 Yang Wang1,2,3
Yang Wang1,2,3 Li-Ying Wang1,3
Li-Ying Wang1,3 Dan Dai4
Dan Dai4 Zheng-Xiang Qi3Zhen-Hao Zhang3Ya-Jie Liu3
Zheng-Xiang Qi3Zhen-Hao Zhang3Ya-Jie Liu3 Jia-Jun Hu3Peng Zhang5
Jia-Jun Hu3Peng Zhang5 Yu Li1,2,3*Bo Zhang1,3*
Yu Li1,2,3*Bo Zhang1,3*- 1Joint International Research Laboratory of Modern Agricultural Technology, Ministry of Education, Jilin Agricultural University, Changchun, China
- 2College of Plant Protection, Shenyang Agricultural University, Shenyang, China
- 3Engineering Research Center of Chinese Ministry of Education for Edible and Medicinal Fungi, College of Plant Protection, Jilin Agricultural University, Changchun, China
- 4Institute of Agricultural Applied Microbiology, Jiangxi Academy of Agricultural Sciences, Nanchang, China
- 5Mudanjiang Sub-Academy, Heilongjiang Academy of Agricultural Sciences, Mudanjiang, Heilongjiang, China
Boletaceae, the largest family in Boletales, has been attracted by mycologists in the world due to its diverse morphology and complex history of evolution. Although considerable work has been done in the past decades, novel taxa are continually described. The current study aimed to introduce three new taxa and one new record of Boletaceae from China. The morphological descriptions, color photographs, phylogenetic trees to show the positions of the taxa, and comparisons with allied taxa are provided. The new genus Hemilanmaoa is unique in the Pulveroboletus group, and Hemilanmaoa retistipitatus was introduced as the type species. It can be distinguished by its bluing basidioma when injured, a decurrent hymenophore, a stipe covered with distinct reticulations, and a fertile stipitipellis. Porphyrellus pseudocyaneotinctus is characterized by its pileipellis consisting of broadly concatenated cells and thin-walled caulocystidia in Porphyrellus. In Phylloporus, Phylloporus biyangensis can be distinguished by its hymenophores that change to blue when injured and yellow basal mycelium. Lanmaoa angustispora, as a new record, is first reported in Northern China. Internal transcribed spacer (ITS), 28S rDNA (28S), translation elongation factor 1-alpha (tef1-α), RNA polymerase II subunit 1 (rpb1), and RNA polymerase II subunit 2 (rpb2) were employed to execute phylogenetic analyses.
Introduction
The symbiotic systems between fungi and plants were recognized to accelerate the process of vascular plants invading land (Treub, 1884; Pirozynski and Malloch, 1975; Malloch et al., 1980). To date, there are four different mycorrhizal association types, viz. vesicular-arbuscular mycorrhizas (VAM), ectomycorrhizas (ECM), orchid mycorrhizas, and ectendo-, arbutoid, and monotropoid associations (Brundrett et al., 1996). The ability that ectomycorrhizal fungi symbiosis with the host can enhance the utilization efficiency of soil nutrition in plants and their ability to resist pests, which consequently improves the survival rates of plants in the ecosystem (Marx, 1972; Trappe, 1977). Meanwhile, competing with saprotrophic fungi in carbon recycling and other nutrients can dramatically influence the balance of forest ecosystems (Tedersoo et al., 2010; Wu et al., 2022). As a species-rich family in the order Boletales E.-J. Gilbert, a majority of species in Boletaceae Chevall. are ectomycorrhizal fungi. Mushrooms in the Boletaceae are characterized by their large, fleshy, and usually brilliantly colored basidioma and hymenophore that are tubulose, lamellate, or loculus.
Over the last few decades, the rapid development of molecular techniques has immensely improved the resolution of the fungal tree of life (He et al., 2019; Naranjo-Ortiz and Gabaldón, 2019; Li et al., 2021). The modern phylogenetic analyses are essential to reassess and resolve the traditional taxonomy of Boletaceae (Skrede et al., 2011; Yang, 2011; Halling et al., 2012b,2015; Nuhn et al., 2013; Arora and Frank, 2014; Wu et al., 2014; Orihara et al., 2016; Kuo and Ortiz-Santana, 2020). In the course of DNA-based research, many new genera were proposed within Boletaceae, and intra-relationships of many complex groups tend to be clarified (Yang et al., 2003; Dentinger et al., 2010; Halling et al., 2012b,2015; Arora and Frank, 2014; Wu et al., 2014, 2021; Cui et al., 2016; Orihara et al., 2016; Frank et al., 2020; Li and Yang, 2021; Biketova et al., 2022). Based on materials collected from China, Chinese mycologists have made huge contributions to the modern taxonomical system of Boletaceae—especially some fantastic studies were done by Wu et al. (2014) and Wu et al., 2016a,b. In 2014, Wu et al. (2014) redefined seven major clades within Boletaceae, viz. Austroboletoideae, Boletoideae, Chalciporoideae, Leccinoideae, Xerocomoideae, Zangioideae, and the Pulveroboletus group. This result was widely recognized by other researchers. Lately, Wu et al. unveiled evolutionary innovations from the genomic respects in the ectomycorrhizal Boletales (Wu et al., 2022). The conclusion similar to Kohler et al. (2015) and Miyauchi et al. (2020) showed that Boletales impressively reduced their plant cell wall-degrading enzymes (PCWDEs). However, a phenomenon that different lineages still retained different set of PCWDEs maybe means moderately ability of cell wall-degrading still existed in Boletales.
Although remarkable results on the taxonomy of boletes were obtained in the past (Li et al., 2011; Zeng et al., 2014; Zhao et al., 2014a,b; Gelardi et al., 2015a,b; Zhu et al., 2015; Wu et al., 2016b; Chen et al., 2019; Vadthanarat et al., 2019; Hosen and Yang, 2021; Ayala-Vásquez et al., 2022; Badou et al., 2022; Biketova et al., 2022), the additions of more new taxa are necessary to reconstruct a high-resolution tree of Boletaceae. More intensive collection are needed to analyze the species diversity of Boletaceae in China. Our study is focused on results from our collection in northern China. In this study, we described the morphological and phylogenetic data of a new genus that is evident to form a distinct lineage in the “Pulveroboletus group.” Meanwhile, two new species and one new record species of Henan province are also reported.
Materials and methods
Samplings and morphological analyses
Specimens were collected from Guizhou and Henan Province, China. Voucher materials were deposited in the Mycology Herbarium of the Jilin Agriculture University (HMJAU). The color of fresh basidiocarps is described following Kornerup and Wanscher’s (1978) method. Tissues of specimens were mounted in 5% KOH and then in 1% Congo Red or Melzer’s solution, and steps of amyloid reactions were followed according to Imler’s procedure (Imler, 1950; Biketova et al., 2022). The observations of microscopic characteristics were performed by Carl Zeiss Lab. A1 optical microscope. The ultrastructure of basidiospores was observed by scanning electron microscope (SEM). Basidiospore dimensions were recorded as length by width, in order of the minimum, the average, and the maximum; the notation (n/m/p) indicated that measurements were made on “p” randomly selected basidiospores from “m” basidiomes of “n” collections. Q is the ratio of length divided by width: Qm = average quotient (length/width ratio) ± standard deviation.
DNA extraction, PCR amplification, and sequencing
The NuClean Plant Genomic DNA kits (CWBIO) are used to extract genomic DNA. The primers LROR/LR5 were used for 28S, RPB1-B-F/RPB1-B-R for rpb1 (Wu et al., 2014), RPB2-B-F1/RPB2-B-R and PRB2-6F/PRB2-7.1R for rpb2 (Matheny and Ammirati, 2003; Matheny, 2005; Wu et al., 2014), and 983F/1567R for tef1-α (Rehner and Buckley, 2005). The polymerase chain reaction (PCR) procedures were executed, referring to Feng et al. (2012) and Wu et al. (2014).
Phylogenetic analyses
The new sequences were uploaded to NCBI,1 and other sequences were downloaded from NCBI (Supplementary Table 1). The raw matrixes (ITS, 28S, rpb1, rpb2, and tef1-α) were spliced in SeqMan (Swindell and Plasterer, 1997) and aligned with MAFFT (Katoh and Standley, 2013) using ‘E-INS-i (accurate)’ strategy and normal alignment mode, respectively. The ITS matrix of Phylloporus was aligned by MEGA 7 with the “Muscle” strategy (Kumar et al., 2016). TrimAL v1.2 was used to trim matrixes with the “gappyout” option (Capella-Gutiérrez et al., 2009). Multi-locus datasets were concatenated by PhyloSuite v1.2.1, and phylogenetic trees were constructed by maximum likelihood (ML) and Bayesian inference (BI) analyses (Zhang et al., 2020). In the multi-locus dataset (28S + rpb1 + rpb2 + tef1) of Hemilanmaoa, 646 bp for 28S, 761 bp for rpb1, 705 bp for rpb2, and 621bp for tef1. In the four-locus dataset of Porphyrellus E.-J. Gilbert and Lanmaoa G. Wu & Zhu L. Yang, 835 bp for 28S, 611 bp for rpb1, 642 bp for rpb2, and 620 bp for tef1. In the three-locus dataset (ITS + 28S + tef1) of Phylloporus Quél. 919 bp for 28S, 586 bp for tef1, and 996 bp for ITS. Best models of matrixes were searched using PartitionFinder 2 integrated into PhyloSuite v1.2.1 (Lanfear et al., 2017; Zhang et al., 2020). Models employed for each locus of Hemilanmaoa were GTR + I + G for 28S and rpb1, SYM+ I + G for rpb2 and tef1, for the locus of Porphyrellus and Lanmaoa, GTR + I + G for 28S, SYM + G for rpb1, SYM + I + G for tef1 and rpb2, and the locus of Phylloporus, GTR + I + G for 28S and ITS, and K80 + I + G for tef1. ML analyses were executed by IQ-tree (Nguyen et al., 2015) using ultrafast bootstrap with 5,000 replicates. BI analyses were executed using MrBayes 3.2.6 (Ronquist et al., 2012), running in 2,000,000 generations, and sampled every 2,000 generations. The initial 25% of the sampled data were discarded as burn-in. Other parameters were kept at default settings.
Results
Molecular phylogeny
In the phylogenetic relationship of Boletaceae, the Bayesian tree (Figure 1) and ML tree (Figure 2) were listed, respectively, due to some differences between the major clades of the Bayesian tree and ML tree. The multi-locus datasets consisted of 203 taxa and 2,733 nucleotide sites (Figures 1, 2). Gyrodon sp. and Paxillus obscurosporus C. Hahn were chosen as outgroups. The molecular phylogenetic analyses showed that Hemilanmaoa retistipitatus is not only embedded in the “Pulveroboletus Group” clade but also sister with Suillellus Murrill. It formed an independent lineage with a strong support value (BP = 0.94, PP = 94).
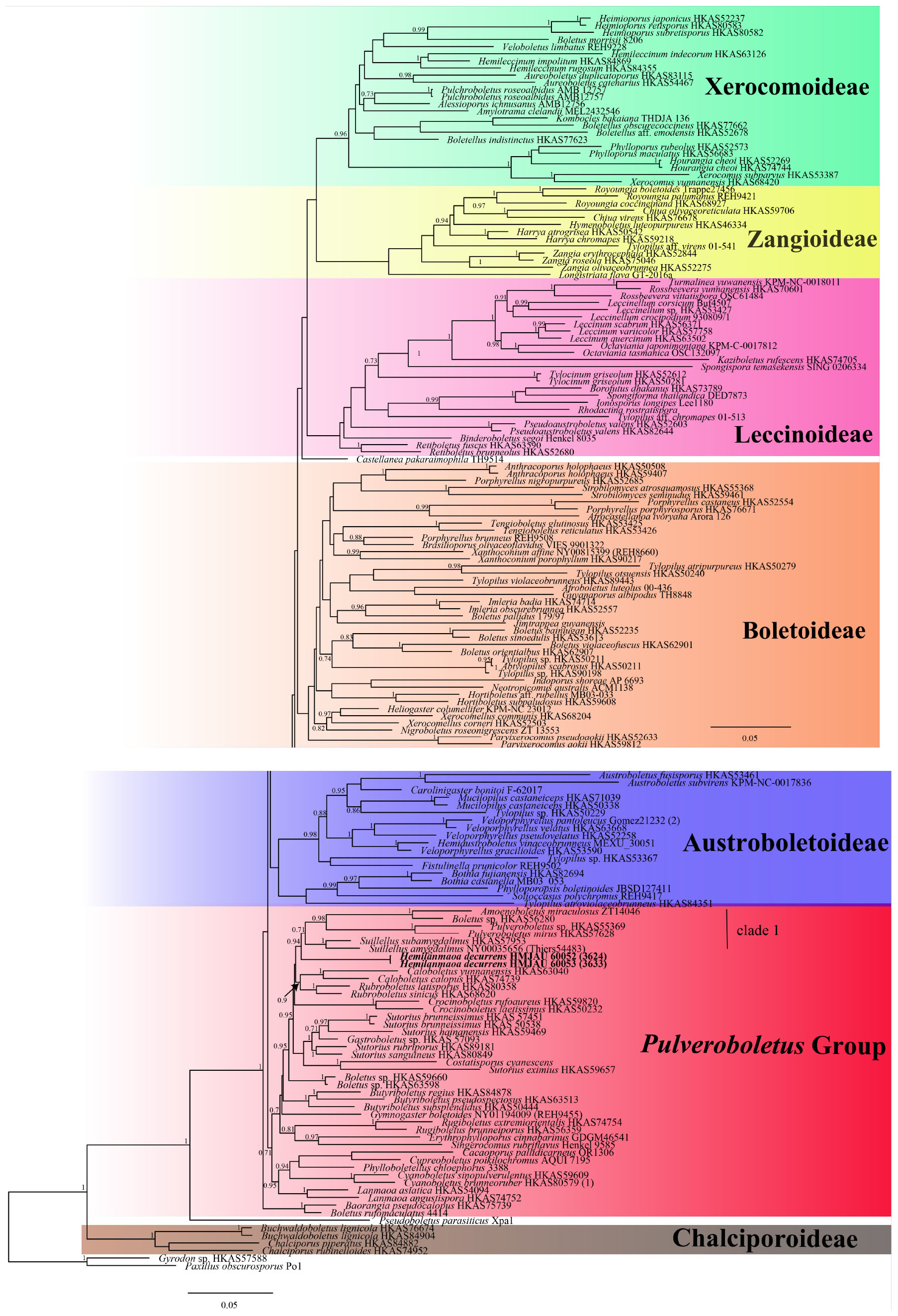
Figure 1. Phylogenetic relationships of Hemilanmaoa in Boletaceae inferred from multi-locus (28S, tef1-α, rpb1, and rpb2) using Bayesian inference. PP ≥ 0.7 are indicated in the phylogram. Newly formed sequences in this study are indicated in bold.
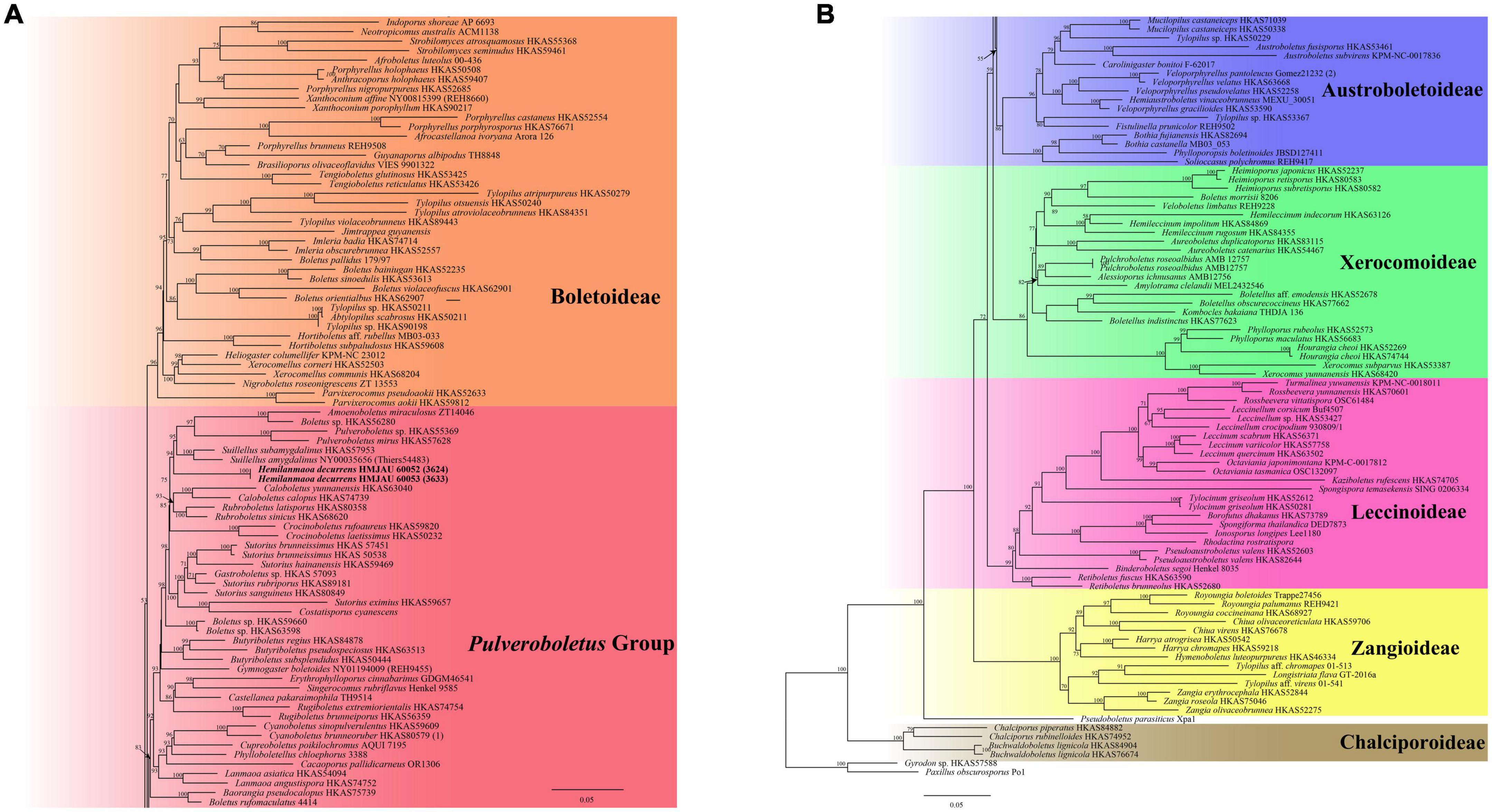
Figure 2. Phylogenetic relationships of Hemilanmaoa in Boletaceae inferred from multi-locus (28S, tef1-α, rpb1, and rpb2) using the maximum likelihood method. BP ≥ 50 is indicated in the phylogram. Newly formed sequences in this study are indicated in bold. (A) Phylogram upper part. (B) Phylogram lower part.
The four-locus datasets of Porphyrellus and Lanmaoa consisted of 89 taxa and 2,709 nucleotide sites. Butyriboletus appendiculatus (Schaeff.) D. Arora & J. L. Frank and Bu. autumniregius D. Arora & J. L. Frank were chosen as outgroups. In the phylogram (Figure 9), our sequences W3013, W3029, and W3022 were clustered together with Lanmaoa angustispora G. Wu & Zhu L. Yang (BP = 1, PP = 100) and formed an independent lineage. In the Porphyrellus, our sequences, namely w3088, w3085, w3054, w3046, w3062, w3019, w3039, and w3091, were clustered together with one previously described Po. cyaneotinctus (A.H. Sm. & Thiers) Singer (BP = 1, PP = 100).

Figure 3. Basidiomata of boletes (A,B,D) Hemilanmaoa retistipitatus, (C,F–H) Porphyrellus pseudocyaneotinctus, (E,I,J) Phylloporus biyangensis, and (K) Lanmaoa angustispora.
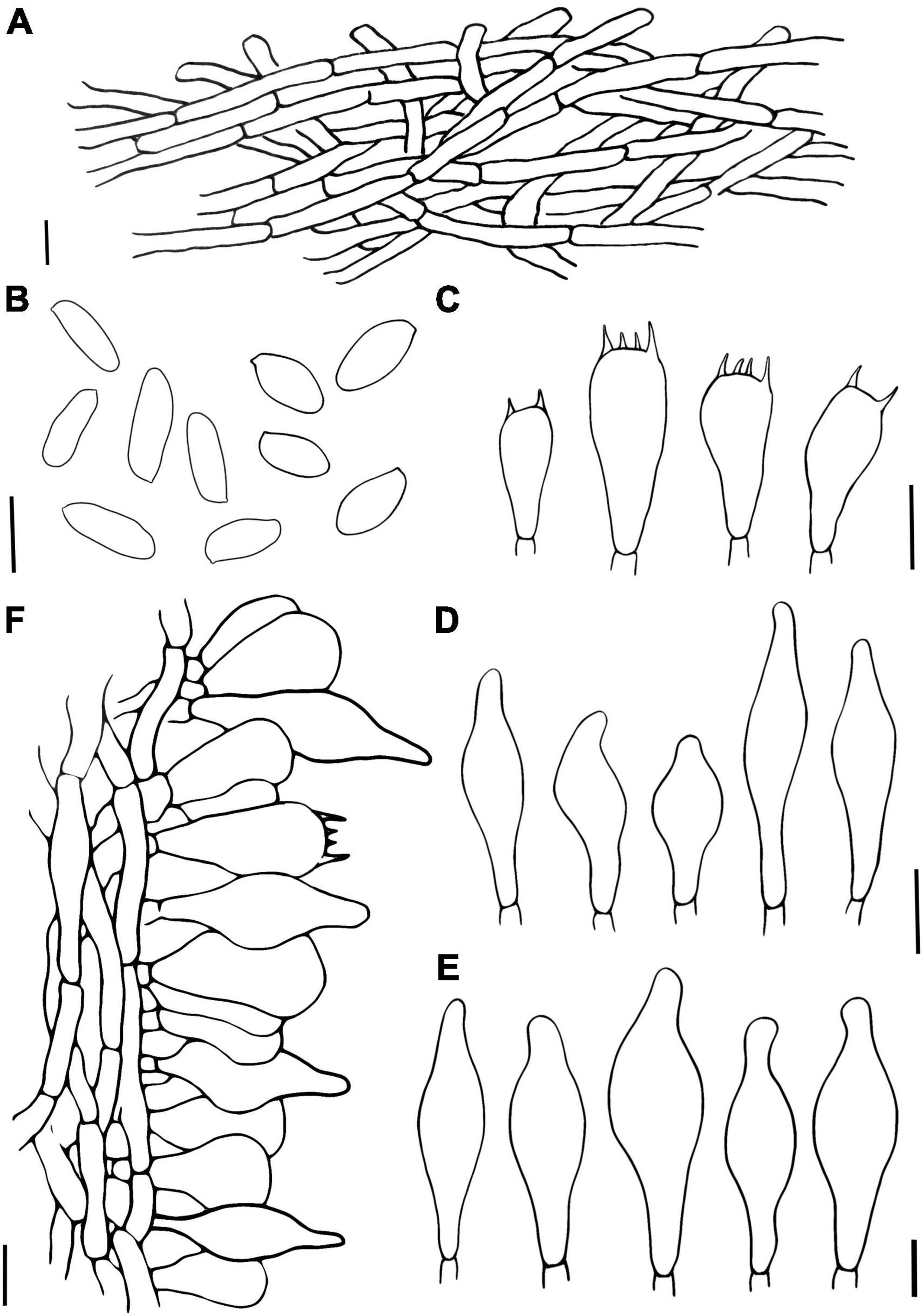
Figure 4. Hemilanmaoa retistipitatus (A) Pileipellis, (B) Basidiospores, (C) Basidia, (D) Cheilocystidia, (E) Pleurocystidia, and (F) Stipitipellis. Scale bars = 10 μm.

Figure 5. Characteristics of basidiospore in SEM. (A–C) Hemilanmaoa retistipitatus, (D,E) Porphyrellus pseudocyaneotinctus, (F,G) Phylloporus biyangensis, and (H) Lanmaoa angustispora.
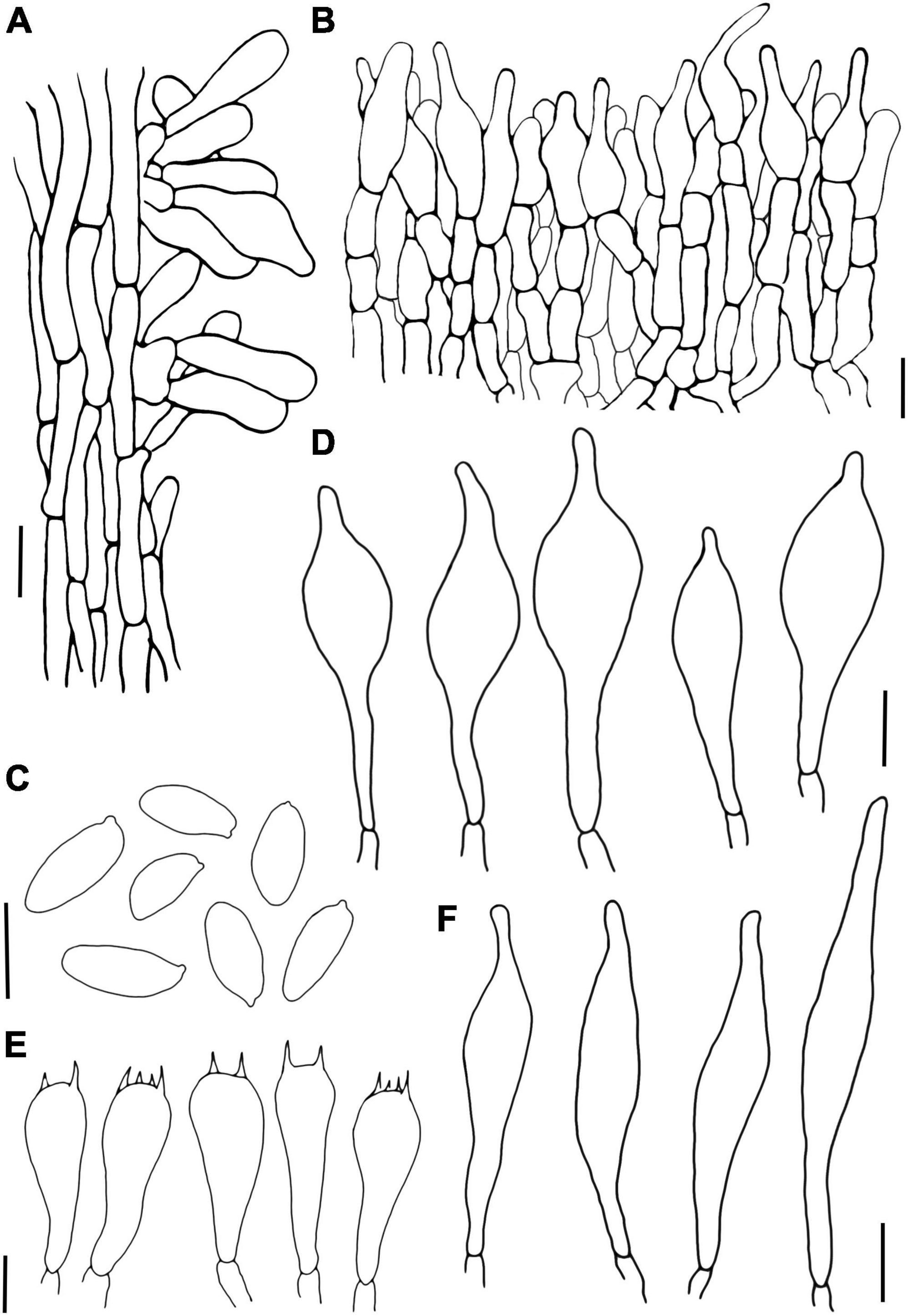
Figure 6. Porphyrellus pseudocyaneotinctus (A) Stipitipellis, (B) Pileipellis, (C) Basidiospores, (D) Cheilocystidia, (E) Basidia, and (F) Pleurocystidia. Scale bars = 10 μm.
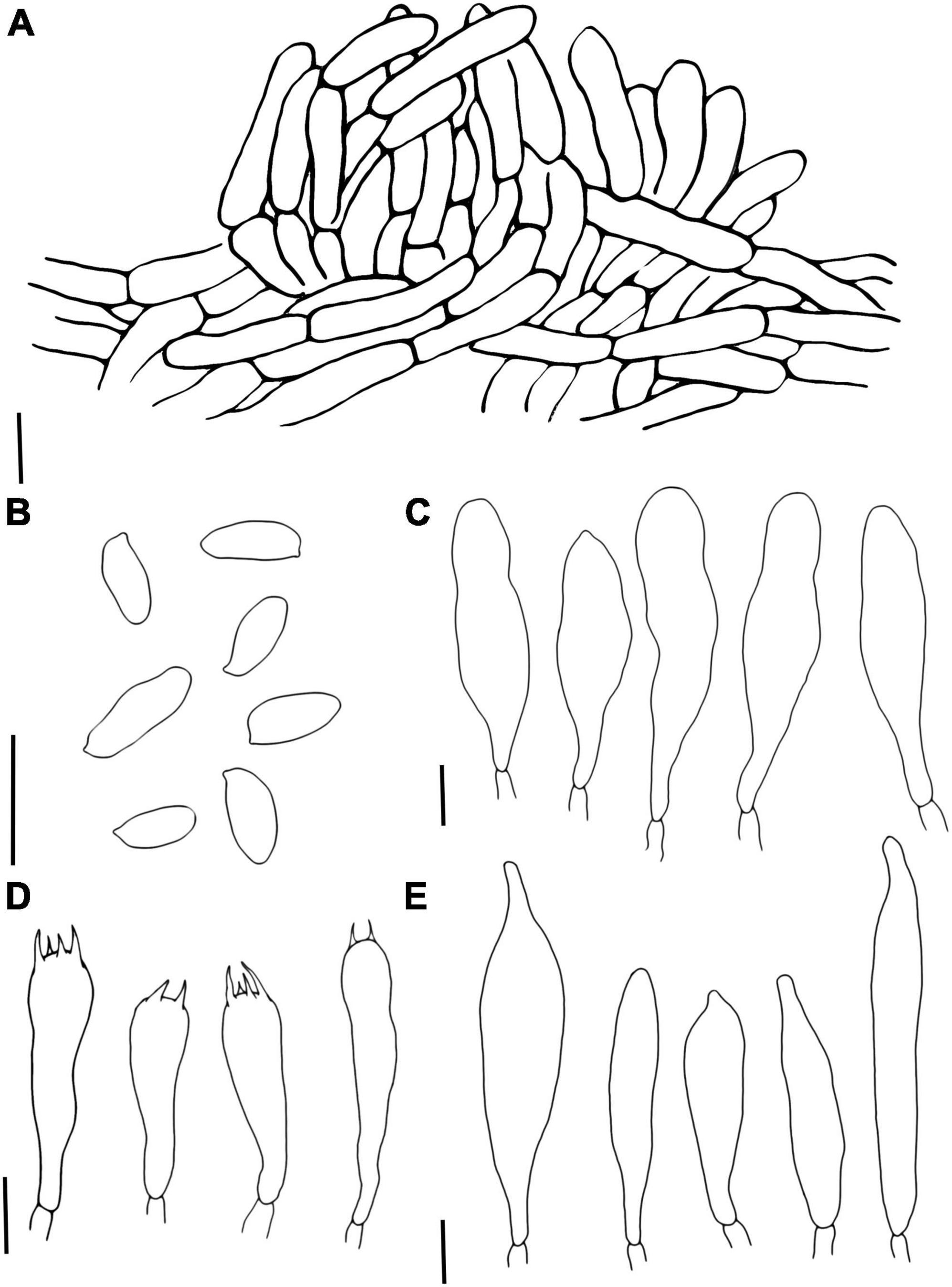
Figure 7. Phylloporus biyangensis (A) Pileipellis, (B) Basidiospores, (C) Cheilocystidia, (D) Basidia, and (E) Pleurocystidia. Scale bars = 10 μm.
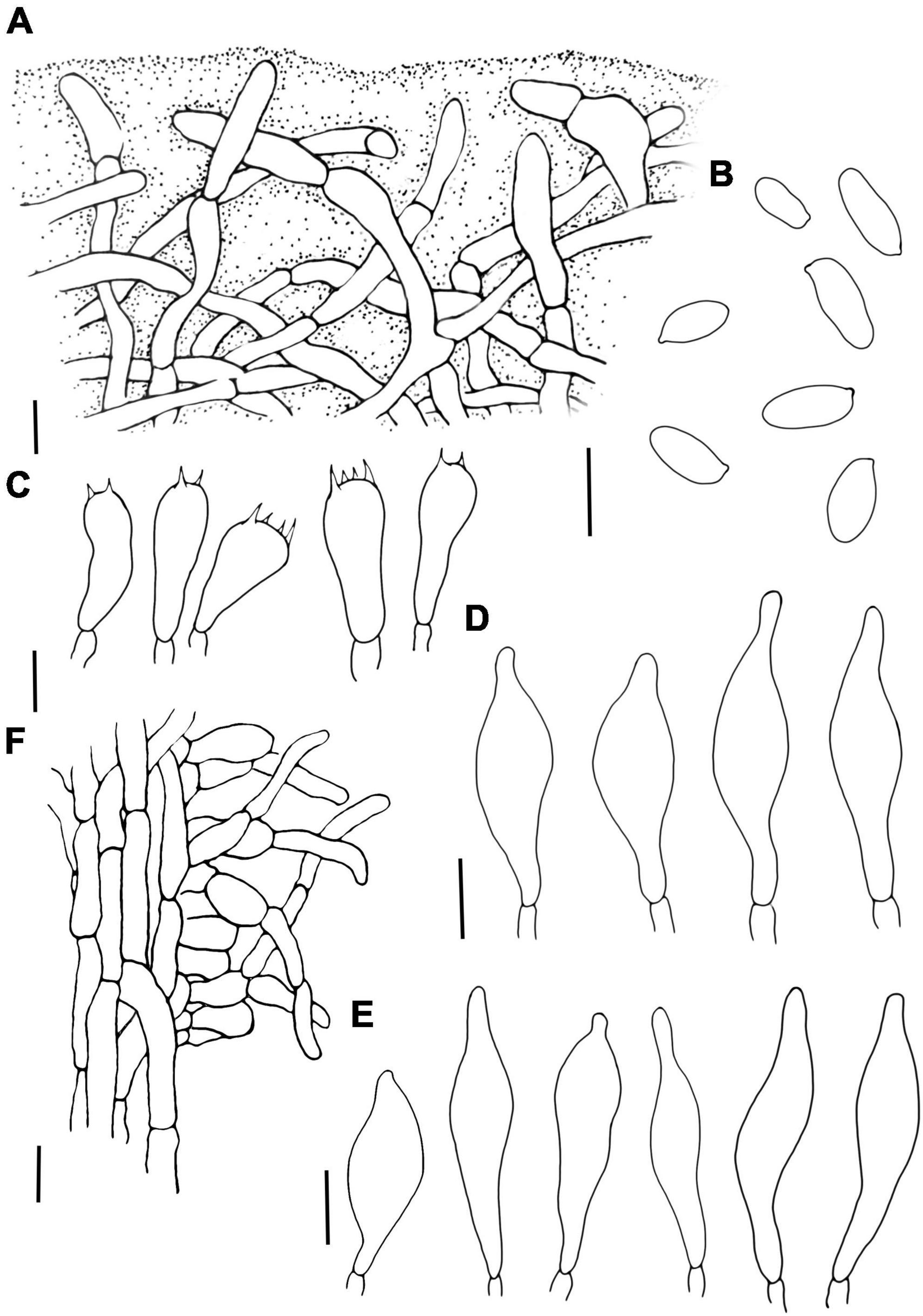
Figure 8. Lanmaoa angustispora (A) Pileipellis, (B) Basidiospores, (C) Basidia, (D) Cheilocystidia, (E) Pleurocystidia, and (F) Stipitipellis. Scale bars = 10 μm.
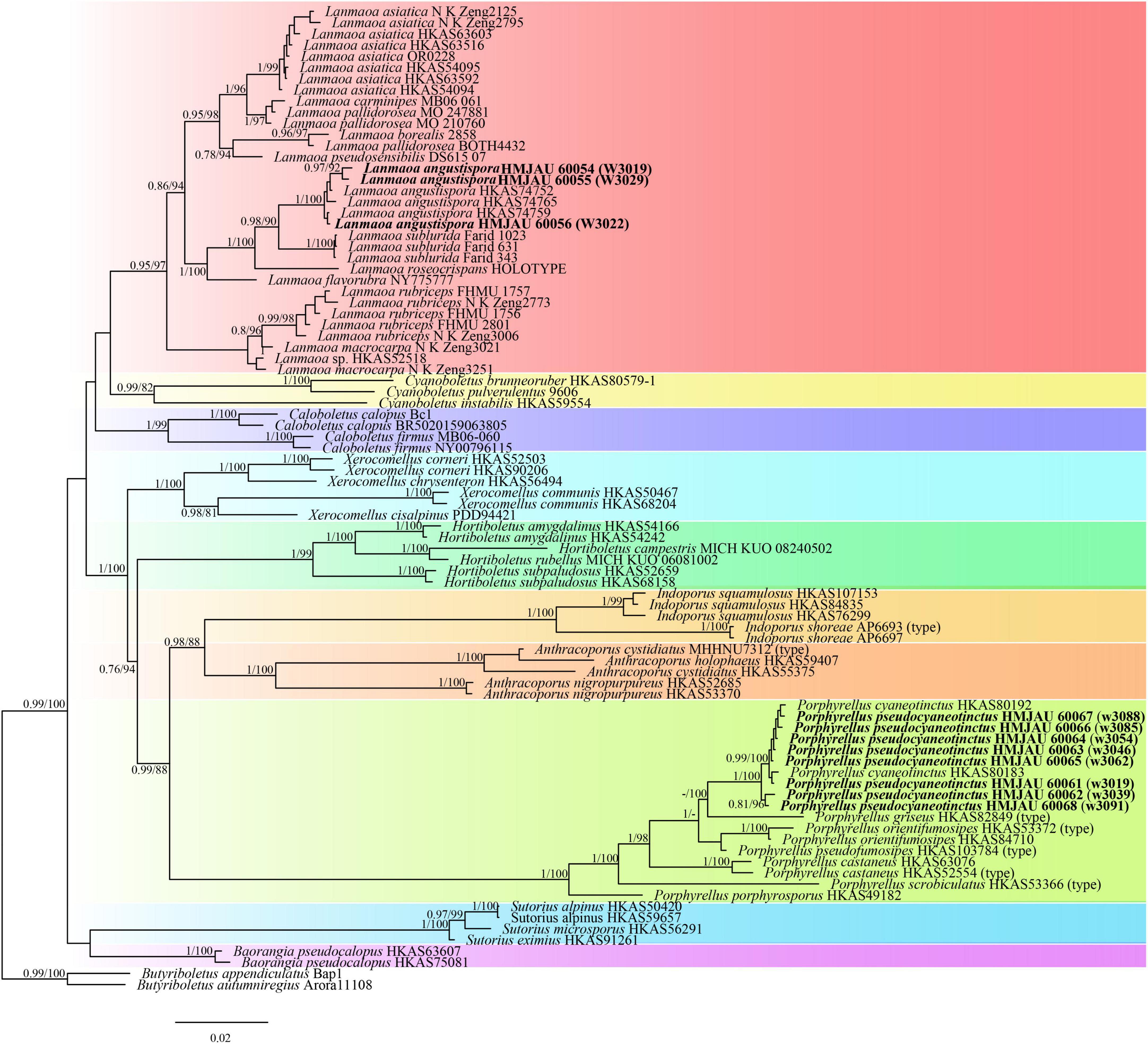
Figure 9. Phylogenetic relationships of Porphyrellus and Lanmaoa in their genera inferred from multi-locus (28S, tef1-α, rpb1, and rpb2) using the maximum likelihood method and Bayesian inference (only the BI tree was shown). BP ≥ 0.8 and PP ≥ 0.7 are indicated in the phylogram. Newly formed sequences in this study are indicated in bold.
The three-locus dataset (28S + ITS + tef1) of Phylloporus consisted of 170 taxa and 2,501 nucleotide sites. Hourangia cheoi (W.F. Chiu) (Xue T. Zhu & Zhu L. Yang) was selected as outgroups. In the phylogram (Figure 10), our sequences w3047, w3048, w3049a, and w3049b formed an independent clade with a high support value (BP = 100, PP = 1).
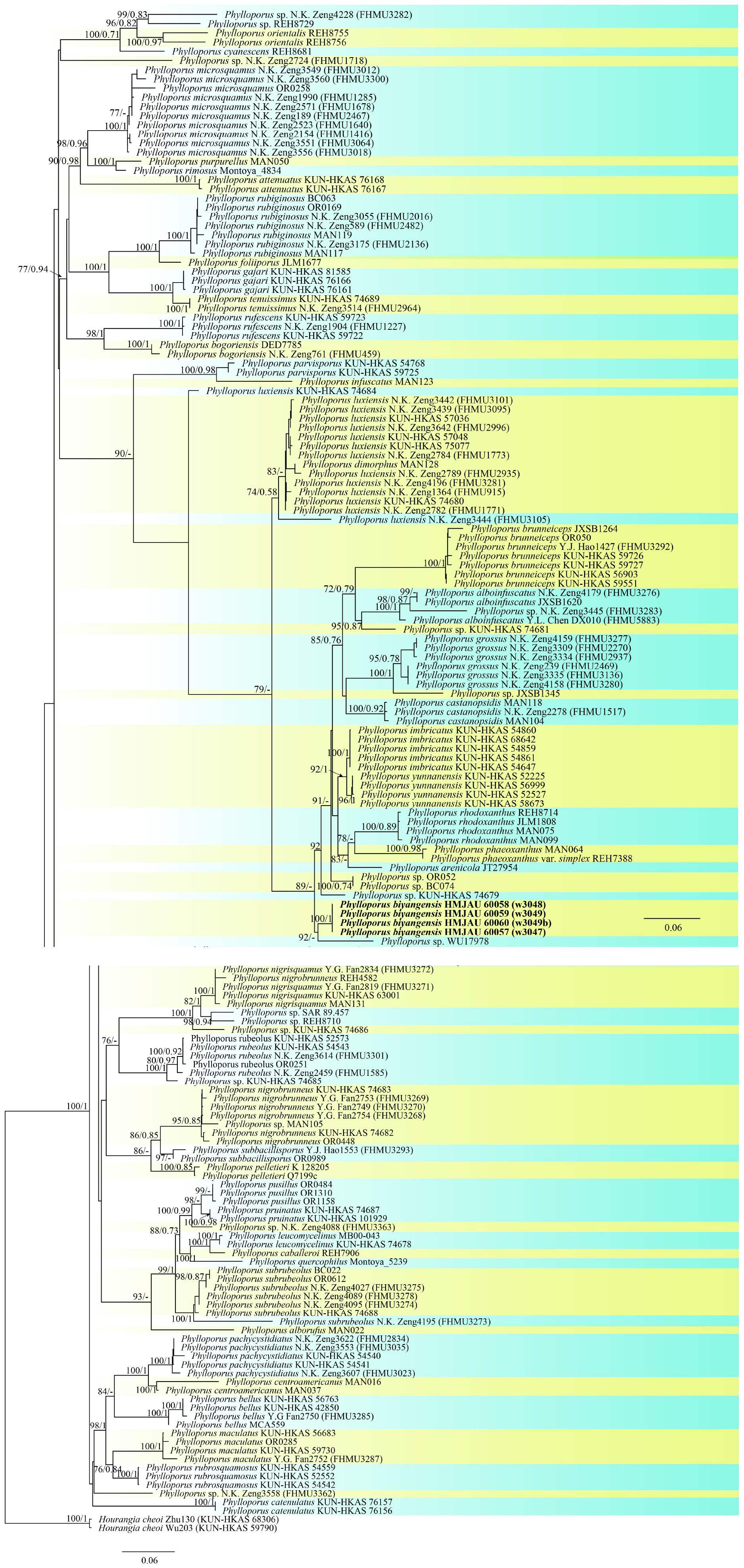
Figure 10. Phylogenetic relationships of Phylloporus in the genus inferred from multi-locus (ITS, 28S, and tef1-α) using maximum likelihood method and Bayesian inference (only the ML tree was shown). BP ≥ 0.7 and PP ≥ 0.7 are indicated in the phylogram. Newly formed sequences in this study are indicated in bold.
Taxonomy
Hemilanmaoa Yang Wang, Bo Zhang & Y. Li gen. nov.
Mycobank No.: MB845571
Etymology. “Hemi” refers to its morphological similarity to Lanmaoa.
Diagnosis. This genus is similar to Lanmaoa but differs from the latter by pores red at the mature, stipe covered with distinctly reticulations and hyphae dextrinoid. Basidioma bluing when bruising, pileus subtomentose, hymenophore decurrent with surface red, stipe covered with reticulations and red dots, and hyphae of context dextrinoid.
Basidioma stipitate-pileate with tubular hymenophore. Pileus hemispherical and depression at the center, subtomentose, dry, margin shortly appendiculate, grayish red to pastel red in the center, pale yellow toward margin; context whitish to pale yellow, discoloring to blue when injured. Hymenophore decurrent, surface orange-red, turning to blue when bruised; pores compound, angular to round, tubes light yellow, changing to blue when cut. Stipe central, yellow at the upper partition, brownish red downwards base, surface reticulate, especially on the upper partition, and erratically covered with brownish red dotted elements, staining blue when touched. Context of stipe brownish red at the base, changing to blue when injured. Basidiospores smooth, ellipsoid, yellowish brown, Pileipellis an interwoven trichodermium. Stipitipellis fertile, caulobasidia scattered. Hyphae of context dextrinoid. Clamp connections absent. Odor mild.
Type species: Hemilanmaoa retistipitatus Yang Wang, Bo Zhang & Y. Li
Hemilanmaoa retistipitatus Yang Wang, Bo Zhang & Y. Li, sp. nov.
Mycobank No.: MB845573
Etymology. “retistipitatus” refers to its’ stipe covered with reticulations.
Holotypus. CHINA. Guizhou Province, Tongren City, Yangxi County, 6 July 2019, 108° 30′ 19.35″ E, 27° 38′ 8.16″ N, 3624 (HMJAU 60052!).
Diagnosis. This species is similar to Lanmaoa macrocarpa but differs from the latter by decurrent hymenophore, red pores, stipe covered with reticulations, and context dextrinoid. Basidiocarps bluish when bruising, pileus tomentose, hymenophore decurrent with surface red, stipe covered with reticulations and red dots, and hyphae of context dextrinoid.
Basidioma medium-sized. Pileus around 9 cm in diam., hemispherical with an indistinct or distinct depression at the center when mature, sterile margin narrow; grayish red to pastel red (7B5–10A4) in the center, pale yellow (2A3) toward margin; surface dry, subtomentose, discoloring into blue when touched. Context firm, whitish to pale yellow, turning to blue when cut. Hymenophore decurrent, surface orange red (8A7), changing to blue when injured; pores nearly round to angular; tubes 2–13 mm, light yellow (3A5), changing to blue when injured. Stipe 7.9–10.3 × 1.4–1.6 cm, solid, central, subcylindrical to slightly obclavate, pale yellow (2A3) at the upper partition, brownish red (10C6) downwards, surface coarse, covered with yellow reticulations especially on the upper partition and brownish red dotted-elements, staining blue when touched; context concolorous with that of pileus on the upper partition, gradually brownish red downwards, turning to blue when cut; basal mycelium white. Odor mild.
Basidia 16.5–34 × 10–14 μm, subcylindrical to clavate, hyaline to pale brown in 5% KOH, 2- and 4-spored. Basidiospores (2/4/117) 10.0–11.6–13.1 × 4.2–4.9–5.8 (6.0) μm, Q = 2–2.7 (2.9), Qm = 2.37 ± 0.17, ellipsoid, yellowish brown in 5% KOH, smooth. Hymenophoral trama boletoid, composed of hyaline to brownish yellow hyphae, 2–12 μm wide. Cheilocystidia 22–40 × 7–14.8 μm, narrowly lageniform to lageniform, thin-walled, hyaline to pale brownish yellow in 5% KOH. Pleurocystidia 33–50 × 7–14 μm, similar to cheilocystidia in shape. Pileipellis is an interwoven trichodermium, composed of hyaline to yellowish brown and filamentous hyphae, 2.5–6 μm wide. Stipitipellis fertile, a hymeniform, with inflated terminal cells, ovoid to obovoid, 22–25 × 12–15 μm, hyaline to brownish yellow, caulocystidia 20.2–53 × 8–14.2 μm, lageniform to broadly lageniform, hyaline to pale brownish yellow, caulobasidia scattered. Context dextrinoid in Melzer’s, especially hyphae of stipe base. Clamp connections absent.
Habitat. Solitary or scattered in a mixed broad-leaf forest, dominated by Cyclobalanopsis sp. and Lauraceae.
Distribution. Currently, only known in Guizhou Province, China.
Additional specimens measured. CHINA. Guizhou Province, Tongren City, Yangxi County, 6 July 2019, 108° 30′ 19.35″ E, 27° 38′ 8.16″ N, 3633 (HMJAU 60053).
Notes. Lanmaoa macrocarpa shares some morphological features with He. retistipitatus, viz. similar color of pileus surface and stipe, bluing when injured. However, it can be distinguished by its hymenophore depressed around the apex of the stipe, tubes concolorous with pore surfaces, basal mycelium yellowish, smaller basidiospores (10–12 × 4.5–5 μm), pileipellis a trichoderm and without reaction in Melzer’s.
Hemilanmaoa retistipitatus resembles Cyanoboletus cyaneitinctus (Murrill) A. Farid, A. R. Franck & J. A. Bolin in its reddish color toward stipe base, decurrent hymenophore, and bluing strongly when handled. However, the latter taxon has a duller pileus, yellow hymenophore, basidiospores larger (11.5–15 × 4–6 μm), and stipitipellis sterile.
Porphyrellus pseudocyaneotinctus Yang Wang, Bo Zhang & Y. Li, sp. nov.
MycoBank No.: MB845570
Etymology. The epithet “pseudocyaneotinctus” refers to its similarity to Po. cyaneotinctus.
Holotypus. CHINA. Henan Province, Zhumadian City, Biyang County, 9 July 2021, W3039 (HMJAU 60062!).
Diagnosis. This species is similar to Po. cyaneotinctus, but differs from the latter by different structures of pileipellis, without reaction in Melzer’s and thin-walled caulocystidia with no thickening in the apex. Basidioma brown, often with distinctly cracked pileus, spores broader, and caulocystidia thin-walled.
Basidioma small to medium-sized. Pileus 4.6–9.9 cm wide, subhemispherical to convex or subconvex, caramel (6C6) to light brown (6D6) or brown (6E4), slightly darker in the center; surface dry, tomentose, with finely or distinctly cracked, sometimes with sterile margin at mature; context 0.5–1.2 cm thick, white (3A1), erratically bluish then reddish brown when injured. Hymenophore adnexed or depressed around the apex of stipe with finely decurrent tooth; surface white (3A1) when young, pale gray (1B1) or light brown (6D8) at the mature, becoming blue when bruised; pores angular, 0.75–3/mm; tubes 0.45–1.70 cm long, concolorous with or a little duller than the hymenophoral surface, changing to blue when injured. Stipe 4.8–12.3 × 0.9–1.9 cm, subcylindrical, sometimes slightly expanded or attenuate to base, concolorous with the pileus, streaked, fibrillose, context white (3A1), sometimes nougat (5D3) at the base, usually turning to red-brown when bruised, changing to blue at the apex when cut; basal mycelium white.
Basidiospores (12/12/150) 9.8–11.8–13.8 (14.5) × 4.5–5.3–6.2 μm, Q = 1.83–2.79, Qm = 2.24 ± 0.19, ellipsoid to elongate ellipsoid, inequilateral with a suprahilar depression in side view, light yellow to reddish brown in 5% KOH, smooth. Basidia 24–42 × 9.9–16.2 μm, clavate, 2-, 4-spored, hyaline in 5% KOH, hyaline to yellow in Melzer’s. Hymenophoral trama boletoid consists of 6–17.5 μm wide hyphae. Pleurocystidia 36.8–85 × 8.5–13 μm, lageniform, hyaline in 5% KOH and Melzer’s. Cheilocystidia 38.2–60.5 × 10.8–17.9 μm, similar to pleurocystidia in shape, hyaline in 5% KOH and Melzer’s. Pileipellis a palisadodermium, composed of broadly concatenated cells, hyaline, sometimes brownish in 5% KOH, terminated cells filamentous or pyriform to subfusiform, 20.5–92.5 × 5–23.8 μm, the lower 2–3 cells broad, 26.3–60 × 12.5–25 μm. Stipitipellis sterile, hymeniform with thin-walled and inflated terminal cells, 16.8–49 × 5–18 μm, without a hyaline (in 5% KOH) refractive thickening in the apex of the cells. Clamp connection absent.
Habitat. Solitary on mixed forests dominated by Quercus spp. and Pinus spp.
Known distribution. Currently, only known from Henan Province, China.
Additional collection examined. CHINA. Henan Province, Zhumadian City, Biyang County, Tongshan Lake, 8 July 2021, 113° 29′ 44.48″ E, 32° 46′ 8.32″ N, W3019 (HMJAU 60061), 9 July 2021, 113° 29′ 47.40″ E, 32°46′9.76″ N, W3046 (HMJAU 60063); Baiyun Mountain, 10 July 2021, 113° 34′ 9.57″ E, 32° 53′ 19.70″ N, W3054 (HMJAU 60064), W3062 (HMJAU 60065); Baiyun Mountain, 11 July 2021, 113° 34′ 0.86″ E, 32° 53′ 19.51″ N, W3083, W3084, W3085 (HMJAU 60066), W3088 (HMJAU 60067), W3091 (HMJAU 60068), W3092.
Notes. Phylogenetically, Porphyrellus pseudocyaneotinctus is a sister of Po. griseus Yan C. Li & Zhu L. Yang and Po. pseudofumosipes Yan C. Li & Zhu L. Yang. However, Po. griseus differs from Po. pseudocyaneotinctus in the context of pileus turning to blue when injured, basidiospores smaller (9.5–11.5 × 4.5–5 μm), pleurocystidia shorter (34–58 × 8–12 μm), terminal cells of pileipellis broadly clavate to cystidioid or pyriform. Porphyrellus pseudofumosipes is different from Po. pseudocyaneotinctus in context of pileus bluish when injured, basidiospores smaller (9–11 × 4.5–5.5 μm), and terminal cells of pileipellis pyriform to subfusiform. Morphologically, Porphyrellus pseudocyaneotinctus is similar to Po. castaneus Y.C. Li & Zhu L. Yang, Po. cyaneotinctus (A.H. Sm. & Thiers) Singer, Po. formosanus K.W. Yeh & Z.C. Chen, Po. orientifumosipes Y.C. Li & Zhu L. Yang, and Po. pseudofumosipes Yan C. Li & Zhu L. Yang. However, Po. castaneus is characterized by its duller pileus, bluish when injured, basidiospores smaller (9–11 × 4–5 μm), terminal cells of pileipellis pyriform or subfusiform. Porphyrellus cyaneotinctus differs from Po. pseudocyaneotinctus in the upper 1-3 cells of pileipellis more or less cylindric and 5–9 μm wide, the lower 3-5 cells of pileipellis somewhat to distinctly inflated (9–25 μm wide), pleurocystidia with dark yellow-brown content in Melzer’s or more rarely with strongly amyloid granules, caulocystidia with a hyaline (in KOH) refractive thickening in the apex of the cell that is often forming a lens or cap and grows on poor sandy soil on hillsides with scattered Quercus spp. trees during the late summer and autumn. Porphyrellus formosanus is characterized by its context of pileus turning red when cut, basidiospores larger (14–25 × 5–6.5 um), and pileipellis consisting of filamentous hyphae. Porphyrellus orientifumosipes is different from Po. pseudocyaneotinctus in its pileus context bluish when injured, basidiospores 9–11 × 4.5–5.5 μm, terminal cells of pileipellis pyriform to subfusiform. Porphyrellus pseudofumosipes differs from Po. pseudocyaneotinctus in its pileus context bluish when injured, basidiospores smaller (9–11 × 4.5–5.5 μm), terminal cells of pileipellis 26–57 × 9–16 μm.
Phylloporus biyangensis Yang Wang, Bo Zhang & Y. Li, sp. nov.
MycoBank No.: MB845572
Etymology. “biyangensis” refers to its type locality Biyang County.
Holotypus. CHINA. Henan Province, Zhumadian City, Biyang County, 9 July 2021, W3049 (HMJAU 60059!).
Diagnosis. This species is close to Ph. luxiensis but differs from the latter by yellowish context, smaller spores, larger cheilocystidia, hyaline to pale brown hyphae of pileipellis. Basidioma dull, context yellowish, hymenophore usually discolored when bruising, basidiospores smaller and hyphae of pileipellis sometimes with granular encrustations.
Basidioma medium-sized. Pileus 7.2–11.9 cm in diam., applanate to slightly depressed at center, margin involute when young; surface dry, tomentose, sometimes cracked into squamulose, reddish brown (8E4–8E7). Context approximately 0.4 cm thick at the position halfway to pileus center, yellow (2A6), color unchanging when injured; Hymenophore lamellate, decurrent, 0.6–0.8 mm broad, subdistant, anastomosing, greenish-yellow (1A8), turning blue or unchanging in color when injured; lamellulae common, concolorous with lamellae. Stipe 3.8–4.7 × 0.6–1.3 cm, central, subcylindrical, tapered downwards base, covered with finely reddish brown squamules; ridged along with decurrent lines of lamellae on the upper partition, context light yellow (4A5), color unchanged when injured. Basal mycelium yellow.
Basidiospores (2/3/124) (6.8) 7–9.47–10.5 (11) × (3.5) 3.8–4.15–4.5 (5.5) μm, Q = (1.56) 1.78–2.63 (2.75), Qm = 2.29 ± 0.23, brownish yellow in 5% KOH, ellipsoid, with a suprahilar depression in side view, smooth under a light microscope but with bacillate ornamentation in scanning electron microscope. Basidia 27.2–44 × 6–11.5 μm, 2–, 4–spored, subcylindrical, hyaline in 5% KOH. Hymenophoral trama phylloporoid, composed of hyphae 4.8–20 μm in diameter. Pleurocystidia 41.5–130 × 10–30 μm, lageniform, subfusiform or subclavate, hyaline to pale brown in 5% KOH. Cheilocystidia 58.5–101 × 12–20.5 μm, similar to pleurocystidia in shape, hyaline to pale brown in 5% KOH. Pileipellis an interwoven trichodermium, consists of filamentous hyphae, 6.3–9.5 μm wide, hyaline to pale brown in 5% KOH, sometimes with granular encrustations. Clamp connection absent.
Habitat. Solitary on mixed forests dominated by Quercus spp. and Pinus spp.
Known distribution. Currently, only known in Henan Province, China.
Additional collections examined. CHINA. Henan Province, Zhumadian City, Biyang County, Tongshan Lake, 9 July 2021, 113° 29′ 47.95″ E, 32° 46′ 8.89″ N, W3047 (HMJAU 60057), W3048 (HMJAU 60058), W3049b (HMJAU 60060).
Notes. Phylloporus luxiensis is similar to Ph. biyangensis and Ph. bellus in morphological characteristics. However, Ph. luxiensis is characterized by its white context, and larger basidiospores (10–12 × 4.5–5 μm) (Zeng et al., 2011). Phylloporus bellus differs from Ph. biyangensis in basal mycelium white, spores longer, and cystidia smaller with encrusting pigment sometimes present.
Lanmaoa angustispora G. Wu & Zhu L. Yang
Figures 3K, 5H, 8
Collection. CHINA. Henan Province, Zhumadian City, Biyang County, 8 July 2021, W3013 (HMJAU 60054).
Basidioma medium to large. Pileus 4.6–11.3 cm wide, plano-convex to convex, reddish brown (8E8) when young, light brown (6D7) to brownish yellow (5C8), surface nearly smooth, incurved at the margin; context 1.0–2.1 cm thick, soft, pastel green (29A4) to white (1A1), staining blue immediately when injured. Hymenophore adnexed with a decurrent tooth around stipe or sinuate, surface orange-red (8A6) to Persian orange (6A7) when young, greenish-yellow (1B6) to yellow (2A6) at mature, staining blue when bruised; pores angular to nearly round, 1–2/mm; tubes short, 0.6–1.0 cm long, greenish-yellow (1B6) to yellow (2A6), staining blue when injured. Stipe 6.2–13 × 1–1.8 cm, subcylindrical, greenish-yellow (1A8) at the apex, pastel red (9A5) toward the base, covered with red dots; context yellowish green (30A8), changing to blue when injured. Basal mycelium white.
Basidiospores (1/2/60) 9.00–10.10–11.50 (15.10) × 3.50–3.96–5.00 μm, Q = 2.25–3.02, Qm = 2.55 ± 0.17, subcylindrical to elongate ellipsoid, inequilateral in side view with slightly suprahilar depression, yellow-brown in 5% KOH, smooth. Basidia 20.5–34.0 × 7.2–11.0 μm, clavate, hyaline, or pale brown in 5% KOH. Hymenophoral trama boletoid type consists of 7.5–14.5 μm wide hyphae. Pleurocystidia 29.2–57.8 × 8.2–16.6 μm, lageniform, thin-walled, brown in 5% KOH. Cheilocystidia 25.0–45.9 × 6.8–14.5 μm, similar to pleurocystidia in shape, thin-walled, usually containing yellow to brownish-yellow pigments. Pileipellis interwoven ixotrichodermium consists of hyaline filamentous hyphae 5–7.5 μm in width, with terminal cells subcylindrical or sometimes with subacute apex. Stipitipellis composed of two layers, outer layer trichodermium, consisting of hyaline interwoven filamentous hyphae; inner layer hymeniform, with 16–25 × 8.8–13 μm thin-walled and inflated terminal cells. Clamp connection not observed.
Habitat. Scattered on the sandy soil in Castanea spp. forest.
Known distribution. Known to be distributed in southwestern China and central China.
Additional collections were examined. CHINA. Henan Province, Zhumadian City, Biyang County, 8 July 2021, 113° 29′ 45.25″ E, 32° 46′ 8.62″ N, Q024), W3022 (HMJAU 60056), 9 July 2021, 113° 29′ 47.40″ E, 32° 46′ 9.76″ N, W3029 (HMJAU 60055).
Notes. The hymenophoral characteristics of Lan. angustispora are different from the original description by Wu et al. (2016b) in hymenophoral surface is reddish when young, and yellow at mature. Morphologically, Lan. angustispora resembles Lan. flavorubra and cluster together in the phylogenetic tree; however, stipe of Lan. angustispora smooth, context soft and spores narrower.
Discussion
Macrofungi in East Asia are well-known to mycologists, because of their highly diverse and endemic nature (Hongo, 1960; Horak et al., 2011; Feng et al., 2012; Zeng et al., 2013; Wu et al., 2016a; Cui et al., 2019). The discovery of the new genus Hemilanmaoa further demonstrated this circumstance. Phylogenetically, Hemilanmaoa is embedded in the Pulveroboletus Group, forming a distinct lineage. Hemilanmaoa is the sister of “clade 1” (Figure 1), including Amoenoboletus G. Wu, E. Horak, & Zhu L. Yang, Pulveroboletus Murrill, and Suillellus However, Hemilanmaoa is different from Amoenoboletus in its context, and hymenophores change to blue when injured (Chen et al., 2019); Pulveroboletus differs in its pulverulent surface, distinctly marginal veil, and adnexed hymenophore (Wu et al., 2016a; Zeng et al., 2017); Suillellus can be distinguished by its usually olive pileus surface and amyloid hyphae of stipe base (Vizzini et al., 2014; Wang et al., 2022). Morphologically, Hemilanmaoa is similar to Rubroboletus Kuan Zhao & Zhu L. Yang, Neoboletus Gelardi, Simonini & Vizzini, Caloboletus Vizzini, Cyanoboletus Gelardi, Vizzini & Simonini and Lanmaoa G. Wu & Zhu L. Yang. However, Rubroboletus differs from Hemilanmaoa in its pileal surface reddish and pink to red reticulations on the surface of the stipe (Zhao et al., 2014b); Neoboletus is different from Hemilanmaoa due to its smooth stipe with no reticulations (Wu et al., 2016a); Caloboletus can be distinguished by paler pileus and smooth or reticulate stipe without dots (Wu et al., 2016a); Cyanoboletus is characterized by its stipe pruinose to the furfuraceous surface, ixosubcutis to subcutis pileipellis (Wu et al., 2016a); Lanmaoa differs from Hemilanmaoa in its adnexed or sinuate hymenophore and much thinner hymenophore (Wu et al., 2016b; Chai et al., 2019).
In our study, the phylogenetic analyses of Boletaceae showed different topologies in the BI and ML trees. None is similar to Wu et al. (2016a). The order of seven major clades in the BI tree was Chalciporoideae, Pseudoboletus, Baorangia, Pulveroboletus group, Austroboletoideae, Boletoideae, Leccinoideae, Zangioideae, and Xerocomoideae. However, the order in the ML tree was Chalciporoideae, Pseudoboletus, Zangioideae, Leccinoideae, Xerocomoideae, Austroboletoideae, Pulveroboletus group, and Boletoideae, and Baorangia clade was embed in Pulveroboletus group. In view of our results and combined with phylogenetic analyses of Zhao et al. (2014b), Badou et al. (2022), and Vadthanarat et al. (2022), the positions of the several major clades are still unstable. To confirm the order of the clades formed, more different species and more genes should be introduced to construct the phylogenetic tree. The phylogenetic trees (Figures 1, 2) showed that some sequestrate boletes were not monophyletic groups, such as Turmalinea Orihara & N. Maek., and Heliogaster Orihara & K. Iwase. Turmalinea and Heliogaster were paraphyletic with Rossbeevera T. Lebel & Orihara and Xerocomellus Šutara, respectively. However, in the study of Orihara et al. (2016), Turmalinea was the sister of Rossbeevera and formed an independent lineage. Given the extremely affair phylogenetic relationships between Turmalinea and Rossbeevera, we are suspicious about the reliability of separating Turmalinea from Rossbeevera. However, the further conclusion needs more detailed and abundant information on species.
In the phylogenetic analyses of Porphyrellus and Lanmaoa, our new species clustered with Po. cyaneotinctus (A.H. Sm. & Thiers) Singer. It was first published by Smith and Thiers based on specimens from Michigan and was now re-described by Li et al. according to Chinese collections in 2022 (Smith and Thiers, 1968; Li and Yang, 2021). Similar to Li et al., we also collected specimens from Henan Province, China with similar habitats. Combined with morphological characteristics and phylogenetic results, we recognized that this clade represented one species. However, compared to the original descriptions of Po. cyaneotinctus, there are four main characteristics, viz. (1) a different pileipellis structure, (2) color of pleurocystidia in Melzer’s, (3) features of caulocystidia, and (4) different habitat. Taken all together, they are sufficient to warrant the elevation of the Po. pseudocyaneotinctus.
In the phylogenetic analyses of Phylloporus, the “Phylloporus luxiensis KUN-HKAS 74684” clade was separated from the major clade of “Phylloporus luxiensis,” which was different from the result of Wu et al. (2021) but similar to that of Zeng et al. (2013).
Data availability statement
The data presented in this study are deposited in the Zenodo repository, accession number 10.5281/zenodo.7538325.
Author contributions
YL and BZ: conceptualization. YW: methodology, writing—original draft preparation, and formal analysis. L-YW, Z-XQ, DD, Z-HZ, Y-JL, J-JH, and PZ: investigation. All authors contributed to the article and approved the submitted version.
Funding
This study was supported by the Scientific and Technological Tackling Plan for the Key Fields of Xinjiang Production and Construction Corps (No. 2021AB004), the Research on the Creation of Excellent Edible Mushroom Resources and High Quality & Efficient Ecological Cultivation Technology in Jiangxi Province (20212BBF61002), the Key Project on R&D of the Ministry of Science and Technology (2019YFD1001905-33), the Natural Science Foundation of China (No. 31970), the “111” program (D17014), the Modern Agricultural Scientific Research Collaborative Innovation Special Project (JXXTCXBSJJ202212), the Modern Agroindustry Technology Research System (CARS20), and China Mycological Valley (Hefei), Jilin Provincial Education Department Industrialisation Research Project (JJKH20220358CY).
Acknowledgments
We sincerely thank Mrs. Xin-Ya Yang and Mr. Lei Yue of the Engineering Research Center of Edible and Medicinal Fungi, Ministry of Education, Jilin Agricultural University for their help in the experiment.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2022.1052948/full#supplementary-material
Footnotes
References
Arora, D., and Frank, J. L. (2014). Clarifying the butter Boletes: a new genus, Butyriboletus, is established to accommodate Boletus sect. Appendiculati, and six new species are described. Mycologia 106, 464–480. doi: 10.3852/13-052
Ayala-Vásquez, O., García-Jiménez, J., Aguirre-Acosta, E., Castro-Rivera, R., Ángeles-Argáiz, R. E., Saldivar, ÁE., et al. (2022). Hemiaustroboletus, a new genus in the subfamily Austroboletoideae (Boletaceae, Boletales). MycoKeys 88:55. doi: 10.3897/mycokeys.88.73951
Badou, S. A., Furneaux, B., De Kesel, A., Khan, F. K., Houdanon, R. D., Ryberg, M., et al. (2022). Paxilloboletus gen. nov., a new lamellate bolete genus from tropical Africa. Mycol. Prog. 21, 243–256. doi: 10.1007/s11557-021-01756-y
Biketova, A. Y., Gelardi, M., Smith, M. E., Simonini, G., Healy, R. A., Taneyama, Y., et al. (2022). Reappraisal of the genus Exsudoporus (Boletaceae) worldwide based on multi–gene phylogeny, morphology and biogeography, and insights on Amoenoboletus. J. Fungi 8:101. doi: 10.3390/jof8020101
Binder, M., and Besl, H. (2000). 28S rDNA sequence data and chemotaxonomical analyses on the generic concept of Leccinum (Boletales). Mycologia 2000, 71–82.
Binder, M., and Hibbett, D. S. (2006). Molecular systematics and biological diversification of Boletales. Mycologia 98, 971–981. doi: 10.1080/15572536.2006.11832626
Brundrett, M., Bougher, N., Dell, B., Grove, T., and Malajczuk, N. (1996). Working with Mycorrhizas in Forestry and Agriculture. Canberra: Australian Centre for International Agricultural Research.
Capella-Gutiérrez, S., Silla-Martínez, J. M., and Gabaldón, T. (2009). trimAl: a tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 25, 1972–1973. doi: 10.1093/bioinformatics/btp348
Castellano, M. A., Elliott, T. F., Truong, C., Séné, O., Dentinger, B., and Henkel, T. W. (2016). Kombocles bakaiana gen. sp. nov. (Boletaceae), a new sequestrate fungus from Cameroon. IMA fungus 7, 239–245. doi: 10.5598/imafungus.2016.07.02.03
Chai, H., Liang, Z. Q., Jiang, S., Fu, X. L., and Zeng, N. K. (2018). Lanmaoa rubriceps, a new bolete from tropical China. Phytotaxa 347, 71–80. doi: 10.11646/phytotaxa.347.1.4
Chai, H., Liang, Z. Q., Xue, R., Jiang, S., Luo, S. H., Wang, Y., et al. (2019). New and noteworthy boletes from subtropical and tropical China. MycoKeys 46:55. doi: 10.3897/mycokeys.46.31470
Chapela, I. H., Rehner, S. A., Schultz, T. R., and Mueller, U. G. (1994). Evolutionary history of the symbiosis between fungus-growing ants and their fungi. Science 266, 1691–1694. doi: 10.1126/science.266.5191.1691
Chen, X. N., Zhang, M., Li, T. H., and Zeng, N. K. (2019). A new species of Heimioporus (Boletaceae) from southern China. Phytotaxa 415, 179–188. doi: 10.11646/phytotaxa.415.4.2
Chuankid, B., Vadthanarat, S., Hyde, K. D., Thongklang, N., Zhao, R., Lumyong, S., et al. (2019). Three new Phylloporus species from tropical China and Thailand. Mycol. Prog. 18, 603–614. doi: 10.1007/s11557-019-01474-6
Crous, P. W., Cowan, D. A., Maggs-Kölling, G., Yilmaz, N., Larsson, E., Angelini, C., et al. (2020). Fungal Planet description sheets: 1112–1181. Persoonia 45:251.
Cui, B. K., Li, H. J., Ji, X., Zhou, J. L., Song, J., Si, J., et al. (2019). Species diversity, taxonomy and phylogeny of Polyporaceae (Basidiomycota) in China. Fungal Divers. 97, 137–392. doi: 10.1007/s13225-019-00427-4
Cui, Y. Y., Feng, B., Wu, G., Xu, J. P., and Yang, Z. L. (2016). Porcini mushrooms (Boletus sect. Boletus) from China. Fungal Divers. 81, 189–212. doi: 10.1007/s13225-015-0336-7
Dentinger, B. T., Ammirati, J. F., Both, E. E., Desjardin, D. E., Halling, R. E., Henkel, T. W., et al. (2010). Molecular phylogenetics of porcini mushrooms (Boletus section Boletus). Mol. Phylogenet. Evol. 57, 1276–1292. doi: 10.1016/j.ympev.2010.10.004
Drehmel, D., James, T., and Vilgalys, R. (2008). Molecular phylogeny and biodiversity of the boletes. Fungi 1, 17–23.
Farid, A., Bessette, A. E., Bessette, A. R., Bolin, J. A., Kudzma, L. V., Franck, A. R., et al. (2021). Investigations in the boletes (Boletaceae) of southeastern USA: four novel species and three novel combinations. Mycosphere 12, 1038–1076. doi: 10.5943/mycosphere/12/1/12
Farid, A., Gelardi, M., Angelini, C., Franck, A., Costanzo, F., Kaminsky, L., et al. (2018). Phylloporus and Phylloboletellus are no longer alone: phylloporopsis gen. nov.(Boletaceae), a new smooth–spored lamellate genus to accommodate the American species Phylloporus boletinoides. Fungal Syst. Evol. 2, 341–359. doi: 10.3114/fuse.2018.02.10
Feng, B., Xu, J., Wu, G., Zeng, N. K., Li, Y. C., Tolgor, B., et al. (2012). DNA sequence analyses reveal abundant diversity, endemism and evidence for Asian origin of the porcini mushrooms. PLoS One 7:e37567. doi: 10.1371/journal.pone.0037567
Frank, J., Siegel, N., Schwarz, C., Araki, B., and Vellinga, E. (2020). Xerocomellus (Boletaceae) in western North America. Fungal Syst. Evol. 6, 265–288. doi: 10.3114/fuse.2020.06.13
Gelardi, M., Simonini, G., Ercole, E., Davoli, P., and Vizzini, A. (2015a). Cupreoboletus (Boletaceae, Boletineae), a new monotypic genus segregated from Boletus sect. Luridi to reassign the Mediterranean species B. poikilochromus. Mycologia 107, 1254–1269. doi: 10.3852/15-070
Gelardi, M., Vizzini, A., Ercole, E., Horak, E., Ming, Z., and Li, T. H. (2015b). Circumscription and taxonomic arrangement of Nigroboletus roseonigrescens gen. et sp. nov., a new member of Boletaceae from tropical South-Eastern China. PLoS One 10:e0134295. doi: 10.1371/journal.pone.0134295
Gelardi, M., Simonini, G., Ercole, E., and Vizzini, A. (2014). Alessioporus and Pulchroboletus (Boletaceae, Boletineae), two novel genera for Xerocomus ichnusanus and X. roseoalbidus from the European Mediterranean basin: molecular and morphological evidence. Mycologia 106, 1168–1187. doi: 10.3852/14-042
Halling, R. E., Fechner, N., Nuhn, M., Osmundson, T., Soytong, K., Arora, D., et al. (2015). Evolutionary relationships of Heimioporus and Boletellus (Boletales), with an emphasis on Australian taxa including new species and new combinations in Aureoboletus, Hemileccinum and Xerocomus. Aust. Syst. Bot. 28, 1–22. doi: 10.1071/SB14049
Halling, R. E., Nuhn, M., Fechner, N. A., Osmundson, T. W., Soytong, K., Arora, D., et al. (2012a). Sutorius: a new genus for Boletus eximius. Mycologia 104, 951–961. doi: 10.3852/11-376
Halling, R. E., Nuhn, M., Osmundson, T., Fechner, N., Trappe, J. M., Soytong, K., et al. (2012b). Affinities of the Boletus chromapes group to Royoungia and the description of two new genera, Harrya and Australopilus. Aust. Syst. Bot. 25, 418–431. doi: 10.1071/SB12028
He, M. Q., Zhao, R. L., Hyde, K. D., Begerow, D., Kemler, M., Yurkov, A., et al. (2019). Notes, outline and divergence times of Basidiomycota. Fungal Divers. 99, 105–367. doi: 10.1007/s13225-019-00435-4
Henkel, T. W., Obase, K., Husbands, D., Uehling, J. K., Bonito, G., Aime, M. C., et al. (2016). New Boletaceae taxa from Guyana: binderoboletus segoi gen. and sp. nov., Guyanaporus albipodus gen. and sp. nov., Singerocomus rubriflavus gen. and sp. nov., and a new combination for Xerocomus inundabilis. Mycologia 108, 157–173. doi: 10.3852/15-075
Hongo, T. (1960). The Agaricales of Japan 1–2: rhodophyllaceae, Paxillaceae, Gomphidiaceae, Boletaceae and Strobilomycetaceae. Acta Phytotaxonomica. et Geobotanica. 18, 97–112.
Horak, E., Horak, E., Horak, E., Biologiste, A., Horak, E., and Biologist, A. (2011). Revision of Malaysian Species of ‘Boletales’ s.l. (’Basidiomycota’) Described by EJH Corner (1972, 1974). Malaysia: Forest Research Institute.
Hosen, I., and Li, T. H. (2017). Two new species of Phylloporus from Bangladesh, with morphological and molecular evidence. Mycologia 109, 277–286. doi: 10.1080/00275514.2017.1312196
Hosen, M., Feng, B., Wu, G., Zhu, X. T., Li, Y. C., and Yang, Z. L. (2013). Borofutus, a new genus of Boletaceae from tropical Asia: phylogeny, morphology and taxonomy. Fungal Divers. 58, 215–226. doi: 10.1007/s13225-012-0211-8
Hosen, M., and Yang, Z. L. (2021). Kaziboletus, a new boletoid genus of Boletaceae associated with Shorea robusta in Bangladesh. Mycol. Prog. 20, 1145–1156. doi: 10.1007/s11557-021-01723-7
Hosen, M. I., and Li, T. H. (2015). Phylloporus gajari, a new species of the family Boletaceae from Bangladesh. Mycoscience 56, 584–589. doi: 10.1016/j.myc.2015.05.006
Katoh, K., and Standley, D. M. (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Khmelnitsky, O., Davoodian, N., Singh, P., Raspé, O., Lee, S. M., Fechner, N., et al. (2019). Ionosporus: a new genus for Boletus longipes (Boletaceae), with a new species, I. australis, from Australia. Mycol. Prog. 18, 439–451. doi: 10.1007/s11557-018-01463-1
Kohler, A., Kuo, A., Nagy, L. G., Morin, E., Barry, K. W., Buscot, F., et al. (2015). Convergent losses of decay mechanisms and rapid turnover of symbiosis genes in mycorrhizal mutualists. Nat. Genet. 47, 410–415. doi: 10.1038/ng.3223
Kumar, S., Stecher, G., and Tamura, K. (2016). MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 33, 1870–1874. doi: 10.1093/molbev/msw054
Kuo, M., and Ortiz-Santana, B. (2020). Revision of leccinoid fungi, with emphasis on North American taxa, based on molecular and morphological data. Mycologia 112, 197–211. doi: 10.1080/00275514.2019.1685351
Lanfear, R., Frandsen, P. B., Wright, A. M., Senfeld, T., and Calcott, B. (2017). PartitionFinder 2: new methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 34, 772–773. doi: 10.1093/molbev/msw260
Li, Y., Steenwyk, J. L., Chang, Y., Wang, Y., James, T. Y., Stajich, J. E., et al. (2021). A genome-scale phylogeny of the kingdom Fungi. Curr. Biol. 31, 1653–1665. doi: 10.1016/j.cub.2021.01.074
Li, Y. C., Feng, B., and Yang, Z. L. (2011). Zangia, a new genus of Boletaceae supported by molecular and morphological evidence. Fungal Divers. 49, 125–143. doi: 10.1007/s13225-011-0096-y
Li, Y. C., Li, F., Zeng, N. K., Cui, Y. Y., and Yang, Z. L. (2014a). A new genus Pseudoaustroboletus (Boletaceae, Boletales) from Asia as inferred from molecular and morphological data. Mycol. Prog. 13, 1207–1216. doi: 10.1007/s11557-014-1011-1
Li, Y. C., Ortiz-Santana, B., Zeng, N. K., Feng, B., and Yang, Z. L. (2014b). Molecular phylogeny and taxonomy of the genus Veloporphyrellus. Mycologia 106, 291–306. doi: 10.3852/106.2.291
Li, Y. C., and Yang, Z. L. (2021). The Boletes of China: Tylopilus s.l. New York, NY: Springer. doi: 10.1007/978-981-16-2986-0
Loizides, M., Bellanger, J. M., Assyov, B., Moreau, P. A., and Richard, F. (2019). Present status and future of boletoid fungi (Boletaceae) on the island of Cyprus: cryptic and threatened diversity unravelled by ten–year study. Fungal Ecol. 41, 65–81. doi: 10.1016/j.funeco.2019.03.008
Magnago, A. C., Alves-Silva, G., Henkel, T. W., and da Silveira, R. M. B. (2022). New genera, species, and combinations of Boletaceae from Brazil and Guyana. Mycologia 114, 607–625. doi: 10.1080/00275514.2022.2037307
Malloch, D., Pirozynski, K., and Raven, P. (1980). Ecological and evolutionary significance of mycorrhizal symbioses in vascular plants (a review). Proc. Natl. Acad. Sci. U S A. 77, 2113–2118. doi: 10.1073/pnas.77.4.2113
Marx, D. H. (1972). Ectomycorrhizae as biological deterrents to pathogenic root infections. Annu. Rev. Phytopathol. 10, 429–454. doi: 10.1146/annurev.py.10.090172.002241
Matheny, P. B. (2005). Improving phylogenetic inference of mushrooms with RPB1 and RPB2 nucleotide sequences (Inocybe; Agaricales). Mol. Phylogenet. Evol. 35, 1–20. doi: 10.1016/j.ympev.2004.11.014
Matheny, P. B., and Ammirati, J. F. (2003). Inocybe angustispora, I. taedophila, and Cortinarius aureifolius: an unusual inocyboid Cortinarius. Mycotaxon 88, 401–407.
Miyauchi, S., Kiss, E., Kuo, A., Drula, E., Kohler, A., Sánchez-García, M., et al. (2020). Large-scale genome sequencing of mycorrhizal fungi provides insights into the early evolution of symbiotic traits. Nat. Commun. 11:5125. doi: 10.1038/s41467-020-18795-w
Montoya, L., Garay-Serrano, E., and Bandala, V. M. (2019). Two new species of Phylloporus (Fungi, Boletales) from tropical Quercus forests in eastern Mexico. MycoKeys 51:107. doi: 10.3897/mycokeys.51.33529
Naranjo-Ortiz, M. A., and Gabaldón, T. (2019). Fungal evolution: diversity, taxonomy and phylogeny of the Fungi. Biol. Rev. 94, 2101–2137. doi: 10.1111/brv.12550
Neves, M. A., Binder, M., Halling, R., Hibbett, D., and Soytong, K. (2012). The phylogeny of selected Phylloporus species, inferred from NUC-LSU and ITS sequences, and descriptions of new species from the Old World. Fungal Divers. 55, 109–123. doi: 10.1007/s13225-012-0154-0
Nguyen, L. T., Schmidt, H. A., Von Haeseler, A., and Minh, B. Q. (2015). IQ–TREE: a fast and effective stochastic algorithm for estimating maximum–likelihood phylogenies. Mol. Biol. Evol. 32, 268–274. doi: 10.1093/molbev/msu300
Nuhn, M. E., Binder, M., Taylor, A. F., Halling, R. E., and Hibbett, D. S. (2013). Phylogenetic overview of the Boletineae. Fungal Biol. 117, 479–511. doi: 10.1016/j.funbio.2013.04.008
Orihara, T., Lebel, T., Ge, Z. W., Smith, M., and Maekawa, N. (2016). Evolutionary history of the sequestrate genus Rossbeevera (Boletaceae) reveals a new genus Turmalinea and highlights the utility of ITS minisatellite–like insertions for molecular identification. Persoonia 37, 173–198. doi: 10.3767/003158516X691212
Orihara, T., Smith, M., Shimomura, N., Iwase, K., and Maekawa, N. (2012). Diversity and systematics of the sequestrate genus Octaviania in Japan: two new subgenera and eleven new species. Persoonia 28, 85–112. doi: 10.3767/003158512X650121
Orihara, T., and Smith, M. E. (2017). Unique phylogenetic position of the African truffle-like fungus, Octaviania ivoryana (Boletaceae, Boletales), and the proposal of a new genus, Afrocastellanoa. Mycologia 109, 323–332. doi: 10.1080/00275514.2017.1301750
Parihar, A., Hembrom, M. E., Vizzini, A., and Das, K. (2018). Indoporus shoreae gen. et sp. nov. (Boletaceae) from tropical India. Cryptogam. Mycol. 39, 447–466. doi: 10.7872/crym/v39.iss4.2018.447
Pirozynski, K., and Malloch, D. (1975). The origin of land plants: a matter of mycotrophism. Biosystems 6, 153–164. doi: 10.1016/0303-2647(75)90023-4
Raspé, O., Vadthanarat, S., De Kesel, A., Degreef, J., Hyde, K. D., and Lumyong, S. (2016). Pulveroboletus fragrans, a new Boletaceae species from Northern Thailand, with a remarkable aromatic odor. Mycol. Prog. 15:38. doi: 10.1007/s11557-016-1179-7
Rehner, S. A., and Buckley, E. (2005). A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 97, 84–98. doi: 10.3852/mycologia.97.1.84
Ronquist, F., Teslenko, M., Van Der Mark, P., Ayres, D. L., Darling, A., Höhna, S., et al. (2012). MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542. doi: 10.1093/sysbio/sys029
Shilova, A., Maksimov, A. Y., and Maksimova, Y. G. (2021). Isolation and identification of alkalitolerant bacteria with hydrolytic activity from a soda sludge storage. Microbiology 90, 166–175. doi: 10.1134/S0026261721020120
Skrede, I., Engh, I. B., Binder, M., Carlsen, T., Kauserud, H., and Bendiksby, M. (2011). Evolutionary history of Serpulaceae (Basidiomycota): molecular phylogeny, historical biogeography and evidence for a single transition of nutritional mode. BMC Evol. Biol. 11:230. doi: 10.1186/1471-2148-11-230
Smith, A. H., and Thiers, H. D. (1968). Notes on Boletes—I 1. the generic position of Boletus subglabripes and Boletus chromapes 2. a comparison of four species of Tylopilus. Mycologia 60, 943–954. doi: 10.1080/00275514.1968.12018651
Smith, M. E., Amses, K. R., Elliott, T. F., Obase, K., Aime, M. C., and Henkel, T. W. (2015). New sequestrate fungi from Guyana: jimtrappea guyanensis gen. sp. nov., Castellanea pakaraimophila gen. sp. nov., and Costatisporus cyanescens gen. sp. nov.(Boletaceae, Boletales). IMA Fungus 6, 297–317. doi: 10.5598/imafungus.2015.06.02.03
Smith, M. E., Henkel, T. W., Uehling, J. K., Fremier, A. K., Clarke, H. D., and Vilgalys, R. (2013). The ectomycorrhizal fungal community in a neotropical forest dominated by the endemic dipterocarp Pakaraimaea dipterocarpacea. PLoS One 8:e55160. doi: 10.1371/journal.pone.0055160
Sulzbacher, M. A., Orihara, T., Grebenc, T., Wartchow, F., Smith, M. E., Martín, M. P., et al. (2020). Longistriataflava (Boletaceae, Basidiomycota)-a new monotypic sequestrate genus and species from Brazilian Atlantic Forest. MycoKeys 62:53. doi: 10.3897/mycokeys.62.39699
Swindell, S. R., and Plasterer, T. N. (1997). ”Seqman,” in Sequence Data Analysis Guidebook, New York, NY: Springer doi: 10.1385/0896033589
Tedersoo, L., May, T. W., and Smith, M. E. (2010). Ectomycorrhizal lifestyle in fungi: global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 20, 217–263. doi: 10.1007/s00572-009-0274-x
Trappe, J. M. (1977). Selection of fungi for ectomycorrhizal inoculation in nurseries. Annu. Rev. Phytopathol. 15, 203–222. doi: 10.1146/annurev.py.15.090177.001223
Trappe, J. M., Castellano, M. A., Halling, R. E., Osmundson, T. W., Binder, M., Fechner, N., et al. (2013). Australasian sequestrate fungi 18: solioccasus polychromus gen. & sp. nov., a richly colored, tropical to subtropical, hypogeous fungus. Mycologia 105, 888–895. doi: 10.3852/12-046
Treub, M. (1884). Recherches sur les Cycadées. 3. Embryogénie du Cycas circinalis. Ann. Jard. Bot. Buitenzorg. 4, 1–11.
Vadthanarat, S., Lumyong, S., and Raspé, O. (2019). Cacaoporus, a new Boletaceae genus, with two new species from Thailand. MycoKeys 54:1. doi: 10.3897/mycokeys.54.35018
Vadthanarat, S., Raspé, O., and Lumyong, S. (2018). Phylogenetic affinities of the sequestrate genus Rhodactina (Boletaceae), with a new species, R. rostratispora from Thailand. MycoKeys 29:63. doi: 10.3897/mycokeys.29.22572
Vadthanarat, S., Raspé, O., and Lumyong, S. (2022). Rubinosporus auriporus gen. et sp. nov. (Boletaceae: xerocomoideae) from tropical forests of Thailand, producing unusual dark ruby spore deposits. J. Fungi 8:278. doi: 10.3390/jof8030278
Vizzini, A., Simonini, G., Ercole, E., and Voyron, S. (2014). Boletus mendax, a new species of Boletus sect. Luridi from Italy and insights on the B. luridus complex. Mycol. Prog. 13, 95–109. doi: 10.1007/s11557-013-0896-4
Wang, Y., Tuo, Y. L., Wu, D. M., Gao, N., Zhang, Z. H., Rao, G., et al. (2022). Exploring the relationships between four new species of boletoid fungi from Northern China and their related species. J. Fungi 8:218. doi: 10.3390/jof8030218
Wu, G., Feng, B., Xu, J. P., Zhu, X. T., Li, Y. C., Zeng, N. K., et al. (2014). Molecular phylogenetic analyses redefine seven major clades and reveal 22 new generic clades in the fungal family Boletaceae. Fungal Divers. 69, 93–115. doi: 10.1007/s13225-014-0283-8
Wu, G., Lee, S. M., Horak, E., and Yang, Z. L. (2018). Spongispora temasekensis, a new boletoid genus and species from Singapore. Mycologia 110, 919–929. doi: 10.1080/00275514.2018.1496387
Wu, G., Li, Y. C., Zhu, X. T., Zhao, K., Han, L. H., Cui, Y. Y., et al. (2016a). One hundred noteworthy boletes from China. Fungal Divers. 81, 25–188. doi: 10.1007/s13225-016-0375-8
Wu, G., Zhao, K., Li, Y. C., Zeng, N. K., Feng, B., Halling, R. E., et al. (2016b). Four new genera of the fungal family Boletaceae. Fungal Divers. 81, 1–24. doi: 10.1007/s13225-015-0322-0
Wu, G., Miyauchi, S., Morin, E., Kuo, A., Drula, E., Varga, T., et al. (2022). Evolutionary innovations through gain and loss of genes in the ectomycorrhizal Boletales. New Phytol. 233, 1383–1400. doi: 10.1111/nph.17858
Wu, K., Wu, G., and Yang, Z. L. (2020). A taxonomic revision of Leccinum rubrum in subalpine coniferous forests, southwestern China. Acta Edulis. Fungi 27:92.
Wu, L. L., Liang, Z. Q., Su, M. S., Fan, Y. G., Zhang, P., Jiang, S., et al. (2021). Updated taxonomy of Chinese Phylloporus (Boletaceae, Boletales): six new taxa and four redescribed species. Mycol. Prog. 20, 1243–1273. doi: 10.1007/s11557-021-01722-8
Yang, Z. L. (2011). Molecular techniques revolutionize knowledge of basidiomycete evolution. Fungal Divers. 50, 47–58. doi: 10.1007/s13225-011-0121-1
Yang, Z. L., Wang, X., and Binder, M. (2003). A study of the type and additional materials of Boletus thibetanus. Mycotaxon 86:e290.
Zeng, N. K., Cai, Q., and Yang, Z. L. (2012). Corneroboletus, a new genus to accommodate the southeastern Asian Boletus indecorus. Mycologia 104, 1420–1432. doi: 10.3852/11-326
Zeng, N. K., and Jiang, S. (2020). Atlas of Macrofungi from Yinggeling of Hainan, China. China: Nanhai Publishing House.
Zeng, N. K., Liang, Z. Q., Tang, L. P., Li, Y. C., and Yang, Z. L. (2017). The genus Pulveroboletus (Boletaceae, Boletales) in China. Mycologia 109, 422–442. doi: 10.1080/00275514.2017.1331689
Zeng, N. K., Su, M. S., Liang, Z. Q., and Yang, Z. L. (2015). A geographical extension of the North American genus Bothia (Boletaceae, Boletales) to East Asia with a new species B. fujianensis from China. Mycol. Prog. 14:1015. doi: 10.1007/s11557-014-1015-x
Zeng, N. K., Tang, L. P., Li, Y. C., Tolgor, B., Zhu, X. T., Zhao, Q., et al. (2013). The genus Phylloporus (Boletaceae, Boletales) from China: morphological and multilocus DNA sequence analyses. Fungal Divers. 58, 73–101. doi: 10.1007/s13225-012-0184-7
Zeng, N. K., Tang, L. P., and Yang, Z. L. (2011). Type studies on two species of Phylloporus (Boletaceae, Boletales) described from southwestern China. Mycotaxon 117, 19–28. doi: 10.5248/117.19
Zeng, N. K., Wu, G., Li, Y. C., Liang, Z. Q., and Yang, Z. L. (2014). Crocinoboletus, a new genus of Boletaceae (Boletales) with unusual boletocrocin polyene pigments. Phytotaxa 175, 133–140. doi: 10.11646/phytotaxa.175.3.2
Zhang, D., Gao, F., Jakovliæ, I., Zou, H., Zhang, J., Li, W. X., et al. (2020). PhyloSuite: an integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 20, 348–355. doi: 10.1111/1755-0998.13096
Zhang, M., and Li, T. H. (2018). Erythrophylloporus (Boletaceae, Boletales), a new genus inferred from morphological and molecular data from subtropical and tropical China. Mycosystema 37, 1111–1126.
Zhang, Y., Xue, R., Su, M. S., Wu, L. L., and Zeng, N. K. (2019). Phylloporus rubiginosus, a noteworthy lamellar bolete from tropical Asia. Guizhou. Sci. 37, 1–5.
Zhao, K., Wu, G., Feng, B., and Yang, Z. L. (2014a). Molecular phylogeny of Caloboletus (Boletaceae) and a new species in East Asia. Mycol. Prog. 13, 1127–1136. doi: 10.1007/s11557-014-1001-3
Zhao, K., Wu, G., and Yang, Z. L. (2014b). A new genus, Rubroboletus, to accommodate Boletus sinicus and its allies. Phytotaxa 188, 61–77.
Keywords: Boletales, new taxa, Hemilanmaoa, Lanmaoa, Phylloporus, Porphyrellus
Citation: Wang Y, Wang L-Y, Dai D, Qi Z-X, Zhang Z-H, Liu Y-J, Hu J-J, Zhang P, Li Y and Zhang B (2023) Boletaceae in China: Taxonomy and phylogeny reveal a new genus, two new species, and a new record. Front. Microbiol. 13:1052948. doi: 10.3389/fmicb.2022.1052948
Received: 24 September 2022; Accepted: 08 December 2022;
Published: 02 February 2023.
Edited by:
Ji-Chuan Kang, Guizhou University, ChinaReviewed by:
Samantha Chandranath Karunarathna, Qujing Normal University, ChinaYusufjon Gafforov, Academy of Sciences of the Republic of Uzbekistan, Uzbekistan
Copyright © 2023 Wang, Wang, Dai, Qi, Zhang, Liu, Hu, Zhang, Li and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yu Li,  ZnVuZ2k5NjZAMTI2LmNvbQ==; Bo Zhang, emhhbmdib2Z1bmdpQDEyNi5jb20=
ZnVuZ2k5NjZAMTI2LmNvbQ==; Bo Zhang, emhhbmdib2Z1bmdpQDEyNi5jb20=