
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 02 June 2022
Sec. Food Microbiology
Volume 13 - 2022 | https://doi.org/10.3389/fmicb.2022.884721
This article is part of the Research TopicGenomics and metagenomics approaches for food value chain quality, safety, and product developmentView all 11 articles
 Julio Parra-Flores1*†
Julio Parra-Flores1*† Ondřej Holý2†
Ondřej Holý2† Sergio Acuña3
Sergio Acuña3 Sarah Lepuschitz4†
Sarah Lepuschitz4† Ariane Pietzka4†Alejandra Contreras-Fernández5Pamela Chavarría-Sepulveda1
Ariane Pietzka4†Alejandra Contreras-Fernández5Pamela Chavarría-Sepulveda1 Ariadnna Cruz-Córdova6†
Ariadnna Cruz-Córdova6† Juan Xicohtencatl-Cortes6†
Juan Xicohtencatl-Cortes6† Jetsi Mancilla-Rojano6,7†
Jetsi Mancilla-Rojano6,7† Alejandro Castillo8†
Alejandro Castillo8† Werner Ruppitsch4†
Werner Ruppitsch4† Stephen Forsythe9*†
Stephen Forsythe9*†This study characterized five Cronobacter spp. and six Salmonella spp. strains that had been isolated from 155 samples of powdered infant formula (PIF) sold in Chile and manufactured in Chile and Mexico in 2018–2020. Two strains of Cronobacter sakazakii sequence type (ST) ST1 and ST31 (serotypes O:1 and O:2) and one strain of Cronobacter malonaticus ST60 (O:1) were identified. All Salmonella strains were identified as Salmonella Typhimurium ST19 (serotype O:4) by average nucleotide identity, ribosomal multilocus sequence typing (rMLST), and core genome MLST (cgMLST). The C. sakazakii and C. malonaticus isolates were resistant to cephalothin, whereas the Salmonella isolates were resistant to oxacillin and ampicillin. Nineteen antibiotic resistance genes were detected in the C. sakazakii and C. malonaticus isolates; the most prevalent were mcr-9.1, blaCSA, and blaCMA. In Salmonella, 30 genes encoding for aminoglycoside and cephalosporin resistance were identified, including aac(6′)-Iaa, β-lactamases ampH, ampC1, and marA. In the Cronobacter isolates, 32 virulence-associated genes were detected by WGS and clustered as flagellar proteins, outer membrane proteins, chemotaxis, hemolysins, invasion, plasminogen activator, colonization, transcriptional regulator, survival in macrophages, use of sialic acid, and toxin-antitoxin genes. In the Salmonella strains, 120 virulence associated genes were detected, adherence, magnesium uptake, resistance to antimicrobial peptides, secretion system, stress protein, toxin, resistance to complement killing, and eight pathogenicity islands. The C. sakazakii and C. malonaticus strains harbored I-E and I-F CRISPR-Cas systems and carried Col(pHHAD28) and IncFIB(pCTU1) plasmids, respectively. The Salmonella strains harbored type I-E CRISPR-Cas systems and carried IncFII(S) plasmids. The presence of C. sakazakii and Salmonella in PIF is a health risk for infants aged less than 6 months. For this reason, sanitary practices should be reinforced for its production and retail surveillance.
The need to ensure the safety of powdered infant formula (PIF) led the FAO/WHO to establish the microbiological or epidemiological relationship of microbial agents found in PIF with infant infection. They identified three categories of microorganisms based on evidence of a causal relationship between the presence of these microorganisms and the disease they cause. The first category of microorganisms with clear causality were identified as Cronobacter spp. and Salmonella enterica. The second consists of microorganisms for which causality is possible but has not yet been demonstrated. These were mainly from the Enterobacteriaceae family, but also included Acinetobacter. Finally, the third involves microorganisms for which causality is less likely or has not yet been shown and has not been identified in PIF. Based on this the FAO/WHO recommended the absence of Cronobacter spp. and Salmonella in PIF for target age less than 6 months (FAO/WHO, 2004, 2006; Forsythe, 2018).
Cronobacter is a genus of bacterial pathogens consisting of seven species: C. sakazakii, C. malonaticus, C. universalis, C. turicensis, C. muytjensii, C. dublinensis, and C. condimenti (Iversen et al., 2008; Joseph et al., 2012; Stephan et al., 2014). The species with the greatest clinical significance are C. sakazakii and C. malonaticus and have been reported in cases and outbreaks associated with PIF in infants (Forsythe, 2018; Parra-Flores et al., 2021a). At present in the United States, three cases of illness caused by PIF contaminated with C. sakazakii, resulting in one fatality, and one case of Salmonella Newport are being investigated; this has prompted an international voluntary recall of these PIFs by the manufacture (U.S FDA, 2022). The severity of the clinical condition has been associated with the presence of virulence factors encoded on plasmids (Shi et al., 2018; Aly et al., 2019), adherence and invasion traits (Cruz et al., 2011; Parra-Flores et al., 2018a; Holý et al., 2021), and various other genes such as aut, cpA, fliC, hly, ompA, sip, plas, and inv (Cruz et al., 2011; Franco et al., 2011; Aldubyan et al., 2017; Holý et al., 2019). Other factors are the use of sialic acid as a carbon source, capsule composition and the presence of its capsule, and endotoxin production (Ogrodzki and Forsythe, 2015). Another important aspect is the resistance to β-lactam antibiotics such as cephalothin, cefotaxime, ceftazidime, and ampicillin in addition to the presence of resistance genes such as marA, glpT, ampH, blaCSA, and mcr (Flores et al., 2011; Lee et al., 2012; Fei et al., 2017; Holý et al., 2021).
Salmonella enterica is a gram-negative, rod-shaped, facultative anaerobic genus. More than 2,600 serotypes belonging to S. enterica have been described worldwide que incluyen, which can cause diseases in humans and animals (Mezal et al., 2014). Salmonella is the most widely studied microbial pathogen, and can be isolated from a variety of foods, including PIF associated with disease outbreaks in infants (Angulo et al., 2008; Carrasco et al., 2012; Jourdan-da Silva et al., 2018). Gastroenteric Salmonella infections usual develop as self-limiting gastroenteritis, and antibiotic treatment is necessary only in severe cases more often associated with immunocompromised patients or those at the extremes of age such as infants (de Toro et al., 2014). Therefore, the emergence of strains that are resistant to β-lactams and cephalosporins is a relevant public health and food safety problem (Güerri et al., 2004; de Toro et al., 2011; Wang et al., 2019). In addition, Salmonella exhibits virulence factors that play a decisive role in systemic infections, such as pathogenicity islands (PAIs), invasion and adherence genes, and enterotoxin coding (Murugkar et al., 2003; Nayak et al., 2004; Huehn et al., 2010; Thung et al., 2018).
Salmonella and Cronobacter species are known persist in low-moisture foods such as milk powder and powdered infant formula for up to 2 years (Caubilla-Barron and Forsythe, 2007). Consequently, outbreaks due to the consumption of contaminated products have been reported (Forsythe, 2018; Jones et al., 2019). Cronobacter can survive spray-drying and persist in the manufacturing environment as biofilms. Genotyping has shown the persistence of specific Salmonella and Cronobacter strains within production facilities for many years (Craven et al., 2010; Jones et al., 2019). Contamination may occur post-pasteurization due to the addition of contaminated ingredients (FAO/WHO, 2008).
Whole-genome sequencing (WGS) has facilitated the in-depth study of pathogenic organisms by generating extensive information that helps to determine relationships and taxonomic differences between them (Leopold et al., 2014). It is not only used for isolate identification, but also extensive profiling and genotyping; such as conventional 7-loci multilocus sequence typing (MLST), core genome MLST (cgMLST) and/or single nucleotide polymorphism (SNP) analysis, molecular serotyping, CRISPR-Cas array profiling, and detection of genes associated with antibiotic resistance and virulence. Consequently, more precise epidemiological links can be established (Marraffini, 2013; Leopold et al., 2014; Uelze et al., 2020). Therefore, the analysis of the complete genomes and their comparison enables a more complete analysis of the pathogenesis process of C. sakazakii (Lehner et al., 2018).
In 2017, a recall of powdered formula samples contaminated with Cronobacter occurred in Chile (Parra-Flores et al., 2018b). This situation led to the incorporation of microbial criteria (n = 30; c = 0) for Cronobacter spp. in PIF intended for consumption by infants aged less than 12 months into the Chilean Food Sanitary Regulations (RSA; Parra-Flores et al., 2018b), given that the microbiological criteria in the RSA for Salmonella was already defined (n = 10; c = 0). This study considers the safety of PIF from 2018 to 2020 with the objective of performing a genomic characterization of five Cronobacter spp. and six Salmonella strains isolated from PIF sold in Chile. These PIF had been and manufactured in Chile and Mexico.
A total of 155 PIF samples from two commercial brands whose main ingredient was casein and whey were analyzed. Of these, 80 PIF samples were made in Chile and 75 PIF samples were made in Mexico. The experimental units, milk cans, were obtained monthly from supermarkets and pharmacies because these products are replenished monthly. In addition, this allowed obtaining greater variability in terms of the origin of the production batch.
Cronobacter were isolated according to the method described by Iversen and Forsythe (2004). For each sample, 225 ml buffered peptone water (BPW) was added to 25 g PIF, homogenized in a stomacher at a mean velocity for 60 s, and incubated at 37°C. For Cronobacter spp., 10 ml of each sample was inoculated after incubation at 37°C for 24 h in 90 ml Enterobacteriaceae enrichment broth (BD Difco, Sparks, MD, United States). A loop was extracted from the culture suspension and striated in Brilliance Chromogenic Agar CM 1035 (OxoidTermo-Fisher, Hampshire, United Kingdom) at 37°C for 20 h. Five strains, presumed to be colonies of Cronobacter spp. (green or blue), were striated in trypticase soy agar (BD Difco, Sparks, MD, United States) to verify their purity prior to future analyses. The isolated strains were maintained in a strain collection and stored at −80°C.
The official method (Instituto Nacional de Normalización, 2002) NCh 2675- ISO 6572-2 rev 2017 for Salmonella in Chile was used. From the initial incubated sample of 25 g PIF with 225 ml BPW for 24 h, 0.1 ml was inoculated in 10 ml of Rappaport-Vassiliadis with soya broth (RVS, Oxoid, Hampshire, United Kingdom) and 1 ml in Muller-Kauffmann-Tetrathionate-Novobiocin broth (MKTTn, Merck, Darmstadt, Germany) incubated for 24 ± 3 h at 41.5°C and 37°C, respectively. The colonies were then isolated in Xylose Lysine Deoxycoholate (XLD, Merck, Darmstadt, Germany) and Salmonella chromogenic agar incubated at 37°C for 24 h. The typical colonies were confirmed by biochemical tests. Both pathogens were identified by Matrix-Assisted Laser Desorption Ionization Time of Flight Mass Spectrometry (MALDI-TOF MS; Bruker, Billerica, MA, United States) and with the MBT Compass IVD software 4.1.60 (Bruker) described by Lepuschitz et al. (2017).
Before WGS, all the Cronobacter spp. and Salmonella spp. strains were cultured in Columbia blood agar plates (bioMérieux, Marcy-l’Étoile, France) at 37°C for 24 h. DNA was isolated from bacterial cultures with the MagAttract HMW DNA Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. The amount of DNA was quantified on a Lunatic instrument (Unchained Labs, Pleasanton, CA, United States). Nextera XT chemistry (Illumina Inc., San Diego, CA, United States) was used to prepare sequencing libraries for a 2 × 300 bp paired-end sequencing run on an Illumina MiSeq sequencer. Samples were sequenced to achieve a minimum of 80-fold coverage using standard protocols by Illumina. The resulting FASTQ files were quality trimmed and de novo assembled with the SPAdes version 3.9.0. Contigs were filtered for a minimum of 5-fold coverage and 200 bp minimum length with Ridom SeqSphere+ software v. 7.8.0 (Ridom, Münster, Germany; Jünemann et al., 2013).
A total of 3,678 targets were used to establish the core genome multilocus sequence typing (cgMLST) scheme of Cronobacter spp. using strain ATCC BAA-894 as a reference using Ridom SeqSphere+ software v. 7.8.0 (Ridom, Münster, Germany; Jünemann et al., 2013). For Salmonella, the cgMLST scheme was performed based on the profile of 2,969 S. enterica target gene loci task template of the Ridom SeqSphere+ software v. 7.8.0 (Ridom, Münster, Germany). According to the cgMLST scheme, isolates were visualized with a minimum spanning tree (MST) to establish their genotypic relationships (Lepuschitz et al., 2019). In addition, the sequences of the seven housekeeping genes of the conventional MLST for Cronobacter spp. and Salmonella were extracted and cross-checked against the Cronobacter MLST database1 (Baldwin et al., 2009) and Salmonella MLST2 (Achtman et al., 2012), respectively. The Cronobacter strains are ID 3409–3413 in the Cronobacter PubMLST database and Salmonella are ID RID389119–RID389124 in the cgMLST database.
The gnd and galF genes that are specific to the Cronobacter serotype O region was determined by WGS sequence analysis with the BIGSdb tool available in the PubMLST database3 and CroTrait WGS analysis (Wang L. et al., 2021). For Salmonella, the SeqSero 1.2 tool available at https://cge.cbs.dtu.dk/services/SeqSero/ was used (Zhang et al., 2015).
The disk diffusion method was used in accordance with the recommendations of the Clinical and Laboratory Standards Institute (CLSI, 2020). The commercial disks that were used consist of ampicillin (10 μg), amikacin (30 μg), cephalothin (30 μg), chloramphenicol (30 μg), ceftriaxone (30 μg), cefotaxime (30 μg), cefepime (30 μg), gentamicin (10 μg), levofloxacin (5 μg), netilmicin (30 μg), oxacillin (1 μg), and sulfamethoxazole-trimethoprim (1.25/23.75 μg). The characterization of the resistance/susceptibility profiles was determined according to the CLSI guidelines. The Escherichia coli ATCC 25922 and Pseudomonas aeruginosa ATCC 27853 strains were used as references.
The existence of virulence genes was confirmed by applying the task template function in SeqSphere+ for the WGS data and the ResFinder tool from the Center of Genomic Epidemiology (CGE).4 Thresholds for the target scanning procedure were set with a required identity of ≥90% to the reference sequence and an aligned reference sequence ≥99%. The Comprehensive Antibiotic Resistance Database (CARD) with the “perfect” and “strict” default settings for sequence analysis (Jia et al., 2017), the Task Template AMRFinderPlus 3.2.3 available in Ridom SeqSphere+ v. 7.8.0 software using the EXACT method at 100%, and BLAST alignment for protein identification available in the AMRFinderPlus database were used for antimicrobial resistance genes.
The PlasmidFinder 2.1 and MobileElementFinder 1.0 tools were used to detect plasmids and mobile genetic elements (MGEs). The selected minimum identity was 95% and 90%, respectively (Carattoli et al., 2014; Johansson et al., 2021).4
The search and characterization of CRISPR arrays and their association with Cas proteins was determined with CRISPRDetect and CRISPRminer (available at http://crispr.otago.ac.nz/CRISPRDetect/predict_crispr_array.html and http://www.microbiome-bigdata.com/CRISPRminer; Biswas et al., 2016; Zhang et al., 2018). The following parameters were applied: 18–55 pb repeated sequence length, 25–60 pb spacer length, 0.6–2.5 spacer sequence size as a function of repeated sequence size, and 60% maximum percentage similarity between spacers. The PHASTER program (available at https://phaster.ca/) was used to identify sequences associated with prophages within the study genomes, and the phages associated with the spacer sequences were determined with the CRISPRminer program. The types of CRISPR systems were determined with the CRISPRmap program (Lange et al., 2013). The CRISPRTarget program was used to determine the PAM (protospacer adjacent motif) sequences associated with each of the repeated sequences of the identified arrays.
Overall positivity for the Cronobacter spp. samples was 6.25% (5/80) and 2.7% (2/75) for Salmonella. Of the five Cronobacter spp. presumptive strains, four were identified as C. sakazakii and one as C. malonaticus. All strains were isolated from different PIF batches from the same manufacturer and country (Chile). The six Salmonella strains were identified as Salmonella Typhimurium. These were from two different batches and tins but same manufacturer and country (Mexico; Table 1).
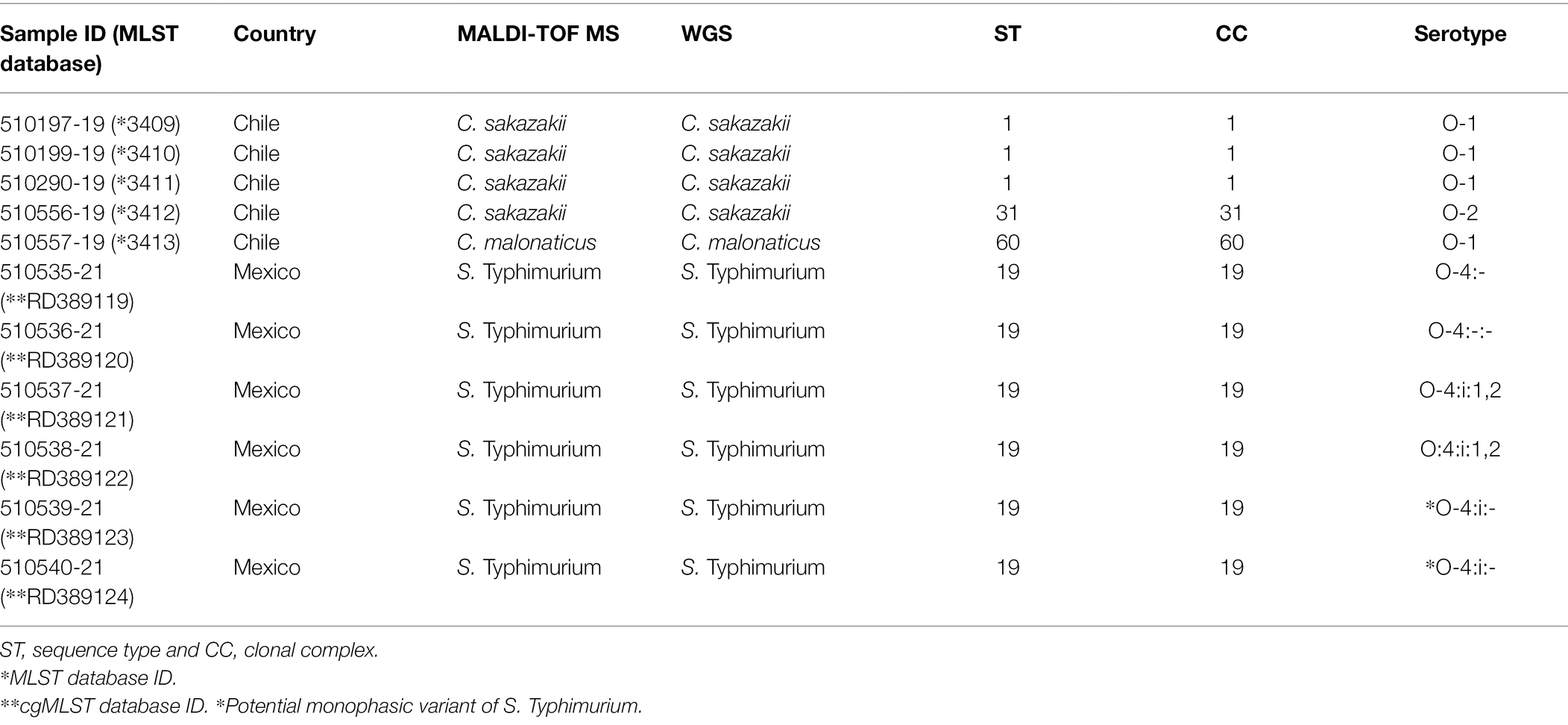
Table 1. Identification of Cronobacter spp. and Salmonella spp. strains isolated from powdered infant formula by matrix-assisted laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF MS) and whole-genome sequencing (WGS).
Three strains of C. sakazakii ST1 (CC1) and ST31, CC31 (serotypes Csak: O:1 and O:2, respectively), and one strain of C. malonaticus ST60, CC60 (O:1) were identified by average nucleotide identity, rMLST, and cgMLST (Figure 1; Table 1).
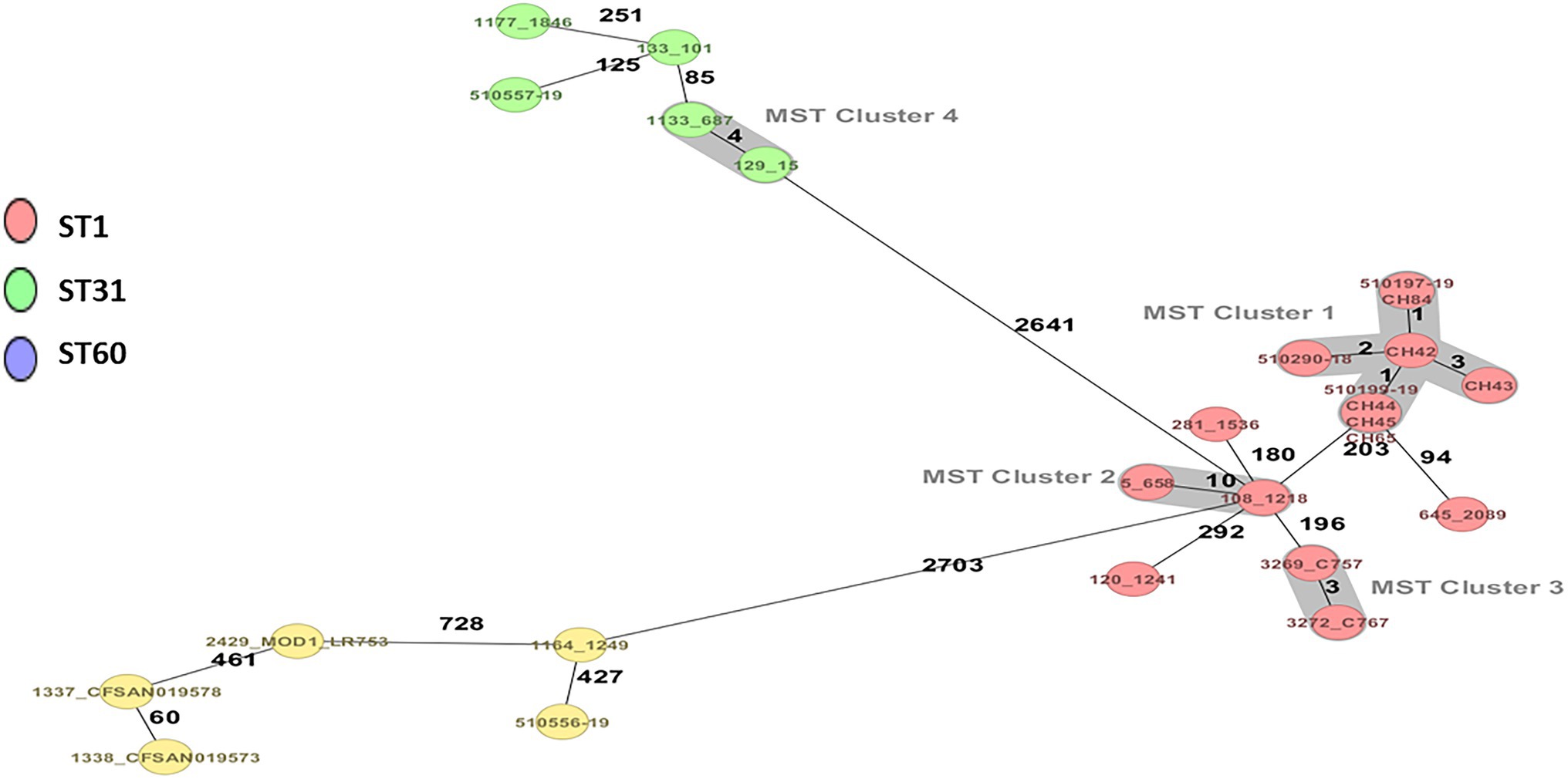
Figure 1. Minimum spanning tree (MST) of five strains of Cronobacter sakazakii and one of Cronobacter malonaticus from powdered infant formula manufactured in Chile, complemented with strains of C. sakazakii and C. malonaticus ST1, ST31, and ST60 of clinical and food origin. Calculation of the MST was based on the defined cgMLST scheme comprising 3,678 target genes for C. sakazakii and C. malonaticus. Isolates are represented as colored circles according to the classical MLST. Black numbers according to the allelic differences between isolates. Isolates with closely related genotypes are marked as Cluster.
All Salmonella strains were identified as S. Typhimurium ST19 (CC19; serotype O:4). Two Salmonella strains (510539-21 and 510540-21) were identified as potential monophasic variants of S. Typhimurium according to SeqSero analysis (Figure 2; Table 1).
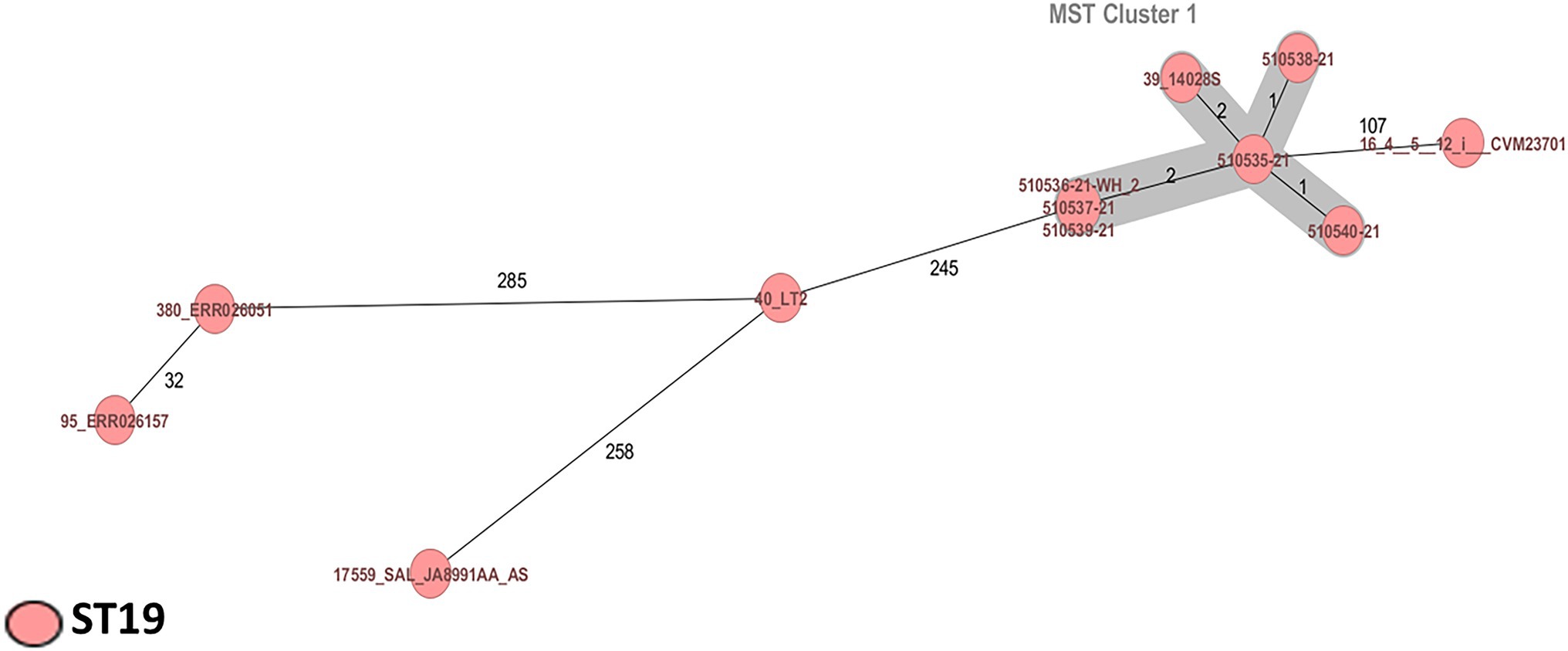
Figure 2. Minimum spanning tree of six Salmonella Typhimurium strains from powdered infant formula manufactured in Mexico and supplemented with other S. Typhimurium ST19 strains of clinical and food origin. Calculation of the MST was based on the defined cgMLST scheme comprising 3,002 target genes for Salmonella. Isolates are represented as colored circles according to the classical MLST. Black numbers according to the allelic differences between isolates. Isolates with closely related genotypes are marked as Cluster.
All the C. sakazakii and C. malonaticus strains were susceptible to 10 of the 12 evaluated antibiotics. However, 100% of the Cronobacter strains were resistant to cephalothin and 40% to ampicillin. Meanwhile, 100% of the Salmonella isolates were resistant to oxacillin, 83% to ampicillin, 66.6% to cephalothin, and 16.6% to gentamicin (Table 2).
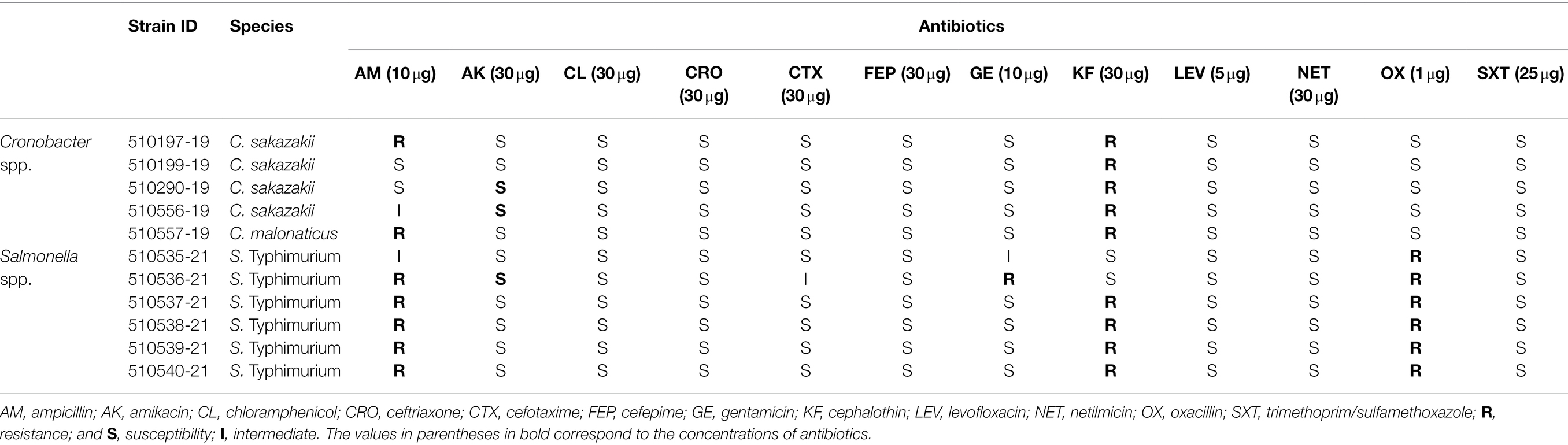
Table 2. Antibiotic resistance profile of Cronobacter spp. and Salmonella spp. strains.
A total of 19 antibiotic resistance genes were detected in the C. sakazakii and C. malonaticus isolates. All the C. sakazakii exhibited blaCSA-1 and the C. malonaticus strain showed blaCMA-1, conferring resistance to cephalosporins. Both C. sakazakii ST1 strains harbored the mcr-9.1 gene, conferring resistance to colistin. All the C. sakazakii and C. malonaticus strains exhibited the same efflux genes (adeF, H-NS, msbA, marA, kpnF, kpnE, emrR, emrB, rsmA, and CRP), antibiotic inactivation gene (ampH), and four antibiotic target alteration genes (pBP3, glpT, eF-Tu, and marR; Table 3).
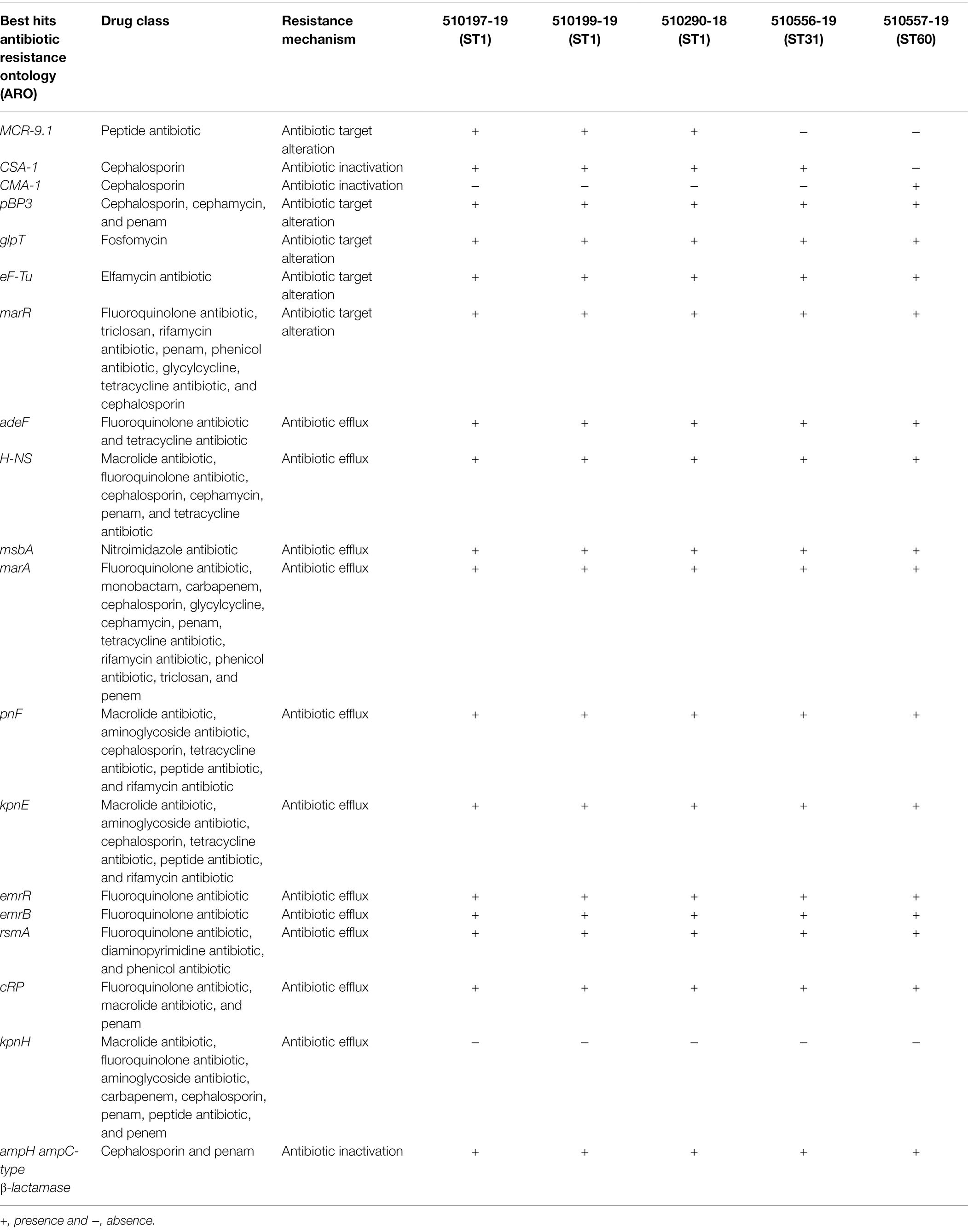
Table 3. Antibiotic-resistant genes of Cronobacter spp. strains identified by Comprehensive Antibiotic Resistance Database (CARD).
In Salmonella, 30 genes that encode for aminoglycoside and cephalosporin resistance were identified, including aac(6′)-Iaa, ampH, ampC1, and marA. All the strains exhibited the same efflux genes (acrAB, golS, mdsA, adeF, marA, kpnF, kpnE, emrRB, rsmA, baeR, H-NS, sdiA, mdfA, mdtK, and kdpE), three antibiotic resistance genes (aac(6′)-Iaa, β-lactamase ampH, and ampC1), and nine antibiotic target alteration genes (bacA, pmrF, uhpT, glpT, PBP3, EF-Tu, soxS, soxR, and marR; Table 4).
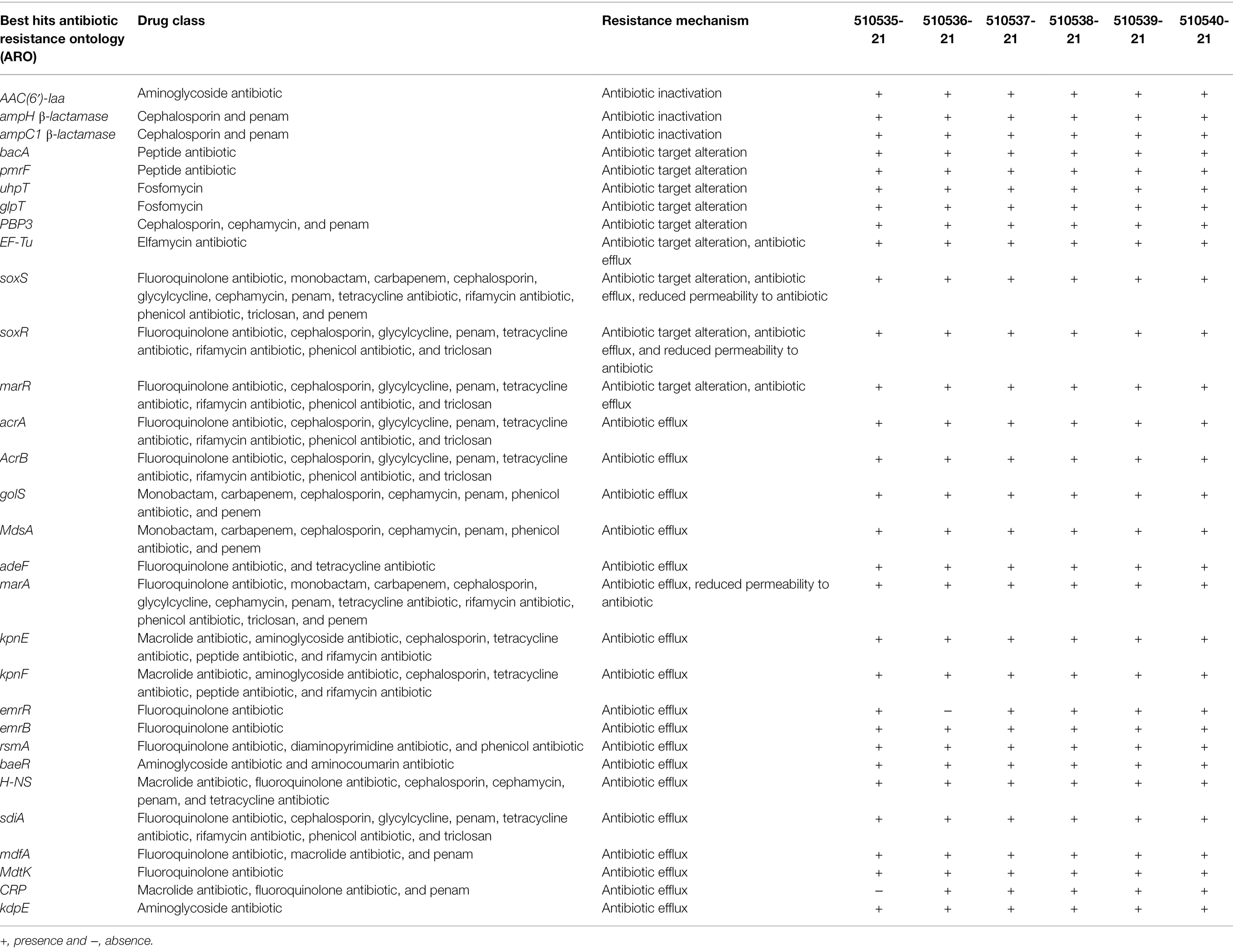
Table 4. Antibiotic-resistant genes of S. Typhimurium strains identified by CARD.
Cronobacter sakazakii isolates showed 32 virulence genes that were detected by WGS and clustered as flagellar proteins, outer membrane proteins, chemotaxis, hemolysins, invasion, plasminogen activator (cpa), colonization, transcriptional regulator, survival in macrophages, utilization of sialic acid (nanA,K,T), desiccation tolerance (cheB, wzzB), and toxin-antitoxin genes (fic, relB). In the C. malonaticus strain, the same virulence genes were detected as found in C. sakazakii, except for the cpa and nanAK,T genes (Table 5).
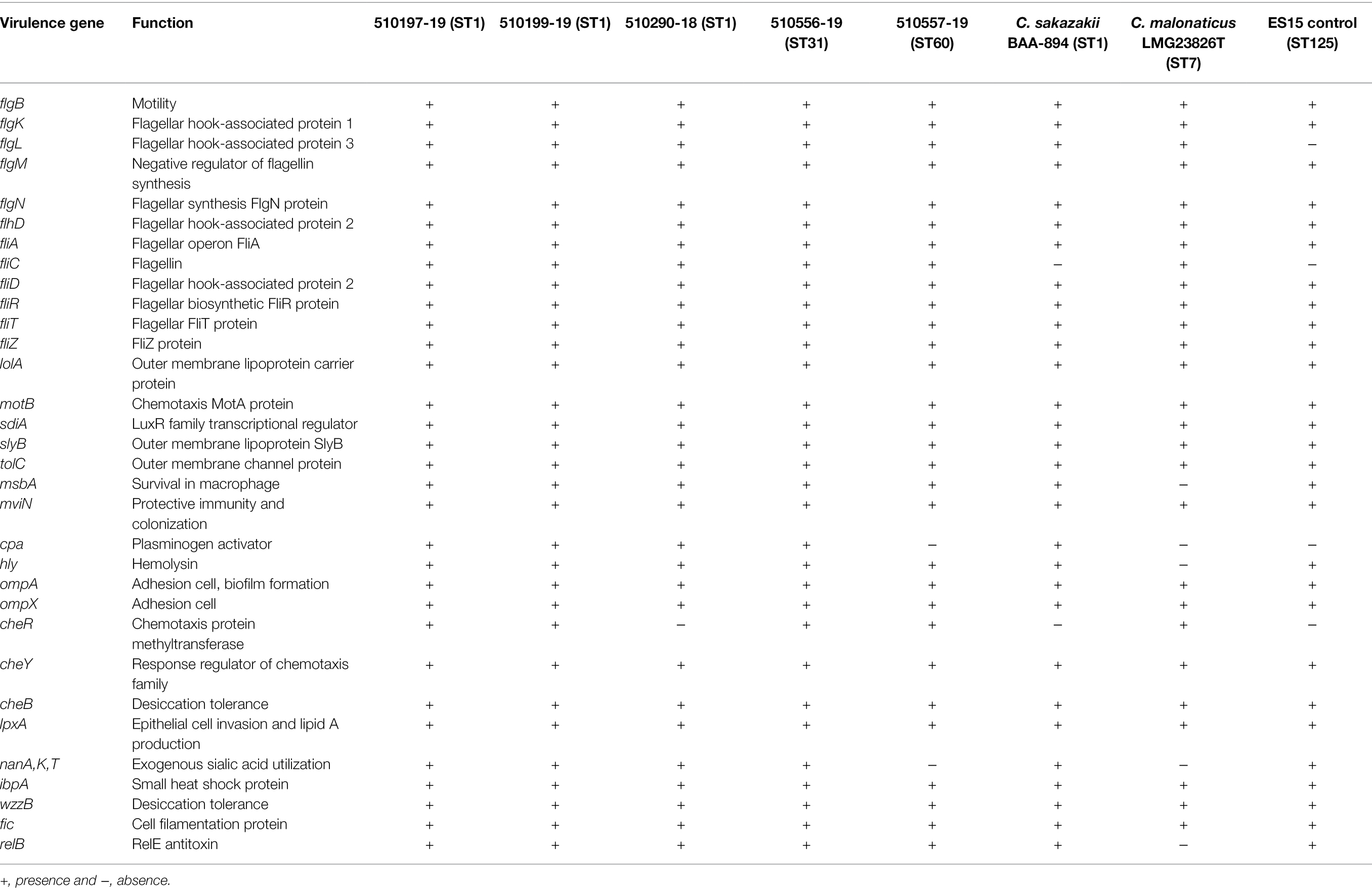
Table 5. Putative virulence and distribution of other genes in seven strains of Cronobacter spp. by WGS.
In the Salmonella strains, 120 virulence genes and eight pathogenicity islands were detected. The virulence genes clustered as adherence, magnesium uptake, resistance to antimicrobial peptides, secretion system, stress protein, toxin, resistance to complement killing. The shdA gene associated with persistence of the bacteria in the intestine was only present in the 510535-21 strain. The gogB, pipB, ssaCTU, ssel/srfH, sseL, sspH2, shdA, sopD2, and sseK1,2 genes were not found in the 510536-21, 510537-21, 510538-21, 510539-21, and 510540-21 strains associated with the secretion system, effector proteins, adherence, and host survival (Table 6).
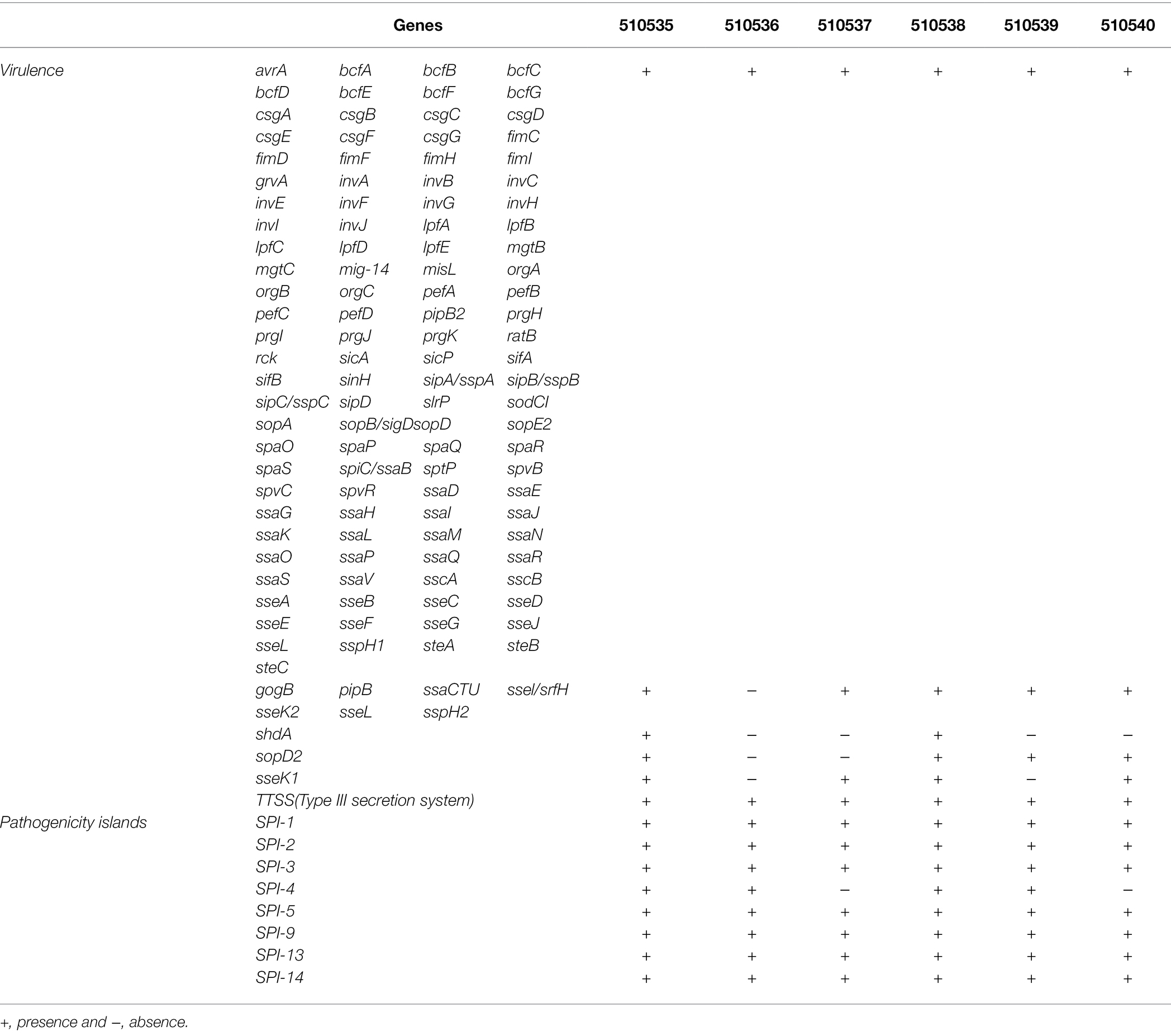
Table 6. Putative virulence and distribution of other genes in six strains of S. Typhimurium by WGS.
The Col(pHHAD28) plasmids and seven MGEs (IS903, IS26, ISEsa2, IS5075, ISEsa1, ISPpu12, and IS102) were detected in only three C. sakazakii strains. The IncFIB(pCTU1) plasmid and one MGE (IS481) were detected in the C. malonaticus strain.
All the Salmonella strains exhibited the IncFII(S) plasmids and five similar MGEs (ISSen7, ISSty2, ISEcI10, MITEEcl, and ISSen1; Table 7).
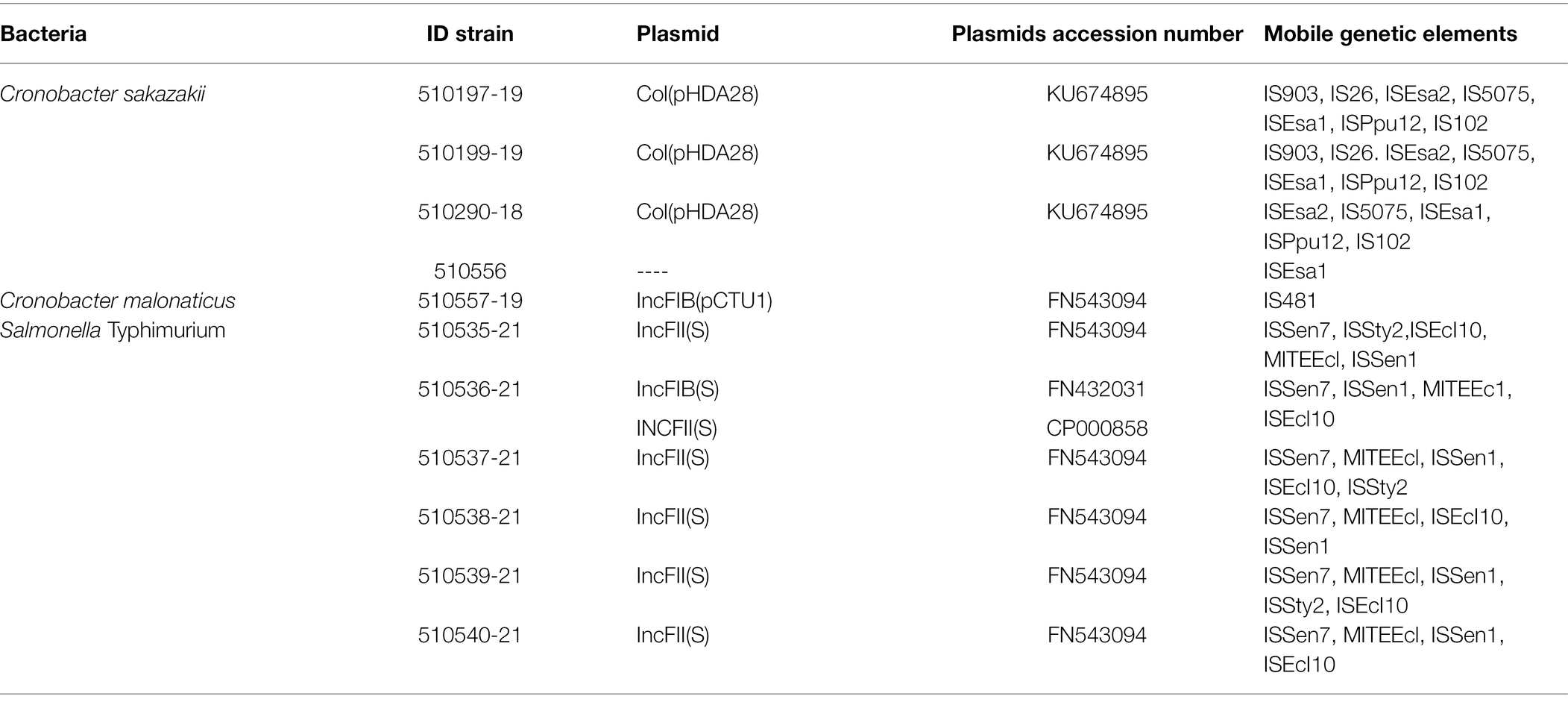
Table 7. Plasmids and mobile genetic elements of Cronobacter spp. and S. Typhimurium.
Genome analysis showed CRISPR-Cas systems in all of the genomes. In Cronobacter spp., 80% (n = 4/5) of the genomes revealed the presence of up to three arrays, which were characterized by the same repeated sequences but at different positions in the genome (Table 8). In the case of Salmonella spp. isolates, 100% (n = 6/6) of the genomes showed two arrays associated with the CRISPR-Cas systems in different positions but characterized by the same number of repeated sequences and spacers, with up to 28 repeated sequences and 27 spacers.
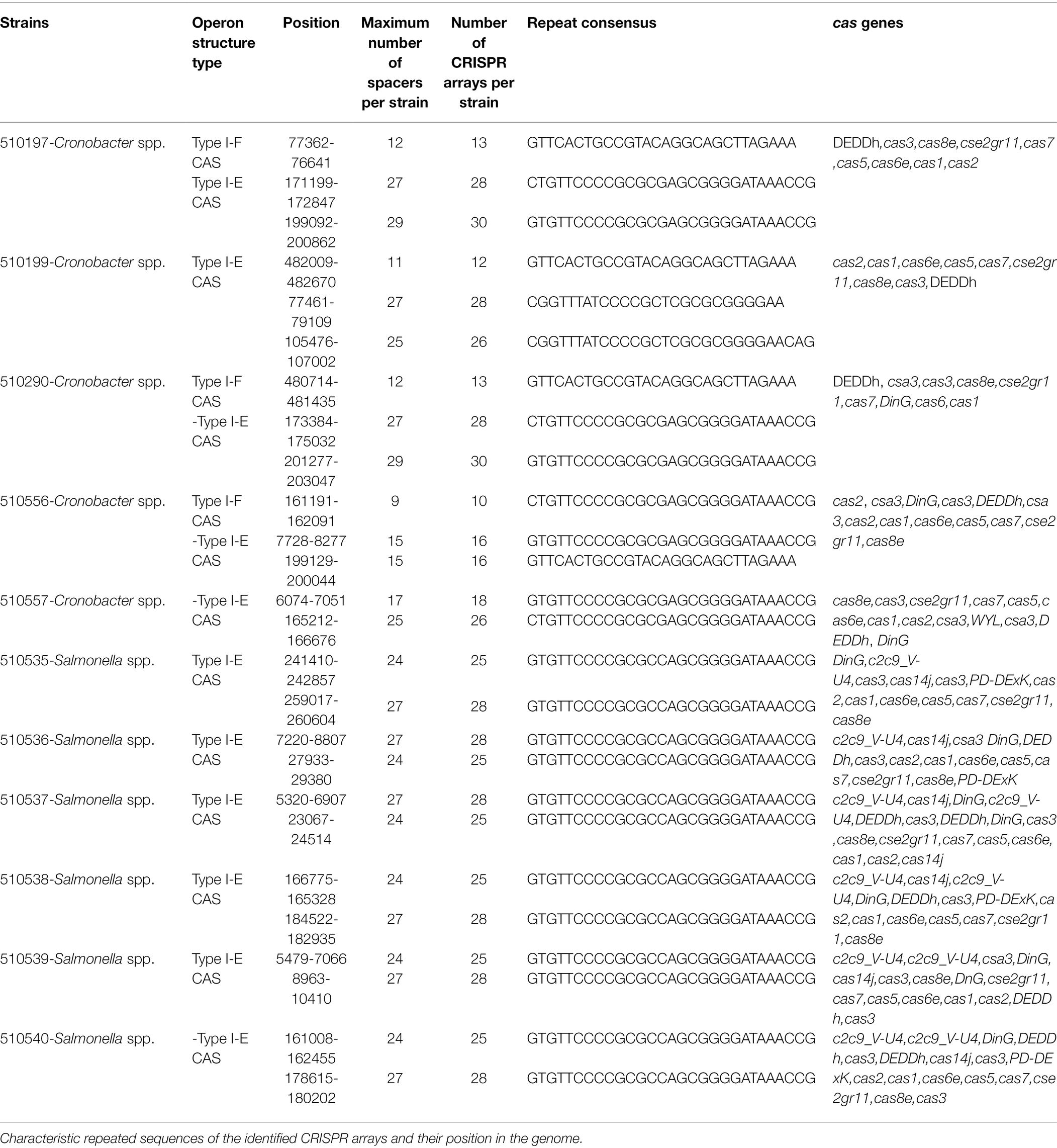
Table 8. CRISPR-Cas systems identified in the Cronobacter spp. and Salmonella spp. genomes.
Using the CRISPRmap program, the repeated sequences and associated cas genes allowed us to determine that the CRISPR systems identified in Cronobacter spp. genomes belonged to type I-E and I-F in which 60% (n = 3/5) were characterized by the presence of both types of CRISPR-Cas systems. However, the opposite was observed in the Salmonella genomes, which were associated with the presence of type I-E systems. As for the associated cas genes, type I-E systems of the Salmonella genomes showed a larger number of genes associated with the CRISPR arrays (Figure 3).
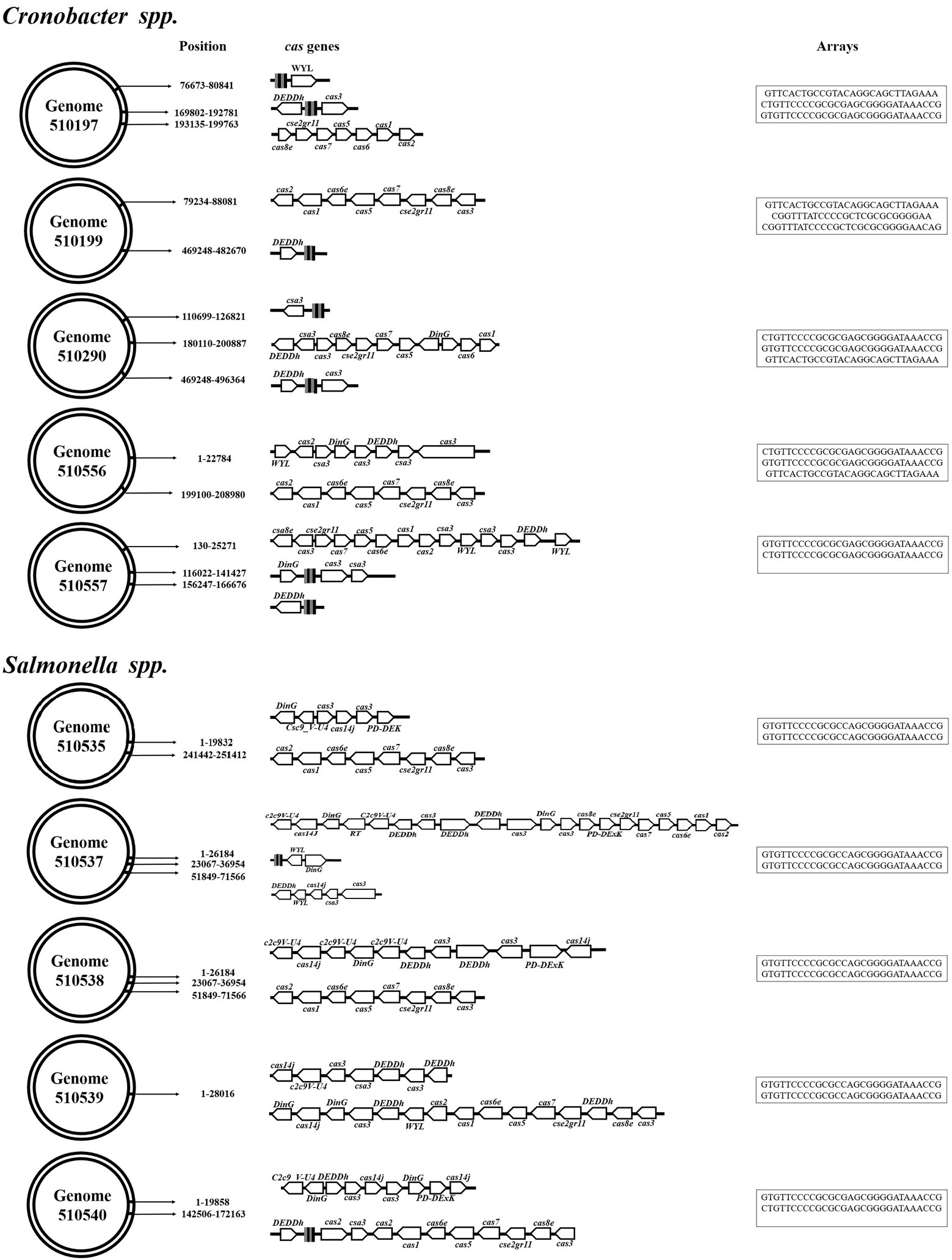
Figure 3. CRISPR-Cas systems identified in Cronobacter spp. and Salmonella spp. genomes. The identified systems belong to the I-E and I-F CRISPR-Cas systems.
The analysis of the spacers with CRISPRminer revealed bacteriophage sequences with bacteriophages that are characteristic of Salmonella and Enterobacteriaceae in three Cronobacter spp. genomes (Supplementary Table 1). However, when searching for the PAM sequences, the spacers of the CRISPR arrays identified in the Cronobacter genomes were also characterized by phages associated with the Klebsiella, Streptococcus, and Acinetobacter genera. The spacers of the Salmonella spp. genomes were associated with the crAss and Acanthamoeba polyphaga moumouvirus phages and bacteriophages that are characteristic of Salmonella and E. coli (Supplementary Table 2). These results are correlated with the phages that were identified in the study genomes (Supplementary Table 3).
WGS enables the molecular typing of bacterial strains on a routine basis for use in epidemiology and real-time infection control. Its usefulness has also been demonstrated in identifying antimicrobial resistance markers, virulence, and the genetic prediction of antibiotic susceptibility test results (Lepuschitz et al., 2020).
In the present study, we identified and characterized C. sakazakii ST1 and ST31, C. malonaticus ST60, and S. Typhimurium ST19 strains from commercially available PIF. The three strains of the pathovar C. sakazakii ST1 were isolated from different batches of PIF which came same manufacturer. This may reflect either the widespread occurrence of this sequence type in PIF manufacturing environment or that a common ingredient was contaminated with the same strain (Sonbol et al., 2013). Cronobacter sakazakii ST1 has frequently been found in PIF commercialized in different countries, in the PIF processing environment, and in invasive clinical cases such as fatal meningitis and septicemia (Joseph and Forsythe, 2012; Sonbol et al., 2013; Fei et al., 2015; Csorba et al., 2021; Holý et al., 2021; Parra-Flores et al., 2021b). A survey of Cronobacter in the Americas showed the majority of reports isolations were from North America (57.4%, n = 465) and Brazil (42.6%, n = 465). There were a total of 75 sequence types, with the most frequently reported being the C. sakazakii pathovars ST4 (CC4) and ST1 (CC1; Costa et al., 2021).
The cgMLST scheme analysis clustered the three C. sakazakii ST1 strains closely to the C. sakazakii ST1 strains isolated from the food alert in Chile on 2017 with one to three alleles differences (Parra-Flores et al., 2018b). When analyzing the isolated strains of human infections in a European multicenter study using cgMLST, eight C. sakazakii ST1 isolates were found among all the C. sakazakii strains. Of these eight ST1 isolates, two strains were analyzed from an outbreak affecting two newborns suffering from necrotizing enterocolitis in Austria in 2009, which differed by only one allele. In addition, three ST1 isolates from Austria and one ST1 from Denmark differed by 203 alleles with the ATCC BAA-894 strain, which was isolated from PIF associated with a fatal case of an infant in the United States in 2001. Cronobacter sakazakii ST31 has also been isolated from clinical cases with fatal outcomes; however, it has been less prevalent in PIF and the environment (Sonbol et al., 2013; Fei et al., 2015; Ogrodzki and Forsythe, 2015; Lepuschitz et al., 2019). In this context, C. sakazakii ST1, ST8, and especially ST4 are the STs with the highest risk of causing disease in infants (Joseph and Forsythe, 2011; Forsythe, 2018; Lachowska et al., 2021). Cronobacter malonaticus ST60 has also been isolated in powdered milk, food, and the environment, but with less significant clinical cases than C. malonaticus ST7 and C. sakazakii ST1 (Forsythe, 2018; Costa et al., 2021).
Salmonella enterica is the most frequently identified cause of food poisoning in the European Union; serotype Typhimurium ST19 is most often associated with disease and death (Carroll et al., 2017; de Frutos et al., 2018) and commonly identified in human clinical samples, animals, food, and the environment (Panzenhagen et al., 2018; Monte et al., 2020). The cgMLST scheme analysis revealed a cluster of six S. Typhimurium ST19 with one to two allele differences. In this context, the present study is the first to identify S. Typhimurium ST19 in PIF, whereas recent reports of Salmonella outbreaks in PIF have involved S. Agona (Brouard et al., 2007; Jourdan-da Silva et al., 2018). We also presumptively identified two Salmonella monophasic variant strains. Monophasic Salmonella has been identified in different human cases in the United States, Spain, Brazil, and Thailand and characterization of these strains revealed resistance to multiple antibiotics (Mossong et al., 2007).
All five C. sakazakii strains analyzed in the present study were resistant to cephalothin and only two strains to ampicillin. The resistance of C. sakazakii to cephalothin and ampicillin has been reported in several studies, also suggesting an almost intrinsic resistance of the Cronobacter spp. genus to cephalothin (Kim et al., 2008; Molloy et al., 2009; Flores et al., 2011; Chon et al., 2012). Parra-Flores et al. (2020) found that 100% of the C. sakazakii strains isolated in powdered infant formula distributed in Latin America were resistant to cefotaxime and ampicillin, 60% to cefepime, 40% to amikacin, and 20% to cephalothin. Furthermore, one of the strains was resistant to six of the 12 evaluated antibiotics and another strain was resistant to five antibiotics. Multiple drug resistance (MDR) is a cause for concern; in a case of neonatal meningitis caused by C. sakazakii in China, the isolated strains were resistant to eight antibiotics (Zeng et al., 2018).
For the antibiotic resistance genes, all C. sakazakii strains and the C. malonaticus strain exhibited the same efflux, antibiotic inactivation, and antibiotic target alteration genes that confer antibiotic resistance to β-lactams, fluoroquinolones, aminoglycosides, imidazoles, and disinfectants such as triclosan. The marA gene, whose transcription function regulates multidrug efflux, modulates membrane permeability and activates the transcription of the AcrAB-TolC efflux pump that plays an important role in antibiotic resistance (Wang X. et al., 2021). Several authors have detected the msbA, emrR, H-NS, emrB, marA, CRP, PBP3, H-NS, and msrB genes that are associated with antibiotic resistance by using efflux pumps, regulatory systems, and antibiotic target protection genes (Aly et al., 2019; Lepuschitz et al., 2019; Parra-Flores et al., 2021b). All strains exhibited the glpT gene that encodes fosfomycin resistance. This is relevant because fosfomycin is considered effective in patients with MDR bacterial infections (Falagas et al., 2019). We also identified the mcr-9.1 gene that confers resistance to colistin and the blaCSA and blaCMA genes that confer resistance to cephalothin. The mcr-9.1 gene can generate resistance to colistin in several enteropathogens and can silently circulate undetected unless induced by colistin (Carroll et al., 2019; Kieffer et al., 2019). The presence of mobile genes resistant to colistin (mcr) is a worldwide public concern because colistin is considered as a last resort to treat infections caused by multidrug-resistant Enterobacteriaceae (Borowiak et al., 2020). The blaCSA and blaCMA genes were first described in 2014 and are associated with β-lactamase class C resistance; they are not inducible and are regarded as cephalosporinases (Müller et al., 2014). Jang et al. (2020) found variants of class C bla resistance genes identified as CSA-2 or CSA-1 and CMA in all the analyzed strains. Holý et al. (2021) encountered blaCSA genes in all the C. sakazakii strains isolated in powdered milk produced in the Czech Republic between 2010 and 2014.
In the present study, all six S. Typhimurium ST19 strains were resistant to oxacillin, five to ampicillin, four to cephalothin, and one to gentamicin. S. Typhimurium ST19 has shown extensive resistance to a variety of critically important antimicrobials (Monte et al., 2020). Jain et al. (2018) reported that all the evaluated S. Typhimurium ST19 strains were resistant to 7 of the 15 tested antibiotics and encountered only strains that were susceptible to ampicillin and gentamicin; this contrasts with our study in which strains were resistant to these two antibiotics.
The Salmonella strains carried many genes such as aac(6′)-Iaa that encodes resistance to aminoglycosides (e.g., gentamicin), which have been found in multidrug-resistant S. Typhimurium strains and caused a foodborne outbreak at a banquet in China in 2017 (Xiang et al., 2020). Wei et al. (2019) reported the presence of β-lactamase ampC genes the same as in our study; in addition to finding β-lactamase ampC-1, we found that it co-harbored with β-lactamase ampH. The β-lactamase ampC is chromosomally encoded in several gram-negative bacteria, including Enterobacter spp., Citrobacter freundii, or Serratia marcescens. Furthermore, plasmid-encoded ampC genes can be horizontally transferred to other Enterobacteriaceae without the presence of chromosomally encoded ampC, such as Klebsiella and Salmonella, resulting in a highly effective and dynamic dissemination mechanism (Ingram et al., 2011). We also found the uhpT and glpT genes that encode fosfomycin resistance. The UhpT and GlpT transporters facilitate fosfomycin incorporation into bacterial cells (Silver, 2017). The reduced expression or introduction of glpT or uhpT mutations and the efflux pump can decrease fosfomycin uptake and thus lower antibiotic susceptibility (Garallah and Al-Jubori, 2020). Several studies have now shown that the overuse of antibiotics in the food chain and the presence of several antibiotic resistance operons promote multiresistance to these drugs (Ferri et al., 2017).
Among the 30 virulence genes detected in C. sakazakii, we identified C. sakazakii ompA and ompX, which encode proteins for basolateral adherence in cell lines and a possible involvement in the blood–brain barrier penetration (Mange et al., 2006; Kim et al., 2010). The Cpa protein is related to C. sakazakii serum resistance and invasion. Recent studies have suggested that the cpa locus could be considered specific for C. sakazakii and C. universalis but is absent in C. malonaticus (Franco et al., 2011). Jang et al. (2020) noted that highly virulent C. sakazakii ST8 clinical strains that carry the pESA3 plasmid do not possess the cpa gene; this indicates that the disease can be associated with other virulence factors. Hemolysins (Hly) are outer membrane proteins occurring in various pathogens belonging to the Enterobacteriaceae family that have hemolytic ability, such as E. coli, Klebsiella, and some gram-positive pathogens (Mare et al., 2020; Mazzantini et al., 2020). Other important genes of the C. sakazakii strains isolated from PIF are nanAKT that encode for exogenous sialic acid utilization. Sialic acid is naturally present in breast milk and supplemented in PIF because of its association with brain development (Forsythe, 2018). Sialic acid also regulates the expression of enzymes, such as sialidase and adhesins, or inhibits transcription factors of the fimB gene that mediates epithelial cell adherence and invasion. Escherichia coli K1 use this compound to modify their cell surface (Severi et al., 2007; Sohanpal et al., 2007).
In our study, we found the fic toxin encoding gene and the relB gene that encodes for the relE antitoxin. The fic gene encodes for the toxin component of the toxin-antitoxin bicistronic operon. The toxin-antitoxin (TA) systems are small genetic elements found in plasmids, phage genomes, and in the chromosomes of different bacterial species. The TA genes play a fundamental role in the physiology of bacterial stress response, such as in stabilizing horizontally acquired mobile genetic elements and participating in a persistence phenotype in some species, including E. coli and Salmonella (Deter et al., 2017; Walling and Butler, 2019). Finkelstein et al. (2019) noted in preliminary studies with C. sakazakii isolates that 2 typical toxin genes, fic and hipA, followed the evolutionary lines of the species and that C. sakazakii ST1 strains were the only strains containing the 22 TA homologs.
In the Salmonella strains, 110 of the 121 detected virulence genes were similar in all the strains, including the invA, ssaAR ssrAB, sipAC, sopBE, spvBC, and rck genes. Genes such as invA, sipA, sopB, and sopE are associated with adherence in epithelial cells and phagocyte invasion. Salmonella pathogenicity islands (SPI) are virulence gene clusters acquired by horizontal transfer that promote virulence in Salmonella spp. The SPI-2 gene encodes genes such as ssaR and ssrA that promote survival and replication within the macrophages and their dissemination in the host (Barilli et al., 2018). The spvC gene enables the survival and rapid growth of Salmonella in the host (Hu et al., 2021), while the spvB gene is known as an intracellular toxin that encodes an enzyme that ADP-ribosylates actin and destabilizes the cytoskeleton of eukaryotic cells (Lesnick et al., 2001).
The rck gene is defined as an invasin that generates a colonization niche or as a cyclomodulin with genotoxic activity (Mambu et al., 2020). To date, 24 SPIs have been identified in Salmonella of which SPI-1 and SPI-2 are the most important for virulence traits. SPI-1 is involved in the entire Salmonella infection process, including invasion, macrophage proliferation, and host responses. Both SPI-1 and SPI-2 encode different secretion systems, such as T3SS, that transport effector proteins to the host cells (Cheng et al., 2019; Lou et al., 2019).
Only the C. sakazakii ST1 strains carried the COL(pHHAD28) plasmids, which are associated with antibiotic resistance genes; these have previously been detected in C. sakazakii strains isolated from dairy products in Chile (Ramsamy et al., 2020; Khezri et al., 2021; Parra-Flores et al., 2021b). The C. malonaticus strain harbored a plasmid homologous to pESA3 called IncFIB (pCTU1), which encodes iron acquisition virulence genes necessary for pathogen survival, but not an external protease known as plasminogen activator (cpa) that enhances pathogen propagation in the host (Franco et al., 2011). Meanwhile, all the Salmonella strains showed IncFII(S) plasmid incompatibility, which is associated with several antibiotic resistance genes, such as aac(6′)-Iaa and mcr-9, and β-lactamase, such as ampC (Cha et al., 2020; Richter et al., 2020).
Bacterial CRISPR-Cas systems are considered as mechanisms of acquired immunity because they provide them with the ability to avoid bacteriophage infection and the acquisition of mobile genetic material from plasmids. This immunity is due to the information stored in the matrices of these systems, specifically in the spacer sequences. It was possible to determine that repeated sequences and cas genes in Cronobacter spp. were associated with two types of systems previously identified as I-E and I-F (Ogrodzki and Forsythe, 2016; Zeng et al., 2017), including cases in which the same strain showed both systems. The opposite was observed in Salmonella spp. genomes that were characterized only by the presence of type I-E (Louwen et al., 2014). Although both genera carry systems of the same type, there are differences between the associated cas gene operons because the Salmonella spp. systems show a larger number of genes. However, these systems are characterized by the presence of the cas1 and cas2 genes, which are involved in integrating and processing the information that is integrated into functional CRISPR arrays. Salmonella enterica and E. coli are closely related and harbor the IE-type CRISPR-Cas system in a similar manner (Makarova et al., 2015). The array sizes of the Cronobacter spp. genomes are smaller. However, when showing more than two arrays, they tend to have a larger number of spacer, suggesting that they have been exposed to a larger number of invasive elements than S. Typhimurium. Although it was not possible to determine that all the spacers of the systems are similar to bacteriophage sequences, there are cases in which they are specific to these genera. Of particular interest, some of the S. Typhimurium spacer were associated with CrAssphages and other phages associated with the human intestinal microbiota (Guerin et al., 2018). It is known that Salmonella is recognized as an intestinal pathogen; therefore, this is an ecosystem where bacteria can acquire external genetic material and integrate it into their genome by various horizontal transfer mechanisms and by other means such as CRISPR-Cas systems (Louwen et al., 2014). The cas1 and cas2 genes are indispensable for integrating and processing the information acquired by the bacteria; in their absence, the system loses the ability to acquire information (Parra-Flores et al., 2021b). The search for prophages in the studied genomes showed that they may have few intact phages. Cronobacter spp. tend to show a larger number of intact and incomplete prophages, which are not only characteristic of this genus but also of Salmonella. Therefore, our data suggest that these genomes carry the necessary information to prevent these bacteriophages from infecting both Cronobacter spp. and Salmonella strains. In addition, this information can be useful in the future when using gene therapy as a therapeutic option for infections caused by these pathogens that are difficult to treat (Gordillo Altamirano and Barr, 2019).
In the present study, WGS allowed us to determine multiple virulence and antibiotic resistance genes in bacterial pathogens isolated from PIF intended for consumption by infants aged less than 6 months and distributed throughout Latin America. In our opinion, the identification of C. sakazakii and S. Typhimurium in PIFs not only violates current health regulations but also endanger the health of infants consuming these products. It is therefore necessary that health authorities conduct more preventive control activities related to these products and carry out campaigns that emphasize the use of rehydration water at 70°C for infant formula. This recommendation by the World Health Organization highlights that the 70°C temperature has a proven effect in significantly decreasing the risk of disease by pathogens in reconstituted PIF (FAO/WHO, 2004, 2006). Control of Salmonella and Cronobacter during the production of milk powder and PIF is through microbiological sampling according to Codex Alimentarius Commission (2007) of finished product, as well as ingredients and intermediate products (International Standards for Organization (ISO), 2017; Podolak and Black, 2017). The use of zoning production facilities to focus environmental sampling according to risk is used (Cordier, 2008; FDA, 2019).
C. sakazakii, C. malonaticus, and S. Typhimurium strains isolated from PIF exhibit antibiotic resistance and various virulence genes and resistance to β-lactam antibiotics. Continuous monitoring of these powdered infant formulas is necessary due to the risk associated with pathogen contamination of the product and consumption by the immunologically vulnerable child population.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
JP-F, OH, SA, SL, AC-C, JM-R, AC, and SF conceived the experiments and prepared the manuscript. JP-F, AC-F, PC-S, AC-C, AP, SL, and WR conducted the laboratory work. JP-F, OH, SA, AC-C, JX-C, JM-R, AC, WR, and SF drafted the manuscript. All authors contributed to the article and approved the submitted version.
This research was funded by the Research Directorate of the Universidad del Bío-Bío, Projects 191520 4/R and GI 195420/EF.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We wish to thank the Research Directorate of the Universidad del Bío-Bío.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2022.884721/full#supplementary-material
1. ^https://pubmlst.org/organisms/cronobacter-spp/
2. ^http://enterobase.warwick.ac.uk/species/index/senterica
Achtman, M., Wain, J., Weill, F. X., Nair, S., Zhou, Z., Sangal, V., et al. (2012). Multilocus sequence typing as a replacement for serotyping in Salmonella enterica. PLoS Pathog. 8:e1002776. doi: 10.1371/journal.ppat.1002776
Aldubyan, M., Almami, I., Benslimane, F., Alsonosi, A., and Forsythe, S. (2017). Comparative outer membrane protein analysis of high and low-invasive strains of Cronobacter malonaticus. Front. Microbiol. 8:2268. doi: 10.3389/fmicb.2017.02268
Aly, M. A., Domig, K. J., Kneifel, W., and Reimhult, E. (2019). Whole genome sequencing-based comparison of food isolates of Cronobacter sakazakii. Front. Microbiol. 10:1464. doi: 10.3389/fmicb.2019.01464
Angulo, F., Cahill, S., Wachsmuth, K., Costarrica, M. L., and Ben Embarek, P. (2008). Powdered infant formula as a source of Salmonella infection in infants. Clin. Infect. Dis. 46, 268–273. doi: 10.1086/524737
Baldwin, A., Loughlin, M., Caubilla-Barron, J., Kucerova, E., Manning, G., Dowson, C., et al. (2009). Multilocus sequence typing of Cronobacter sakazakii and Cronobacter malonaticus reveals stable clonal structures with clinical significance which do not correlate with biotypes. BMC Microbiol. 9:223. doi: 10.1186/1471-2180-9-223
Barilli, E., Bacci, C., StellaVilla, Z., Merialdi, G., D'Incau, M., Brindani, F., et al. (2018). Antimicrobial resistance, biofilm synthesis and virulence genes in Salmonella isolated from pigs bred on intensive farms. Ital. J. Food Saf. 7:7223. doi: 10.4081/ijfs.2018.7223
Biswas, A., Staals, R., Morales, S., Fineran, P., and Brown, C. (2016). CRISPRDetect: a flexible algorithm to define CRISPR arrays. BMC Genomics 17:356. doi: 10.1186/s12864-016-2627-0
Borowiak, M., Baumann, B., Fischer, J., Thomas, K., Deneke, C., Hammerl, J. A., et al. (2020). Development of a novel mcr-6 to mcr-9 multiplex PCR and assessment of mcr-1 to mcr-9 occurrence in colistin-resistant Salmonella enterica isolates from environment, feed, animals and food (2011–2018) in Germany. Front. Microbiol. 11:80. doi: 10.3389/fmicb.2020.00080
Brouard, C., Espié, E., Weill, F. X., Kérouanton, A., Brisabois, A., Forgue, A. M., et al. (2007). Two consecutive large outbreaks of Salmonella enterica serotype Agona infections in infants linked to the consumption of powdered infant formula. Pediatr. Infect. Dis. J. 26, 148–152. doi: 10.1097/01.inf.0000253219.06258.23
Carattoli, A., Zankari, E., García-Fernández, A., Voldby Larsen, M., Lund, O., Villa, L., et al. (2014). In silico detection and typing of plasmids using plasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 58, 3895–3903. doi: 10.1128/AAC.02412-14
Carrasco, E., Morales-Rueda, A., and García-Gimeno, R. M. (2012). Cross-contamination and recontamination by salmonella in foods: a review. Food Res. Int. 45, 545–556. doi: 10.1016/j.foodres.2011.11.004
Carroll, L., Gaballa, A., Guldimann, C., Sullivan, G., Henderson, L., and Wiedmann, M. (2019). Identification of novel mobilized colistin resistance gene mcr-9 in a multidrug-resistant, colistin-susceptible Salmonella enterica serotype Typhimurium isolate. MBio 10, e00853–e00919. doi: 10.1128/mBio.00853-19
Carroll, L. M., Wiedmann, M., den Bakker, H., Siler, J., Warchocki, S., Kent, D., et al. (2017). Whole-genome sequencing of drug-resistant Salmonella enterica isolates from dairy cattle and humans in New York and Washington States reveals source and geographic associations. Appl. Environ. Microbiol. 83, e00140–e00217. doi: 10.1128/AEM.00140-17
Caubilla-Barron, J., and Forsythe, S. (2007). Dry stress and survival time of Enterobacter sakazakii and other Enterobacteriaceae in dehydrated powdered infant formula. J. Food Prot. 70, 2111–2117. doi: 10.4315/0362-028x-70.9.2111
Cha, M. H., Woo, G. J., Lee, W., Kim, S. H., Woo, J. H., Kim, J., et al. (2020). Emergence of transferable mcr-9 gene-carrying colistin-resistant Salmonella enterica Dessau ST14 isolated from retail chicken meat in Korea. Foodborne Pathog. Dis. 17, 720–727. doi: 10.1089/fpd.2020.2810
Cheng, R. A., Eade, C. R., and Wiedmann, M. (2019). Embracing diversity: differences in virulence mechanisms, disease severity, and host adaptations contribute to the success of Nontyphoidal Salmonella as a foodborne pathogen. Front. Microbiol. 10:1368. doi: 10.3389/fmicb.2019.01368
Chon, J., Song, K., Kim, S., Hyeon, J., and Seo, K. (2012). Isolation and characterization of Cronobacter from desiccated foods in Korea. J. Food Sci. 77, 354–358. doi: 10.1111/j.1750-3841.2012.02750.x
CLSI. (2020). Performance Standards for Antimicrobial Susceptibility Testing. 30th Edn. Wayne, PA: CLSI.
Codex Alimentarius Commission (2007). Standard for Infant Formula and Formulas for Special Medical Purposes Intended for Infants. CXS 72-1981 Revised 2007, 119–11.
Cordier, J.-L. (2008). “Chapter 6: Production of powdered infant formulae and microbiological control measures,” in Enterobacter sakazakii. eds. J. Farber and S. J. Forsythe (Washington, D.C.: ASM Press).
Costa, P. V., Vasconcellos, L., Forsythe, S. J., and Brandão, M. L. L. (2021). Diversity of Cronobacter genus isolated between 1970 and 2019 on the American continent and genotyped using multi-locus sequence typing. FEMS Microbiol. Lett. 368, 1–9. doi: 10.1093/femsle/fnab027
Craven, H. H., McAuley, C. M., Duffy, L. L., and Fegan, N. (2010). Distribution, prevalence and persistence of Cronobacter (Enterobacter sakazakii) in the non-processing and processing environments of five milk powder factories. J. Appl. Microbiol. 109, 1044–1052. doi: 10.1111/j.1365-2672.2010.04733.x
Cruz, A., Xicohtencatl, J., Gonzalez, B., Bobadilla, M., Eslava, C., and Rosas, I. (2011). Virulence traits in Cronobacter species isolated from different sources. Can. J. Microbiol. 57, 735–744. doi: 10.1139/w11-063
Csorba, C., Pajić, M., Blagojević, B., Forsythe, S., Radinović, M., and Velebit, B. (2021). Prevalence, characterization, and antibiotic susceptibility of Cronobacter spp. in a milk powder processing environment: the first reported case in Serbia. Food Sci. Nutr. 10, 554–563. doi: 10.1002/fsn3.2681
de Frutos, M., López-Urrutia, L., Berbel, C., Allue, M., Herrera, S., Azcona, J. M., et al. (2018). Brote de Salmonella Typhimurium monofásica asociada al consumo de carne asada de cerdo [Monophasic Salmonella Typhimurium outbreak due to the consumption of roast pork meat]. Rev. Esp. Quimioter. 31, 156–159.
de Toro, M., Sáenz, Y., Cercenado, E., Rojo-Bezares, B., García-Campello, M., Undabeitia, E., et al. (2011). Genetic characterization of the mechanisms of resistance to amoxicillin/clavulanate and third-generation cephalosporins in Salmonella enterica from three Spanish hospitals. Int. Microbiol. 14, 173–181. doi: 10.2436/20.1501.01.146
de Toro, M., Seral, C., Rojo-Bezares, B., Torres, C., Castillo, F. J., and Sáenz, Y. (2014). Resistencia a antibióticos y factores de virulencia en aislados clínicos de Salmonella enterica. Enferm. Infecc. Microbiol. Clin. 32, 4–10. doi: 10.1016/j.eimc.2013.03.006
Deter, H. S., Jensen, R. V., Mather, W. H., and Butzin, N. C. (2017). Mechanisms for differential protein production in toxin–antitoxin systems. Toxins 9:211. doi: 10.3390/toxins9070211
Falagas, M., Athanasaki, F., Voulgaris, G., Triarides, N., and Vardakas, K. (2019). Resistance to fosfomycin: mechanisms, frequency and clinical consequences. Int. J. Antimicrob. Agents 53, 22–28. doi: 10.1016/j.ijantimicag.2018.09.013
FAO/WHO (2004). Enterobacter sakazakii and Other Microorganisms in Powdered Infant Formula: Meeting Report. Microbiological Risk Assessment. Series No. 6 Geneva: WHO.
FAO/WHO (2006). Enterobacter Sakazakii and Salmonella Powdered Infant Formula. Microbiological Risk Assessment. Series No. 10. Rome: FAO
FAO/WHO (2008). Enterobacter sakazakii and other microorganisms in powdered follow-up formula. Meeting report. Microbiological Risk Assessment. Series No. 15. Rome: FAO.
FDA (2019). Investigations Operation Manual. Available at: https://www.fda.gov/inspections-compliance-enforcement-and-criminal-investigations/inspection-references/investigations-operations-manual (Accessed March, 2021).
Fei, P., Jiang, Y., Feng, J., Forsythe, S. J., Li, R., Zhou, Y., et al. (2017). Antibiotic and desiccation resistance of Cronobacter sakazakii and C. malonaticus isolates from powdered infant formula and processing environments. Front. Microbiol. 8:316. doi: 10.3389/fmicb.2017.00316
Fei, P., Man, C., Lou, B., Forsythe, S., Chai, Y., Li, R., et al. (2015). Genotyping and source tracking of the Cronobacter sakazakii and C. malonaticus isolated from powdered infant formula and an infant formula production factory in China. Appl. Environ. Microbiol. 81, 5430–5439. doi: 10.1128/AEM.01390-15
Ferri, M., Ranucci, E., Romagnoli, P., and Giaccone, V. (2017). Antimicrobial resistance: a global emerging threat to public health systems. Crit. Rev. Food Sci. Nutr. 57, 2857–2876. doi: 10.1080/10408398.2015.1077192
Finkelstein, S., Negrete, F., Jang, H., Gangiredla, J., Mammel, M., Patel, I. R., et al. (2019). Prevalence, distribution, and phylogeny of type two toxin-antitoxin genes possessed by Cronobacter species where C. sakazakii homologs follow sequence type lineages. Microorganisms 7:554. doi: 10.3390/microorganisms7110554
Flores, J. P., Arvizu, S., Silva, J., and Fernández-Escartín, E. (2011). Two cases of hemorrhagic diarrhea caused by Cronobacter sakazakii in hospitalized nursing infants associated with the consumption of powdered infant formula. J. Food Prot. 74, 2177–2181. doi: 10.4315/0362-028X.JFP-11-257
Forsythe, S. J. (2018). Updates on the Cronobacter genus. Annu. Rev. Food Sci. Technol. 9, 23–44. doi: 10.1146/annurev-food-030117-012246
Franco, A. A., Kothary, M., Gopinath, G., Jarvis, K., Grim, C. J., Hu, L., et al. (2011). Cpa, the outer membrane protease of Cronobacter sakazakii, activates plasminogen and mediates resistance to serum bactericidal activity. Infect. Immun. 79, 1578–1587. doi: 10.1128/IAI.01165-10
Garallah, E. T., and Al-Jubori, S. (2020). Molecular detection of glpT and uhpT genes as fosfomycin pathways in UTI infection patients. Gene Rep. 21:100930. doi: 10.1016/j.genrep.2020.100930
Gordillo Altamirano, F. L., and Barr, J. J. (2019). Phage therapy in the postantibiotic era. Clin. Microbiol. Rev. 32, e00066–e00068. doi: 10.1128/CMR.00066-18
Guerin, E., Shkoporov, A., Stockdale, S. R., Clooney, A. G., Ryan, F. J., Sutton, T., et al. (2018). Biology and taxonomy of crAss-like bacteriophages, the Most abundant virus in the human gut. Cell Host Microbe 24, 653.e6–664.e6. doi: 10.1016/j.chom.2018
Güerri, M. L., Aladueña, A., Echeíta, A., and Rotger, R. (2004). Detection of integrons and antibiotic-resistance genes in Salmonella enterica serovar Typhimurium isolates with resistance to ampicillin and variable susceptibility to amoxicillin-clavulanate. Int. J. Antimicrob. Agents 24, 327–333. doi: 10.1016/j.ijantimicag.2004.04.009
Holý, O., Cruz-Cordova, A., Xicohtencatl-Cortés, J., Hochel, I., Parra-Flores, J., Petrzelova, J., et al. (2019). Occurrence of virulence factors in Cronobacter sakazakii and Cronobacter malonaticus originated from clinical samples. Microb. Pathog. 127, 250–256. doi: 10.1016/j.micpath.2018.12.011
Holý, O., Parra-Flores, J., Lepuschitz, S., Alarcón-Lavín, M. P., Cruz-Córdova, A., Xicohtencatl-Cortes, J., et al. (2021). Molecular characterization of Cronobacter sakazakii strains isolated from powdered milk. Foods 10:20. doi: 10.3390/foods10010020
Hu, L., Cao, G., Brown, E. W., Allard, M., Ma, L., and Zhang, G. (2021). Whole genome sequencing and protein structure analyses of target genes for the detection of Salmonella. Sci. Rep. 11:20887. doi: 10.1038/s41598-021-00224-7
Huehn, S., La Ragione, R. M., Anjum, M., Saunders, M., Woodward, M. J., Bunge, C., et al. (2010). Virulotyping and antimicrobial resistance typing of Salmonella enterica serovars relevant to human health in Europe. Foodborne Pathog. Dis. 7, 523–535. doi: 10.1089/fpd.2009.0447
Ingram, P., Inglis, T., Vanzetti, T., Henderson, B., Harnett, G., and Murray, R. (2011). Comparison of methods for AmpC β-lactamase detection in Enterobacteriaceae. J. Med. Microbiol. 60, 715–721. doi: 10.1099/jmm.0.029140-0.
Instituto Nacional de Normalización (2002). Chile. Norma Chilena NCh 2675 de detección de Salmonella. Available at: https://ecommerce.inn.cl/nch2675200244470 (Accessed October 10, 2020).
International Standards for Organization (ISO) (2017). ISO 22964:2017: microbiology of the food chain—horizontal method for the detection of Cronobacter spp. international standards Organization. Available at: https://www.iso.org/standard/64708.htm
Iversen, C., and Forsythe, S. (2004). Isolation of Enterobacter sakazakii and other Enterobacteriaceae from powdered infant formula milk and related products. Food Microbiol. 21, 771–777. doi: 10.1016/j.fm.2004.01.009
Iversen, C., Mullane, N., Mc Cardell, B., Tall, B. D., Lehner, A., Fanning, S., et al. (2008). Cronobacter gen. nov., a new genus to accommodate the biogroups of Enterobacter sakazakii, and proposal of Cronobacter sakazakii gen. Nov. comb. nov., C. malonaticus sp. nov., C. turicensis sp. nov., C. muytjensii sp. nov., C. dublinensis sp. nov., Cronobacter genomospecies 1, and of three subspecies, C. dublinensis sp. nov. subsp. dublinensis subsp. nov., C. dublinensis sp. nov. subsp. lausannensis subsp. nov., and C. dublinensis sp. nov. subsp. lactaridi subsp. nov. Int. J. Syst. Evol. Microbiol. 58, 1442–1447. doi: 10.1099/ijs.0.65577-0
Jain, P., Sudhanthirakodi, S., Chowdhury, G., Joshi, S., Anandan, S., Ray, U., et al. (2018). Antimicrobial resistance, plasmid, virulence, multilocus sequence typing and pulsed-field gel electrophoresis profiles of Salmonella enterica serovar Typhimurium clinical and environmental isolates from India. PLoS One 13:e0207954. doi: 10.1371/journal.pone.0207954
Jang, H., Chase, H. R., Gangiredla, J., Grim, C. J., Patel, I. R., Kothary, M. H., et al. (2020). Analysis of the molecular diversity among Cronobacter species isolated from filth flies using targeted PCR, pan genomic DNA microarray, and whole genome sequencing analyses. Front. Microbiol. 11:561204. doi: 10.3389/fmicb.2020.561204
Jia, B., Raphenya, A. R., Alcock, B., Waglechner, N., Guo, P., Tsang, K., et al. (2017). CARD 2017: expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 45, D566–D573. doi: 10.1093/nar/gkw1004
Johansson, M., Bortolaia, V., Tansirichaiya, S., Aarestrup, F. M., Roberts, A. P., and Petersen, T. N. (2021). Detection of mobile genetic elements associated with antibiotic resistance in Salmonella enterica using a newly developed web tool: MobileElementFinder. J. Antimicrob. Chemother. 76, 101–109. doi: 10.1093/jac/dkaa390
Jones, G., Pardos de la Gandara, M., Herrera-Leon, L., Herrera-Leon, S., Varela Martinez, C., Hureaux-Roy, R., et al. (2019). Outbreak of Salmonella enterica serotype Poona in infants linked to persistent Salmonella contamination in an infant formula manufacturing facility, France, august 2018 to February 2019. Euro Surveill. 24:1900161. doi: 10.2807/1560-7917.ES.2019.24.13.1900161
Joseph, S., Cetinkaya, E., Drahovska, H., Levican, A., Figueras, M., and Forsythe, S. (2012). Cronobacter condimenti sp. nov., isolated from spiced meat, and Cronobacter universalis sp. nov., a species designation for Cronobacter sp. Geneomoespecies 1, recovered from a leg infection, water and food ingredients. Int. J. Syst. Evol. Microbiol. 62, 1277–1283. doi: 10.1099/ijs.0.032292-0
Joseph, S., and Forsythe, S. J. (2011). Predominance of Cronobacter sakazakii sequence type 4 in neonatal infections. Emerg. Infect. Dis. 17, 1713–1715. doi: 10.3201/eid1709.110260
Joseph, S., and Forsythe, S. (2012). Insights into the emergent bacterial pathogen Cronobacter spp., generated by multilocus sequence typing and analysis. Front. Microbiol. 3:397. doi: 10.3389/fmicb.2012.00397
Jourdan-da Silva, N., Fabre, L., Robinson, E., Fournet, N., Nisavanh, A., Bruyand, M., et al. (2018). Ongoing nationwide outbreak of Salmonella Agona associated with internationally distributed infant milk products, France, December 2017. Euro Surveill. 23, 17–00852. doi: 10.2807/1560-7917.ES.2018.23.2.17-00852
Jünemann, S., Sedlazeck, F. J., Prior, K., Albersmeier, A., John, U., Kalinowski, J., et al. (2013). Updating benchtop sequencing performance comparison. Nat. Biotechnol. 31, 294–296. doi: 10.1038/nbt.2522
Khezri, A., Avershina, E., and Ahmad, R. (2021). Plasmid identification and plasmid-mediated antimicrobial gene detection in norwegian isolates. Microorganisms 9:52. doi: 10.3390/microorganisms9010052
Kieffer, N., Royer, G., Decousser, J. W., Bourrel, A. S., Palmieri, M., Ortiz De La Rosa, J. M., et al. (2019). Mcr-9, an inducible gene encoding an acquired phosphoethano-lamine transferase in Escherichia coli, and its origin. Antimicrob. Agents Chemother. 63, e00965–e001019. doi: 10.1128/AAC.00965-19
Kim, K., Jang, S., Kim, S., Park, J., Heu, S., and Ryu, S. (2008). Prevalence and genetic diversity of Enterobacter sakazakii in ingredients of infant foods. Int. J. Food Microbiol. 122, 196–203. doi: 10.1016/j.ijfoodmicro.2007.11.072
Kim, K., Kim, K., Choi, J., Lim-Jeong, A., Lee, J., Hwang, S., et al. (2010). Outer membrane proteins a (OmpA) and x (OmpX) are essential for basolateral invasion of Cronobacter sakazakii. Appl. Environ. Microbiol. 76, 5188–5198. doi: 10.1128/AEM.02498-09
Lachowska, M., Izdebski, R., Urbanowicz, P., Żabicka, D., and Królak-Olejnik, B. (2021). Infection of Cronobacter sakazakii ST1 producing SHV-12 in a premature infant born from triplet pregnancy. Microorganisms 9:1878. doi: 10.3390/microorganisms9091878
Lange, S., Alkhnbashi, O., Rose, D., Will, S., and Backofen, R. (2013). CRISPRmap: an automated classification of repeat conservation in prokaryotic adaptive immune systems. Nucleic Acids Res. 41, 8034–8044. doi: 10.1093/nar/gkt606
Lee, Y.-D., Park, J., and Chang, H. (2012). Detection, antibiotic susceptibility and biofilm formation of Cronobacter spp. from various foods in Korea. Food Control 24, 225–230. doi: 10.1016/j.foodcont.2011.09.023
Lehner, A., Tall, B. D., Fanning, S., and Srikumar, S. (2018). Cronobacter spp.—opportunistic foodborne pathogens: an update on evolution, osmotic adaptation and pathogenesis. Curr. Clin. Microbiol. Rep. 5, 97–105. doi: 10.1007/s40588-018-0089-7
Leopold, S., Goering, R., Witten, A., Harmsen, D., and Mellmann, A. (2014). Bacterial whole-genome sequencing revisited: portable, scalable, and standardized analysis for typing and detection of virulence and antibiotic resistance genes. J. Clin. Microbiol. 52, 2365–2370. doi: 10.1128/JCM.00262-14
Lepuschitz, S., Ruppitsch, W., Pekard-Amenitsch, S., Forsythe, S. J., Cormican, M., Mach, R. L., et al. (2019). Multicenter study of Cronobacter sakazakii infections in humans, Europe, 2017. Emerg. Infect. Dis. 25, 515–522. doi: 10.3201/eid2503.181652
Lepuschitz, S., Sorschag, S., Springer, B., Allerberger, F., and Ruppitsch, W. (2017). Draft genome sequence of carbapenemase-producing Serratia marcescens isolated from a patient with chronic obstructive pulmonary disease. Genome Announc. 5, e01288–e01317. doi: 10.1128/genomeA.01288-1
Lepuschitz, S., Weinmaier, T., Mrazek, K., Beisken, S., Weinberger, J., and Posch, A. E. (2020). Analytical performance validation of next-generation sequencing based clinical microbiology assays using a K-mer analysis workflow. Front. Microbiol. 11:1883. doi: 10.3389/fmicb.2020.01883
Lesnick, M. L., Reiner, N. E., Fierer, J., and Guiney, D. G. (2001). The Salmonella spvB virulence gene encodes an enzyme that ADP-ribosylates actin and destabilizes the cytoskeleton of eukaryotic cells. Mol. Microbiol. 39, 1464–1470. doi: 10.1046/j.1365-2958.2001.02360.x
Lou, L., Zhang, P., Piao, R., and Wang, Y. (2019). Salmonella Pathogenicity Island 1 (SPI-1) and its complex regulatory network. Front. Cell. Infect. Microbiol. 9:270. doi: 10.3389/fcimb.2019.00270
Louwen, R., Staals, R. H., Endtz, H. P., van Baarlen, P., and van der Oost, J. (2014). The role of CRISPR-Cas systems in virulence of pathogenic bacteria. Microbiol. Mol. Biol. Rev. 78, 74–88. doi: 10.1128/MMBR.00039-13
Makarova, K. S., Wolf, Y. I., Alkhnbashi, O. S., Costa, F., Shah, S. A., Saunders, S. J., et al. (2015). An updated evolutionary classification of CRISPR-Cas systems. Nat. Rev. Microbiol. 13, 722–736. doi: 10.1038/nrmicro3569
Mambu, J., Barilleau, E., Fragnet-Trapp, L., Le Vern, Y., Olivier, M., Sadrin, G., et al. (2020). Rck of Salmonella Typhimurium delays the host cell cycle to facilitate bacterial invasion. Front. Cell. Infect. Microbiol. 10:586934. doi: 10.3389/fcimb.2020.586934
Mange, J. P., Stephan, R., Borel, L., Wild, P., Kim, K. S., Pospischil, A., et al. (2006). Adhesive properties of Enterobacter sakazakii to human epithelial and brain microvascular endothelial cells. BMC Microbiol. 6, 58–68. doi: 10.1186/1471-2180-6-58
Mare, A., Man, A., Toma, F., Ciurea, C. N., Coşeriu, R. L., Vintilă, C., et al. (2020). Hemolysin-producing strains among diarrheagenic Escherichia coli isolated from children under 2 years old with diarrheal disease. Pathogens 9:1022. doi: 10.3390/pathogens9121022
Marraffini, L. A. (2013). CRISPR-Cas immunity against phages: its effects on the evolution and survival of bacterial pathogens. PLoS Pathog. 9:e1003765. doi: 10.1371/journal.ppat.1003765
Mazzantini, D., Fonnesu, R., Celandroni, F., Calvigioni, M., Vecchione, A., Mrusek, D., et al. (2020). GTP-dependent FlhF homodimer supports secretion of a hemolysin in Bacillus cereus. Front. Microbiol. 11:879. doi: 10.3389/fmicb.2020.00879
Mezal, E. H., Sabol, A., Khan, M. A., Ali, N., Stefanova, R., and Khan, A. A. (2014). Isolation and molecular characterization of salmonella enterica serovar Enteritidis from poultry house and clinical samples during 2010. Food Microbiol. 38, 67–74. doi: 10.1016/j.fm.2013.08.003
Molloy, C., Cagney, C., O’Brien, S., Iversen, C., Fanning, S., and Duffy, G. (2009). Surveillance and characterization by pulsed-field gel electrophoresis of Cronobacter spp in farming and domestic environments, food production animals and retails foods. Int. J. Food Microbiol. 136, 198–203. doi: 10.1016/j.ijfoodmicro.2009.07.007
Monte, D. F. M., Sellera, F. P., Lopes, R., Keelara, S., Landgraf, M., Greene, S., et al. (2020). Class 1 integron-borne cassettes harboring blaCARB-2 gene in multidrug-resistant and virulent Salmonella Typhimurium ST19 strains recovered from clinical human stool samples, United States. PLoS One 15:e0240978. doi: 10.1371/journal.pone.0240978
Mossong, J., Marques, P., Ragimbeau, C., Huberty-Krau, P., Losch, S., Meyer, G., et al. (2007). Outbreaks of monophasic Salmonella enterica serovar 4, [5],12: i: - in Luxembourg, 2006. Euro Surveill. 12, E11–E12. doi: 10.2807/esm.12.06.00719-en
Müller, A., Hächler, H., Stephan, R., and Lehner, A. (2014). Presence of AmpC beta-lactamases, CSA-1, CSA-2, CMA-1, and CMA-2 conferring an unusual resistance phenotype in Cronobacter sakazakii and Cronobacter malonaticus. Microb. Drug Resist. 20, 275–280. doi: 10.1089/mdr.2013.0188
Murugkar, H. V., Rahman, H., and Dutta, P. K. (2003). Distribution of virulence genes in Salmonella serovars isolated from man & animals. Indian J. Med. Res. 117, 66–70.
Nayak, R., Stewart, T., Wang, R. F., Lin, J., Cerniglia, C. E., and Kenney, P. B. (2004). Genetic diversity and virulence gene determinants of antibiotic-resistant Salmonella isolated from preharvest Turkey production sources. Int. J. Food Microbiol. 91, 51–62. doi: 10.1016/S0168-1605(03)00330-1
Ogrodzki, P., and Forsythe, S. (2015). Capsular profiling of the Cronobacter genus and the association of specific Cronobacter sakazakii and C. malonaticus capsule types with neonatal meningitis and necrotizing enterocolitis. BMC Genomics 16:758. doi: 10.1186/s12864-015-1960-z
Ogrodzki, P., and Forsythe, S. (2016). CRISPR–cas loci profiling of Cronobacter sakazakii pathovars. Future Microbiol. 11, 1507–1519. doi: 10.2217/fmb-2016-0070
Panzenhagen, P. H. N., Paul, N. C., Conte, C. A., Costa, R. G., Rodrigues, D. P., and Shah, D. H. (2018). Genetically distinct lineages of Salmonella Typhimurium ST313 and ST19 are present in Brazil. Int. J. Med. Microbiol. 308, 306–316. doi: 10.1016/j.ijmm.2018.01.005
Parra-Flores, J., Cruz-Córdova, A., Acuña, S., Riffo-Sepúlveda, F., Maury-Sintjago, E., Rodriguez-Fernández, A., et al. (2021a). “Cronobacter spp. in milk,” in Reference Module in Food Science. eds. P. L. H. McSweeney and J. P. McNamara (Amsterdam, Netherlands: Elsevier), 1–9.
Parra-Flores, J., Aguirre, J., Juneja, V., Jackson, E., Cruz-Córdova, A., Silva-Sanchez, J., et al. (2018a). Virulence and antibiotic resistance profiles of Cronobacter sakazakii and Enterobacter spp. involved in the diarrheic hemorrhagic outbreak in Mexico. Front. Microbiol. 9:2206. doi: 10.3389/fmicb.2018.02206
Parra-Flores, J., Cerda-Leal, F., Contreras, A., Valenzuela-Riffo, N., Rodriguez, A., and Aguirre, J. (2018b). Cronobacter sakazakii and microbiological parameters in dairy formulas associated with a food alert in Chile. Front. Microbiol. 9:1708. doi: 10.3389/fmicb.2018.01708.
Parra-Flores, J., Holý, O., Riffo, F., Lepuschitz, S., Maury-Sintjago, E., Rodríguez-Fernández, A., et al. (2021b). Profiling the virulence and antibiotic resistance genes of Cronobacter sakazakii strains isolated from powdered and dairy formulas by whole-genome sequencing. Front. Microbiol. 12:694922. doi: 10.3389/fmicb.2021.694922
Parra-Flores, J., Maury-Sintjago, E., Rodríguez-Fernández, A., Acuña, S., Cerda, F., Aguirre, J., et al. (2020). Microbiological quality of powdered infant formula in Latin America. J. Food Prot. 83, 534–541. doi: 10.4315/0362-028X.JFP-19-399
Podolak, R., and Black, D. G. (2017). “Introduction and overview,” in In Control of Salmonella and Other Bacterial Pathogens in Low-Moisture Foods. 1st Edn. eds. R. Podolak and D. G. Black (West Sussex, UK: John Wiley and Sons), 1–27.
Ramsamy, Y., Mlisana, K. P., Allam, M., Amoako, D. G., Abia, A. L. K., Ismail, A., et al. (2020). Genomic analysis of carbapenemase-producing extensively drug-resistant Klebsiella pneumoniae isolates reveals the horizontal spread of p18-43_01 plasmid encoding blaNDM-1 in South Africa. Microorganisms 8:137. doi: 10.3390/microorganisms8010137
Richter, L., du Plessis, E. M., Duvenage, S., and Korsten, L. (2020). Occurrence, phenotypic and molecular characterization of extended-Spectrum- and AmpC- β-lactamase producing Enterobacteriaceae isolated from selected commercial spinach supply chains in South Africa. Front. Microbiol. 11:638. doi: 10.3389/fmicb.2020.00638
Severi, E., Hood, D. W., and Thomas, G. H. (2007). Sialic acid utilization by bacterial pathogens. Microbiology 153, 2817–2822. doi: 10.1099/mic.0.2007/009480-0
Shi, L., Liang, Q., Zhan, Z., Feng, J., Zhao, Y., Chen, Y., et al. (2018). Co-occurrence of 3 different resistance plasmids in a multi-drug resistant Cronobacter sakazakii isolate causing neonatal infections. Virulence 9, 110–120. doi: 10.1080/21505594.2017.1356537
Silver, L. L. (2017). Fosfomycin: mechanism and resistance. Cold Spring Harb. Perspect. Med. 7:a025262. doi: 10.1101/cshperspect.a025262
Sohanpal, B. K., Friar, S., Roobol, J., Plumbridge, J. A., and Blomfield, I. C. (2007). Multiple co-regulatory elements and IHF are necessary for the control of fimB expression in response to sialic acid and N-acetylglucosamine in Escherichia coli K-12. Mol. Microbiol. 63, 1223–1236. doi: 10.1111/j.1365-2958.2006.05583.x
Sonbol, H., Joseph, S., McAuley, C., Craven, H., and Forsythe, S. (2013). Multilocus sequence typing of Cronobacter spp. from powdered infant formula and milk powder production factories. Int. Dairy J. 30, 1–7. doi: 10.1016/j.idairyj.2012.11.004
Stephan, R., Grim, C., Gopinath, G., Mammel, M., Sathyamoorthy, V., Trach, L., et al. (2014). Re-examination of the taxonomic status of Enterobacter helveticus, Enterobacter pulveris and Enterobacter turicensis as members of the genus Cronobacter and their reclassification in the genera Franconibacter gen. nov. and Siccibacter gen. nov. as Franconibacter helveticus comb. nov., Franconibacter pulveris comb. nov. and Siccibacter turicensis comb. nov., respectively. Int. J. Syst. Evol. Microbiol. 64, 3402–3410. doi: 10.1099/ijs.0.059832-0
Thung, T. Y., Radu, S., Mahyudin, N. A., Rukayadi, Y., Zakaria, Z., Mazlan, N., et al. (2018). Prevalence, virulence genes and antimicrobial resistance profiles of Salmonella Serovars from retail beef in Selangor, Malaysia. Front. Microbiol. 8:2697. doi: 10.3389/fmicb.2017.02697
U.S FDA (2022). FDA Investigation of Cronobacter and Salmonella Complaints: Powdered Infant Formula (February 2022). Available at: https://www.fda.gov/food/outbreaks-foodborne-illness/fda-investigation-cronobacter-and-salmonella-complaints-powdered-infant-formula-february-2022 (Accessed February 21, 2022).
Uelze, L., Borowiak, M., Deneke, C., Szabó, I., Fischer, J., Tausch, S. H., et al. (2020). Performance and accuracy of four open-source tools for in silico serotyping of salmonella spp. based on whole-genome short-read sequencing data. Appl. Environ. Microbiol. 86, e02265–e02319. doi: 10.1128/AEM.02265-19
Walling, L. R., and Butler, J. S. (2019). Toxins targeting transfer RNAs: translation inhibition by bacterial toxin–antitoxin systems. Wiley Interdiscip. Rev. RNA 10:e1506. doi: 10.1002/wrna.1506
Wang, X., Biswas, S., Paudyal, N., Pan, H., Li, X., Fang, W., et al. (2019). Antibiotic resistance in Salmonella Typhimurium isolates recovered from the food chain through National Antimicrobial Resistance Monitoring System Between 1996 and 2016. Front. Microbiol. 10:985. doi: 10.3389/fmicb.2019.00985
Wang, X., Wang, Y., Ling, N., Shen, Y., Zhang, D., Liu, D., et al. (2021). Effects of tolC on tolerance to bile salts and biofilm formation in Cronobacter malonaticus. J. Dairy Sci. 104, 9521–9531. doi: 10.3168/jds.2021-20128
Wang, L., Zhu, W., Lu, G., Wu, P., Wei, Y., Su, Y., et al. (2021). In silico species identification and serotyping for Cronobacter isolates by use of whole-genome sequencing data. Int. J. Food Microbiol. 358:109405. doi: 10.1016/j.ijfoodmicro.2021.109405
Wei, Z., Xu, X., Yan, M., Chang, H., Li, Y., Kan, B., et al. (2019). Salmonella Typhimurium and salmonella Enteritidis infections in sporadic diarrhea in children: source tracing and resistance to third-generation Cephalosporins and ciprofloxacin. Foodborne Pathog. Dis. 16, 244–255. doi: 10.1089/fpd.2018.2557
Xiang, Y., Li, F., Dong, N., Tian, S., Zhang, H., Du, X., et al. (2020). Investigation of a salmonellosis outbreak caused by multidrug resistant salmonella Typhimurium in China. Front. Microbiol. 11:801. doi: 10.3389/fmicb.2020.00801
Zeng, H., Lei, T., He, W., Zhang, J., Liang, B., Li, C., et al. (2018). Novel multidrug resistant Cronobacter sakazakii causing meningitis in neonate, China, 2015. Emerg. Infect. Dis. 24, 2121–2124. doi: 10.3201/eid2411.180718
Zeng, H., Zhang, J., Li, C., Xie, T., Ling, N., Wu, Q., et al. (2017). The driving force of prophages and CRISPR-Cas system in the evolution of Cronobacter sakazakii. Sci. Rep. 7:40206. doi: 10.1038/srep46783
Zhang, S., Yin, Y., Jones, M. B., Zhang, Z., Deatherage Kaiser, B. L., Dinsmore, B. A., et al. (2015). Salmonella serotype determination utilizing high-throughput genome sequencing data. J. Clin. Microbiol. 53, 1685–1692. doi: 10.1128/JCM.00323-15
Keywords: Cronobacter sakazakii, Cronobacter malonaticus, Salmonella Typhimurium, powdered infant formula, virulence, resistance genes, whole-genome sequencing, CRISPR-Cas
Citation: Parra-Flores J, Holý O, Acuña S, Lepuschitz S, Pietzka A, Contreras-Fernández A, Chavarría-Sepulveda P, Cruz-Córdova A, Xicohtencatl-Cortes J, Mancilla-Rojano J, Castillo A, Ruppitsch W and Forsythe S (2022) Genomic Characterization of Cronobacter spp. and Salmonella spp. Strains Isolated From Powdered Infant Formula in Chile. Front. Microbiol. 13:884721. doi: 10.3389/fmicb.2022.884721
Edited by:
Jerome Combrisson, Mars, United StatesReviewed by:
Luxin Wang, University of California, Davis, United StatesCopyright © 2022 Parra-Flores, Holý, Acuña, Lepuschitz, Pietzka, Contreras-Fernández, Chavarría-Sepulveda, Cruz-Córdova, Xicohtencatl-Cortes, Mancilla-Rojano, Castillo, Ruppitsch and Forsythe. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Julio Parra-Flores, anVwYXJyYUB1YmlvYmlvLmNs; Stephen Forsythe, c2ZvcnN5dGhlNGpAZ21haWwuY29t
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.