 Xuemei Yang1
Xuemei Yang1 Ning Dong1
Ning Dong1 Xiaoxuan Liu1
Xiaoxuan Liu1 Chen Yang1Lianwei Ye1
Chen Yang1Lianwei Ye1 Edward Wai-Chi Chan1,2
Edward Wai-Chi Chan1,2 Rong Zhang3
Rong Zhang3 Sheng Chen1*
Sheng Chen1*- 1Department of Infectious Diseases and Public Health, Jockey Club College of Veterinary Medicine and Life Sciences, City University of Hong Kong, Kowloon, Hong Kong SAR, China
- 2State Key Lab of Chemical Biology and Drug Discovery, Department of Applied Biology and Chemical Technology, The Hong Kong Polytechnic University, Kowloon, Hong Kong SAR, China
- 3Department of Clinical Laboratory, School of Medicine, Second Affiliated Hospital of Zhejiang University, Hangzhou, China
Carbapenem-resistant hypervirulent Klebsiella pneumoniae (CR-HvKP) strains have been increasingly reported, and it is important to understand the evolutionary mechanisms of these highly pathogenic and resistant bacterial pathogens. In this study, we characterized a ST11 carbapenem-resistant K. pneumoniae strain which harbored an IncFIB/IncHI1B type virulence plasmid and an IncFII/IncR type blaKPC–2-bearing plasmid. The virulence plasmid was found to be conjugative and harbored a 35-kbp fragment including aerobactin encoding cluster from virulence plasmid pLVPK and multiple resistance genes, resulting in a mosaic multi-drug resistance and virulence plasmid. This virulence plasmid could be transferred via conjugation to Escherichia coli and K. pneumoniae strains alone as well as together with the blaKPC–2-bearing plasmid. Co-transmission of virulence and blaKPC–2-bearing plasmids would directly convert a classic K. pneumoniae strain into CR-HvKP strain, leading to a sharp increase in the prevalence of CR-HvKP in clinical settings, which poses a great threat to human health.
Introduction
Klebsiella pneumoniae is a Gram-negative pathogen that has become a major causative agent of hospital infections worldwide in recent years (Paczosa and Mecsas, 2016; Grundmann et al., 2017; Gu et al., 2018). K. pneumoniae could acquire an accessory genome which makes the strain either hypervirulent or antibiotic resistant (Lin et al., 2008; Yang et al., 2020). The hypervirulent K. pneumoniae (hvKP) pathotype is undergoing global dissemination and was found to be highly associated with genes peg-344, iroB, iucA, rmpA, and rmpA2, all of which were typically encoded by a pLVPK-like virulence plasmid (Russo et al., 2018). These genes are involved in the aerobactin (Iuc) and salmochelin (Iro) siderophores synthesis and the upregulation of capsule production. The best characterized virulence plasmids are plasmid pLVPK recovered from K2 hvKP strain CG43 and plasmid pK2044 from K1 hvKP strain NTUH-K2044 which shared 96% coverage and 99% identity (Chen et al., 2004; Wu et al., 2009). Another well-characterized K1 hvKP strain, SGH10, was also found to harbor a 231,583-bp virulence plasmid, similar to plasmids pK2044 and pLVPK (Lam et al., 2018). Plasmid recombination between such virulence plasmid and other plasmids is increasingly being observed, resulting in mosaic plasmids encoding both virulence and resistance elements or conjugative virulence plasmids (Lam et al., 2019; Turton et al., 2019; Yang et al., 2019). Such genetic changes during the transmission process indicates that virulence plasmids have rapidly adapted to co-exist with K. pneumoniae strains (Lev et al., 2018; Russo et al., 2018). In this study, we characterized a CRKP strain which harbored a conjugative virulence and resistance co-encoding plasmid. Such plasmid was found to be able to conjugate to Escherichia coli and K. pneumoniae strains together with the KPC plasmid.
Materials and Methods
Bacterial Strains
K. pneumoniae strain SH12 was recovered from a clinical specimen from a patient at a hospital located in Shanghai in 2015 in a nationwide surveillance project in mainland China. This strain was isolated from a sputum sample of the hospitalized patient, who was diagnosed with pneumonia. A string test was performed on blood agar as previously described (Gu et al., 2018). K. pneumoniae strains CR-HvKP4 and FJ8 were reported in our previous study (Gu et al., 2018). Ciprofloxacin-resistant K. pneumoniae strain PM23 was a clinical isolate in Hong Kong provided by Prof. Kwok-yung Yuen from The University of Hong Kong. E. coli strains J53 and ATCC 25922 were from our laboratory stock.
Antibiotic Susceptibility Testing
Antimicrobial susceptibility testing was performed by the micro dilution method. E. coli strain ATCC 25922 served as quality control strain for susceptibility testing. Antimicrobial agents including aztreonam, cefotaxime, ceftazidime, meropenem, imipenem, ertapenem, ciprofloxacin, amikacin, polymyxin B, tigecycline, ceftazidime-avibactam, and cefiderocol were tested. All tests were performed in duplicate, and each test included three biological replicates per strain. The susceptibility was interpreted according to both Performance Standards for Antimicrobial Susceptibility Testing by the Clinical and Laboratory Standards Institute (CLSI) of 2021 (CLSI, 2021) and Clinical Breakpoints and Guidance by European Committee on Antimicrobial Susceptibility Testing (EUCAST) of 2021 (The European Committee on Antimicrobial Susceptibility Testing, 2021).
DNA Sequencing and Bioinformatics
Genomic DNA of strain SH12 was extracted using the PureLink Genomic DNA Mini Kit (Invitrogen, United States) according to the manufacturer’s instructions. The extracted DNA was then subjected to library preparation by NEBNext Ultra II DNA Library Prep Kit for Illumina (New England Biolabs, United States) and sequenced via the 150-bp paired-end NextSeq 500 platform (Illumina, San Diego, CA). Genomic DNA was also subjected to the long-read MinION platform (Oxford Nanopore Technologies, Oxford, United Kingdom). MinION libraries were prepared using the SQK-RBK004 nanopore sequencing kit according to the manufacturer’s instructions. The library was then added to a MinION flow cell (R9. 4.1) and sequenced. Base calling was performed using Guppy v5.0.11. Read trimming and filtering was performed using NanoFilt v2.8.0 (De Coster et al., 2018). Both short and long reads were de novo hybrid assembled using Unicycler v0.4.7 (Wick et al., 2017). Assembled genome sequences were annotated with RAST v2.0 (Brettin et al., 2015). Multi-locus sequence typing (MLST) were determined by the Kleborate software based on genetic variation in the seven housekeeping genes (Lam et al., 2020). Capsular typing and O-antigen typing on the assembled sequences were performed using Kaptive (Wyres et al., 2016). Virulence genes were identified by searching against the BIGSdb Klebsiella genome database.1 The BLAST command lines, with an 80% coverage and identity cutoff, were used to map genome sequences against antibiotic resistance genes and plasmid replicons. The resistance genes and plasmid replicons databases were obtained from the Center for Genomic Epidemiology.2 The insertion sequences (ISs) were identified using Isfinder and ISsaga3 (Siguier et al., 2006). Alignment of plasmids with similar structures were generated by Easyfig win_2.1 (Sullivan et al., 2011).
Conjugation Assay
To determine the transferability of the virulence plasmid and KPC plasmid of strain SH12, conjugation was performed using azide-resistant E. coli strain J53 as recipient. Transfer of the plasmids from the donor strain to the recipient strain was determined by plating samples of a mating mixture onto selective media as previously described with modifications (Yang et al., 2019). As the virulence plasmid carried resistance gene blaDHA–1, the transconjugants carrying the virulence plasmid were selected by spreading the culture on eosin-methylene blue (EMB) agar plates containing 2 μg/ml cefotaxime and 100 μg/ml sodium azide. As transconjugants carrying the KPC plasmid can also grow on the EMB agar plates containing 2 μg/ml cefotaxime and 100 μg/ml sodium azide, colonies were further tested for growth on EMB plates containing 2 μg/ml meropenem. Those that could grow on both plates containing 2 μg/ml cefotaxime and 2 μg/ml meropenem were treated as KPC plasmid transconjugants, and those that could grow on plates containing 2 μg/ml cefotaxime rather than plates containing 2 μg/ml meropenem were treated as virulence plasmid transconjugants. The presence of rmpA2 and blaKPC–2 genes as marker genes of virulence plasmid and KPC plasmid in transconjugants was determined by polymerase chain reaction (PCR). The minimum inhibitory concentration (MIC) profiles of transconjugants were also determined to differentiate from the donor. Successful virulence plasmid transconjugants of E. coli strain J53 were then treated as donor and the ciprofloxacin-resistant K. pneumoniae strain PM23 as recipient. MacConkey agar plates containing 2 μg/ml cefotaxime together with 2 μg/ml ciprofloxacin were used to select transconjugants. S1 nuclease pulsed-field gel electrophoresis (S1-PFGE) was performed to confirm the transfer of plasmids. To calculate the conjugation efficiency, the diluted culture after conjugation was also plated on plates containing only 100 μg/ml sodium azide for strain J53 as recipient and 2 μg/ml ciprofloxacin for strain PM23 as recipient to count the total recipient cells, respectively. The conjugation efficiency was calculated as the number of transconjugants cells divided by the number of recipient cells. The conjugation efficiency was determined for transconjugants carrying virulence plasmid only, blaKPC–2-bearing plasmid only, and both plasmids.
Plasmid Curing
Curing of the large virulence plasmid of strain SH12, pSH12_Vir, was performed using sodium dodecyl sulfate (SDS) (El-Mansi et al., 2000). Briefly, an SDS solution (10% w/v, pH 7.4) was added to Luria broth (LB) to give 5, 4, 3, 2, 1, and 0.5% SDS. An inoculum of 100 μl overnight-cultured strain SH12 was then used to seed the SDS-containing LB, and the cultures were incubated overnight with shaking at 27°C. After SDS treatment, the diluted culture of each concentration was spread on LB agar plates, respectively, to select virulence plasmid cured colonies. About 50 colonies from each concentration were passed on LB agar plates and tested for the presence of rmpA2 and iucA genes, and those which proved negative were treated as plasmid cured strains. The plasmid-curing rate for the plasmid is 5.00E–03. Plasmid curing was confirmed by S1-PFGE analysis.
Mouse Infection Model
Mouse bacteremia model was used to test the potential virulence of K. pneumoniae strain SH12. In this experiment, eight male Crl:CD1(ICR) mice in each group were infected intraperitoneally with an inoculum of 2.0 × 105 and 1.0 × 106 CFUs of different strains of K. pneumoniae, respectively. The mortality rate of the test mice was observed and recorded for 1 week post infection. Carbapenem-resistant, hypervirulent K. pneumoniae strain CR-HvKP4 reported in our previous study was used as control for high virulence, while CRKP strain FJ8 was used as control for low virulence (Gu et al., 2018; Yang et al., 2019). Strain CR-HvKP4 was a ST11 CRKP strain that harbored a pLVPK-like virulence plasmid, exhibiting hyper virulence. FJ8 was a classical CRKP strain which exhibited low virulence. All animal experiments were approved by the Animal Research Ethics Sub-Committee, City University of Hong Kong. Animal experiments were repeated twice to assess the consistency of the data.
Results and Discussion
A strain suspected to be K. pneumoniae, SH12, was recovered from a sputum sample of a hospitalized patient in 2015 in Shanghai, China. Results of string test indicated that strain SH12 was string test negative. Antimicrobial susceptibility testing performed on SH12 showed that it was resistant to all β-lactam antibiotics, amikacin, and ciprofloxacin but remained susceptible to polymyxin B and ceftazidime-avibactam according to both CLSI and EUCAST standards and resistant to cefiderocol but susceptible to tigecycline according to EUCAST standards (Table 1).
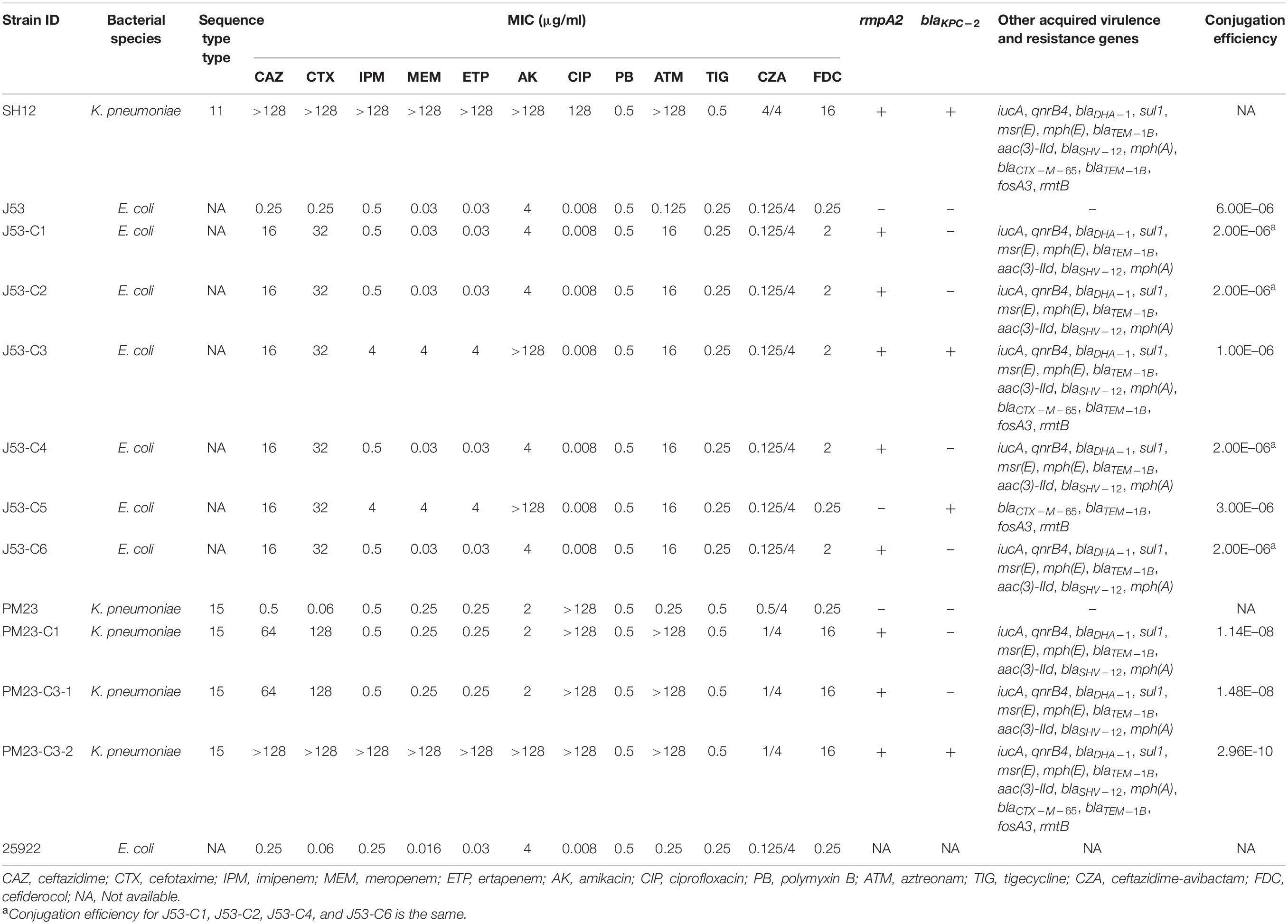
Table 1. Phenotypic and genotypic characteristics of K. pneumoniae strain SH12 and its transconjugants.
Strain SH12 was then subjected to whole-genome sequencing using both the 150-bp paired-end Illumina NextSeq 500 platform and the long-read Oxford Nanopore Technologies MinION platform. The genome size of strain SH12 was found to be 5,922,385 bp, including a 5.19-Mb chromosome and six plasmids with size of 284,671 bp (IncFIB/IncHI1B), 167,468 bp (IncFII/IncR), 10,060 bp (ColRNAI), 6,135 bp (IncQ1), 5,596 bp (ColRNAI), and 3,138 bp, respectively. Strain SH12 was found to belong to ST11 based on MLST, KL47 serotype based on capsular typing, and OL101 O-antigen type. The 6,135-bp IncQ1 plasmid was designated pSH12_4 which carried ermT gene coding for macrolide resistance. The chromosome of strain SH12 also harbored blaSHV–11 and fosA genes.
Strain SH12 harbored a number of virulence factors, including the regulator of mucoid phenotype 2 (rmpA2), aerobactin (iucABCD,iutA), and yersiniabactin (ybt 9; ICEKp3). However, the RmpA2 was found to be truncated at position 126 (nucleotide C301A&GGG284–286 delete&A353 delete), which might cause the negative string test. The rmpA2 and iucABCDiutA genes were found to be located in a 284,671-bp IncFIB/IncHI1B plasmid, designated as pSH12_Vir (Figure 1). Using Blast against the National Center for Biotechnology Information (NCBI) database, this plasmid was found to show highest similarity (90% coverage and 99% identity) to plasmid p17-16-vir (GenBank accession no. MK191024.1), a 290,451-bp IncFIB/IncHI1B plasmid recovered from a K. pneumoniae strain isolated from sputum in China. Plasmid p17-16-vir was found to have a 66-kbp insertion from virulence plasmid pLVPK, including the tellurium resistance cluster, rmpA2 and iucABCDiutA genes (Figure 1 and Table 2). Plasmid pSH12_Vir was found to reserve 35 kbp of this fragment, including rmpA2 and iucABCDiutA genes, with the tellurium resistance cluster excluded. These genetic changes indicated the adaption of virulence plasmids with K. pneumoniae strains during the transmission process. These virulence plasmids lacked virulence genes rmpA and iroBCDN when compared to typical virulence plasmid pLVPK, which may affect the virulence potentials of these CRKP strains. However, 96.2% of the virulence plasmids in the GenBank carried iucA, while only 46.8% carried iroB (Tian et al., 2021). The virulence effects and any benefits of the iro deletion remains to be studied. There were also several mobile genetic elements with antimicrobial resistance (AMR) genes found in pSH12_Vir, including a qnrB4-blaDHA–1-sul1 integron and an IS5-msr(E)-mph(E)-ISKpn21 region which was similar to plasmid p17-16-vir. Other AMR regions were an IS26-mer-IS5075-blaTEM–1B-ISCfr1-aac(3)-IId-IS26-blaSHV–12-IS26-blaSHV–12-IS26 region and an ISKpn28-mph(A)-IS6100-IS5075 region. Such virulence-resistance hybrid plasmids have been reported recently, which can be generated by integrating the AMR gene-bearing DNA fragment into the virulence plasmid backbone (Zhang et al., 2016; Shen et al., 2019), as well as integrating the virulence-bearing DNA fragment into the AMR plasmid (Lam et al., 2019; Turton et al., 2019). Strain SH12 was also shown to harbor the carbapenemase gene blaKPC–2, which was found to be located in a NTEKPC-Ib transposon in a 167,468-bp IncFII/IncR plasmid, pSH12_KPC (Figure 2). Plasmid pSH12_KPC was highly similar (98% coverage and 99% identity) to the 177,516-bp IncFII/IncR plasmid pKPC2_020002 (GenBank accession no. CP028541.2) recovered from K. pneumoniae strain WCHKP2 (Figure 2 and Table 2). Plasmid pSH12_KPC was also found to harbor blaCTX–M–65, blaTEM–1B, fosA3, and rmtB resistance genes.
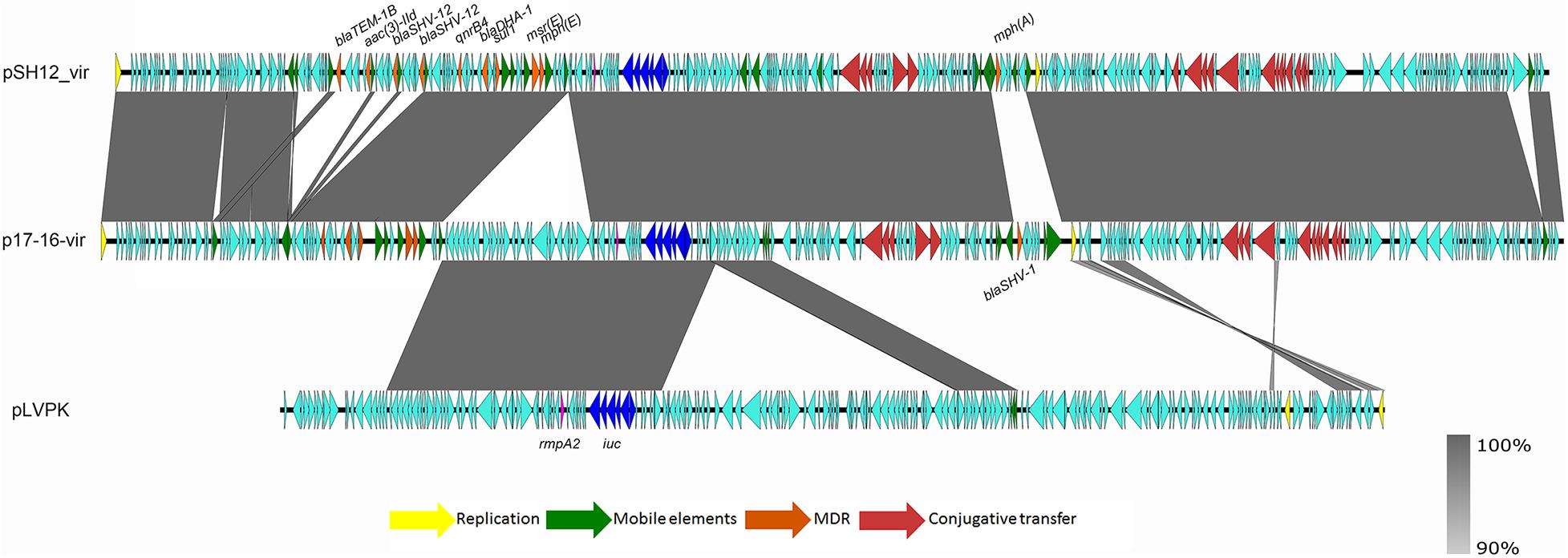
Figure 1. Alignment of pSH12_Vir and plasmids with similar structures. Plasmid pSH12_Vir was an IncFIB/IncHI1B plasmid with several mosaic MDR regions and virulence factors. Genetic regions associated with virulence and resistance are highlighted.

Table 2. Comparison of plasmids pSH12_Vir and pSH12_KPC with similar plasmids in the GenBank database.
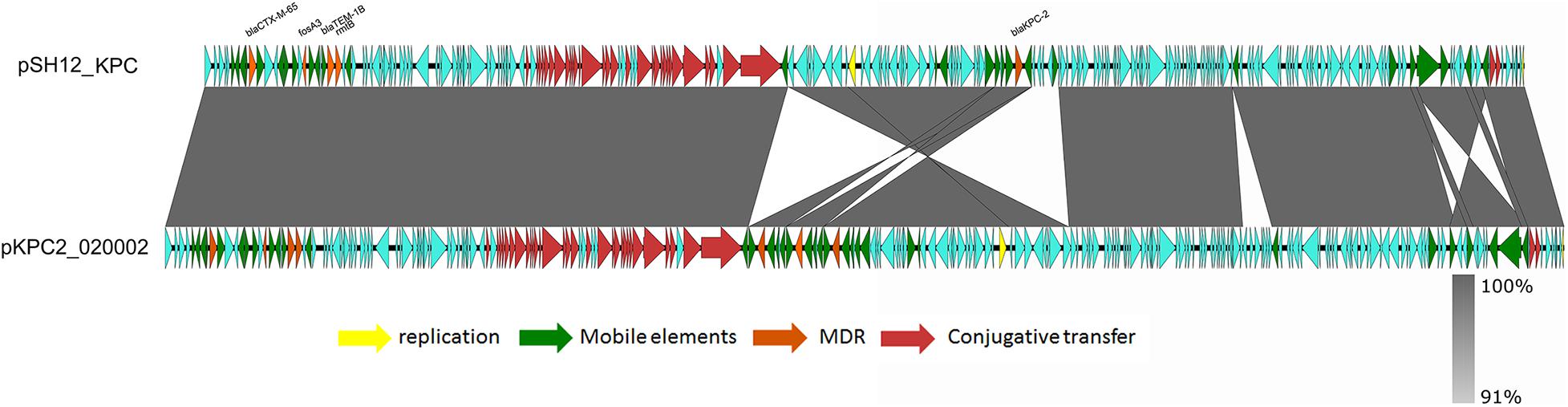
Figure 2. Alignment of pSH12_KPC and plasmid with similar structures. Plasmid pSH12_KPC was highly similar (98% coverage and 99% identity) to plasmid pKPC2_020002 (GenBank accession no. CP028541.2) recovered from a K. pneumoniae strain.
More importantly, carrying the self-transmission systems, mosaic multidrug resistance (MDR)-virulence plasmids could be transferred into hvKP or cKP strains, resulting in MDR-HvKP strains (Tang et al., 2020). As plasmid pSH12_Vir and pSH12_KPC were both shown to harbor conjugative transfer genes, the transferability of these two plasmids were determined by conjugation. Firstly, E. coli strain J53 was used as recipient strain. Both the virulence and KPC plasmid were found to be able to transfer to strain J53 (Table 1 and Figure 3). Transconjugants J53-C1 showed resistance to β-lactam antibiotics, ceftazidime, cefotaxime, and aztreonam and was positive for rmpA2, a marker gene of virulence plasmid pSH12_Vir by PCR, indicating that plasmid pSH12_Vir was acquired. Transconjugants J53-C5 showed resistance to β-lactam antibiotics, including carbapenem and amikacin, and was positive for blaKPC–2, a marker gene of the KPC plasmid pSH12_KPC by PCR, indicating that plasmid pSH12_KPC was acquired. Interestingly, transconjugant J53-C3 was positive for both rmpA2 and blaKPC–2 genes, indicating that transconjugants received these two plasmids simultaneously. The KPC plasmid was confirmed in the according transconjugants J53-C3 and J53-C5 by S1-PFGE (Figure 3). The virulence plasmid in the E. coli transconjugants was hardly observed even though several colonies were tried, namely, J53-C1, J53-C2, J53-C4, and J53-C6, which were identified as acquiring virulence plasmid pSH12_Vir by MIC and PCR assays; a weak band was observed of transconjugant J53-C6 (Figure 3). This might be caused by low copy numbers of this plasmid in E. coli. To further determine the transferability of the virulence plasmid, transconjugant strain J53-C1 and J53-C3 were used as donors, respectively, and ciprofloxacin-resistant K. pneumoniae strain PM23 was used as recipient. It was found that virulence plasmid pSH12_Vir could be transferred to K. pneumoniae strain PM23 alone as well as together with pSH12_KPC from those E. coli transconjugants. When J53-C3 was used as donor, transconjugant PM23-C3-1 was found to receive only the virulence plasmid; transconjugant PM23-C3-2 was found to receive both the virulence and KPC plasmid (Table 1 and Figure 3). Interestingly, strain SH12 was resistant to cefiderocol with a MIC of 16 μg/ml, and strains that received plasmid pSH12_Vir also exhibited increased resistance to cefiderocol (Table 1), indicating possible resistance determinants on plasmid pSH12_Vir. Limited data of cefiderocol resistance mechanisms are currently available, and deficiencies in the iron transporter systems (Ito et al., 2018), serine β-lactamases, and metallo-β-lactamases (MBLs) (Kohira et al., 2020) might play roles in conferring resistance to cefiderocol. Thus, serine β-lactamases, DHA-1 and TEM-1B, encoded by plasmid pSH12_Vir, may play a role in mediating resistance to cefiderocol, which needs further confirmation.
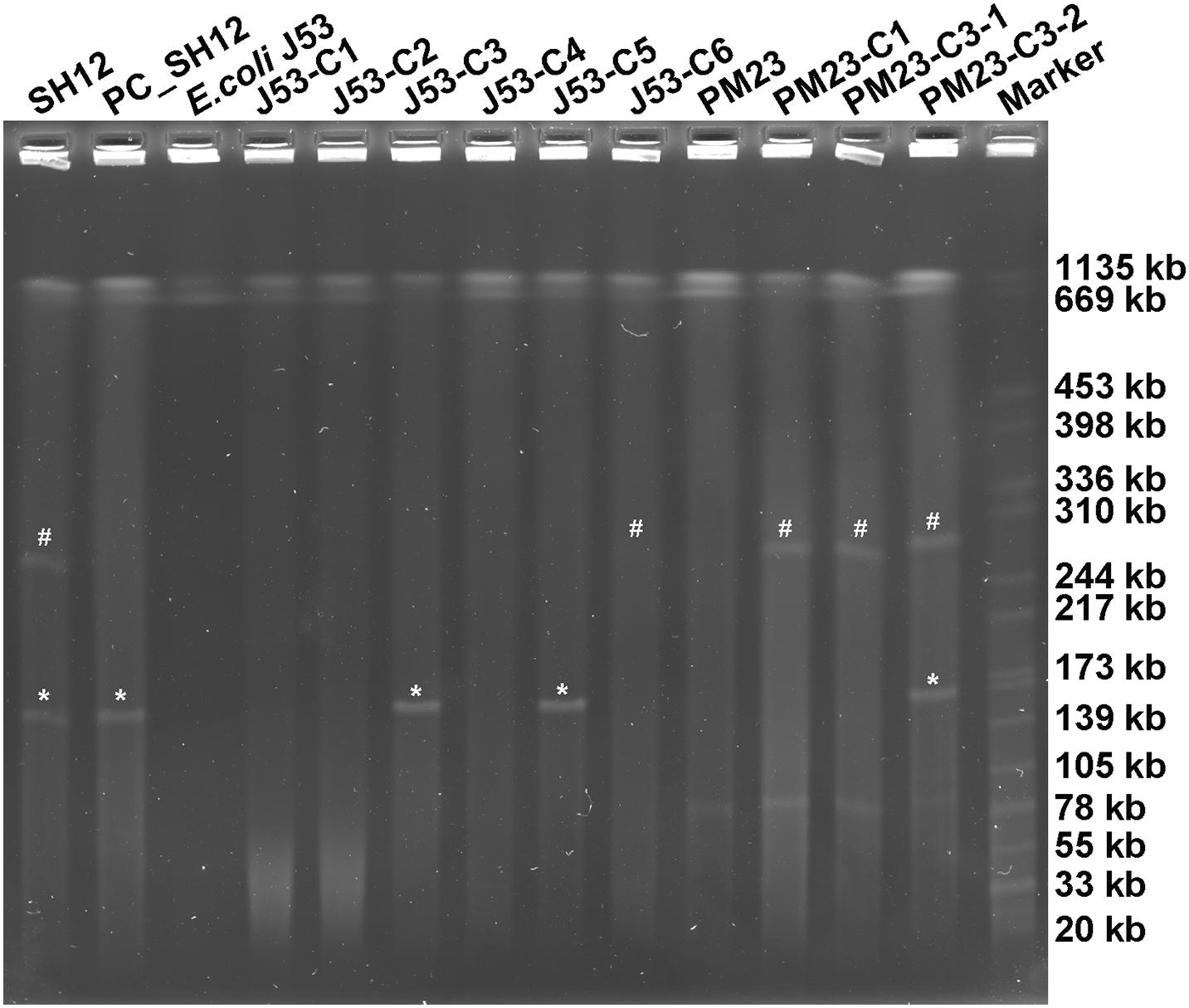
Figure 3. S1-PFGE of strain SH12 and its transconjugants. Transconjugants J53-C1, J53-C2, J53-C4, and J53-C6 were found to receive virulence plasmid only; J53-C5 was found to receive the KPC plasmid only; J53-C3 was found to receive both plasmids. Transconjugant PM23-C1 was generated using J53-C1 as donor. Transconjugants PM23-C3-1 and PM23-C3-2 were generated using J53-C3 as donor. “#” denotes virulence plasmid pSH12_Vir, and “*” denotes KPC plasmid pSH12_KPC.
The virulence level of strain SH12 was tested in male ICR mouse. Upon being infected for 1 week at an inoculum of 1 × 106 CFU, survival of mice was 0% with strain CR-HvKP4, 0% with SH12, and 75% with strain FJ8 (Figure 4B). While at an inoculum of 2 × 105 CFU, survival of mice was 0% with strain CR-HvKP4, 50% with SH12, and 100% with strain FJ8 (Figure 4A). These data all showed that strain SH12 possessed moderate virulence potential. To determine virulence contribution of the virulence plasmid to strain SH12, plasmid curing was performed. The plasmid-cured strain, designated as PC_SH12, was then subjected to mice infection model. Upon being infected for 1 week at inoculums of 1 × 106 and 2 × 105 CFU, survival of mice was 50 and 100% with strain PC_SH12, respectively, indicating that virulence plasmid pSH12_Vir played an important role in determining virulence of strain SH12 (Figure 4).
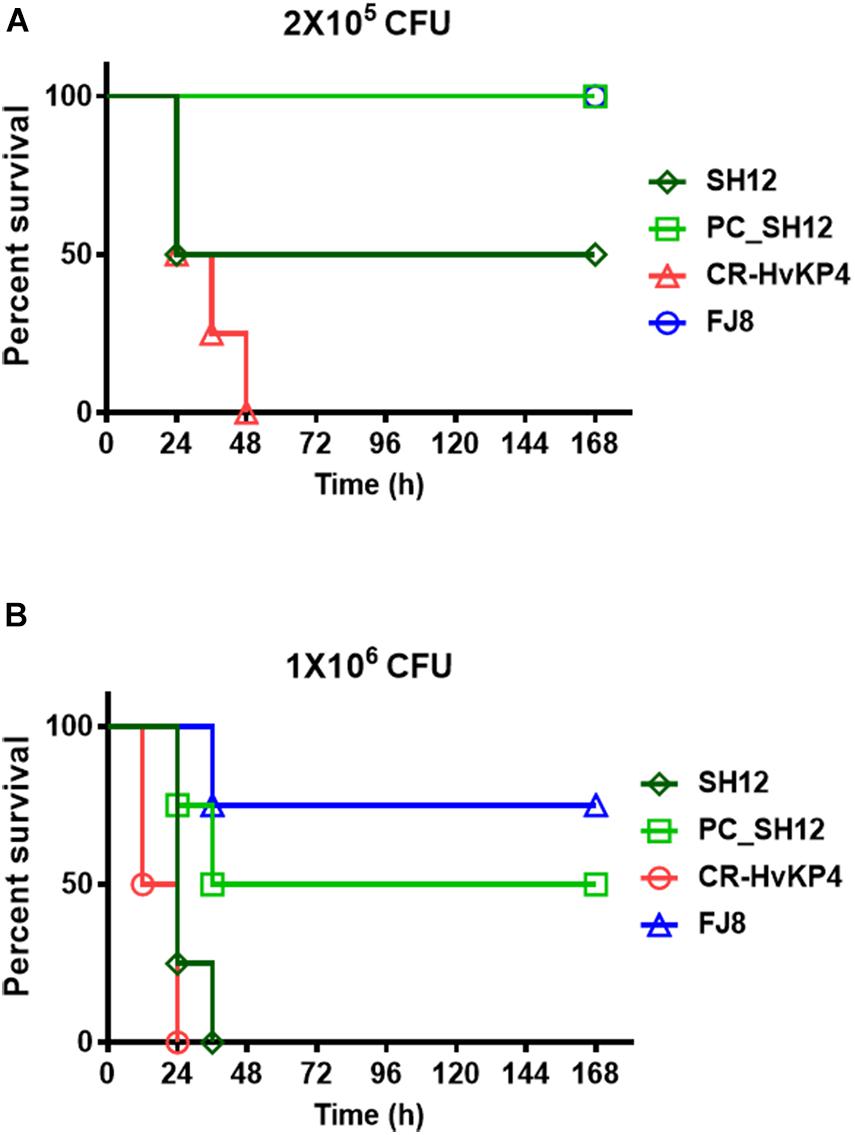
Figure 4. Virulence level of different bacterial strains as depicted in mouse infection model. Survival of mice (n = 8) infected by 2 × 105 CFU (A) and 1 × 106 CFU (B) of each K. pneumoniae strain at 168 h. The test strains included K. pneumoniae strain SH12, its virulence plasmid cured strain PC_SH12, CR-HvKP strain CR-HvKP4 (hypervirulence control), and the classic CRKP strain FJ8 (low virulence control).
Conclusion
In conclusion, this study characterized a CRKP strain which harbored a conjugative virulence plasmid. This virulence plasmid was generated by integration of a 35-kbp fragment containing rmpA2 gene and aerobactin encoding genes from a pLVPK-like virulence plasmid into a resistance plasmid. Generation of conjugative plasmid simultaneously carrying virulence and resistance-encoding genes will increase the spread of these elements among pathogens. This virulence plasmid was found to be able to transfer together with the KPC plasmid, which may promote prevalence of CR-HvKP infections. Findings in this work therefore provide important insight into the evolution of virulence-encoding elements during transmission in K. pneumoniae strains.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/genbank/, CP040833-CP040839.
Ethics Statement
The animal study was reviewed and approved by the Animal Research Ethics Sub-Committee, City University of Hong Kong.
Author Contributions
XY performed the experiment and drafted the manuscript. ND helped with the bioinformatic analysis. XL and CY performed the S1-PFGE. LY performed the DNA sequencing. EC edited the manuscript and contributed to experimental design. RZ and SC designed, supervised the study, and interpreted the data. SC wrote the manuscript. All authors contributed to the article and approved the submitted version.
Funding
The research was supported by SGP grant from City University of Hong Kong (9380110).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
- ^ http://bigsdb.pasteur.fr/klebsiella/klebsiella.html
- ^ http://www.genomicepidemiology.org/
- ^ https://www-is.biotoul.fr/index.php
References
Brettin, T., Davis, J. J., Disz, T., Edwards, R. A., Gerdes, S., Olsen, G. J., et al. (2015). RASTtk: a modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 5:8365. doi: 10.1038/srep08365
Chen, Y.-T., Chang, H. Y., Lai, Y. C., Pan, C. C., Tsai, S. F., and Peng, H. L. (2004). Sequencing and analysis of the large virulence plasmid pLVPK of Klebsiella pneumoniae CG43. Gene 337, 189–198. doi: 10.1016/j.gene.2004.05.008
CLSI (2021). Performance Standards for Antimicrobial Susceptibility Testing. 31th ed. CLSI Supplement M100. Wayne, PA: Clinical and Laboratory Standards Institute.
De Coster, W., D’Hert, S., Schultz, D. T., Cruts, M., and Van Broeckhoven, C. (2018). NanoPack: visualizing and processing long-read sequencing data. Bioinformatics 34, 2666–2669. doi: 10.1093/bioinformatics/bty149
El-Mansi, M., Anderson, K. J., Inche, C. A., Knowles, L. K., and Platt, D. J. (2000). Isolation and curing of the Klebsiella pneumoniae large indigenous plasmid using sodium dodecyl sulphate. Res. Microbiol. 151, 201–208. doi: 10.1016/s0923-2508(00)00140-6
Grundmann, H., Glasner, C., Albiger, B., Aanensen, D. M., Tomlinson, C. T., Andrasević, A. T., et al. (2017). Occurrence of carbapenemase-producing Klebsiella pneumoniae and Escherichia coli in the European survey of carbapenemase-producing Enterobacteriaceae (EuSCAPE): a prospective, multinational study. Lancet Infect. Dis. 17, 153–163. doi: 10.1016/S1473-3099(16)30257-2
Gu, D., Dong, N., Zheng, Z., Lin, D., Huang, M., Wang, L., et al. (2018). A fatal outbreak of ST11 carbapenem-resistant hypervirulent Klebsiella pneumoniae in a Chinese hospital: a molecular epidemiological study. Lancet Infect. Dis. 18, 37–46. doi: 10.1016/S1473-3099(17)30489-9
Ito, A., Sato, T., Ota, M., Takemura, M., Nishikawa, T., Toba, S., et al. (2018). In vitro antibacterial properties of cefiderocol, a novel siderophore cephalosporin, against gram-negative bacteria. Antimicrob. Agents Chemother. 62:e01454-17.
Kohira, N., Hackel, M. A., Ishioka, Y., Kuroiwa, M., Sahm, D. F., and Sato, T. (2020). Reduced susceptibility mechanism to cefiderocol, a siderophore cephalosporin, among clinical isolates from a global surveillance programme (SIDERO-WT-2014). J. Glob. Antimicrob. Resist. 22, 738–741. doi: 10.1016/j.jgar.2020.07.009
Lam, M. M. C., Wick, R. R., Wyres, K. L., and Holt, K. E. (2020). Genomic surveillance framework and global population structure for Klebsiella pneumoniae. bioRxiv [Preprint] doi: 10.1101/2020.12.14.422303
Lam, M. M. C., Wyres, K. L., Duchêne, S., Wick, R. R., Judd, L. M., Gan, Y.-H., et al. (2018). Population genomics of hypervirulent Klebsiella pneumoniae clonal-group 23 reveals early emergence and rapid global dissemination. Nat. Commun. 9:2703. doi: 10.1038/s41467-018-05114-7
Lam, M. M. C., Wyres, K. L., Wick, R. R., Judd, L. M., Fostervold, A., Holt, K. E., et al. (2019). Convergence of virulence and MDR in a single plasmid vector in MDR Klebsiella pneumoniae ST15. J. Antimicrob. Chemother. 74, 1218–1222. doi: 10.1093/jac/dkz028
Lev, A. I., Astashkin, E. I., Kislichkina, A. A., Solovieva, E. V., Kombarova, T. I., Korobova, O. V., et al. (2018). Comparative analysis of Klebsiella pneumoniae strains isolated in 2012-2016 that differ by antibiotic resistance genes and virulence genes profiles. Pathog. Glob. Health 112, 142–151. doi: 10.1080/20477724.2018.1460949
Lin, T. L., Lee, C. Z., Hsieh, P. F., Tsai, S. F., and Wang, J. T. (2008). Characterization of integrative and conjugative element ICEKp1-associated genomic heterogeneity in a Klebsiella pneumoniae strain isolated from a primary liver abscess. J. Bacteriol. 190, 515–526. doi: 10.1128/JB.01219-07
Paczosa, M. K., and Mecsas, J. (2016). Klebsiella pneumoniae: going on the offense with a strong defense. Microbiol. Mol. Biol. Rev. 80, 629–661. doi: 10.1128/MMBR.00078-15
Russo, T. A., Olson, R., Fang, C. T., Stoesser, N., Miller, M., MacDonald, U., et al. (2018). Identification of biomarkers for differentiation of hypervirulent Klebsiella pneumoniae from Classical K. pneumoniae. J. Clin. Microbiol. 56:e00776-18.
Shen, D., Ma, G., Li, C., Jia, X., Qin, C., Yang, T., et al. (2019). Emergence of a multidrug-resistant hypervirulent Klebsiella pneumoniae sequence type 23 strain with a rare bla CTX-M-24-harboring virulence plasmid. Antimicrob. Agents Chemother. 63:e02273-18. doi: 10.1128/AAC.02273-18
Siguier, P., Perochon, J., Lestrade, L., Mahillon, J., and Chandler, M. (2006). ISfinder: the reference centre for bacterial insertion sequences. Nucleic Acids Res. 34, D32–D36.
Sullivan, M. J., Petty, N. K., and Beatson, S. A. (2011). Easyfig: a genome comparison visualizer. Bioinformatics 27, 1009–1010. doi: 10.1093/bioinformatics/btr039
Tang, M., Kong, X., Hao, J., and Liu, J. (2020). Epidemiological characteristics and formation mechanisms of multidrug-resistant hypervirulent Klebsiella pneumoniae. Front. Microbiol. 11:581543. doi: 10.3389/fmicb.2020.581543
The European Committee on Antimicrobial Susceptibility Testing (2021). Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 11.0. Available online at: http://www.eucast.org (accessed January 6, 2021).
Tian, D., Wang, M., Zhou, Y., Hu, D., Ou, H. Y., and Jiang, X. (2021). Genetic diversity and evolution of the virulence plasmids encoding aerobactin and salmochelin in Klebsiella pneumoniae. Virulence 12, 1323–1333. doi: 10.1080/21505594.2021.1924019
Turton, J., Davies, F., Turton, J., Perry, C., Payne, Z., and Pike, R. (2019). Hybrid resistance and virulence plasmids in “High-Risk” clones of Klebsiella pneumoniae, including those carrying blaNDM-5. Microorganisms 7:326. doi: 10.3390/microorganisms7090326
Wick, R. R., Judd, L. M., Gorrie, C. L., and Holt, K. E. (2017). Unicycler: resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 13:e1005595. doi: 10.1371/journal.pcbi.1005595
Wu, K. M., Li, L. H., Yan, J. J., Tsao, N., Liao, T. L., Tsai, H. C., et al. (2009). Genome sequencing and comparative analysis of Klebsiella pneumoniae NTUH-K2044, a strain causing liver abscess and meningitis. J. Bacteriol. 191, 4492–4501. doi: 10.1128/JB.00315-09
Wyres, K. L., Wick, R. R., Gorrie, C., Jenney, A., Follador, R., Thomson, N. R., et al. (2016). Identification of Klebsiella capsule synthesis loci from whole genome data. Microb. Genom. 2:e000102. doi: 10.1099/mgen.0.000102
Yang, X., Chan, E. W. C., Zhang, R., and Chen, S. A. (2019). conjugative plasmid that augments virulence in Klebsiella pneumoniae. Nat. Microbiol. 4, 2039–2043. doi: 10.1038/s41564-019-0566-7
Yang, X., Ye, L., Li, Y., Chan, E. W., Zhang, R., and Chen, S. (2020). Identification of a chromosomal integrated DNA fragment containing the rmpA2 and iucABCDiutA virulence genes in Klebsiella pneumoniae. mSphere 5:e01179-20. doi: 10.1128/mSphere.01179-20
Keywords: Klebsiella pneumonia, conjugative virulence plasmid, blaKPC-bearing plasmid, transmission, CR-HvKP, IncFIB/IncHI1B, IncFII/IncR
Citation: Yang X, Dong N, Liu X, Yang C, Ye L, Chan EW-C, Zhang R and Chen S (2021) Co-conjugation of Virulence Plasmid and KPC Plasmid in a Clinical Klebsiella pneumoniae Strain. Front. Microbiol. 12:739461. doi: 10.3389/fmicb.2021.739461
Received: 11 July 2021; Accepted: 13 October 2021;
Published: 08 November 2021.
Edited by:
Basil Britto Xavier, University of Antwerp, BelgiumReviewed by:
Kotaro Aoki, Toho University, JapanCarla Fontana, Policlinico Tor Vergata, Italy
Sulagna Basu, National Institute of Cholera and Enteric Diseases (ICMR), India
Yi-Tsung Lin, Taipei Veterans General Hospital, Taiwan
Copyright © 2021 Yang, Dong, Liu, Yang, Ye, Chan, Zhang and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sheng Chen, c2hlY2hlbkBjaXR5dS5lZHUuaGs=