
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol. , 14 January 2022
Sec. Virology
Volume 12 - 2021 | https://doi.org/10.3389/fmicb.2021.817200
 Marcela Salazar-García1,2
Marcela Salazar-García1,2 Samyr Acosta-Contreras2
Samyr Acosta-Contreras2 Griselda Rodríguez-Martínez2
Griselda Rodríguez-Martínez2 Armando Cruz-Rangel3
Armando Cruz-Rangel3 Alejandro Flores-Alanis4
Alejandro Flores-Alanis4 Genaro Patiño-López5
Genaro Patiño-López5 Victor M. Luna-Pineda2,5*
Victor M. Luna-Pineda2,5*
World Health Organization (WHO) has prioritized the infectious emerging diseases such as Coronavirus Disease (COVID-19) in terms of research and development of effective tests, vaccines, antivirals, and other treatments. Severe Acute Respiratory Syndrome-Coronavirus-2 (SARS-CoV-2), the etiological causative agent of COVID-19, is a virus belonging to risk group 3 that requires Biosafety Level (BSL)-3 laboratories and the corresponding facilities for handling. An alternative to these BSL-3/-4 laboratories is to use a pseudotyped virus that can be handled in a BSL-2 laboratory for study purposes. Recombinant Vesicular Stomatitis Virus (VSV) can be generated with complementary DNA from complete negative-stranded genomic RNA, with deleted G glycoprotein and, instead, incorporation of other fusion protein, like SARS-CoV-2 Spike (S protein). Accordingly, it is called pseudotyped VSV-SARS-CoV-2 S. In this review, we have described the generation of pseudotyped VSV with a focus on the optimization and application of pseudotyped VSV-SARS-CoV-2 S. The application of this pseudovirus has been addressed by its use in neutralizing antibody assays in order to evaluate a new vaccine, emergent SARS-CoV-2 variants (delta and omicron), and approved vaccine efficacy against variants of concern as well as in viral fusion-focused treatment analysis that can be performed under BSL-2 conditions.
Infectious diseases develop and re-emerge regularly, triggering epidemics and pandemics (Morens et al., 2008). Human Immunodeficiency Virus/Acquired Immunodeficiency Syndrome (HIV/AIDS; 1981), Nipah virus (1999), Severe Acute Respiratory Syndrome (SARS; 2002), Middle East Respiratory Syndrome (MERS; 2012), and Coronavirus Disease (COVID-19; 2019) are examples of newly emerging infectious diseases while re-emerging infectious diseases have reappeared in new locations, such as West Nile in the United States and Russia (Morens and Fauci, 2020). The World Health Organization’s (WHO) Research and Development Blueprint Initiative has prioritized infection diseases such as COVID-19, Crimean-Congo hemorrhagic fever, Ebola virus and Marburg virus diseases, Lassa fever, MERS and SARS, Nipah and henipaviral diseases, Rift Valley fever, and Zika for the development of effective tests, vaccines, antivirals, and other treatments (Kieny et al., 2016). To be sure, the pathogens that cause these diseases are classified as risk group 3 (high individual risk and low risk to the community) or risk group 4 (high individual risk and high risk to the community, without treatment), which necessitates the use of Biosafety Level (BSL)-3 and BSL-4 laboratories and facilities for handling and propagation (Artika and Ma’roef, 2017).
Many nations across the globe lack the infrastructure and resources needed to research these emerging and re-emerging pathogens. There are just seven BSL-3 laboratories in Mexico, and they are typically overworked. From these, the Biosafety Level 3 Laboratory from Institute for Epidemiological Diagnosis and Reference1, Biosecurity Unit from Institute of Biomedical Research from UNAM2, Animal Health Laboratory from National Service of Agrifood Health, Safety and Quality3, National Laboratory for Maximum Biological Safety from National Institute of Medical Sciences and Nutrition “Salvador Zubirán,”4 and Emerging and Non-Emerging Pathogens Research Tower from National Institute of Respiratory Diseases5 are located in Mexico City, whereas the other two BSL-3: the Center for Research and Assistance in Technology and Design of the State of Jalisco6 and the University of Monterrey/Autonomous University of Nuevo León7 are located in Guadalajara, Jalisco and Nuevo León, Monterrey, respectively. However, because Mexico has no BSL-4 laboratories, research with WHO-designated priority diseases such as Ebola is hampered (Figure 1).
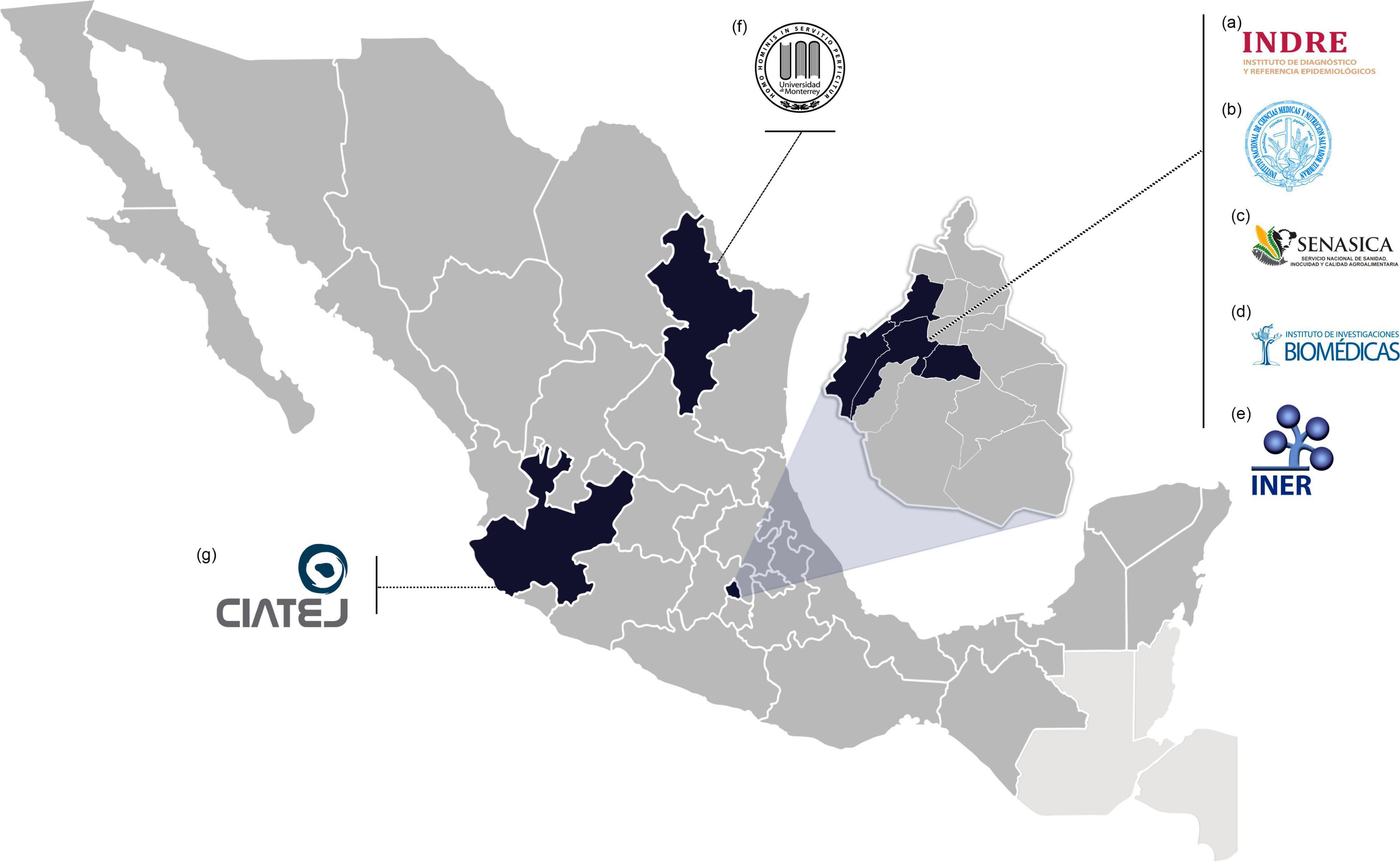
Figure 1. Mexican BSL-3 laboratories. (a) BSL-3 Laboratory from the Institute for Epidemiological Diagnosis and Reference (Mexico City); (b) The Biosecurity Unit from Institute of Biomedical Research (UNAM, Mexico City); (c) Animal Health Laboratory from National Service of Agrifood Health, Safety, and Quality (Mexico City); (d) National Laboratory for Maximum Biological Safety from National Institute of Medical Sciences and Nutrition “Salvador Zubirán” (Mexico City); (e) Emerging and Non-Emerging Pathogens Research Tower from National Institute of Respiratory Diseases (Mexico City); (f) Center for Research and Assistance in Technology and Design of the State of Jalisco (Guadalajara City), and (g) University of Monterrey/Autonomous University of Nuevo León (Monterrey City).
To enable the study of BSL-3/-4 pathogens under BSL-2 laboratory conditions, one can use pseudotyped virus (PV) to study virus entry (Millet et al., 2019). PV, sometimes known as “pseudoviruses” or “pseudoparticles” (pp), are amplification-defective viruses capable of just one cycle of replication that can infect host cells in the same way as wild-type viruses do (Takada et al., 1997). PVs are primarily derived from retroviruses [HIV and Murine Leukemia Virus (MLV)] and rhabdoviruses (VSV) and are used to investigate the function of viral fusion proteins in enveloped viruses such as lifecycle initiation, host and cellular tropism, interspecies transmission, viral pathogenesis, and host cell entry pathways (Moore et al., 2004; Whitt, 2010; Mendenhall et al., 2012). VSV is a negative polarity enveloped RNA virus with a genome size of 11 kb that contains five main viral proteins: nucleoprotein (N), phosphoprotein (P), matrix protein (M), glycoprotein (G), and large polymerase protein (L). VSV has been frequently utilized as an enveloped virus for the creation of efficient PV harboring a foreign virus’s surface protein.
Pseudotyped viruses are useful tools for studying the function of viral fusion proteins (Moore et al., 2004; Whitt, 2010; Mendenhall et al., 2012). PVs have been used in phenotypic mixing since the 1970s, in which two encapsulated viruses “share” coat proteins while having distinct genetic material (RNA or DNA) (Choppin and Compans, 1970; Závada, 1972; Huang et al., 1974). Thus, a temperature-sensitive (ts) mutant of VSV was reported, which was deficient in the synthesis of its G protein at nonpermissive temperatures (Schnitzer et al., 1979). Several PVs were supplemented with foreign viral glycoproteins (GPs) using VSV strain ts045, including VSV-Avian sarcoma viruses, VSV-Rous sarcoma virus, VSV-Murine leukemia virus, VSV-Murine oncoviruses and -murine cytomegalovirus, and Rous-associated virus 1 is a VSV-Avian retrovirus (Weiss et al., 1977; Lodish and Weiss, 1979; Schnitzer and Gonczol, 1979; Weiss and Bennett, 1980). Furthermore, an infectious defective interfering (DI) particle was characterized as a VSV strain with faulty replication and amplification of its RNA genome but proper viral packaging (Pattnaik and Wertz, 1990). These studies demonstrated RNAs produced from non-viral origins could be packaged into VSV particles. When co-expressed with the other VSV proteins, a full negative-stranded genomic RNA from a cDNA clone of a VSV DI RNA was replicated, transcribed, and packed into infectious particles (Pattnaik et al., 1992; Stillman et al., 1995). Full-length positive-sense RNA, complementary to the VSV genome, can be produced using the bacteriophage T7 RNA polymerase. Production of the full-length anti-genome along with proteins required for RNA replication (N, P, and L) enable the recovery of replication-competent recombinant (r)VSV from DNA plasmids (Lawson et al., 1995; Whelan et al., 1995). Recovery of rVSVs lacking the glycoprotein open reading frame from the genome (ppVSVΔG) is accomplished by supplying the VSV G in trans and they are named ppVSVΔG-G (Schnell et al., 1996). Several reporter genes have been cloned into ppVSVΔG permitting various experimental read-outs, and these included green and red fluorescent protein (GFP/RFP/DsRed), secreted Embryonic Alkaline Phosphatase (SEAP), and firefly luciferase (fLuc), generating ppVSVΔG-reporter (Takada et al., 1997; Fukushi et al., 2008; Tani et al., 2010; Muik et al., 2012).
Enveloped viruses have fusion proteins that enable attachment and fusion into host cells (Barrett and Dutch, 2020). These viral fusion proteins are classified structurally as class I (e.g., HIV Env Glycoprotein), class II (e.g., Rift Valley fever virus glycoprotein C), and class III (e.g., VSV G glycoprotein), with all of them exhibiting both pre- and post-fusion static conformations (Blumenthal et al., 2012; Dessau and Modis, 2013; Kim et al., 2017). Lassa, Ebola, HIV, MERS, SARS, and SARS-CoV-2 viruses all feature class I viral fusion glycoproteins that have two domains, the C- and N-terminal domains located between the furin-like protease cleavage site (Figure 2). In their pre- and post-fusion states, they form homotrimers, and their C terminal domain contains two heptad repeats (HR), a single-pass transmembrane motif, and a cytoplasmic tail (CT) (Muñoz-Barroso et al., 1999; Lee et al., 2008; Hastie et al., 2017; Pallesen et al., 2017; Yuan et al., 2017; Wrapp et al., 2020).
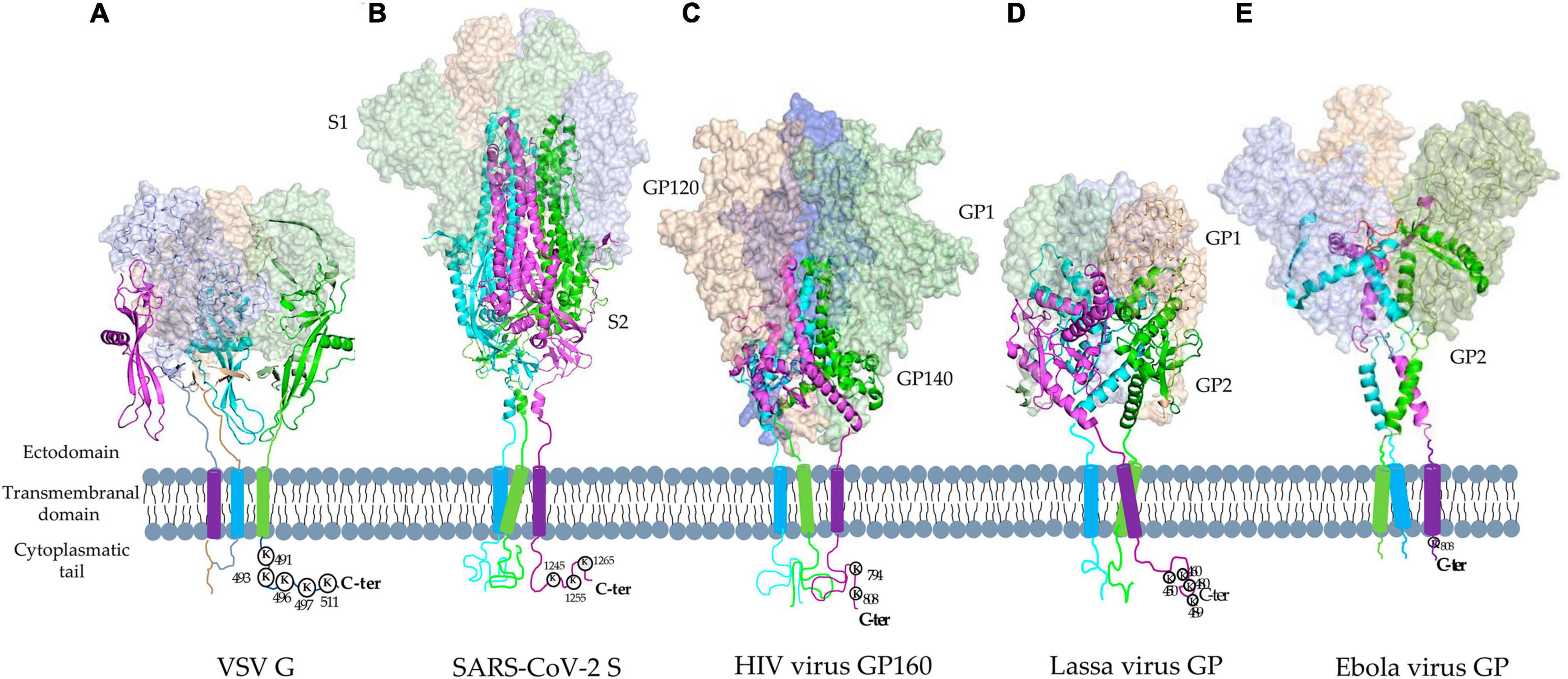
Figure 2. Tridimensional structures of Vesicular Stomatitis Virus G protein and the main class I viral fusion proteins in pre-fusion static conformations. (A) Vesicular Stomatitis Virus (VSV) class III fusion glycoprotein (PDB: 6TIT) and representative class I viral fusion proteins: (B) SARS-CoV-2 S (PDB: 6VXX), (C) HIV glycoprotein (GP) 160 (PDB: 6ULC), (D) Lassa virus GP (PDB: 6P91), and (E) Ebola virus GP (PDB: 6QD7). They form homotrimers with two domains, the C- and N-terminal domains (Magenta, green, and cyan represent each monomer). N terminal contains the receptor binding site and C terminal domain contains two heptad repeats (HR), a single-pass transmembrane motif, and a cytoplasmic tail (CT).
The membrane-associated RING-CH (MARCH)-8, a RING (really interesting new gene)-finger E3 ubiquitin ligase, has been reported to downregulate human transmembrane proteins, including the enveloped viral glycoproteins SARS-CoV-2 spike (S), HIV-1 Env, and EboV-GP (Tada et al., 2015). The CTs found in these viral glycoproteins are made up of Lys residues that can vary in quantity and serve as targets for MARCH-mediated ubiquitination (Figure 3A). Interestingly, expression of MARCH8 in the virus-producing cells reduced the levels of viral glycoprotein and compromised infection of cells was obtained by replacing Lys residues with Ala (K to A) in the CTs of these glycoproteins (Lun et al., 2021). Although S glycoprotein CTs of Coronaviruses (CoVs) features cysteine-rich motifs (six conserved residues) play an important role in S glycoprotein function, these cysteines are palmitoylated and their substitution with Ala (Cys-to-A) affect the S-mediated cell fusion of these viruses (Figure 3; Chang et al., 2000; Petit et al., 2007).
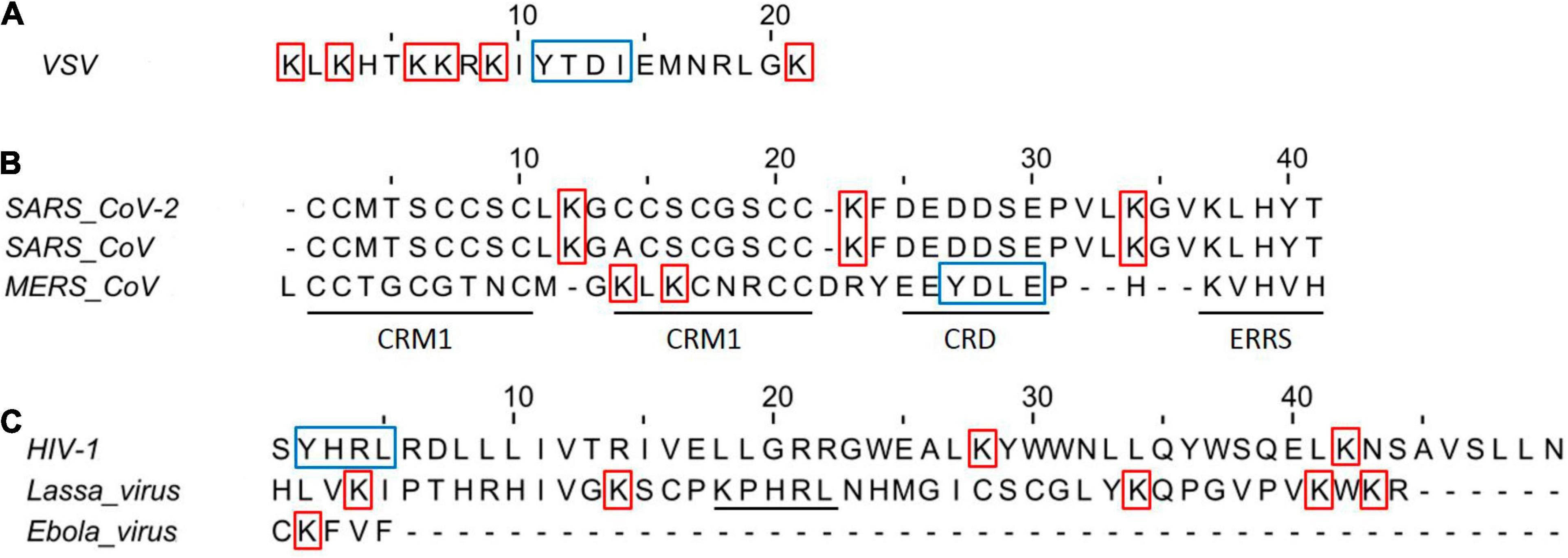
Figure 3. Amino acid sequences of viral glycoproteins class I CT domains. (A) Vesicular stomatitis virus CT domain Indian strain. (B) Amino acid sequence alignment of CoVs Spike protein in the CT domains. CRM, cysteine-rich motif; CRD, charge-rich domain; ERRS, endoplasmatic reticulum retrieval signal (KxHxx). The amino acid alignment was performed with the Jalview v2.11.1.4 using CLUSTAL W. (C) Amino acid of HIV, Lassa, and Ebola virus glycoproteins CT domains. Underline in Lassa virus indicates the ERRS motif. The HIV CT domain was trimmed to 60 and 40 amino acids in the N and C terminal, respectively. The red squares indicate Lys that could be implicated in efficient infectivity, while the blue squares indicate the tyrosine-dependent internalization signals (YxxΦ motif, where Φ is F, I, L, M, or V).
Coronaviruses S CT comprises a dilysine (KKxx-COOH) or a dibasic (KxHxx-COOH) endoplasmic reticulum retrieval signal, as well as a tyrosine-dependent localization signal (YxxF or YxxI motif) that interacts with the CoVsM protein for virions incorporation (Figure 3; Lontok et al., 2004; McBride et al., 2007; Winter et al., 2008; Shirato et al., 2011). CoVs S CT with a nonsense mutation-generated 21- or 24-amino acid deletions (conserved KxHxx motif) showed the highest viral spread and the appearance of non-syncytium-forming infectious centers, likely driving rVSV-SARS-CoV-2 S adaptation for the efficient spread in tissue culture (Case et al., 2020b; Dieterle et al., 2020). In addition, rVSV-Hantaan virus Gn and Gc glycoproteins also acquired a substitution in CT (I532K) of Gn, and a substitution (S1094L) in the stem region of Gc following three serial passages in Vero cells, that have the highest viral spread likely for relocalization of Gn/Gc from the Golgi complex to the cell surface (Slough et al., 2019). Overall, all findings emphasize the relevance of CT of these glycoproteins in the production of virus-like particles and virion, including pseudotyped viruses. Pseudotyped VSV virions have been used to assess cellular tropism, glycoprotein function, receptor recognition, and neutralization antibody assay in viruses from risk groups 3 and 4 (Table 1), including Crimean-Congo hemorrhagic fever virus, Ebola virus, Marburg virus, Lassa fever virus, Nipah virus, Rift Valley virus, MERS-CoV, and SARS-CoV-2 virus (Kunz et al., 2005; Fukushi et al., 2008; Kaku et al., 2012; Bukbuk et al., 2014; Suda et al., 2016; Lennemann et al., 2017; Lester et al., 2019; Saito et al., 2020; Zettl et al., 2020).
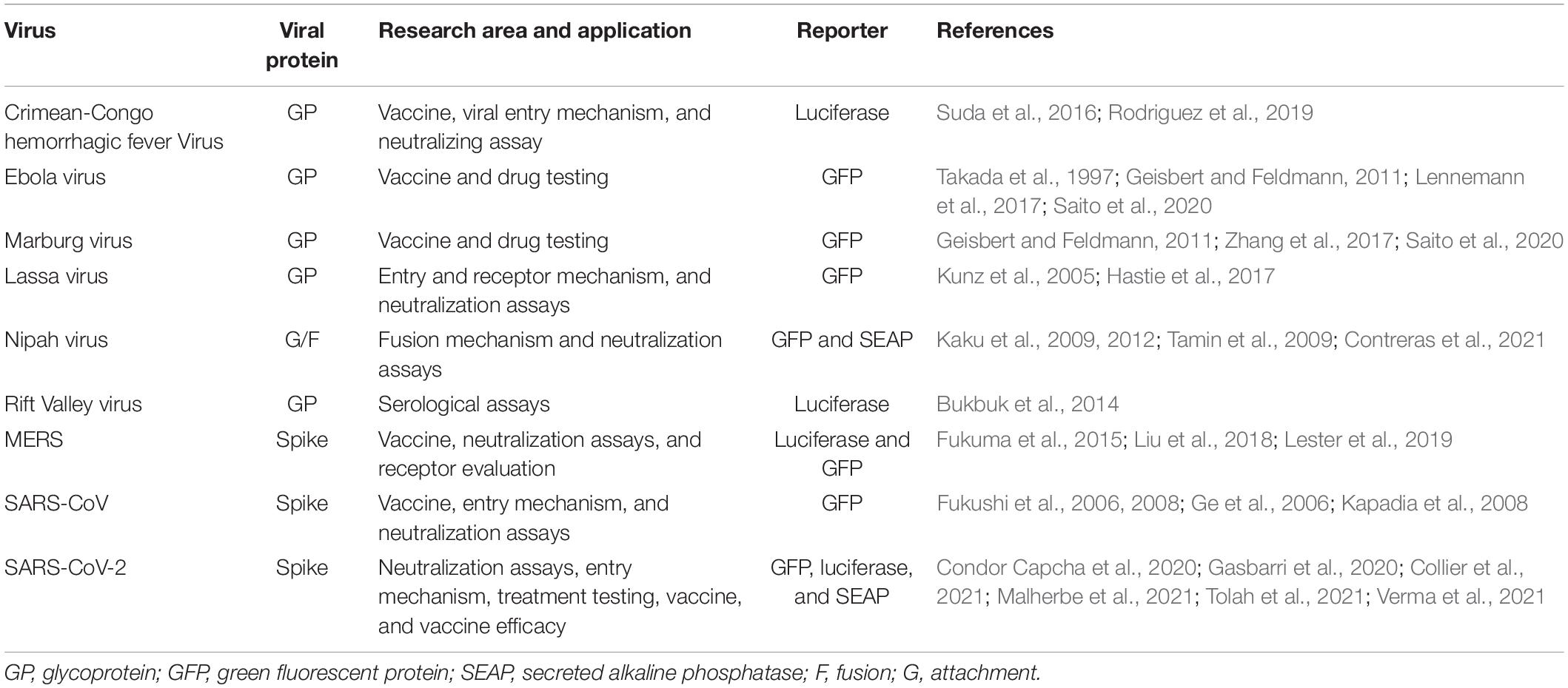
Table 1. Application of pseudotyped Vesicular Stomatitis Virus (VSV).
Pneumonia caused by the SARS-CoV-2 virus was recognized as COVID-19 by the WHO and is now considered a pandemic, with over 200 million cases and over four million fatalities globally8. More than three million COVID-19 infections have been reported in Mexico, with over 250 thousand deaths9. Because of the SARS-CoV-2 virus’s high infectivity, toxicity, and lack of therapies, BSL-3 laboratories are necessary for its handling in drug testing, neutralizing antibodies, and authorized vaccination effectiveness (Kaufer et al., 2020). As a result, most research laboratories have been unable to conduct SARS-CoV-2 research; however, the use of ppVSVΔG-SARS-CoV-2 S provides a feasible option for studying this virus at BSL-2 facilities (Plescia et al., 2021).
The generation of ppVSVΔG-SARS-CoV-2 S may be classified as (a) ppVSVΔG packaging and (b) SARS-CoV-2 S protein production (Figure 4). The ppVSVΔG is a viral particle that can be packaged in vitro by transfection of six plasmids into the bacteriophage T7 RNA polymerase-harboring cell. Although these particles lack a glycoprotein in their genome, they are coated in VSV-G in trans that result in ppVSVΔG-G (G-complemented particles). In a new passage of cells, it may incorporate the SARS-CoV-2 S protein onto the virus’s surface by cotransfection with the S gene-containing plasmid, producing ppVSVΔG-SARS-CoV-2-S particles or pseudotyped VSV-SARS-CoV-2-S (Lawson et al., 1995; Whelan et al., 1995; Schnell et al., 1996).
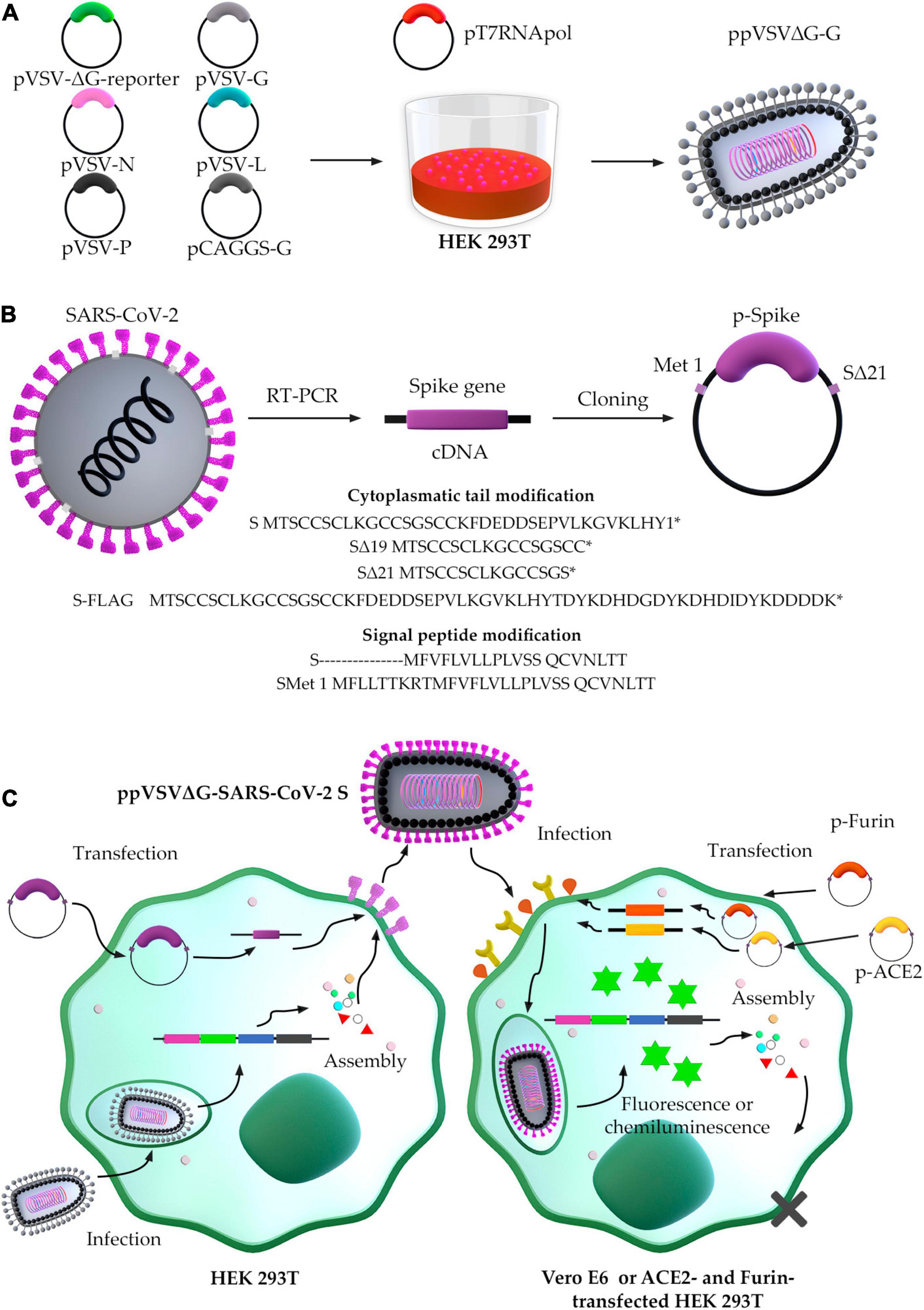
Figure 4. Scheme of the generation of pseudotyped vesicular stomatitis virus-severe acute respiratory syndrome-coronavirus-2 (SARS-CoV-2) S. (A) Packaging of ppVSV. (B) Optimized conditions for cloning Spike gene. (C) ppVSVΔG-SARS-CoV-2 S assembly in HEK 293T cells, and infection assay in Vero E6 cells by ppVSVΔG-SARS-CoV-2 S.
In primary transfection, five plasmids are used to accomplish ppVSVΔG packaging: (1) pVSV-ΔG-reporter containing all VSV antigenome directed by the T7 promoter, excepting the GP gene, which was deleted and replaced by a reporter gene-GFP, -RFP/DsRed, -SEAP, or -fLuc; (2) pVSV-G containing the VSV GP gene directed by the T7 promoter; (3) pVSV-L containing the VSV polymerase (L) gene directed by T7 promoter; (4) pVSV-N containing VSV nucleocapsid (N) gene directed by T7 promoter; and (5) pVSV-P containing VSV phosphoprotein (F) gene directed by T7 promoter. During cellular passages, ppVSVΔG-G is generated with pCAGGS-G containing VSV GP gene directed by pol II promoter as chicken beta-actin promoter (Figure 4A).
To be able to transcribe the VSV antigenome and the N, P, G, and L genes, the HEK293T cell line must first be infected with the vaccinia virus or transfected with pT7pol (expressing the bacteriophage T7 RNA polymerase). The VSV polymerase complex (VSV L and P proteins) and the envelope nucleoprotein N produce genome from the antigenome RNA produced by T7. The matrix (M) protein is encoded in the VSV RNA genome and is produced after the genome is made and transcription occurs. Because the genome lacks glycoprotein, the cell line must be transfected with pCAGGS-G to produce G in trans, and make infectious G pseudotyped ppVSVΔG-G virions (Figure 4A; Whitt, 2010).
The SARS-CoV-2 S protein interacts with the host cell receptor, allowing viral and cellular membranes to fuse. The SARS-CoV-2 S protein comprises a signal peptide, an N-terminal S1 domain, and a C terminal S2 domain and has 1,274 amino acids. Replacing rare codons with abundant cognate tRNA in the cytosol can affect protein translation from mRNA, resulting in guanine and cytosine enrichment as a process of sequence optimization with higher steady-state mRNA levels in vitro and protein expression in vivo (Kudla et al., 2006; Mauro and Chappell, 2014). Furthermore, the SARS-CoV-2 S protein comprises a short signal peptide with weak Sec61 recognition, but the inclusion of the nine upstream residues improves recognition and increases protein levels (Havranek et al., 2020).
Severe Acute Respiratory Syndrome-Coronavirus-2 isolates from the initial human cases in Wuhan had an S protein with the D614 form; however, the viruses that are currently circulating in the human population have the G614 form. SARS-CoV-2 viruses with the G614 mutation in the S protein have greater infectious titers in vitro than those with the D614 mutation (Korber et al., 2020). Another R682Q mutation in the S protein has been characterized as a fast adaptation of SARS-CoV-2 after successive passaging in Vero E6 cells (Ogando et al., 2020). Both the D614G and R682Q alterations improve VSV-SARS-CoV-2 pseudotyping, with the S protein containing 19 deleted residues into the C terminus (19) (Johnson et al., 2020). CT alterations of various viral envelope GPs, including truncation and point mutation in this tail, may be necessary to facilitate appropriate integration into the ppVSVΔG-SARS-CoV-2 S. The greater incorporation into ppVSVΔG-SARS-CoV-2 S is most likely related to a change in cellular localization that produces additional S at sites of assembly (Zingler and Littman, 1993; Mammano et al., 1997; Fukushi et al., 2006; Slough et al., 2019). When compared to WT Spike, first research utilizing shortened SARS-CoV-2 S protein with a deletion (Δ) of 8 to 39 amino acids in CT demonstrated a high titer of ppVSVΔG-SARS-CoV-2 S in Δ19 and Δ26 (Giroglou et al., 2004). When the deletions Δ19 and Δ21 into CT were examined, additional investigations revealed a high titer of ppVSVΔG-SARS-CoV-2 S with increased cell-to-cell fusion (Case et al., 2020b; Dieterle et al., 2020; Havranek et al., 2020; Johnson et al., 2020; Schmidt et al., 2020). If necessary, the SARS-CoV-2 S protein can be fused with a C terminal 3XFLAG tag to detect full-length S without interfering with fusion, surface expression, translation, and VSV incorporation of SARS-CoV-2 S protein (Havranek et al., 2020). All findings have proposed the cloning of Met1-S-Δ21 with usage codons for efficient production of SARS-CoV-2 S protein and ppVSV assembly into the optimal cell line (Figure 4B). The obtained ppVSV viral particles are used to infect a cell line previously transfected with the plasmid containing the optimized S gene that will provide S protein for its assembly onto the virus’s surface.
By attaching to a cellular receptor, like the angiotensin-converting enzyme 2 (ACE2), the SARS-CoV-2 S protein promotes viral entry into target cells (Datta et al., 2020; Wrapp et al., 2020). The lungs and other tissues, including the nasal and oral mucosa, vasculature, kidney, heart, gastrointestinal tract, pancreas, and brain, express ACE2 (Gembardt et al., 2005). Thus, cell lines from human and animal origin, such as 293T (human kidney cells), BHK-21 (Syrian hamster kidney cells), Huh-7 (human liver cells), LLC-PK1 (pig kidney cells), MRC-5 (human lung cells), MyDauLu/47.1 [Daubenton’s bat (Myotis daubentonii) lung cells], NIH/3T3 (Mouse embryonic fibroblast cells), RhiLu/1.1 [Halcyon horseshoe bat (Rhinolophus alcyone) lung cells], Vero (African green monkey kidney cells), Calu-3 (human lung cells), Caco-2 (human colon cells), MDBK (cattle kidney cells), MDCKII (Dog kidney cells), A549 (human lung cells), BEAS-2B (human bronchus cells), and NCI-H1299 (human lung cells) have been evaluated for ppVSVΔG-SARS-CoV-2 S entry (Hoffmann et al., 2020). Preliminary investigations revealed that Vero cells were highly susceptible to ppVSVΔG-SARS-CoV-2 S, along with Caco-2 and Calus-3, while other cell lines tested were not efficient for ppVSVΔG-SARS-CoV-2 S entry (Hoffmann et al., 2020). Vero E6 cells were also initially utilized in cell-culture-based infection models for SARS-CoV-1 studies (Keyaerts et al., 2005; Yamate et al., 2005). Several studies have compared different cell lines for ppVSVΔG-SARS-CoV-2-S entry efficiency. When ppVSV-SARS-CoV-2-S was titrated in both Vero E6 and MA104 (Monkey African Green kidney) cell lines, the virus entry was increased (Case et al., 2020b). In addition, transfecting non-susceptible 293T cells with ACE2, Furin, and TMPRSS2 greatly enhanced the entry of ppVSV-SARS-CoV-2-S particles, supporting the fact that ACE2, Furin, and TMPRSS2 are required for optimal spike infectivity of kidney cells (Condor Capcha et al., 2020; Johnson et al., 2020; Xiong et al., 2020).
Although the enzyme-linked immunosorbent test is extensively used for detecting SARS-CoV-2 specific antibodies, it does not offer information on virus-neutralizing antibody titers (Roy et al., 2020). The neutralization test is a technique for determining the presence of neutralizing antibodies. The traditional viral neutralization experiment necessitates the use of a live SARS-CoV-2 virus, which must be handled in a BSL-3 facility. The downside of using a live viral test is that it is labor-intensive, taking up to 4 days to complete (Ogando et al., 2020). To address these challenges, the pseudovirus neutralization assay (PVNA) is a viable option since it can be conducted under BSL-2 settings and is a safe, rapid, and scalable test. ppVSVΔG-SARS-CoV-2 S can be kept for an extended period (≥6 months) with negligible titer loss at −20 or −80, 4°C (4 weeks), or even at room temperature (1 week). Furthermore, ppVSV-SARS-CoV-1-S may withstand many freeze-thaw cycles without decreasing their infectivity (Wright et al., 2009; Molesti et al., 2014). Another advantage of using PVNA is that most ppVSVΔG-SARS-CoV-2 S contain a marker gene that can be detected by a fluorescence or luminescence signal, with a linear correlation between the value of fluorescence or chemiluminescence and the number of infected ppVSVΔG-SARS-CoV-2 S, allowing for easier and more accurate quantification (Case et al., 2020b; Nie et al., 2020a).
Microplate luminometer is a sensitive, ready-to-use device for gene reporter, cell-based, and biochemical experiments in 96-well plates with luminous responses (Alam and Cook, 1990). Fluorescent imaging and quantitation may be assessed using a variety of microscopes that must automatically scan 96-well plates and include quick focusing, image collection, and big data processing (Case et al., 2020b). Flow cytometers can also be used to assess fluorescent-based techniques. ppVSVΔG-SARS-CoV-2 S titration is performed in 96-well plates with 10-fold serial dilution until a total of nine dilutions with six duplicates are obtained (Figure 5; Crawford et al., 2020).
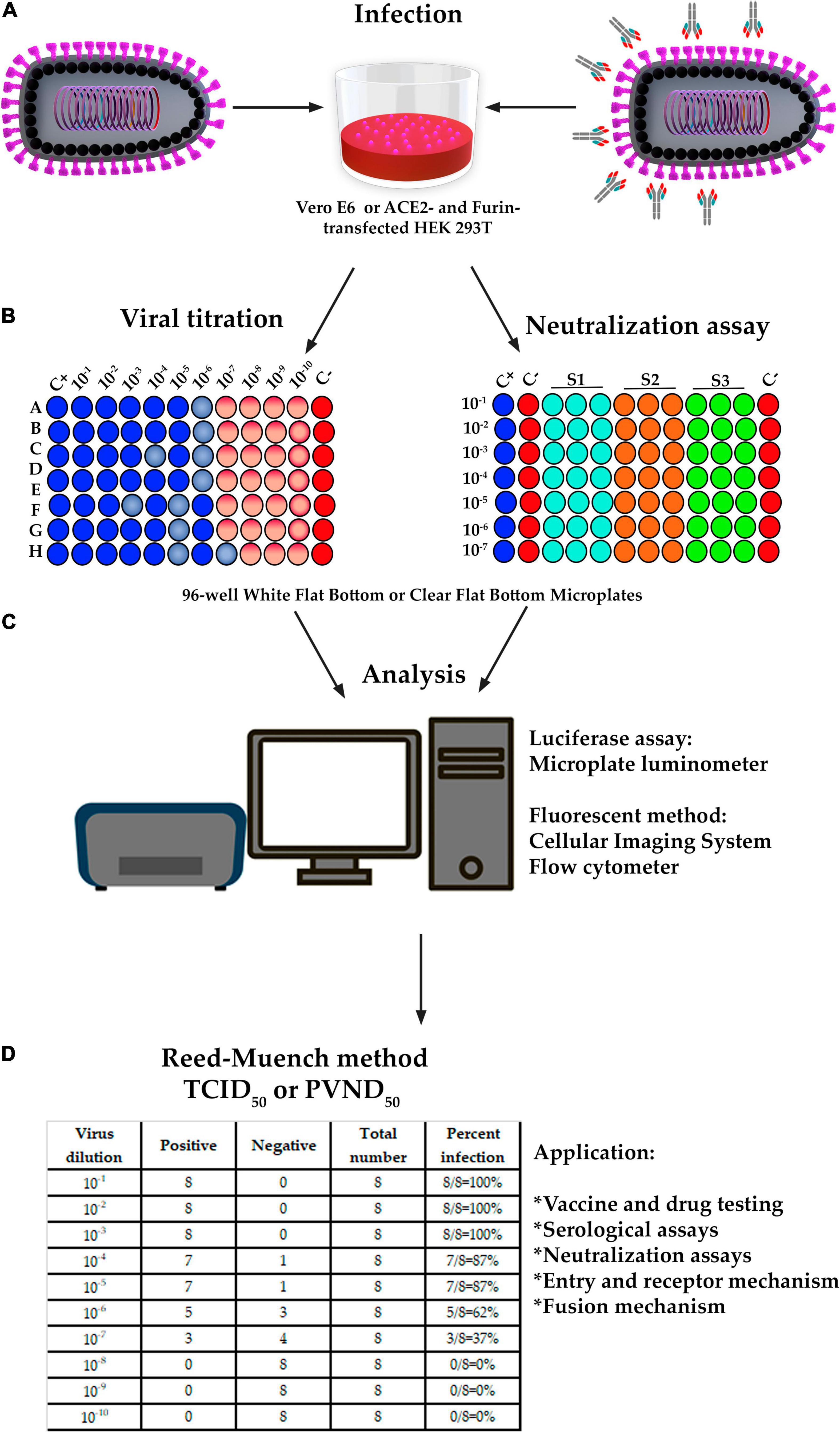
Figure 5. The design of titration and pseudovirus neutralization assay (PVNA). (A,B) Infection, titration and PVNA was performed in a 96-well plate with 10-fold serial dilution of ppVSVΔG-SARS-CoV-2 S until a total of 8 or 10 dilutions and 6 or 8 replicates were obtained. Neutralization assay was also performed in a 96-well plate with 6 dilutions and 2 replicates; however, the sera samples were previously diluted and then mixed with ppVSVΔG-SARS-CoV-2 S 325 to 1,300 TCID50/mL and incubated for 1 h at 37°C. (C) In titration, the cells containing the luciferase were lysed to perform the luciferase assay and a 96-well-plate luminometer was used. In the fluorescence method, the DAPI-stained cells containing GFP were analyzed by fluorescence microscope, and the images were analyzed with specialized software. An alternative method is by using a flow cytometer equipped with a 96-well-autosampler. In PVNA, the luciferase activity was determined by the relative light units (RLU) and the fluorescence by the number of GFP-positive cells. Percent neutralization must be normalized considering uninfected cells as 100% neutralization and infected cells with ppVSVΔG-SARS-CoV-2 S alone as 0% neutralization. (D) The 50% tissue culture infectious dose (TCID50) and/or the 50% ppVSVΔG-SARS-CoV-2 S neutralizing doses (PVND50) were calculated according to the Reed-Muench method and reference table.
The Reed-Muench method is used to determine the 50% tissue culture infectious dose (TCID50) or pseudotyped virus neutralization doses (PVND50) of the ppVSVΔG-SARS-CoV-2 S, which includes calculating the proportional distance (PD) between dilutions above and below the 50% endpoint (Reed, 2010; Gauger and Vincent, 2014; Manenti et al., 2020). The dilution factor is defined as the fold difference between two inoculum titers that are above and below a 50% response rate. Based on the results of Figure 5, the equation must be applied as follows:
Sera samples must be diluted before being combined with ppVSVΔG-SARS-CoV-2 S 325 to 1,300 TCID50/mL and incubated at 37°C for 1 h. Pre-incubated ppVSVΔG-SARS-CoV-2 S are used to infect cell lines for 24–72 h at 37°C and 5% CO2, and the quantification of ppVSVΔG-SARS-CoV-2 S infecting the target cells is estimated by measuring the production of luciferase or fluorescence as stated in the section “Pseudotyped Vesicular Stomatitis Virus-Severe Acute Respiratory Syndrome-Coronavirus-2 Spike titration” (Figure 5B). The luciferase activity is determined by relative light units (RLU), whereas the fluorescence is determined by the number of GFP-positive cells. Percent neutralization must be adjusted by assuming uninfected cells to be 100% neutralized and infected cells with ppVSVΔG-SARS-CoV-2 S alone to be 0% neutralized (Crawford et al., 2020). The PVND50 (50% ppVSVΔG-SARS-CoV-2 S neutralizing doses) is determined as described in the section “Pseudotyped Vesicular Stomatitis Virus-Severe Acute Respiratory Syndrome-Coronavirus-2 Spike Titration” (Reed, 2010; Gauger and Vincent, 2014; Manenti et al., 2020).
For some groups, there is a good connection between the experimental data acquired using authentic SARS-CoV-2 and the PVNA. Neutralizing activity was detected with a remarkable connection between the two tests, as demonstrated by Spearman’s correlation with R values ranging from 0.76 to 0.939 and p values of <0.001 (Case et al., 2020b; Dieterle et al., 2020; Hyseni et al., 2020; Nie et al., 2020a; Xiong et al., 2020; Zettl et al., 2020; Tolah et al., 2021). Furthermore, once a ppVSVΔG-SARS-CoV-2 S stock is produced, the PVNA may be completed in 1 day with a GFP or RLU readout of 7.5 h, whereas the live SARS-CoV-2 assay takes 30 h (Case et al., 2020b; Nie et al., 2020b).
A SARS-CoV-2 variant is identified by alterations in receptor binding, decreased antibody neutralization (post-infection or post-vaccination), decreased treatment effectiveness, or even a possible influence on diagnostics, as well as increased anticipated transmissibility or disease severity (WHO). ppVSVΔG-SARS-CoV-2 S particles can rapidly be produced with the changes in newly define variants and tested for alterations in neutralizing capacity and entry efficiencies.
South Africa was the first to describe the SARS-CoV-2 B.1.351 variety (Beta), which was characterized by N501Y, K417N, and E484K alterations. This variant is more transmissible and resistant to neutralizing antibodies. ppVSVΔG-SARS-CoV-2 S containing the S protein (B.1.351)-associated mutations can quickly infect cell cultures and tolerate neutralizing action of monoclonal antibodies against SARS-CoV-2 S RBD (Kim et al., 2021).
The SARS-CoV-2 B.1.427/B.1.429 variant (epsilon), a novel variant with S13I, W152C, and L452R amino acid changes, was reported for the first time in California (34 nations, beginning May 2021) with enhanced transmissibility and infectivity. When compared to the D614G change, ppVSVΔG-SARS-CoV-2 S bearing the L452R change showed a 6.7- to 22.5-fold higher infectivity in cell cultures and lung organoids, while the W152C change showed just a little increase in infection in these cells. Furthermore, ppVSVΔG-SARS-CoV-2 S with S13I, W152C, and L452R changes demonstrated considerable resistance to neutralization by post-infection (4.0- to 6.7-fold) and vaccination-elicited antibodies (2-fold) as well as monoclonal antibodies (Deng et al., 2021; McCallum et al., 2021).
In the New York region, the SARS-CoV-2 B.1.526 variation (Iota), defined by L5F, T95I, D253G, E484K, or S477N, D614G, and A701V change, was identified. ppVSVΔG-SARS-CoV-2 S harboring L5F, T95I, D253G, E484K, D614G, and A701V, as well as L5F, T95I, D253G, S477N, D614G, and Q957R changes, showed reduced neutralization by post-infection- (2- to 4.5-fold) and vaccine-elicited antibodies (2- to 6-fold) (West et al., 2021).
Recently, a new SARS-CoV-2 VOC was identified in November 2021 and named as Omicron (B.1.1.529) variant by the WHO10. This variant was first detected in Botswana (Gauteng Province) on November 11, 2021, and 3 days later in South Africa. The Omicron variant is characterized by 26–32 changes in the S protein, particularly within the RBD, as well as three deletions and one insertion in the S protein, along with mutations outside of the S protein [Network for Genomic Surveillance in South Africa (NGS-SA)]. Many of these mutations are either known or predicted to contribute not only to increase infectivity and transmissibility, but also to confer therapeutic and neutralization resistance (Schmidt et al., 2021). Indeed, Pulliam et al. (2021), reported that reinfections in South Africa have increased as Omicron has spread. Therefore, studies comparing the SARS-CoV-2 Omicron variant vs. ppVSVΔG-SARS-CoV-2 S are urgently needed to get a better understanding of how the immune system is reacting to this variant.
We propose comparing the SARS-CoV-2 B.1.617.2 (delta) variant to ppVSVΔG-SARS-CoV-2 S with L452R, D614G, and P681R amino acid changes. This variety, which is the most common SARS-CoV-2 strain in Mexico and across the world, necessitates immediate monitoring for vaccination effectiveness. In addition, given the fast spread of the Omicron variant, we also propose to compare the SARS-CoV-2 Omicron variant to ppVSVΔG-SARS-CoV-2 S.
The ongoing development of SARS-CoV-2 variants of concern (VOC) across the world emphasizes the need of monitoring the effectiveness of approved vaccinations for human use. So, far, six very effective COVID-19 vaccines have been approved for human use: Moderna mRNA1273, BioNTech BNT162b2, Janssen Ad26.COV2.S, Gamaleya’s Sputnik V, AstraZeneca’s AZD1222, and CoronaVac. All of these vaccines, albeit in various forms, are based on the Spike protein. Because VOC contains polymorphisms on the S gene, there is an urgent need to evaluate vaccination effectiveness against prevalent SARS-CoV-2 VOC in all geographic areas. As a result, ppVSVΔG-SARS-CoV-2 S have played an important role in determining vaccination effectiveness for SARS-CoV-2 VOC. ppVSVΔG-SARS-CoV-2 S, in particular, has been frequently used in assessing neutralizing antibody titers of vaccinated persons to evaluate if vaccinations provide enough protection against SARS-CoV-2 VOC infection (Table 2; Collier et al., 2021; Ikegame et al., 2021; McCallum et al., 2021; Muik et al., 2021; Shen et al., 2021a,b; Wang G.-L. et al., 2021; Wang P. et al., 2021).
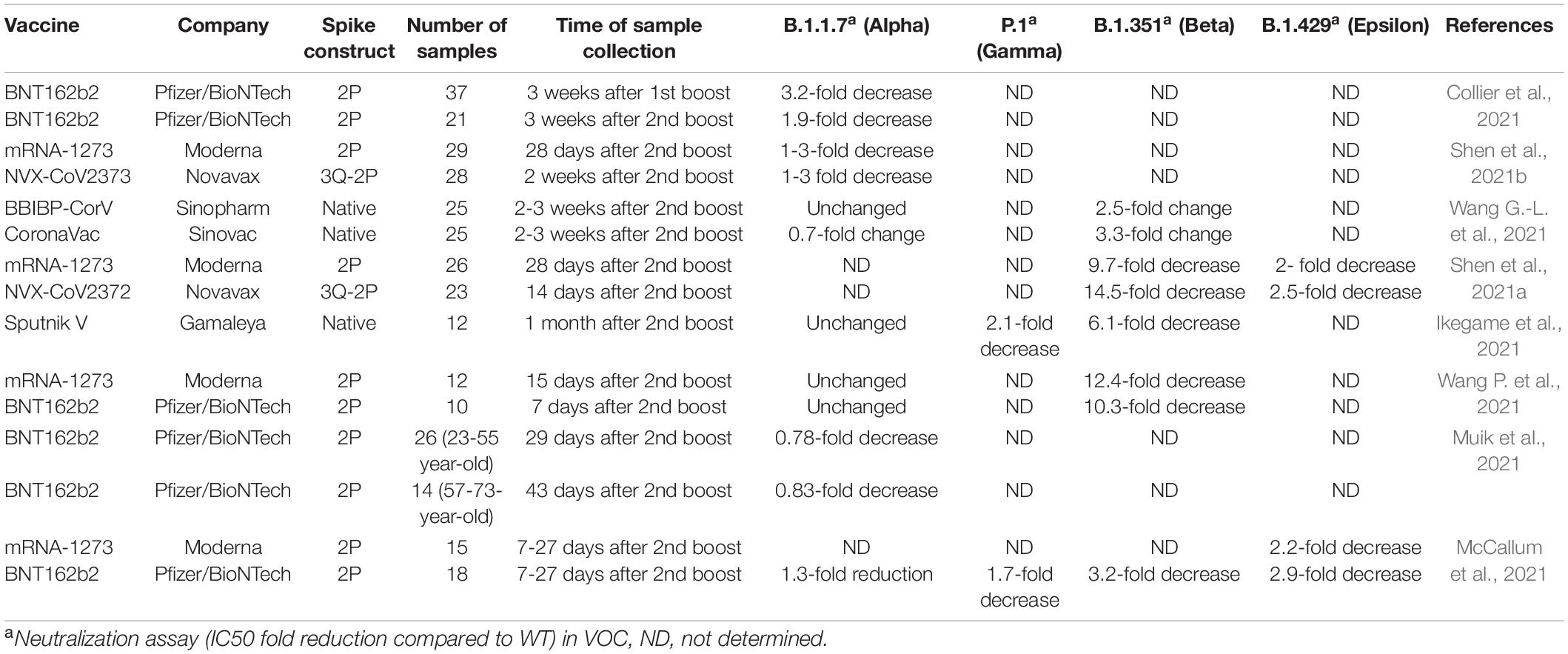
Table 2. Summary of post-vaccine sera evaluated for neutralization potency by using pseudotyped VSV-Severe Acute Respiratory Syndrome-Coronavirus-2 (SARS-CoV-2) Spike variants of concern (VOC).
Overall, the data show that SARS-CoV-2 VOC can reduce neutralization potency in vaccinated people’s sera. To note, the SARS-CoV-2 beta variant exhibited the greatest decrease in PVNAs in sera from persons who had been vaccinated with Moderna, Novavax, or PfizerBioNTech (Table 2). Despite the apparently worrying results obtained in the neutralization tests, it is important to consider that vaccines also induce cell mediated immunity; thus, even when neutralization levels decrease, vaccine effectiveness is still high. As a result, monitoring the neutralizing activity evoked by vaccine sera will be required to assess whether a vaccination update is required to limit the establishment and spread of new SARS-CoV-2 variations, such as the delta and omicron variants. Furthermore, ppVSVΔG-SARS-CoV-2 S are being utilized in the development of a vaccine against SARS-CoV-2. Thus, various VSV-SARS-CoV-2 vaccines have been shown in animal models to be effective in both generating neutralizing antibodies at high titers and protecting against the SARS-CoV-2 challenge (Case et al., 2020a; Yahalom-Ronen et al., 2020; Lu et al., 2021; Malherbe et al., 2021). Although the evidence provided by these vector vaccines is encouraging, it is still early in the research process, and additional study is needed before moving forward with human trials.
Some COVID-19 treatment drugs are directed against the SARS-CoV-2 S protein and its receptor ACE2, which are found on the membranes of different human cells. Through ACE2 attachment, SARS-CoV-2 causes cell-membrane fusion, facilitating viral entrance (Hoffmann et al., 2020). Although preclinical research for effective drugs against SARS-CoV-2 necessitates the use of live SARS-CoV-2, ppVSVΔG-SARS-CoV-2 S can be utilized for S protein-focused therapy evaluation by blocking or down-regulating ACE2 or preventing viral fusion.
MEK inhibitors (VS-6766, trametinib, and selumetinib) have been used to reduce ACE2 cellular expression as a method to prevent early SARS-CoV-2 infection. ppVSVΔG-SARS-CoV-2 S and human bronchial epithelial, small airway epithelial, and lung cancer cells were utilized to assess infectivity alterations caused by MEK inhibitors (Zhou et al., 2020). Furthermore, PVs were used to test the creation of fusion inhibitor peptides against SARS-CoV-2, and micromolar concentrations of peptides suppressed ppVSVΔG-SARS-CoV-2 S infection by inhibiting viral fusion (Kandeel et al., 2021). Demethylzeylasteral, another inhibiting drug, can interact with hACE2 and the RBD of the SARS-CoV-2 S protein, thus when tested in ppVSVΔG-SARS-CoV-2 S, it inhibited ppVSVΔG-SARS-CoV-2 S entrance into 293T cells (Zhu et al., 2020). Polyunsaturated-3 fatty acids limit SARS-CoV-2 binding and cellular entrance, while linolenic and eicosapentaenoic acids prevent ppVSVΔG-SARS-CoV-2 S penetration (Goc et al., 2021). Inhalable nano catchers containing hACE2 are a proposal for SARS-CoV-2 suppression, which was tested with ppVSVΔG-SARS-CoV-2 S and shown a strong capability for infection inhibition in a hACE2-expressing mouse model (Zhang et al., 2021). Furthermore, when tested with ppVSVΔG-SARS-CoV-2 S and live SARS-CoV-2, hACE2-Fc inhibited Vero E6 cells (Case et al., 2020b).
There are only seven BSL-3 laboratories in Mexico, four of which are in Mexico City. Because of the scarcity of BSL-3 facilities, research into existing and newly emerging infectious diseases are hampered. Furthermore, there is no BSL-4 laboratory in Mexico, making it impossible to research the viruses that are classified as risk group 4 and are designated priority diseases by the WHO. This is an invitation to the Mexican government to develop research policies and infrastructure for the construction of BSL3 and BSL4 facilities. ppVSVΔG-SARS-CoV-2 S is an excellent choice for studies of host and cellular tropism, interspecies transmission, viral pathogenesis, and host cell entry pathways with risk group 3 and 4 viruses such as SARS-CoV-2. It can be used in BSL-2 facilities for studies of host and cellular tropism, interspecies transmission, viral pathogenesis, and host cell entry pathways. ppVSVΔG-SARS-CoV-2 S can be supplemented with SARS-CoV-2 S protein via changes that improve expression efficiency, and we propose the cloning of Met1-S-Δ21 with usage codons. Once the S gen has been refined and cloned, we propose using site-directed mutagenesis to generate novel variations of the SARS-CoV-2 S protein. ppVSVΔG-G and ppVSVΔG-SARS-CoV-2 S particles must be produced in HEK 293T cells, and infection assays with ppVSVΔG-SARS-CoV-2 S in Vero E6 or ACE2- and Furin-transfected HEK 293T cells are suggested. The ppVSV-SARS-CoV-1-S are stable once produced, and they may be kept with low activity loss for up to 6 months (−20 or −80°C), 4 weeks (4°C), and just 1 week (room temperature) with up to four freeze-thaw cycles. PVNA, surprisingly, is a safe (BSL-2), efficient (7.5 h), and scalable (r ≥ 0.9 and p ≤ 0.001) method that may be utilized to evaluate novel SARS-CoV-2 variants, post-infection- and vaccine-elicited neutralizing antibodies, and S protein-based COVID-19 therapies. ppVSVΔG-SARS-CoV-2 S can be detected by a fluorescence or luminescence signal with a linear correlation between the value of fluorescence or chemiluminescence and the number of infective ppVSVΔG-SARS-CoV-2 S, and these particles can be easily calculated using the TCID50/mL or PVID50 equations, which are included in this review. We are the first Mexican organization to employ ppVSVΔG-SARS-CoV-2 S for preclinical assessment of post-infection-neutralizing antibodies, evaluation of a possible COVID-19 vaccine and decontamination equipment, and evaluation of vaccinated volunteers against SARS-CoV-2 delta variants.
VL-P and MS-G: conceptualization, resources, project administration, and funding acquisition. AC-R and AF-A: software and formal analysis. SA-C, GP-L, GR-M, AC-R, and VL-P: investigation. VL-P, GP-L, and GR-M: writing—original draft preparation. VL-P, GP-L, and MS-G: writing—review and editing. SA-C, AC-R, and AF-A: visualization. VL-P: supervision. All authors have read and agreed to the published version of the manuscript.
This review was funded by the Public Federal Funds from HIMFG with grant numbers HIM/2020/029 SSA. 1664, HIM/2020/060 SSA. 1686, and HIM/2021/007.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We want to acknowledge the research department of HIMFG for the support given in this review.
Alam, J., and Cook, J. L. (1990). Reporter genes: application to the study of mammalian gene transcription. Anal. Biochem. 188, 245–254. doi: 10.1016/0003-2697(90)90601-5
Artika, I. M., and Ma’roef, C. N. (2017). Laboratory biosafety for handling emerging viruses. Asian Pac. J. Trop. Biomed. 7, 483–491. doi: 10.1016/j.apjtb.2017.01.020
Barrett, C. T., and Dutch, R. E. (2020). Viral membrane fusion and the transmembrane domain. Viruses 12:E693. doi: 10.3390/v12070693
Blumenthal, R., Durell, S., and Viard, M. (2012). HIV entry and envelope glycoprotein-mediated fusion. J. Biol. Chem. 287, 40841–40849. doi: 10.1074/jbc.R112.406272
Bukbuk, D. N., Fukushi, S., Tani, H., Yoshikawa, T., Taniguchi, S., Iha, K., et al. (2014). Development and validation of serological assays for viral hemorrhagic fevers and determination of the prevalence of Rift Valley fever in Borno State, Nigeria. Trans. R. Soc. Trop. Med. Hyg. 108, 768–773. doi: 10.1093/trstmh/tru163
Case, J. B., Rothlauf, P. W., Chen, R. E., Kafai, N. M., Fox, J. M., Smith, B. K., et al. (2020a). Replication-competent vesicular stomatitis virus vaccine vector protects against SARS-CoV-2-mediated pathogenesis in mice. Cell Host Microbe 28, 465–474.e4. doi: 10.1016/j.chom.2020.07.018
Case, J. B., Rothlauf, P. W., Chen, R. E., Liu, Z., Zhao, H., Kim, A. S., et al. (2020b). Neutralizing antibody and soluble ACE2 inhibition of a replication-competent VSV-SARS-CoV-2 and a clinical isolate of SARS-CoV-2. Cell Host Microbe 28, 475–485.e5. doi: 10.1016/j.chom.2020.06.021
Chang, K. W., Sheng, Y., and Gombold, J. L. (2000). Coronavirus-induced membrane fusion requires the cysteine-rich domain in the spike protein. Virology 269, 212–224. doi: 10.1006/viro.2000.0219
Choppin, P. W., and Compans, R. W. (1970). Phenotypic mixing of envelope proteins of the parainfluenza virus SV5 and vesicular stomatitis virus. J. Virol. 5, 609–616. doi: 10.1128/JVI.5.5.609-616.1970
Collier, D. A., De Marco, A., Ferreira, I. A. T. M., Meng, B., Datir, R. P., Walls, A. C., et al. (2021). Sensitivity of SARS-CoV-2 B.1.1.7 to mRNA vaccine-elicited antibodies. Nature 593, 136–141. doi: 10.1038/s41586-021-03412-7
Condor Capcha, J. M., Lambert, G., Dykxhoorn, D. M., Salerno, A. G., Hare, J. M., Whitt, M. A., et al. (2020). Generation of SARS-CoV-2 spike pseudotyped virus for viral entry and neutralization assays: a 1-week protocol. Front. Cardiovasc. Med. 7:618651. doi: 10.3389/fcvm.2020.618651
Contreras, E. M., Johnston, G. P., Buchholz, D. W., Ortega, V., Monreal, I. A., Zamora, J. L. R., et al. (2021). Roles of cholesterol in early and late steps of the nipah virus membrane fusion cascade. J. Virol. 95, e02323–20. doi: 10.1128/JVI.02323-20
Crawford, K. H. D., Eguia, R., Dingens, A. S., Loes, A. N., Malone, K. D., Wolf, C. R., et al. (2020). Protocol and reagents for pseudotyping lentiviral particles with SARS-CoV-2 Spike protein for neutralization assays. Viruses 12:E513. doi: 10.3390/v12050513
Datta, P. K., Liu, F., Fischer, T., Rappaport, J., and Qin, X. (2020). SARS-CoV-2 pandemic and research gaps: understanding SARS-CoV-2 interaction with the ACE2 receptor and implications for therapy. Theranostics 10, 7448–7464. doi: 10.7150/thno.48076
Deng, X., Garcia-Knight, M. A., Khalid, M. M., Servellita, V., Wang, C., Morris, M. K., et al. (2021). Transmission, infectivity, and neutralization of a spike L452R SARS-CoV-2 variant. Cell 184, 3426–3437.e8. doi: 10.1016/j.cell.2021.04.025
Dessau, M., and Modis, Y. (2013). Crystal structure of glycoprotein C from Rift Valley fever virus. Proc. Natl. Acad. Sci. U.S.A. 110, 1696–1701. doi: 10.1073/pnas.1217780110
Dieterle, M. E., Haslwanter, D., Bortz, R. H., Wirchnianski, A. S., Lasso, G., Vergnolle, O., et al. (2020). A replication-competent vesicular stomatitis virus for studies of SARS-CoV-2 spike-mediated cell entry and its inhibition. Cell Host Microbe 28, 486–496.e6. doi: 10.1016/j.chom.2020.06.020
Fukuma, A., Tani, H., Taniguchi, S., Shimojima, M., Saijo, M., and Fukushi, S. (2015). Inability of rat DPP4 to allow MERS-CoV infection revealed by using a VSV pseudotype bearing truncated MERS-CoV spike protein. Arch. Virol. 160, 2293–2300. doi: 10.1007/s00705-015-2506-z
Fukushi, S., Mizutani, T., Saijo, M., Kurane, I., Taguchi, F., Tashiro, M., et al. (2006). Evaluation of a novel vesicular stomatitis virus pseudotype-based assay for detection of neutralizing antibody responses to SARS-CoV. J. Med. Virol. 78, 1509–1512. doi: 10.1002/jmv.20732
Fukushi, S., Watanabe, R., and Taguchi, F. (2008). Pseudotyped vesicular stomatitis virus for analysis of virus entry mediated by SARS coronavirus spike proteins. Methods Mol. Biol. 454, 331–338. doi: 10.1007/978-1-59745-181-9_23
Gasbarri, M., V’kovski, P., Torriani, G., Thiel, V., Stellacci, F., Tapparel, C., et al. (2020). SARS-CoV-2 inhibition by sulfonated compounds. Microorganisms 8:E1894. doi: 10.3390/microorganisms8121894
Gauger, P. C., and Vincent, A. L. (2014). Serum virus neutralization assay for detection and quantitation of serum-neutralizing antibodies to influenza A virus in swine. Methods Mol. Biol. 1161, 313–324. doi: 10.1007/978-1-4939-0758-8_26
Ge, J., Wen, Z., Wang, X., Hu, S., Liu, Y., Kong, X., et al. (2006). Generating vesicular stomatitis virus pseudotype bearing the severe acute respiratory syndrome coronavirus spike envelope glycoprotein for rapid and safe neutralization test or cell-entry assay. Ann. N. Y. Acad. Sci. 1081, 246–248. doi: 10.1196/annals.1373.030
Geisbert, T. W., and Feldmann, H. (2011). Recombinant vesicular stomatitis virus–based vaccines against Ebola and Marburg virus infections. J. Infect. Dis. 204, S1075–S1081. doi: 10.1093/infdis/jir349
Gembardt, F., Sterner-Kock, A., Imboden, H., Spalteholz, M., Reibitz, F., Schultheiss, H.-P., et al. (2005). Organ-specific distribution of ACE2 mRNA and correlating peptidase activity in rodents. Peptides 26, 1270–1277. doi: 10.1016/j.peptides.2005.01.009
Giroglou, T., Cinatl, J., Rabenau, H., Drosten, C., Schwalbe, H., Doerr, H. W., et al. (2004). Retroviral vectors pseudotyped with severe acute respiratory syndrome coronavirus S protein. J. Virol. 78, 9007–9015. doi: 10.1128/JVI.78.17.9007-9015.2004
Goc, A., Niedzwiecki, A., and Rath, M. (2021). Polyunsaturated ω-3 fatty acids inhibit ACE2-controlled SARS-CoV-2 binding and cellular entry. Sci. Rep. 11:5207. doi: 10.1038/s41598-021-84850-1
Hastie, K. M., Zandonatti, M. A., Kleinfelter, L. M., Heinrich, M. L., Rowland, M. M., Chandran, K., et al. (2017). Structural basis for antibody-mediated neutralization of Lassa virus. Science 356, 923–928. doi: 10.1126/science.aam7260
Havranek, K. E., Jimenez, A. R., Acciani, M. D., Lay Mendoza, M. F., Reyes Ballista, J. M., Diaz, D. A., et al. (2020). SARS-CoV-2 spike alterations enhance pseudoparticle titers and replication-competent VSV-SARS-CoV-2 virus. Viruses 12:1465. doi: 10.3390/v12121465
Hoffmann, M., Kleine-Weber, H., Schroeder, S., Krüger, N., Herrler, T., Erichsen, S., et al. (2020). SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 181, 271–280.e8. doi: 10.1016/j.cell.2020.02.052
Huang, A. S., Palma, E. L., Hewlett, N., and Roizman, B. (1974). Pseudotype formation between enveloped RNA and DNA viruses. Nature 252, 743–745. doi: 10.1038/252743a0
Hyseni, I., Molesti, E., Benincasa, L., Piu, P., Casa, E., Temperton, N. J., et al. (2020). Characterisation of SARS-CoV-2 lentiviral pseudotypes and correlation between pseudotype-based neutralisation assays and live virus-based micro neutralisation assays. Viruses 12:E1011. doi: 10.3390/v12091011
Ikegame, S., Siddiquey, M. N. A., Hung, C.-T., Haas, G., Brambilla, L., Oguntuyo, K. Y., et al. (2021). Neutralizing activity of Sputnik V vaccine sera against SARS-CoV-2 variants. Nat. Commun. 12:4598. doi: 10.1038/s41467-021-24909-9
Johnson, M. C., Lyddon, T. D., Suarez, R., Salcedo, B., LePique, M., Graham, M., et al. (2020). Optimized pseudotyping conditions for the SARS-COV-2 spike glycoprotein. J. Virol. 94, e01062–20. doi: 10.1128/JVI.01062-20
Kaku, Y., Noguchi, A., Marsh, G. A., Barr, J. A., Okutani, A., Hotta, K., et al. (2012). Second generation of pseudotype-based serum neutralization assay for Nipah virus antibodies: sensitive and high-throughput analysis utilizing secreted alkaline phosphatase. J. Virol. Methods 179, 226–232. doi: 10.1016/j.jviromet.2011.11.003
Kaku, Y., Noguchi, A., Marsh, G. A., McEachern, J. A., Okutani, A., Hotta, K., et al. (2009). A neutralization test for specific detection of Nipah virus antibodies using Pseudotyped Vesicular Stomatitis virus expressing green fluorescent protein. J. Virol. Methods 160, 7–13. doi: 10.1016/j.jviromet.2009.04.037
Kandeel, M., Yamamoto, M., Tani, H., Kobayashi, A., Gohda, J., Kawaguchi, Y., et al. (2021). Discovery of new fusion inhibitor peptides against SARS-CoV-2 by targeting the Spike S2 subunit. Biomol. Ther. (Seoul) 29, 282–289. doi: 10.4062/biomolther.2020.201
Kapadia, S. U., Simon, I. D., and Rose, J. K. (2008). SARS vaccine based on a replication-defective recombinant vesicular stomatitis virus is more potent than one based on a replication-competent vector. Virology 376, 165–172. doi: 10.1016/j.virol.2008.03.002
Kaufer, A. M., Theis, T., Lau, K. A., Gray, J. L., and Rawlinson, W. D. (2020). Laboratory biosafety measures involving SARS-CoV-2 and the classification as a Risk Group 3 biological agent. Pathology 52, 790–795. doi: 10.1016/j.pathol.2020.09.006
Keyaerts, E., Vijgen, L., Maes, P., Neyts, J., and Van Ranst, M. (2005). Growth kinetics of SARS-coronavirus in Vero E6 cells. Biochem. Biophys. Res. Commun. 329, 1147–1151. doi: 10.1016/j.bbrc.2005.02.085
Kieny, M. P., Rottingen, J.-A., and Farrar, J. (2016). The need for global R&D coordination for infectious diseases with epidemic potential. Lancet 388, 460–461. doi: 10.1016/S0140-6736(16)31152-7
Kim, I. S., Jenni, S., Stanifer, M. L., Roth, E., Whelan, S. P. J., van Oijen, A. M., et al. (2017). Mechanism of membrane fusion induced by vesicular stomatitis virus G protein. Proc. Natl. Acad. Sci. U.S.A. 114, E28–E36. doi: 10.1073/pnas.1618883114
Kim, Y. J., Jang, U. S., Soh, S. M., Lee, J.-Y., and Lee, H.-R. (2021). The impact on infectivity and neutralization efficiency of SARS-CoV-2 lineage B.1.351 pseudovirus. Viruses 13:633. doi: 10.3390/v13040633
Korber, B., Fischer, W. M., Gnanakaran, S., Yoon, H., Theiler, J., Abfalterer, W., et al. (2020). Tracking changes in SARS-CoV-2 spike: evidence that D614G increases infectivity of the COVID-19 virus. Cell 182, 812–827.e19. doi: 10.1016/j.cell.2020.06.043
Kudla, G., Lipinski, L., Caffin, F., Helwak, A., and Zylicz, M. (2006). High guanine and cytosine content increases mRNA levels in mammalian cells. PLoS Biol. 4:e180. doi: 10.1371/journal.pbio.0040180
Kunz, S., Rojek, J. M., Perez, M., Spiropoulou, C. F., and Oldstone, M. B. A. (2005). Characterization of the interaction of Lassa fever virus with its cellular receptor alpha-dystroglycan. J. Virol. 79, 5979–5987. doi: 10.1128/JVI.79.10.5979-5987.2005
Lawson, N. D., Stillman, E. A., Whitt, M. A., and Rose, J. K. (1995). Recombinant vesicular stomatitis viruses from DNA. Proc. Natl. Acad. Sci. U.S.A. 92, 4477–4481. doi: 10.1073/pnas.92.10.4477
Lee, J. E., Fusco, M. L., Hessell, A. J., Oswald, W. B., Burton, D. R., and Saphire, E. O. (2008). Structure of the Ebola virus glycoprotein bound to an antibody from a human survivor. Nature 454, 177–182. doi: 10.1038/nature07082
Lennemann, N. J., Herbert, A. S., Brouillette, R., Rhein, B., Bakken, R. A., Perschbacher, K. J., et al. (2017). Vesicular Stomatitis Virus Pseudotyped with Ebola Virus glycoprotein serves as a protective, noninfectious vaccine against Ebola virus challenge in mice. J. Virol. 91, e00479–17. doi: 10.1128/JVI.00479-17
Lester, S., Harcourt, J., Whitt, M., Al-Abdely, H. M., Midgley, C. M., Alkhamis, A. M., et al. (2019). Middle East respiratory coronavirus (MERS-CoV) spike (S) protein vesicular stomatitis virus pseudoparticle neutralization assays offer a reliable alternative to the conventional neutralization assay in human seroepidemiological studies. Access Microbiol. 1:e000057. doi: 10.1099/acmi.0.000057
Liu, R., Wang, J., Shao, Y., Wang, X., Zhang, H., Shuai, L., et al. (2018). A recombinant VSV-vectored MERS-CoV vaccine induces neutralizing antibody and T cell responses in rhesus monkeys after single dose immunization. Antiviral Res. 150, 30–38. doi: 10.1016/j.antiviral.2017.12.007
Lodish, H. F., and Weiss, R. A. (1979). Selective isolation of mutants of Vesicular Stomatitis virus defective in production of the viral glycoprotein. J. Virol. 30, 177–189. doi: 10.1128/JVI.30.1.177-189.1979
Lontok, E., Corse, E., and Machamer, C. E. (2004). Intracellular targeting signals contribute to localization of Coronavirus spike proteins near the virus assembly site. J. Virol. 78, 5913–5922. doi: 10.1128/JVI.78.11.5913-5922.2004
Lu, M., Zhang, Y., Dravid, P., Li, A., Zeng, C., Kc, M., et al. (2021). A methyltransferase-defective VSV-based SARS-CoV-2 vaccine candidate provides complete protection against SARS-CoV-2 infection in hamsters. J. Virol. 95:e0059221. doi: 10.1128/JVI.00592-21
Lun, C. M., Waheed, A. A., Majadly, A., Powell, N., and Freed, E. O. (2021). Mechanism of viral glycoprotein targeting by membrane-associated RING-CH proteins. mBio 12, e219–e221. doi: 10.1128/mBio.00219-21
Malherbe, D. C., Kurup, D., Wirblich, C., Ronk, A. J., Mire, C., Kuzmina, N., et al. (2021). A single dose of replication-competent VSV-vectored vaccine expressing SARS-CoV-2 S1 protects against virus replication in a hamster model of severe COVID-19. NPJ Vaccines 6:91. doi: 10.1038/s41541-021-00352-1
Mammano, F., Salvatori, F., Indraccolo, S., De Rossi, A., Chieco-Bianchi, L., and Göttlinger, H. G. (1997). Truncation of the human immunodeficiency virus type 1 envelope glycoprotein allows efficient pseudotyping of Moloney Murine Leukemia virus particles and gene transfer into CD4+ cells. J. Virol. 71, 3341–3345. doi: 10.1128/JVI.71.4.3341-3345.1997
Manenti, A., Maggetti, M., Casa, E., Martinuzzi, D., Torelli, A., Trombetta, C. M., et al. (2020). Evaluation of SARS-CoV-2 neutralizing antibodies using of a CPE-based Colorimetric live virus micro-neutralization assay in human serum samples. J. Med. Virol. 92, 2096–2104. doi: 10.1002/jmv.25986
Mauro, V. P., and Chappell, S. A. (2014). A critical analysis of codon optimization in human therapeutics. Trends Mol. Med. 20, 604–613. doi: 10.1016/j.molmed.2014.09.003
McBride, C. E., Li, J., and Machamer, C. E. (2007). The cytoplasmic tail of the severe acute respiratory syndrome coronavirus spike protein contains a novel endoplasmic reticulum retrieval signal that binds COPI and promotes interaction with membrane protein. J. Virol. 81, 2418–2428. doi: 10.1128/JVI.02146-06
McCallum, M., Bassi, J., De Marco, A., Chen, A., Walls, A. C., Di Iulio, J., et al. (2021). SARS-CoV-2 immune evasion by the B.1.427/B.1.429 variant of concern. Science 373, 648–654. doi: 10.1126/science.abi7994
Mendenhall, A., Lesnik, J., Mukherjee, C., Antes, T., and Sengupta, R. (2012). Packaging HIV- or FIV-based lentivector expression constructs and transduction of VSV-G pseudotyped viral particles. J. Vis. Exp. 63:e3171. doi: 10.3791/3171
Millet, J. K., Tang, T., Nathan, L., Jaimes, J. A., Hsu, H.-L., Daniel, S., et al. (2019). Production of pseudotyped particles to study highly pathogenic coronaviruses in a biosafety level 2 setting. J. Vis. Exp. doi: 10.3791/59010
Molesti, E., Wright, E., Terregino, C., Rahman, R., Cattoli, G., and Temperton, N. J. (2014). Multiplex evaluation of influenza neutralizing antibodies with potential applicability to in-field serological studies. J. Immunol. Res. 2014:457932. doi: 10.1155/2014/457932
Moore, M. J., Dorfman, T., Li, W., Wong, S. K., Li, Y., Kuhn, J. H., et al. (2004). Retroviruses pseudotyped with the severe acute respiratory syndrome coronavirus spike protein efficiently infect cells expressing angiotensin-converting enzyme 2. J. Virol. 78, 10628–10635. doi: 10.1128/JVI.78.19.10628-10635.2004
Morens, D. M., and Fauci, A. S. (2020). Emerging pandemic diseases: how we got to COVID-19. Cell 182, 1077–1092. doi: 10.1016/j.cell.2020.08.021
Morens, D. M., Folkers, G. K., and Fauci, A. S. (2008). Emerging infections: a perpetual challenge. Lancet Infect. Dis. 8, 710–719. doi: 10.1016/S1473-3099(08)70256-1
Muik, A., Dold, C., Geiß, Y., Volk, A., Werbizki, M., Dietrich, U., et al. (2012). Semireplication-competent Vesicular Stomatitis virus as a novel platform for oncolytic virotherapy. J. Mol. Med. (Berl.) 90, 959–970. doi: 10.1007/s00109-012-0863-6
Muik, A., Wallisch, A.-K., Sänger, B., Swanson, K. A., Mühl, J., Chen, W., et al. (2021). Neutralization of SARS-CoV-2 lineage B.1.1.7 pseudovirus by BNT162b2 vaccine-elicited human sera. Science 371, 1152–1153. doi: 10.1126/science.abg6105
Muñoz-Barroso, I., Salzwedel, K., Hunter, E., and Blumenthal, R. (1999). Role of the membrane-proximal domain in the initial stages of human immunodeficiency virus type 1 envelope glycoprotein-mediated membrane fusion. J. Virol. 73, 6089–6092. doi: 10.1128/JVI.73.7.6089-6092.1999
Nie, J., Li, Q., Wu, J., Zhao, C., Hao, H., Liu, H., et al. (2020a). Establishment and validation of a pseudovirus neutralization assay for SARS-CoV-2. Emerg. Microbes Infect. 9, 680–686. doi: 10.1080/22221751.2020.1743767
Nie, J., Li, Q., Wu, J., Zhao, C., Hao, H., Liu, H., et al. (2020b). Quantification of SARS-CoV-2 neutralizing antibody by a pseudotyped virus-based assay. Nat. Protoc. 15, 3699–3715. doi: 10.1038/s41596-020-0394-5
Ogando, N. S., Dalebout, T. J., Zevenhoven-Dobbe, J. C., Limpens, R. W. A. L., van der Meer, Y., Caly, L., et al. (2020). SARS-coronavirus-2 replication in Vero E6 cells: replication kinetics, rapid adaptation and cytopathology. J. Gen. Virol. 101, 925–940. doi: 10.1099/jgv.0.001453
Pallesen, J., Wang, N., Corbett, K. S., Wrapp, D., Kirchdoerfer, R. N., Turner, H. L., et al. (2017). Immunogenicity and structures of a rationally designed prefusion MERS-CoV spike antigen. Proc. Natl. Acad. Sci. U.S.A. 114, E7348–E7357. doi: 10.1073/pnas.1707304114
Pattnaik, A. K., Ball, L. A., LeGrone, A. W., and Wertz, G. W. (1992). Infectious defective interfering particles of VSV from transcripts of a cDNA clone. Cell 69, 1011–1020. doi: 10.1016/0092-8674(92)90619-n
Pattnaik, A. K., and Wertz, G. W. (1990). Replication and amplification of defective interfering particle RNAs of vesicular stomatitis virus in cells expressing viral proteins from vectors containing cloned cDNAs. J. Virol. 64, 2948–2957. doi: 10.1128/JVI.64.6.2948-2957.1990
Petit, C. M., Chouljenko, V. N., Iyer, A., Colgrove, R., Farzan, M., Knipe, D. M., et al. (2007). Palmitoylation of the cysteine-rich endodomain of the SARS-coronavirus spike glycoprotein is important for spike-mediated cell fusion. Virology 360, 264–274. doi: 10.1016/j.virol.2006.10.034
Plescia, C. B., David, E. A., Patra, D., Sengupta, R., Amiar, S., Su, Y., et al. (2021). SARS-CoV-2 viral budding and entry can be modeled using BSL-2 level virus-like particles. J. Biol. Chem. 296:100103. doi: 10.1074/jbc.RA120.016148
Pulliam, J. R. C., van Schalkwyk, C., Govender, N., von Gottberg, A., Cohen, C., Groome, M. J., et al. (2021). Increased risk of SARS-CoV-2 reinfection associated with emergence of the Omicron variant in South Africa. medRxiv [Preprint]. doi: 10.1101/2021.11.11.21266068
Reed, N. G. (2010). The history of ultraviolet germicidal irradiation for air disinfection. Public Health Rep. 125, 15–27. doi: 10.1177/003335491012500105
Rodriguez, S. E., Cross, R. W., Fenton, K. A., Bente, D. A., Mire, C. E., and Geisbert, T. W. (2019). Vesicular Stomatitis virus-based vaccine protects mice against Crimean-Congo hemorrhagic fever. Sci. Rep. 9:7755. doi: 10.1038/s41598-019-44210-6
Roy, V., Fischinger, S., Atyeo, C., Slein, M., Loos, C., Balazs, A., et al. (2020). SARS-CoV-2-specific ELISA development. J. Immunol. Methods 484–485:112832. doi: 10.1016/j.jim.2020.112832
Saito, T., Maruyama, J., Nagata, N., Isono, M., Okuya, K., Takadate, Y., et al. (2020). A surrogate animal model for screening of Ebola and Marburg Glycoprotein-targeting drugs using pseudotyped Vesicular Stomatitis viruses. Viruses 12:E923. doi: 10.3390/v12090923
Schmidt, F., Weisblum, Y., Muecksch, F., Hoffmann, H.-H., Michailidis, E., Lorenzi, J. C. C., et al. (2020). Measuring SARS-CoV-2 neutralizing antibody activity using pseudotyped and chimeric viruses. J. Exp. Med. 217:e20201181. doi: 10.1084/jem.20201181
Schmidt, F., Weisblum, Y., Rutkowska, M., Poston, D., DaSilva, J., Zhang, F., et al. (2021). High genetic barrier to SARS-CoV-2 polyclonal neutralizing antibody escape. Nature 600, 512–516. doi: 10.1038/s41586-021-04005-0
Schnell, M. J., Buonocore, L., Kretzschmar, E., Johnson, E., and Rose, J. K. (1996). Foreign glycoproteins expressed from recombinant vesicular stomatitis viruses are incorporated efficiently into virus particles. Proc. Natl. Acad. Sci. U.S.A. 93, 11359–11365. doi: 10.1073/pnas.93.21.11359
Schnitzer, T. J., Dickson, C., and Weiss, R. A. (1979). Morphological and biochemical characterization of viral particles produced by the tsO45 mutant of vesicular stomatitis virus at restrictive temperature. J. Virol. 29, 185–195. doi: 10.1128/JVI.29.1.185-195.1979
Schnitzer, T. J., and Gonczol, E. (1979). Phenotypic mixing between murine oncoviruses and murine cytomegalovirus. J. Gen. Virol. 43, 691–695. doi: 10.1099/0022-1317-43-3-691
Shen, X., Tang, H., McDanal, C., Wagh, K., Fischer, W., Theiler, J., et al. (2021a). SARS-CoV-2 variant B.1.1.7 is susceptible to neutralizing antibodies elicited by ancestral spike vaccines. Cell Host Microbe 29, 529–539.e3. doi: 10.1016/j.chom.2021.03.002
Shen, X., Tang, H., Pajon, R., Smith, G., Glenn, G. M., Shi, W., et al. (2021b). Neutralization of SARS-CoV-2 Variants B.1.429 and B.1.351. N. Engl. J. Med. 384, 2352–2354. doi: 10.1056/NEJMc2103740
Shirato, K., Maejima, M., Matsuyama, S., Ujike, M., Miyazaki, A., Takeyama, N., et al. (2011). Mutation in the cytoplasmic retrieval signal of porcine epidemic diarrhea virus spike (S) protein is responsible for enhanced fusion activity. Virus Res. 161, 188–193. doi: 10.1016/j.virusres.2011.07.019
Slough, M. M., Chandran, K., and Jangra, R. K. (2019). Two Point mutations in old world hantavirus glycoproteins afford the generation of highly infectious recombinant Vesicular Stomatitis virus vectors. mBio 10, e02372–18. doi: 10.1128/mBio.02372-18
Stillman, E. A., Rose, J. K., and Whitt, M. A. (1995). Replication and amplification of novel vesicular stomatitis virus minigenomes encoding viral structural proteins. J. Virol. 69, 2946–2953. doi: 10.1128/JVI.69.5.2946-2953.1995
Suda, Y., Fukushi, S., Tani, H., Murakami, S., Saijo, M., Horimoto, T., et al. (2016). Analysis of the entry mechanism of Crimean-Congo hemorrhagic fever virus, using a Vesicular Stomatitis virus pseudotyping system. Arch. Virol. 161, 1447–1454. doi: 10.1007/s00705-016-2803-1
Tada, T., Zhang, Y., Koyama, T., Tobiume, M., Tsunetsugu-Yokota, Y., Yamaoka, S., et al. (2015). MARCH8 inhibits HIV-1 infection by reducing virion incorporation of envelope glycoproteins. Nat. Med. 21, 1502–1507. doi: 10.1038/nm.3956
Takada, A., Robison, C., Goto, H., Sanchez, A., Murti, K. G., Whitt, M. A., et al. (1997). A system for functional analysis of Ebola virus glycoprotein. Proc. Natl. Acad. Sci. U.S.A. 94, 14764–14769. doi: 10.1073/pnas.94.26.14764
Tamin, A., Harcourt, B. H., Lo, M. K., Roth, J. A., Wolf, M. C., Lee, B., et al. (2009). Development of a neutralization assay for Nipah virus using pseudotype particles. J. Virol. Methods 160, 1–6. doi: 10.1016/j.jviromet.2009.02.025
Tani, H., Shiokawa, M., Kaname, Y., Kambara, H., Mori, Y., Abe, T., et al. (2010). Involvement of ceramide in the propagation of Japanese encephalitis virus. J. Virol. 84, 2798–2807. doi: 10.1128/JVI.02499-09
Tolah, A. M. K., Sohrab, S. S., Tolah, K. M. K., Hassan, A. M., El-Kafrawy, S. A., and Azhar, E. I. (2021). Evaluation of a pseudovirus neutralization assay for SARS-CoV-2 and correlation with live virus-based micro neutralization assay. Diagnostics (Basel) 11:994. doi: 10.3390/diagnostics11060994
Verma, D., Church, T. M., and Swaminathan, S. (2021). Epstein-barr virus lytic replication induces ACE2 expression and enhances SARS-CoV-2 pseudotyped virus entry in epithelial cells. J. Virol. 95:e0019221. doi: 10.1128/JVI.00192-21
Wang, G.-L., Wang, Z.-Y., Duan, L.-J., Meng, Q.-C., Jiang, M.-D., Cao, J., et al. (2021). Susceptibility of circulating SARS-CoV-2 variants to neutralization. N. Engl. J. Med. 384, 2354–2356. doi: 10.1056/NEJMc2103022
Wang, P., Nair, M. S., Liu, L., Iketani, S., Luo, Y., Guo, Y., et al. (2021). Antibody resistance of SARS-CoV-2 variants B.1.351 and B.1.1.7. Nature 593, 130–135. doi: 10.1038/s41586-021-03398-2
Weiss, R. A., and Bennett, P. L. (1980). Assembly of membrane glycoproteins studied by phenotypic mixing between mutants of vesicular stomatitis virus and retroviruses. Virology 100, 252–274. doi: 10.1016/0042-6822(80)90518-8
Weiss, R. A., Boettiger, D., and Murphy, H. M. (1977). Pseudotypes of avian sarcoma viruses with the envelope properties of vesicular stomatitis virus. Virology 76, 808–825. doi: 10.1016/0042-6822(77)90261-6
West, A. P., Wertheim, J. O., Wang, J. C., Vasylyeva, T. I., Havens, J. L., Chowdhury, M. A., et al. (2021). Detection and characterization of the SARS-CoV-2 lineage B.1.526 in New York. Nat. Commun. 12:4886. doi: 10.1038/s41467-021-25168-4
Whelan, S. P., Ball, L. A., Barr, J. N., and Wertz, G. T. (1995). Efficient recovery of infectious vesicular stomatitis virus entirely from cDNA clones. Proc. Natl. Acad. Sci. U.S.A. 92, 8388–8392. doi: 10.1073/pnas.92.18.8388
Whitt, M. A. (2010). Generation of VSV pseudotypes using recombinant ΔG-VSV for studies on virus entry, identification of entry inhibitors, and immune responses to vaccines. J. Virol. Methods 169, 365–374. doi: 10.1016/j.jviromet.2010.08.006
Winter, C., Schwegmann-Wessels, C., Neumann, U., and Herrler, G. (2008). The spike protein of infectious bronchitis virus is retained intracellularly by a tyrosine motif. J. Virol. 82, 2765–2771. doi: 10.1128/JVI.02064-07
Wrapp, D., Wang, N., Corbett, K. S., Goldsmith, J. A., Hsieh, C.-L., Abiona, O., et al. (2020). Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 367, 1260–1263. doi: 10.1126/science.abb2507
Wright, E., McNabb, S., Goddard, T., Horton, D. L., Lembo, T., Nel, L. H., et al. (2009). A robust lentiviral pseudotype neutralisation assay for in-field serosurveillance of Rabies and Lyssaviruses in Africa. Vaccine 27, 7178–7186. doi: 10.1016/j.vaccine.2009.09.024
Xiong, H.-L., Wu, Y.-T., Cao, J.-L., Yang, R., Liu, Y.-X., Ma, J., et al. (2020). Robust neutralization assay based on SARS-CoV-2 S-protein-bearing vesicular stomatitis virus (VSV) pseudovirus and ACE2-overexpressing BHK21 cells. Emerg. Microbes Infect. 9, 2105–2113. doi: 10.1080/22221751.2020.1815589
Yahalom-Ronen, Y., Tamir, H., Melamed, S., Politi, B., Shifman, O., Achdout, H., et al. (2020). A single dose of recombinant VSV-ΔG-spike vaccine provides protection against SARS-CoV-2 challenge. Nat. Commun. 11:6402. doi: 10.1038/s41467-020-20228-7
Yamate, M., Yamashita, M., Goto, T., Tsuji, S., Li, Y.-G., Warachit, J., et al. (2005). Establishment of Vero E6 cell clones persistently infected with severe acute respiratory syndrome coronavirus. Microbes Infect. 7, 1530–1540. doi: 10.1016/j.micinf.2005.05.013
Yuan, Y., Cao, D., Zhang, Y., Ma, J., Qi, J., Wang, Q., et al. (2017). Cryo-EM structures of MERS-CoV and SARS-CoV spike glycoproteins reveal the dynamic receptor binding domains. Nat. Commun. 8:15092. doi: 10.1038/ncomms15092
Závada, J. (1972). Pseudotypes of vesicular stomatitis virus with the coat of murine Leukaemia and of Avian Myeloblastosis viruses. J. Gen. Virol. 15, 183–191. doi: 10.1099/0022-1317-15-3-183
Zettl, F., Meister, T. L., Vollmer, T., Fischer, B., Steinmann, J., Krawczyk, A., et al. (2020). Rapid quantification of SARS-CoV-2-neutralizing antibodies using propagation-defective Vesicular Stomatitis virus pseudotypes. Vaccines (Basel) 8:E386. doi: 10.3390/vaccines8030386
Zhang, H., Zhu, W., Jin, Q., Pan, F., Zhu, J., Liu, Y., et al. (2021). Inhalable nanocatchers for SARS-CoV-2 inhibition. Proc. Natl. Acad. Sci. U.S.A. 118:e2102957118. doi: 10.1073/pnas.2102957118
Zhang, L., Li, Q., Liu, Q., Huang, W., Nie, J., and Wang, Y. (2017). A bioluminescent imaging mouse model for Marburg virus based on a pseudovirus system. Hum. Vaccin. Immunother. 13, 1811–1817. doi: 10.1080/21645515.2017.1325050
Zhou, L., Huntington, K., Zhang, S., Carlsen, L., So, E.-Y., Parker, C., et al. (2020). MEK inhibitors reduce cellular expression of ACE2, pERK, pRb while stimulating NK-mediated cytotoxicity and attenuating inflammatory cytokines relevant to SARS-CoV-2 infection. Oncotarget 11, 4201–4223. doi: 10.18632/oncotarget.27799
Zhu, Z.-L., Qiu, X.-D., Wu, S., Liu, Y.-T., Zhao, T., Sun, Z.-H., et al. (2020). Blocking Effect of demethylzeylasteral on the interaction between human ACE2 Protein and SARS-CoV-2 RBD protein discovered using SPR technology. Molecules 26:57. doi: 10.3390/molecules26010057
Keywords: vesicular stomatitis virus, SARS-CoV-2, Biosafety Level 3, pseudotyped viruses, pseudovirus, glycoprotein, ppVSVΔG-SARS-CoV-2 S
Citation: Salazar-García M, Acosta-Contreras S, Rodríguez-Martínez G, Cruz-Rangel A, Flores-Alanis A, Patiño-López G and Luna-Pineda VM (2022) Pseudotyped Vesicular Stomatitis Virus-Severe Acute Respiratory Syndrome-Coronavirus-2 Spike for the Study of Variants, Vaccines, and Therapeutics Against Coronavirus Disease 2019. Front. Microbiol. 12:817200. doi: 10.3389/fmicb.2021.817200
Received: 17 November 2021; Accepted: 20 December 2021;
Published: 14 January 2022.
Edited by:
Miguel Angel Martinez, IrsiCaixa, SpainReviewed by:
Daniel Smrz, Charles University in Prague, CzechiaCopyright © 2022 Salazar-García, Acosta-Contreras, Rodríguez-Martínez, Cruz-Rangel, Flores-Alanis, Patiño-López and Luna-Pineda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Victor M. Luna-Pineda, bHVuYS5waW5lZGFAaG90bWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.