
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 29 September 2021
Sec. Extreme Microbiology
Volume 12 - 2021 | https://doi.org/10.3389/fmicb.2021.728977
 Pauline Bergsten1,2
Pauline Bergsten1,2 Pauline Vannier1
Pauline Vannier1 Alexandra María Klonowski1
Alexandra María Klonowski1 Stephen Knobloch1
Stephen Knobloch1 Magnús Tumi Gudmundsson3
Magnús Tumi Gudmundsson3 Marie Dolores Jackson4
Marie Dolores Jackson4 Viggó Thor Marteinsson1,5*
Viggó Thor Marteinsson1,5*The island of Surtsey was formed in 1963–1967 on the offshore Icelandic volcanic rift zone. It offers a unique opportunity to study the subsurface biosphere in newly formed oceanic crust and an associated hydrothermal-seawater system, whose maximum temperature is currently above 120°C at about 100m below surface. Here, we present new insights into the diversity, distribution, and abundance of microorganisms in the subsurface of the island, 50years after its creation. Samples, including basaltic tuff drill cores and associated fluids acquired at successive depths as well as surface fumes from fumaroles, were collected during expedition 5059 of the International Continental Scientific Drilling Program specifically designed to collect microbiological samples. Results of this microbial survey are investigated with 16S rRNA gene amplicon sequencing and scanning electron microscopy. To distinguish endemic microbial taxa of subsurface rocks from potential contaminants present in the drilling fluid, we use both methodological and computational strategies. Our 16S rRNA gene analysis results expose diverse and distinct microbial communities in the drill cores and the borehole fluid samples, which harbor thermophiles in high abundance. Whereas some taxonomic lineages detected across these habitats remain uncharacterized (e.g., Acetothermiia, Ammonifexales), our results highlight potential residents of the subsurface that could be identified at lower taxonomic rank such as Thermaerobacter, BRH-c8a (Desulfallas-Sporotomaculum), Thioalkalimicrobium, and Sulfurospirillum. Microscopy images reveal possible biotic structures attached to the basaltic substrate. Finally, microbial colonization of the newly formed basaltic crust and the metabolic potential are discussed on the basis of the data.
The subsurface biosphere, defined as an ecosystem encompassing regions beneath soils and sediments, occupies roughly twice the volume of the oceans and holds about 15% of the total biomass on Earth (Pedersen, 2000; Heberling et al., 2010; Bar-On et al., 2018; Orcutt et al., 2019). Recent estimates suggest that these zones (i.e., continental subsurface, subseafloor sediments, and oceanic crust) contain ~70% of all prokaryotic cells and possibly more than 80% of all bacterial and archaeal species (Magnabosco et al., 2018). This large biome has only recently become the focus of research studies (Gold, 1992), and most microbial surveys have, to date, focused on subseafloor sediments (Parkes et al., 2000; D’Hondt et al., 2004; Inagaki et al., 2015). The deep biosphere hosted in the basaltic ocean crust, on the contrary, has been understudied because of various challenges: the relative inaccessibility of the samples (drilling operation often required), the low biomass (high risk of external contamination), the presence of minerals which generates DNA binding and inhibitions (Direito et al., 2012; Lever et al., 2015), and the difficulties in handling the inevitable microbial contamination from the surface (Baross et al., 2004; Sheik et al., 2018; Ramírez et al., 2019).
Over the past decades, many microorganisms have been discovered within Earth’s igneous oceanic crust despite extreme conditions, such as oligotrophy, temperature gradients, active circulation, and limited available space within the rock, which were once thought unsuitable to supporting life (Cowen et al., 2003; Nakagawa et al., 2006; Orcutt et al., 2011b; Jungbluth et al., 2013, 2014; Lever et al., 2013; Baquiran et al., 2016). Studies of ridge-flank systems have demonstrated that crustal aquifers harbor aerobic mesophiles and anaerobic thermophiles, involved in hydrogen, nitrogen, carbon, and sulfur cycling (Cowen et al., 2003; Nakagawa et al., 2006; Smith et al., 2011; Lever et al., 2013; Orcutt et al., 2013; Robador et al., 2015; Jungbluth et al., 2016). The comparison of biomes in very young (<3Ma) and old (80Ma) oceanic crust indicates that microbial diversity increases with the age of the basalt and the community compositions converge toward similar profiles over time (Lee et al., 2015). The colonization of rocks and the succession of microbial communities mainly depend on the temperature and reduction-oxidation conditions (Baquiran et al., 2016; Ramírez et al., 2019). While microbial communities hosted by young basalt (<10Ma) have been detected (Konhauser et al., 2002; Templeton et al., 2005), our knowledge of the pioneering communities inhabiting newly erupted oceanic basalt (<100years) is extremely limited.
A recent drilling operation has addressed this gap in knowledge through the study of the volcanic island, Surtsey, located on the southern offshore extension of the Icelandic volcanic rift zone (Figure 1). The island is the visible part of a volcano formed by underwater and basaltic eruptions from the seafloor between 1963 and 1967 (Þórarinsson, 1965; Jakobsson and Moore, 1982; Jakobsson et al., 2009). Since the initiation of eruptive activity, the entire area has been accessible only to scientific investigations. Surtsey is the site of long-term longitudinal studies, which have provided a unique record of pioneering species of plants and animals colonizing the surface of the basaltic deposits (Baldursson and Ingadóttir, 2007; Magnússon et al., 2014). In 1979, a 181m core (SE-01) drilled through the eastern sector of the Surtur vent (Figure 1), probed the hydrothermal system in the subaerial and the submarine deposits – above and below coastal sea level, respectively (Jakobsson and Moore, 1982, 1986; Jackson et al., 2019a). In 2017, three new cored boreholes (SE-02a, SE-02b and SE-03) were acquired through the International Continental Scientific Drilling Program (ICDP) 5,059 expedition, SUSTAIN drilling operation (Jackson et al., 2015, 2019b; Weisenberger et al., 2019). Annual monitoring of temperatures in the 1979 borehole, SE-01, indicates that the hydrothermal system has cooled down over the years. The maximal temperature in 1980 was 141.3°C at 100m depth, and it has decreased gradually to 124.6°C in 2017 (Figure 1; Jakobsson and Moore, 1986; Ólafsson and Jakobsson, 2009; Marteinsson et al., 2015; Jackson et al., 2019b). The current temperatures exceed a presumed upper limit for functional microbial life (122°C; Prieur and Marteinsson, 1998; Kashefi and Lovley, 2003; Takai et al., 2008). The highly porous subaerial and submarine tephra deposits were largely transformed to palagonitized lapilli tuff by 1979, described in studies of the SE-01 drill core (Jakobsson and Moore, 1982; Jakobsson and Moore, 1986). Progressive alteration was recorded in samples from a parallel, time-lapse drill core acquired in 2017, SE-02b (Figure 1; Prause et al., 2020). A recent study of geothermal water chemistries in the 1979 and 2017 Surtsey boreholes indicates depletion in boron, magnesium, iron, carbon dioxide, and sulfate concentrations, suggesting that the fluid compositions in the subsurface deposits are controlled by seawater-basalt interactions and temperature (Kleine et al., 2020). This further suggests that fluid-rock interactions in the submarine Surtsey basaltic deposits behave similarly to those interactions in basaltic oceanic crust, where the chemical composition of rocks and fluids changes and introduces organic matter and oxygen into the system (Furnes and Staudigel, 1999; Edwards et al., 2005; Orcutt et al., 2011a). The Surtsey volcano geothermal system represents an exceptional natural laboratory for studying fluid-rock-microbe interactions at temperatures approaching the presumed thermal limit for functional life on Earth. Its boreholes can be viewed as windows opened from the land surface that allow the study of subsurface processes at high temperature associated with the basaltic oceanic crust.
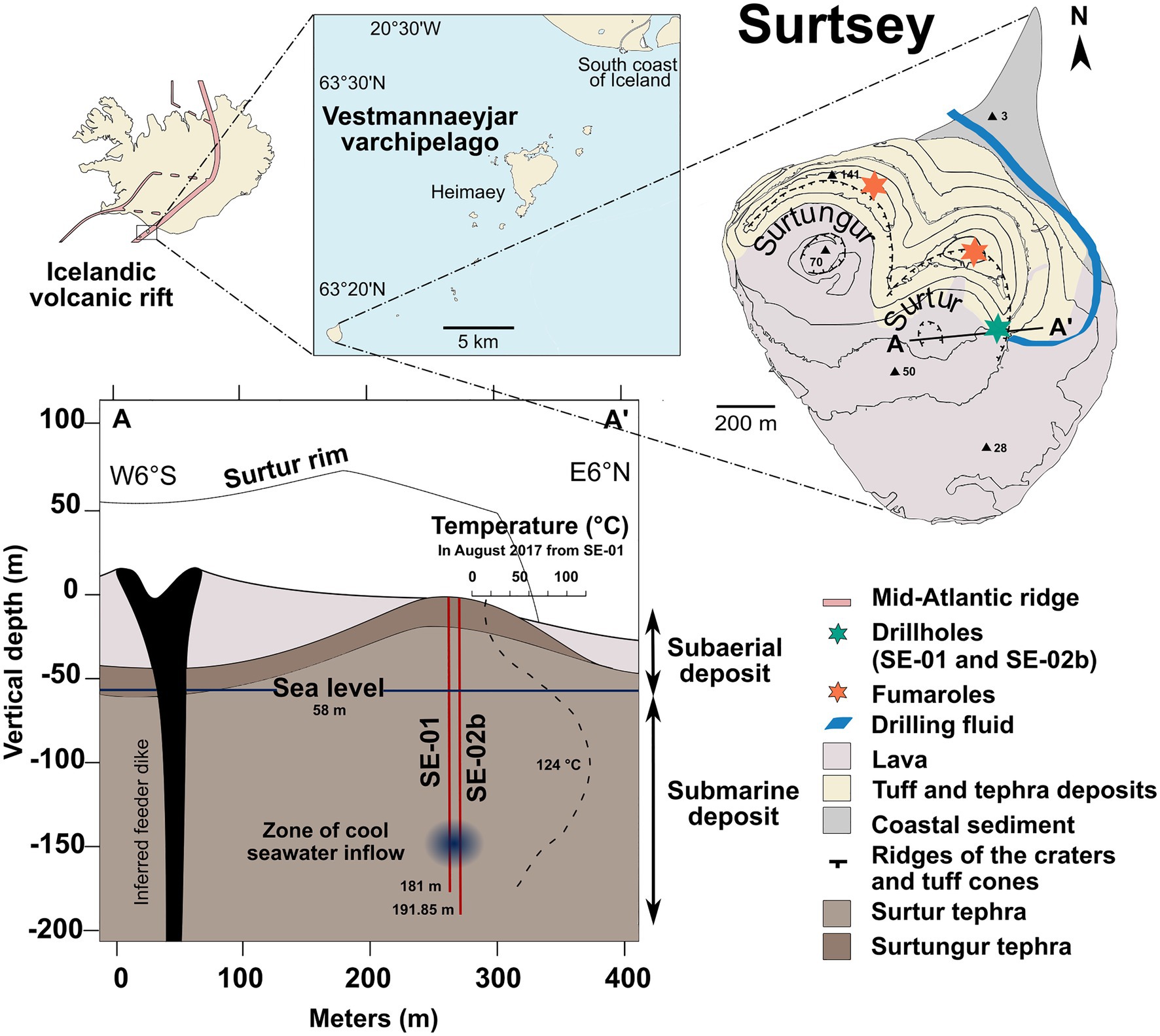
Figure 1. Location of the study area of the south coast of Iceland. Map of the Surtsey volcanic island within the Westman Islands (Vestmannaeyjar) archipelago and location of sampling sites: 1979 borehole SE-01 and 2017 borehole SE-02b (green star), drilling fluid (blue line), and fumaroles (orange stars). Schematic cross section of Surtur crater showing the geological setting of the subsurface deposits and the location of the time-lapse cored boreholes (see Jackson et al., 2019b) and a zone of seawater inflow.
Several studies have described the volcanic structure of Surtsey and the geochemical, chemical, and mineralogical changes in the altered basaltic deposits (Jakobsson and Moore, 1982; Jakobsson and Moore, 1986; Schipper et al., 2016; McPhie et al., 2020; Moore and Jackson, 2020; Prause et al., 2020). Little is known, however, about the microorganisms inhabiting the subsurface and their metabolic potential. A pioneering study by Marteinsson et al. (2015) detected archaea of the Archaeoglobus genus at 145m depth (80°C) and one taxon from the Methanobacteriales order in the SE-01 borehole at depths to 170m (55°C; Marteinsson et al., 2015). Furthermore, recent studies of 1979 and 2017 drill core samples show abundant microtubules in basaltic glass that resemble endolithic microborings (Jackson et al., 2019a; Jackson, 2020); these are thought to indicate microbial microboring into glass (Fisk et al., 2003; Staudigel et al., 2008; Walton, 2008; Mcloughlin et al., 2010). These investigations suggest that Surtsey’s basalt and fluids could reveal new information about the microbial communities associated with the young hydrothermal-seawater system and the newly formed oceanic basaltic crust.
This work focuses on the following questions: (i) how can we distinguish the true residents of the subsurface from the marine microbial taxa derived from the drilling fluid? (ii) which microorganisms inhabit Surtsey’s basalt and fluids? (iii) does life persist in the subsurface zones crossing the presumed thermal limit for life? and (iv) can we detect actual microbial structures or biotic signatures in the basaltic cores only 50years after eruptions terminated? We used high-throughput sequencing (16S rRNA gene amplicon sequence analysis) and scanning electron microscopy (SEM) to address these questions and enable an in-depth characterization of the basalt-hosted microbial communities within the volcanic system. Microbial communities from drill core samples collected at successive depths from the subaerial and submarine basaltic tephra deposits were compared to the communities in borehole fluid samples associated with the active hydrothermal system and in fumarole samples from the surface of the island and seawater samples collected several kilometers offshore. In this study, it was of crucial importance to collect drilling fluids, sequence control samples representing potential sources of contamination, and implement computational methods to distinguish the endemic microbial taxa of the subsurface rocks from the microorganisms found in the drilling fluid and other potential contaminants. Here, we report our strategy to identify and minimize contaminants in the data sets; we describe the archaeal and bacterial taxa that are candidate residents of the oceanic crust; we discuss the microbial colonization and dissemination from the surrounding ecosystems, and we explore microbial metabolic potentials. The results provide unique data on subsurface microbial life in one of the newest and most pristine oceanic basalt environments on Earth.
The volcanic island of Surtsey is located in the Vestmannaeyjar archipelago, approximately 35km from the south coast of Iceland (63°18'10.8'N; 20°36'16.9'W; Figure 1) within the southern offshore extension of Iceland’s Eastern Volcanic Zone. The ocean depth was 130m below sea level prior to eruption. After 3.5years of submarine, explosive, and effusive lava-producing eruptions of basalts from the seafloor in 1963–1967, Surtsey had a subaerial area of 2.6km2, and the highest point on the island was 174m above sea level. The island has now eroded to less than 1.2km2 and a height of 150m above sea level (Baldursson and Ingadóttir, 2007; Óskarsson et al., 2020).
The cores extracted from the SE-01 borehole in 1979 and the SE-02b borehole in 2017 indicate that the subsurface of Surtsey consists mainly of lithified tephra, mainly lapilli tuff, and minor amounts of weakly consolidated tephra and alkali olivine basalt intrusions (Jakobsson and Moore, 1982; McPhie et al., 2020; Figure 1). The principal authigenic minerals in the lapilli tuff are smectitic clay mineral (nontronite and clinochlore), analcite, phillipsite, Al-tobermorite, and anhydrite (Jakobsson and Moore, 1986; Jackson et al., 2019a; Prause et al., 2020). Alteration through palagonitization and production of authigenic cementitious minerals has progressed during the past 38years, from 1979 to 2017 (Jackson, 2020; Prause et al., 2020). In situ subsurface fluid temperatures have been measured annually since 1980 in SE-01, usually at 1-m intervals from the surface to the bottom of the borehole (e.g., Jakobsson and Moore, 1986; Marteinsson et al., 2015). Geochemical analyses and pH measurements of borehole fluids and pore water extracted from the 2017 drill cores are described by Kleine et al. (2020).
One of the principal objectives of the 2017 Surtsey Underwater volcanic System for Thermophiles, Alteration processes and Innovative concretes (SUSTAIN) drilling project at Surtsey volcano, sponsored in part by the ICDP, was to investigate microbial diversity within the basaltic tephra. All possible precautions were made to avoid microbial contamination (Jackson et al., 2019b). Water from the sea was pumped to the drill site and used as drilling fluid since no fresh water is available on the island (Figure 1). The conventional methods used to track contamination during drilling operations, such as the addition of tracer compounds in the circulating fluid, could not be applied during drilling at Surtsey due to the strict environmental protection of the island (see discussion). To avoid contamination, drilling fluid was sterilized using two filtration units with a pore size of 30μm (Pentek Big Blue, R30-BB 30 Micron cartridge filter, Lenntech, The Netherlands) and two ultraviolet sterilization devices (AQUA4ALT from WEDECO, Aquaculture systems, Xylem Water Solutions Herford GmbH, Germany) with a maximum flow rate of 1.58ls−1. The decontaminated drilling fluid was stored in 1000L containers before pumping into the borehole. To track potential problems with the sterilization system, 1l of drilling fluid was collected regularly at different drilling depths to assess potential microbial contaminants (Figure 1).
Four types of samples were collected for molecular investigation and comparison of microbial diversity among these sample types: drill cores from the 2017 SE-02b borehole, borehole fluids from the 1979 SE-01 borehole, steam from surface cracks of fumaroles, and seawater samples collected few kilometers from the northwest coast of Surtsey (Figure 1). A description of the samples is shown in Table 1.
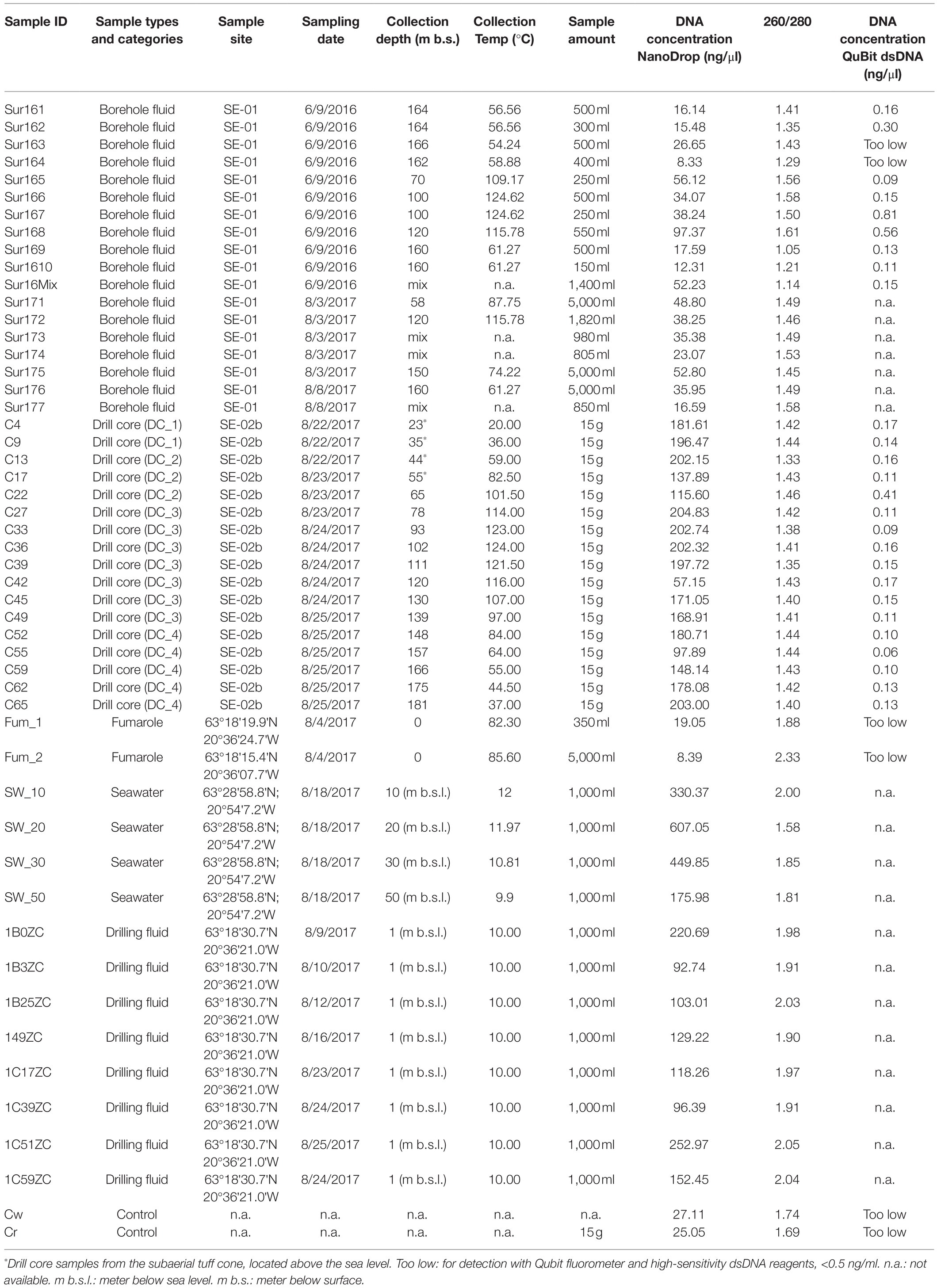
Table 1. Sample description table: locations, sampling date, depth, temperature, and DNA concentration.
Seventeen drill core samples were collected for microbial analyses from the vertical SE-02b cored borehole that extends to 192m below surface and terminates in poorly consolidated tephra a few meters above the presumed depth of the pre-eruption seafloor (Jackson et al., 2019b). At the drill site, drill core samples were collected from every third 3-m core run for molecular analyses by cutting a 10-cm section at 70cm from the top of the core run. Immediately after sampling, each section was kept in the plastic core liner, taped at both ends, wrapped in a plastic bag, kept in liquid nitrogen on site, and at −80°C for long-term laboratory storage. Fluid samples from SE-01 borehole were collected in 2016 and in 2017, before drilling operations started, using a custom sampler made of stainless steel, as described in Marteinsson et al. (2015). The sampler was rinsed with 70% ethanol before each sampling. Eighteen fluid samples were collected from the SE-01 borehole: 18 in 2016 and seven in 2017 (Table 1). Steam from fumaroles, located on the summit of Surtur, the eastern tephra cone (~150m above sea level; 63°18'15.4'N 20°36'07.7'W) and Surtlungur, the western tephra cone (63°18'19.9'N 20°36'24.7'W) were collected in 2017 by introducing a sterile rubber hose into the outlet of the fumarole, with the other end connected to a sterile plastic container. This generated 5,350ml of condensed water from the fumaroles over 12h of continuous sampling. Four liters of seawater samples were collected 25km offshore during the drilling operation (63°28'58.8'N; 20°54'7.2'W). All water samples (drilling fluid, borehole fluid, fumarole, and seawater samples) were immediately filtrated through 0.22-μm Sterivex™ filters (Merck Millipore). The filters were stored in liquid nitrogen on site and at −80°C for long-term storage in the laboratory.
A modified PowerMax® Soil DNA Isolation Kit protocol (MO BIO Laboratories, Inc.) was applied to extract the DNA from the drill core samples. Small fragments of tuff from the interior of the 17 frozen core samples from SE-02b were broken aseptically. After 2min pre-cooling on ice, 15g were cryo-ground at an impact rate of 8cycles per second for 1min using a 6,700 Freezer/Mill cryogenic grinder (SPEX). Phosphate-ethanol solution (1M phosphate buffer, 15% ethanol, pH 8.0; Direito et al., 2012) and proteinase K (20mg/ml) were added to the lysis solution provided by the kit. Samples were vortexed twice for 30s with a 1min cooling step in between instead of the bead-beating step, followed by incubations at 55°C for 1h and at 80°C for 40min. Subsequent DNA isolation was carried out according to the standard PowerMax® Soil DNA isolation protocol. DNA precipitation was done overnight with isopropanol (0.7 volume) and glycogen (20mg/ml). After a washing step with 70% ethanol, DNA was resuspended in Tris buffer (10mM, pH 8) and quantified using both NanoDrop® ND-1000 UV-Vis Spectrophotometer (Thermo Fisher Scientific) and a Qubit fluorometer and high-sensitivity dsDNA reagents (Invitrogen™) before being stored at −20°C. Attempts to estimate the microbial biomass within the drill core samples using fluorescence microscopy were unsuccessful because of the non-biological background signals that occurred during recognition of cells for counting. Two controls were carried out to test for contamination during DNA extraction of the core samples. One used sterile water (instead of the basalt powder) to test for contaminants derived from the kit and reagents. The other used 15g of basalt powder from a drill core that had been treated with 70% ethanol and heated at 180°C for 24h. Both controls resulted in amplifications with primers targeting bacterial and archaeal 16S rRNA genes, which were then sequenced.
DNA was extracted from Sterivex™ filters containing biomass from borehole water, condensed fumarole water and drilling fluid samples following a modified protocol by Neufeld et al. (2007). Sucrose EDTA Tris buffer (SET buffer: 40mM ethylenediaminetetraacetic acid (EDTA), 50mM Tris-HCl pH 9, and 0.75M sucrose) and 20mg/ml lysozyme solution were added to the Sterivex™ filters. Filters were incubated at 37°C for 30min. After the addition of 10% (w/v) sodium dodecyl sulfate (SDS) and proteinase K (20mg/ml), the filters were incubated with rotation at 55°C for 2h. Lysates were collected into syringes, while the filters were rinsed twice with SET buffer and the rinsed buffer was combined to the lysate. One volume of phenol:chloroform:isoamyl alcohol (PCI: 25:24:1, pH 8) was added, and the aqueous phase was transferred to a new tube after a 15-min centrifugation at 10,000×g at 4°C. The phenol was removed from the aqueous phase by adding 1 volume of chloroform. The cleaned aqueous phase was transferred to a new tube after a 5-min centrifugation at 10,000×g at 4°C, and 0.7 volume of cold isopropanol was added. After inverting the tube several times, the sample was incubated for 15min at room temperature and then overnight at −20°C. After a 20-min centrifugation at 16,000×g at 4°C, the DNA pellet was washed twice with 75% (v/v) ethanol, dried for 5min using a SpeedVac™ and 15min at room temperature, and finally resuspended in sterile Tris-HCl buffer (10mM, pH 8). DNA was quantified using a NanoDrop® ND-1000 UV-Vis Spectrophotometer (Thermo Fisher Scientific) and Qubit fluorometer (Invitrogen, Quant-iT™ dsDNA HS) and stored at −20°C. One negative extraction control was carried out for each round of extractions by rinsing a sterile Sterivex™ filter with sterile laboratory-grade water.
Illumina MiSeq paired-end (2×300 base pair) tag sequencing was carried out using the Earth Microbiome Project universal primers 515f (5'-GTG CCA GCM GCC GCG GTA A-3') and 806r (5'-GGA CTA CHV GGG TWT CTA AT-3'), which amplify the V4 region of the bacterial and archaeal 16S rRNA genes (Caporaso et al., 2012). Since this primer pair does not show a high coverage for Archaea (Parada et al., 2016), specific archaeal primer sets were used with a nested PCR approach. The first-round PCR was performed to amplify the V3-V5 region of archaeal 16S rRNA gene with the primer set Parch340F (5'- CCC TAY GGG GYG CAS CAG -3'; Øvreås et al., 1997) and Arch958VR (5'- YCC GGC GTT GAV TCC AAT T -3'; Klindworth et al., 2013). Then, a second round was performed on the first PCR product to amplify the V3 region of archaeal 16S rRNA gene with the primer set Arch349F (5'- GYGCASCAGKCGMGAAW -3'; Takai and Horikoshi, 2000) and Parch519R (5'- TTACCGCGGCKGCTG -3'; Øvreås et al., 1997).
All PCR reactions were carried out in 25μl reactions with 20μl of Q5® High-Fidelity PCR Master Mix (New England Biolabs, MA, United States) following the manufacturer’s amplification protocol and 5μl of DNA at 10ng/μl (NanoDrop quantification). The second round of the nested PCR used 5μl of amplified DNA from the first round. Bovine serum albumin was added to the master mix at a final concentration of 0.5ng/μl for rock samples that could not be amplified. Thermal cycling consisted of an initial denaturation step at 98°C for 30s, followed by 30cycles of denaturation at 98°C for 10s, annealing at 52°C (using the bacterial primer set) or at 55°C (using the archaeal primer sets) for 30s, and elongation at 72°C for 60s. Final elongation was set at 72°C for 2min. Amplification products were visualized on 1% (w/v) agarose gels.
Sequencing libraries were generated using the “16S Metagenomic Sequencing Library Preparation guide” from Illumina and barcodes from the Illumina Nextera®XT DNA Sample Preparation Kit (8cycles for index PCR). The libraries were assessed on a Qubit Fluorometer (Invitrogen, Quant-iT™ dsDNA HS) and a Bioanalyzer system (Agilent Technologies). After normalization and quantification, the final pooled library was loaded on a MiSeq Desktop sequencer (Illumina) and sequenced with V3 chemistry and 2×300cycles across two sequencing runs. Raw sequences have been deposited in the European Nucleotide Archive (ENA) at EMBL-EBI under accession number ERP126178.
Bioinformatic analysis was conducted in RStudio running R version 4.0.2 (R Core Team, 2019; Rstudio, 2020). Sequence variants were inferred using the R Package DADA2 (Callahan et al., 2016) version 1.4, available at https://benjjneb.github.io/dada2/tutorial.html. The following trimming parameters were used: trimLeft=10, maxN=0, maxEE=c(2,5), truncQ=2, and truncLen=c(230,215) for the universal primer set and truncLen=c(115,110) for the archaeal primer sets. After quality screening and trimming, forward and reverse reads were merged to remove chimeric variants and singletons and to identify amplicon sequence variants (ASV). Non-target-length sequences were removed, and only amplicons of 270–275bp length were kept with the universal primer set and amplicons of 130–165bp for the archaeal primer sets. Data sets from three MiSeq sequencing runs (universal primer set) were processed separately using the same pipeline and same parameters and merged into a unique ASV table using the function “mergeSequenceTables.” The taxonomy was assigned using the function “assignTaxonomy” (minimum bootstrap confidence at 50) and the SILVA SSU database release 138 (Quast et al., 2013).
In total, all 51 samples were successfully sequenced using the universal primer set, including extraction controls and drilling fluid samples. The R package Decontam (version 1.10.0; Davis et al., 20181) was used to identify contaminant ASVs in the universal primer data set. We identified 160 contaminant ASVs (Supplementary Table S1) using the Decontam prevalence method (threshold value of 0.5), based on the prevalence comparison of each sequence in true samples and negative controls (Supplementary Figure S1). In addition, sequences that were identified as chloroplast at the order level, mitochondria at the family level, Eukaryote at the kingdom level, and those that could not be identified at the kingdom level were subtracted from the data set prior to analysis, as well as putative contaminants identified by taxonomic affiliation at the genus level by a study on common contaminants from the Census of Deep Life data set (Sheik et al., 2018). Supplementary Table S2 gives a list of 95 potentially contaminant genera removed from the analyses. A total of 588,510 sequences were removed from the samples by the abovementioned procedures (Details are available in Supplementary Table S3).
From the archaeal nested PCR, 17 samples were analyzed. Sequences identified at the kingdom level as Eukaryote, Bacteria, or not assigned were removed from libraries prior to analysis, as well as sequences detected in the DNA extraction blanks (Supplementary Table S4, 26 ASVs). Using the archaeal primer sets, the number of sequences removed from the samples was 74,315 (Details available in Supplementary Table S3).
Microbial community analysis (α and β diversity, community composition, and statistical analysis) was conducted in R (R Core Team, 2019) with the Phyloseq (McMurdie and Holmes, 2013) and Vegan (Oksanen et al., 2007) packages. ANOVA and Tukey’s HSD (Honestly Significant Difference) test were conducted to evaluate the differences in α diversity values. For β diversity assessment, the data were normalized using “rarefy_even_depth” function prior to performing a non-metric multidimensional scaling (NMDS) ordination on Bray-Curtis dissimilarities. The significance of sample type variable was assess using permutational multivariate analysis of variance (PERMANOVA) using distance matrices and multilevel pairwise comparison (Martinez Arbizu, 2020). The command envfit was used to investigate the correlation between the community structure and environmental variables (depth and temperature). Finally, DESeq2 was used to identify ASVs significantly different among sample types (F, fumarole; BF, borehole fluid; DC, drill core; SW, seawater samples) and categories of drill cores (DC_1, DC_2, DC_3 and DC_4; Love et al., 2014). Details of the data analyses can be found in the Supplementary Material. Predictive functional analyses of the prokaryotic communities were performed using Phylogenetic Investigation of Communities by Reconstruction of Unobserved States 2 (PICRUSt2) on both data sets, obtained with the universal primer set and the archaeal primer sets, separately (Douglas et al., 2020). From the universal data set, a total of 2,333 PICRUSt2 predicted KEGG orthologs (enzymes) were collapsed into 425 MetaCyc pathways, while 828 predicted KEGG orthologs and 124 MetaCyc pathways were obtained from the archaeal data set. Only few MetaCyc pathways were selected to represent sulfur, nitrogen, methane, and carbon metabolism (Supplementary Figures S8, S9).
For SEM images, drill core samples were crushed in a sterile mortar to obtain rock grains <0.5mm diameter, which were dehydrated by four 10-min wash steps in increasing concentrations of ethanol (30, 50, 80, and 100%). After drying, samples were placed on carbon conductive tabs (PELCO Tabs™, 9mm) and gold-coated. SEM used a Zeiss Auriga 40 Focused Ion Beam Field Emission Scanning Electron Microscope coupled with an energy dispersive X-ray spectroscopy analyzer (EDX) at the Institute de Physique du Globe de Paris (University Sorbonne Paris Cité, Paris, France) using two types of secondary electron detectors: In-Lens and SESI, and a backscattered electron detector: EsB. The acceleration voltage (EHT) ranged from 5 to 15kV.
Based on Qubit quantification, DNA concentration extracted from 15g of each drill core sample yields from 0.2 to 1.36ng.g−1 of rocks with an average of 0.48ng.g−1. However, no significant correlations in DNA yield were apparent with sampling depth or in situ temperature (Supplementary Figure S2).
The island of Surtsey is subjected to strict environmental protection and is a UNESCO World Heritage site (Baldursson and Ingadóttir, 2007). For this reason (and other reasons mentioned later in the study), the use of tracer compounds to track contamination could not be considered during the design of the 2017 SUSTAIN drilling operation. Instead, a different approach was used to assess contamination. Filtered and UV-sterilized water pumped from the sea was used as drilling fluid (see above). Aliquots of those fluids were then collected to track microbial DNA potentially contaminating the core. DNA extracted from 1l of each drilling fluid sample yields ~96 to 252ng/μL of DNA (Table 1).
The 16s rRNA gene sequencing of the drilling fluid samples thus showed, unfortunately, that the sterilization system failed (Supplementary Table S3, Supplementary Figure S3). Consequently, marine microbial taxa from the drilling fluid were introduced into the subsurface. The failure can possibly be explained by the clogging of filters and an overly high flow rate for the UV system.
As the contamination of the samples was evidently pervasive, the data were evaluated with caution. A NMDS ordination plot conducted using Bray-Curtis dissimilarity metrics (stress value=0.124) revealed that some of the drill core samples (e.g., C45, C52, C65) showed a microbial community composition similar to the drilling fluid and seawater samples (Supplementary Figure S4). Whereas differences in microbial community structure between drill core and drilling fluid and between drill core and seawater samples proved to be significant by a Tukey HSD test (p adjusted values of 0.01 and 0.0008, respectively), a strategy was adopted to distinguish endemic microbial taxa of Surtsey subsurface rocks from the marine residents introduced to the subsurface by the drilling fluid. The distributions of individual ASVs were evaluated by a simple overlap approach using a Venn diagram to compare ASVs shared among the seawater samples, drilling fluid, drill core from the subaerial deposits, and from the submarine deposits (Figure 2).
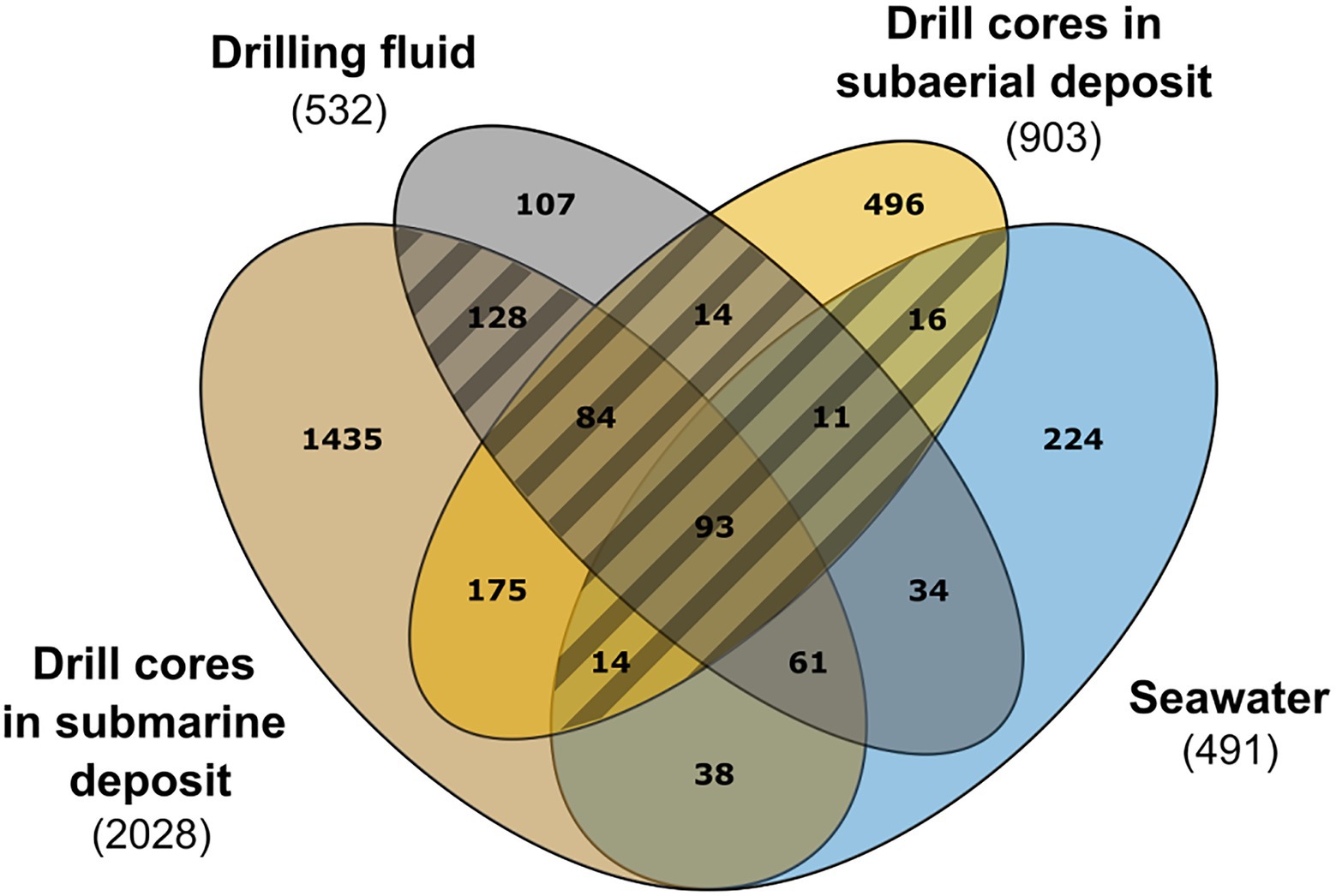
Figure 2. Contamination assessment by an overlap approach between drill cores from the subaerial and submarine deposits, drilling fluid, and seawater samples. Five drilling fluid samples (in grey): 1B49ZC, 1C17ZC, 1C39ZC, 1C51ZC, 1C59ZC, 4 sea samples (in blue): SW_10, SW_20, SW_30, SW_50, 4 samples from the subaerial deposit (in yellow): C4, C9, C13 and C17, and 13 samples from the submarine deposit (in light brown): C22, C27, C33, C36, C39, C42, C45, C49, C52, C55, C59, C62, C65. Hatched area: 360 ASVs considered as contaminants.
In the subaerial tephra deposits, located above coastal sea level, meteoric water was presented before drilling. In the zone of tidal flux at ~58m b.s., temperatures up to 100°C could operate as a natural autoclave or biological barrier, preventing transfer of live microorganisms between the subaerial and submarine basaltic deposits (Ólafsson and Jakobsson, 2009; Marteinsson et al., 2015). Hence considering that no marine taxa should be detected in the subaerial deposit, all ASVs shared between the subaerial samples (C4, C9, C13, and C17; 23–55m b.s.; 20–82.5°C), the drilling fluid, and the seawater samples (Figure 2: 93+84+14+11+16+14 ASVs, Supplementary Table S6) were considered as potential marine contaminants; these sequences were removed from the data set. In a second iteration, ASVs shared only between the drill core samples located in the submarine deposits and the drilling fluid (Figure 2, Supplementary Table S6, 128 ASVs) were also considered as marine contaminants and were removed from the data set. Natural infiltration of cool seawater occurs in the subsurface of Surtsey at 144–155m b.s (Jakobsson and Moore, 1986; Jackson et al., 2019b; Kleine et al., 2020). This suggests that marine taxa detected in the submarine deposit could derive from the inflow of seawater that infiltrate the subsurface. Therefore, ASVs shared between the submarine drill core, seawater samples, and the drilling fluid were retained (Figure 2, 61 ASVs). The total number of sequences removed from the drill core samples by the abovementioned procedures was 138,913 (Supplementary Table S3), which represents on average 47% of the reads per drill core samples. The subsequent analyses were performed on the decontaminated data set, excluding the drilling fluid samples.
Sequencing provided enough reads to capture the total richness of the samples as all libraries reached saturation in rarefaction curves (Supplementary Figure S5). The 41 samples were catalogued by sample type: 18 samples from borehole fluids, 17 samples from drill cores, 2 samples from fumaroles, and 4 seawater samples (Table 1, Supplementary Table S3). Amplification of the partial 16S rRNA gene using the universal primer set was successful for all samples, while only 17 samples could be amplified using the archaeal primer sets.
We obtained a total of 455,545 and 848,053 high-quality sequences using universal and archaeal 16S rRNA primer sets, respectively (Supplementary Table S3). A total of 4,222 ASVs ranging from 41 (Sur168) to 317 (SW50) ASVs per sample were obtained using the universal primer set, whereas a total of 157 ASVs ranging between 3 (Sur4a) and 41 (Sur1a) ASVs per sample were obtained using the archaeal primer sets.
Analysis of variance on species richness showed significant differences between sample types (ANOVA, F value=9.082, Pr(>F)=0.000123). A Tukey HSD test highlighted significant differences among them between seawater samples and borehole fluid (<0.001), seawater samples and drill core (0.0339), and between drill core and borehole fluid (0.0184). Shannon diversity also differed significantly among them (ANOVA, F value=5.998, Pr(>F)=0.00195), and Tukey’s HSD test revealed significant differences between seawater samples and borehole fluid (0.00323) and between seawater samples and drill core (0.02570; Figure 3A, Supplementary Table S7). The SE-01 borehole fluids displayed a significantly lower observed diversity than the drill cores, but Shannon diversity was not significantly different, indicating that the evenness of the species present in these sample types was comparable. Observed diversity and evenness were significantly higher in the seawater samples than in drill core and borehole fluid samples.
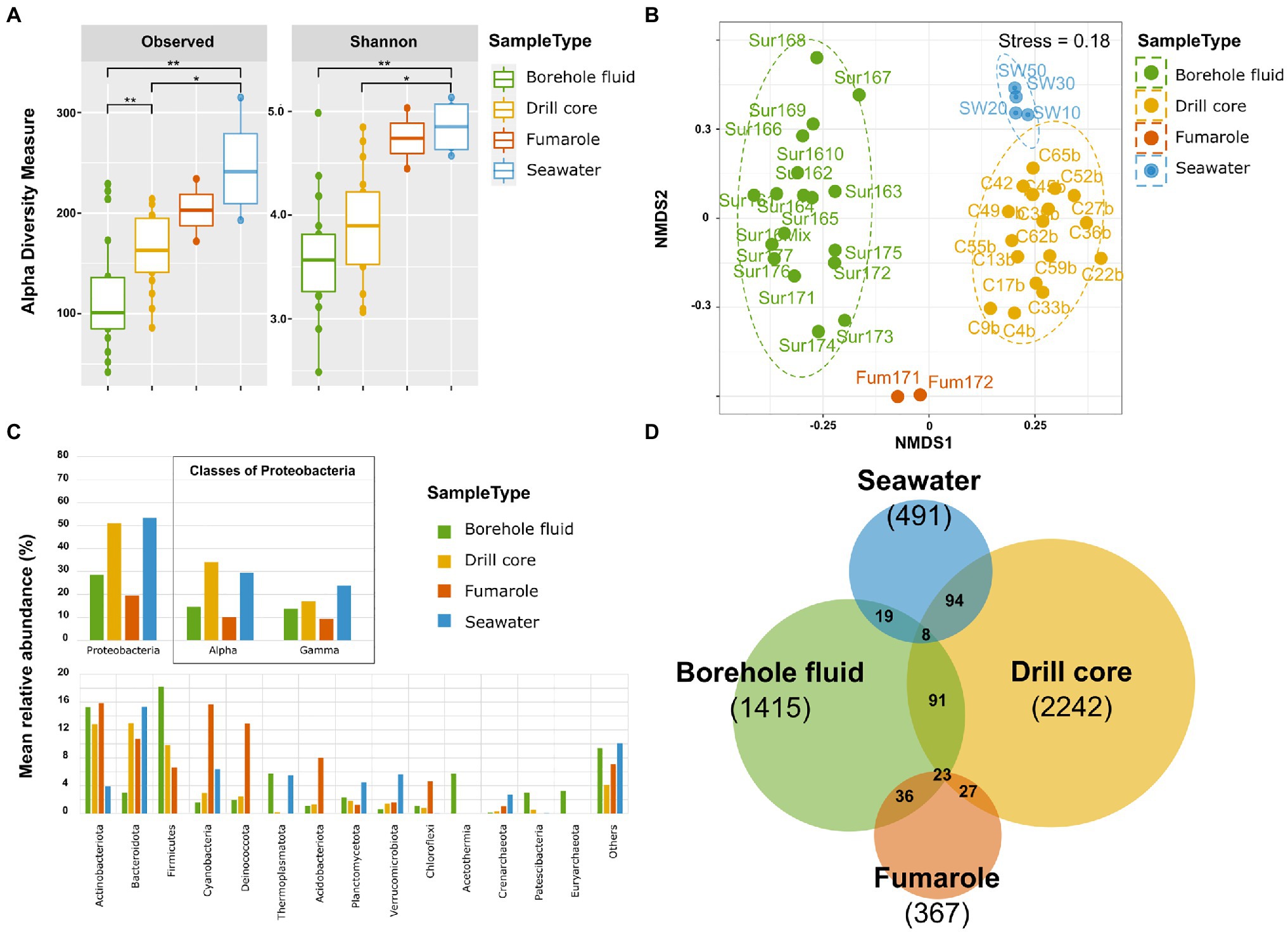
Figure 3. Overview of the microbial community diversity, structure, and composition of the samples types: borehole fluid, drill core, fumarole, and seawater samples. (A) α diversity indices: observed diversity and Shannon’s diversity indexes. Signif. codes: 0.001 “**“0.01 “*”. (B) β diversity assessed with non-metric multidimensional scaling ordination using Bray-Curtis dissimilarity metrics of microbial community within sample types. The stress value equaled 0.18. (C) Mean relative abundance of the 14 most abundant phyla. Proteobacteria are divided into classes. (D) Venn diagram showing the number of shared and unique ASVs among the sample types.
The resulting NMDS ordination conducted using Bray-Curtis dissimilarity metrics revealed distinct microbial communities specific to each sample types, which clustered individually (Figure 3B). The stress value equaled 0.18. This was further confirmed by a PERMANOVA analysis showing that sample types differed significantly [Pr(>F)=0.001, r2=274], and a multilevel pairwise comparison showed significant differences among drill cores and borehole fluids (adjusted p value=0.0184), seawater samples and borehole fluids (<0.001), and between seawater samples and drill cores (0.0338). This indicates that the samples within a given sample type show microbial communities that are more similar to one another than to samples from a different sample type. The cluster of SE-01 borehole fluid samples appears wider than the drill core cluster, indicating more differences in microbial community (Figure 3B).
At the phylum level, some taxa were common across the four sample types (Figure 3C). The phylum Proteobacteria dominated all sample types, for example, whereas its relative proportion varied greatly among them. Within Proteobacteria, Alphaproteobacteria was the most abundant class followed by Gammaproteobacteria. Fumarole samples showed a higher relative proportion of Cyanobacteria (15.3%), Deinococcota (12.6%), Acidobacteriota (7.84%), and Chloroflexi (4.52%) than borehole fluids and drill cores. Borehole fluids displayed the highest occurrence of the bacterial phyla Acetothermia (5.56%) and Patescibacteria (2.26%), as well as the archaeal phyla Thermoplasmatota (5.60%) and Euryarchaeota (3.13%), in comparison with the other sample types. The class Desulfotomaculia represented more than 5% of the borehole fluid community (data not shown). Nevertheless, due to the compositional nature of the data, lineage-specific enrichments necessitate further investigation, using for example a qPCR approach (Jian et al., 2020). The drill core samples show the highest sequence variants (2,242 ASVs), followed by the borehole fluids (1,415 ASVs), seawater samples (494 ASVs), and fumarole samples (367 ASVs). Most ASVs were not shared between the sample types (Figure 3D).
The bacterial sequences from the drill core samples are classified into 35 phyla, with the ten phyla Proteobacteria, Bacteroidota, Actinobacteriota, Firmicutes, Cyanobacteria, Deinococcota, Planctomycetota, Verrucomicrobiota, Acidobacteriota, and Chloroflexi, comprising more than 95% of the sequences (Figure 4). The 29 ASVs classified as Archaea using the universal primer set fall into the phyla Thermoplasmatota, Crenarchaeota, Nanoarchaeota, and Iainarchaeota.
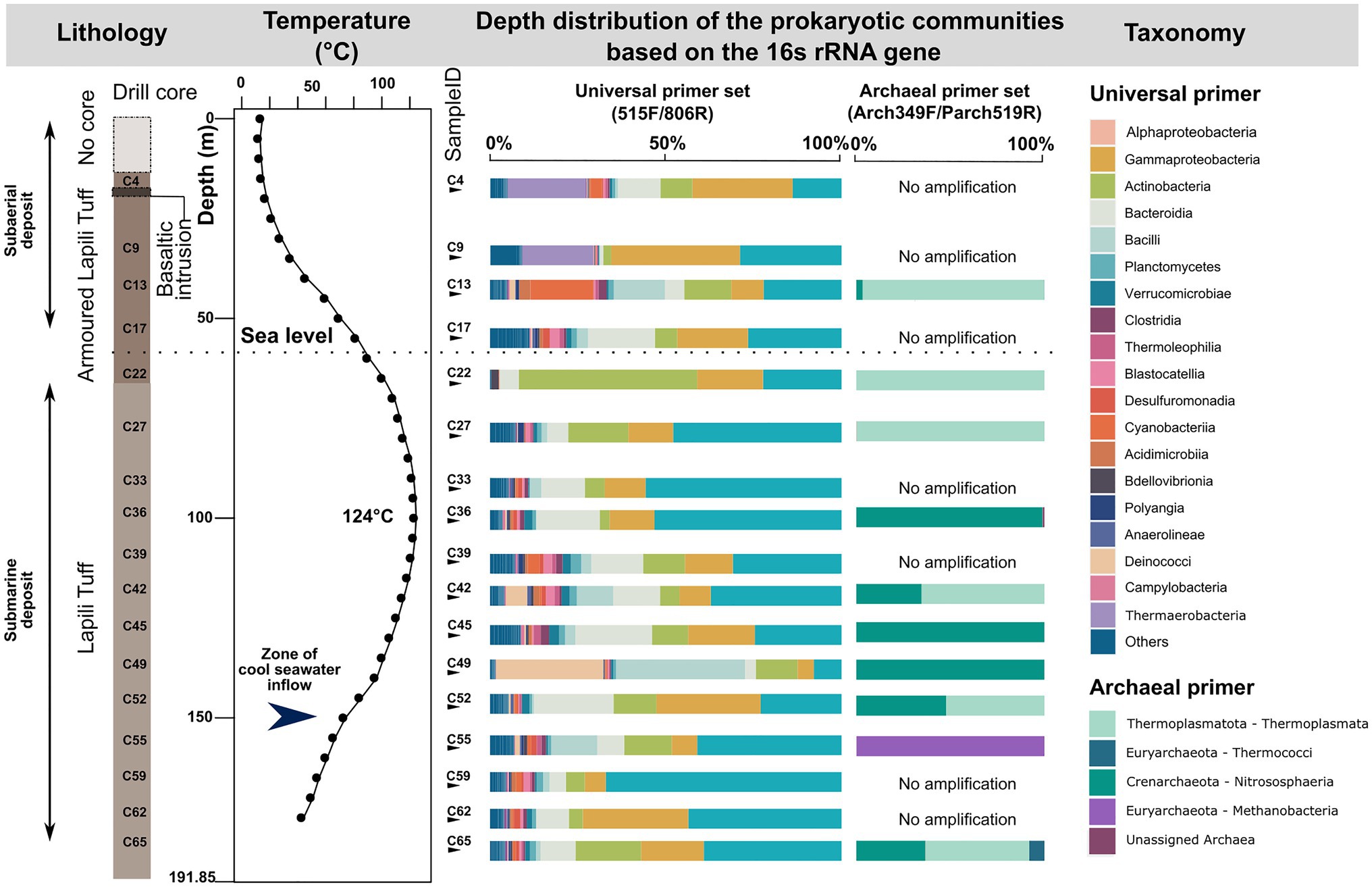
Figure 4. Vertical distribution of the microbial communities at the phylum or class level detected in the 17 drill core samples from the subsurface of the Surtsey volcano using universal primer set and archaeal primer sets.
Archaeal sequences obtained by nested PCR from 10 of the drill cores were more diverse. The 88 unique ASVs include the phyla Crenarchaeota (mainly Nitrososphaeria), Thermoplasmatota (mainly Thermoplasmata), Euryarchaeota, Halobacterota, Hydrothermarchaeota, Nanoarchaeota, and unassigned Archaea (Figure 4). Remarkably, the sample C55 (157m b.s.; 64°C) below the submarine inflow zone is dominated by the archaeal genus Methanobacterium, while the sample C65 (181m b.s.; 37°C) in weakly consolidated tephra near the pre-eruption seafloor shows the highest abundance of the genus Thermococcus (Figure 4).
Canonical correspondence analysis (CCA) ordination (ANOVA, p=0.018) and envfit analyses demonstrate that both depth and in situ temperature are significantly correlated with microbial community structure of the drill core samples (Supplementary Figure S6). However, it is unclear which variable has the most influence on the microbial community structure, since both are linked. Observed and Shannon diversity indices showed no significant difference between the four categories of drill core samples (data not shown).
To identify the ASVs causing the dissimilarity of community structure in the CCA ordination plot, the drill core samples are grouped in four categories as follows (Supplementary Figure S6): (DC_1) drill core samples from the subaerial deposits (23–35m b.s.; 20–36°C in 2017; n=2, C4, C9) (DC_2) drill core samples from the zone of daily intertidal fluctuations at coastal sea level (44–65m b.s.; 59–101.5°C; n=3, C13, C17, C22), (DC_3) drill core samples from the submarine deposits near the hydrothermal temperature maximum (78–139m b.s.; 97–124°C in 2017; n=7, C27, C33, C36, C39, C42, C45, C49), and (DC_4) drill core samples from the submarine deposits, below the zone of seawater inflow (157–181m b.s.; 37–84°C in 2017; n=5, C52, C55, C65, C62, C59).
To identify differences in ASVs abundance between the four sample types (F, fumarole; BF, borehole fluid; DC, drill core; SW, seawater samples), we performed differential abundance analyses (two-by-two comparisons) using separately the universal primer and the archaeal primer data sets (Supplementary Table S5). Additionally, the same analyses were performed for the four categories of drill core samples to highlight the ASVs contributing to the dissimilarity in the CCA plot (Supplementary Figure S6). Due to the compositional features of the data, the identification of differentially abundant taxa between the different groups of samples must be assessed carefully, since it is based on a count-based method. Being aware of the limitations of this approach, a total of 95 and 26 unique ASVs were proved to be significantly represented using the universal primer data set (grouping into 59 taxonomic bins) and the archaeal primer sets (7 taxonomic bins), respectively (Figure 5), in accordance with the log fold change of the mean normalized read counts (p<0.01).
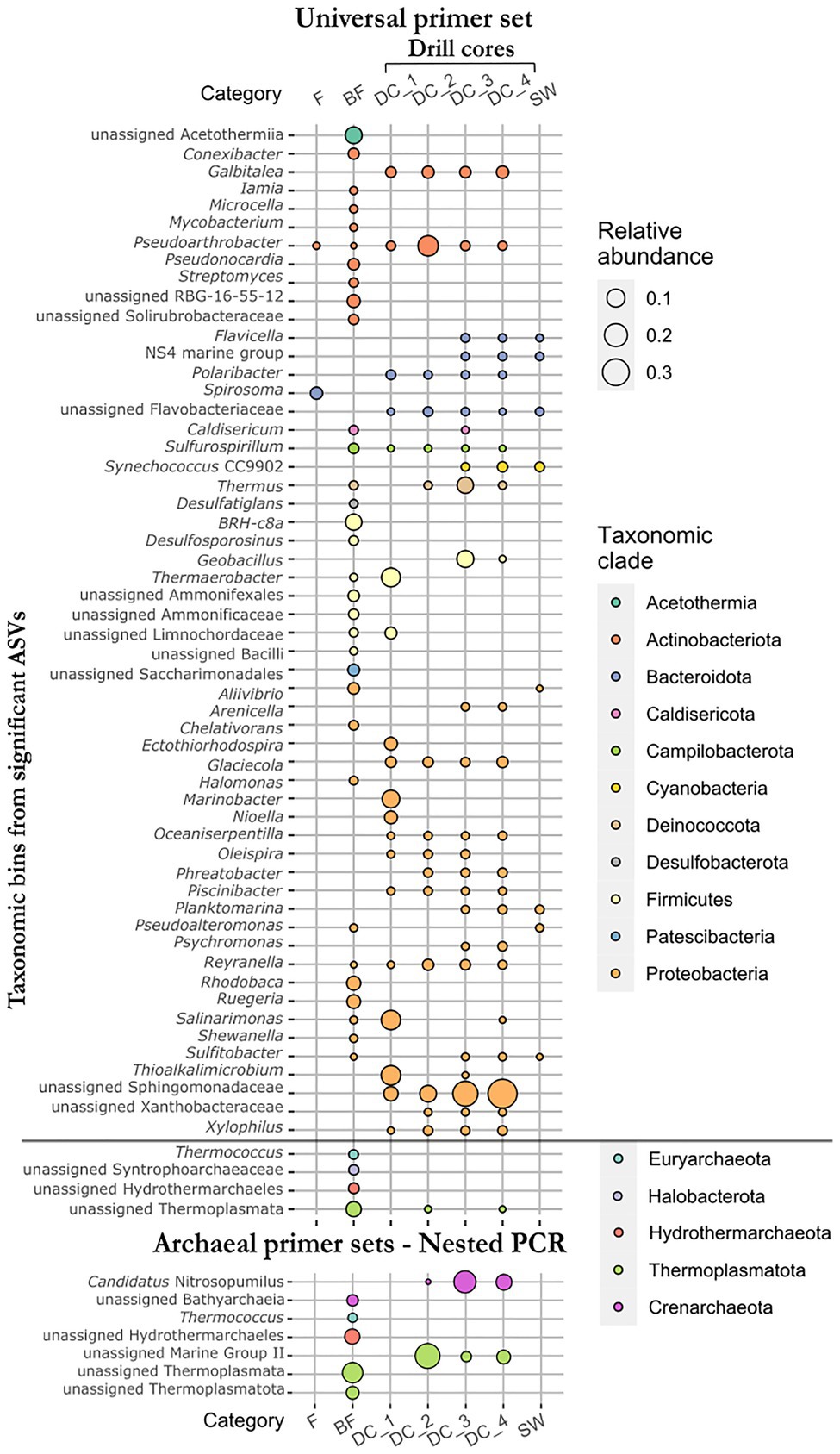
Figure 5. Relative abundance of taxonomic bins from all significant ASVs identified by differential abundance analyses using both universal and archaeal primer data sets. Sample types: F, fumarole (n=2); BF, borehole fluid (n=18); DC, drill core (n=17); SW, seawater samples (n=4). Categories of drill cores: DC_1, drill cores from the subaerial deposit (n=2); DC_2, just above and at a zone of daily intertidal fluctuations at coastal sea level (n=3); DC_3, in the submarine deposit at the hydrothermal temperature maximum (n=7); DC_4, in the submarine deposit below the zone of seawater inflow (n=5).
Compared to the seawater samples, the drill cores showed overrepresented ASVs assigned to the taxa Caldisericum, Galbitalea, Geobacillus, Glaciecola, Oceaniserpentilla, Oleispira, Piscinibacter, Pseudoarthrobacter, Psychromonas, Reyranella, Sulfurospirillum, Thermaerobacter, Thermus, unassigned bacteria from the Sphingomonadaceae family, and unassigned Thermoplasmata. ASVs assigned to Aliivibrio and Pseudoalteromonas were significantly underrepresented in the drill cores compared to the seawater samples. Although the latter genera are generally detected in seawater, they were significantly overrepresented in the borehole fluids collected before drilling, indicating that marine microorganisms (or their DNA) naturally infiltrate the subsurface basaltic deposits. Other overrepresented ASVs in the borehole fluids, as compared to the drill core samples, included unassigned Acetothermiia, BRH-c8a (Desulfallas-Sporotomaculum), Ruegeria, Rhodobaca, unassigned RBG-16-55-12 (Actinobacteriota), Conexibacter, Desulfosporosinus, Desulfatiglans, unassigned Ammonifexales, and Ammonificaceae, among others (Supplementary Table S5). Archaeal taxa that proved to be overrepresented in the borehole fluids included unassigned Thermoplasmata, Hydrothermarchaeales, Syntrophoarchaeaceae, and the genus Thermococcus. Using the archaeal primer sets, unassigned Thermoplasmatota and Bathyarchaeia were added to the list of overrepresented archaeal taxa detected using the universal primer set (Figure 5, Supplementary Table S5).
Comparing the four categories of drill cores, ASVs overrepresented in the drill core samples from the subaerial tuff cone (DC_1; 23–35m b.s.; 20–36°C in 2017) were assigned to the genera Thermaerobacter, Thioalkalimicrobium, Salinarimonas, Marinobacter, Nioella, Ectothiorhodospira, Polaribacter, and unassigned Limnochordaceae (Figure 5). No ASVs were significantly overrepresented in drill core samples from the zone of daily intertidal fluctuations (DC_2; 44–65m b.s.; 59–101.5°C in 2017) compared with the other categories of drill core samples. Although one ASV assigned to Geobacillus is significantly overrepresented in the drill core samples from the hydrothermal temperature maximum (DC_3; 78–139m b.s.; 97–124°C in 2017), this is similar to the drill core samples located below the zone of seawater inflow (DC_4; 157–181m b.s.; 37–84°C in 2017). These samples shared significant ASVs with the seawater samples, including Flavicella, NS4 marine group from the Flavobacteriaceae family, Synechococcus CC9902, Planktomarina, and Sulfitobacter. The presence of these taxa further suggests an infiltration of marine microorganisms associated with the infiltration of cool seawater at 145–155m depth, if we presume that no drilling fluid contamination remains.
To determine whether the detected DNA sequences could be derived from planktonic cells in interstitial pore fluids or, alternatively, attached to surfaces of the lapilli tuff and to investigate their organization on the basaltic substrate, SEM studies were undertaken on six drill core samples, differing in depth (C4, C9, C22, C49, C55, and C65).
All six lapilli tuff samples are porous with high water absorption (Jackson et al., 2019b). Vesicles with spherical shapes 10–80μm in diameter occur in all samples, and platy clay mineral structures, with flower petal morphologies, indicate alteration of the original volcanic glass (Jakobsson and Moore, 1986; Figure 6B). The distribution of microstructures varies with sampling depth; here, we focus on instructive samples at 32m b.s (C9) and 65m b.s (C22). Some vesicle surfaces are covered by a net of thin filaments with an approximate diameter of 10nm. These features are closely intertwined with one other and could possibly correspond to extracellular polymeric substances-like (EPS) structures (Thorseth et al., 2001, 2003; Sudek et al., 2017), perhaps representing a “relic” of past biofilm activity (Figures 6A,B). In other vesicles, spheroidal elements correspond to putative microbial cells 2μm in diameter (Figure 6C) as previously described (Thorseth et al., 2001, 2003). Furthermore, these vesicles are covered by fibrillar-like microstructures (Figure 6D), that appear texturally different from the EPS-like structures (Figure 6B). They occur as nets of oriented fibers with ramifications; some seem to be attached to the spheroidal structures. Energy dispersive X-ray analysis on the spheroidal elements does not detect carbon or phosphate but, instead, detects magnesium, aluminum, and iron. They therefore seem to have an inorganic origin.
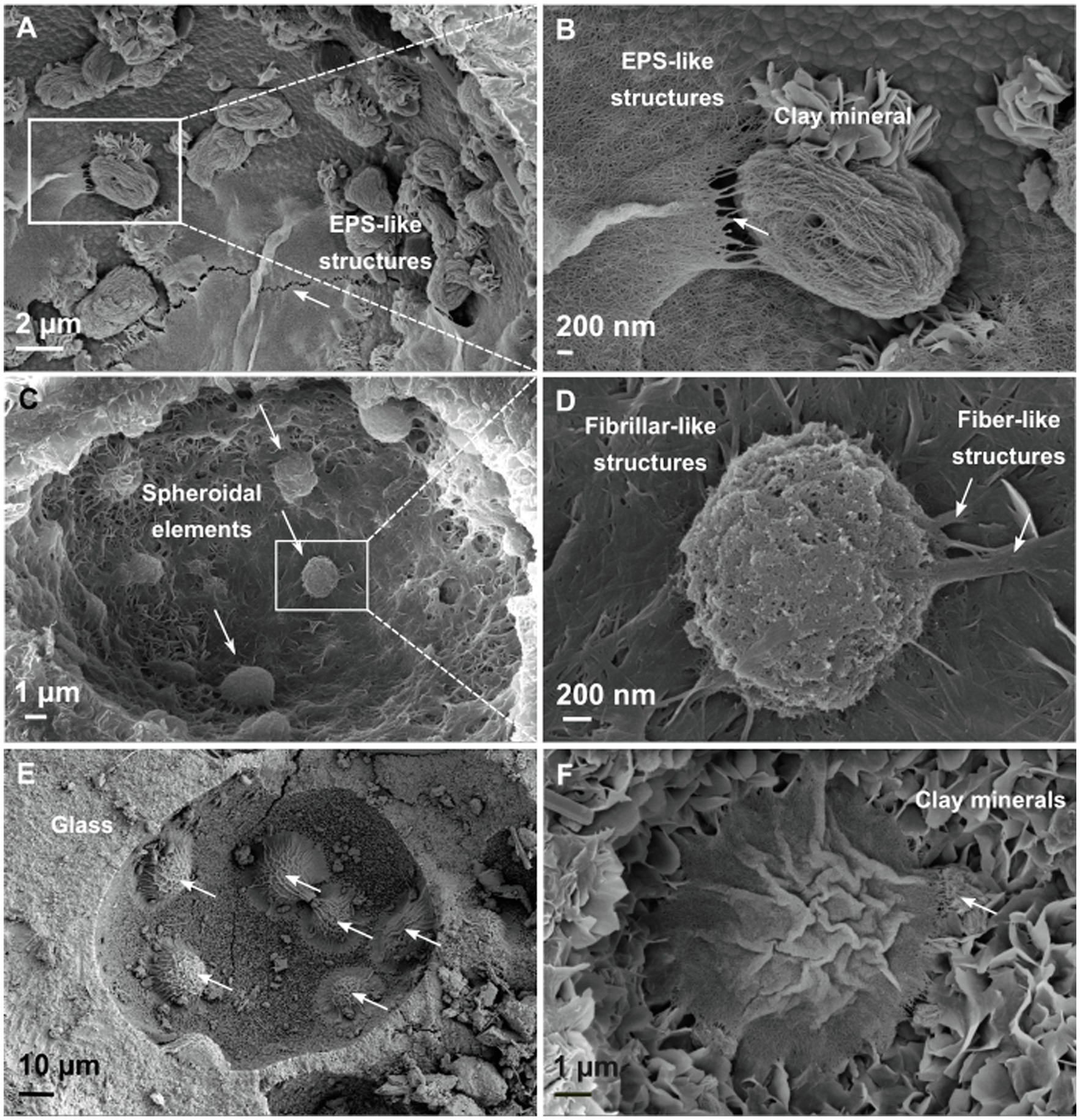
Figure 6. Scanning electron micrographs of Surtsey tephra surfaces in drill core samples. (A,B) Platey clay mineral and EPS-like structures, 65m b.s., sample C22. (C,D) Vesicle surfaces in altered basaltic glass are covered by a net of fibrillar-like structures that appears to be attached to spheroidal elements, 32m b.s, sample C9. (E,F) Wrinkled dome structures, or mounds, are covered by a network of filaments, 65m b.s, sample C22.
Wrinkled dome structures are observed in many vesicles at 65m b.s (Figure 6E) with a diameter size of 10–20μm. A network of intercrossed filaments covers the surface of the dome or mound (Figure 6F). Their wrinkled appearance could be explained by the sample preparation for SEM images, which includes gradual dehydration. EDX analyses investigate the chemical signature of the wrinkled dome structures (Supplementary Figure S7). The superposition of the spectra corresponding to the adjacent clay mineral(s) (Supplementary Figure S7, red area) and the wrinkled dome structures (Supplementary Figure S7, blue area) shows enrichment in carbon, oxygen, sodium, phosphate, and calcium in the crinkled ridge of the mound. These could possibly indicate elements associated with the cells and biomass of a biofilm.
Microbial colonization by pioneer communities can occur within months on freshly deposited erupted volcanic rocks in geothermally active environments (e.g., Konhauser et al., 2002; Kelly et al., 2014). The Surtsey geothermal system offers a highly unusual site to study rapid microbial colonization in newly formed oceanic crust at <100years and>50°C. Results of the analysis described in Figures 3, 4, and 6 suggest that diverse microbial communities have developed in the Surtsey deposits only 50years after the eruptions ended. Communities attached to the lapilli tuff of drill core samples are significantly different from communities in associated fluids (Figures 3A,B). Similar observations have previously been reported in crustal environments (Ramírez et al., 2019). The Surtsey data suggest that fumaroles, borehole fluids, drill cores, and seawater samples harbor significantly distinct microbial communities (Figures 3B,C). Yet, taxa are shared between these sample types (Figure 3D), suggesting possible transfer of microorganisms among these habitats. The hot fumarole samples from the surface of Surtsey at 82.3 and 85.6°C harbor microbial communities that are more similar to those of the drill core samples located above sea level in the subaerial deposits (e.g., samples C4, C9, 23–35m b.s.; Figure 2B). This suggests that some species could disperse through the subaerial tuff cone above sea level up to the surface through fumarole activity (Tin et al., 2011). Excluding all the drilling fluid ASVs contaminants, microbial communities detected in the seawater samples are closer in community structure to the drill core samples from the zone of seawater inflow (sample C52, 148m b.s., Figure 2B) and to the samples in proximity to the seafloor (sample C65, 181m b.s., Figure 2B). This suggests that microorganisms from the seawater surrounding the island (i) infiltrate the submarine tephra deposits, following the seawater inflow at about 148m b.s (Jakobsson and Moore, 1986; Jackson et al., 2019b; Kleine et al., 2020), and (ii) occupy the deeper zone of poorly consolidated tuff through indirect infiltration of the seawater and its circulation via the seafloor. The ability of some species to expand across ecosystems and adapt to new environmental conditions (Sriswasdi et al., 2017) has possibly driven the subsurface colonization of the Surtsey volcano. Species that cannot survive might serve as supply of fermentable organic molecules for the subsurface heterotrophic microorganisms (Li et al., 2020).
The problem of subsurface sample contamination during drilling operations is well-known (Barton et al., 2006; Lever et al., 2006; Kieft, 2010; Santelli et al., 2010; Sheik et al., 2018) since drilling operations require the use of circulating fluid that inevitably infiltrates into the drill core. As a result, the recovery of uncontaminated rock is nearly impossible (Friese et al., 2017; Kallmeyer, 2017). Conventional methods to assess contamination include the addition of tracer compounds to the drilling fluid. These include fluorescent dyes (Pellizzari et al., 2013), perfluorocarbon tracers (PFT; Lever et al., 2006; Inagaki et al., 2016; Orcutt et al., 2017), and microsphere tracers (Kallmeyer et al., 2006; Yanagawa et al., 2013). The use of these tracer compounds is prohibited on Surtsey due to the protected environmental status of the island (Baldursson and Ingadóttir, 2007). Furthermore, their performance in the Surtsey system would have been quite problematic. Although fluorescent dyes have the advantages of high sensitivity for detection, low cost, and ease of use (Pellizzari et al., 2013), they are unstable at low pH (Zhu et al., 2005) and are also susceptible to degradation in the presence of light (Diehl and Horchak-Morris, 1987). Measurements of Surtsey subsurface borehole fluids extracted from SE-01 in 2016 showed pH values decreasing to 5 at some depths (Kleine et al., 2020). Water tanks containing the drilling fluid were exposed to sunlight during the long Icelandic summer days. The detection of PFTs must be performed immediately on fresh cores because of the high volatility of the PFTs and requires elaborate equipment (Lever et al., 2006) whose transport by helicopter to Surtsey would have been very difficult. Finally, microsphere tracers decompose under high-temperature conditions (Yanagawa et al., 2013) and would not have performed well in the Surtsey hydrothermal system, which currently exceeds 120°C at some depths (Marteinsson et al., 2015; Jackson et al., 2019b). Because of the unique circumstances of drilling on Surtsey, the filtration and UV sterilization of the drilling fluid was the most effective strategy to manage contamination. We then mitigated this strategy with a simple overlap approach that is commonly used in environmental microbiology studies to identify and remove contaminants (Sheik et al., 2018). At the ASVs level, this approach enabled the distinction between marine contaminants from the drilling fluid and possible endemic residents, providing a firm basis for the exploration of microbial life in this extreme and unusual habitat (Figure 2).
The DNA concentration in the drill core samples, measured using a QuBit fluorometer, ranged from 0.2–1.36ng.g−1 of basalt, which is in good correlation with other basaltic environments (Fisk et al., 2003; Herrera and Cockell, 2007). Surprisingly, DNA concentration did not correlate with variations in temperature (Supplementary Figure S2a). It does decrease with depth (Supplementary Figure S2b), as previously observed in terrestrial and marine subsurface habitats (Cockell et al., 2012; Ciobanu et al., 2014; Mcmahon and Parnell, 2014), but not significantly. The presence of contaminant DNA in the low-biomass basaltic core samples quite possibly distorts these results. Indeed, low DNA concentration exacerbates issues of external contamination (Sheik et al., 2018). When using 16S rRNA gene sequencing to analyze low-biomass samples, the amplification and sequencing of contaminant DNA are introduced during extraction and library preparation steps, and the risk of well-to-well contamination is high (Minich et al., 2019). The low biomass and the presence of external contamination also influence the sequencing results by affecting the magnitude and biological provenance of analyzed sequences. Therefore, it is of crucial importance to use negative controls and optimal decontamination approaches. The inclusion of bacterial mock communities as extraction and sequencing controls would have been beneficial for the study (Pollock et al., 2018). We emphasize that all samples were handled with extreme care during all steps of the project to minimize, identify and remove contaminants using a combination of experimental and computational methods. The lists of contaminants that have been identified based on the presence/absence of ASVs across samples, their relative abundances, and their taxonomic assignment (Supplementary Tables S1, S2 and S4) should be valuable for future microbial explorations of the oceanic crust. In addition to the low biomass and the presence of external contamination affecting the results, we should keep in mind that the compositional nature of the data has limits. Indeed, the data set reported in this study was obtained by 16S rRNA gene amplicon sequencing; thus, the data are compositional being based on relative abundances, which sum to a constant. Therefore, the analytical approaches (e.g., rarefaction, normalization) and statistical methods (e.g., ANOVA) used to study microbiome data influence the results and can potentially be are subjected to inflated false discovery rates (Gloor et al., 2017). This can lead to the lack of reproducibility among microbiome studies and misinterpretations of microbial community structures (e.g., alpha diversity).
Many of the taxa detected in the SE-01 borehole fluid samples match DNA sequences previously identified in hot springs, hydrothermal vents, and subsurface environments (Figure 5). For example, sequences assigned to the class Acetothermiia closely match with DNA sequences previously found in hydrothermal sediments (GenBank: FM868292). Members of this class were previously detected in anaerobic digesters, hot springs, and other deep biosphere studies (Takami et al., 2012; Jungbluth et al., 2017; Zaitseva et al., 2017; Hao et al., 2018; Korzhenkov et al., 2018). Sequences assigned to the genus BRH-c8a from the family Desulfallas-Sporotomaculum match closely with sequences retrieved from deep groundwater (LC179584) and group with the genus Desulfotomaculum that can be found in deep subsurface environments (Sousa et al., 2018; Watanabe et al., 2018). Other sequences belonging to the class Desulfotomaculia are assigned to Ammonifexales and Ammonificaceae; these are related to DNA sequences retrieved from petroleum reservoirs (MF470409). In addition, sequences assigned to Desulfosporosinus closely match sequences retrieved from coal formation waters (KC215435), while sequences assigned to Desulfatiglans match with sequences found in hydrothermal vents (AB294892). Among the abundant ASVs detected in the borehole fluids, an early-branching, uncultivated actinobacterial clade identified as RBG-16-55-12 in the SILVA database release 138 has been previously detected in serpentinite-hosted systems (Merino et al., 2020). Uncharacterized Thermoplasmata are also detected in serpentinite subsurface deposits (Motamedi et al., 2020), yet numerous sequences assigned to this class from the SE-01 borehole fluid samples showed less than 90% of sequence similarity with the first match on the NCBI Nucleotide collection database (AB327321). These results suggest that all the latter taxa could be endemic in the Surtsey subsurface deposits and perhaps could be common in other oceanic or continental subsurface habitats, as well. Other taxa detected in the SE-01 borehole fluid samples are usually found in seawater, including Halomonas, Pseudoalteromonas, Sulfitobacter, Aliivibrio, and Shewanella (Figure 5). The presence of marine microorganisms in the SE-01 borehole fluid before new drilling began in 2017 further demonstrates that the infiltration of seawater transports marine microorganisms into the subsurface and its hydrothermal system. These species are also frequently detected in cool basaltic oceanic crust (Templeton et al., 2005; Mason et al., 2007; Zhang et al., 2016), and the question of whether or not these species survive in the Surtsey hydrothermal system could depend on the influence of temperature (Edwards et al., 2012; Baquiran et al., 2016; Ramírez et al., 2019).
Scanning electron microscopy studies of instructive drill core samples reveal possible biotic structures whose size, morphology, and fabrics seem to be consistent with EPS-microcolony complexes that are attached to the basaltic substrate (Figure 6, Supplementary Figure S7; Thorseth et al., 2001, 2003; Nakagawa et al., 2006). Indeed, some taxa detected in the drill core samples could represent endemic inhabitant of the basaltic subsurface of the island, which can be divided into two distinct habitats: the subaerial tuff cone and the submarine tuff deposits. In the subaerial tuff cone (DC_1; 23–35m b.s.; 20–36°C in 2017), the taxa include the genera Thermaerobacter, Thioalkalimicrobium, Salinaromonas, Marinobacter, and Ectothiorhodospira (Figure 5). Thermaerobacter are thermophilic to extremely thermophilic bacteria found in terrestrial and oceanic subsurface environments (Spanevello et al., 2002; Takai et al., 1999). Thioalkalimicrobium, Ectothiorhodospira, and Salinarimonas are detected in alkaline and saline habitats such as soda lakes (Sorokin et al., 2001), saline soil (Cai et al., 2011), or salt mines (Liu et al., 2010). The presence of Marinobacter only in the tuff deposits above sea level is curious since it is a common lineage found in marine basaltic habitats, including ridge-flank systems and seamount (Templeton et al., 2005; Zhang et al., 2016). Nevertheless, the presence of these taxa in the subaerial deposits suggests adaptation of the microbial communities to more extreme environmental conditions, such as high temperature, high salt concentration produced by NaCl saturation from seawater evaporation or from alkaline pH produced by basaltic glass dissolution at low fluid-rock ratios (Kleine et al., 2020). No hyperthermophilic species were significantly enriched in the drill core samples from the hydrothermal temperature maximum (DC_3; 78–139m b.s.; 97–124°C in 2017 and 100–141°C in 1979), yet two thermophilic species Thermus and Geobacillus from the Thermoleovorans group were detected (Figure 5). Based on these results and the taxa detected in the SE-01 borehole fluids, it seems that microbial life may persist in subsurface deposits that have experienced temperatures >120°C, the presumed temperature for functional microbial life. The hydrothermal zone at about 100m b.s. could act as a dispersal barrier that provides an obstacle to the transfer of live cells from the zone of tidal flux and upper submarine deposits to the deeper submarine deposits (e.g., samples C27; 78m b.s. and C52; 148m b.s.). Furthermore, no significant difference exists between the observed microbial communities from the lowermost submarine deposits below the zone of cool seawater inflow (DC_4; 157–181m b.s.; 37–84°C in 2017) and the other submarine deposits (Supplementary Figure S6). Shared taxa mainly include mesophilic Proteobacteria such as unassigned Sphingomonadaceae, Glaciecola, Arenicella, Oceaniserpentilla, and Piscinibacter that are commonly found in marine habitats. In addition, Candidatus Nitrosopumilus and unassigned Thermoplasmata from the Marine Group II were detected in great abundance (Figure 5). Those taxa have been previously detected in marine basalts and deep seawater circulation through oceanic crust (Mason et al., 2007; Singer et al., 2015; Suzuki et al., 2020; Bergo et al., 2021). Although the deep ocean is typically enriched in archaeal cells, other marine environments usually show a dominance of bacterial lineages (Karner et al., 2001). This has been previously observed in the basaltic crust. For example, Gammaproteobacteria and Alphaproteobacteria are detected in great abundance in the seafloor basaltic glass of the East Pacific Rise (Santelli et al., 2008, 2009), the Arctic spreading ridges (Lysnes et al., 2004), altered basalts from the Hawaiian Loihi Seamount (Templeton et al., 2005; Santelli et al., 2008; Jacobson Meyers et al., 2014), and the Mid-Atlantic Ridge (Rathsack et al., 2009; Mason et al., 2010). Deltaproteobacteria, Firmicutes, Gammaproteobacteria, and Bacteroidetes are also detected in great abundance in the Juan de Fuca Ridge flank and the Costa Rica Rift (Nigro et al., 2012; Jungbluth et al., 2013, 2014). Our results support the dominance of bacterial lineages in marine basalts.
While most taxa could be identified at a low taxonomic rank, many others were not assigned to a known genus, family, or even order. For example, many sequences detected in this study fell into unknown clades, including Acetothermiia, Ammonifexales, Bacilli, RBG-16-55-12 from the phylum Actinobacteriota, Sphingomonadaceae, Limnochordaceae, and Saccharimonadales, among others. Likewise, many archaeal sequences could not be identified further than the phylum level, including unassigned Thermoplasmatota and Halobacterota, or the class level, including unassigned Thermoplasmata and Bathyarchaeia. Hence, the subsurface biosphere of Surtsey lapilli tuff and tephra, as well as fluids, could have high potential for discoveries of new microbial clades.
The metabolic potential of the endemic subsurface microbial communities can be discussed with some degree of certainty, while remaining mindful of the difficulties inherent to identifications based on 16S rRNA gene sequence analysis. To support our hypotheses, predictive functional analyses were performed using PICRUSt2. A few MetaCyc pathways were selected to represent the functional potential of the bacterial and archaeal communities for carbon, sulfur, nitrogen, and methane metabolism (Supplementary Figures S8, S9). Many genera reported in this study belong to taxonomic clades with known metabolisms that are involved in both heterotrophy and chemolithoautotrophy (Figure 7). The presence of putative sulfate-reducing bacteria strongly suggests a potential for active sulfate reduction, including Desulfosporosinus, Desulfatiglans, and the Desulfotomaculia class, among others. In addition, sulfur oxidizers were detected, such as the genera Thioalkalimicrobium, Sulfurospirillum, Sulfurimonas, Ectothiorhodospira, and Sulfurihydrogenibium. The observation of these bacterial taxa possibly involved in sulfate reduction and sulfur oxidation suggests an active sulfur cycle in the subsurface of Surtsey (Supplementary Figure S8), as has previously been reported in similar ecosystems (Bach and Edwards, 2003; Lever et al., 2013; Suzuki et al., 2020). This is further reinforced by the detection of archaea possibly involved in the sulfur cycle, such as Thermococcus, Pyrococcus, and Archaeoglobus. Also, characterized members of the Thermoplasmatales are typically involved in sulfur cycling (Barton et al., 2014; Arce-Rodríguez et al., 2019). These taxa coincide with deposits that contain sulfate minerals, principally anhydrite and gypsum (Jakobsson and Moore, 1986; Kleine et al., 2020; Prause et al., 2020).
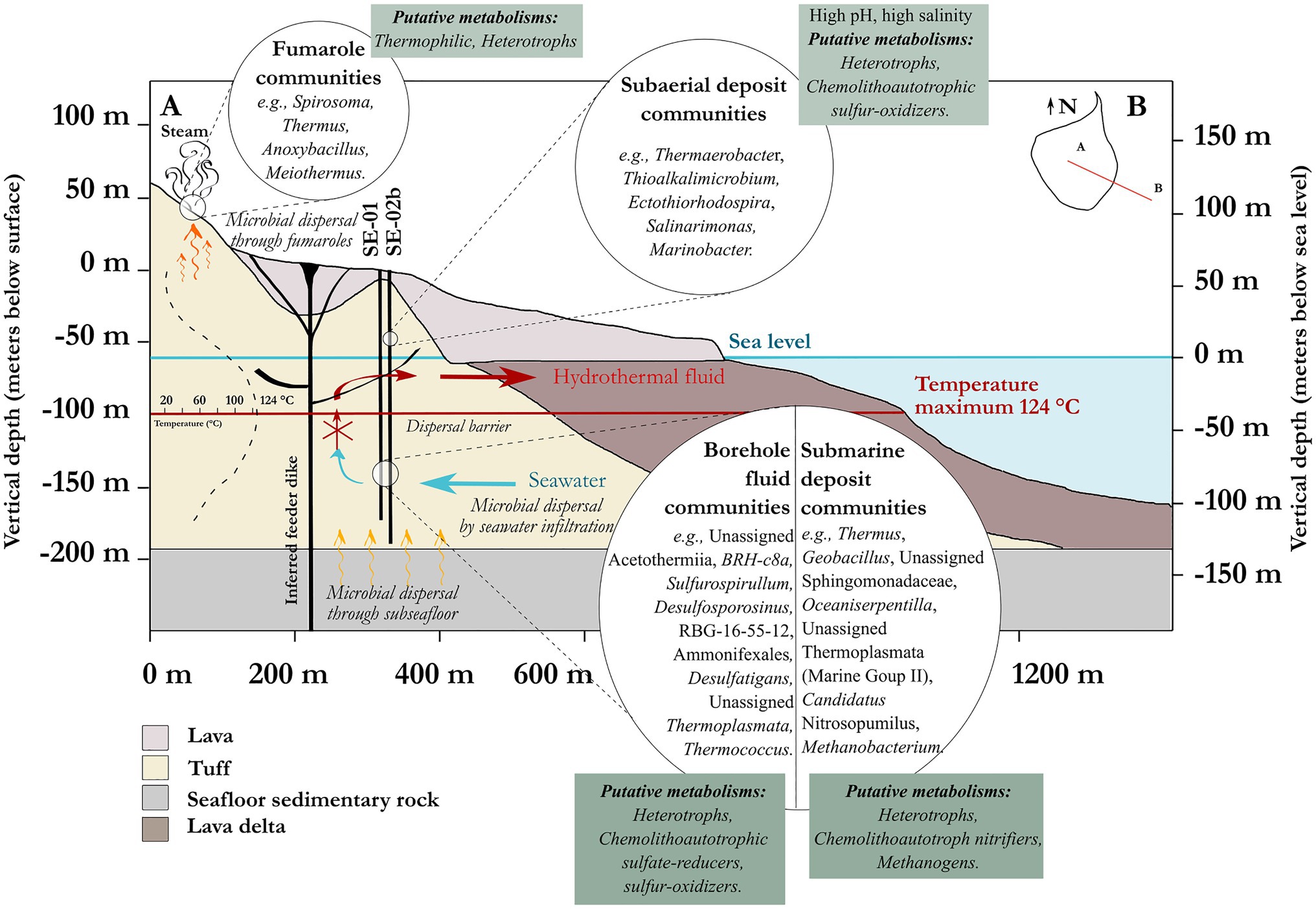
Figure 7. Overview of the putative metabolisms and microbial dispersion scenario of the subsurface of Surtsey volcano.
In addition, the genus Methanobacterium from the order Methanobacteriales dominates the archaeal sequences of drill core sample C55, at 157m depth (Figure 3). The MetaCyc pathways detected in this sample using the archaeal data set were mainly involved in methane metabolism (e.g., methanogenesis from H2 and CO2, coenzyme B biosynthesis, coenzyme M biosynthesis I; Supplementary Figure S9). One taxon from the same order of Methanobacteriales was previously reported to dominate the SE-01 borehole fluid microbial communities sampled in 2009 at similar depth (Marteinsson et al., 2015). Methanobacterium spp. grows by reducing carbon dioxide to methane and uses molecular hydrogen as the electron donor (Kern et al., 2015). Hence, these taxa could play an important role as primary producers in this ecosystem, at least at certain depths. The possible occurrence of an active methane cycle is supported by the presence of other methanogens (e.g., Methanosarcinia (Syntrophoarchaeaceae), Methanomassiliicoccales (Methanothermus, Methanoregula), as well as methanotrophic (e.g., Methylocella, Methylacidiphilaceae), and methylotrophic (e.g., Hansschlegelia, Methylopila, Methylophilaceae (OM43 clade), Methylophaga) bacteria, despite their relatively low abundances. The observation of genera such as Geobacter, Rhodoferax, Marinobacter, Shewanella, and Ferruginibacter, as well as Hydrogenophilus and Hydrogenophaga, could indicate that iron and hydrogen are electron donors within the ecosystem, as previously reported for other subsurface habitats (Bach and Edwards, 2003; Bach, 2016; Zhang et al., 2016). In addition, putative ammonium-oxidizing archaea belonging to the Marine Group II and Candidatus Nitrosopumilus suggest an ability to transform nitrogen compounds as previously reported in the oceanic crust (Daae et al., 2013; Orcutt et al., 2015; Jørgensen and Zhao, 2016; Zhao et al., 2020).
The 1979 and 2017 drilling projects at Surtsey volcano provide a rare opportunity to explore the subsurface microbial diversity of a very young basaltic island associated with an active hydrothermal-seawater system in newly formed oceanic crust. A cored borehole dedicated to microbiology research in the 2017 drilling operation probes a low biomass but highly diverse habitat that hosts bacterial and archaeal clades, including extremophiles, that have been previously detected in other terrestrial and marine environments. Many clades, however, fall into as-yet-unknown lineages. The 16s rRNA gene amplicon sequencing data provide insights into diverse sources of microbial colonization in newly formed oceanic crust, with potential dissemination from the deep subsurface, surrounding seawater, and surface ecosystems. The island of Surtsey may thus be regarded as a porous, sponge-like basaltic structure that absorbs cells from the surrounding environments and selects microorganisms that can adapt to the extreme environmental conditions that exist within the volcano. The data also provide a baseline for long-term observations of the microbial communities inhabiting the subsurface of Surtsey and their temporal succession amid a changing hydrothermal environment, in which poorly consolidated tephra lithify to form well-consolidated lapilli tuff. Further research, including metagenomic sequencing, will increase our knowledge of the metabolism and function of the microbiome in this very young basaltic environment.
The data sets (accession number ERP126178) for this study can be found in the European Nucleotide Archive (ENA) at EMBL-EBI (https://www.ebi.ac.uk/ena/browser/view/PRJEB42339).
VM (PI of the microbiology part of drilling operation), MJ, and MG conceived the study. PB, AK, VM, and members of the SUSTAIN onsite, science teams conducted field operations and sampling. PB and PV processed the samples and conducted the molecular biology experiments. PB, PV, and SK performed the data analysis. PB wrote the original draft. All authors took part in writing the manuscript.
Sample collection and sample processing were funded with a grant of excellence from the Icelandic Science fund, ICF-RANNÍS IceSUSTAIN (163083-051). The drilling operation was funded with International Continental Scientific Drilling Program (ICDP) through the SUSTAIN project and the IceSUSTAIN grant. A doctoral student grant from RANNÍS (206582-051) was attributed to PB.
The pipeline used to analyze the data set reported in this manuscript can be found at https://benjjneb.github.io/dada2/tutorial.html. The code used to identify contaminants can be found at https://benjjneb.github.io/decontam/vignettes/decontam_intro.html, and the code used to build the figures can be found in the supplementary material under the section code.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The authors would like to thank all members of the SUSTAIN onsite and science teams, and the Surtsey Research Society for their contributions to the drilling project and sampling, especially B. Kleine, T. Weisenberger, S. Prause, C.F. Gorny, and S. Couper. The Icelandic Science fund, ICF-RANNÍS Jules Verne fund (scientific and technical cooperation between Iceland and France) contributed additional funds. The authors thank E. Gérard, A. Lecoeuvre, B. Ménez, and S. Borensztajn for their contributions to sample preparation and SEM images.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2021.728977/full#supplementary-material
Arce-Rodríguez, A., Puente-Sánchez, F., Avendaño, R., Martínez-Cruz, M., de Moor, J. M., Pieper, D. H., et al. (2019). Thermoplasmatales and sulfur-oxidizing bacteria dominate the microbial community at the surface water of a CO2-rich hydrothermal spring located in Tenorio volcano National Park, Costa Rica. Extremophiles 23, 177–187. doi: 10.1007/s00792-018-01072-6
Bach, W. (2016). Some compositional and kinetic controls on the bioenergetic landscapes in oceanic basement. Front. Microbiol. 7:107. doi: 10.3389/fmicb.2016.00107
Bach, W., and Edwards, K. J. (2003). Iron and sulfide oxidation within the basaltic ocean crust: implications for chemolithoautotrophic microbial biomass production. Geochim. Cosmochim. Acta 67, 3871–3887. doi: 10.1016/S0016-7037(03)00304-1
Baldursson, S., and Ingadóttir, Á. (2007). Nomination of Surtsey for the UNESCO world heritage list. Icelandic Inst. Nat. Hist. Reykjavik Available at: https://whc.unesco.org/uploads/nominations/1267.pdf
Baquiran, J. P. M., Ramírez, G. A., Haddad, A. G., Toner, B. M., Hulme, S., Wheat, C. G., et al. (2016). Temperature and redox effect on mineral colonization in Juan de Fuca Ridge Flank subsurface crustal fluids. Front. Microbiol. 7:396. doi: 10.3389/fmicb.2016.00396
Bar-On, Y. M., Phillips, R., and Milo, R. (2018). The biomass distribution on earth. Proc. Natl. Acad. Sci. U. S. A. 115, 6506–6511. doi: 10.1073/pnas.1711842115
Baross, J. A., Wilcock, W. S. D., Kelley, D. S., Delong, E. F., and Cary, S. C. (2004). The subsurface biosphere at mid-ocean ridges: issues and challenges. Geophys. Monogr. Ser. doi: 10.1029/GM144
Barton, L. L., Fardeau, M. L., and Fauque, G. D. (2014). Hydrogen sulfide: A toxic gas produced by dissimilatory sulfate and sulfur reduction and consumed by microbial oxidation. Met. Ions Life Sci. 14, 237–277. doi: 10.1007/978-94-017-9269-1_10
Barton, H. A., Taylor, N. M., Lubbers, B. R., and Pemberton, A. C. (2006). DNA extraction from low-biomass carbonate rock: An improved method with reduced contamination and the low-biomass contaminant database. J. Microbiol. Methods 66, 21–31. doi: 10.1016/j.mimet.2005.10.005
Bergo, N. M., Bendia, A. G., Ferreira, J. C. N., Murton, B. J., Brandini, F. P., and Pellizari, V. H. (2021). Microbial diversity of deep-sea ferromanganese crust field in the Rio Grande Rise, Southwestern Atlantic Ocean. Microb. Ecol. 82, 344–355. doi: 10.1007/s00248-020-01670-y
Cai, M., Wang, L., Cai, H., Li, Y., Wang, Y. N., Tang, Y. Q., et al. (2011). Salinarimonas ramus sp. nov. and Tessaracoccus oleiagri sp. nov., isolated from a crude oil-contaminated saline soil. Int. J. Syst. Evol. Microbiol. 61, 1767–1775. doi: 10.1099/ijs.0.025932-0
Callahan, B. J., McMurdie, P. J., Rosen, M. J., Han, A. W., Johnson, A. J. A., and Holmes, S. P. (2016). DADA2: high-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583. doi: 10.1038/nmeth.3869
Caporaso, J. G., Lauber, C. L., Walters, W. A., Berg-Lyons, D., Huntley, J., Fierer, N., et al. (2012). Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 6, 1621–1624. doi: 10.1038/ismej.2012.8
Ciobanu, M. C., Burgaud, G., Dufresne, A., Breuker, A., Rédou, V., Maamar, S. B., et al. (2014). Microorganisms persist at record depths in the subseafloor of the Canterbury Basin. ISME J. 8, 1370–1380. doi: 10.1038/ismej.2013.250
Cockell, C. S., Voytek, M. A., Gronstal, A. L., Finster, K., Kirshtein, J. D., Howard, K., et al. (2012). Impact disruption and recovery of the deep subsurface biosphere. Astrobiology 12, 231–246. doi: 10.1089/ast.2011.0722
Cowen, J. P., Giovannoni, S. J., Kenig, F., Johnson, H. P., Butterfield, D., Rappé, M. S., et al. (2003). Fluids from aging ocean crust that support microbial life. Science 299, 120–123. doi: 10.1126/science.1075653
D’Hondt, S., Jørgensen, B. B., Miller, D. J., Batzke, A., Blake, R., Cragg, B. A., et al. (2004). Distributions of microbial activities in deep subseafloor sediments. Science 306, 2216–2221. doi: 10.1126/science.1101155
Daae, F. L., Økland, I., Dahle, H., Jørgensen, S. L., Thorseth, I. H., and Pedersen, R. B. (2013). Microbial life associated with low-temperature alteration of ultramafic rocks in the Leka ophiolite complex. Geobiology 11, 318–339. doi: 10.1111/gbi.12035
Davis, N. M., Proctor, D. M., Holmes, S. P., Relman, D. A., and Callahan, B. J. (2018). Simple statistical identification and removal of contaminant sequences in marker-gene and metagenomics data. Microbiome 6:226. doi: 10.1186/s40168-018-0605-2
Diehl, H., and Horchak-Morris, N. (1987). Studies on fluorescein-V The absorbance of fluorescein in the ultraviolet, as a function of pH. Talanta 34, 739–741. doi: 10.1016/0039-9140(87)80232-1
Direito, S. O. L., Marees, A., and Röling, W. F. M. (2012). Sensitive life detection strategies for low-biomass environments: optimizing extraction of nucleic acids adsorbing to terrestrial and Mars analogue minerals. FEMS Microbiol. Ecol. 81, 111–123. doi: 10.1111/j.1574-6941.2012.01325.x
Douglas, G. M., Maffei, V. J., Zaneveld, J. R., Yurgel, S. N., Brown, J. R., Taylor, C. M., et al. (2020). PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 38, 685–688. doi: 10.1038/s41587-020-0548-6
Edwards, K. J., Bach, W., and McCollom, T. M. (2005). Geomicrobiology in oceanography: microbe-mineral interactions at and below the seafloor. Trends Microbiol. 13, 449–456. doi: 10.1016/j.tim.2005.07.005
Edwards, K. J., Fisher, A. T., and Wheat, C. G. (2012). The deep subsurface biosphere in igneous ocean crust: frontier habitats for microbiological exploration. Front. Microbiol. 3:8. doi: 10.3389/fmicb.2012.00008
Fisk, M. R., Storrie-Lombardi, M. C., Douglas, S., Popa, R., McDonald, G., and Di Meo-Savoie, C. (2003). Evidence of biological activity in Hawaiian subsurface basalts. Geochem. Geophys. Geosyst, 12:1103. doi: 10.1029/2002GC000387
Friese, A., Kallmeyer, J., Kitte, J. A., Martínez, I. M., Bijaksana, S., and Wagner, D. (2017). A simple and inexpensive technique for assessing contamination during drilling operations. Limnol. Oceanogr. Methods 15, 200–211. doi: 10.1002/lom3.10159
Furnes, H., and Staudigel, H. (1999). Biological mediation in ocean crust alteration: how deep is the deep biosphere? Earth Planet. Sci. Lett. 166, 97–103. doi: 10.1016/S0012-821X(99)00005-9
Gloor, G. B., Macklaim, J. M., Pawlowsky-Glahn, V., and Egozcue, J. J. (2017). Microbiome datasets are compositional: And this is not optional. Front. Microbiol. 8:2224. doi: 10.3389/fmicb.2017.02224
Gold, T. (1992). The deep, hot biosphere. Proc. Natl. Acad. Sci. U. S. A. 89, 6045–6049. doi: 10.1073/pnas.89.13.6045
Hao, L., McIlroy, S. J., Kirkegaard, R. H., Karst, S. M., Fernando, W. E. Y., Aslan, H., et al. (2018). Novel prosthecate bacteria from the candidate phylum Acetothermia. ISME J. 12, 2225–2237. doi: 10.1038/s41396-018-0187-9
Heberling, C., Lowell, R. P., Liu, L., and Fisk, M. R. (2010). Extent of the microbial biosphere in the oceanic crust. Geochem. Geophys. Geosyst. 8, 1525–2027. doi: 10.1029/2009GC002968
Herrera, A., and Cockell, C. (2007). Exploring microbial diversity in volcanic environments: A review of methods in DNA extraction. J. Microbiol. Method. 70, 1–12. doi: 10.1016/j.mimet.2007.04.005
Inagaki, F., Hinrichs, K. U., Kubo, Y., Bihan, M., Bowden, S. A., Bowles, M., et al. (2016). IODP expedition 337: deep coalbed biosphere off Shimokita – microbial processes and hydrocarbon system associated with deeply buried coalbed in the ocean. Sci. Drill. 21, 17–28. doi: 10.5194/sd-21-17-2016
Inagaki, F., Hinrichs, K. U., Kubo, Y., Bowles, M. W., Heuer, V. B., Hong, W. L., et al. (2015). Exploring deep microbial life in coal-bearing sediment down to ~2.5 km below the ocean floor. Science 349, 420–424. doi: 10.1126/science.aaa6882
Jackson, M. D. (2020). Petrograpic and material observations of basaltic lapilli tuff, 1979 and 2017 Surtsey drill cores, Iceland. Surtsey Res. 14, 47–62. doi: 10.33112/surtsey.14.4
Jackson, M. D., Couper, S., Stan, C. V., Ivarsson, M., Czabaj, M. W., Tamura, N., et al. (2019a). Authigenic mineral texture in submarine 1979 basalt drill Core, Surtsey volcano, Iceland. Geochem. Geophys. Geosyst. 20, 3751–3773. doi: 10.1029/2019GC008304
Jackson, M. D., Gudmundsson, M. T., Bach, W., Cappelletti, P., Coleman, N. J., Ivarsson, M., et al. (2015). Time-lapse characterization of hydrothermal seawater and microbial interactions with basaltic tephra at Surtsey volcano. Sci. Drill. 20, 51–58. doi: 10.5194/sd-20-51-2015
Jackson, M. D., Gudmundsson, M. T., Weisenberger, T. B., Michael Rhodes, J., Stefánsson, A., Kleine, B. I., et al. (2019b). SUSTAIN drilling at Surtsey volcano, Iceland, tracks hydrothermal and microbiological interactions in basalt 50 years after eruption. Sci. Drill. 25, 35–46. doi: 10.5194/sd-25-35-2019
Jacobson Meyers, M. E., Sylvan, J. B., and Edwards, K. J. (2014). Extracellular enzyme activity and microbial diversity measured on seafloor exposed basalts from Loihi seamount indicate the importance of basalts to global biogeochemical cycling. Appl. Environ. Microbiol. 80, 4854–4864. doi: 10.1128/AEM.01038-14
Jakobsson, S. P., and Moore, J. G. (1982). The Surtsey research drilling project of 1979. Surtsey Res. 9, 76–93.
Jakobsson, S. P., and Moore, J. G. (1986). Hydrothermal minerals and alteration rates at Surtsey volcano, Iceland. Geol. Soc. Am. Bull. 97, 648–659. doi: 10.1130/0016-7606(1986)97<648:HMAARA>2.0.CO;2
Jakobsson, S. P., Thors, K., Vésteinsson, Á. T., and Ásbjörnsdóttir, L. (2009). Some aspects of the seafloor morphology at Surtsey volcano: The new multibeam bathymetric survey of 2007. Surtsey Res. 12,9–20.
Jian, C., Luukkonen, P., Yki-Järvinen, H., Salonen, A., and Korpela, K. (2020). Quantitative PCR provides a simple and accessible method for quantitative microbiota profiling. PLoS One 15:e0227285. doi: 10.1371/journal.pone.0227285
Jørgensen, S. L., and Zhao, R. (2016). Microbial inventory of deeply buried oceanic crust from a young ridge flank. Front. Microbiol. 7:820. doi: 10.3389/fmicb.2016.00820
Jungbluth, S. P., Bowers, R. M., Lin, H. T., Cowen, J. P., and Rappé, M. S. (2016). Novel microbial assemblages inhabiting crustal fluids within mid-ocean ridge flank subsurface basalt. ISME J. 10, 2033–2047. doi: 10.1038/ismej.2015.248
Jungbluth, S. P., del Rio, T. G., Tringe, S. G., Stepanauskas, R., and Rappé, M. S. (2017). Genomic comparisons of a bacterial lineage that inhabits both marine and terrestrial deep subsurface systems. PeerJ 5:e3134. doi: 10.7717/peerj.3134
Jungbluth, S. P., Grote, J., Lin, H. T., Cowen, J. P., and Rappé, M. S. (2013). Microbial diversity within basement fluids of the sediment-buried Juan de Fuca Ridge flank. ISME J. 7, 161–172. doi: 10.1038/ismej.2012.73
Jungbluth, S. P., Lin, H. T., Cowen, J. P., Glazer, B. T., and Rappé, M. S. (2014). Phylogenetic diversity of microorganisms in subseafloor crustal fluids from holes 1025C and 1026B along the Juan de Fuca Ridge flank. Front. Microbiol. 5:119. doi: 10.3389/fmicb.2014.00119
Kallmeyer, J. (2017). Contamination control for scientific drilling operations. Adv. Appl. Microbiol. 98, 61–91. doi: 10.1016/bs.aambs.2016.09.003
Kallmeyer, J., Mangelsdorf, K., Cragg, B., and Horsfield, B. (2006). Techniques for contamination assessment during drilling for terrestrial subsurface sediments. Geomicrobiol J. 23, 227–239. doi: 10.1080/01490450600724258
Karner, M. B., Delong, E. F., and Karl, D. M. (2001). Archaeal dominance in the mesopelagic zone of the Pacific Ocean. Nature 409, 507–510. doi: 10.1038/35054051
Kashefi, K., and Lovley, D. R. (2003). Extending the upper temperature limit for life. Science 301:934. doi: 10.1126/science.1086823
Kelly, L. C., Cockell, C. S., Thorsteinsson, T., Marteinsson, V., and Stevenson, J. (2014). Pioneer microbial communities of the Fimmvörðuháls lava flow, Eyjafjallajökull, Iceland. Microb. Ecol. 68, 504–518. doi: 10.1007/s00248-014-0432-3
Kern, T., Linge, M., and Rother, M. (2015). Methanobacterium Aggregans sp. nov., a hydrogenotrophic methanogenic archaeon isolated from an anaerobic digester. Int. J. Syst. Evol. Microbiol. 65, 1975–1980. doi: 10.1099/ijs.0.000210
Kieft, T. L. (2010). “Sampling the deep sub-surface using drilling and coring techniques,” in Handbook of Hydrocarbon and Lipid Microbiology. ed. K. N. Timmis (Berlin/Heidelberg: Springer).
Kleine, B. I., Stefánsson, A., Kjartansdóttir, R., Prause, S., Weisenberger, T. B., Reynolds, H. I., et al. (2020). The Surtsey volcano geothermal system: An analogue for seawater-oceanic crust interaction with implications for the elemental budget of the oceanic crust. Chem. Geol. 550:119702. doi: 10.1016/j.chemgeo.2020.119702
Klindworth, A., Pruesse, E., Schweer, T., Peplies, J., Quast, C., Horn, M., et al. (2013). Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 41:e1. doi: 10.1093/nar/gks808
Konhauser, K. O., Schiffman, P., and Fisher, Q. J. (2002). Microbial mediation of authigenic clays during hydrothermal alteration of basaltic tephra, Kilauea volcano. Geochem. Geophys. Geosyst. 3, 1–13. doi: 10.1029/2002GC000317
Korzhenkov, A. A., Teplyuk, A. V., Lebedinsky, A. V., Khvashchevskaya, A. A., Kopylova, Y. G., Arakchaa, K. D., et al. (2018). Members of the uncultured taxon OP1 (“Acetothermia”) predominate in the microbial community of an alkaline hot spring at East-Tuvinian upland. Microbiol. 87, 783–795. doi: 10.1134/S0026261718060115
Lee, M. D., Walworth, N. G., Sylvan, J. B., Edwards, K. J., and Orcutt, B. N. (2015). Microbial communities on seafloor basalts at Dorado Outcrop reflect level of alteration and highlight global lithic clades. Front. Microbiol. 6:1470. doi: 10.3389/fmicb.2015.01470
Lever, M. A., Alperin, M., Engelen, B., Inagaki, F., Nakagawa, S., Steinsbu, B. O., et al. (2006). Trends in basalt and sediment core contamination during IODP expedition 301. Geomicrobiol J. 23, 517–530. doi: 10.1080/01490450600897245
Lever, M. A., Rogers, K. L., Lloyd, K. G., Overmann, J., Schink, B., Thauer, R. K., et al. (2015). Life under extreme energy limitation: A synthesis of laboratory- and field-based investigations. FEMS Microbiol. Rev. 39, 688–728. doi: 10.1093/femsre/fuv020
Lever, M. A., Rouxel, O., Alt, J. C., Shimizu, N., Ono, S., Coggon, R. M., et al. (2013). Evidence for microbial carbon and sulfur cycling in deeply buried ridge flank basalt. Science 339, 1305–1308. doi: 10.1126/science.1229240
Li, J., Mara, P., Schubotz, F., Sylvan, J. B., Burgaud, G., Klein, F., et al. (2020). Recycling and metabolic flexibility dictate life in the lower oceanic crust. Nature 579, 250–255. doi: 10.1038/s41586-020-2075-5
Liu, J. H., Wang, Y. X., Zhang, X. X., Wang, Z. G., Chen, Y. G., Wen, M. L., et al. (2010). Salinarimonas rosea gen. nov., sp. nov., a new member of the α-2 subgroup of the proteobacteria. Int. J. Syst. Evol. Microbiol. 60, 55–60. doi: 10.1099/ijs.0.006981-0
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15:550. doi: 10.1186/s13059-014-0550-8
Lysnes, K., Torsvik, T., Thorseth, I. H., and Pedersen, R. B. (2004). “Microbial populations in ocean floor basalt: Results from ODP leg 187.” in Proc. ODP, Sci. Results. eds. R. B. Pedersen, D. M. Christie, and D. J. Miller 187.
Magnabosco, C., Lin, L. H., Dong, H., Bomberg, M., Ghiorse, W., Stan-Lotter, H., et al. (2018). The biomass and biodiversity of the continental subsurface. Nat. Geosci. 11, 707–717. doi: 10.1038/s41561-018-0221-6
Magnússon, B., Magnússon, S. H., Ólafsson, E., and Sigurdsson, B. D. (2014). Plant colonization, succession and ecosystem development on Surtsey with reference to neighbouring islands. Biogeosciences 11, 5521–5537. doi: 10.5194/bg-11-5521-2014
Marteinsson, V., Klonowski, A., Reynisson, E., Vannier, P., Sigurdsson, B. D., and Ólafsson, M. (2015). Microbial colonization in diverse surface soil types in Surtsey and diversity analysis of its subsurface microbiota. Biogeosciences 12, 1191–1203. doi: 10.5194/bg-12-1191-2015
Martinez Arbizu, P. (2020). pairwiseAdonis: pairwise multilevel comparison using Adonis. R Packag version 04. Available at: https://github.com/pmartinezarbizu/pairwiseAdonis
Mason, O. U., Nakagawa, T., Rosner, M., van Nostrand, J. D., Zhou, J., Maruyama, A., et al. (2010). First investigation of the microbiology of the deepest layer of ocean crust. PLoS One 5:e15399. doi: 10.1371/journal.pone.0015399
Mason, O. U., Stingl, U., Wilhelm, L. J., Moeseneder, M. M., Di Meo-Savoie, C. A., Fisk, M. R., et al. (2007). The phylogeny of endolithic microbes associated with marine basalts. Environ. Microbiol. 9, 2539–2550. doi: 10.1111/j.1462-2920.2007.01372.x
Mcloughlin, N., Staudigel, H., Furnes, H., Eickmann, B., and Ivarsson, M. (2010). Mechanisms of microtunneling in rock substrates: distinguishing endolithic biosignatures from abiotic microtunnels. Geobiology 8, 245–255. doi: 10.1111/j.1472-4669.2010.00243.x
Mcmahon, S., and Parnell, J. (2014). Weighing the deep continental biosphere. FEMS Microbiol. Ecol. 87, 113–120. doi: 10.1111/1574-6941.12196
McMurdie, P. J., and Holmes, S. (2013). Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One 8:e61217. doi: 10.1371/journal.pone.0061217
McPhie, J., White, J. D. L., Gorny, C., Jackson, M. D., Gudmundsson, M. T., and Couper, S. (2020). Lithofacies from the 1963-1967 Surtsey eruption in SUSTAIN drill cores SE-2a, SE-2b and SE-03. Surtsey Res. 14, 19–32. doi: 10.33112/surtsey.14.2
Merino, N., Kawai, M., Boyd, E. S., Colman, D. R., McGlynn, S. E., Nealson, K. H., et al. (2020). Single-cell genomics of novel actinobacteria with the Wood–Ljungdahl pathway discovered in a serpentinizing system. Front. Microbiol. 11:1031. doi: 10.3389/fmicb.2020.01031
Minich, J. J., Sanders, J. G., Amir, A., Humphrey, G., Gilbert, J. A., and Knight, R. (2019). Quantifying and understanding well-to-well contamination in microbiome research. mSystems. 4:e00186-19. doi: 10.1128/mSystems.00186-19
Moore, J. C., and Jackson, M. D. (2020). Observations on the structure of Surtsey. Surtsey Res. 14, 33–45. doi: 10.33112/surtsey.14.3
Motamedi, S., Orcutt, B. N., Früh-Green, G. L., Twing, K. I., Pendleton, H. L., and Brazelton, W. J. (2020). Microbial residents of the Atlantis massif’s shallow serpentinite subsurface. Appl. Environ. Microbiol. 86:e00356-20. doi: 10.1128/AEM.00356-20
Nakagawa, S., Inagaki, F., Suzuki, Y., Steinsbu, B. O., Lever, M. A., Takai, K., et al. (2006). Microbial community in black rust exposed to hot ridge flank crustal fluids. Appl. Environ. Microbiol. 72, 6789–6799. doi: 10.1128/AEM.01238-06
Neufeld, J. D., Schäfer, H., Cox, M. J., Boden, R., McDonald, I. R., and Murrell, J. C. (2007). Stable-isotope probing implicates Methylophaga spp and novel Gammaproteobacteria in marine methanol and methylamine metabolism. ISME J. 1, 480–491. doi: 10.1038/ismej.2007.65
Nigro, L. M., Harris, K., Orcutt, B. N., Hyde, A., Clayton-Luce, S., Becker, K., et al. (2012). Microbial communities at the borehole observatory on the Costa Rica Rift flank (ocean drilling program hole 896A). Front. Microbiol. 3:232. doi: 10.3389/fmicb.2012.00232
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O’Hara, R. B., et al. (2007). The vegan package. Commun. Ecol. Packag 10, 631–637.
Ólafsson, M., and Jakobsson, S. P. (2009). Chemical composition of hydrothermal water and water-rock interactions on Surtsey volcanic island: A preliminary report. Surtsey Res, 12, 29–38.
Orcutt, B. N., Bach, W., Becker, K., Fisher, A. T., Hentscher, M., Toner, B. M., et al. (2011b). Colonization of subsurface microbial observatories deployed in young ocean crust. ISME J. 5, 692–703. doi: 10.1038/ismej.2010.157
Orcutt, B. N., Bergenthal, M., Freudenthal, T., Smith, D., Lilley, M. D., Schnieders, L., et al. (2017). Contamination tracer testing with seabed drills: IODP expedition 357. Sci. Drill. 23, 39–46. doi: 10.5194/sd-23-39-2017
Orcutt, B. N., Daniel, I., and Dasgupta, R. (2019). Book deep carbon: past to present. Cambridge: Cambridge University Press.
Orcutt, B. N., Sylvan, J. B., Knab, N. J., and Edwards, K. J. (2011a). Microbial ecology of the Dark Ocean above, at, and below the seafloor. Microbiol. Mol. Biol. Rev. 75, 361–422. doi: 10.1128/MMBR.00039-10
Orcutt, B. N., Sylvan, J. B., Rogers, D. R., Delaney, J., Lee, R. W., and Girguis, P. R. (2015). Carbon fixation by basalt-hosted microbial communities. Front. Microbiol. 6:904. doi: 10.3389/fmicb.2015.00904
Orcutt, B. N., Wheat, C. G., Rouxel, O., Hulme, S., Edwards, K. J., and Bach, W. (2013). Oxygen consumption rates in subseafloor basaltic crust derived from a reaction transport model. Nat. Commun. 4:2539. doi: 10.1038/ncomms3539
Óskarsson, B. V., Jónasson, K., Valsson, G., and Belart, J. M. C. (2020). Erosion and sedimentation in Surtsey island quantified from new DEMs. Surtsey Res. 4, 63–77. doi: 10.33112/surtsey.14.5
Øvreås, L., Forney, L., Daae, F. L., and Torsvik, V. (1997). Distribution of bacterioplankton in meromictic lake Saelenvannet, as determined by denaturing gradient gel electrophoresis of PCR-amplified gene fragments coding for 16S rRNA. Appl. Environ. Microbiol. 63, 3367–3373. doi: 10.1128/aem.63.9.3367-3373.1997
Parada, A. E., Needham, D. M., and Fuhrman, J. A. (2016). Every base matters: assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 18, 1403–1414. doi: 10.1111/1462-2920.13023
Parkes, J. R., Cragg, B. A., and Wellsbury, P. (2000). Recent studies on bacterial populations and processes in subseafloor sediments. A review. Hydrogeol. J. 8, 11–28. doi: 10.1007/PL00010971
Pedersen, K. (2000). Exploration of deep intraterrestrial microbial life: current perspectives. FEMS Microbiol. Lett. 185, 9–16. doi: 10.1111/j.1574-6968.2000.tb09033.x
Pellizzari, L., Neumann, D., Alawi, M., Voigt, D., Norden, B., and Würdemann, H. (2013). The use of tracers to assess drill-mud penetration depth into sandstone cores during deep drilling: method development and application. Environ. Earth Sci. 70, 3727–3738. doi: 10.1007/s12665-013-2715-2
Pollock, J., Glendinning, L., Wisedchanwet, T., and Watson, M. (2018). The madness of microbiome: attempting to find consensus “best practice” for 16S microbiome studies. Appl. Environ. Microbiol. 84:e02627-17. doi: 10.1128/AEM.02627-17
Prause, S., Weisenberger, T. B., Cappelletti, P., Grimaldi, C., Rispoli, C., Jónasson, K., et al. (2020). Alteration progress within the Surtsey hydrothermal system, SW Iceland – A time-lapse petrographic study of cores drilled in 1979and 2017. J. Volcanol. Geotherm. Res. 392:106754. doi: 10.1016/j.jvolgeores.2019.106754
Prieur, D., and Marteinsson, V. T. (1998). “Prokaryotes living under elevated hydrostatic pressure,” in Biotechnology of Extremophiles. Advances in Biochemical Engineering/Biotechnology. Vol 61. ed. G. Antranikian (Berlin, Heidelberg: Springer).
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., et al. (2013). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41, D590–D596. doi: 10.1093/nar/gks1219
R Core Team (2019). R: A language and environment for statistical computing. R Found Stat Comput. Vienna. Available at: https://www.R-project.org/
Ramírez, G. A., Garber, A. I., Lecoeuvre, A., D’angelo, T., Wheat, C. G., and Orcutt, B. N. (2019). Ecology of subseafloor crustal biofilms. Front. Microbiol. 10:1983. doi: 10.3389/fmicb.2019.01983
Rathsack, K., Stackebrandt, E., Reitner, J., and Schumann, G. (2009). Microorganisms isolated from deep sea low-temperature influenced oceanic crust basalts and sediment samples collected along the Mid-Atlantic Ridge. Geomicrobiol J. 26, 264–274. doi: 10.1080/01490450902892456
Robador, A., Jungbluth, S. P., LaRowe, D. E., Bowers, R. M., Rappé, M. S., Amend, J. P., et al. (2015). Activity and phylogenetic diversity of sulfate-reducing microorganisms in low-temperature subsurface fluids within the upper oceanic crust. Front. Microbiol. 5:748. doi: 10.3389/fmicb.2014.00748
Rstudio, T. (2020). RStudio: Integrated Development for R. Rstudio Team, PBC, Boston, MA. Available at: http://www.rstudio.com/
Santelli, C. M., Banerjee, N., Bach, W., and Edwards, K. J. (2010). Tapping the subsurface ocean crust biosphere: low biomass and drilling-related contamination calls for improved quality controls. Geomicrobiol J. 27, 158–169. doi: 10.1080/01490450903456780
Santelli, C. M., Edgcomb, V. P., Bach, W., and Edwards, K. J. (2009). The diversity and abundance of bacteria inhabiting seafloor lavas positively correlate with rock alteration. Environ. Microbiol. 11, 86–98. doi: 10.1111/j.1462-2920.2008.01743.x
Santelli, C. M., Orcutt, B. N., Banning, E., Bach, W., Moyer, C. L., Sogin, M. L., et al. (2008). Abundance and diversity of microbial life in ocean crust. Nature 453, 653–656. doi: 10.1038/nature06899
Schipper, C. I., Le Voyer, M., Moussallam, Y., White, J. D. L., Thordarson, T., Kimura, J. I., et al. (2016). Degassing and magma mixing during the eruption of Surtsey volcano (Iceland, 1963–1967): the signatures of a dynamic and discrete rift propagation event. Bull. Volcanol. 78:33. doi: 10.1007/s00445-016-1025-4
Sheik, C. S., Reese, B. K., Twing, K. I., Sylvan, J. B., Grim, S. L., Schrenk, M. O., et al. (2018). Identification and removal of contaminant sequences from ribosomal gene databases: lessons from the census of deep life. Front. Microbiol. 9:840. doi: 10.3389/fmicb.2018.00840
Singer, E., Chong, L. S., Heidelberg, J. F., and Edwards, K. J. (2015). Similar microbial communities found on two distant seafloor basalts. Front. Microbiol. 6:1409. doi: 10.3389/fmicb.2015.01409
Smith, A., Popa, R., Fisk, M., Nielsen, M., Wheat, C. G., Jannasch, H. W., et al. (2011). In situ enrichment of ocean crust microbes on igneous minerals and glasses using an osmotic flow-through device. Geochem. Geophys. Geosyst. 6, 1525–2027. doi: 10.1029/2010GC003424
Sorokin, D. Y., Lysenko, A. M., Mityushina, L. L., Tourova, T. P., Jones, B. E., Rainey, F. A., et al. (2001). Thioalkalimicrobium aerophilum gen. nov., sp. nov. and Thioalkalimicrobium sibericum sp. nov., and Thioalkalivibrio versutus gen. nov., sp. nov., Thioalkalivibrio nitratis sp. nov. and Thioalkalivibrio denitrificans sp. nov., novel obligately alkaliphilic. Int. J. Syst. Evol. Microbiol. 51, 565–580. doi: 10.1099/00207713-51-2-565
Sousa, D. Z., Visser, M., Van Gelder, A. H., Boeren, S., Pieterse, M. M., Pinkse, M. W. H., et al. (2018). The deep-subsurface sulfate reducer Desulfotomaculum kuznetsovii employs two methanol-degrading pathways. Nat. Commun. 9:239. doi: 10.1038/s41467-017-02518-9
Spanevello, M. D., Yamamoto, H., and Patel, B. K. C. (2002). Thermaerobacter subterraneus sp. nov., a novel aerobic bacterium from the Great Artesian Basin of Australia, and emendation of the genus Thermaerobacter. Int. J. Syst. Evol. Microbiol. doi: 10.1099/ijs.0.01959-0
Sriswasdi, S., Yang, C. C., and Iwasaki, W. (2017). Generalist species drive microbial dispersion and evolution. Nat. Commun. 8:1162. doi: 10.1038/s41467-017-01265-1
Staudigel, H., Furnes, H., McLoughlin, N., Banerjee, N. R., Connell, L. B., and Templeton, A. (2008). 3.5 billion years of glass bioalteration: volcanic rocks as a basis for microbial life? Earth Sci. Rev. 89, 156–176. doi: 10.1016/j.earscirev.2008.04.005
Sudek, L. A., Wanger, G., Templeton, A. S., Staudigel, H., and Tebo, B. M. (2017). Submarine basaltic glass colonization by the heterotrophic Fe(II)-oxidizing and siderophore-producing deep-sea bacterium pseudomonas stutzeri VS-10: The potential role of basalt in enhancing growth. Front. Microbiol. 8:363. doi: 10.3389/fmicb.2017.00363
Suzuki, Y., Yamashita, S., Kouduka, M., Ao, Y., Mukai, H., Mitsunobu, S., et al. (2020). Deep microbial proliferation at the basalt interface in 33.5–104 million-year-old oceanic crust. Commun. Biol. 3:136. doi: 10.1038/s42003-020-0860-1
Takai, K., and Horikoshi, K. (2000). Rapid detection and quantification of members of the archaeal community by quantitative PCR using fluorogenic probes. Appl. Environ. Microbiol. 66, 5066–5072. doi: 10.1128/AEM.66.11.5066-5072.2000
Takai, K., Inoue, A., and Horikoshi, K. (1999). Thermaerobacter marianensis gen. nov., sp. nov., an aerobic extremely thermophilic marine bacterium from the 11 000 m deep Mariana Trench. Int. J. Syst. Bacteriol. 2, 619–628. doi: 10.1099/00207713-49-2-619
Takai, K., Nakamura, K., Toki, T., Tsunogai, U., Miyazaki, M., Miyazaki, J., et al. (2008). Cell proliferation at 122°C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation. Proc. Natl. Acad. Sci. U. S. A. 105, 10949–10954. doi: 10.1073/pnas.0712334105
Takami, H., Noguchi, H., Takaki, Y., Uchiyama, I., Toyoda, A., Nishi, S., et al. (2012). A deeply branching thermophilic bacterium with an ancient acetyl-CoA pathway dominates a subsurface ecosystem. PLoS One 7:e30559. doi: 10.1371/journal.pone.0030559
Templeton, A. S., Staudigel, H., and Tebo, B. M. (2005). Diverse Mn(II)-oxidizing bacteria isolated from submarine basalts at Loihi seamount. Geomicrobiol J. 22, 127–139. doi: 10.1080/01490450590945951
Þórarinsson, S. (1965). The Surtsey eruption: course of events and the development of the new island. Surtsey Res 1, 51–55.
Thorseth, I. H., Pedersen, R. B., and Christie, D. M. (2003). Microbial alteration of 0-30-Ma seafloor and sub-seafloor basaltic glasses from the Australian Antarctic Discordance. Earth Planet. Sci. Lett. 215, 237–247. doi: 10.1016/S0012-821X(03)00427-8
Thorseth, I. H., Torsvik, T., Torsvik, V., Daae, F. L., and Pedersen, R. B. (2001). Diversity of life in ocean floor basalt. Earth Planet. Sci. Lett. 194, 31–37. doi: 10.1016/S0012-821X(01)00537-4
Tin, S., Bizzoco, R. W., and Kelley, S. T. (2011). Role of the terrestrial subsurface in shaping geothermal spring microbial communities. Environ. Microbiol. Rep. 3, 491–499. doi: 10.1111/j.1758-2229.2011.00248.x
Walton, A. W. (2008). Microtubules in basalt glass from Hawaii Scientific Driling Project #2 phase 1 core and Hilina slope, Hawaii: evidence of the occurrence and behavior of endolithic microorganisms. Geobiology 6, 351–364. doi: 10.1111/j.1472-4669.2008.00149.x
Watanabe, M., Kojima, H., and Fukui, M. (2018). Review of Desulfotomaculum species and proposal of the genera Desulfallas gen. Nov., Desulfofundulus gen. nov., Desulfofarcimen gen. nov. and Desulfohalotomaculum gen. nov. Int. J. Syst. Evol. Microbiol. 68, 2891–2899. doi: 10.1099/ijsem.0.002915
Weisenberger, T. B., Gudmundsson, M. T., Jackson, M. D., Türke, A., Kleine, B. I., and Marshall, B. (2019). Operational report for the 2017 Surtsey underwater volcanic system for thermophiles, alteration processes and INnovative concretes (SUSTAIN) drilling project at Surtsey volcano, Iceland. Geo. Forschungs Zent Rep. Available at: https://gfzpublic.gfz-potsdam.de/rest/items/item_4342891_3/component/file_4342889/content
Yanagawa, K., Nunoura, T., McAllister, S. M., Hirai, M., Breuker, A., Brandt, L., et al. (2013). The first microbiological contamination assessment by deep-sea drilling and coring by the D/V chikyu at the Iheya North hydrothermal field in the Mid-Okinawa trough (IODP Expedition 331). Front. Microbiol. 4:327. doi: 10.3389/fmicb.2013.00327
Zaitseva, S., Lavrentieva, E., Radnagurueva, A., Baturina, O., Kabilov, M., and Barkhutova, D. (2017). Distribution of Acetothermia-dominated microbial communities in alkaline hot springs of Baikal Rift Zone. Peer. J. Preprints. 5:e3492v1. doi: 10.7287/peerj.preprints.3492v1
Zhang, X., Feng, X., and Wang, F. (2016). Diversity and metabolic potentials of subsurface crustal microorganisms from the western flank of the Mid-Atlantic Ridge. Front. Microbiol. 7:363. doi: 10.3389/fmicb.2016.02094
Zhao, R., Dahle, H., Ramírez, G. A., and Jørgensen, S. L. (2020). Indigenous ammonia-oxidizing archaea in oxic subseafloor oceanic crust. mSystems. 5:e00758-19. doi: 10.1128/mSystems.00758-19
Keywords: 16S rRNA gene amplicon sequencing, bacterial and archaeal communities, microbial diversity, extreme environment, subsurface, oceanic basaltic crust, Iceland
Citation: Bergsten P, Vannier P, Klonowski AM, Knobloch S, Gudmundsson MT, Jackson MD and Marteinsson VT (2021) Basalt-Hosted Microbial Communities in the Subsurface of the Young Volcanic Island of Surtsey, Iceland. Front. Microbiol. 12:728977. doi: 10.3389/fmicb.2021.728977
Edited by:
Mark Alexander Lever, ETH Zürich, SwitzerlandReviewed by:
Gustavo Antonio Ramírez, University of North Carolina at Chapel Hill, United StatesCopyright © 2021 Bergsten, Vannier, Klonowski, Knobloch, Gudmundsson, Jackson and Marteinsson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Viggó Thor Marteinsson, dmlnZ29AbWF0aXMuaXM=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.