 5). To date, protein expression of all eight miniproteins from the top-eight ranking predicted AMPs (8/8 positive AMP predictions, mean probability score range 98.9–92.6%) has not been demonstrated in rabbit models of infection (McGill et al., 2010; Osbak et al., 2016), and only three of the top-eight ranking predicted AMPs have been shown to be expressed at the transcript level (Smajs et al., 2005). The strong experimental evidence confirming expression of Tp0749 at both the RNA and protein levels in rabbit infections increased the prioritization of this predicted treponemal AMP over higher-ranking predicted AMPs for further bioinformatics and functional characterization studies.
Reverse Transcription-PCR Analysis of tp0451a
To confirm expression of tp0451a, we analyzed RNA isolated from T. pallidum by reverse transcription PCR (RT-PCR) using sense and antisense primers. When reverse transcriptase was present (RT+), the primer pair amplified a 198 base pair product, matching a similarly sized amplicon generated from T. pallidum genomic DNA (Supplementary Figure 3 lanes 2 and 4, respectively). In comparison, only a very faint amplicon was detected when reverse transcriptase was omitted from the RT-PCR reaction (RT-) (Supplementary Figure 3 lane 3), indicating that the 198 base pair product from the RT+ reaction was amplified from RNA and not contaminating DNA. Together with the previous finding that showed expression of Tp0749 at the protein level (Osbak et al., 2016), this result allowed us to proceed with investigations into potential AMP activity in T. pallidum by focusing on two miniproteins, Tp0451a and Tp0749, that are known to be expressed at either the transcript or protein level.
Bioinformatic Identification of Potential Critical Core Regions in Two Putative T. pallidum AMPs
The first step for mapping AMPCCRs in Tp0451a and Tp0749 involved using four prediction servers [AMPA (one algorithm), CAMP (three algorithms), AntiBP (three algorithms), and AntiBP2 (one algorithm)] to identify the amino acid boundaries of potential antimicrobial active regions (critical core regions, CCRs) within the two T. pallidum proteins based on clusters of high probability scoring regions predicted by at least three of the four servers. For both Tp0451a and Tp0749, two potential active regions were identified in the N-and C-termini of each protein (Figures 5A,B).
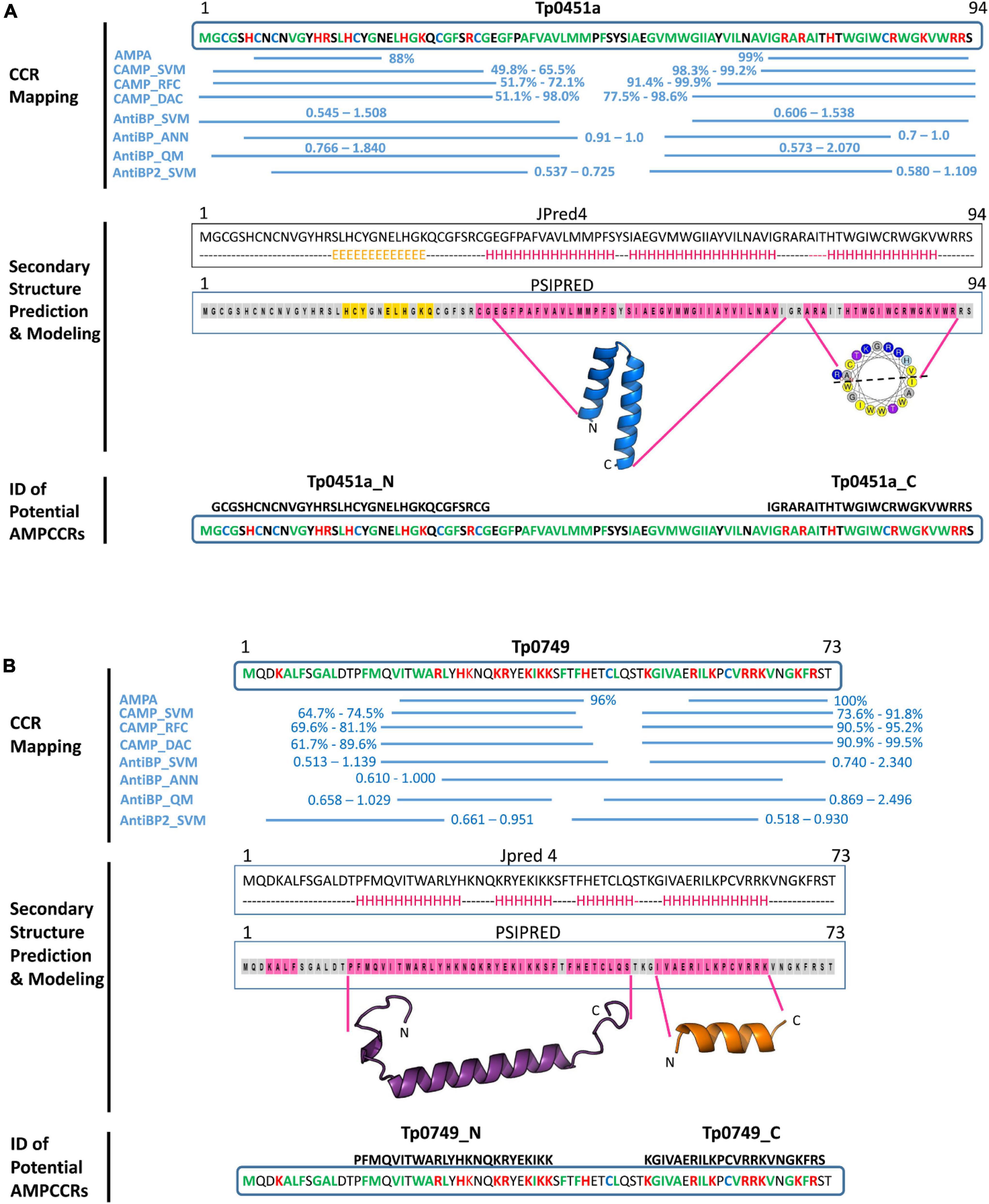
Figure 5. In silico identification of potential T. pallidum AMPCCRs. The critical core regions of two candidate AMPs, (A) Tp0451a and (B) Tp0749, were predicted using our bioinformatics pipeline. The first step of the pipeline involved CCR mapping (A,B, top): four prediction servers [AMPA (one algorithm), CAMP (three algorithms), AntiBP (three algorithms), and AntiBP2 (one algorithm)] were used to identify the amino acid boundaries of potential antimicrobial active regions (critical core regions, CCRs). High probability/scoring regions predicted by at least three of the four servers are shown with their corresponding probabilities (AMPA and CAMP algorithms) or scores (AntiBP and AntiBP2 algorithms). Hydrophobic residues: green; Positively-charged residues: red; Cysteines: blue. In the second step of the pipeline, secondary structure analyses and modeling were performed (A and B panels, middle): secondary structure analyses of the full-length proteins were performed using Jpred 4 (H: alpha helix; E: beta strand; dashed line: coiled) and PSIPRED (pink highlight: alpha helix; orange highlight: beta strand; gray highlight: coiled). HeliQuest was used to generate helical wheel diagrams for potential alpha helices (yellow: hydrophobic residues; purple: serine or threonine; blue: positively charged residues: gray: glycine or alanine). Structure modeling using Modeller generated a confident model for the central region of Tp0451a (residues E36-I71), but confident models were not generated for the N- or C-terminal regions. Structure modeling using a combination of PEP-FOLD-2, Swiss-Model, Molsoft ICM, and Modeller generated models for the N- and C-terminal regions of Tp0749 (residues P14-S48 and I52-K65, respectively). Together, these findings allowed for the identification of two potential critical core regions within the N-terminus (Tp0451a_N and Tp0749_N) and C-terminus (Tp0451a_C and Tp0749_C) of Tp0451a and Tp0749 (A and B panels, bottom).
In the second step of our pipeline, secondary structure analyses and structure modeling of the two proteins and their identified potential active regions were performed. For Tp0451a, Jpred 4, and PSIPRED analyses predicted a predominantly coiled/beta strand structure that corresponded to the N-terminal predicted active region and an alpha helical structure that corresponded to the C-terminal predicted active region (Figure 5A). HeliQuest analysis showed that the predicted C-terminal alpha helix exhibited amphipathic properties (Figure 5A), a common structural characteristic in AMPs that is important for membrane integration and pore formation. Structure modeling using a combination of PEP-FOLD 2, Swiss Model, Modeller and Molsoft ICM was unable to generate high confidence models for either the N- or C-terminal regions that correspond to the predicted antimicrobial active regions. However, a robust model was generated for the intervening central region (residues E36-I71) (Figure 5A and Supplementary Figure 4). In agreement with the secondary structure predictions, this central region was modeled as two hydrophobic alpha helices. The structure prediction analyses were consistent with the multi-server AMPCCR mapping predictions by defining potential structural elements, one of which is important for AMP function, that corresponded to high-scoring predicted active regions. Together, these findings allowed for the identification of two potential critical core regions within the N- (Tp0451a_N) and C-terminus (Tp0451a_C) of Tp0451a (Figure 5A).
For Tp0749, both Jpred 4 and PSIPRED analyses predicted alpha helices that corresponded to the N- and C-terminal predicted active regions (Figure 5B). Consistently, a high confidence (92%) alpha helix was modeled for the N-terminal region that was predicted to exhibit antimicrobial activity (Figures 5B, 6A). Importantly for potential AMP function, this modeled N-terminal alpha helix was also shown to be amphipathic with one face of the helix rich in positively-charged residues and the opposing face rich in hydrophobic residues (Figures 6A,B). In addition, a structure-based alignment using PROMALS3D of the Tp0749 N-terminal alpha helix model and a solved structure from the known AMP, human cathelicidin LL-37 (PDB:5NMN), predicted structural similarity between the two peptides (RMSD—0.31 Å over 19 Cα atoms) and conservation of 3/6 positively charged residues involved in binding target cell membrane lipids (Sancho-Vaello et al., 2017) (Figure 6C). In agreement with secondary structure predictions, a lower confidence (69%) partially amphipathic alpha helix model was also modeled for the C-terminal predicted antimicrobial active region (Figures 5B, 6D,E). The combined approach of multi-server AMPCCR mapping, secondary structure prediction, and modeling allowed for the identification of two potential critical core regions located in the N- (Tp0749_N) and C- (Tp0749_C) terminus of Tp0749 (Figure 5B).
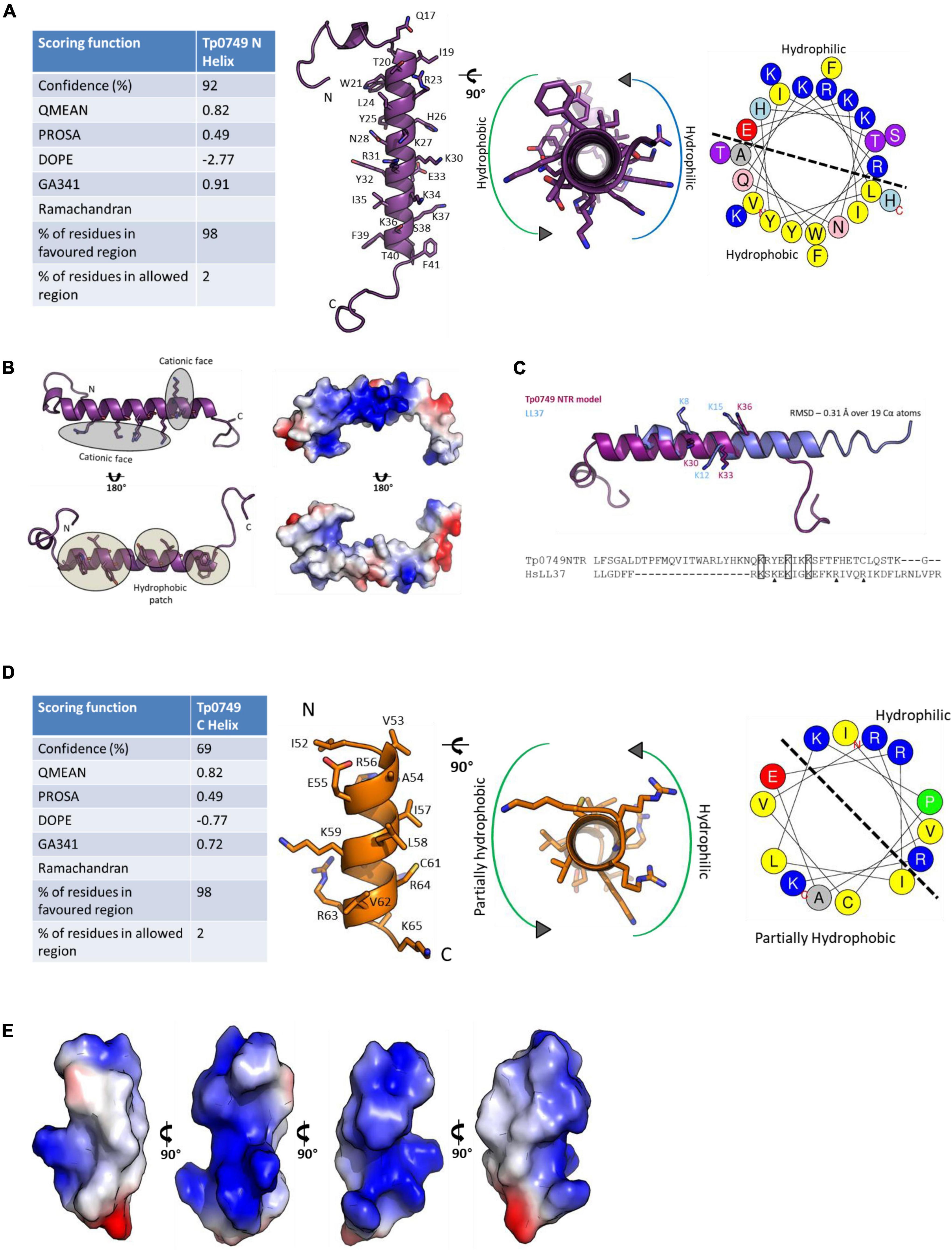
Figure 6. Structure modeling of the candidate T. pallidum AMP, Tp0749. (A, Left): Table showing the scoring functions of the Tp0749 N-terminal model (residues P14-S48) generated by a combination of PEP-FOLD-2, Swiss-Model, Molsoft ICM, and Modeller. (Middle) Model ribbon structure of Tp0749 residues P14-S48 and rotated view showing amphipathicity. (Right) Helical wheel schematic of Tp0749 (P14-S48) generated using HeliQuest. Dashed line separates the hydrophilic/polar and hydrophobic/non-polar faces of the predicted alpha helix. (B) Ribbon and surface/charge distribution images of the Tp0749 (P14-S48) model showing one face of the alpha helix rich in positively-charged/polar residues (blue) and the opposing face rich in hydrophobic/non-polar residues (white). Red: negatively-charged/polar residues. (C) PROMALS3D was used to generate a structure-based comparative model of the Tp0749 N-terminal alpha helix model (P14-S48) using the structure of the known AMP, human cathelicidin LL-37 (PDB:5NMN) as a template. RMSD—0.31 Å over 19 Cα atoms and 3/6 positively charged residues involved in binding target cell membrane lipids are conserved (indicated by rectangles; triangles show non-conserved residues). (D) Left: Table showing the scoring functions of the Tp0749 C-terminal model (residues I52-K65) generated by using a combination of PEP-FOLD-2, Swiss-Model, Molsoft ICM, and Modeller. (Middle) Model ribbon structure of Tp0749 residues I52-K65 and rotated view showing partial amphipathicity (red: negatively charged/polar residue side chains; blue: positively charged/polar residue side chains; yellow: cysteine residue side chain). (Right) Helical wheel schematic of Tp0749 (I52-K65) generated using HeliQuest. Dashed line separates the hydrophilic/polar and partially hydrophobic/non-polar faces of the predicted alpha helix. (E) Surface and charge distribution views of Tp0749 (I52-K65) (red: negatively charged/polar residues; blue: positively charged/polar residues; white: hydrophobic/non-polar residues).
To help further resolve the potential active regions identified above, the four candidate AMPCCRs were then analyzed for similarity with known, experimentally-validated AMPs from the APD3 database and for their predicted cell penetrating capabilities using CellPPD. Amino acid sequence-based homology searches identified similarities of each of the four AMPCCRs with established AMPs with homologies ranging between 37 and 44% (Table 2). CellPPD analysis also predicted that three of the four potential AMPCCRs with predicted amphipathic alpha helices (Tp0451a_C, Tp0749_N, and Tp0749_C) (Figures 5A, B, 6) contain peptide stretches that may have the ability to penetrate cell membranes (Table 3), a key functional feature of AMPs. The positive control peptide (LL-37) was also predicted to contain cell penetrating peptide regions, unlike the negative control peptide, Tp0751_p5 (Table 3). Together, these results bolstered the initial AMP predictions generated by the four prediction servers and identified two potential AMPCCRs within each of Tp0451a and Tp0749. These putative functionally-active core regions were prioritized for peptide synthesis and AMP functional characterization studies.
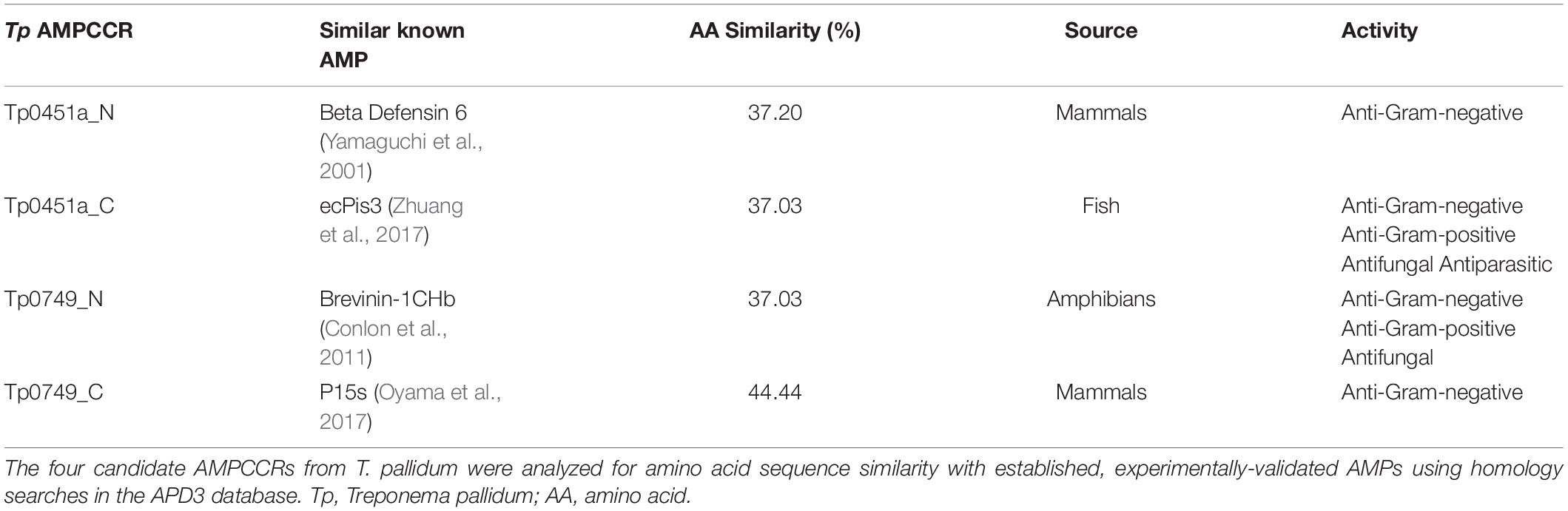
Table 2. Known AMPs with the highest similarity to four T. pallidum candidate AMPCCRs.

Table 3. Treponema pallidum candidate AMPCCRs with predicted cell penetrating abilities.
In vitro Antimicrobial Activity of T. pallidum AMPCCR Candidates
To evaluate the potential antimicrobial activity of the four predicted T. pallidum AMPCCRs identified via our bioinformatics pipeline, synthetic peptides were produced and antimicrobial susceptibility assays were performed to test for bacteriostatic and bactericidal activities against a panel of biologically and clinically relevant Gram-negative and Gram-positive bacteria. In broth microdilution assays, all four candidate AMPCCRs were active against M. smegmatis (Table 4). Consistent with our bioinformatics pipeline analyses, the three T. pallidum peptides with predicted amphipathic alpha helices (Tp0451a_C, Tp0749_N, and Tp0749_C) (Figures 5A,B, 6) and highest AMPCCR mapping scores (Figures 5A,B) all exhibited robust anti-mycobacterial activity, unlike the lower-scoring Tp0451a_N. The lack of activity in the latter peptide aligns with the observation that this was the only peptide that lacked predicted (amphipathic) helical structure (Figure 5A). With the exception of Tp0451a_N, the treponemal peptides exhibited anti-mycobacterial activity that was similar to the positive control AMPs, LL-37 and RaCa-2. Tp0451a_N showed no antimicrobial activity against any of the other tested bacteria, whereas Tp0451a_C showed considerable bacteriostatic and bactericidal potency toward the Gram-positive bacterium, S. pyogenes, to a level that exceeded that observed with the positive control AMPs, LL-37 and RaCa-2. None of the other three treponemal peptides exhibited anti-streptococcal activity and all four treponemal peptides were inactive against S. enterica and S. aureus. Together, these findings demonstrate that each of the four treponemal peptides identified in our AMP discovery bioinformatics pipeline are capable of exhibiting both bacteriostatic and bactericidal activity.
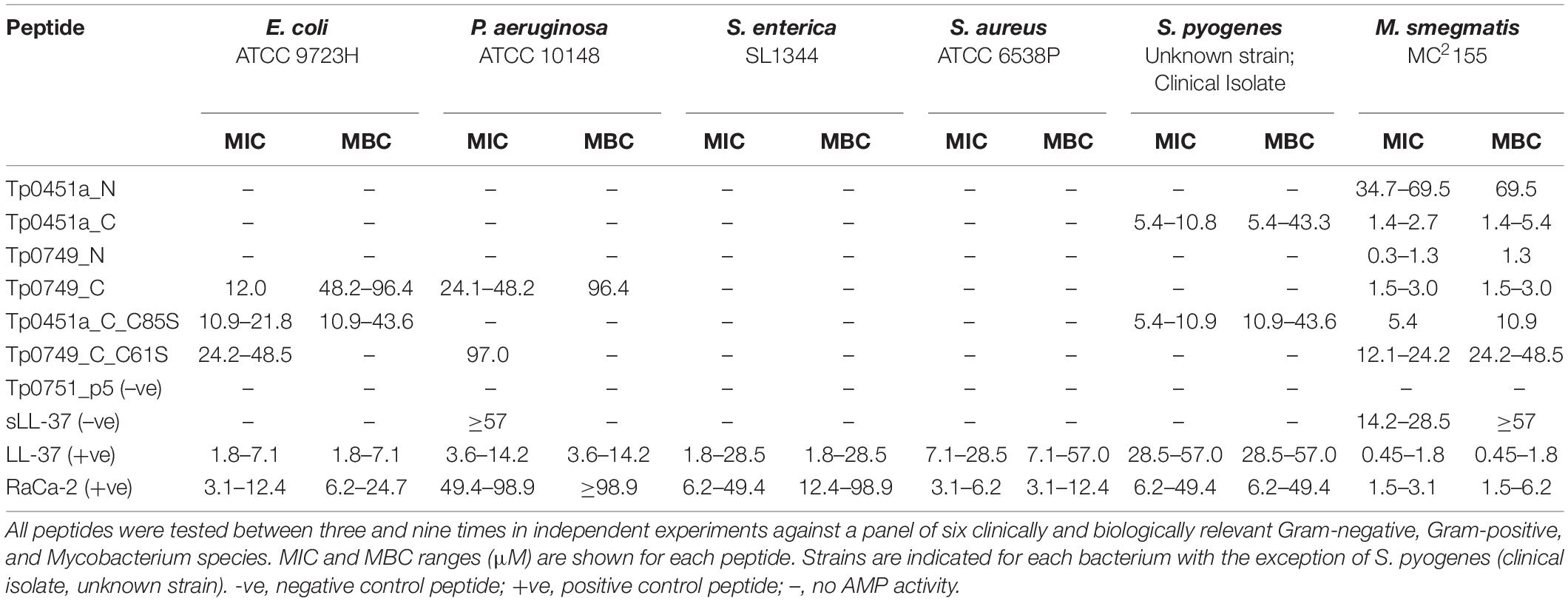
Table 4. Antimicrobial susceptibility testing of T. pallidum candidate AMPCCRs using broth microdilution.
Based on our six-member panel of bacteria, we found Tp0749_C to be the most broad-spectrum AMP of the four treponemal peptides. In addition to its anti-mycobacterial properties, it also showed moderate bacteriostatic activity towards E. coli, moderate-low bacteriostatic activity against P. aeruginosa, and low bactericidal activity against both E. coli and P. aeruginosa (Table 4). The positive control peptide, LL-37, was more active against E. coli and P. aeruginosa than Tp0749_C. The positive control peptide, RaCa-2, was more active against E. coli than Tp0749_C but exhibited lower anti-pseudomonal activity. The other three treponemal peptides showed no activity against these two Gram-negative bacteria. The negative control peptide, Tp0751_p5, a 24-mer peptide from the T. pallidum adhesin Tp0751 with similar physicochemical properties to AMPs, was inactive against all six bacteria in all experiments. Surprisingly, the negative control peptide, sLL-37, a scrambled version of LL-37, exhibited a low degree of antimicrobial activity against M. smegmatis and minimal bacteriostatic activity against P. aeruginosa (Table 4). Since sLL-37 retains many of the same physicochemical properties, including overall charge and amino acid composition, as LL-37, but is predicted to lack the helical content found in native LL-37, this may explain the low-level activity observed with the scrambled version of this peptide. Indeed, a similar low level of antimicrobial activity for sLL-37 has been demonstrated previously in an independent study (Gordon et al., 2005). Together, the Tp0451a and Tp0749 results suggest that T. pallidum is capable of producing AMPs that target Gram-negative bacteria, Gram-positive bacteria, and mycobacteria, and established proof-of-concept for our AMP discovery bioinformatics pipeline.
To evaluate if cysteine residues are important for T. pallidum AMP function, cysteine-to-serine substituted versions of Tp0451a_C (Tp0451a_C_C85S) and Tp0749_C (Tp0749_C_C61S) were synthesized and tested for antimicrobial activity using our panel of six bacteria and broth microdilution assays. These peptides were chosen as they have broad-spectrum AMP activity, and only contain one cysteine residue each. As shown in Table 4, compared to the unmodified, cysteine-containing AMPCCR, Tp0451a_C, the antimicrobial activity of Tp0451a_C85S against M. smegmatis was reduced two- to eight-fold, whereas the cysteine substitution had no effect on anti-streptococcal activity. Interestingly, the cysteine substituted version of Tp0451a_C exhibited moderate antimicrobial activity against E. coli, unlike the unmodified version. Compared to the unmodified, cysteine-containing AMPCCR, Tp0749_C, the bacteriostatic activity of Tp0749_C_C61S against E. coli and P. aeruginosa was reduced two- to four-fold, and reduced eight-fold against M. smegmatis (Table 4). Furthermore, the low bactericidal activity of Tp0749_C against E. coli and P. aeruginosa was abolished in Tp0749_C_C61S and the strong bactericidal activity against M. smegmatis was reduced 16-fold. Consistent with the high abundance of cysteines in the top-ranking predicted AMPs, these findings show that the cysteine residues are important for the AMP activity.
To investigate the potential antimicrobial activity of the T. pallidum AMPCCRs against the frequently co-infecting sexually transmitted pathogen, N. gonorrhoeae, an antimicrobial susceptibility assay based on agar dilution was developed and performed to test for bacteriostatic and bactericidal activities. Although growth was visibly inhibited by Tp0451a_C, none of the four treponemal AMPCCRs completely inhibited Neisseria growth on the agar plates at any of the peptide concentrations. The visible inhibition of growth by Tp0451a_C prompted us to test whether any of the treponemal peptides were bactericidal against N. gonorrhoeae by comparing TVCs of the 3 h-incubated peptide/bacteria mixtures with TVCs from the corresponding positive control growth wells to give the percentage of bacteria killed by the peptides. In agreement with our modified agar dilution results, we found Tp0451a_C to be the only treponemal peptide capable of exhibiting bactericidal activity against N. gonorrhoeae, with Neisseria-killing activity consistently observed at peptide concentrations of 64 μg/mL and higher (Table 5). The loss of anti-Neisseria activity in the cysteine substituted version of Tp0451a_C (Tp0451a_C_C85S) further suggested the importance of cysteine residues in treponemal AMP function. These findings suggest that T. pallidum may express proteins that are capable of killing Neisseria during co-infections involving these two sexually transmitted pathogens.
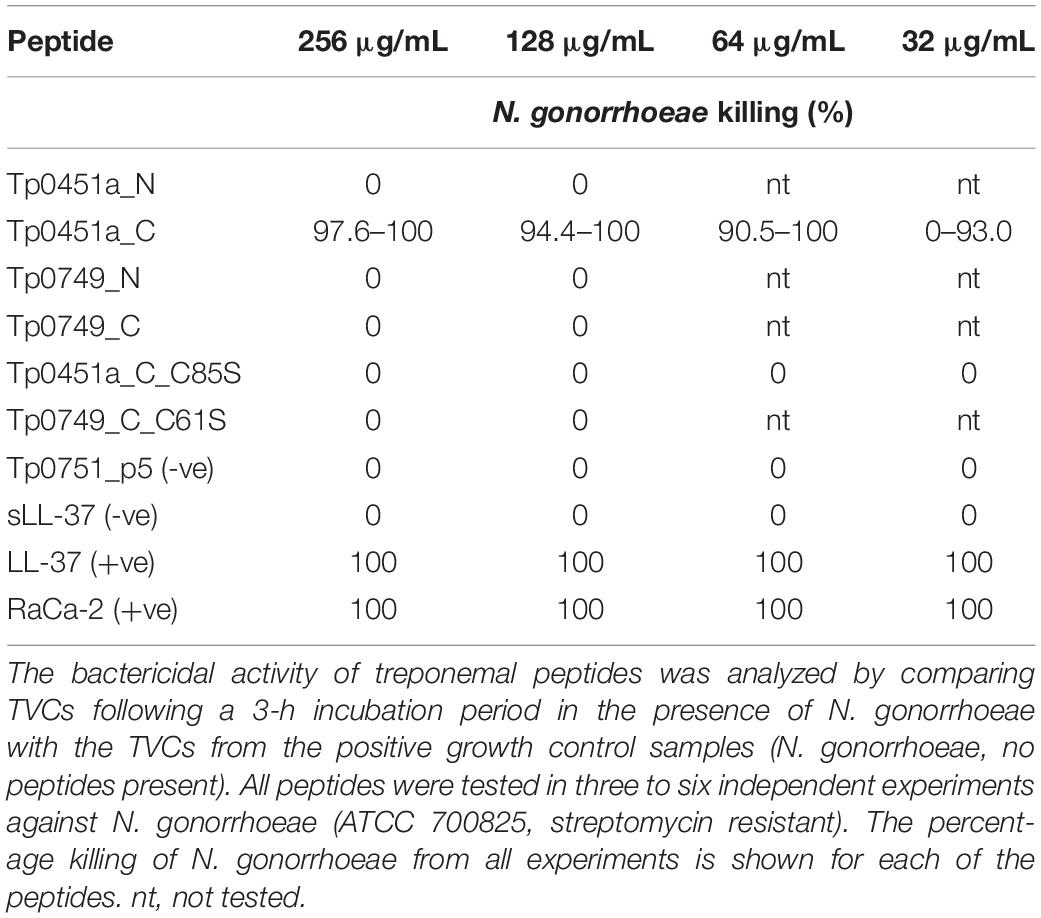
Table 5. Antimicrobial susceptibility testing of T. pallidum candidate AMPCCRs against N. gonorrhoeae using a modified agar dilution method.
AMPCCR Susceptibility Testing of T. pallidum
Treponema pallidum exhibits vigorous motility which can be used as an indication of bacterial viability. To assess the activity of the treponemal peptides against T. pallidum, in vitro-cultured T. pallidum was incubated with each peptide at three different concentrations. Treponeme viability was then determined by counting motile treponemes using darkfield microscopy. As shown in Supplementary Figure 5, only Tp451a_C showed antimicrobial activity against T. pallidum at 64, 16, and 4 μg/ml peptide concentrations, when compared to the negative control peptide, Tp0751_p5. The positive control peptide, LL-37, and Tp0749_N also showed a low level of inhibitory activity against T. pallidum at 64 μg/ml. These findings demonstrate that, similar to other bacteria (Beis and Rebuffat, 2019; Smits et al., 2020), T. pallidum is susceptible to the antimicrobial activity of some of the AMPs it produces, at least when added exogenously.
Immunomodulatory Capabilities of T. pallidum AMPCCRs
Based upon the well-established propensity for AMPs to suppress or induce cytokine production and the persistent nature of T. pallidum infection, we assessed potential immunomodulatory functions of the T. pallidum AMPCCRs through examining their capacity to influence cytokine production (IL-1β, IL-6, IL-8, IL-10, MCP-1, and TNF) from a human monocyte/macrophage cell line. Initially, we quantified cytokine production from monocytes (THP-1 cells) stimulated with the treponemal peptides under non-inflammatory conditions. Stimulation with treponemal peptides resulted in the production of several cytokines from monocytes, but the levels of cytokine were low (Supplementary Figure 6). No statistically significant differences in cytokine production between monocytes stimulated with Tp0751_p5 (peptide with no antimicrobial activity in the present study), and the treponemal AMPCCR, Tp0451a_C, was detected. Exposure of each of the peptides to macrophages (differentiated THP-1 cells) failed to induce a significant difference in cytokine expression when compared to the unstimulated control (Supplementary Figure 7). In summary, no treponemal AMPCCR-specific effect on cytokine secretion by undifferentiated monocytes or macrophages was observed in these non-inflammatory immunomodulation assays.
Previously, it has been shown that LL-37 modulates macrophage cytokine production under conditions where macrophages have been highly activated by exposure to the pro-inflammatory cytokine IL-32γ (Choi et al., 2014). IL-32γ also plays key roles in pathogen defense and persistence of chronic infection (Bai et al., 2010; Dos Santos et al., 2017; Li et al., 2018). To assess whether the T. pallidum AMPCCRs identified in the current study also modulate cytokine expression levels under cytokine-induced pro-inflammatory conditions, macrophages were activated with IL-32γ and immediately additionally stimulated with one of our control peptides (LL-37, sLL-37, and Tp0751_p5), or one of the four treponemal AMPCCRs (Tp0451a_N, Tp0451a_C, Tp0749_N, and Tp0749_C), or not exposed to a peptide (IL-32γ only) or IL-32γ (no stimulation). IL-32γ elicited robust production of the chemokine MCP-1 from macrophages compared to non-IL-32γ stimulated cells, demonstrating that the cells were successfully activated, and notably, both LL-37 and the treponemal AMPCCR Tp0451a_C were found to be the only peptides that significantly downregulated (p < 0.0001 for both peptides) IL-32γ-induced expression of MCP-1, in three independent experiments (Figure 7A). Additionally, LL-37 and Tp0451a_C were able to modulate macrophage-production of the chemokine IL-8 following IL-32γ stimulation: IL-32γ stimulation resulted in lower levels of IL-8 release compared to unstimulated cells, and co-stimulation of macrophages with IL-32γ and LL-37 or Tp0451a_C caused significantly higher levels of IL-8 release compared to from macrophages stimulated with IL-32γ alone (p < 0.0001 for both peptides in three independent experiments (Figure 7B). Co-stimulation of macrophages with each of the peptides failed to significantly affect the expression levels of IL-1β, IL-6, TNF, or IL-10 (Supplementary Figure 8). In addition to the antimicrobial activities described above, these findings demonstrate that the T. pallidum AMPCCR Tp0451a_C is also capable of immunomodulatory activities in certain inflammatory contexts.
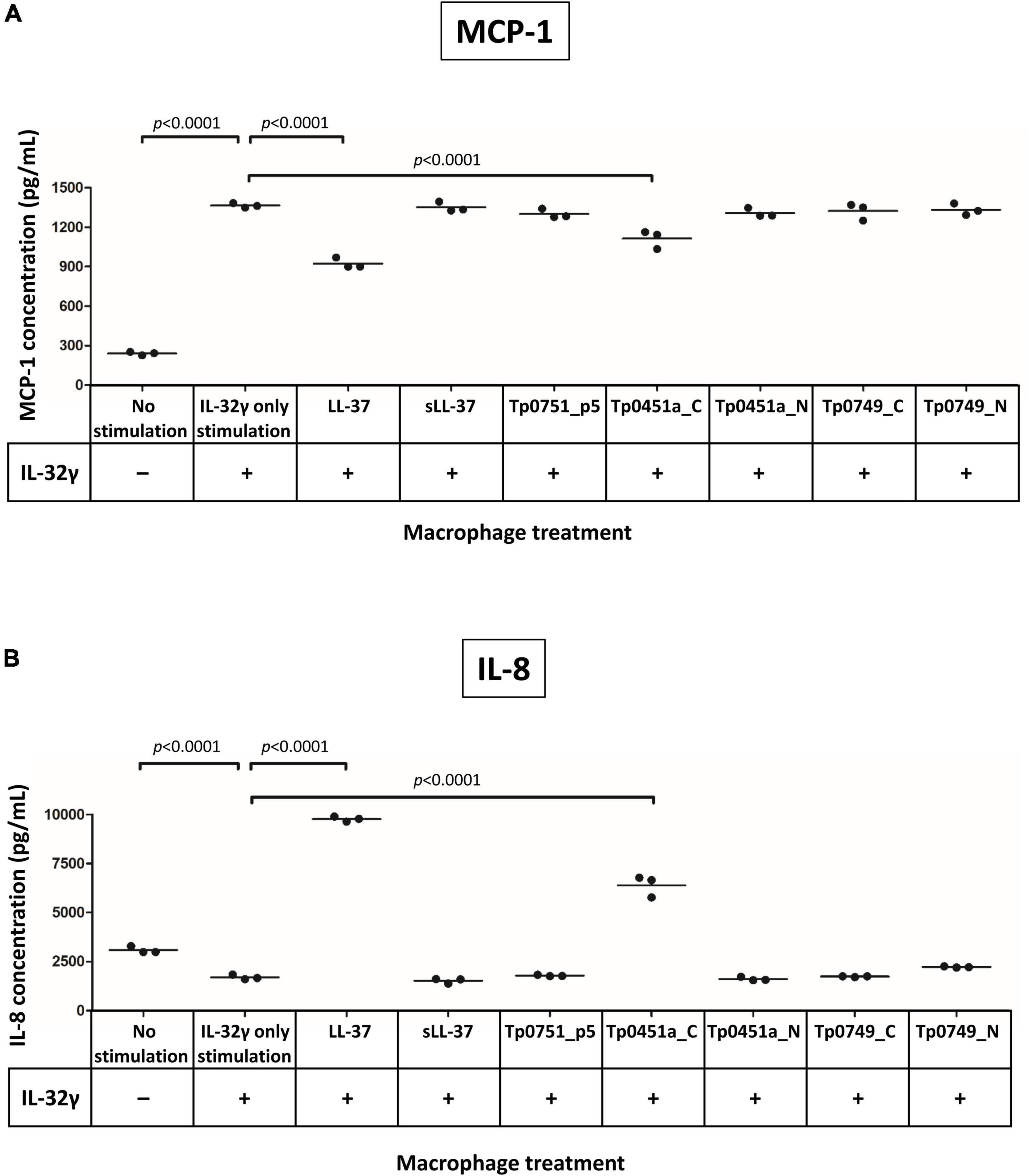
Figure 7. Analysis of the immunomodulatory capacities of the T. pallidum AMPCCRs. Human THP-1 cells were differentiated to macrophages and then stimulated with or without IL-32γ. Cells were then immediately exposed to either LL-37, sLL-37, Tp0751_p5, Tp0451a_C, Tp0451a_N, Tp0749_C, or Tp0749_N and analyzed for expression of (A) MCP-1 and (B) IL-8. Each data point is representative of cells from one well of a 12-well plate. Data shown is representative of three independent experiments. A Dunnett’s multiple comparisons test was used for normally distributed data and a Dunn’s multiple comparisons test was used for data that was not normally distributed. For statistical analyses, mean values from each peptide were compared to the mean of the unstimulated control (IL-32γ only stimulation); p-values (Dunnett’s multiple comparisons test) indicating statistically significant differences observed in three independent experiments are indicated.
Discussion
In this study, we investigated the potential for T. pallidum to express AMPs as a previously unrecognized strategy to defend against competing bacteria and the host immune response. To conduct these studies, we developed a bioinformatics pipeline for the identification of candidate T. pallidum AMPs. By using AMPCCR mapping, followed by a combination of structure, modeling, homology, and cell penetration prediction analyses, potential AMPCCRs were identified in two T. pallidum proteins, Tp0451a and Tp0749. Together, these findings enabled the design and synthesis of four putative AMPCCRs to evaluate these T. pallidum proteins for antibacterial and immunomodulatory functions.
Using antimicrobial susceptibility assays, we demonstrated bacteriostatic and bactericidal activity for the four predicted T. pallidum AMPCCRs. All four peptides were active against the model Mycobacterium sp., M. smegmatis, three of which (Tp0451a_C, Tp0749_N, and Tp0749_C) exhibited robust anti-mycobacterial activity that was similar or more potent than the activity of many antimycobacterial AMPs from other organisms (Abedinzadeh et al., 2015; Gupta et al., 2015; Helbing et al., 2019). Mycobacterium smegmatis is generally considered as nonpathogenic, however, it has been shown on rare occasions to cause disease in humans (Wallace et al., 1988; Newton et al., 1993; Pennekamp et al., 1997; Hong et al., 2003; Ergan et al., 2004). Interestingly, M. smegmatis was originally isolated from syphilitic chancres and gummas (Bloom, 1885) and is known to be present in normal genital (smegma) secretions. Mycobacterium smegmatis is neither a true Gram-negative nor Gram-positive due to its unusual cell envelope ultrastructure and composition, which is similar in all known mycobacteria species, including the important human pathogen Mycobacterium tuberculosis (Brennan and Nikaido, 1995; Cook et al., 2009). Although few cases have been documented, T. pallidum/M. tuberculosis coinfections in HIV patients have been reported (Latif et al., 2020). Given the cell envelope conservation observed within mycobacteria, the T. pallidum proteins demonstrated in this study to possess AMP activity may also be capable of targeting M. tuberculosis during situations of co-infection.
One of the treponemal AMPCCRs (Tp0451a_C) exhibited bacteriostatic and bactericidal activity against a clinical isolate of the Gram-positive extracellular bacterial pathogen, S. pyogenes. At the time of writing, searches of the AMP database, APD3 (see footnote 26) (Wang et al., 2016), identified only 71 known AMPs (out of a total of 3324 known AMPs) with antimicrobial activity against S. pyogenes, including only 12 from other bacteria, most of which are non-pathogenic members of the microbiota. In line with these results, few reports have been published describing this rare antimicrobial activity, making Tp0451a_C a new addition to this select group of S. pyogenes-targeting AMPs. Furthermore, the anti-S. pyogenes activity of Tp0451a_C was found to be generally comparable (Cogen et al., 2010; Uhlmann et al., 2016; Ma et al., 2020) or more potent (Sornwatana et al., 2018; Li et al., 2022) than known AMPs from other organisms. The relatively small number of AMPs that show activity against S. pyogenes is attributed to the expression of several proteins involved in AMP resistance, including the streptococcal cysteine protease SpeB (Schmidtchen et al., 2002) which is involved in the proteolytic degradation and inactivation of AMPs, the M1 protein, streptokinase, and the streptococcal inhibitor of complement, the latter three of which are involved in resistance against defensins and/or LL-37 (Frick et al., 2003; Lauth et al., 2009; Hollands et al., 2012). Our findings show that the AMP-activity identified in Tp0451a_C is capable of circumventing the resistance mechanisms developed by pathogenic S. pyogenes. Importantly, S. pyogenes colonizes and infects several host sites that T. pallidum also encounters, particularly during T. pallidum transmission. These shared host sites include the skin and the mucosal membranes of the oropharynx, rectum, and genitals (Stamm et al., 1978; Mead and Winn, 2000; Lafond and Lukehart, 2006; Sobel et al., 2007; Minami et al., 2010; Verstraelen et al., 2011; Nelson et al., 2016; Norimatsu and Ohno, 2020). Taken together, these findings suggest that Streptococcus-killing AMPs may comprise a defense strategy that facilitates survival during infection by protecting T. pallidum against this clinically relevant, competing microbe in key host infection sites such as the genital mucosa, via direct inhibition and killing.
Tp0451a_C was also demonstrated to be the only treponemal peptide capable of exhibiting bactericidal activity against the frequently co-infecting sexually transmitted pathogen N. gonorrhoeae (Bala et al., 2011). This finding is consistent with the concept that T. pallidum and N. gonorrhoeae localize to the same environmental niches within the urogenital region, particularly during transmission and primary stage syphilis when treponemes are localized within the chancre at the initial site of infection. In light of the increasing global public health threat posed by N. gonorrhoeae due to the rapid emergence of multiple drug resistant strains and with an estimated 78 million new cases per year (Alirol et al., 2017), our finding of a Neisseria-active AMP is relevant to the goal of developing novel therapeutics against this pathogen.
Most AMPs from bacteria exhibit narrow antimicrobial spectra; they are produced in order to defend themselves and their environmental niche from a few genus/species-specific competing species that are often also closely related to the AMP-producing bacterium (Cleveland et al., 2001; Cotter et al., 2005; De Vuyst and Leroy, 2007). This is most likely because closely related bacterial species often reside within the same environmental niches. However, identification of broad spectrum AMPs has become increasingly common, particularly amongst non-pathogenic Gram-positive bacteria (Kemperman et al., 2003a,b; Zendo, 2013; Todorov et al., 2019). Here we found Tp0749_C and Tp0451a_C to exhibit the most broad-spectrum activity of the four treponemal peptides tested. Interestingly, Tp0749_C exhibited bacteriostatic and bactericidal activity against a non-pathogenic strain of the Gram-negative bacterium, E. coli. Non-pathogenic E. coli strains are normal, prevalent residents of the human lower gastrointestinal tract, including the rectum (Zhang et al., 2002; Tenaillon et al., 2010). The fact that syphilis chancres commonly occur in the rectum and anal canal of men-who-have-sex-with-men infected with T. pallidum (Lafond and Lukehart, 2006), suggests (i) the likelihood of a close association between non-pathogenic E. coli and T. pallidum during primary stage syphilis in these individuals and (ii) the possibility that T. pallidum AMPs are expressed that target non-pathogenic E. coli in this host infection site. It is also likely that T. pallidum encounters pathogenic / uropathogenic strains of E. coli during infection. Although these strains were not tested in our antimicrobial susceptibility assays, previous studies have shown that AMPs from other organisms exhibit similar levels of antimicrobial activity against both non-pathogenic and pathogenic / uropathogenic E. coli strains (Fedders et al., 2010; Aghazadeh et al., 2019; Mardirossian et al., 2019; Moazzezy et al., 2020; Li et al., 2022; Lin et al., 2022). Tp0749_C also exhibited antimicrobial activity against the Gram-negative bacterium, P. aeruginosa. P. aeruginosa is an opportunistic pathogen that is the causative agent of both severe acute and chronic infections in immunocompromised individuals, and is one of the major etiological agents of urinary tract infections (Ronald, 2002; Shigemura et al., 2006). Given that T. pallidum / HIV co-infections are common (Shockman et al., 2014; Burchell et al., 2015), and the potential for co-localization of T. pallidum and P. aeruginosa to the same environmental niches within the urogenital region in immunocompromised patients, these findings support the concept that T. pallidum may produce anti-pseudomonal AMPs as a defense mechanism during infection. Together, T0451a and Tp0749 were capable of inhibition and/or killing of five of the seven diverse bacterial species tested, including three Gram-negative bacteria, a Gram-positive bacterium, and a mycobacterium. This finding is consistent with the ability of T. pallidum to invade and disseminate to any host organ or tissue during the different stages of syphilis and the variable, and often complex, polymicrobial environments that exist within different sites of infection.
As part of our physicochemical analyses of T. pallidum miniproteins, we determined that the rare amino acid cysteine appears frequently in high-ranking predicted AMPs. Substitution of the single cysteine residue in Tp0451a_C with the structurally similar residue serine reduced its antimicrobial activity against N. gonorrhoeae and M. smegmatis, but resulted in a gain of antimicrobial activity against E. coli. Substitution of the single cysteine residue in Tp0749_C with serine greatly reduced the antimicrobial activity against E. coli and P. aeruginosa, and even more so against M. smegmatis. This unusual physicochemical property is shared with specific classes of AMPs, in particular certain eukaryotic AMPs, including defensins which typically contain six to twelve cysteines (Shafee et al., 2016), protegrins, which are active against several sexually transmitted pathogens and often contain four cysteines (Tamamura et al., 1995; Fahrner et al., 1996; Qu et al., 1996; Yasin et al., 1996), and brevinins which usually contain two cysteine residues (Simmaco et al., 1994). More recently, increasing numbers of cysteine-rich and defensin-like AMPs are being discovered in bacteria (Baindara et al., 2017; Sugrue et al., 2020). The even number of cysteine residues in these AMPs allows for the formation of intra-molecular disulfide bonds, which are important for AMP stability (Dias Rde and Franco, 2015). Literature and PDB searches indicate that most cysteine-containing AMP structures that have been solved are monomers that contain even numbers of cysteines / intra-molecular disulfide bonds. However, some AMPs with an odd number of cysteines form inter-molecular disulfide bonds resulting in dimers that can increase activity and stability compared to monomeric forms (Campopiano et al., 2004; Min et al., 2017). The AMPCCRs, Tp0451a_C and Tp0749_C, should only be capable of forming inter-molecular disulfide bonds / multimers, as they only contain one cysteine each. However, full-length Tp0451a_C contains seven cysteine resides and Tp0749 contains two cysteine residues, suggesting the additional possibility of intra-molecular disulfide bond formation. Similar to the aforementioned cysteine-rich AMPs, our findings are in agreement with the hypothesis that cysteines are important for the demonstrated treponemal AMP activity, possibly via the formation of inter- and/or intra-molecular disulfide bonds and miniprotein multimerization.
Due to the antimicrobial findings described herein, the fact that many AMPs have the ability to modulate immune functions (Lai and Gallo, 2009; Kindrachuk et al., 2013; Dicks et al., 2018; Malaczewska et al., 2019), and the stealth nature of T. pallidum, we investigated the immunomodulatory capabilities of the four treponemal peptides under both non-inflammatory and cytokine-induced inflammatory conditions, through assaying cytokine production from a human monocyte/macrophage cell line. Given that IL-32γ plays key roles in pathogen defense and persistence of chronic infection (Bai et al., 2010; Dos Santos et al., 2017; Li et al., 2018), and that AMP-mediated immunomodulatory activities have been shown to modulate IL-32γ-induced cytokine production (Choi et al., 2014), IL-32γ was selected as a biologically relevant co-stimulatory agent for our assays to mimic the pro-inflammatory environment present during T. pallidum infection (Lafond and Lukehart, 2006). Interleukin-32 is a recently discovered pro-inflammatory cytokine that functions in the persistence of inflammation via induction of other pro-inflammatory cytokines and in the control of infectious and chronic diseases (Khawar et al., 2016; Ribeiro-Dias et al., 2017). It has been found to be elevated in human infections, where it can have a protective (Mycobacterium tuberculosis, HIV) or detrimental (Helicobacter pylori) effect on the host (Rasool et al., 2008; Bai et al., 2010; Peng et al., 2014). Although the up-regulation and involvement of IL-32 isoforms in T. pallidum infection has yet to be demonstrated, the inflammation associated with T. pallidum infection is consistent with this possibility. Furthermore, IL-32 is upregulated in HIV infection (Rasool et al., 2008), a pathogen that is frequently involved in co-infections with T. pallidum.
Under IL-32γ-induced pro-inflammatory conditions in macrophages, we found that co-stimulation with Tp0451a_C resulted in decreased production of the pro-inflammatory chemokine MCP-1 and increased production of the pro-inflammatory chemokine, IL-8, compared to macrophages not exposed to peptides. MCP-1 is produced by a variety of cell types, several of which are relevant to T. pallidum infection, including monocytes, macrophages, epithelial, and endothelial cells (Yoshimura et al., 1989a,b; Cushing et al., 1990; Standiford et al., 1991). Monocyte chemoattractant protein-1 is the main chemoattractant for monocytes and macrophages to sites of inflammation where recruited monocytes undergo transformation into macrophages (Deshmane et al., 2009). During infection, clearance of T. pallidum is dependent on macrophages via antibody-mediated opsonophagocytosis and subsequent killing by macrophages (Lukehart and Miller, 1978; Baker-Zander and Lukehart, 1992; Baker-Zander et al., 1993). However, in the absence of antibiotic intervention, treponemes are never fully cleared from the host. The ability of Tp0451a to down-regulate expression of pro-inflammatory cytokines, including MCP-1, may contribute to treponemal survival via local suppression of the inflammatory response and dampening of macrophage recruitment.
In the present study, we also found Tp0451a_C capable of significantly increasing production of IL-8 in co-stimulated macrophages. This pro-inflammatory cytokine is a potent neutrophil chemoattractant that can induce neutrophil morphology changes and degranulation, and neutrophil migration to sites of infection and inflammation (Baggiolini and Clark-Lewis, 1992; Kolaczkowska and Kubes, 2013; Arango Duque and Descoteaux, 2014). Similar to the findings described herein, IL-8 has been shown to be up-regulated in macrophages stimulated with T. pallidum peptides derived from the major immunogenic lipoprotein, TpN47 (Sellati et al., 1998). If IL-8 up-regulation, by Tp0451a_C or other treponemal proteins, is recapitulated at the level of viable whole T. pallidum, the following consequences may be envisioned that could promote T. pallidum pathogenesis. Since neutrophils are largely ineffective against T. pallidum during early infection (Lafond and Lukehart, 2006), IL-8 production may function as an early infection immune diversion mechanism. Alternatively, an increase in IL-8 expression may redirect the immune response towards competing neutrophil-sensitive microorganisms in polymicrobial host sites, such as uropathogenic E. coli (Svanborg-Eden et al., 1987). As IL-8 is also a pro-angiogenic chemokine (Li et al., 2003), up-regulation may contribute to the vascular inflammation and increased angiogenesis observed during secondary syphilis (French, 2007; Gao et al., 2019) and may promote treponemal accessibility to the host’s circulatory system via proliferation of endothelial cells and formation of new blood vessels, similar to the mechanism proposed for the IL-8-inducing T. pallidum protein TpF1 (Pozzobon et al., 2016).
Here we employed a combination of bioinformatics, antimicrobial susceptibility testing, and immunomodulation assays that allowed for the experimental identification of the first proteins with AMP activity from a spirochete bacterium. However, there are limitations with our approach. First, due to difficulties with expression, full-length versions of the treponemal proteins were not used in antimicrobial susceptibility or immunomodulation experiments. Confirmation of AMP activity with the full-length proteins would be optimal, and may uncover an enhanced AMP activity due to the additive effects of the AMPCCRs. Second, in our antimicrobial susceptibility assays, we only tested for antibacterial activity and thus may have missed antifungal, antiviral, and/or antiparasitic activities. Third, although most AMPs are smaller than 150 amino acids, our bioinformatics pipeline omitted larger proteins from our analyses thereby excluding potential colicin-like bacteriocins. Fourth, the expression status of approximately three-quarters of all T. pallidum miniproteins of unknown function remain to be determined, due in part to the experimental difficulty with identifying small, positively-charged proteins via mass spectrometry-based proteomic analyses. Furthermore, the identification of potentially secreted T. pallidum proteins from in vivo and in vitro cultures by mass spectrometry and other methodologies is not possible due to the overwhelming abundance of contaminating rabbit/host proteins and the complexity of the culture medium, respectively. Fifth, similar to other AMP studies, the in vitro nature of the antimicrobial susceptibility assays, which were based on established, standardized protocols (Wiegand et al., 2008), are not representative of the complex host environments encountered during infection. Therefore, it remains to be determined if the concentrations of T. pallidum peptides that resulted in bacteriostatic and bactericidal activities are representative of the concentrations that are expressed and secreted into diverse host sites during infection.
Finally, and most significantly, similar to other recently discovered AMPs, our functional characterization of the treponemal proteins with AMP activity has yet to determine if/how they are processed/activated, and the mechanisms involved in self-immunity and export. Most bacterial AMPs that have been functionally characterized are synthesized as inactive protein precursors comprised of a N-terminal leader peptide/signal peptide, that is required for export, and a C-terminal proprotein that contains the critical core regions (Torrent et al., 2009, 2012; Chang et al., 2015; Perez et al., 2018). Following specific protease-mediated cleavage of the leader / signal peptide, the mature, functionally-active AMP is exported from the AMP-producing bacteria to the external environment via ATP-binding cassette (ABC) transporters (Beis and Rebuffat, 2019). In the present study, we showed that several T. pallidum candidate AMPs contain N-terminal amino acid sequences with similarity to the Gram-negative Sec-independent double-glycine/glycine-alanine (GG/GA) leader peptides, suggesting that a subset of T. pallidum AMPs may also use this recognition signal to direct secretion and allow for activation of precursor AMP forms. In addition, proteome functional annotation analyses and Phyre2 structure modeling of open reading frames located close to several potential AMPs identified (1) putative homologs and structural orthologs of proteases with potential roles in AMP leader peptide cleavage and activation and (2) ABC transporters that may mediate export of the mature active treponemal AMPs to the external host environment during infection. Notably, an emerging class of bacterial AMPs with varying characteristics, referred to as leaderless AMPs, do not undergo post-translational processing or modification, are produced without an export-mediating N-terminal leader/signal peptide, and are fully active immediately following expression (Perez et al., 2018). Leaderless AMPs are the most poorly understood group of bacterial AMPs, and the molecular mechanisms underlying self-immunity, secretion, and export remain largely unknown (Perez et al., 2018). Although no common secretion or export mechanism has been discovered for leaderless AMPs, an ABC transporter protein (LmrB) has been shown to be involved in the immunity and export of a leaderless AMP from Lactococcus lactis, LsbB (Gajic et al., 2003). Additionally, an ABC transporter protein (DDHIJ) from Enterococcus faecalis has been implicated in the active export of the leaderless AMP, Enterocin DD14 (Ladjouzi et al., 2020). It is possible that Tp0451a and Tp0749 belong to, or are related to, the leaderless group of bacterial AMPs. In this situation, the two T. pallidum AMPs would be expressed without processing/proteolytic cleavage, followed by export via one or more of the many ABC transporter proteins located within the T. pallidum proteome (Fraser et al., 1998). This mechanism would result in the N- and C-terminal AMPCCRs identified in the current study to remain fully intact and associated with each other, potentially allowing for enhanced, broad spectrum AMP activity due to the additive effects of the AMPCCRs. This is consistent with the knowledge that AMP activity is often localized to more than one discrete peptide region, or domain (AMPCCRs), within the full-length protein (Todorov, 2009; Torrent et al., 2012; Snyder and Worobo, 2014).
In the present study, we showed that the viability of T. pallidum was reduced following incubation with Tp0451a_C. This result was not surprising, given that AMPs from other bacteria are highly toxic to the AMP-producing strain, necessitating the expression of self-immunity proteins and dedicated ABC transporters that protect the AMP-producing bacteria during expression within the cell through extrusion outside and away from the cell (Smits et al., 2020). The unique composition of the T. pallidum cell envelope may confer a degree of protection against AMPs, as suggested in a previous study that investigated the activity of the full-length mammalian AMP LL-37 against T. pallidum (Cox et al., 2003). Specifically, the T. pallidum outer membrane has low anionic phospholipid content compared to other Gram-negative bacteria, contains few outer membrane proteins including porins and negatively-charged proteins, lacks LPS, and contains cholesterol. Thus, T. pallidum possesses a cell envelope that is reminiscent of the host cell membrane with regards to a neutral surface charge and the presence of the AMP inhibitory molecule, cholesterol (Feigin et al., 1995; Matsuzaki et al., 1995; Glukhov et al., 2005). Together, these may help prevent the electrostatic binding of cationic AMPs to the T. pallidum surface, which is consistent with our findings in the current study that showed only minor or no decrease in T. pallidum viability following incubation with all but one of the AMPs that were tested.
In conclusion, this study has established proof-of-concept for our AMP discovery bioinformatics pipeline via the experimental identification of proteins with AMP activity in T. pallidum. The ability of T. pallidum to produce proteins with dual antimicrobial and immunomodulatory activities may contribute to treponeme survival by eliminating competing microbes via direct inhibition and killing effects, and by modulating the host immune response to promote both indirect inhibition and killing of competing bacterial species and immune evasion. This research has the potential to enhance our understanding of the unique pathogenesis of T. pallidum and reveal novel defense and survival mechanisms with broad applicability to bacterial pathogens, including other pathogenic spirochetes.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics Statement
The animal study was reviewed and approved by the local institutional review board at the University of Victoria, and conducted in strict accordance with standard accepted principles as set forth by the Canadian Council on Animal Care, National Institutes of Health and the United States Department of Agriculture in a facility accredited by the Canadian Council on Animal Care and the American Association for the Accreditation of Laboratory Animal Care.
Author Contributions
SH, CC, LR, KC, ES, RR, and MB contributed to the experimental design. SH, KC, ES, AG, SM, AH, and RR conducted the experiments. SH, KC, RR, ES, LR, MB, and CC were involved in the analysis and interpretation of the data. CC and LR acquired financial support for the project. SH wrote the first draft of the manuscript with contributions from KC. ES, RR, AG, SM, AH, CC, LR, and MB reviewed the manuscript before submission for accuracy and intellectual content. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the National Institutes of Health, National Institute of Allergy and Infectious Diseases (R37AI051334; CC), the Natural Sciences and Engineering Research Council of Canada (327196; CC and LR), and the Canadian Institutes of Health Research (388288, LR).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
Escherichia coli ATCC 9723H, P. aeruginosa ATCC 10148, S. aureus ATCC 6538P, and S. pyogenes (hospital isolate, strain unknown) were supplied by the Department of Biochemistry and Microbiology, University of Victoria, BC. Mycobacterium smegmatis MC2155 was kindly supplied by Francis Nano, University of Victoria, BC, and S. enterica subsp. enterica serovar Typhimurium SL1344 was kindly supplied by Lisa Reynolds, University of Victoria, BC. We thank Caren Helbing (University of Victoria) and Inanc Birol (University of British Columbia) for permitting the use of RaCa-2 peptide as a positive control in antimicrobial susceptibility assays.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2022.888525/full#supplementary-material
Footnotes
- ^ https://www.ncbi.nlm.nih.gov/genome/?term=treponema+pallidum
- ^ https://www.dveltri.com/ascan/
- ^ http://cabgrid.res.in:8080/amppred/
- ^ http://www.camp.bicnirrh.res.in/predict
- ^ https://services.healthtech.dtu.dk/service.php?SignalP-5.0
- ^ https://services.healthtech.dtu.dk/service.php?LipoP-1.0
- ^ https://www.ebi.ac.uk/Tools/msa/clustalo/
- ^ https://weblogo.berkeley.edu/logo.cgi
- ^ https://aps.unmc.edu/prediction
- ^ http://www.sbg.bio.ic.ac.uk/~phyre2/html/page.cgi?id=index
- ^ http://tcoffee.crg.cat/apps/ampa/do
- ^ http://www.camp.bicnirrh.res.in/predict_c/
- ^ https://webs.iiitd.edu.in/raghava/antibp/submit.html
- ^ https://webs.iiitd.edu.in/raghava/antibp2/submit.html
- ^ http://www.compbio.dundee.ac.uk/jpred/
- ^ http://bioinf.cs.ucl.ac.uk/psipred/
- ^ http://heliquest.ipmc.cnrs.fr/
- ^ https://mobyle.rpbs.univ-paris-diderot.fr/cgi-bin/portal.py#forms::PEP-FOLD
- ^ https://swissmodel.expasy.org/
- ^ https://salilab.org/modeller/
- ^ https://www.molsoft.com/index.html
- ^ http://prodata.swmed.edu/promals3d/promals3d.php
- ^ https://blast.ncbi.nlm.nih.gov/Blast.cgi
- ^ https://webs.iiitd.edu.in/raghava/cellppd/submit_prot.php
- ^ https://aps.unmc.edu/
References
Abedinzadeh, M., Gaeini, M., and Sardari, S. (2015). Natural antimicrobial peptides against Mycobacterium tuberculosis. J. Antimicrob. Chemother. 70, 1285–1289. doi: 10.1093/jac/dku570
Aghazadeh, H., Memariani, H., Ranjbar, R., and Pooshang Bagheri, K. (2019). The activity and action mechanism of novel short selective LL-37-derived anticancer peptides against clinical isolates of Escherichia coli. Chem. Biol. Drug. Des. 93, 75–83. doi: 10.1111/cbdd.13381
Alirol, E., Wi, T. E., Bala, M., Bazzo, M. L., Chen, X. S., Deal, C., et al. (2017). Multidrug-resistant gonorrhea: a research and development roadmap to discover new medicines. PLoS Med. 14:e1002366.
Almagro Armenteros, J. J., Tsirigos, K. D., Sonderby, C. K., Petersen, T. N., Winther, O., Brunak, S., et al. (2019). SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 37, 420–423. doi: 10.1038/s41587-019-0036-z
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410.
Arango Duque, G., and Descoteaux, A. (2014). Macrophage cytokines: involvement in immunity and infectious diseases. Front. Immunol. 5:491. doi: 10.3389/fimmu.2014.00491
Baggiolini, M., and Clark-Lewis, I. (1992). Interleukin-8, a chemotactic and inflammatory cytokine. FEBS Lett. 307, 97–101. doi: 10.1016/0014-5793(92)80909-z
Bai, X., Kim, S. H., Azam, T., Mcgibney, M. T., Huang, H., Dinarello, C. A., et al. (2010). IL-32 is a host protective cytokine against Mycobacterium tuberculosis in differentiated THP-1 human macrophages. J. Immunol. 184, 3830–3840. doi: 10.4049/jimmunol.0901913
Baindara, P., Kapoor, A., Korpole, S., and Grover, V. (2017). Cysteine-rich low molecular weight antimicrobial peptides from Brevibacillus and related genera for biotechnological applications. World J. Microbiol. Biotechnol. 33:124. doi: 10.1007/s11274-017-2291-9
Baker-Zander, S. A., and Lukehart, S. A. (1992). Macrophage-mediated killing of opsonized Treponema pallidum. J. Infect. Dis. 165, 69–74. doi: 10.1093/infdis/165.1.69
Baker-Zander, S. A., Shaffer, J. M., and Lukehart, S. A. (1993). Characterization of the serum requirement for macrophage-mediated killing of Treponema pallidum ssp. pallidum: relationship to the development of opsonizing antibodies. FEMS Immunol. Med. Microbiol. 6, 273–279. doi: 10.1111/j.1574-695X.1993.tb00339.x
Bala, M., Mullick, J. B., Muralidhar, S., Kumar, J., and Ramesh, V. (2011). Gonorrhoea & its co-infection with other ulcerative, non-ulcerative sexually transmitted & HIV infection in a regional STD centre. Indian J. Med. Res. 133, 346–349.
Beis, K., and Rebuffat, S. (2019). Multifaceted ABC transporters associated to microcin and bacteriocin export. Res. Microbiol. 170, 399–406. doi: 10.1016/j.resmic.2019.07.002
Bienert, S., Waterhouse, A., De Beer, T. A., Tauriello, G., Studer, G., Bordoli, L., et al. (2017). The SWISS-MODEL repository-new features and functionality. Nucleic Acids Res. 45, D313–D319. doi: 10.1093/nar/gkw1132
Bloom, J. N. (1885). The bacillus of syphilis by DR. Sigmund Lustgarten (Translation of lustgarten s. ueber stezifische bacillen in syphilitischen. wiener medizinische wochenschrift 1884; 34:1389). Lancet 1, 609–610. doi: 10.1016/s0140-6736(02)17741-5
Brennan, P. J., and Nikaido, H. (1995). The envelope of mycobacteria. Annu. Rev. Biochem. 64, 29–63. doi: 10.1146/annurev.bi.64.070195.000333
Brodsky, I. E., and Gunn, J. S. (2005). A bacterial sensory system that activates resistance to innate immune defenses: Potential targets for antimicrobial therapeutics. Mol. Interv. 5, 335–337. doi: 10.1124/mi.5.6.4
Burchell, A. N., Allen, V. G., Gardner, S. L., Moravan, V., Tan, D. H., Grewal, R., et al. (2015). High incidence of diagnosis with syphilis co-infection among men who have sex with men in an HIV cohort in Ontario. Canada. BMC Infect. Dis. 15:356. doi: 10.1186/s12879-015-1098-2
Cameron, C. E., Brouwer, N. L., Tisch, L. M., and Kuroiwa, J. M. Y. (2005). Defining the interaction of the Treponema pallidum adhesin Tp0751 with laminin. Infect. Immun. 73, 7485–7494. doi: 10.1128/IAI.73.11.7485-7494.2005
Campopiano, D. J., Clarke, D. J., Polfer, N. C., Barran, P. E., Langley, R. J., Govan, J. R., et al. (2004). Structure-activity relationships in defensin dimers: a novel link between beta-defensin tertiary structure and antimicrobial activity. J. Biol. Chem. 279, 48671–48679. doi: 10.1074/jbc.M404690200
Cardozo, T., Totrov, M., and Abagyan, R. (1995). Homology modeling by the ICM method. Proteins 23, 403–414. doi: 10.1002/prot.340230314
Chang, K. Y., Lin, T. P., Shih, L. Y., and Wang, C. K. (2015). Analysis and prediction of the critical regions of antimicrobial peptides based on conditional random fields. PLoS One 10:e0119490. doi: 10.1371/journal.pone.0119490
Chanput, W., Mes, J. J., and Wichers, H. J. (2014). THP-1 cell line: an in vitro cell model for immune modulation approach. Int. Immunopharmacol. 23, 37–45. doi: 10.1016/j.intimp.2014.08.002
Chiorean, S., Vederas, J. C., and Van Belkum, M. J. (2018). Identification and heterologous expression of the sec-dependent bacteriocin Faerocin MK from Enterococcus faecium M3K31. Probiotics Antimicrob. Proteins 10, 142–147. doi: 10.1007/s12602-017-9374-7
Choi, K. Y., Napper, S., and Mookherjee, N. (2014). Human cathelicidin LL-37 and its derivative IG-19 regulate interleukin-32-induced inflammation. Immunology 143, 68–80. doi: 10.1111/imm.12291
Cleveland, J., Montville, T. J., Nes, I. F., and Chikindas, M. L. (2001). Bacteriocins: safe, natural antimicrobials for food preservation. Int. J. Food Microbiol. 71, 1–20. doi: 10.1016/s0168-1605(01)00560-8
Coelho, E. C., Souza, S. B., Costa, C. C. S., Costa, L. M., Pinheiro, L. M. L., Machado, L. F. A., et al. (2021). Treponema pallidum in female sex workers from the brazilian marajo archipelago: prevalence, risk factors, drug-resistant mutations and coinfections. Trans. R. Soc. Trop. Med. Hyg. 115, 792–800. doi: 10.1093/trstmh/traa127
Cogen, A. L., Yamasaki, K., Sanchez, K. M., Dorschner, R. A., Lai, Y., Macleod, D. T., et al. (2010). Selective antimicrobial action is provided by phenol-soluble modulins derived from Staphylococcus epidermidis, a normal resident of the skin. J. Invest. Dermatol. 130, 192–200. doi: 10.1038/jid.2009.243
Collart, P., Franceschini, P., and Durel, P. (1971). Experimental rabbit syphilis. Br. J. Vener. Dis. 47, 389–400.
Conlon, J. M., Mechkarska, M., Coquet, L., Jouenne, T., Leprince, J., Vaudry, H., et al. (2011). Characterization of antimicrobial peptides in skin secretions from discrete populations of Lithobates chiricahuensis (Ranidae) from central and southern Arizona. Peptides 32, 664–669. doi: 10.1016/j.peptides.2011.01.018
Cook, G. M., Berney, M., Gebhard, S., Heinemann, M., Cox, R. A., Danilchanka, O., et al. (2009). Physiology of mycobacteria. Adv. Microb. Physiol. 55, 81–182, 318–189.
Cotter, P. D., Hill, C., and Ross, R. P. (2005). Bacteriocins: developing innate immunity for food. Nat. Rev. Microbiol. 3, 777–788. doi: 10.1038/nrmicro1273
Cox, D. L., Sun, Y., Liu, H., Lehrer, R. I., and Shafer, W. M. (2003). Susceptibility of Treponema pallidum to host-derived antimicrobial peptides. Peptides 24, 1741–1746. doi: 10.1016/j.peptides.2003.07.026
Crooks, G. E., Hon, G., Chandonia, J. M., and Brenner, S. E. (2004). WebLogo: a sequence logo generator. Genome Res. 14, 1188–1190. doi: 10.1101/gr.849004
Cumberland, M. C., and Turner, T. B. (1949). Rate of multiplication of Treponema pallidum in normal and immune rabbits. Amer. J. Syph. 33, 201–212.
Cushing, S. D., Berliner, J. A., Valente, A. J., Territo, M. C., Navab, M., Parhami, F., et al. (1990). Minimally modified low density lipoprotein induces monocyte chemotactic protein 1 in human endothelial cells and smooth muscle cells. Proc. Natl. Acad. Sci. U.S.A. 87, 5134–5138. doi: 10.1073/pnas.87.13.5134
De Vuyst, L., and Leroy, F. (2007). Bacteriocins from lactic acid bacteria: Production, purification, and food applications. J. Mol. Microbiol. Biotechnol. 13, 194–199. doi: 10.1159/000104752
Deshmane, S. L., Kremlev, S., Amini, S., and Sawaya, B. E. (2009). Monocyte chemoattractant protein-1 (MCP-1): an overview. J. Interferon Cytokine Res. 29, 313–326. doi: 10.1089/jir.2008.0027
Dias Rde, O., and Franco, O. L. (2015). Cysteine-stabilized alphabeta defensins: from a common fold to antibacterial activity. Peptides 72, 64–72. doi: 10.1016/j.peptides.2015.04.017
Dicks, L. M. T., Dreyer, L., Smith, C., and Van Staden, A. D. (2018). A review: the fate of bacteriocins in the human gastro-intestinal tract: do they cross the gut-blood barrier? Front Microbiol. 9:2297. doi: 10.3389/fmicb.2018.02297
Dirix, G., Monsieurs, P., Dombrecht, B., Daniels, R., Marchal, K., Vanderleyden, J., et al. (2004). Peptide signal molecules and bacteriocins in Gram-negative bacteria: a genome-wide in silico screening for peptides containing a double-glycine leader sequence and their cognate transporters. Peptides 25, 1425–1440. doi: 10.1016/j.peptides.2003.10.028
Dos Santos, J. C., Heinhuis, B., Gomes, R. S., Damen, M. S., Real, F., Mortara, R. A., et al. (2017). Cytokines and microbicidal molecules regulated by IL-32 in THP-1-derived human macrophages infected with New World Leishmania species. PLoS Negl. Trop. Dis. 11:e0005413. doi: 10.1371/journal.pntd.0005413
Drozdetskiy, A., Cole, C., Procter, J., and Barton, G. J. (2015). JPred4: a protein secondary structure prediction server. Nucleic Acids Res. 43, W389–W394. doi: 10.1093/nar/gkv332
Edmondson, D. G., and Norris, S. J. (2021). In vitro cultivation of the syphilis spirochete Treponema pallidum. Curr. Protoc. 1:e44. doi: 10.1002/cpz1.44
Edmondson, D. G., Hu, B., and Norris, S. J. (2018). Long-term in vitro culture of the syphilis spirochete Treponema pallidum subsp. pallidum. mBio 9:e01153–18. doi: 10.1128/mBio.01153-18
Ergan, B., Coplu, L., Alp, A., and Artvinli, M. (2004). Mycobacterium smegmatis pneumonia. Respirology 9, 283–285. doi: 10.1111/j.1440-1843.2004.00570.x
Fahrner, R. L., Dieckmann, T., Harwig, S. S., Lehrer, R. I., Eisenberg, D., and Feigon, J. (1996). Solution structure of protegrin-1, a broad-spectrum antimicrobial peptide from porcine leukocytes. Chem. Biol. 3, 543–550. doi: 10.1016/s1074-5521(96)90145-3
Fedders, H., Podschun, R., and Leippe, M. (2010). The antimicrobial peptide Ci-MAM-A24 is highly active against multidrug-resistant and anaerobic bacteria pathogenic for humans. Int. J. Antimicrob. Agents 36, 264–266. doi: 10.1016/j.ijantimicag.2010.04.008
Feigin, A. M., Teeter, J. H., and Brand, J. G. (1995). The influence of sterols on the sensitivity of lipid bilayers to melittin. Biochem. Biophys. Res. Commun. 211, 312–317. doi: 10.1006/bbrc.1995.1812
Fraser, C. M., Norris, S. J., Weinstock, G. M., White, O., Sutton, G. G., Dodson, R., et al. (1998). Complete genome sequence of Treponema pallidum, the syphilis spirochete. Science 281, 375–388.
Frick, I. M., Akesson, P., Rasmussen, M., Schmidtchen, A., and Bjorck, L. (2003). SIC, a secreted protein of Streptococcus pyogenes that inactivates antibacterial peptides. J. Biol. Chem. 278, 16561–16566. doi: 10.1074/jbc.M301995200
Gajic, O., Buist, G., Kojic, M., Topisirovic, L., Kuipers, O. P., and Kok, J. (2003). Novel mechanism of bacteriocin secretion and immunity carried out by lactococcal multidrug resistance proteins. J. Biol. Chem. 278, 34291–34298. doi: 10.1074/jbc.M211100200
Gao, Z. X., Liu, L. L., Lin, L. R., Tong, M. L., Liu, F., and Yang, T. C. (2019). Treponema pallidum induces the secretion of HDVSMC inflammatory cytokines to promote the migration and adhesion of THP-1 cells. Front. Cell. Infect. Microbiol. 9:220. doi: 10.3389/fcimb.2019.00220
Gautam, A., Chaudhary, K., Kumar, R., Sharma, A., Kapoor, P., Tyagi, A., et al. (2013). In silico approaches for designing highly effective cell penetrating peptides. J. Transl. Med. 11:74. doi: 10.1186/1479-5876-11-74
Gautier, R., Douguet, D., Antonny, B., and Drin, G. (2008). HELIQUEST: a web server to screen sequences with specific alpha-helical properties. Bioinformatics 24, 2101–2102. doi: 10.1093/bioinformatics/btn392
Gilson, L., Mahanty, H. K., and Kolter, R. (1990). Genetic analysis of an MDR-like export system: the secretion of colicin V. EMBO J. 9, 3875–3884. doi: 10.1002/j.1460-2075.1990.tb07606.x
Glukhov, E., Stark, M., Burrows, L. L., and Deber, C. M. (2005). Basis for selectivity of cationic antimicrobial peptides for bacterial versus mammalian membranes. J. Biol. Chem. 280, 33960–33967. doi: 10.1074/jbc.M507042200
Gonzalez-Pastor, J. E., San Millan, J. L., Castilla, M. A., and Moreno, F. (1995). Structure and organization of plasmid genes required to produce the translation inhibitor microcin C7. J. Bacteriol. 177, 7131–7140. doi: 10.1128/jb.177.24.7131-7140.1995
Gordon, Y. J., Huang, L. C., Romanowski, E. G., Yates, K. A., Proske, R. J., and Mcdermott, A. M. (2005). Human cathelicidin (LL-37), a multifunctional peptide, is expressed by ocular surface epithelia and has potent antibacterial and antiviral activity. Curr. Eye Res. 30, 385–394. doi: 10.1080/02713680590934111
Gupta, K., Singh, S., and Van Hoek, M. L. (2015). Short, synthetic cationic peptides have antibacterial activity against Mycobacterium smegmatis by forming pores in membrane and synergizing with antibiotics. Antibiotics 4, 358–378. doi: 10.3390/antibiotics4030358
Havarstein, L. S., Holo, H., and Nes, I. F. (1994). The leader peptide of colicin V shares consensus sequences with leader peptides that are common among peptide bacteriocins produced by Gram-positive bacteria. Microbiology 140, 2383–2389. doi: 10.1099/13500872-140-9-2383
Hazlett, K. R., Cox, D. L., Decaffmeyer, M., Bennett, M. P., Desrosiers, D. C., La Vake, C. J., et al. (2005). Tp0453, a concealed outer membrane protein of Treponema pallidum, enhances membrane permeability. J. Bacteriol. 187, 6499–6508. doi: 10.1128/JB.187.18.6499-6508.2005
Helbing, C. C., Hammond, S. A., Jackman, S. H., Houston, S., Warren, R. L., Cameron, C. E., et al. (2019). Antimicrobial peptides from Rana [Lithobates] catesbeiana: gene structure and bioinformatic identification of novel forms from tadpoles. Sci. Rep. 9:1529. doi: 10.1038/s41598-018-38442-1
Hilchie, A. L., Wuerth, K., and Hancock, R. E. (2013). Immune modulation by multifaceted cationic host defense (antimicrobial) peptides. Nat. Chem. Biol. 9, 761–768. doi: 10.1038/nchembio.1393
Hollands, A., Gonzalez, D., Leire, E., Donald, C., Gallo, R. L., Sanderson-Smith, M., et al. (2012). A bacterial pathogen co-opts host plasmin to resist killing by cathelicidin antimicrobial peptides. J. Biol. Chem. 287, 40891–40897. doi: 10.1074/jbc.M112.404582
Hong, T., Butler, W. R., Hollis, F., Floyd, M. M., Toney, S. R., Tang, Y. W., et al. (2003). Characterization of a novel rapidly growing Mycobacterium species associated with sepsis. J. Clin. Microbiol. 41, 5650–5653. doi: 10.1128/JCM.41.12.5650-5653.2003
Jones, D. T. (1999). Protein secondary structure prediction based on position-specific scoring matrices. J. Mol. Biol. 292, 195–202. doi: 10.1006/jmbi.1999.3091
Juncker, A. S., Willenbrock, H., Von, H. G., Brunak, S., Nielsen, H., and Krogh, A. (2003). Prediction of lipoprotein signal peptides in Gram-negative bacteria. Protein Sci. 12, 1652–1662. doi: 10.1110/ps.0303703
Kelley, L. A., Mezulis, S., Yates, C. M., Wass, M. N., and Sternberg, M. J. (2015). The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 10, 845–858. doi: 10.1038/nprot.2015.053
Kemperman, R., Jonker, M., Nauta, A., Kuipers, O. P., and Kok, J. (2003a). Functional analysis of the gene cluster involved in production of the bacteriocin circularin A by Clostridium beijerinckii ATCC 25752. Appl. Environ. Microbiol. 69, 5839–5848. doi: 10.1128/AEM.69.10.5839-5848.2003
Kemperman, R., Kuipers, A., Karsens, H., Nauta, A., Kuipers, O., and Kok, J. (2003b). Identification and characterization of two novel clostridial bacteriocins, circularin A and closticin 574. Appl. Environ. Microbiol. 69, 1589–1597. doi: 10.1128/AEM.69.3.1589-1597.2003
Khawar, M. B., Abbasi, M. H., and Sheikh, N. (2016). IL-32: a novel pluripotent inflammatory interleukin, towards gastric inflammation, gastric cancer, and chronic rhino sinusitis. Mediators Inflamm. 2016:8413768. doi: 10.1155/2016/8413768
Kindrachuk, J., Jenssen, H., Elliott, M., Nijnik, A., Magrangeas-Janot, L., Pasupuleti, M., et al. (2013). Manipulation of innate immunity by a bacterial secreted peptide: Lantibiotic Nisin Z is selectively immunomodulatory. Innate Immun. 19, 315–327. doi: 10.1177/1753425912461456
Kolaczkowska, E., and Kubes, P. (2013). Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 13, 159–175. doi: 10.1038/nri3399
Kumar, P., Kizhakkedathu, J. N., and Straus, S. K. (2018). Antimicrobial peptides: diversity, mechanism of action and strategies to improve the activity and biocompatibility in vivo. Biomolecules 8:4. doi: 10.3390/biom8010004
Ladjouzi, R., Lucau-Danila, A., Benachour, A., and Drider, D. (2020). A Leaderless two-peptide bacteriocin, Enterocin DD14, is involved in its own self-immunity: evidence and insights. Front. Bioeng. Biotechnol. 8:644. doi: 10.3389/fbioe.2020.00644
Lafond, R. E., and Lukehart, S. A. (2006). Biological basis for syphilis. Clin. Microbiol. Rev. 19, 29–49. doi: 10.1128/CMR.19.1.29-49.2006
Lai, Y., and Gallo, R. L. (2009). AMPed up immunity: how antimicrobial peptides have multiple roles in immune defense. Trends Immunol. 30, 131–141. doi: 10.1016/j.it.2008.12.003
Lata, S., Mishra, N. K., and Raghava, G. P. (2010). AntiBP2: improved version of antibacterial peptide prediction. BMC Bioinformatics 11 Suppl 1:S19. doi: 10.1186/1471-2105-11-S1-S19
Lata, S., Sharma, B. K., and Raghava, G. P. (2007). Analysis and prediction of antibacterial peptides. BMC Bioinformatics 8:263. doi: 10.1186/1471-2105-8-263
Latif, N., Janani, M. K., Sudharshan, Selvamuthu, P., and Dutta Majumder, P. (2020). Triple trouble: A case of retinochoroiditis in a patient with syphilis, tuberculosis, and human immunodeficiency virus infection. Indian J. Ophthalmol. 68, 1995–1997. doi: 10.4103/ijo.IJO_2170_19
Lauth, X., Von Kockritz-Blickwede, M., Mcnamara, C. W., Myskowski, S., Zinkernagel, A. S., Beall, B., et al. (2009). M1 protein allows Group A streptococcal survival in phagocyte extracellular traps through cathelicidin inhibition. J. Innate Immun. 1, 202–214. doi: 10.1159/000203645
Leer, R. J., Van Der Vossen, J. M., Van Giezen, M., Van Noort, J. M., and Pouwels, P. H. (1995). Genetic analysis of Acidocin B, a novel bacteriocin produced by Lactobacillus acidophilus. Microbiology 141, 1629–1635. doi: 10.1099/13500872-141-7-1629
Li, A., Dubey, S., Varney, M. L., Dave, B. J., and Singh, R. K. (2003). IL-8 directly enhanced endothelial cell survival, proliferation, and matrix metalloproteinases production and regulated angiogenesis. J. Immunol. 170, 3369–3376. doi: 10.4049/jimmunol.170.6.3369
Li, C., Sutherland, D., Hammond, S. A., Yang, C., Taho, F., Bergman, L., et al. (2022). AMPlify: Attentive deep learning model for discovery of novel antimicrobial peptides effective against WHO priority pathogens. BMC Genomics 23:77. doi: 10.1186/s12864-022-08310-4
Li, M. F., Zhang, B. C., Li, J., and Sun, L. (2014). Sil: a Streptococcus iniae bacteriocin with dual role as an antimicrobial and an immunomodulator that inhibits innate immune response and promotes S. iniae infection. PLoS One 9:e96222. doi: 10.1371/journal.pone.0096222
Li, W., Deng, W., and Xie, J. (2018). The biology and role of interleukin-32 in tuberculosis. J. Immunol. Res. 2018:1535194. doi: 10.1155/2018/1535194
Lin, B., Hung, A., Li, R., Barlow, A., Singleton, W., Matthyssen, T., et al. (2022). Systematic comparison of activity and mechanism of antimicrobial peptides against nosocomial pathogens. Eur. J. Med. Chem. 231:114135. doi: 10.1016/j.ejmech.2022.114135
Lukehart, S. A., and Marra, C. M. (2007). Isolation and laboratory maintenance of Treponema pallidum. Curr. Protoc. Microbiol. Chapter 12, 12A.1.1–12A.1.18.
Lukehart, S. A., and Miller, J. N. (1978). Demonstration of the in vitro phagocytosis of Treponema pallidum by rabbit peritoneal macrophages. J. Immunol. 121, 2014–2024.
Lukehart, S. A., Hook, E. W., Baker-Zander, S. A., Collier, A. C., Critchlow, C. W., and Handsfield, H. H. (1988). Invasion of the central nervous system by Treponema pallidum: implications for diagnosis and treatment. Ann. Intern. Med. 109, 855–862. doi: 10.7326/0003-4819-109-11-855
Luthra, A., Zhu, G., Desrosiers, D. C., Eggers, C. H., Mulay, V., Anand, A., et al. (2011). The transition from closed to open conformation of Treponema pallidum outer membrane-associated lipoprotein TP0453 involves membrane sensing and integration by two amphipathic helices. J. Biol. Chem. 286, 41656–41668. doi: 10.1074/jbc.M111.305284
Ma, L., Xie, X., Liu, H., Huang, Y., Wu, H., Jiang, M., et al. (2020). Potent antibacterial activity of MSI-1 derived from the magainin 2 peptide against drug-resistant bacteria. Theranostics 10, 1373–1390. doi: 10.7150/thno.39157
Madeira, F., Park, Y. M., Lee, J., Buso, N., Gur, T., Madhusoodanan, N., et al. (2019). The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 47, W636–W641. doi: 10.1093/nar/gkz268
Magnuson, H. J., and Eagle, H. (1948). The minimal infectious inoculum of Spirochaeta pallida (Nichols strain), and a consideration of its rate of multiplication in vivo. Amer. J. Syph. 32, 1–18. doi: 10.1007/978-3-319-23534-9_1
Malaczewska, J., Kaczorek-Lukowska, E., Wojcik, R., Rekawek, W., and Siwicki, A. K. (2019). In vitro immunomodulatory effect of Nisin on porcine leucocytes. J. Anim. Physiol. Anim. Nutr. 103, 882–893. doi: 10.1111/jpn.13085
Mardirossian, M., Sola, R., Beckert, B., Collis, D. W. P., Di Stasi, A., Armas, F., et al. (2019). Proline-rich peptides with improved antimicrobial activity against E. coli, K. pneumoniae, and A. baumannii. Chem. Med. Chem. 14, 2025–2033.
Marra, C., Baker-Zander, S. A., Hook, E. W. D., and Lukehart, S. A. (1991). An experimental model of early central nervous system syphilis. J. Infect. Dis. 163, 825–829. doi: 10.1093/infdis/163.4.825
Matsuzaki, K., Sugishita, K., Fujii, N., and Miyajima, K. (1995). Molecular basis for membrane selectivity of an antimicrobial peptide, Magainin 2. Biochemistry 34, 3423–3429. doi: 10.1021/bi00010a034
McGill, M. A., Edmondson, D. G., Carroll, J. A., Cook, R. G., Orkiszewski, R. S., and Norris, S. J. (2010). Characterization and serologic analysis of the Treponema pallidum proteome. Infect. Immun. 78, 2631–2643. doi: 10.1128/IAI.00173-10
Mead, P. B., and Winn, W. C. (2000). Vaginal-rectal colonization with group A streptococci in late pregnancy. Infect. Dis. Obstet. Gynecol. 8, 217–219. doi: 10.1155/S1064744900000302
Meade, E., Slattery, M. A., and Garvey, M. (2020). Bacteriocins, potent antimicrobial peptides and the fight against multi drug resistant species: resistance is futile? Antibiotics 9:32. doi: 10.3390/antibiotics9010032
Meher, P. K., Sahu, T. K., Saini, V., and Rao, A. R. (2017). Predicting antimicrobial peptides with improved accuracy by incorporating the compositional, physico-chemical and structural features into Chou’s general PseAAC. Sci. Rep. 7:42362.
Michiels, J., Dirix, G., Vanderleyden, J., and Xi, C. (2001). Processing and export of peptide pheromones and bacteriocins in Gram-negative bacteria. Trends Microbiol. 9, 164–168. doi: 10.1016/s0966-842x(01)01979-5
Min, H. J., Yun, H., Ji, S., Rajasekaran, G., Kim, J. I., Kim, J. S., et al. (2017). Rattusin structure reveals a novel defensin scaffold formed by intermolecular disulfide exchanges. Sci. Rep. 7:45282. doi: 10.1038/srep45282
Minami, M., Wakimoto, Y., Matsumoto, M., Matsui, H., Kubota, Y., Okada, A., et al. (2010). Characterization of Streptococcus pyogenes isolated from balanoposthitis patients presumably transmitted by penile-oral sexual intercourse. Curr. Microbiol. 61, 101–105. doi: 10.1007/s00284-010-9581-x
Mishra, A. K., Choi, J., Moon, E., and Baek, K. H. (2018). Tryptophan-rich and proline-rich antimicrobial peptides. Molecules 23:815. doi: 10.3390/molecules23040815
Moazzezy, N., Asadi Karam, M. R., Rafati, S., Bouzari, S., and Oloomi, M. (2020). Inhibition and eradication activity of truncated alpha-defensin analogs against multidrug resistant uropathogenic Escherichia coli biofilm. PLoS One 15:e0235892. doi: 10.1371/journal.pone.0235892
Muller, M., Ewert, I., Hansmann, F., Tiemann, C., Hagedorn, H. J., Solbach, W., et al. (2007). Detection of Treponema pallidum in the vitreous by PCR. Br. J. Ophthalmol. 91, 592–595. doi: 10.1136/bjo.2006.110288
Nelson, G. E., Pondo, T., Toews, K. A., Farley, M. M., Lindegren, M. L., Lynfield, R., et al. (2016). Epidemiology of invasive group A streptococcal infections in the United States, 2005-2012. Clin. Infect. Dis. 63, 478–486. doi: 10.1093/cid/ciw248
Newton, J. A. Jr., Weiss, P. J., Bowler, W. A., and Oldfield, E. C. III (1993). Soft-tissue infection due to Mycobacterium smegmatis: report of two cases. Clin. Infect. Dis. 16, 531–533. doi: 10.1093/clind/16.4.531
Norimatsu, Y., and Ohno, Y. (2020). Streptococcus pyogenes balanoposthitis. ID Cases 21:e00832. doi: 10.1016/j.idcr.2020.e00832
Oman, T. J., and van der Donk, W. A. (2010). Follow the leader: the use of leader peptides to guide natural product biosynthesis. Nat. Chem. Biol. 6, 9–18. doi: 10.1038/nchembio.286
Osbak, K. K., Houston, S., Lithgow, K. V., Meehan, C. J., Strouhal, M., Smajs, D., et al. (2016). Characterizing the syphilis-causing Treponema pallidum ssp. pallidum proteome using complementary mass spectrometry. PLoS Negl. Trop. Dis. 10:e0004988. doi: 10.1371/journal.pntd.0004988
Oyama, L. B., Girdwood, S. E., Cookson, A. R., Fernandez-Fuentes, N., Prive, F., Vallin, H. E., et al. (2017). The rumen microbiome: an underexplored resource for novel antimicrobial discovery. NPJ Biofilms Microbiomes 3:33. doi: 10.1038/s41522-017-0042-1
Pei, J., Kim, B. H., and Grishin, N. V. (2008). PROMALS3D: a tool for multiple protein sequence and structure alignments. Nucleic Acids Res. 36, 2295–2300. doi: 10.1093/nar/gkn072
Pei, J., Mitchell, D. A., Dixon, J. E., and Grishin, N. V. (2011). Expansion of type II CAAX proteases reveals evolutionary origin of gamma-secretase subunit APH-1. J. Mol. Biol. 410, 18–26. doi: 10.1016/j.jmb.2011.04.066
Peng, L. S., Zhuang, Y., Li, W. H., Zhou, Y. Y., Wang, T. T., Chen, N., et al. (2014). Elevated interleukin-32 expression is associated with Helicobacter pylori-related gastritis. PLoS One 9:e88270. doi: 10.1371/journal.pone.0088270
Pennekamp, A., Pfyffer, G. E., Wuest, J., George, C. A., and Ruef, C. (1997). Mycobacterium smegmatis infection in a healthy woman following a facelift: case report and review of the literature. Ann. Plast. Surg. 39, 80–83. doi: 10.1097/00000637-199707000-00014
Perez, R. H., Zendo, T., and Sonomoto, K. (2018). Circular and leaderless bacteriocins: biosynthesis, mode of action, applications, and prospects. Front. Microbiol. 9:2085. doi: 10.3389/fmicb.2018.02085
Petrosova, H., Pospisilova, P., Strouhal, M., Cejkova, D., Zobanikova, M., Mikalova, L., et al. (2013). Resequencing of Treponema pallidum ssp. pallidum strains Nichols and SS14: correction of sequencing errors resulted in increased separation of syphilis treponeme subclusters. PLoS. One 8:e74319. doi: 10.1371/journal.pone.0074319
Pinho-Bandeira, T., Ricoca Peixoto, V., Dias, S., and Sá Machado, R. (2020). Factors associated with coinfection and reinfection by chlamydia, gonorrhoea and syphilis in Portugal. Eur. J. Public Health 30:ckaa165.539. doi: 10.1093/eurpub/ckaa165.539
Pozzobon, T., Facchinello, N., Bossi, F., Capitani, N., Benagiano, M., Di Benedetto, G., et al. (2016). Treponema pallidum (syphilis) antigen TpF1 induces angiogenesis through the activation of the IL-8 pathway. Sci. Rep. 6:18785. doi: 10.1038/srep18785
Qu, X. D., Harwig, S. S., Oren, A. M., Shafer, W. M., and Lehrer, R. I. (1996). Susceptibility of Neisseria gonorrhoeae to protegrins. Infect. Immun. 64, 1240–1245. doi: 10.1128/iai.64.4.1240-1245.1996
Radolf, J. D. (1994). Role of outer membrane architecture in immune evasion by Treponema pallidum and Borrelia burgdorferi. Trends Microbiol. 2, 307–311. doi: 10.1016/0966-842x(94)90446-4
Rasool, S. T., Tang, H., Wu, J., Li, W., Mukhtar, M. M., Zhang, J., et al. (2008). Increased level of IL-32 during human immunodeficiency virus infection suppresses HIV replication. Immunol. Lett. 117, 161–167. doi: 10.1016/j.imlet.2008.01.007
Ribeiro-Dias, F., Saar Gomes, R., De Lima Silva, L. L., Dos Santos, J. C., and Joosten, L. A. (2017). Interleukin 32: a novel player in the control of infectious diseases. J. Leukoc. Biol. 101, 39–52. doi: 10.1189/jlb.4RU0416-175RR
Ronald, A. (2002). The etiology of urinary tract infection: traditional and emerging pathogens. Am. J. Med. 113 Suppl 1A, 14S–19S.
San Millan, J. L., Hernandez-Chico, C., Pereda, P., and Moreno, F. (1985). Cloning and mapping of the genetic determinants for microcin B17 production and immunity. J. Bacteriol. 163, 275–281. doi: 10.1128/jb.163.1.275-281.1985
Sancho-Vaello, E., Francois, P., Bonetti, E. J., Lilie, H., Finger, S., Gil-Ortiz, F., et al. (2017). Structural remodeling and oligomerization of human cathelicidin on membranes suggest fibril-like structures as active species. Sci. Rep. 7:15371. doi: 10.1038/s41598-017-14206-1
Schmidtchen, A., Frick, I. M., Andersson, E., Tapper, H., and Bjorck, L. (2002). Proteinases of common pathogenic bacteria degrade and inactivate the antibacterial peptide LL-37. Mol. Microbiol. 46, 157–168. doi: 10.1046/j.1365-2958.2002.03146.x
Sell, S., Gamboa, D., Baker-Zander, S. A., Lukehart, S. A., and Miller, J. N. (1980). Host response to Treponema pallidum in intradermally-infected rabbits: evidence for persistence of infection at local and distant sites. J. Invest. Dermatol. 75, 470–475. doi: 10.1111/1523-1747.ep12524230
Sellati, T. J., Bouis, D. A., Kitchens, R. L., Darveau, R. P., Pugin, J., Ulevitch, R. J., et al. (1998). Treponema pallidum and Borrelia burgdorferi lipoproteins and synthetic lipopeptides activate monocytic cells via a CD14-dependent pathway distinct from that used by lipopolysaccharide. J. Immunol. 160, 5455–5464.
Shafee, T. M., Lay, F. T., Hulett, M. D., and Anderson, M. A. (2016). The defensins consist of two independent, convergent protein superfamilies. Mol. Biol. Evol. 33, 2345–2356. doi: 10.1093/molbev/msw106
Shen, Y., Maupetit, J., Derreumaux, P., and Tuffery, P. (2014). Improved PEP-FOLD approach for peptide and miniprotein structure prediction. J. Chem. Theory Comput. 10, 4745–4758. doi: 10.1021/ct500592m
Shigemura, K., Arakawa, S., Sakai, Y., Kinoshita, S., Tanaka, K., and Fujisawa, M. (2006). Complicated urinary tract infection caused by Pseudomonas aeruginosa in a single institution (1999-2003). Int. J. Urol. 13, 538–542. doi: 10.1111/j.1442-2042.2006.01359.x
Shockman, S., Buescher, L. S., and Stone, S. P. (2014). Syphilis in the United States. Clin. Dermatol. 32, 213–218.
Simmaco, M., Mignogna, G., Barra, D., and Bossa, F. (1994). Antimicrobial peptides from skin secretions of Rana esculenta. Molecular cloning of cDNAs encoding esculentin and brevinins and isolation of new active peptides. J. Biol. Chem. 269, 11956–11961. doi: 10.1016/s0021-9258(17)32666-2
Smajs, D., Mckevitt, M., Howell, J. K., Norris, S. J., Cai, W. W., Palzkill, T., et al. (2005). Transcriptome of Treponema pallidum: gene expression profile during experimental rabbit infection. J. Bacteriol. 187, 1866–1874. doi: 10.1128/JB.187.5.1866-1874.2005
Smits, S. H. J., Schmitt, L., and Beis, K. (2020). Self-immunity to antibacterial peptides by ABC transporters. FEBS Lett. 594, 3920–3942. doi: 10.1002/1873-3468.13953
Snyder, A. B., and Worobo, R. W. (2014). Chemical and genetic characterization of bacteriocins: antimicrobial peptides for food safety. J. Sci. Food. Agric. 94, 28–44. doi: 10.1002/jsfa.6293
Sobel, J. D., Funaro, D., and Kaplan, E. L. (2007). Recurrent group A streptococcal vulvovaginitis in adult women: family epidemiology. Clin. Infect. Dis. 44, e43–e45. doi: 10.1086/510678
Sornwatana, T., Arpornsuwan, T., Roytrakul, S., and Wetprasit, N. (2018). M-Brucin, an antibacterial peptide against Staphylococcus epidermidis and Streptococcus pyogenes. J. App. Pharm. Sci. 8, 27–32.
Stamm, W. E., Feeley, J. C., and Facklam, R. R. (1978). Wound infections due to group A streptococcus traced to a vaginal carrier. J. Infect. Dis. 138, 287–292. doi: 10.1093/infdis/138.3.287
Standiford, T. J., Kunkel, S. L., Phan, S. H., Rollins, B. J., and Strieter, R. M. (1991). Alveolar macrophage-derived cytokines induce monocyte chemoattractant protein-1 expression from human pulmonary type II-like epithelial cells. J. Biol. Chem. 266, 9912–9918. doi: 10.1016/s0021-9258(18)92905-4
Sugrue, I., O’connor, P. M., Hill, C., Stanton, C., and Ross, R. P. (2020). Actinomyces produces defensin-like bacteriocins (actifensins) with a highly degenerate structure and broad antimicrobial activity. J. Bacteriol. 202:e00529–19. doi: 10.1128/JB.00529-19
Svanborg-Eden, C., Hagberg, L., Hull, R., Hull, S., Magnusson, K. E., and Ohman, L. (1987). Bacterial virulence versus host resistance in the urinary tracts of mice. Infect. Immun. 55, 1224–1232. doi: 10.1128/iai.55.5.1224-1232.1987
Tamames, J., Casari, G., Ouzounis, C., and Valencia, A. (1997). Conserved clusters of functionally related genes in two bacterial genomes. J. Mol. Evol. 44, 66–73. doi: 10.1007/pl00006122
Tamamura, H., Murakami, T., Horiuchi, S., Sugihara, K., Otaka, A., Takada, W., et al. (1995). Synthesis of protegrin-related peptides and their antibacterial and anti-human immunodeficiency virus activity. Chem. Pharm. Bull. 43, 853–858. doi: 10.1248/cpb.43.853
Tenaillon, O., Skurnik, D., Picard, B., and Denamur, E. (2010). The population genetics of commensal Escherichia coli. Nat. Rev. Microbiol. 8, 207–217. doi: 10.1038/nrmicro2298
Thevenet, P., Shen, Y., Maupetit, J., Guyon, F., Derreumaux, P., and Tuffery, P. (2012). PEP-FOLD: an updated de novo structure prediction server for both linear and disulfide bonded cyclic peptides. Nucleic Acids Res. 40, W288–W293. doi: 10.1093/nar/gks419
Todorov, S. D. (2009). Bacteriocins from Lactobacillus plantarum – production, genetic organization and mode of action: produção, organização genética e modo de ação. Braz. J. Microbiol. 40, 209–221. doi: 10.1590/s1517-83822009000200001
Todorov, S. D., De Melo Franco, B. D. G., and Tagg, J. R. (2019). Bacteriocins of Gram-positive bacteria having activity spectra extending beyond closely-related species. Benef. Microbes 10, 315–328. doi: 10.3920/BM2018.0126
Torrent, M., Di Tommaso, P., Pulido, D., Nogues, M. V., Notredame, C., Boix, E., et al. (2012). AMPA: An automated web server for prediction of protein antimicrobial regions. Bioinformatics 28, 130–131. doi: 10.1093/bioinformatics/btr604
Torrent, M., Nogues, V. M., and Boix, E. (2009). A theoretical approach to spot active regions in antimicrobial proteins. BMC Bioinformatics 10:373. doi: 10.1186/1471-2105-10-373
Turner, J., Cho, Y., Dinh, N. N., Waring, A. J., and Lehrer, R. I. (1998). Activities of LL-37, a cathelin-associated antimicrobial peptide of human neutrophils. Antimicrob. Agents Chemother. 42, 2206–2214. doi: 10.1128/AAC.42.9.2206
Uhlmann, J., Rohde, M., Siemens, N., Kreikemeyer, B., Bergman, P., Johansson, L., et al. (2016). LL-37 Triggers formation of Streptococcus pyogenes extracellular vesicle-like structures with immune stimulatory properties. J. Innate Immun. 8, 243–257. doi: 10.1159/000441896
Veltri, D., Kamath, U., and Shehu, A. (2018). Deep learning improves antimicrobial peptide recognition. Bioinformatics 34, 2740–2747. doi: 10.1093/bioinformatics/bty179
Verstraelen, H., Verhelst, R., Vaneechoutte, M., and Temmerman, M. (2011). Group A streptococcal vaginitis: an unrecognized cause of vaginal symptoms in adult women. Arch. Gynecol. Obstet. 284, 95–98. doi: 10.1007/s00404-011-1861-6
Waghu, F. H., Barai, R. S., Gurung, P., and Idicula-Thomas, S. (2016). CAMPR3: a database on sequences, structures and signatures of antimicrobial peptides. Nucleic Acids Res. 44, D1094–D1097. doi: 10.1093/nar/gkv1051
Wallace, R. J. Jr., Nash, D. R., Tsukamura, M., Blacklock, Z. M., and Silcox, V. A. (1988). Human disease due to Mycobacterium smegmatis. J. Infect. Dis. 158, 52–59. doi: 10.1093/infdis/158.1.52
Wang, G., Li, X., and Wang, Z. (2016). APD3: the antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 44, D1087–D1093. doi: 10.1093/nar/gkv1278
Waterhouse, A., Bertoni, M., Bienert, S., Studer, G., Tauriello, G., Gumienny, R., et al. (2018). SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res. 46, W296–W303. doi: 10.1093/nar/gky427
Webb, B., and Sali, A. (2016). Comparative protein structure modeling using MODELLER. Curr. Protoc. Bioinformatics 54, 5.6.1–5.6.37.
Wiegand, I., Hilpert, K., and Hancock, R. E. (2008). Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 3, 163–175. doi: 10.1038/nprot.2007.521
World Health Organization (2019). Sexually Transmitted Infections (STIs). Available online at: https://www.who.int/news-room/fact-sheets/detail/sexually-transmitted-infections-(stis). [Accessed August 16, 2021].
Yamaguchi, Y., Fukuhara, S., Nagase, T., Tomita, T., Hitomi, S., Kimura, S., et al. (2001). A novel mouse beta-defensin, mBD-6, predominantly expressed in skeletal muscle. J. Biol. Chem. 276, 31510–31514. doi: 10.1074/jbc.M104149200
Yasin, B., Harwig, S. S., Lehrer, R. I., and Wagar, E. A. (1996). Susceptibility of Chlamydia trachomatis to protegrins and defensins. Infect. Immun. 64, 709–713. doi: 10.1128/iai.64.3.709-713.1996
Yoshimura, T., Robinson, E. A., Tanaka, S., Appella, E., and Leonard, E. J. (1989a). Purification and amino acid analysis of two human monocyte chemoattractants produced by phytohemagglutinin-stimulated human blood mononuclear leukocytes. J. Immunol. 142, 1956–1962.
Yoshimura, T., Yuhki, N., Moore, S. K., Appella, E., Lerman, M. I., and Leonard, E. J. (1989b). Human monocyte chemoattractant protein-1 (MCP-1). Full-length cDNA cloning, expression in mitogen-stimulated blood mononuclear leukocytes, and sequence similarity to mouse competence gene JE. FEBS Lett. 244, 487–493. doi: 10.1016/0014-5793(89)80590-3
Zendo, T. (2013). Screening and characterization of novel bacteriocins from lactic acid bacteria. Biosci. Biotechnol. Biochem. 77, 893–899. doi: 10.1271/bbb.130014
Zhang, L., Foxman, B., and Marrs, C. (2002). Both urinary and rectal Escherichia coli isolates are dominated by strains of phylogenetic group B2. J. Clin. Microbiol. 40, 3951–3955. doi: 10.1128/JCM.40.11.3951-3955.2002
Keywords: antimicrobial peptides, syphilis, Treponema pallidum, bacteriostatic, bactericidal
Citation: Houston S, Schovanek E, Conway KME, Mustafa S, Gomez A, Ramaswamy R, Haimour A, Boulanger MJ, Reynolds LA and Cameron CE (2022) Identification and Functional Characterization of Peptides With Antimicrobial Activity From the Syphilis Spirochete, Treponema pallidum. Front. Microbiol. 13:888525. doi: 10.3389/fmicb.2022.888525
Received: 03 March 2022; Accepted: 08 April 2022;
Published: 03 May 2022.
Edited by:
Justin David Radolf, UCONN Health, United StatesReviewed by:
Diane Edmondson, University of Texas Health Science Center at Houston, United StatesDarrin Akins, University of Oklahoma Health Sciences Center, United States
Copyright © 2022 Houston, Schovanek, Conway, Mustafa, Gomez, Ramaswamy, Haimour, Boulanger, Reynolds and Cameron. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Caroline E. Cameron, Y2Fyb2NAdXZpYy5jYQ==