
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 21 May 2021
Sec. Terrestrial Microbiology
Volume 12 - 2021 | https://doi.org/10.3389/fmicb.2021.658668
This article is part of the Research Topic Exploring Plant Rhizosphere, Phyllosphere and Endosphere Microbial Communities to Improve the Management of Polluted Sites View all 28 articles
 Songhe Chen1†
Songhe Chen1† Xiaoling Xiang1†
Xiaoling Xiang1† Hongliang Ma1
Hongliang Ma1 Petri Penttinen2
Petri Penttinen2 Jiarong Zhao1Han Li1
Jiarong Zhao1Han Li1 Rencai Gao1
Rencai Gao1 Ting Zheng1
Ting Zheng1 Gaoqiong Fan1*
Gaoqiong Fan1*Diazotrophs that carry out the biological fixation of atmospheric dinitrogen (N2) replenish biologically available nitrogen (N) in soil and are influenced by the input of inorganic and organic substrates. To date, little is known about the effects of combined organic substrate addition and N fertilization on the diazotroph community composition and structure in purple soils. We investigated the effects of N fertilization and straw mulching on diazotroph communities by quantifying and sequencing the nifH gene in wheat rhizosphere. The abundance and richness of diazotrophs were greater the higher the fertilization level in the mulched treatments, whereas in the nonmulched treatments (NSMs), richness was lowest with the highest N fertilization level. The abundance and α-diversity of diazotrophs correlated with most of the soil properties but not with pH. At the genus level, the relative abundances of Azospirillum, Bacillus, and Geobacter were higher in the NSMs and those of Pseudacidovorax, Skermanella, Azospira, Paraburkholderia, Azotobacter, Desulfovibrio, Klebsiella, and Pelomonas in the mulched treatments. The differences in community composition between the mulched and the NSMs were associated with differences in soil temperature and soil organic carbon and available potassium contents and C:N ratio. Overall, straw mulching and N fertilization were associated with changes in diazotroph community composition and higher abundance of nifH gene in alkaline purple soils.
Nitrogen (N) is an essential element that plays a crucial role in agroecosystems (Feng et al., 2018). N transformation processes in the N cycle, e.g., N fixation (dinitrogen fixation), nitrification, and organic N mineralization, provide inorganic N to fulfill the requirements of plants (Luo et al., 2018; Pereg et al., 2018). Microbes play an important role in the soil N cycle by regulating N availability and transformation (Canfield et al., 2010). N2-fixing bacteria and archaea, diazotrophs, convert atmospheric N to bioavailable N (Levy-Booth et al., 2014; Chen et al., 2021). Diazotrophs may promote the growth of cereals, even though this has been mostly attributed to their other plant growth-promoting abilities instead of N fixation (Rosenblueth et al., 2018). Diazotrophs are highly diverse and include members of several bacterial phyla, e.g., Proteobacteria, Firmicutes and Cyanobacteria, and Archaea (Rösch et al., 2002). The nifH gene in the diazotrophs encodes a subunit of the nitrogenase enzyme and is highly conserved across the bacterial and archaeal domains, making nifH a suitable molecular biomarker to detect diazotrophic microbes (Zehr et al., 2003; Collavino et al., 2014).
Agricultural management practices, e.g., mulching and fertilization, affect the physiochemical properties of soil (Wang Y.S. et al., 2017). Diazotrophs are sensitive to soil physiochemical properties, including soil pH (Wang Y.S. et al., 2017; Fan et al., 2018), soil organic carbon (SOC) content (Collavino et al., 2014; Yang et al., 2019), C and N availability (Wang C. et al., 2017; Hu et al., 2019; Yang et al., 2021), phosphorus content (Tang et al., 2017), soil moisture (Penton et al., 2016), and C:N ratio (Wang C. et al., 2017; Liu et al., 2019). However, the effects of soil physiochemical properties on the diazotrophs have been variable. For example, the effects of soil pH on the diversity and abundance of soil diazotrophs were inconsistent in different soil types (Wang Y.S. et al., 2017; Fan et al., 2018; Lin et al., 2018). Likewise, N availability has been found to have both stimulatory (Perez et al., 2014; Reardon et al., 2014) and inhibitory effects (Zhalnina et al., 2015; Wang C. et al., 2017) on the diversity of diazotrophs.
The effects of fertilization on diazotroph communities are a growing concern (Meng et al., 2012; Wang C. et al., 2017). Long-term experiments have shown that the application of N fertilizers has a great impact on the diazotroph community composition (Meng et al., 2012; Reardon et al., 2014; Fan et al., 2019). For example, long-term synthetic fertilizer application can decrease the diversity and change the community structure of diazotrophs (Wang C. et al., 2017). However, in several studies, N fertilization did not affect the abundance and community composition of nifH-containing microbes in agricultural soils (Wakelin et al., 2007; Mårtensson et al., 2009; Sun et al., 2015). In general, the addition of organic substrates, e.g., by straw mulching, into soil provides a source of energy and nutrients to support the growth of diazotrophs (Tang et al., 2012; Liao et al., 2018). To date, little is known about the effects of combined straw mulching and N fertilization on the diazotroph community composition in purple soils. Thus, it is important to explore the effects of long-term N application alone and in combination with organic substrates, e.g., straw, on the abundance, diversity, and community composition of diazotrophs to develop sustainable cultivation practices.
The nutrient-rich, neutral or alkaline purple soils in the Sichuan Basin of southwestern China have a complex mineral composition and are characterized with high fertility and suitability to various crops (Wang et al., 2015). However, nitrification activity is high in purple soils, which has resulted in severe soil erosion and N loss via nitrate leaching and runoff (Zhou et al., 2012; Zhao et al., 2018; Zhong et al., 2019). Therefore, increasing N availability is essential for the sustainability of agriculture in the purple soil regions (Wang et al., 2015). This could be at least partially achieved through biological N fixation by diazotrophs.
For this study, we established a field experiment to monitor the effects of inorganic N fertilizer and straw mulching on the community composition of diazotrophs in purple soil. We applied quantitative PCR (qPCR) and high-throughput sequencing targeting the nifH gene to assess variation in the diazotroph community. Our objectives were to (1) evaluate the effects of N fertilization and straw mulching on the abundance, diversity, and community structure of diazotrophs; (2) identify the diazotroph genera affected by N fertilization and straw mulching; and (3) determine the relationships between dominant diazotroph genera and soil properties. We hypothesized that fertilization and straw incorporation would change soil properties, in particular the C and N levels, that would further affect the diazotroph communities; higher C to N ratios were expected to be associated with higher abundance and α-diversity of diazotrophs.
The straw mulch no-tillage experiment was established in 2015 at Renshou experimental base (30°04′N, 104°13′E) of the Agricultural College of Sichuan Agricultural University, China. The station is located in an area with a humid subtropical monsoon climate, an annual average temperature of 17.4°C and a mean annual precipitation of 1,009 mm. The soil is a typical alkaline purple soil. The basic physicochemical properties of the 0–20-cm soil layer in 2015 were as follows: pH 7.82 (soil:water = 1:2.5), organic carbon content 9.78 g kg–1, total nitrogen (TN) content 0.83 g kg–1, total phosphorus (TP) content 0.86 g kg–1, and total potassium content 13.96 g kg–1.
The split plot design experiment had straw mulching as the main factor and nitrogen level as the subplot factor with four 30 m2 (6 m × 5 m) replicate plots per treatment. The straw mulching rates were 0 (NSM: no straw mulching) and 8,000 (SM: straw mulching) kg ha–1. N fertilizer levels were 0 (N0), 120 (N1), and 180 (N2) kg ha–1. P and K fertilizers were applied to all the plots as 75 kg P2O5 ha–1 and 75 kg K2O ha–1, respectively.
Soil samples were collected on March 20, 2019, at the wheat anthesis stage. Fifteen wheat plants per plot were collected gently using a spade, and the soil attached to the roots was pooled into one composite rhizosphere soil sample per plot. The samples were transported on ice in a constant-temperature box to the laboratory and stored at −80°C.
Soil pH and SOC, TN, TP, available P (AP), and available potassium (AK) contents were analyzed as described previously (Liu et al., 2020). NH4+-N and NO3–-N were extracted from fresh soil with 2 mol L–1 KCl and determined using colorimetry (P7 Double Beam UV–Visible Spectrophotometer, Mapada Inc. Shanghai, China) (Liu et al., 2020). Soil temperature was measured using a soil moisture and water potential instrument (Em50G, Decagon Devices Inc. Pullman, United States).
DNA was extracted from 0.5 g of soil using the Fast DNA Spin Kit for Soil (MP Biomedicals, CA, United States) according to the manufacturer’s instructions. The purity and concentration of the extracted DNA were detected by using 2% agarose gel electrophoresis and a Nanodrop 2000 spectrophotometer (Thermo Scientific, United States). The extracted DNA was stored at −20°C.
The nifH gene copy number was determined using real-time qPCR on an ABI7500 Fast Real-Time PCR System (Applied Biosystems Inc. United States). The nifH fragments were amplified in triplicate using primers PolF (5′-TGCGAYCCSAARGCBGACTC-3′) and PolR (5′-ATSGCCATCATYTCRCCGGA-3′) (Poly et al., 2001). Amplification was done in a 20-μl reaction including 10 μl SYBR Green Master Mix (Applied Biosystems, United States), 0.25 μl (10 μM) of each primer, 1 μl (1–10 ng) DNA template or 1 μl sterilized distilled water in the negative control, and 8.5 μl double-distilled water (ddH2O). Amplification was initiated by denaturation at 95°C for 3 min, followed by 35 cycles of denaturation at 95°C for 10 s, annealing at 55°C for 30 s, extension at 72°C for 30 s, and reading the plate at 80°C (Lin et al., 2018).
The standards for qPCR were made by cloning a nifH gene fragment from mixed soil DNA samples. The nifH fragment was amplified as described above using pmD®18-T Vector (TaKaRa, Dalian, China) according to the manufacturer’s instructions. Based on a translated nucleotide query (Altschul et al., 1997), the fragment was 88% similar with nifH from Ruminococcus sp. (sequence ID MBE6845760.1). Plasmid DNA was extracted using a Plasmid Miniprep Kit (Biomiga, Santiago, United States), and the plasmid concentration was measured with a spectrophotometer (Nanodrop 2000, Thermo Scientific, Wilmington, United States) (Liao et al., 2018). As the sequences of the vector and PCR inserts were known, the copy number of nifH was calculated directly from the concentration of extracted nonlinearized plasmid DNA. A standard curve was done using 10-fold serial dilutions from 1.13 × 103 to 1.13 × 109 nifH copies (Wang et al., 2012). Since Nanodrop measurements are prone to overestimate DNA concentration (Kapp et al., 2015), the plasmid copy numbers were likely overestimated. However, this overestimation was not expected to affect comparisons between treatments. The amplification efficiency was 81.8% with R2-value of 0.999 and slope of −3.85. No amplification was detected in the negative controls. The detected CT values in samples were all within the range of the standard curve.
The nifH amplicons of approximately 360 bp were amplified using the primers PolF with a unique sample identifying barcode and PolR. Amplification was done in triplicate 25 μl reactions including 1 μl of each 10 μM primer, 2 μl (20 ng) of template DNA or 2 μl sterilized distilled water in the negative control, and 12.5 μl of MasterMix containing Taq DNA polymerase, PCR buffer, Mg2+ and dNTPs (CWBIO, China), and 8.5 μl double distilled water (ddH2O). The amplification program included initial denaturation at 95°C for 3 min, followed by 40 cycles of 95°C for 30 s, 60°C for 60 s, and 72°C for 60 s, and a final extension at 72°C for 10 min (Gaby et al., 2018). The triplicate PCR products of each sample were mixed and purified with a gel extraction kit (TIANGEN, Inc., Beijing, China) according to the manufacturer’s instructions and quantified using a Qubit 2.0 fluorimeter (Invitrogen, Carlsbad, CA, United States). No amplification was detected in the negative controls. Purified amplicons were pooled in equimolar concentrations and sequenced using MiSeq Reagent Kit V3 for paired-end sequencing on an Illumina MiSeq platform at Shanghai Personal Biotechnology Co., Ltd (Shanghai, China).
The raw sequences were demultiplexed and low-quality sequences were removed using the QIIME2 pipeline (Bolyen et al., 2019). Reads with an average quality score below 20 were filtered out and the remaining reads were trimmed at 200 and 450 bp as minimum and maximum length, respectively. Chimeric sequences were removed using UCHIME against a reference alignment (Edgar et al., 2011). A total of 1,965,677 high-quality sequences were obtained from 24 samples. The UPARSE pipeline was used to cluster sequences into operational taxonomic units (OTUs) (Edgar, 2013). Sequences with 97% similarity were assigned to an OTU (Edgar, 2010), after removing the singleton OTUs, generating 5,283 OTUs.
The statistical analyses were conducted using SPSS 22.0 software (International Business Machine, Armonk, New York, United States). Differences in soil properties, nifH abundance, and α-diversity were tested using two-way ANOVA followed by Fisher’s least significant difference (LSD) test. The nifH abundance data were log10 transformed and checked for normality prior to the test. The relationships between soil properties, nifH abundance, and α-diversity were estimated using Pearson correlation analysis. β-Diversity was visualized using weighted UniFrac distance-based principal coordinate analysis (PCoA) in R package “ape” (Hu et al., 2020). Hierarchical cluster analysis was performed using the “hclust” function with the average linkage algorithm in R version 3.5.1 (R Development Core Team, 2012). Corrplot in R was used to visualize the results of Pearson correlation analysis. Differences in diazotroph community structures were tested using permutational multivariate analysis of variance (PERMANOVA) in the R package vegan (Oksanen et al., 2012). The relationships between community variation and environmental factors were tested using redundancy analysis (RDA) with 999 permutations (Legendre and Anderson, 1999). Linear discriminant analysis effect size (LEfSe) analysis with default parameters was used to detect differentially abundant taxa (Segata et al., 2011).
The nifH gene copy numbers ranged from 3.2 × 107 to 4.7 × 108 per g soil, with the lowest and the highest numbers in the NSMN2 and SMN2 treatment, respectively (Figure 1). In the mulched treatment (SM), the abundance of nifH gene was higher, the higher the nitrogen fertilization level (P < 0.05) (Figure 1). Compared with the nonmulched treatment (NSM), the diversity and richness of diazotrophs were higher in the SM (P < 0.05) (Figure 2). Furthermore, N fertilization resulted in higher diazotroph diversity and richness in the SM. In the NSM, richness was lowest in the highest fertilization level (P < 0.05) (Figure 2).
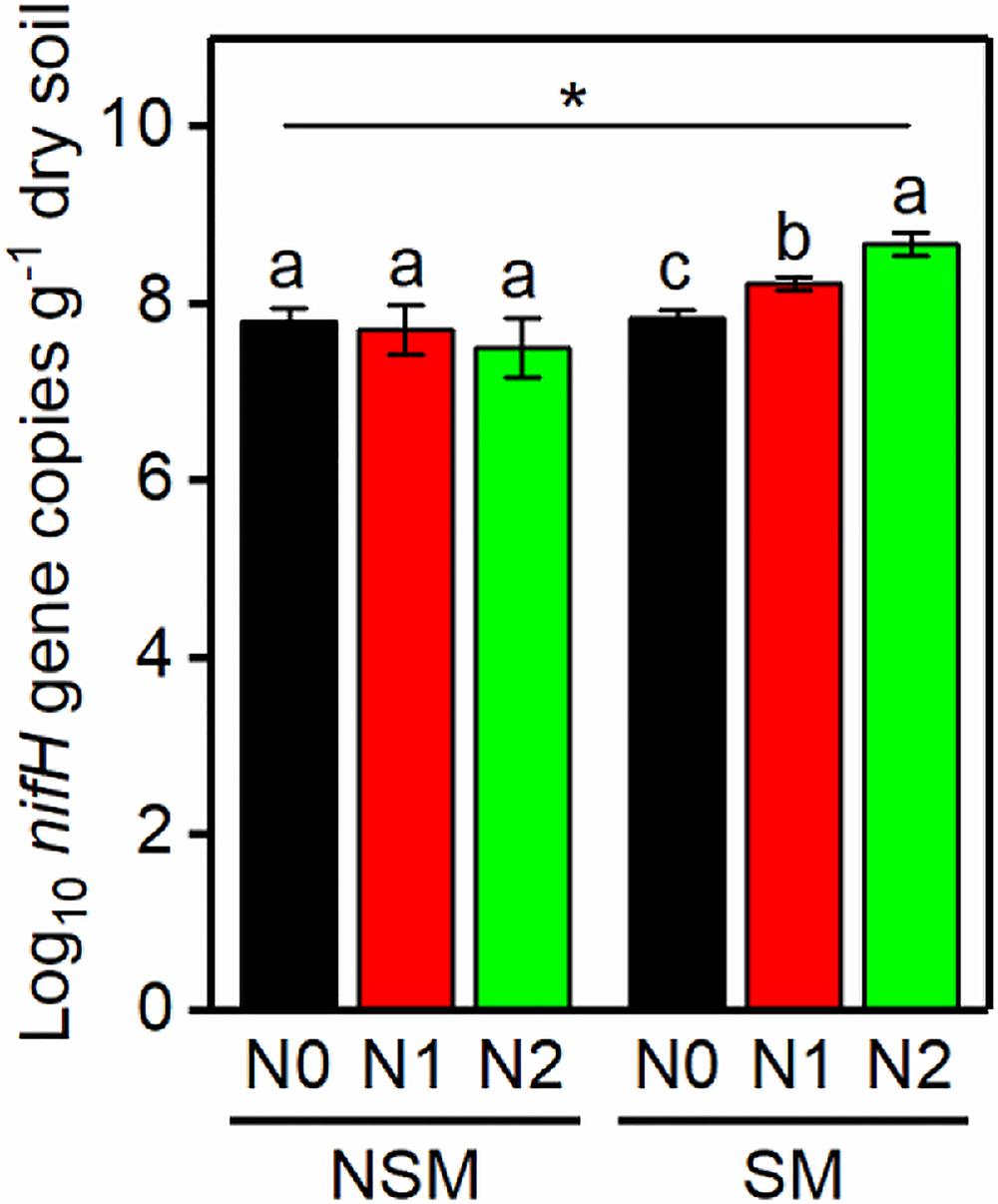
Figure 1. The abundance of nifH gene copies in wheat rhizosphere soil under straw mulching and nitrogen fertilization treatments. NSM, no straw mulching; SM, straw mulching; N0, no nitrogen; N1, 120 kg N ha− 1; N2, 180 kg N ha− 1. Data shown as mean ± SD. *indicates statistically significant difference (P < 0.05) between NSM and SM. Different letters above columns indicate statistically significant difference (P < 0.05) between N fertilizer levels within NSM and SM.
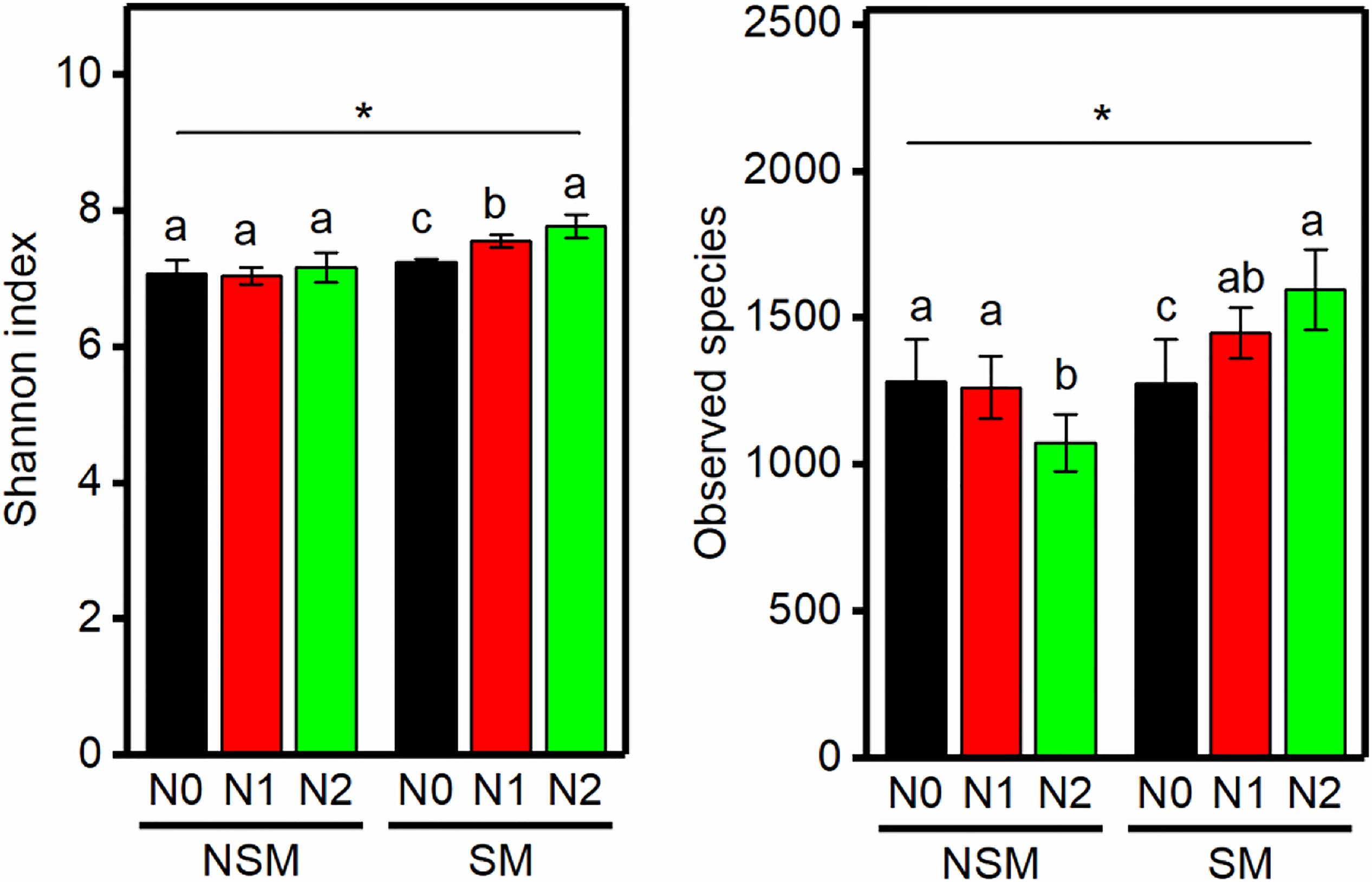
Figure 2. The α-diversity of the diazotrophs in wheat rhizosphere soil under straw mulching and nitrogen fertilization treatments. NSM, no straw mulching; SM, straw mulching; N0, no nitrogen; N1, 120 kg N ha− 1; N2, 180 kg N ha− 1. Data shown as mean ± SD. *indicates statistically significant difference (P < 0.05) between NSM and SM. Different letters above columns indicate statistically significant difference (P < 0.05) between N fertilizer levels within NSM and SM.
The nifH communities were distinct in all the six mulching–fertilizer level combinations (P < 0.05) (Supplementary Table 1). The first and the second principal coordinates explained 50.5% of the variation in the community composition in the PCoA (Figure 3A). The mulched and NSMs were clearly separated along axis 1, and the nonfertilized treatments NSM0 and SMN0 were separated from the fertilized treatments (P < 0.05) (Figure 3A). Similarly, the samples were divided into three clusters in the hierarchical cluster analysis: the mulched and fertilized, the nonmulched and fertilized, and the nonfertilized (Figure 3B), showing that both mulching and N fertilization affected the diazotroph community structure.
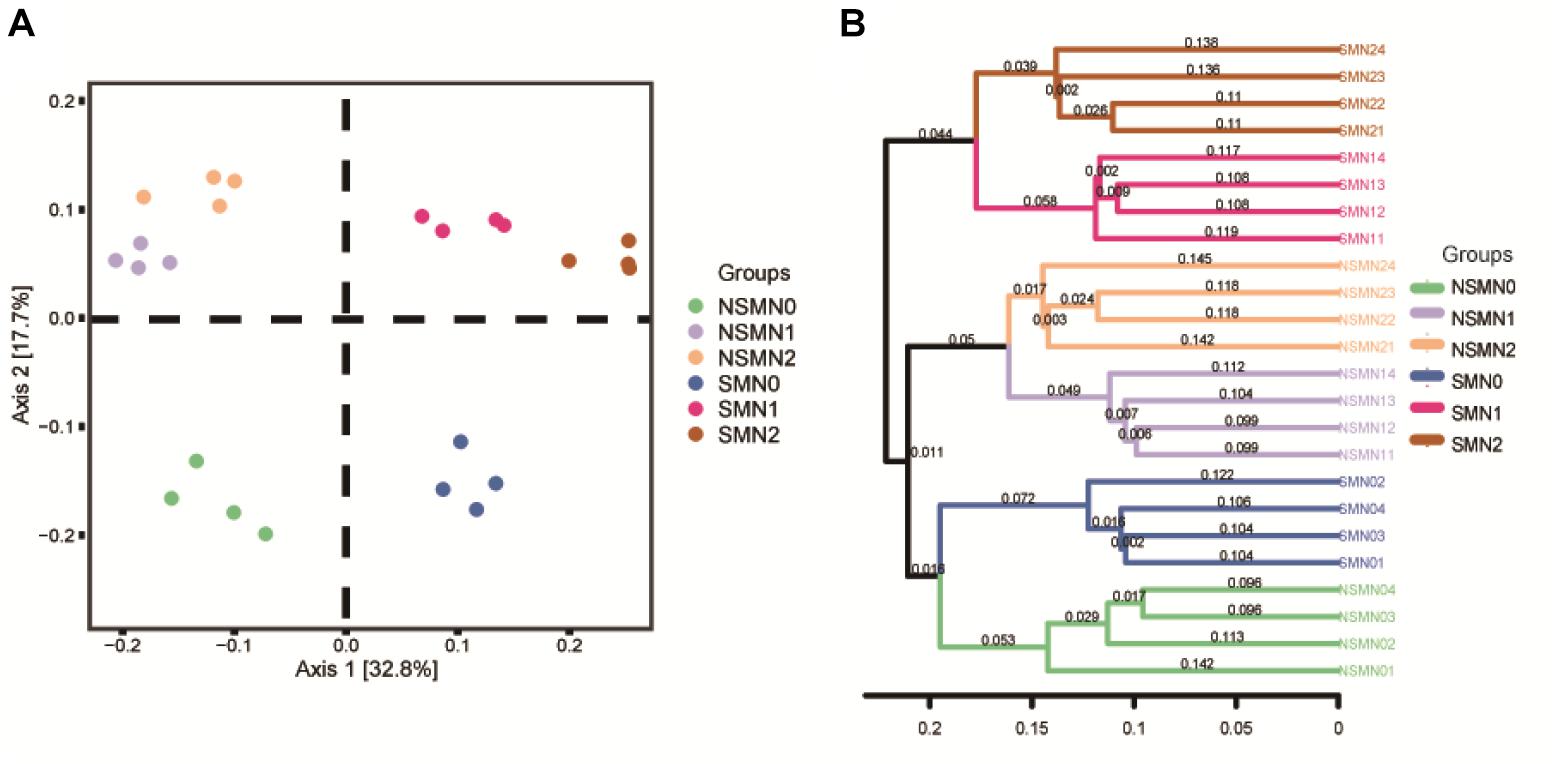
Figure 3. Principal coordinates analysis (A) and hierarchical clustering (B) of diazotroph communities in wheat rhizosphere soil under straw mulching and nitrogen fertilization treatments. NSM, no straw mulching; SM, straw mulching; N0, no nitrogen; N1, 120 kg N ha− 1; N2, 180 kg N ha− 1.
The relative abundance of the phylum Proteobacteria accounted for 88.5–92.9% of the total relative abundance in all the treatments (Figure 4A). The relative abundances of the classes Deltaproteobacteria, Betaproteobacteria, Gammaproteobacteria, and Bacilli and those of the genera Azospirillum, Bradyrhizobium, and Desulfovibrio were high in all treatments (Figures 4B,C). In the LEfSe analysis, 17 and 21 taxa in the nonmulched and the mulched treatments, respectively, had large effect sizes with LDA score >2.5 (Supplementary Table 2). At the genus level, the relative abundances of Azospirillum, Bacillus, and Geobacter were higher in the NSMs, and those of Pseudacidovorax, Skermanella, Azospira, Paraburkholderia, Azotobacter, Desulfovibrio, Klebsiella, and Pelomonas were higher in the mulched treatments (LDA score > 3.5) (Figure 5 and Supplementary Table 2).
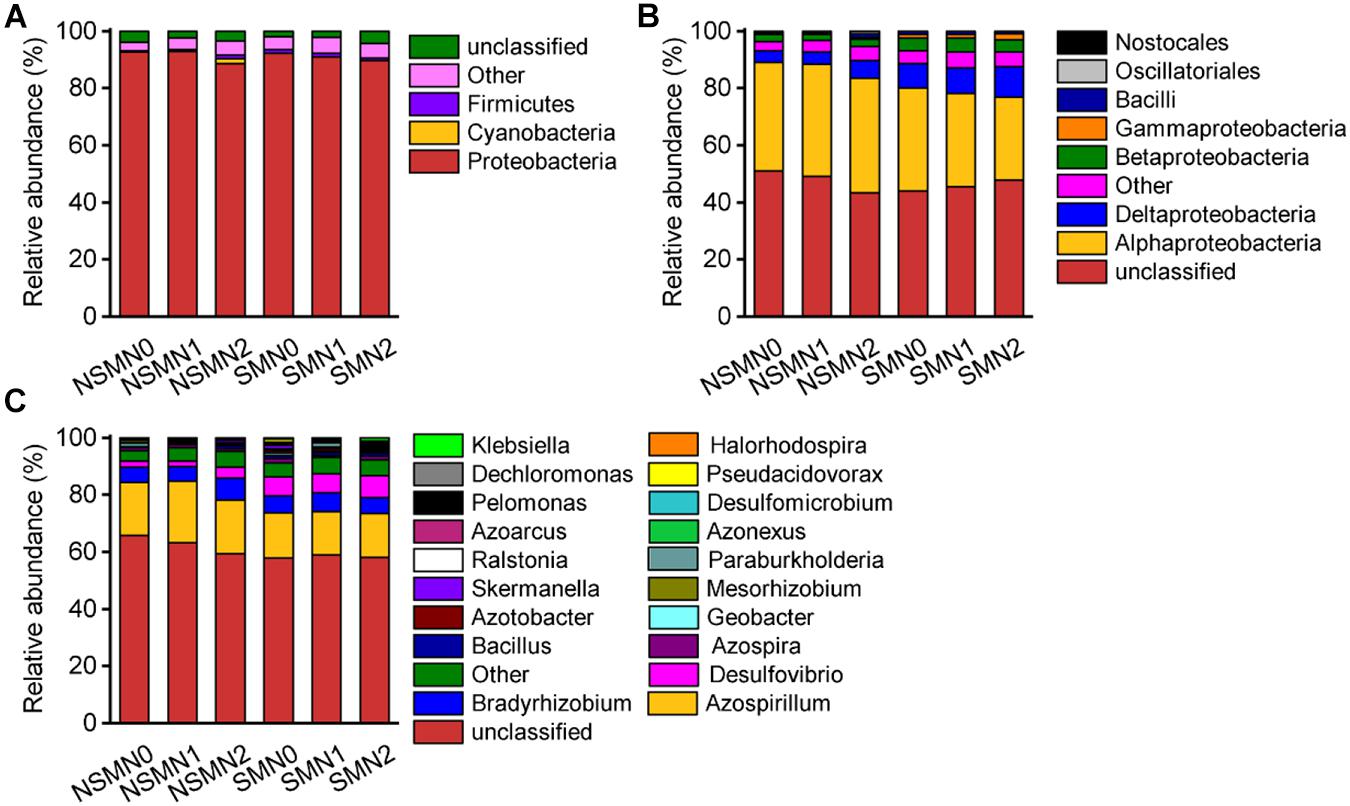
Figure 4. The community composition of diazotrophs in wheat rhizosphere soil under straw mulching and nitrogen fertilization treatments. (A) Phylum level; (B) class level; (C) genus level. NSM, no straw mulching; SM, straw mulching; N0, no nitrogen; N1, 120 kg N ha− 1; N2, 180 kg N ha− 1.
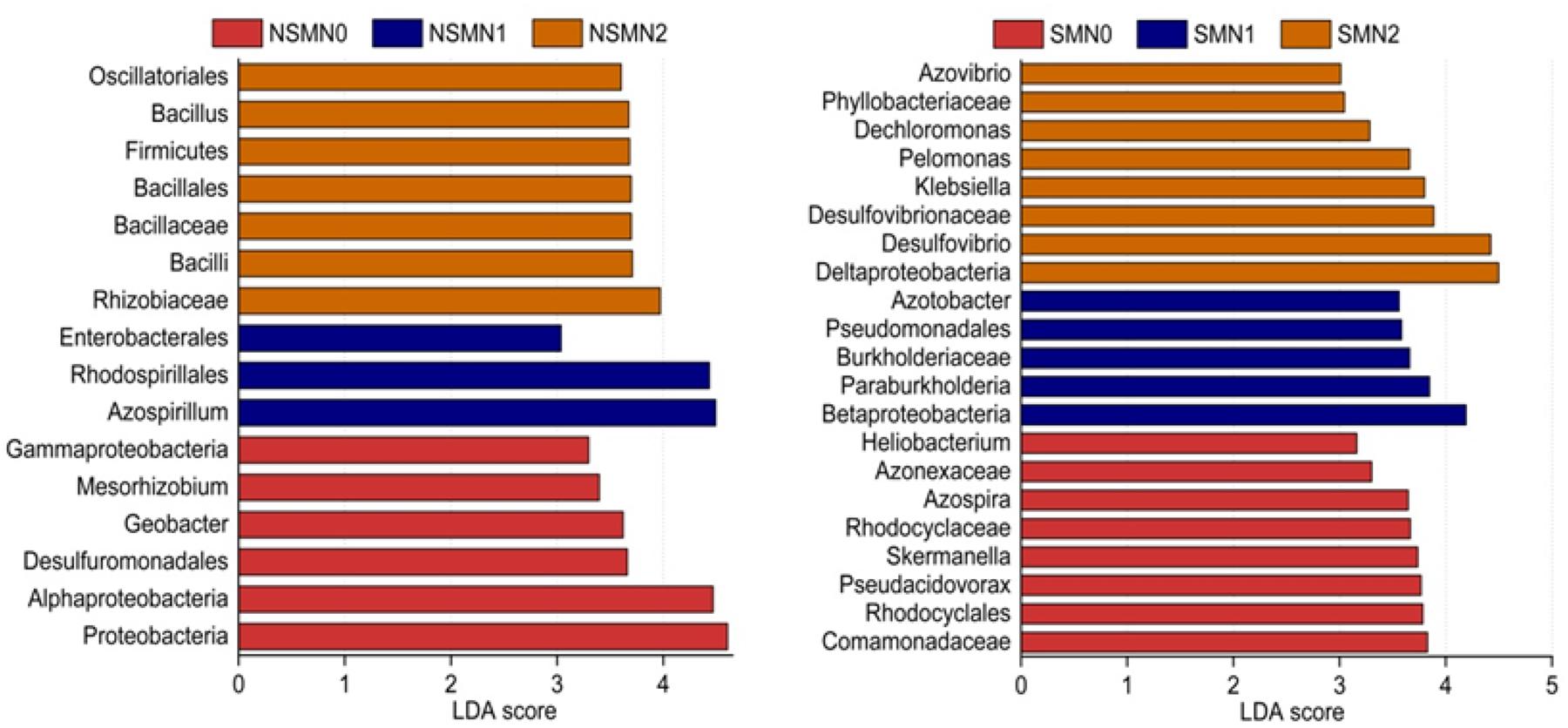
Figure 5. Differentially abundant diazotroph taxa in wheat rhizosphere soil under straw mulching and nitrogen fertilization treatments. Detected using linear discriminant analysis effect size analysis. NSM, no straw mulching; SM, straw mulching; N0, no nitrogen; N1, 120 kg N ha− 1; N2, 180 kg N ha− 1.
Shannon diversity correlated positively with SOC, TN, AN, NH4+-N, NO3–-N, AP, and AK contents and C:N ratio and negatively with soil temperature (P < 0.05), and the number of observed species correlated positively with SOC, AN, NO3–-N, AP, and AK contents and C:N ratio and negatively with soil temperature (P < 0.05) (Table 1). The abundance of nifH gene correlated positively with SOC, AK, AP, AN, NH4+-N, and NO3–-N contents and C:N ratio and negatively with soil temperature (P < 0.05) (Table 1). The relationships between soil physicochemical properties and the diazotroph community composition at the genus level were visualized using RDA (Figure 6 and Supplementary Table 3). Differences in community composition between the mulched and the NSMs were associated with differences in soil temperature, C:N ratio, and SOC and AK contents (Figure 6). The genus Azospirillum correlated positively with soil temperature, whereas the genera Dechloromonas and Desulfovibrio correlated positively with C:N ratio and SOC and AK contents. The differences between the fertilized and the nonfertilized treatments were associated with differences in NH4+-N, AN, AP, and TN contents. The genus Azorhizobium correlated positively and Mesorhizobium negatively with soil TN, NH4+-N, AN, and AP contents (Figure 6).
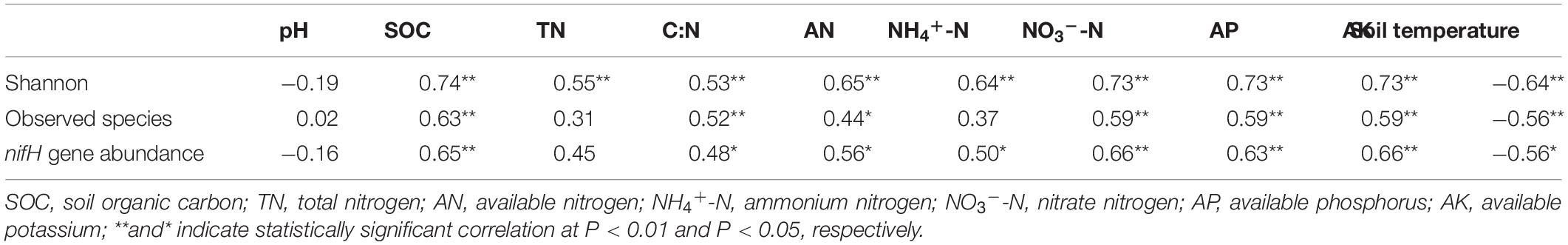
Table 1. The correlations of soil properties with α-diversity and nifH gene abundance in the wheat rhizosphere.
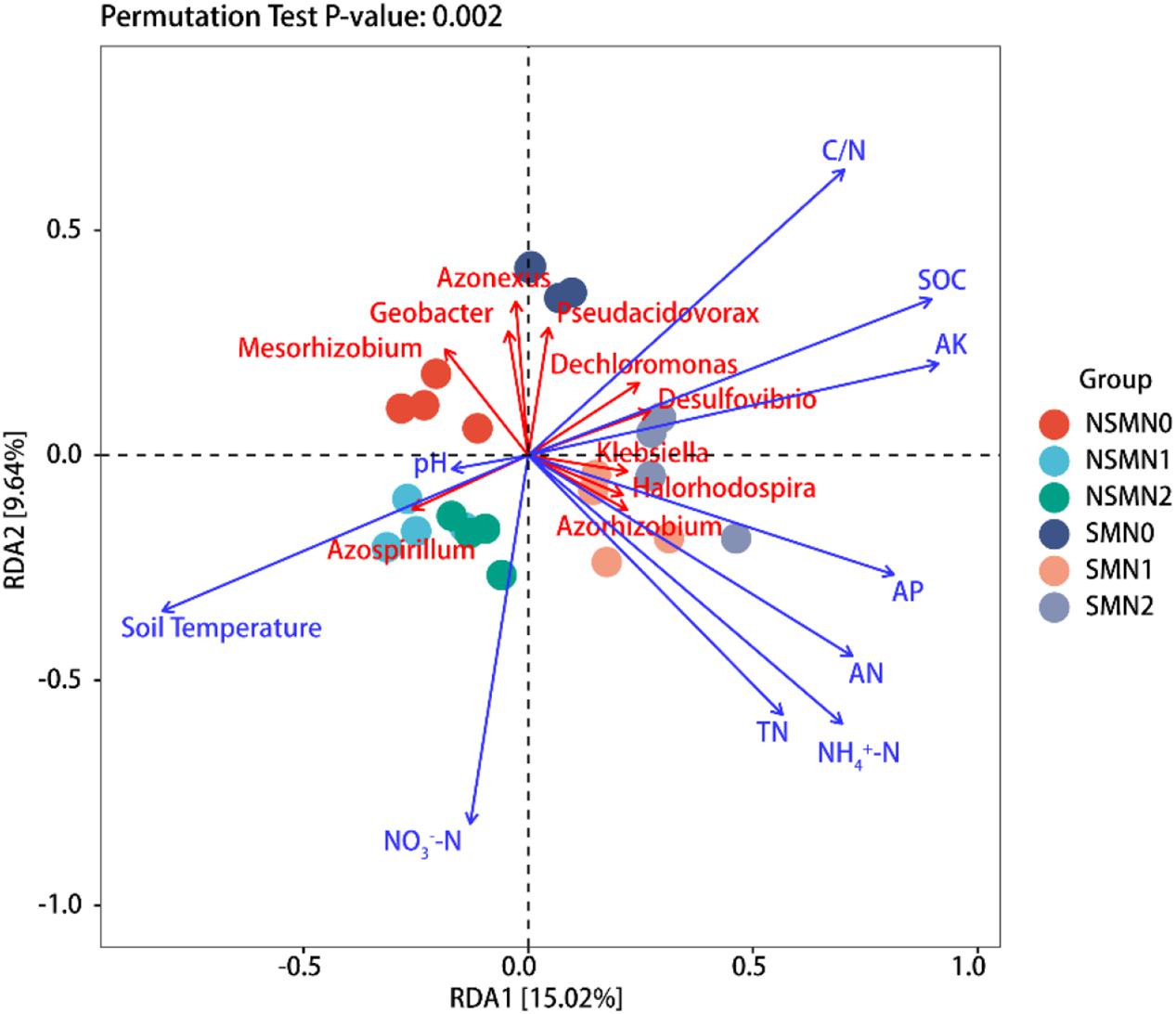
Figure 6. Redundancy analysis (RDA) of the nifH community and soil environmental factors in the wheat rhizosphere. SOC, soil organic carbon; TN, total nitrogen; AN, available nitrogen; NH4+-N, ammonium nitrogen; NO3–-N, nitrate nitrogen; AP, available phosphorus; AK, available potassium; NSM, no straw mulching; SM, straw mulching; N0, no nitrogen; N1, 120 kg N ha− 1; N2, 180 kg N ha− 1.
Soil diazotrophs include Bacteria and Archaea that fix atmospheric nitrogen as free-living (e.g., Azospirillum and Azotobacter) or in symbiosis (e.g., Bradyrhizobium and Rhizobium) (De Bruijn, 2015). The abundance, diversity, and community composition of soil diazotrophs have been associated with various factors such as soil physicochemical characteristics and management strategies (Reardon et al., 2014). Fertilization and straw return change the environmental conditions and may affect the diazotrophs due to their individual responses to the changes in soil properties (Romero et al., 2012; Wang et al., 2016; Yang et al., 2019). We assessed the response of diazotrophs to straw mulching and N fertilization by quantifying and sequencing the nifH gene and noticed that the variation in diazotroph communities was more associated with straw mulching than with N fertilization. In previous studies, stubble retention and straw incorporation increased the abundance, diversity, and richness of the nifH carrying diazotrophs, which is possibly associated with the input of labile carbohydrates; monosaccharides and polysaccharides in straw provided an energy and nutrient source for the generally heterotrophic diazotrophs (Park et al., 2009). The N fertilization-accompanied soil acidification might in turn decrease the abundance and diversity of diazotrophs (Meng et al., 2012; Reardon et al., 2014). We found that the abundance and α-diversity of diazotrophs were greater the higher the fertilization level in the mulched treatments, whereas in the NSMs, richness was lowest with the highest N fertilization level, indicating that the slightly negative effect of N fertilization had been offset by mulching.
The availability of soil nutrients, which is highly responsive to straw and fertilizer input, is crucial for the establishment of soil diazotroph community structure (Reardon et al., 2014; Hu et al., 2019; Yang et al., 2019). In our study, Pearson correlation analysis showed that Shannon diversity was associated with differences in soil temperature, SOC, TN, AN, NH4+-N, NO3–-N, AP, and AK contents and C:N ratio, and the number of observed species was associated with differences in soil temperature; SOC, AN, NO3–-N, AP, and AK contents; and C:N ratio. In agreement, the results of RDA suggested that the differences in diazotroph communities were associated with differences in soil temperature; NH4+-N, AN, NO3–-N, AP, and TN contents; and C:N ratio. Especially, the differences between the mulched and NSMs possibly resulted from the higher SOC and AK contents in the mulched treatments.
Consistent with previous studies (Reardon et al., 2014; Lin et al., 2018; Hu et al., 2019, 2020), we observed that the relative abundances of Proteobacteria that play a vital role in carbon cycle and nitrogen mineralization (Wu et al., 2020) were high. However, it is worth noting that concluding absolute abundances directly from the relative abundance data is not possible, for example, due to bias introduced by the primers (McLaren et al., 2019). Consistent with Hu et al. (2019), the relative abundances of the class α-Proteobacteria and the genera Azospirillum within this class were high. Azospirillum is the best characterized genus of plant growth-promoting rhizobacteria and increased the yield and dry weight of wheat (Lin et al., 2018). The diazotroph communities may respond differentially to environmental factors and cultivation practices in different soil types (Wang C. et al., 2017; Lin et al., 2018). Contrary to Wang C. et al. (2017), in our study, Azospirillum correlated negatively with SOC content, possibly due to the differential response.
The soil N cycle is generally coupled with sulfur and iron cycles, and high N availability can increase the iron and sulfur cycle-related genes (Bao et al., 2014; Minamisawa et al., 2016). N limitation inhibited the growth of sulfate-reducing bacteria and the relative abundance of Desulfovibrionales that are mostly sulfate reducers increased with N fertilizer input (Sim et al., 2012; Li et al., 2018). In agreement, we found that the relative abundance of Desulfovibrio was the highest in the mulched and high N fertilization level treatment. In addition, straw mulching combined with N fertilization led to higher relative abundances of Azotobacter, Klebsiella, and Paraburkholderia that participate in the N cycle in soil ecosystems and have plant growth-promoting properties (Li et al., 2019; Patel et al., 2019; Castellano-Hinojosa and Strauss, 2020; Liu et al., 2020; Mukherjee et al., 2020; Ravi et al., 2020), suggesting that straw return may further contribute to the growth of plants.
In summary, our study demonstrated that straw mulching and N fertilization lead to changes in soil diazotroph community composition and to higher abundance, diversity, and richness of the nifH gene. The differences in community composition between the mulched and NSMs were associated with differences in soil temperature, C/N, SOC, and AK contents. Straw mulching combined with N fertilization was characterized by higher relative abundances of plant growth-promoting Azotobacter, Klebsiella, and Paraburkholderia genera. Overall, straw mulching and N fertilization were associated with changes in diazotroph community composition and increased the abundance of the nifH gene.
The nifH amplicon sequences were deposited in the National Center for Biotechnology Information (NCBI) GenBank database with the accession number SRP299232. The sequence of the nifH qPCR standard was deposited in the NCBI GenBank database with the accession number MW817080.
All authors listed have made substantial, direct and intellectual contribution to the work, and approved it for publication.
The study was funded by the Special Fund for National Key Research and Development Program (grant number 2016YFD0300406, 2016) and Agro-scientific Research in Public Interest (grant number 20150312705, 2015).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2021.658668/full#supplementary-material
Altschul, S. F., Madden, T. L., Schäffer, A. A., Zhang, J., Zhang, Z., Miller, W., et al. (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25, 3389–3402. doi: 10.1093/nar/25.17.3389
Bao, Z. H., Okubo, T., Kubota, K., Kasahara, Y., Tsurumaru, H., Anda, M., et al. (2014). Metaproteomic identification of diazotrophic methanotrophs and their localization in root tissues of field-grown rice plants. Appl. Environ. Microbiol. 80, 5043–5052. doi: 10.1128/aem.00969-14
Bolyen, E., Rideout, J. R., Dillon, M. R., Bokulich, N. A., Abnet, C. C., Al-Ghalith, G. A., et al. (2019). Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857.
Canfield, D. E., Glazer, A. N., and Falkowski, P. G. (2010). The evolution and future of Earth’s nitrogen cycle. Science 330, 192–196.
Castellano-Hinojosa, A., and Strauss, S. L. (2020). Impact of cover crops on the soil microbiome of tree crops. Microorganisms 8:328. doi: 10.3390/microorganisms8030328
Chen, H., Zheng, C. Y., Qiao, Y. Q., Du, S. Z., Li, W., Zhang, X. Q., et al. (2021). Long-term organic and inorganic fertilization alters the diazotrophic abundance, community structure, and co-occurrence patterns in a vertisol. Sci. Total Environ. 766:142441. doi: 10.1016/j.scitotenv.2020.142441
Collavino, M. M., Tripp, H. J., Frank, I. E., Vidoz, M. L., Calderoli, P. A., Donato, M., et al. (2014). nifH pyrosequencing reveals the potential for location-specific soil chemistry to influence N2-fixing community dynamics. Environ. Microbiol. 16, 3211–3223. doi: 10.1111/1462-2920.12423
De Bruijn, F. J. (2015). Biological Nitrogen Fixation. In Principles of Plant-Microbe Interactions. Cham: Springer, 215–224.
Edgar, R. C. (2010). Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26, 2460–2461. doi: 10.1093/bioinformatics/btq461
Edgar, R. C. (2013). UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 10, 996–998. doi: 10.1038/nmeth.2604
Edgar, R. C., Haas, B. J., Clemente, J. C., Quince, C., and Knight, R. (2011). UCHIME improves sensitivity and speed of chimera detection. Bionformatics 27, 2194–2200. doi: 10.1093/bioinformatics/btr381
Fan, K. K., Delgado-Baquerizo, M., Guo, X. S., Wang, D. Z., Wu, Y. Y., Zhu, M., et al. (2019). Suppressed N fixation and diazotrophs after four decades of fertilization. Microbiome 7:143.
Fan, K. K., Weisenhorn, P., Gilbert, J. A., Shi, Y., Bai, Y., and Chu, H. Y. (2018). Soil pH correlates with the co-occurrence and assemblage process of diazotrophic communities in rhizosphere and bulk soils of wheat fields. Soil Biol. Biochem. 121, 185–192. doi: 10.1016/j.soilbio.2018.03.017
Feng, M. M., Adams, J. M., Fan, K. K., Shi, Y., Sun, R. B., Wang, D. Z., et al. (2018). Long-term fertilization influences community assembly processes of soil diazotrophs. Soil Biol. Biochem. 126, 151–158. doi: 10.1016/j.soilbio.2018.08.021
Gaby, J. C., Rishishwar, L., Valderrama-Aguirre, L. C., Green, S. J., Valderrama-Aguirre, A., Jordan, I. K., et al. (2018). Diazotroph community characterization via a high-throughput nifH amplicon sequencing and analysis pipeline. Appl. Environ. Microbiol. 84:e00713.
Hu, X. J., Liang, A. Z., Yao, Q., Liu, Z. X., Yu, Z. H., Wang, G. H., et al. (2020). Ridge tillage improves soil properties, sustains diazotrophic communities, and enhances extensively cooperative interactions among diazotrophs in a clay loam soil. Front. Microbiol. 11:1333. doi: 10.3389/fmicb.2020.01333
Hu, X. J., Liu, J. J., Wei, D., Zhou, B. K., Chen, X. L., Jin, J., et al. (2019). Long-term application of nitrogen, not phosphate or potassium, significantly alters the diazotrophic community compositions and structures in a Mollisol in northeast China. Res. Microbiol. 170, 147–155. doi: 10.1016/j.resmic.2019.02.002
Kapp, J. R., Diss, T., Spicer, J., Gandy, M., Schrijver, I., Jennings, L. J., et al. (2015). Variation in pre-PCR processing of FFPE samples leads to discrepancies in BRAF and EGFR mutation detection: a diagnostic RING trial. J. Clin. Pathol. 68, 111–118. doi: 10.1136/jclinpath-2014-202644
Legendre, P., and Anderson, M. J. (1999). Distance-based redundancy analysis: testing multispecies responses in multifactorial ecological experiments. Ecol. Monographs 69, 1–24. doi: 10.1890/0012-9615(1999)069[0001:dbratm]2.0.co;2
Levy-Booth, D. J., Prescott, C. E., and Grayston, S. J. (2014). Microbial functional genes involved in nitrogen fixation, nitrification and denitrification in forest ecosystems. Soil Biol. Biochem. 75, 11–25. doi: 10.1016/j.soilbio.2014.03.021
Li, Y., Sun, H., Wu, Z. J., Li, H., and Sun, Q. Y. (2019). Urban traffic changes the biodiversity, abundance, and activity of phyllospheric nitrogen-fixing bacteria. Environ. Sci. Pollution Res. 26, 16097–16104. doi: 10.1007/s11356-019-05008-1
Li, Y. Y., Pan, F. X., and Yao, H. Y. (2018). Response of symbiotic and asymbiotic nitrogen-fixing microorganisms to nitrogen fertilizer application. J. Soils Sediments 19, 1948–1958. doi: 10.1007/s11368-018-2192-z
Liao, H. K., Li, Y. Y., and Yao, H. Y. (2018). Fertilization with inorganic and organic nutrients changes diazotroph community composition and N-fixation rates. J. Soils Sediments 18, 1076–1086. doi: 10.1007/s11368-017-1836-8
Lin, Y. X., Ye, G. P., Liu, D. Y., Ledgard, S., Luo, J. F., Fan, J. B., et al. (2018). Long-term application of lime or pig manure rather than plant residues suppressed diazotroph abundance and diversity and altered community structure in an acidic Ultisol. Soil Biol. Biochem. 123, 218–228. doi: 10.1016/j.soilbio.2018.05.018
Liu, C., Li, L. L., Xie, J. H., Coulter, J. A., Zhang, R. Z., Luo, Z. Z., et al. (2020). Soil bacterial diversity and potential functions are regulated by long-term conservation tillage and straw mulching. Microorganisms 8:836. doi: 10.3390/microorganisms8060836
Liu, X. Y., Liu, C., Gao, W. H., Xue, C., Guo, Z. H., Jiang, L., et al. (2019). Impact of biochar amendment on the abundance and structure of diazotrophic community in an alkaline soil. Sci. Total Environ. 688, 944–951. doi: 10.1016/j.scitotenv.2019.06.293
Luo, G. W., Friman, V. P., Chen, H., Liu, M. Q., Wang, M., Guo, S. W., et al. (2018). Long-term fertilization regimes drive the abundance and composition of N-cycling-related prokaryotic groups via soil particle-size differentiation. Soil Biol. Biochem. 116, 213–223. doi: 10.1016/j.soilbio.2017.10.015
Mårtensson, L., Díez, B., Wartiainen, I., Zheng, W., El-Shehawy, R., and Rasmussen, U. (2009). Diazotrophic diversity, nifH gene expression and nitrogenase activity in a rice paddy field in Fujian. China. Plant Soil 325, 207–218. doi: 10.1007/s11104-009-9970-8
McLaren, M. R., Willis, A. D., and Callahan, B. J. (2019). Consistent and correctable bias in metagenomic sequencing experiments. Elife 8:e46923.
Meng, X. F., Wang, L., Long, X. H., Liu, Z. P., Zhang, Z. H., and Zed, R. (2012). Influence of nitrogen fertilization on diazotrophic communities in the rhizosphere of the Jerusalem artichoke (Helianthus tuberosus L.). Res. Microbiol. 163, 349–356. doi: 10.1016/j.resmic.2012.03.005
Minamisawa, K., Imaizumi-Anraku, H., Bao, Z. H., Shinoda, R., Okubo, T., and Ikeda, S. (2016). Are symbiotic methanotrophs key microbes for N acquisition in paddy rice root? Microbes Environ. 31, 4–10. doi: 10.1264/jsme2.me15180
Mukherjee, T., Banik, A., and Mukhopadhyay, S. K. (2020). Plant growth-promoting traits of a thermophilic strain of the klebsiella group with its effect on rice plant growth. Curr. Microbiol. 77, 2613–2622. doi: 10.1007/s00284-020-02032-0
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O’Hara, R., et al. (2012). Vegan: Community Ecology Package. R Package, Version 2.
Park, J. Y., Seyama, T., Shiroma, R., Ike, M., Srichuwong, S., Nagata, K., et al. (2009). Efficient recovery of glucose and fructose via enzymatic saccharification of rice straw with soft carbohydrates. Biosci. Biotechnol. Biochem. 73, 1072–1077. doi: 10.1271/bbb.80840
Patel, N. P., Raju, M., Haldar, S., and Chatterjee, P. B. (2019). Characterization of phenazine-1-carboxylic acid by Klebsiella sp. NP-C49 from the coral environment in Gulf of Kutch, India. Arch. Microbiol. 202, 351–359. doi: 10.1007/s00203-019-01742-9
Penton, C. R., Yang, C., Wu, L., Wang, Q., Zhang, J., Liu, F., et al. (2016). NifH-harboring bacterial community composition across an Alaskan permafrost thaw gradient. Front. Microbiol. 7:1894. doi: 10.3389/fmicb.2016.01894
Pereg, L., Morugán-Coronado, A., McMillan, M., and García-Orenes, F. (2018). Restoration of nitrogen cycling community in grapevine soil by a decade of organic fertilization. Soil Tillage Res. 179, 11–19. doi: 10.1016/j.still.2018.01.007
Perez, P. P., Ye, J., Wang, S., Wang, X. L., and Huang, D. F. (2014). Analysis of the occurrence and activity of diazotrophic communities in organic and conventional horticultural soils. Appl. Soil Ecol. 79, 37–48. doi: 10.1016/j.apsoil.2014.03.006
Poly, F., Monrozier, L. J., and Bally, R. (2001). Improvement in the RFLP procedure for studying the diversity of nifH genes in communities of nitrogen fixers in soil. Res. Microbiol. 152, 95–103. doi: 10.1016/s0923-2508(00)01172-4
R Development Core Team. (2012). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Austria ISBN 3-900051-07-0.
Ravi, A., Theresa, M., Nandayipurath, V. V. T., Rajan, S., Khalid, N. K., Thankappanpillai, A. C., et al. (2020). Plant beneficial features and application of Paraburkholderia sp. NhPBG1 isolated from pitcher of Nepenthes hamblack. Probiotics Anti. Proteins 33, 32–39. doi: 10.1007/s12602-020-09665-4
Reardon, C. L., Gollany, H. T., and Wuest, S. B. (2014). Diazotroph community structure and abundance in wheat-fallow and wheat-pea crop rotations. Soil Biol. Biochem. 69, 406–412. doi: 10.1016/j.soilbio.2013.10.038
Romero, I. C., Jacobson, M., Fuhrman, J. A., Fogel, M., and Capone, D. G. (2012). Long-term nitrogen and phosphorus fertilization effects on N2 fixation rates and nifH gene community patterns in mangrove sediments. Marine Ecol. 33, 117–127. doi: 10.1111/j.1439-0485.2011.00465.x
Rosenblueth, M., Ormeño-Orrillo, E., López-López, A., Rogel, M. A., Reyes-Hernández, B. J., Martínez-Romero, J. C., et al. (2018). Nitrogen fixation in cereals. Front. Microbiol. 9:1794. doi: 10.3389/fmicb.2018.01794
Rösch, C., Mergel, A., and Bothe, H. (2002). Biodiversity of denitrifying and dinitrogen-fixing bacteria in an acid forest soil. Appl. Environ. Microbiol. 68, 3818–3829. doi: 10.1128/aem.68.8.3818-3829.2002
Sanaullah, M., Usman, M., Wakeel, A., Cheema, S. A., Ashraf, I., and Farooq, M. (2020). Terrestrial ecosystem functioning affected by agricultural management systems: a review. Soil Tillage Res. 196:104464. doi: 10.1016/j.still.2019.104464
Segata, N., Izard, J., Waldron, L., Gevers, D., Miropolsky, L., Garrett, W. S., et al. (2011). Metagenomic biomarker discovery and explanation. Geno. Biol. 12:R60.
Sim, M. S., Ono, S., and Bosak, T. (2012). Effects of iron and nitrogen limitation on sulfur isotope fractionation during microbial sulfate reduction. Appl. Environ. Microbiol. 78, 8368–8376. doi: 10.1128/aem.01842-12
Sun, R. B., Guo, X. S., Wang, D. Z., and Chu, H. Y. (2015). Effects of long-term application of chemical and organic fertilizers on the abundance of microbial communities involved in the nitrogen cycle. Appl. Soil Ecol. 95, 171–178. doi: 10.1016/j.apsoil.2015.06.010
Tang, H., Yu, M., Wang, Y. Y., Han, X. W., Wang, X. G., Jin, W., et al. (2012). Effects of long-term fertilization on nifH gene diversity in agricultural black soil. Afr. J. Microbiol. Res. 6, 2659–2666.
Tang, Y. F., Zhang, M. M., Chen, A. L., Zhang, W. Z., Wei, W. X., and Sheng, R. (2017). Impact of fertilization regimes on diazotroph community compositions and N2-fixation activity in paddy soil. Agric. Ecosystems Environ. 247, 1–8. doi: 10.1016/j.agee.2017.06.009
Wakelin, S. A., Colloff, M. J., Harvey, P. R., Marschner, P., Gregg, A. L., and Rogers, S. L. (2007). The effects of stubble retention and nitrogen application on soil microbial community structure and functional gene abundance under irrigated maize. FEMS Microbiol. Ecol. 59, 661–670. doi: 10.1111/j.1574-6941.2006.00235.x
Wang, C., Zheng, M. M., Song, W. F., Wen, S. L., Wang, B. R., Zhu, C. Q., et al. (2017). Impact of 25 years of inorganic fertilization on diazotrophic abundance and community structure in an acidic soil in southern China. Soil Biol. Biochem. 113, 240–249. doi: 10.1016/j.soilbio.2017.06.019
Wang, J., Zhu, B., Zhang, J. B., Müller, C., and Cai, Z. C. (2015). Mechanisms of soil N dynamics following long-term application of organic fertilizers to subtropical rain-fed purple soil in China. Soil Biol. Biochem. 91, 222–231. doi: 10.1016/j.soilbio.2015.08.039
Wang, J. C., Zhang, D., Zhang, L., Li, J., Raza, W., Huang, Q. W., et al. (2016). Temporal variation of diazotrophic community abundance and structure in surface and subsoil under four fertilization regimes during a wheat growing season. Agric. Ecosystems Environ. 216, 116–124. doi: 10.1016/j.agee.2015.09.039
Wang, S., Gonzalez, P. P., Ye, J., and Huang, D. F. (2012). Abundance and diversity of nitrogen-fixing bacteria in rhizosphere and bulk paddy soil under different duration of organic management. World J. Microbiol. Biotechnol. 28, 493–503. doi: 10.1007/s11274-011-0840-1
Wang, Y. S., Li, C. N., Kou, Y. P., Wang, J. J., Tu, B., Li, H., et al. (2017). Soil pH is a major driver of soil diazotrophic community assembly in Qinghai-Tibet alpine meadows. Soil Biol. Biochem. 115, 547–555. doi: 10.1016/j.soilbio.2017.09.024
Wu, X. T., Sun, Y., Deng, L. T., Meng, Q. X., Jiang, X., Bello, A., et al. (2020). Insight to key diazotrophic community during composting of dairy manure with biochar and its role in nitrogen transformation. Waste Mange. 105, 190–197. doi: 10.1016/j.wasman.2020.02.010
Yang, L., Bai, J. S., Zeng, N. H., Zhou, X., Liao, Y. L., Lu, Y. H., et al. (2019). Diazotroph abundance and community structure are reshaped by straw return and mineral fertilizer in rice-rice-green manure rotation. Appl. Soil Ecol. 136, 11–20. doi: 10.1016/j.apsoil.2018.12.015
Yang, L., Rees, R. M., Nie, J., Xu, C. X., and Cao, W. D. (2021). Carbon substrates exert a stronger role than mineral nitrogen application in structuring soil diazotroph communities during Chinese milk vetch growth. Appl. Soil Ecol. 158:103778. doi: 10.1016/j.apsoil.2020.103778
Zehr, J. P., Jenkins, B. D., Short, S. M., and Steward, G. F. (2003). Nitrogenase gene diversity and microbial community structure: a cross-system comparison. Environ. Microbiol. 5, 539–554. doi: 10.1046/j.1462-2920.2003.00451.x
Zhalnina, K., Dias, R., de Quadros, P. D., Davis-Richardson, A., Camargo, F. A. O., Clark, I. M., et al. (2015). Soil pH determines microbial diversity and composition in the park grass experiment. Microbial Ecol. 69, 395–406. doi: 10.1007/s00248-014-0530-2
Zhao, Y., Zhang, J. B., Müller, C., and Cai, Z. C. (2018). Temporal variations of crop residue effects on soil N transformation depend on soil properties as well as residue qualities. Biol. Fertility Soils 54, 659–669. doi: 10.1007/s00374-018-1291-8
Zhong, S. Q., Han, Z., Du, J., Ci, E., Ni, J. P., Xie, D. T., et al. (2019). Relationships between the lithology of purple rocks and the pedogenesis of purple soils in the Sichuan Basin, China. Sci. Rep. 9:13272.
Keywords: straw mulching, nitrogen fertilization, diazotroph community, nifH gene, abundance
Citation: Chen S, Xiang X, Ma H, Penttinen P, Zhao J, Li H, Gao R, Zheng T and Fan G (2021) Straw Mulching and Nitrogen Fertilization Affect Diazotroph Communities in Wheat Rhizosphere. Front. Microbiol. 12:658668. doi: 10.3389/fmicb.2021.658668
Received: 26 January 2021; Accepted: 09 April 2021;
Published: 21 May 2021.
Edited by:
Markus Puschenreiter, University of Natural Resources and Life Sciences, AustriaReviewed by:
Pia H. Moisander, University of Massachusetts Dartmouth, United StatesCopyright © 2021 Chen, Xiang, Ma, Penttinen, Zhao, Li, Gao, Zheng and Fan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gaoqiong Fan, ZmFuZ2FvMjAwNTZAMTI2LmNvbQ==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.