
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 29 April 2022
Sec. Microbial Physiology and Metabolism
Volume 13 - 2022 | https://doi.org/10.3389/fmicb.2022.871077
 Ana Laura Ramos1
Ana Laura Ramos1 Maria Aquino2
Maria Aquino2 Gema García3Miriam Gaspar2
Gema García3Miriam Gaspar2 Cristina de la Cruz4Anaid Saavedra-Flores5
Cristina de la Cruz4Anaid Saavedra-Flores5 Susana Brom4
Susana Brom4 Ramón Cervantes-Rivera6,7,8
Ramón Cervantes-Rivera6,7,8 Clara Elizabeth Galindo-Sánchez5
Clara Elizabeth Galindo-Sánchez5 Rufina Hernandez9
Rufina Hernandez9 Andrea Puhar6,7,8
Andrea Puhar6,7,8 Andrei N. Lupas10
Andrei N. Lupas10 Edgardo Sepulveda11*
Edgardo Sepulveda11*The SLC5/STAC histidine kinases comprise a recently identified family of sensor proteins in two-component signal transduction systems (TCSTS), in which the signaling domain is fused to an SLC5 solute symporter domain through a STAC domain. Only two members of this family have been characterized experimentally, the CrbS/R system that regulates acetate utilization in Vibrio and Pseudomonas, and the CbrA/B system that regulates the utilization of histidine in Pseudomonas and glucose in Azotobacter. In an attempt to expand the characterized members of this family beyond the Gammaproteobacteria, we identified two putative TCSTS in the Alphaproteobacterium Sinorhizobium fredii NGR234 whose sensor histidine kinases belong to the SLC5/STAC family. Using reverse genetics, we were able to identify the first TCSTS as a CrbS/R homolog that is also needed for growth on acetate, while the second TCSTS, RpuS/R, is a novel system required for optimal growth on pyruvate. Using RNAseq and transcriptional fusions, we determined that in S. fredii the RpuS/R system upregulates the expression of an operon coding for the pyruvate symporter MctP when pyruvate is the sole carbon source. In addition, we identified a conserved DNA sequence motif in the putative promoter region of the mctP operon that is essential for the RpuR-mediated transcriptional activation of genes under pyruvate-utilizing conditions. Finally, we show that S. fredii mutants lacking these TCSTS are affected in nodulation, producing fewer nodules than the parent strain and at a slower rate.
Two-component signal transduction systems (TCSTS) are regulatory systems that allow bacteria to respond to environmental stimuli by regulating different cellular processes like chemotaxis, nutrient acquisition and utilization, virulence, and antibiotic resistance. As their name implies, they are comprised of two elements, a histidine kinase that acts as the sensing and signal transducing component, and a response regulator, that usually activates or represses gene expression (Ann M. Stock et al., 2000; Tiwari et al., 2017). Most histidine kinases are membrane-bound proteins, detecting signals through extracellular sensory domains, which trigger a conformational change that is propagated across the membrane to the intracellular region of the protein, very often via a HAMP domain (Hulko et al., 2006). The conformational change induces the phosphorylation of a conserved histidine in the dimerization and histidine phosphotransfer domain (DHp) by the catalytic ATP-binding domain (CA). In many kinases, this process has additional levels of regulation by the action of other cytoplasmic domains, such as PAS and GAF (Gao and Stock, 2009). Finally, the phosphate group is transferred to a conserved aspartate in the receiver domain (REC) of response regulator proteins, translating the phosphorylation signal into a physiological response, often by modulating transcription (Galperin, 2010). In hybrid histidine kinases, a REC domain occurs on the same polypeptide with the core histidine kinase. This arrangement, that often is accompanied by the addition of extra elements in the phosphorelay, is believed to provide extra-fine-tuning mechanisms to the regulatory pathways involved (Borland et al., 2016).
CbrA and CrbS are histidine kinases belonging to a family of membrane-bound TCSTS sensor proteins in which the cytosolic histidine kinase domains are fused to a transmembrane SLC5 solute symporter domain through a STAC domain (Henriquez et al., 2021). The STAC domain is a recently described protein domain associated with bacterial signal transduction. Although its role is still unknown, it has been speculated that it could mediate interactions with other proteins or regulate the flow of substrates through the SLC5 domain (Korycinski et al., 2015).
CbrA is a histidine kinase that has been characterized as a global regulator of metabolism in the Pseudomonadaceae. Together with its cognate response regulator, CbrB, it regulates the utilization of histidine in P. fluorescens and P. putida (Zhang and Rainey, 2008; Zhang et al., 2015; Monteagudo-Cascales et al., 2019), and glucose in Azotobacter vinelandii (Quiroz-Rocha et al., 2017). Interestingly, it has been found that neither is the kinase domain of CbrA required for histidine uptake by the SLC5 domain nor the SLC5 domain essential for signal perception, strongly suggesting that stimulus sensing in this family occurs in the cytoplasm (Monteagudo-Cascales et al., 2019; Wirtz et al., 2020). Recently, it has been shown in P. putida that cbrX, a small ORF that is co-transcribed with cbrA, modulates CbrA translation by a mechanism of translational coupling (Monteagudo-Cascales et al., 2019).
CrbS is a hybrid histidine kinase, which regulates acetate utilization in Vibrio cholerae and the Pseudomonadaceae (Jacob et al., 2017; Henriquez and Jung, 2021). CrbR, its cognate response regulator, is a LuxR transcriptional activator that, in Gammaproteobacteria, induces expression of acetate utilization genes by binding to a conserved DNA sequence motif in their promoter region (Sepulveda and Lupas, 2017). Recently, it has been demonstrated that in V. cholerae the cAMP receptor protein (CRP)-cAMP system activates the expression of the crbS and crbR genes (Muzhingi et al., 2018).
Although SLC5/STAC-containing signaling proteins can be found among all phyla of bacteria (Korycinski et al., 2015), the work on this TCSTS family has been restricted mainly to the Gammaproteobacteria. Only the PrlS/PrlR system from Brucella melitensis, which plays a role in bacterial adaptation to ionic strength and persistence in mice (Mirabella et al., 2012), has been genetically characterized outside this phylum. In this work, we identified the CrbS/R system in the Alphaproteobacterium Sinorhizobium fredii and determined that, as in the Gammaproteobacteria, it is involved in acetate utilization. Moreover, we report the characterization of RpuS from Sinorhizobium fredii, which together with RpuR forms a two-component system that is required for optimal growth on pyruvate as the sole carbon source. Using RNAseq, we identified a plasmid-encoded sodium-solute symporter as its primary target and identified the minimal promoter region required for RpuS/RpuR regulation. Finally, we determined the effect that a deletion of this system has on nodulation.
Sinorhizobium fredii and Escherichia coli strains were grown in Peptone-Yeast medium (PY; Bravo and Mora, 1988). S. fredii NGR234 was obtained from the Culture Collection of the Centro de Ciencias Genómicas-UNAM. Experiments that required a defined carbon source were performed in Minimal Media (MM; NaNO3 2.5 g/l; KH2PO4 1 g/l; K2HPO4 2.0 g/l; MgSO4-7H2O 0.3 g/l; NaCl 0.1 g/l; FeCl3 0.01 g/l) supplemented with 1 ml/l of Nitsch’s trace elements (Atlas, 2010), and a carbon source at a final concentration of 20 mm. When required, antibiotics were added at the following concentrations: gentamicin (Gm), 25 μg/ml; kanamycin (Km), 30 μg/ml; nalidixic acid (Nal), 20 μg/ml; rifampicin (Rif), 25 μg/ml. For selection against pK18mobSacB, PY agar plates were supplemented with 10% sucrose. Genes cloned in pSRKgm were induced by the addition of 100 μm of isopropyl-β-D-thiogalactopyranoside (IPTG).
Pre-cultures for assays in MM were prepared in 5 ml PY and washed with MM as described previously (Zhang et al., 2015). Growth curves were performed by incubating 25 ml cultures at 30°C in an orbital shaker incubator at 150 rpm and monitored by reading 100 μl samples using a Multiskan Sky plate reader and the SkanIt 6 software (Thermo Fisher, Waltham, United States). Replicates for growth curves, transcriptional fusion measurements, and RNAseq are biological, derived from independent cultures.
Bacterial strains and plasmids used in this study are listed in Table 1; primer sequences are listed in Supplementary Table S1. All molecular techniques were conducted following standard protocols (Sambrook et al., 2012). S. fredii NGR234 (Stanley et al., 1988) was transformed by electroporation as described previously (Ferri et al., 2010). All constructs were verified by sequencing (Eton Bioscience, Inc., Sand Diego, United States) and all mutants were confirmed by PCR.
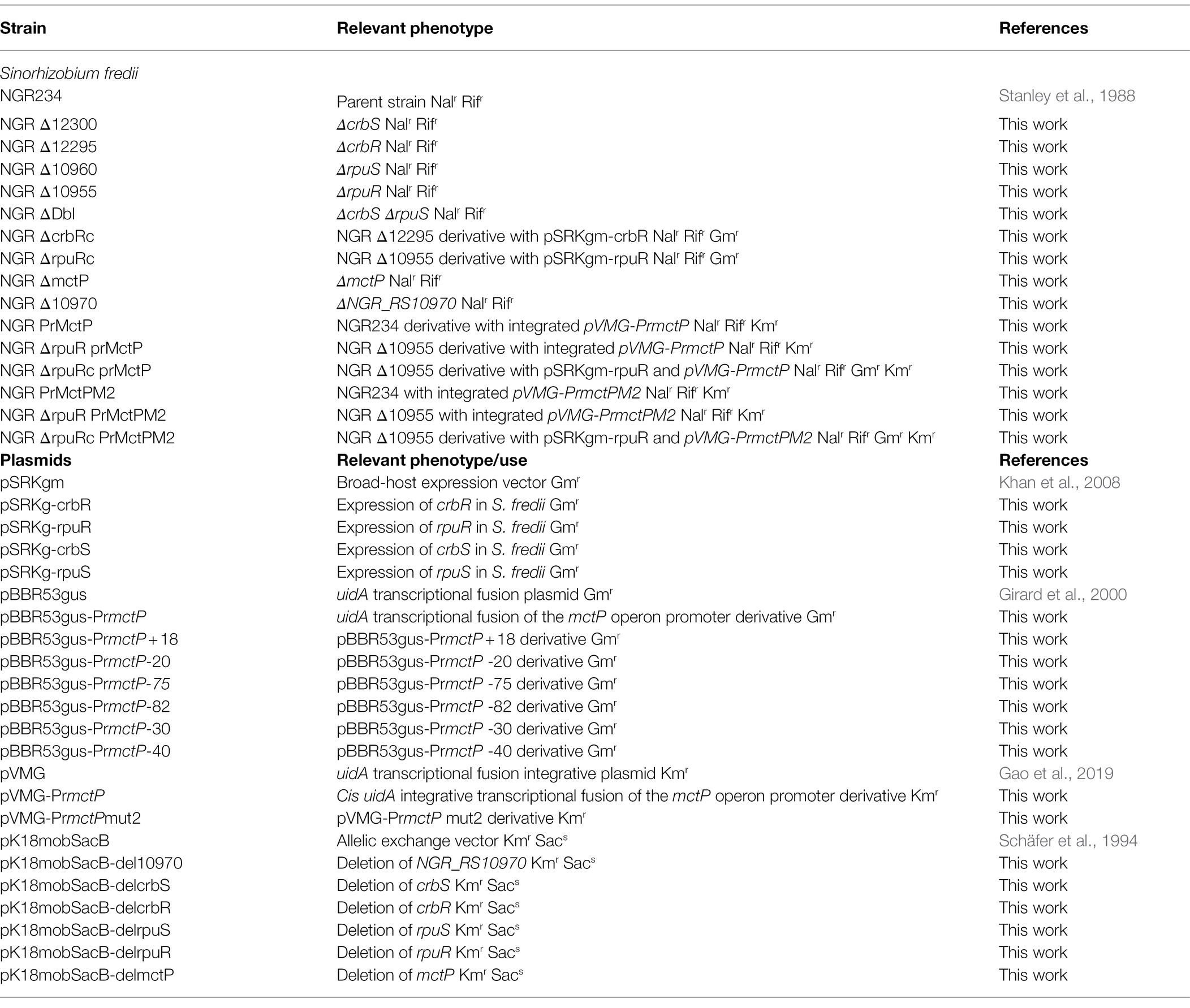
Table 1. Strains and plasmids used in this work.
Strains with deletions in specific genes were constructed by allelic exchange and selected by sucrose sensitivity, using plasmid pK18mobSacB (Schäfer et al., 1994). The pK18mobSacB derivatives for the deletion of rpuS and crbS were constructed by overlap extension (Lee et al., 2010) of two PCR products corresponding to ~750 bp of the upstream and downstream sequence of the targeted gene, and the resulting product was re/amplified and cloned in the HindIII/EcoRI and HindIII/SmaI sites of plasmid pK18mobSacB, respectively. The pK18mobSacB derivatives for the deletion of rpuR, cbrR, NGR_RS10965, and mctP were obtained by cloning in the HindIII/EcoRI sites of plasmid pK18mobSacB a PCR product containing the gene to be deleted, together with ~750 bp of upstream and downstream sequence. Then the targeted gene was deleted by total PCR of the plasmid with phosphorylated divergent primers (Imai et al., 1991). The resulting PCR product was ligated, and the religated plasmid was recovered by transformation.
Plasmids pSRKgm-rpuR and pSRKgm-crbR were built by cloning the ORF of each gene into the NdeI/BamHI sites of plasmid pSRKgm (Khan et al., 2008). Plasmids pSRKgm-rpuS and pSRKgm-crbS were built by cloning the ORF of each gene in plasmid pSRKgm by Gibson cloning using the GeneArt Gibson Assembly HiFi Master Mix (Thermo Fisher, Waltham, United States).
The in-cis uidA transcriptional fusion of the mctP operon was constructed by cloning a PCR product of the putative promoter region in the SalI/BamHI sites of plasmid pVMG (Gao et al., 2019) producing plasmid pVMG-PrmctP. Mutagenesis of the mctP operon promoter was achieved by the quick-change strategy (Agilent, Santa Clara, United States) using plasmid pVMG-PrmctP as a template. After electroporation, plasmid cointegration and orientation were verified by PCR as described previously (Gao et al., 2019).
The in-trans uidA transcriptional fusion of the mctP operon promoter was constructed by cloning a PCR product of the putative promoter region in the BcuI/BamHI sites of plasmid pBBR53Gus (Girard et al., 2000). Transcriptional fusions used to map the NGR_RS10970 operon promoter were built by amplifying plasmid pBBR53g-PrmctP with divergent primers carrying a BcuI site or a BamHI site. The resulting PCR product was digested with the corresponding restriction enzyme, ligated, and transformed for the recovery of the reconstituted plasmid.
Two 20-ml cultures of the appropriate strains were inoculated from independent pre-cultures to an A620 of 0.1 in minimal medium supplemented with pyruvate and incubated at 150 rpm and 30°C for 12 h. RNA was isolated from 500 μl aliquots using the RiboPure RNA Purification Kit for bacteria (Thermo Fisher Scientific, Waltham, United States) and RNA purity and integrity were checked by agarose gel electrophoresis. Then, to eliminate the residual DNA, the samples were treated with DNase-I using the RQ1 RNase-Free DNase Kit (Promega, Madison, United States).
The RNA samples treated with DNase-I were used to prepare high throughput sequencing (HTS) libraries. First, rRNA depletion was performed using the QIAseq FastSelect—5S/16S/23S Kit (Qiagen, Hilden, Germany). Then, the resulting depleted and fragmented mRNA was used as input for the first-strand cDNA synthesis step of the TruSeq Stranded Kit (Illumina, San Diego, United States) and sequenced on an Illumina MySeq platform at the CICESE Functional Genomics Laboratory facility (Ensenada, Baja California, México), with a paired-end protocol and read length of 75 nt (PE 2 × 75), resulting in a total output of approximately 3.6 to 5.2 million reads per sample.
All read outputs were checked to satisfy Illumina quality standards (Ewels et al., 2016; Wingett and Andrews, 2018). Raw data were cleaned up with trimmomatic/0.36 (Bolger et al., 2014) to remove sequences originating from Illumina adaptors and low-quality reads. Files were aligned to the reference genome [RefSeq accession numbers NC_012587 (chromosome), NC_000914 (plasmid a), and NC_012586 (plasmid b)] with bowtie2/2.3.5.1(Langmead and Salzberg, 2012) using -N 0 (no mismatches accepted in the alignment) and --no-unal (suppress SAM records for unaligned reads) settings. All the other settings were implemented with default options. Once the alignment was completed, samtools/1.12 (Li et al., 2009) was used to select the reads that were aligned in proper pairs, and to sort them. The reads counting per gene was performed with htseq/0.9.1 (Anders et al., 2014) using stranded mode and gene as a feature. Differential expression analysis was performed with DESeq2/1.28.1(Love et al., 2014). Genes with adjusted p-values <0.05 and log2 fold changes ≥1 were considered differentially expressed. Differential expression plots were prepared in R/4.0.3 (R Core Team, 2021) using the packages ggplot2/3.2.5 (Wickham and Sievert, 2016) and EnhancedVolcano/1.6.0 (Blighe and Lewis, 2021).
Five-milliliter cultures of the appropriate strains were inoculated from three independent pre-cultures to an A620 of 0.1 in MM supplemented with succinate or pyruvate and incubated at 30°C for 8 h. β-glucuronidase activity expressed in modified Miller units was quantified as described previously (Jefferson et al., 1986). Photometric measurements were performed with a Multiskan Sky plate reader (Thermo Fisher, Waltham, United States).
Common bean Phaseolus vulgaris cv. Negro Jamapa seeds were surface-sterilized by sequential immersions in 96% (v/v) ethanol for 30 s and 6% sodium hypochlorite for 7 min. Seeds were subsequently rinsed several times with sterilized water and placed for germination on plates containing 1% water agar at 30°C for 48 h in the dark. Nodulation kinetic assays were performed as described previously (Rodríguez et al., 2020). Briefly, 2-day-old seedlings were transferred to growth pouches (CYGTM) containing 10 ml of nitrogen-free Fahräeus nutrient solution. Bacterial inocula were grown overnight in PY liquid medium, concentrated by centrifugation, and washed once in a solution of MgSO4 10 mm. Bacterial suspensions were adjusted at an A600 of 0.3 and seedlings were inoculated by direct addition to each root of appropriate amounts of the bacterial solution, to give a final bacterial density of 108 cells ml − 1 in the growth pouch. Pouches were kept under controlled environmental conditions (14/10-h light/dark cycle, 22°C/16°C, and 60 to 70% relative humidity). For each bacterial strain, eight growth pouches, containing one seedling each, were monitored daily for nodule appearance and the number of nodules was registered daily for 30 days.
The global search for SLC5/STAC proteins was performed as described previously (Korycinski et al., 2015). GeConT (Martinez-Guerrero et al., 2008) was used to search for homologs of the rpuR and rpuS to retrieve their putative promoter region. MEME (Bailey et al., 2009) and FIMO (Grant et al., 2011) searches were run with the default options searching for a motif between 6 and 30 nucleotides long. Sequence alignments and analyses were performed in the MPI bioinformatics toolkit (Zimmermann et al., 2018; Gabler et al., 2020) using MUSCLE (Edgar, 2004), Quick2D, HHpred (Hildebrand et al., 2009), PCOILS (Lupas et al., 1991), and MARCOIL (Delorenzi and Speed, 2002). Transmembrane segments were predicted with FMHMM (Sonnhammer et al., 1998) and Phobius (Käll et al., 2004). For transcriptional fusions, statistical comparisons between samples were performed by ANOVA and Tukey’s multiple comparison test using SPSS version 25 (IBM).
The raw RNA-sequencing data are deposited in SRA under the accession numbers SRR16684185 and SRR16684186 for wild type, and SRR16684183 and SRR16684184 for mutant samples. The total reads and the differential expression analysis tables are available as Supplementary Tables S3 and S4, respectively.
A global search for SLC5/STAC proteins allowed us to identify two proteins belonging to this family in Sinorhizobium fredii NGR234, NGR_RS12300, and NGR_RS10960. Both are hybrid histidine kinases, with a REC domain at the C-terminal end as found in CrbR. The SLC5 domain is composed of 13 transmembrane segments, linked to a histidine kinase via STAC, then PAS, then a coiled-coil segment (Supplementary Figure S1). The NGR_RS12300/NGR_RS12295 system is located on the chromosome. Both genes are adjacent but are transcribed convergent from each other and NGR_RS12295 codes for a response regulator of the LuxR family, like CrbR. On the other hand, the NGR_RS10960/NGR_RS10955 system is located on the pNGR234b megaplasmid. Both genes are predicted to form an operon, and gene NGR_RS10955 also codes for a response regulator of the LuxR family (Figure 1).
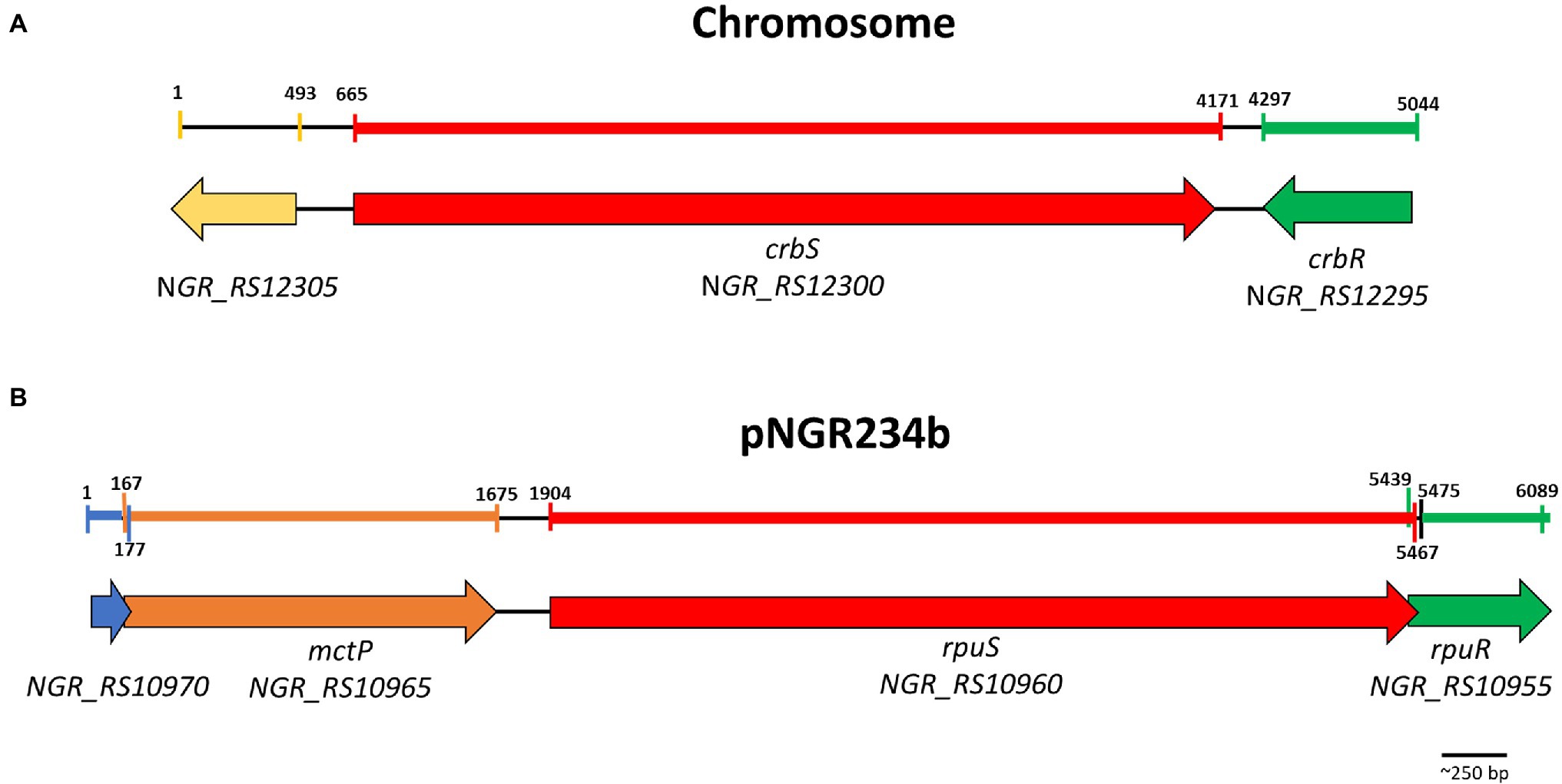
Figure 1. Genomic context of the CrbS/R and RpuS/R TCSTS. Genes are annotated as follows Red, SLC5/STAC Histidine kinase; Green, LuxR family response regulator; Orange, monocarboxylate permease; Blue, DUF3311 domain-containing protein; Yellow, Large-conductance mechanosensitive channels mscL family. Colored areas in the scale bars indicate the region deleted for the construction of the mutant strain in the corresponding gene.
To determine the biological role that NGR_RS12300 and its cognate response regulator play in S. fredii, we first constructed a ΔNGR_RS12300 mutant and screened it for its ability to grow in solid minimal media supplemented with different carbon sources. Our results showed that the ΔNGR_RS12300 strain was unable to grow on acetate as the sole carbon source (Figure 2; Supplementary Figure S3). These results suggested that the NGR_RS12300 and NGR_RS12295 genes are the S. fredii homologs of CrbS and CrbR, respectively. To further test this hypothesis, we constructed ΔNGR_RS12295 (ΔCrbR) strains that, as expected, was unable to grow on acetate. When both mutants were complemented, growth on acetate was successfully recovered (Figure 2A). Our results are congruent with the characteristics of the CrbS/R system in the Gammaproteobacteria, therefore we renamed NGR_RS12300 and NGR_RS12295 to crbS and crbR, respectively (Supplementary Table S2).
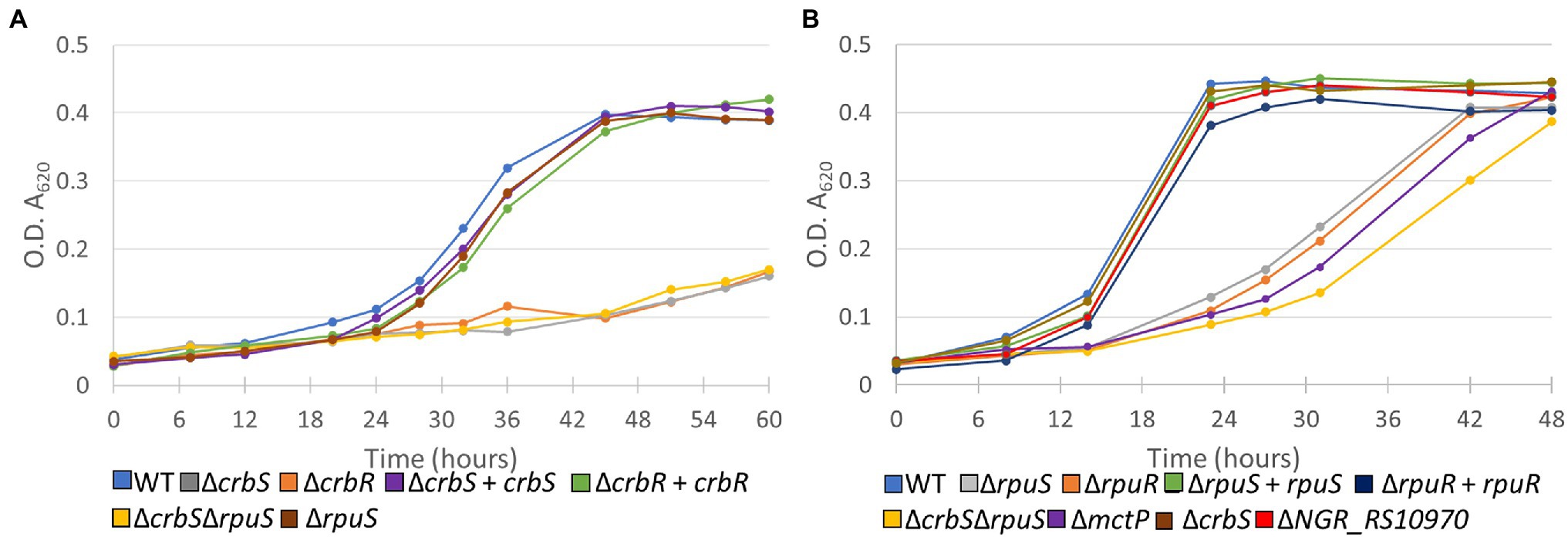
Figure 2. Growth curves of deletion mutants of (A) CrbS/R and (B) RpuS/R TCSTS. Bacteria were grown in minimal medium complemented with (A) acetate or (B) pyruvate as sole carbon sources. Medium was not complemented with Kanamycin in any case. Expression from plasmid pSRKgm was induced with 100 mm IPTG. Results are means for six independent cultures. Empty pSRKgm controls for deleted mutants and non-induced controls for complemented strains were also tested but showed a negligible effect on growth phenotype and are not depicted.
The putative NGR_RS10960/NGR_RS10955 system was of particular interest to us because all the previously characterized two-component systems of the family are located on the chromosome, while this one is located on the pNGR234b megaplasmid. Located 220 bp upstream of NGR_RS10960 is another operon comprised of two genes, NGR_RS10965 and NGR_RS10970, of which NGR_RS10965 codes for a sodium:solute symporter family protein (Figure 1). This was evocative because in P. fluorescens the CrbR/S system regulates the actP gene, which also encodes a sodium-solute symporter family protein (Jacob et al., 2017) but this is not necessary for optimal growth in acetate (Sepulveda and Lupas, 2017). Rather, a BLASTP similarity search of NGR_RS10965 indicated that it is highly similar to the Rhizobium leguminosarum permease MctP, which transports alanine, pyruvate, and lactate, and is required for optimal growth on pyruvate as sole carbon source (Hosie et al., 2002). In R. leguminosarum and other species of the genus Rhizobium, mctP is regulated by the two-component system MctS/R. MctS is a transcriptional regulator integrated by a molecule-binding Cache2 domain and a Histidine kinase domain. These observations suggested to us that the NGR_RS10960 and NGR_RS10955 system is a new two/component system regulating mctP.
To test our hypothesis, we first constructed a ΔNGR_RS10960 mutant and tested it for its ability to utilize alanine, pyruvate, and lactate as sole carbon sources. Our results showed that the mutant strain was able to grow on alanine and lactate but showed impaired growth on pyruvate (Supplementary Figure S2). Next, we constructed a ΔNGR_RS10955 strain and compared its growth and that of ΔNGR_RS10960 with that of the wild-type strain in liquid minimal medium supplemented with pyruvate. Our results showed that the parent strain reached the stationary phase ~24 h earlier than both mutants. Moreover, when both strains were complemented, growth rate was almost recovered to parent strain levels (Figure 2B). Overall, these results showed that the NGR_RS10960 and NGR_RS10955 constitute a two-component signal transduction system that is involved in the regulation of pyruvate uptake and therefore we renamed the genes to rpuS and rpuR, respectively (Supplementary Table S2). As expected, a double ΔcrbSΔrpuS mutant was not able to utilize acetate and showed growth retardation when growing on pyruvate (Figure 2; Supplementary Figure S2). Additionally, our results show that the ΔcrbS strain is able to grow normally in pyruvate and the ΔrpuS mutant was able to utilize acetate. Thus, the two systems appear to operate independently without substantial crosstalk (Figure 2; Supplementary Figure S2).
Interestingly, although the rpuS deletion also deleted the original ATG start-codon and ribosomal binding site (RBS) as well as the 5′-sequence of rpuR (Figure 2), in-trans complementation of the rpuS deleted strain was able to restore growth in pyruvate (Figure 3). Therefore, it can be concluded that in the rpuS strain rpuR is being translated from the SD of rpuS and that the missing amino acids are not essential for RpuR function.
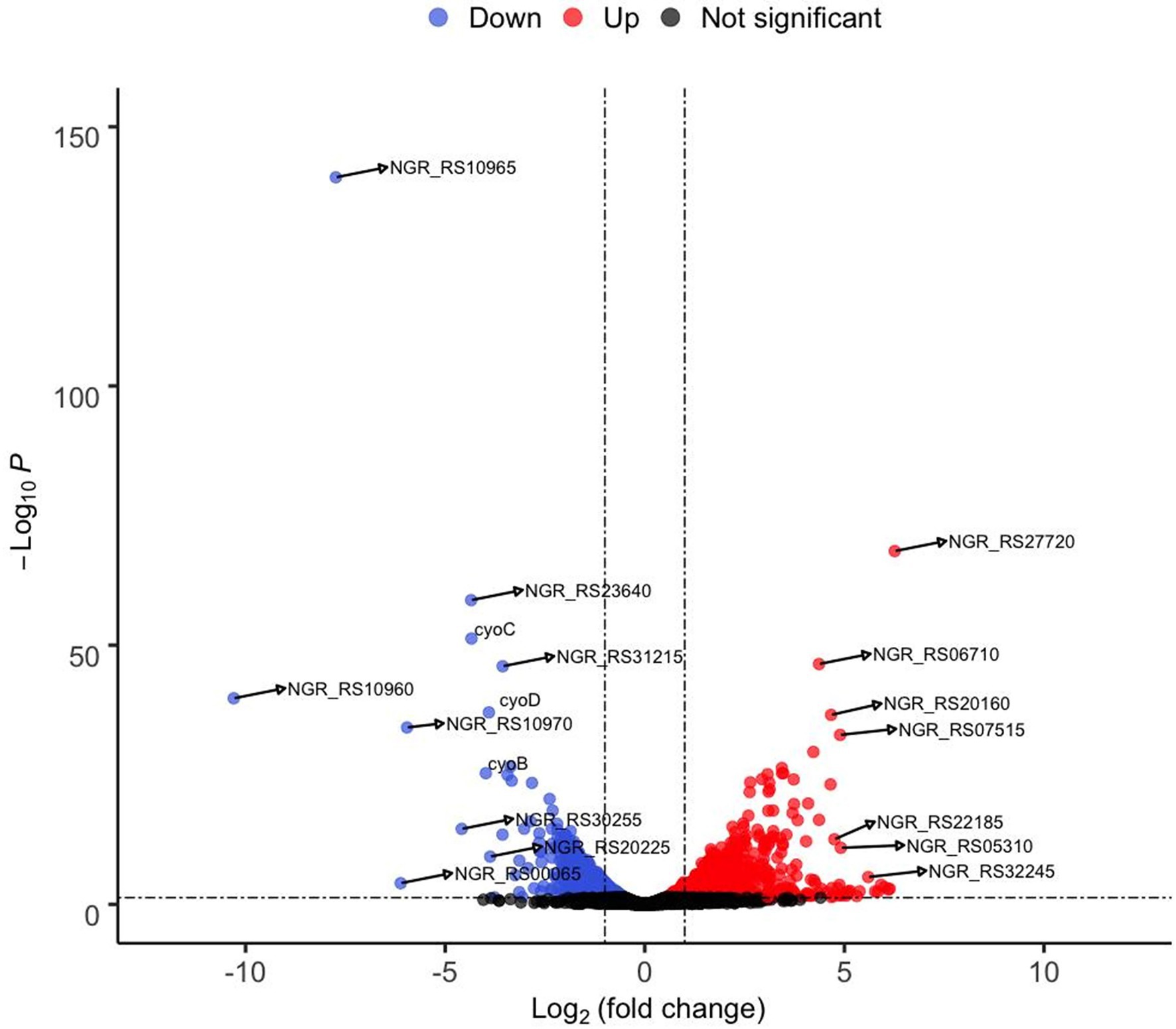
Figure 3. Volcano graph showing fold change in expression of genes in the ΔrpuS S. fredii strain compared to the parental strain (NGR234) when growing on pyruvate. Cultures were inoculated to a A620 of 0.1 in minimal medium supplemented with pyruvate and incubated at 150 rpm and 30°C for 12 h. RNA was isolated from 500 μl aliquots. Colored dots indicate upregulated (red) and downregulated (blue) genes. Labeled dots correspond to some of the genes listed in Table 2.
To confirm if RpuS/R regulates the expression of mctP and to identify additional genes involved in pyruvate utilization that are regulated by this new system, we performed an RNAseq analysis to compare strains NGR234 (parent strain) and ΔrpuS after incubation for 12 h in minimal medium supplemented with pyruvate. Our results show a clear transcriptome profile specific to the mutant strain in a sample distance analysis against the parent strain (Supplementary Figure S3). The main characteristics are the repression of genes involved in energy generation and the induction of stress-related proteins and transmembrane transporters (Table 2; Figure 3). These results are consistent with the cells being under starvation, by the lack of a carbon source. Among other genes that show a small but not significant decrease in expression in the mutant strain when compared to the wild-type strain, we found pycar (Supplementary Table S4). This gene codes for the Pyruvate carboxylase enzyme (EC 6.4.1.1,) a central component of pyruvate metabolism that transforms pyruvate into oxaloacetate, which then can be fed into the TCA cycle to produce ATP (Dunn, 1998).
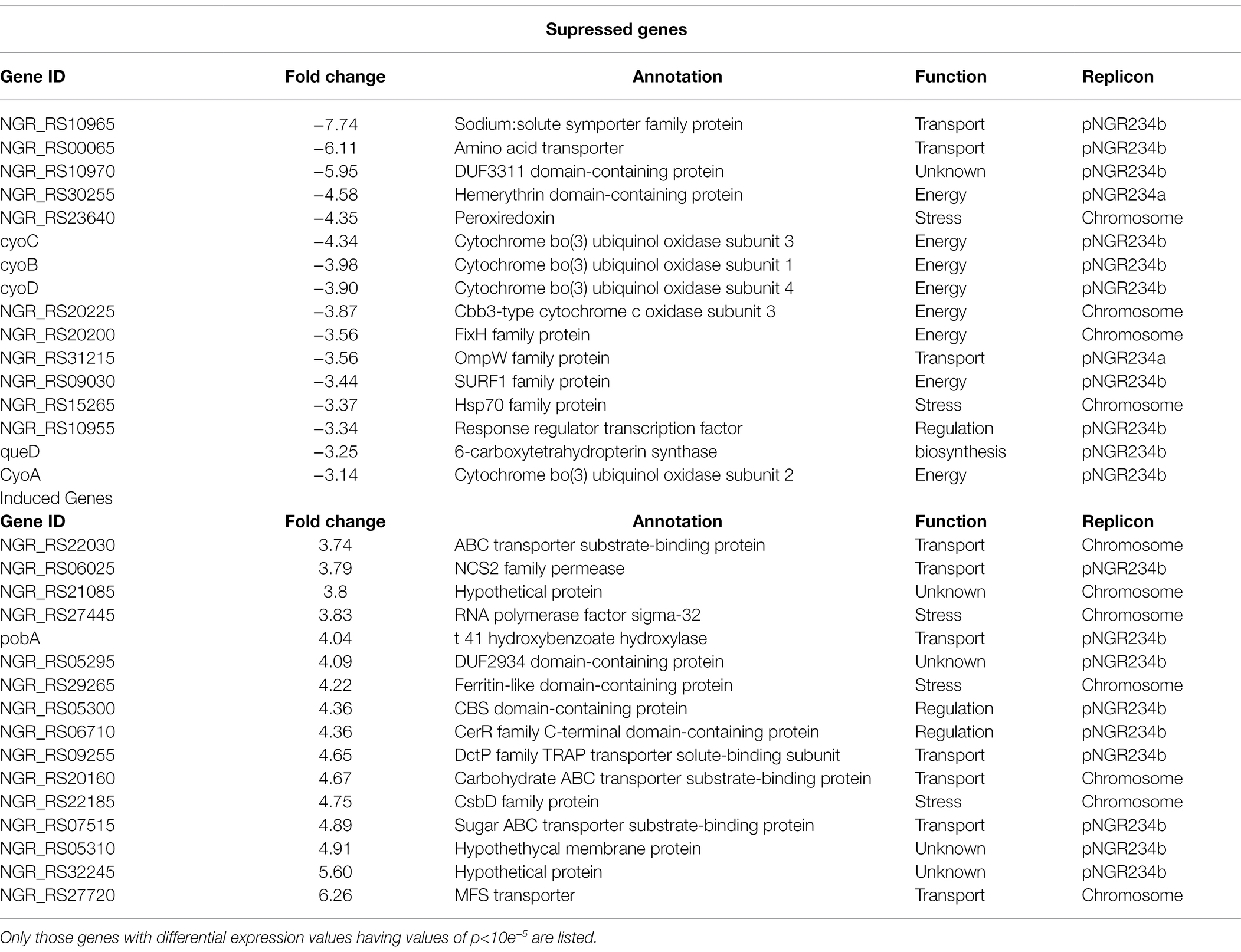
Table 2. Selection of differentially expressed genes in the S. fredii NGR234 ΔrpuS strain under pyruvate-utilizing conditions.
Surprisingly, only the operon containing the genes mctP and NGR_RS10970, showed a relevant decrease in expression (Figure 3). To understand the role that these proteins have in pyruvate utilization we constructed deletion mutants of each of the genes and challenged them to grow on minimal medium supplemented with pyruvate. While the ΔNGR_RS10970 mutation showed a negligible effect on growth, the ΔmctP strain displayed a ~ 24 h delay in reaching the stationary phase, just like the ΔrpuS and ΔrpuR strains (Figure 2B).
Next, we studied the sequence properties of the RpuS/R-responsive mctP promoter. We used the RNAseq data to predict the transcription initiation site (TIS) to be located ~100 base pairs upstream from the first codon of NGR_RS10970 (Cervantes-Rivera and Puhar, 2020). Approximately 22 bp upstream of the TIS lies a sequence consistent with the −10 box consensus but we were not able to identify a − 35 box. To begin the characterization of the putative pyruvate-induced promoter, we built an in-cis β-glucuronidase (uidA) transcriptional fusion of the 220 bp intergenic region between the mctP operon and the next upstream gene and measured its activity in minimal medium supplemented with pyruvate or succinate in a parent or a ΔrpuR background. Congruently with the RNAseq results, the transcriptional fusion was activated in the parental background only under pyruvate-utilizing conditions but not on the ΔrpuR strain. When the mutant strain was complemented with rpuR cloned in an IPTG-inducible plasmid, activation of the promoter was restored upon induction of rpuR expression with IPTG (Figure 4, blue bars). To determine the minimal region necessary for RpuR-dependent regulation, we built in-trans transcriptional fusions from several derivatives of the mctP promoter and measured their activity in minimal medium supplemented with pyruvate in a parent or a ΔrpuR background. While none of the derivatives show induction in the ΔrpuR strain (data not shown) our results in the parent strain outlined a minimal inducible promoter encompassing the region between the −82 and + 1 nucleotides from the TIS (Figure 5A). Moreover, we observed a ~ 65% drop in the induction of the PrmctP-75 derivative compared to the Prmctp-82 derivative, but not a complete loss of induction as with other fragments. These results indicated that an important element for RpuS/R-mediated induction could be located around nucleotide −75 (Figure 5A). A closer examination unveiled an imperfect direct repeat located between the −80 and − 43 bases from the TIS (Figure 5B). To determine its relevance to the RpuS/R-mediated mctP induction, we performed a search for rpuS in the Rhizobiaceae. Our results led to the identification of four Bradirhizobium and five Sinorhizobium strains in which this gene is also neighboring a mctP homolog (Supplementary Figure S4). Notably, the direct repeat is conserved in the mctP promoter region of these strains (Figure 5B), suggesting it may play a relevant role in the regulation of the mctP operon. This hypothesis is strengthened when comparing the complete loss of activity of the fusion PrmctP-40 in which both repeats are absent, with the reduced induction of the PrmctP-72 fusion in which the cloning of the mctP promoter derivative for the construction of the transcriptional fusion resulted in a change of 6 bp of the first repeat, but not in its complete deletion (Figure 5B).
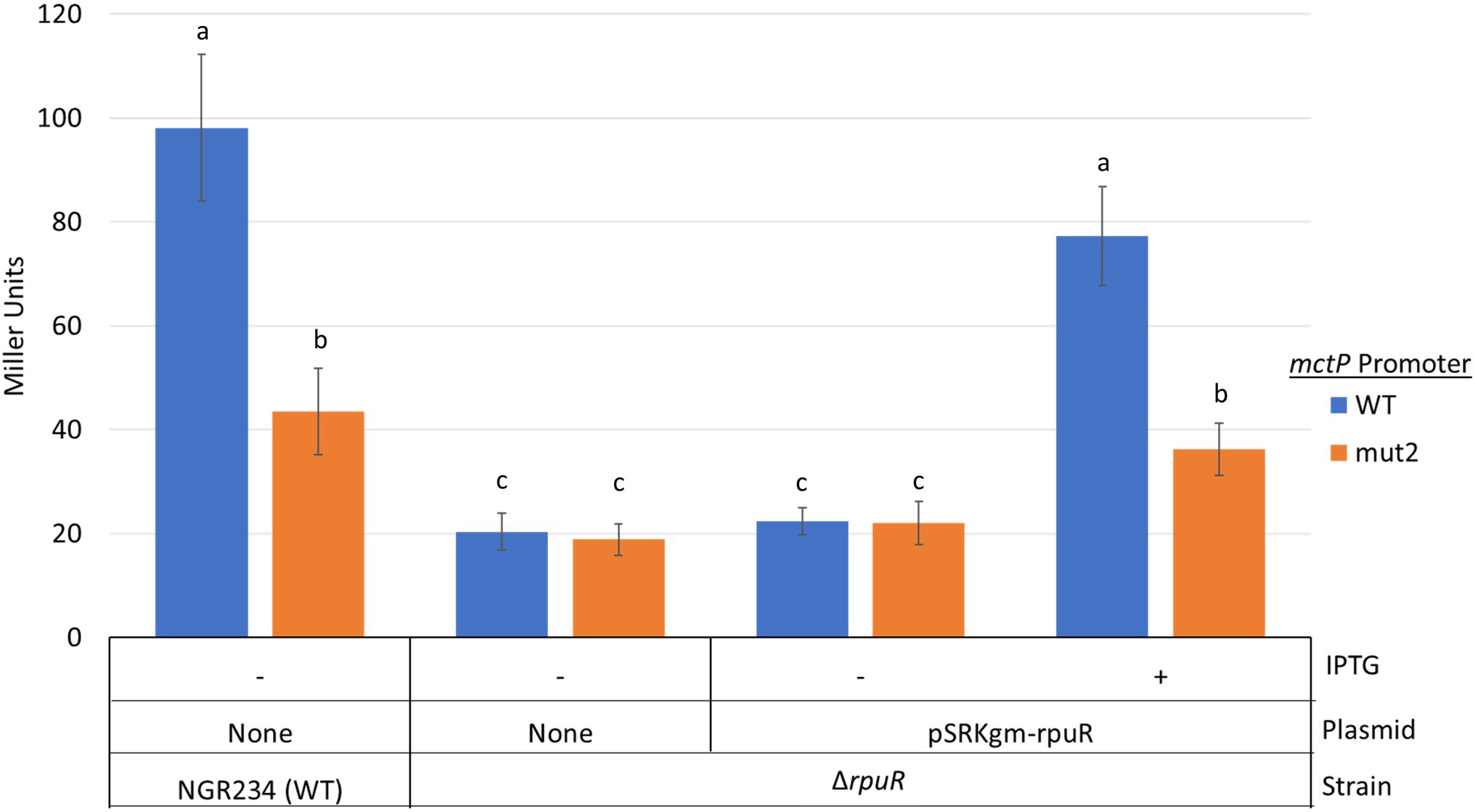
Figure 4. Activity of in-cis transcriptional fusions of the of the NGR_RS10970-mctP operon promoter, and its mut2 derivative, in various genetic backgrounds under pyruvate-utilizing conditions. Miller units are expressed as means ± Standard deviations (error bars) of results from at least three independent experiments. Means accompanied by the same letter are not significantly different (p < 0.05). NGR234, parent strain; ΔrpuR, rpuR deleted strain; +, induction of rpuR expression from plasmid pSRKgm with 100 mm IPTG. Cultures were inoculated to a A620 of 0.1 in minimal medium supplemented with pyruvate and incubated at 150 rpm and 30°C for 12 h.
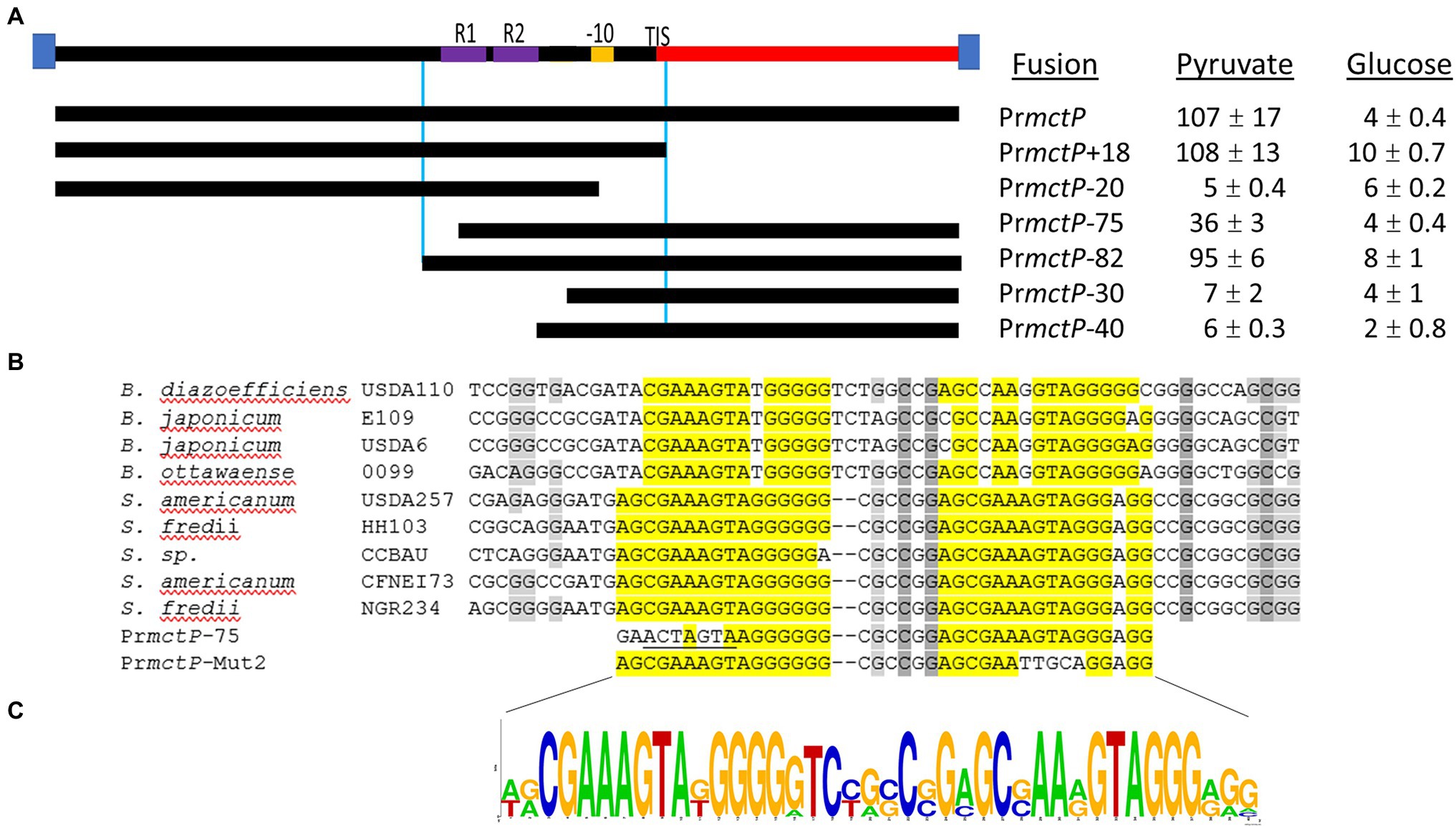
Figure 5. Mapping of the NGR_RS10970-mctP operon promoter. (A) Activities of in-trans transcriptional fusions of the NGR_RS10970-mctP operon promoter and its derivatives in a wild-type strain growing on pyruvate or glucose as sole carbon sources. Miller units are expressed as means ± Standard deviations of results from at least three independent experiments. (B) Sequence of the mctP operon promoter in different members of the Rhizobiaceae and of the mctP derivatives with mutations in the first (PrmctP-75) and second (PrmctP-Mut2) repeat. (C) Sequence Logo of the identified direct repeat obtained from the aligned sequences of the NGR_RS10970-mctP operon promoter of different members of the Rhizobiaceae.
To further demonstrate the relevance of the direct repeat in the activation of the mctP promoter by the RpuS/R we built an in-cis uidA transcriptional fusion with a fragment of the PrmctP derivative in which 4 bp of the second repeat were changed from AGTAG to TTGCA (Figure 5B, PrmctP-mut2). Then, we measured its activity under pyruvate-utilizing conditions in a parent or a ΔrpuR background. In concordance with our previous results, the transcriptional fusion was activated in the parent and in the complemented ΔrpuR strain (upon induction with IPTG), but only at ~40% rate when compared with the parental promoter; as expected, no activation was detected in the ΔrpuR background (Figure 4, orange bars).
Finally, we used MEME to search for this motif in a 500 bp putative promoter region immediately upstream of the other top differentially expressed genes identified through our RNAseq assay (Table 2). Neither pycar nor the other genes contained the imperfect direct repeat in the searched region.
Altogether, our results confirm the role of the direct repeat in the pyruvate-dependent induction of the mctP promoter and demonstrate that this induction is contingent on the presence of the RpuS/R system.
Finally, we performed nodulation kinetic assays in beans to assess the effect that the deletion of the CrbS/R or the RpuS/R systems may have on this relevant biological process. Our results showed that the ΔcrbS mutant strain had a significant delay of approximately 7 days on nodulation in all the treated plants and at the end of the experiment produced ~50% fewer nodules than the parent strain. On the other hand, the deletion of rpuS had a milder effect on nodulation delay. Plants inoculated with the ΔrpuS strain showed ~33% less nodules than the parent strain after 30 days (Figure 6).

Figure 6. Common bean nodulation kinetics of S. fredii NGR234 (blue) and its ΔrpuS (red) and ΔcrbS (green) derivative mutant strains. Bars represent the average number of nodules per condition per day of 8 plants ± SD per condition. An asterisk over bars indicates significant differences (p < 0.05).
The study of TCSTS with SLC5/STAC histidine kinases as the sensing and signal transmission component has focused mainly on the Pseudomonadales and other Gammaproteobacteria. In this work, we have expanded the characterization of this family into the Alphaproteobacteria by identifying a homolog of the CrbS/R system and characterizing the novel RpuS/R system as a mctP regulator in S. fredii NGR234.
The identification of the RpuS/R system shows that there are two lineages of rhizobia dependent on MctP for optimal pyruvate utilization, but that these are under the control of different regulatory systems, specifically the MctS/R system and the RpuS/R system identified here. A search of the GeConT database for MctP homologs in nitrogen-fixing Alphaproteobacteria returned 26 hits (Table 3; Supplementary Figure S4). Seventeen are adjacent to a MctS/R system and are found in bacteria of the genus Rhizobium (family Rhizobiaceae). Nine systems are adjacent to a RpuS/R system (Supplementary Figure S4) and are found in bacteria belonging to the promiscuous genus Sinorhizobium (family Rhizobiaceae) and the slow-growing genus Bradyrhizobium (family Nitrobacteraceae). From the MctS/R associated hits, very few are located on the chromosome, only those from R. tropici and Agrobacterium, while from the MctS/R associated hits, all those that come from Bradyrhizobium are located on the chromosome; Bradyrhizobia are known for having characteristic large genomes, and very few plasmids (Ormeño-Orrillo and Martínez-Romero, 2019). From the Sinorhizobium genus, only strain USDA257 that is missing one repABC plasmid (Schuldes et al., 2012) has a chromosomally encoded mctP gene. The rest of the hits, in Rhizobium and Sinorhizobium, are located on repABC megaplasmids. In the Rhizobiaceae, all the plasmids with a MctS/R associated mctP gene belong to the p42c family, as defined by their RepC protein (González et al., 2019). Although no detailed genealogy has been determined for the plasmids of Sinorhizobium, it has been shown that the non-symbiotic repABC plasmids from the genus differ significantly from those of the Rhizobiaceae (Wang et al., 2018; González et al., 2019). Moreover, plasmid pNGR234b of S. fredii is considered a hot spot for lateral transfer because of the presence of 142 transposases, as well as the determination that ≥40% of its encoded genes are not strain-specific and were probably acquired from a wide variety of other microorganisms (Streit et al., 2004; Schmeisser et al., 2009). Overall, these observations suggest a crucial role of recombination and lateral transfer in the acquisition of the mctP gene by rhizobia and its association with different regulatory systems.
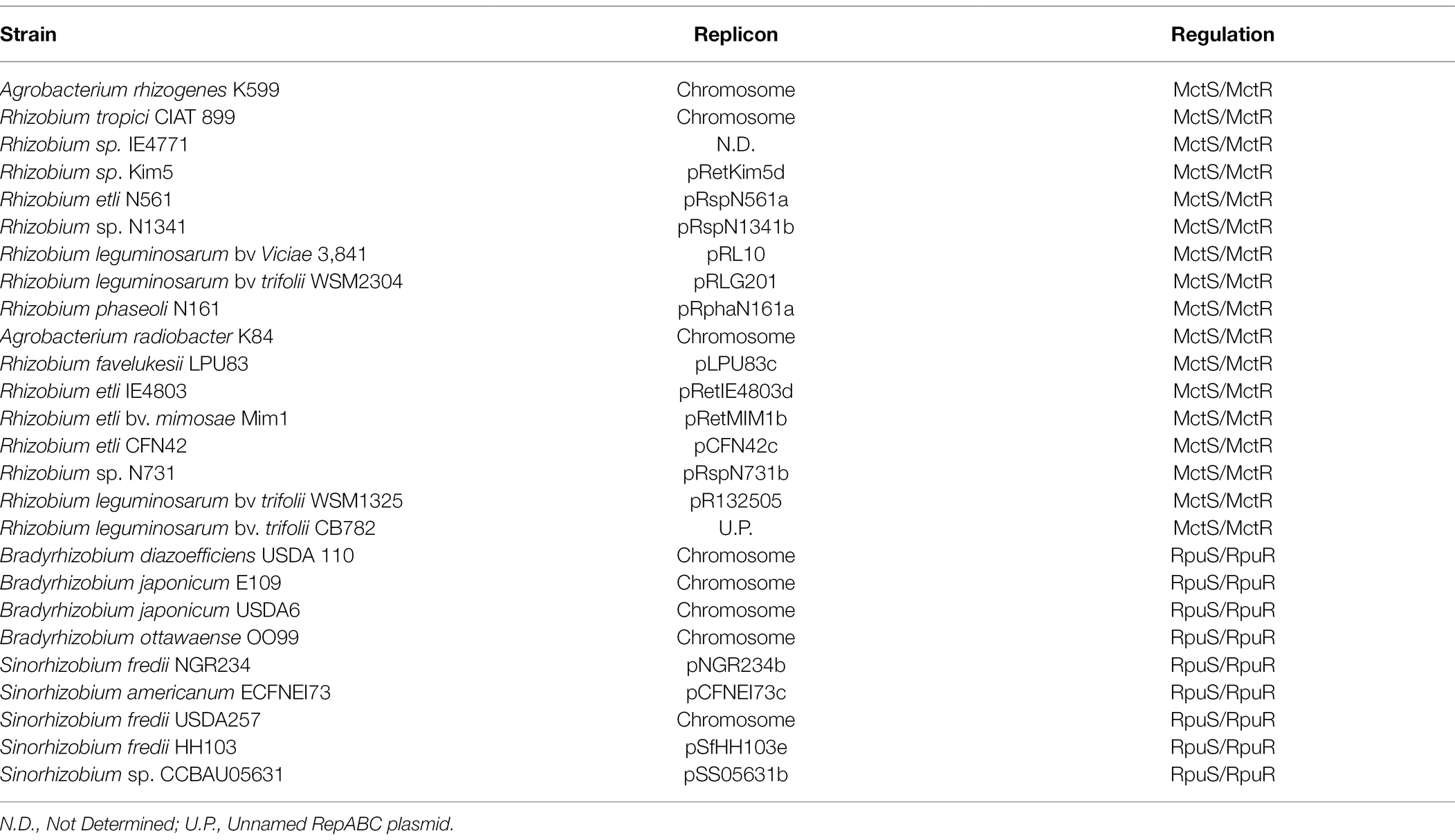
Table 3. Rhizobia with mctP neighbored by a TCSTS.
Although surprising, the finding that mctP is one of the genes under the control of the RpuS/R system is biologically sound. In rhizobia, the regulation of pyruvate utilization through the enzymes pyruvate carboxylase (EC 6.4.1.1) and pyruvate dehydrogenase (EC 1.2.4.1) is complex, including transcriptional and post-translational mechanisms, depending on the availability of different compounds, and varies among different organisms (Dunn, 1998; Bolger et al., 2014; Geddes and Oresnik, 2014). Therefore, acquiring a modular improvement that enhances substrate acquisition and provides an ecological advantage over other microorganisms is a straightforward addition to an intricate network that otherwise may have little flexibility for modification. These observations also highlight the adaptation of TCST sensors, particularly those belonging to the SL5/STAC family, as regulators of genes requiring early detection and fast response to subtle concentration changes of substrates in a highly competitive environment. Interestingly, the ΔmctP, ΔrpuS, and ΔrpuR strains showed remanent growth on pyruvate as the sole carbon source after 24 and 72 h of incubation in liquid and solid MM, respectively (Figure 2; Supplementary Figure S3). This observation is consistent with previous reports in Rhizobium leguminosarum that show that ΔmctP, ΔmctS, and ΔmctR strains were still able to grow on alanine and pyruvate, although at a lower rate (Hosie et al., 2002). These results could be explained by a leaky expression of the mctP promoter in the ΔrpuS, and ΔrpuR strains or by the slow assimilation of pyruvate by less efficient transporters. It is not uncommon for bacteria to have several transporters for pyruvate transport; for example, in E. coli at least two uptake systems and one excretion system for pyruvate have been identified (Kreth et al., 2013). Finally, the fact that we were unable to locate the imperfect direct repeat in the putative promoter region of any of the other genes that showed differential expression in our RNAseq assays suggests that they are not part of the RpuS/R regulon. Nonetheless, more experimental evidence is required to confirm this hypothesis.
Of particular interest was our finding that both the ΔcrbS and ΔrpuS strains start nodulation later than the parental strain. Several works have shown different impacts on bacteria–host interactions caused by the deletion of the SLC5/STAC TCSTS. For example, it was found that V. cholerae strains lacking CrbS or CrbR are less virulent when compared with the parent strain. At the same time, the equivalent mutants in P. aeruginosa and P. entomophila are fully pathogenic in Drosophila (Jacob et al., 2017). It was also recently reported that the root colonization efficiency of a plant growth-promoting strain of P. aeruginosa was severely impeded in its ΔcbrA and ΔcbrB derivatives compared to the parental strain (Sivakumar et al., 2021). Since the RpuS/R system of R. fredii was characterized for the first time in this work, no previous information of its involvement in nodulation is available. However, it was shown previously that in R. leguminosarum, a ΔmctP deletion mutant was still capable of forming nitrogen-fixing nodules on peas, although the authors mention no details on nodulation kinetics (Hosie et al., 2002). Overall, these observations suggest that the effects elicited by the deletion of SLC5/STAC TCSTS on host–microbe interactions depend significantly on each microorganism’s particular environmental conditions and adaptations. For S. fredii NGR234, a plausible explanation could be the dependence on plant exudates as a carbon source by the bacteria, of which pyruvate and acetate are relevant components (Koo et al., 2005). However, since S. fredii NGR234 has an extraordinary nodulation-host diversity, nodulating more than 112 genera of legumes (Pueppke and Broughton, 1999), it could be interesting to further characterize the role of the CrbS/R and RpuS/R systems in nodulation in other plant species.
Finally, the identification of the CrbS/R system in S. fredii and the confirmation of its participation in the regulation of acetate utilization highlight its importance in bacteria. Previously, when characterizing this system in P. fluorescens we failed to obtain a phenotype for the mutants on several of the genes it regulated, probably because our experimental conditions failed to reproduce the natural conditions during which their activity is important (Sepulveda and Lupas, 2017). Currently, we are characterizing the regulon of different homologs of the CrbS/R system, which also seem to be involved in the regulation of acetate utilization, in different families of bacteria with various lifestyles. We expect that this multi-organism comparative characterization will allow us to better understand not only the sensing and signaling mechanisms of the SLC5/STAC sensors, but also the metabolic role of the genes regulated by these systems.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below:https://www.ncbi.nlm.nih.gov/, SRR16684185https://www.ncbi.nlm.nih.gov/, SRR16684186https://www.ncbi.nlm.nih.gov/, SRR16684183https://www.ncbi.nlm.nih.gov/, SRR16684184.
AR, MA, GG, RH, MG, and ES constructed the plasmids and strains, performed the biochemical and microbiological assays, and analyzed the data. CC and SB performed the nodulation assays and analyzed the data. AS-F and CG-S performed the RNAseq assays. RC-R and AP analyzed the RNAseq data. AL and RC performed the bioinformatic analyses. ES, AR, RC, and AL contributed to conception and design of the study. ES, AL, and AP provided funding. ES wrote the first draft of the manuscript. RC-R, CG-S, AL, and AP wrote sections of the manuscript. All authors contributed to manuscript revision, read, and approved the submitted version.
This work was funded by the Max Planck Society through the Max Planck Partner Group in Signal Transduction and Agricultural Microbiology, by the Mexican Research Council (SEP-CONACYT project no. A1-S-7797), and by the Knut and Alice Wallenberg Foundation grant KAW 2015.0225. RNAseq analysis was performed on resources provided by SNIC through the Uppsala Multidisciplinary Center for Advanced Computational Science (UPPMAX, Uppsala University, Sweden), via projects SNIC 2017-7-258, uppstore2017093, and SNIC 2020/16-261 to Andrea Puhar.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb. 2022.871077/full#supplementary-material
Anders, S., Pyl, P. T., and Huber, W. (2014). HTSeq--a python framework to work with high throughput sequencing data. Bioinformatics 31, 166–169. doi: 10.1093/bioinformatics/btu638
Bailey, T. L., Boden, M., Buske, F. A., Frith, M., Grant, C. E., Clementi, L., et al. (2009). MEME SUITE: tools for motif discovery and searching. Nucleic Acids Res. 37, W202–W208. doi: 10.1093/nar/gkp335
Blighe, K. R., and Lewis, M. (2021). EnhancedVolcano: publication-ready volcano plots with enhanced colouring and labeling. R Package Version 1.12.0.
Bolger, A. M., Lohse, M., and Usadel, B. (2014). Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
Borland, S., Prigent-Combaret, C., and Wisniewski-Dyé, F. (2016). Bacterial hybrid histidine kinases in plant–bacteria interactions. Microbiology 162, 1715–1734. doi: 10.1099/mic.0.000370
Bravo, A., and Mora, J. (1988). Ammonium assimilation in rhizobium phaseoli by the glutamine synthetase-glutamate synthase pathway. J. Bacteriol. 170, 980–984. doi: 10.1128/jb.170.2.980-984.1988
Cervantes-Rivera, R., and Puhar, A. (2020). Whole-genome identification of transcriptional start sites by differential RNA-seq in bacteria. BIO-PROTOCOL 10:e3757. doi: 10.21769/bioprotoc.3757
Delorenzi, M., and Speed, T. (2002). An HMM model for coiled-coil domains and a comparison with PSSM-based predictions. Bioinformatics 18, 617–625. doi: 10.1093/bioinformatics/18.4
Dunn, M. F. (1998). Tricarboxylic acid cycle and anaplerotic enzymes in rhizobia. FEMS Microbiol. Rev. 22, 105–123. doi: 10.1111/j.1574-6976.1998.tb00363.x
Edgar, R. C. (2004). Muscle: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797. doi: 10.1093/nar/gkh340
Ewels, P., Magnusson, M., Lundin, S., and Käller, M. (2016). MultiQC: summarize analysis results for multiple tools and samples in a single report. Bioinformatics 32, 3047–3048. doi: 10.1093/bioinformatics/btw354
Ferri, L., Gori, A., Biondi, E. G., Mengoni, A., and Bazzicalupo, M. (2010). Plasmid electroporation of sinorhizobium strains: the role of the restriction gene HSDR in type strain RM1021. Plasmid 63, 128–135. doi: 10.1016/j.plasmid.2010.01.001
Gabler, F., Nam, S. Z., Till, S., Mirdita, M., Steinegger, M., Söding, J., et al. (2020). Protein sequence analysis using the MPI bioinformatics toolkit. Curr. Protoc. Bioinformatics 72:e108. doi: 10.1002/cpbi.108
Galperin, M. Y. (2010). Diversity of structure and function of response regulator output domains. Curr. Opin. Microbiol. 13, 150–159. doi: 10.1016/j.mib.2010.01.005
Gao, M., Benge, A., Wu, T.-J., and Javier, R. (2019). Use of plasmid pVMG to make transcriptional ß-glucuronidase reporter gene fusions in the rhizobium genome for monitoring the expression of rhizobial genes in vivo. Biol. Proced. Online 21:8. doi: 10.1186/s12575-019-0096-y
Gao, R., and Stock, A. M. (2009). Biological insights from structures of two-component proteins. Annu. Rev. Microbiol. 63, 133–154. doi: 10.1146/annurev.micro.091208.073214
Geddes, B. A., and Oresnik, I. J. (2014). Physiology, genetics, and biochemistry of carbon metabolism in the alphaproteobacteriumsinorhizobium meliloti. Can. J. Microbiol. 60, 491–507. doi: 10.1139/cjm-2014-0306
Girard, L., Brom, S., Dávalos, A., López, O., Soberón, M., and Romero, D. (2000). Differential regulation of fixN-reiterated genes in rhizobium Etli by a novel fixL—fixK cascade. Mol. Plant-Microbe Interact. 13, 1283–1292. doi: 10.1094/mpmi.2000.13.12.1283
González, V., Santamaría, R. I., Bustos, P., Pérez-Carrascal, O. M., Vinuesa, P., Juárez, S., et al. (2019). Phylogenomic rhizobium species are structured by a continuum of diversity and genomic clusters. Front. Microbiol. 10:910. doi: 10.3389/fmicb.2019.00910
Grant, C. E., Bailey, T. L., and Noble, W. S. (2011). FIMO: scanning for occurrences of a given motif. Bioinformatics 27, 1017–1018. doi: 10.1093/bioinformatics/btr064
Henriquez, T., and Jung, H. (2021). Involvement of the MXTR/ERDR (CrbS/CrbR) two-component system in acetate metabolism in pseudomonas Putida KT2440. Microorganisms 9:1558. doi: 10.3390/microorganisms9081558
Henriquez, T., Wirtz, L., Su, D., and Jung, H. (2021). Prokaryotic solute/sodium symporters: versatile functions and mechanisms of a transporter family. Int. J. Mol. Sci. 22:1880. doi: 10.3390/ijms22041880
Hildebrand, A., Remmert, M., Biegert, A., and Söding, J. (2009). Fast and accurate automatic structure prediction with HHpred. Proteins Struct. Funct. Bioinformatics 77, 128–132. doi: 10.1002/prot.22499
Hosie, A. H., Allaway, D., and Poole, P. S. (2002). A monocarboxylate permease of rhizobium Leguminosarum is the first member of a new subfamily of transporters. J. Bacteriol. 184, 5436–5448. doi: 10.1128/jb.184.19.5436-5448.2002
Hulko, M., Berndt, F., Gruber, M., Linder, J. U., Truffault, V., Schultz, A., et al. (2006). The HAMP domain structure implies helix rotation in transmembrane signaling. Cell 126, 929–940. doi: 10.1016/j.cell.2006.06.058
Imai, Y., Matsushima, Y., Sugimura, T., and Terada, M. (1991). A simple and rapid method for generating a deletion by PCR. Nucleic Acids Res. 19:2785. doi: 10.1093/nar/19.10.2785
Jacob, K., Rasmussen, A., Tyler, P., Servos, M. M., Sylla, M., Prado, C., et al. (2017). Regulation of acetyl-COA synthetase transcription by the CrbS/R two-component system is conserved in genetically diverse environmental pathogens. PLoS One 12:e0177825. doi: 10.1371/journal.pone.0177825
Jefferson, R. A., Burgess, S. M., and Hirsh, D. (1986). beta-Glucuronidase from Escherichia coli as a gene-fusion marker. Proc. Natl. Acad. Sci. U.S.A. 83, 8447–8451. doi: 10.1073/pnas.83.22.8447
Käll, L., Krogh, A., and Sonnhammer, E. L. (2004). A Combined transmembrane topology and signal peptide prediction method. J. Mol. Biol. 338, 1027–1036. doi: 10.1016/j.jmb.2004.03.016
Khan, S. R., Gaines, J., Roop, R. M., and Farrand, S. K. (2008). Broad-host-range expression vectors with tightly regulated promoters and their use to examine the influence of Trar and tram expression on Ti plasmid quorum sensing. Appl. Environ. Microbiol. 74, 5053–5062. doi: 10.1128/aem.01098-08
Koo, B. J., Adriano, D. C., Bolan, N. S., and Barton, C. D. (2005). Root exudates and microorganisms. Encycl. Soils Environ. 6, 421–428. doi: 10.1016/b0-12-348530-4/00461-6
Korycinski, M., Albrecht, R., Ursinus, A., Hartmann, M. D., Coles, M., Martin, J., et al. (2015). Stac—a new domain associated with transmembrane solute transport and two-component signal transduction systems. J. Mol. Biol. 427, 3327–3339. doi: 10.1016/j.jmb.2015.08.017
Kreth, J., Lengeler, J. W., and Jahreis, K. (2013). Characterization of pyruvate uptake in escherichia coli K-12. PLoS One 8:e67125. doi: 10.1371/journal.pone.0067125
Langmead, B., and Salzberg, S. L. (2012). Fast gapped-read alignment with bowtie 2. Nat. Methods 9, 357–359. doi: 10.1038/nmeth.1923
Lee, J., Shin, M.-K., Ryu, D.-K., Kim, S., and Ryu, W.-S. (2010). Insertion and deletion mutagenesis by overlap extension PCR. Methods Mol. Biol. 634, 137–146. doi: 10.1007/978-1-60761-652-8_10
Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N., et al. (2009). The sequence alignment/map format and SAMtools. Bioinformatics 25, 2078–2079. doi: 10.1093/bioinformatics/btp352
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with deseq2. Genome Biol. 15:550. doi: 10.1186/s13059-014-0550-8
Lupas, A., Van Dyke, M., and Stock, J. (1991). Predicting coiled coils from protein sequences. Science 252, 1162–1164. doi: 10.1126/science.252.5009.1162
Martinez-Guerrero, C. E., Ciria, R., Abreu-Goodger, C., Moreno-Hagelsieb, G., and Merino, E. (2008). GeConT 2: gene context analysis for orthologous proteins, conserved domains and metabolic pathways. Nucleic Acids Res. 36, W176–W180. doi: 10.1093/nar/gkn330
Mirabella, A., Yañez Villanueva, R.-M., Delrue, R.-M., Uzureau, S., Zygmunt, M. S., Cloeckaert, A., et al. (2012). The two-component system prls/PRLR of brucella melitensis is required for persistence in mice and appears to respond to ionic strength. Microbiology 158, 2642–2651. doi: 10.1099/mic.0.060863-0
Monteagudo-Cascales, E., García-Mauriño, S. M., Santero, E., and Canosa, I. (2019). Unraveling the role of the CBRA histidine kinase in the signal transduction of the CBRAB two-component system in pseudomonas putida. Sci. Rep. 9:9110. doi: 10.1038/s41598-019-45554-9
Muzhingi, I., Prado, C., Sylla, M., Diehl, F. F., Nguyen, D. K., Servos, M. M., et al. (2018). Modulation of CrbS-dependent activation of the acetate switch in vibrio cholerae. J. Bacteriol. 200:e00380-18. doi: 10.1128/jb.00380-18
Ormeño-Orrillo, E., and Martínez-Romero, E. (2019). A genomotaxonomy view of the bradyrhizobium genus. Front. Microbiol. 10:1334. doi: 10.3389/fmicb.2019.01334
Pueppke, S. G., and Broughton, W. J. (1999). Rhizobium sp. strain NGR234 and R. Fredii USDA257 share exceptionally broad, nested host ranges. Mol. Plant-Microbe Interact. 12, 293–318. doi: 10.1094/mpmi.1999.12.4.293
Quiroz-Rocha, E., Moreno, R., Hernández-Ortíz, A., Fragoso-Jiménez, J. C., Muriel-Millán, L. F., Guzmán, J., et al. (2017). Glucose uptake in Azotobacter vinelandii occurs through a glup transporter that is under the control of the CBRA/CBRB and Hfq-CRC systems. Sci. Rep. 7:858. doi: 10.1038/s41598-017-00980-5
R Core Team (2021). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria.
Rodríguez, S., Correa-Galeote, D., Sánchez-Pérez, M., Ramírez, M., Isidra-Arellano, M. C., del Reyero-Saavedra, M. R., et al. (2020). A novel OMPR-type response regulator controls multiple stages of the rhizobium etli – phaseolus vulgaris N2-fixing symbiosis. Front. Microbiol. 11:3250. doi: 10.3389/fmicb.2020.615775
Sambrook, J., Fritsch, E. F., and Maniatis, T. (2012). Molecular Cloning: A Laboratory Manual. Cold Spring Harbor: Cold Spring Harbor Laboratory Press.
Schäfer, A., Tauch, A., Jäger, W., Kalinowski, J., Thierbach, G., and Pühler, A. (1994). Small mobilizable multi-purpose cloning vectors derived from the escherichia coli plasmids PK18 and PK19: selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 145, 69–73. doi: 10.1016/0378-1119(94)90324-7
Schmeisser, C., Liesegang, H., Krysciak, D., Bakkou, N., Antoine, L. Q., Wollherr, A., et al. (2009). Rrhizobium SP. strain NGR234 possesses a remarkable number of secretion systems. Appl. Environ. Microbiol. 75, 4035–4045. doi: 10.1128/aem.00515-09
Schuldes, J., Rodriguez Orbegoso, M., Schmeisser, C., Krishnan, H. B., Daniel, R., and Streit, W. R. (2012). Complete genome sequence of the broad-host-range strain sinorhizobium fredii USDA257. J. Bacteriol. 194:4483. doi: 10.1128/jb.00966-12
Sepulveda, E., and Lupas, A. N. (2017). Characterization of the CrbS/R two-component system in pseudomonas fluorescens reveals a new set of genes under its control and a DNA motif required for CrbR-mediated transcriptional activation. Front. Microbiol. 8:2287. doi: 10.3389/fmicb.2017.02287
Sivakumar, R., Gunasekaran, P., and Rajendhran, J. (2021). Inactivation of CbrAB two-component system hampers root colonization in rhizospheric strain of pseudomonas aeruginosa PGPR2. Biochimica et Biophysica Acta (BBA) – Gene Regul. Mech. 1864:194763. doi: 10.1016/j.bbagrm.2021.194763
Sonnhammer, E. L., von Heijne, G., and Krogh, A. (1998). A hidden Markov model for predicting transmembrane helices in protein sequences. Proc. Int. Conf. Intell. Syst. Mol. Biol. 6, 175–182.
Stanley, J., Dowling, D. N., and Broughton, W. J. (1988). Cloning of Hema from rhizobium sp. NGR234 and symbiotic phenotype of a gene-directed mutant in diverse legume genera. Mol. Gen. Genet. MGG 215, 32–37. doi: 10.1007/bf00331299
Stock, A. M., Robinson, V. L., and Goudreau, P. N. (2000). Two-component signal transduction. Annu. Rev. Biochem. 69, 183–215. doi: 10.1146/annurev.biochem.69.1.183
Streit, W. R., Schmitz, R. A., Perret, X., Staehelin, C., Deakin, W. J., Raasch, C., et al. (2004). An evolutionary hot spot: the PNGR234 B replicon of rhizobium sp. strain NGR234. J. Bacteriol. 186, 535–542. doi: 10.1128/jb.186.2.535-542.2004
Tiwari, S., Jamal, S. B., Hassan, S. S., Carvalho, P. V., Almeida, S., Barh, D., et al. (2017). Two-component signal transduction systems of pathogenic bacteria as targets for antimicrobial therapy: an overview. Front. Microbiol. 8:1878. doi: 10.3389/fmicb.2017.01878
Wang, X., Liu, D., Luo, Y., Zhao, L., Liu, Z., Chou, M., et al. (2018). Comparative analysis of rhizobial chromosomes and plasmids to estimate their evolutionary relationships. Plasmid 96–97, 13–24. doi: 10.1016/j.plasmid.2018.03.001
Wickham, H., and Sievert, C. (2016). GGPLOT2: Elegant Graphics for Data Analysis. Cham, Schweiz: Springer.
Wingett, S. W., and Andrews, S. (2018). FastQ Screen: a tool for multi-genome mapping and quality control. F1000Research 7:1338. doi: 10.12688/f1000research.15931.2
Wirtz, L., Eder, M., Schipper, K., Rohrer, S., and Jung, H. (2020). Transport and kinase activities of CBRA of pseudomonas Putida KT2440. Sci. Rep. 10:5400. doi: 10.1038/s41598-020-62337-9
Zhang, X.-X., Gauntlett, J. C., Oldenburg, D. G., Cook, G. M., and Rainey, P. B. (2015). Role of the transporter-like sensor kinase CBRA in histidine uptake and signal transduction. J. Bacteriol. 197, 2867–2878. doi: 10.1128/jb.00361-15
Zhang, X.-X., and Rainey, P. B. (2008). Dual involvement of CBRAB and NTRBC in the regulation of histidine utilization in pseudomonas fluorescens SBW25. Genetics 178, 185–195. doi: 10.1534/genetics.107.081984
Keywords: TCSTSs, two-component signal transduction systems, piruvate, STAC, Sinorhizobium fredii
Citation: Ramos AL, Aquino M, García G, Gaspar M, de la Cruz C, Saavedra-Flores A, Brom S, Cervantes-Rivera R, Galindo-Sánchez CE, Hernandez R, Puhar A, Lupas AN and Sepulveda E (2022) RpuS/R Is a Novel Two-Component Signal Transduction System That Regulates the Expression of the Pyruvate Symporter MctP in Sinorhizobium fredii NGR234. Front. Microbiol. 13:871077. doi: 10.3389/fmicb.2022.871077
Edited by:
Marie-Joelle Virolle, Centre National de la Recherche Scientifique (CNRS), FranceReviewed by:
MIguel Castañeda, Instituto de Ciencias Benemérita Universidad Autónoma de Puebla, MexicoCopyright © 2022 Ramos, Aquino, García, Gaspar, de la Cruz, Saavedra-Flores, Brom, Cervantes-Rivera, Galindo-Sánchez, Hernandez, Puhar, Lupas and Sepulveda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Edgardo Sepulveda, ZXNlcHVsdmVkYUBjaWNlc2UubXg=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.