 Liliana Medina-Aparicio1
Liliana Medina-Aparicio1 Sarahí Rodriguez-Gutierrez1
Sarahí Rodriguez-Gutierrez1 Javier E. Rebollar-Flores1Ángel G. Martínez-Batallar2
Javier E. Rebollar-Flores1Ángel G. Martínez-Batallar2 Blanca D. Mendoza-Mejía1
Blanca D. Mendoza-Mejía1 Eira D. Aguirre-Partida1
Eira D. Aguirre-Partida1 Alejandra Vázquez1
Alejandra Vázquez1 Sergio Encarnación2
Sergio Encarnación2 Edmundo Calva1
Edmundo Calva1 Ismael Hernández-Lucas1*
Ismael Hernández-Lucas1*- 1Departamento de Microbiología Molecular, Instituto de Biotecnología, Universidad Nacional Autónoma de México, Cuernavaca, Mexico
- 2Centro de Ciencias Genómicas, Universidad Nacional Autónoma de México, Cuernavaca, Mexico
The CRISPR-Cas cluster is found in many prokaryotic genomes including those of the Enterobacteriaceae family. Salmonella enterica serovar Typhi (S. Typhi) harbors a Type I-E CRISPR-Cas locus composed of cas3, cse1, cse2, cas7, cas5, cas6e, cas1, cas2, and a CRISPR1 array. In this work, it was determined that, in the absence of cas5 or cas2, the amount of the OmpC porin decreased substantially, whereas in individual cse2, cas6e, cas1, or cas3 null mutants, the OmpF porin was not observed in an electrophoretic profile of outer membrane proteins. Furthermore, the LysR-type transcriptional regulator LeuO was unable to positively regulate the expression of the quiescent OmpS2 porin, in individual S. Typhi cse2, cas5, cas6e, cas1, cas2, and cas3 mutants. Remarkably, the expression of the master porin regulator OmpR was dependent on the Cse2, Cas5, Cas6e, Cas1, Cas2, and Cas3 proteins. Therefore, the data suggest that the CRISPR-Cas system acts hierarchically on OmpR to control the synthesis of outer membrane proteins in S. Typhi.
Introduction
Microorganisms are constantly exposed to multiple viral infections and have developed many strategies to survive phage attack and invasion by foreign DNA. One such strategy is the CRISPR-Cas bacterial immunological system (Barrangou et al., 2007). This system is classified according to the presence of signature Cas proteins (Makarova et al., 2011, 2015). The hallmark of the CRISPR-Cas Type I system is the presence of the endonuclease Cas3. This protein is involved in cleavage of exogenous target nucleic acids (Sinkunas et al., 2011; Westra et al., 2012). The Type II system requires Cas9 and a trans-activating CRISPR RNA (tracrRNA) for DNA recognition and degradation (Deltcheva et al., 2011). The Type III system uses the RAMP proteins and Cas10 nuclease to silence the invader (Samai et al., 2015; Elmore et al., 2016).
In the Enterobacteriaceae family, the Type I CRISPR-Cas is the predominant system. The analysis of 228 enterobacterial genomes, corresponding to 38 genera, showed that 55% present, at least, one Type I CRISPR-Cas system (Medina-Aparicio et al., 2018). In the Salmonella genus, two CRISPR arrays (CRISPR1 and CRISPR2) have been identified, and only CRISPR1 is associated with a Type I-E set of cas genes (Touchon and Rocha, 2010). In 35 of 38 Salmonella genomes analyzed so far, the Type I-E CRISPR-Cas system was present, whereas S. enterica serovars Pullorum S06004, Javiana and Paratyphi B did not have any cas genes (Medina-Aparicio et al., 2018).
Salmonella Typhi IMSS-1 harbors a Type I-E CRISPR-Cas cluster composed of cas3, cse1-cse2-cas7-cas5-cas6e-cas1-cas2, an 84-bp leader sequence, seven 29-bp repeats and six 32-bp spacers with no homologous sequences reported in the DDBJ data bank (Medina-Aparicio et al., 2011). This locus contains five transcriptional units, two of them are the cse1-cse2-cas7-cas5-cas6e-cas1-cas2-CRISPR (cas-CRISPR operon) and scse2 (sense cse2 RNA), are transcribed from the sense strand, whereas ascse2-1 (antisense RNA of cse2 to cse1) and ascas2-1 (antisense RNA of cas2 to cas1) are present on the antisense strand (Medina-Aparicio et al., 2017). Additionally, the S. Typhi cas3 gene is transcribed as an independent unit divergent from the cas-CRISPR operon (Figure 1). The transcription of the cse1-cse2-cas7-cas5-cas6e-cas1-cas2-CRISPR polycistronic mRNA is induced by LeuO and negatively regulated by H-NS and Lrp (Hernández-Lucas et al., 2008; Medina-Aparicio et al., 2011). The role of H-NS in silencing the expression of the cas3 and ascse2-1 transcriptional units has also been demonstrated. The transcriptional activities of the five transcriptional units present in the S. Typhi CRISPR-Cas locus are induced by basic pH (Medina-Aparicio et al., 2017).
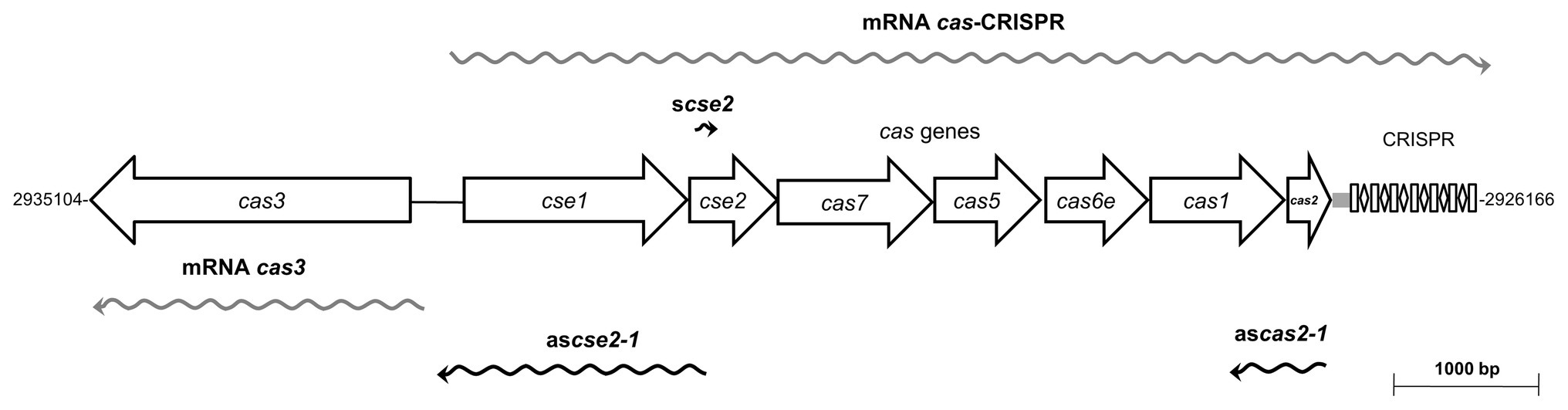
Figure 1. Transcriptional organization of the Type I-E CRISPR-Cas system of Salmonella enterica serovar Typhi IMSS-1. The CRISPR-Cas system is composed of eight cas genes (cas3 and cse1-cse2-cas7-cas5-cas6e-cas1-cas2), a leader of 84 bp and the CRISPR1 array, containing seven 29-bp repeats and six 32-bp spacers. Five transcriptional units are transcribed from this locus, wavy gray arrows represent mRNAs of cas-CRISPR operon and cas3, whereas the RNA scse2, the antisense ascse2-1 and ascas2-1 are shown as wavy black arrows. The Salmonella Typhi ΔCRISPR-cas strain is devoided of the entire Type I-E CRISPR-Cas system (from cas3 to CRISPR locus).
Relevant work on the regulation and the signals that induce the S. Typhi CRISPR-Cas system has been reported by our group (Hernández-Lucas et al., 2008; Medina-Aparicio et al., 2011, 2017). However, its biological function remains to be determined. In this regard, the low number of spacers in the CRISPR sequences, as well as their non-homology with bacteriophage and plasmid sequences, suggest that the CRISPR-Cas system does not provide an immune function in Salmonella. However, the conserved genetic organization of the cas genes in different Salmonella serovars is consistent with the system having a biological function in these bacteria (Touchon and Rocha, 2010; Shariat et al., 2015). In this work, it is reported that, in S. Typhi, CRISPR-Cas positively regulates OmpR, a two-component system regulator that induces the synthesis of the OmpC, OmpF, and OmpS2 porins. Additionally, it is demonstrated that the CRISPR-Cas system is involved in the resistance to bile salts and biofilm formation in S. Typhi.
Materials and Methods
Bacterial Strains, Plasmids, and Culture Conditions
The bacterial strains and plasmids used in this work are listed in Supplementary Table S1. Salmonella Typhi IMSS-1 (Puente et al., 1987) and Escherichia coli strains were grown aerobically at 37°C in LB (10 g tryptone, 5 g yeast extract, and 10 g NaCl per liter), MA (7 g nutrient broth, 1 g yeast extract, 2 ml glycerol, 3.75 g K2HPO4, and 1.3 g KH2PO4 per liter; Kawaji et al., 1979) or N-MM media [0.37 g KCl, 0.99 g (NH4)2SO4, 0.087 g K2SO4, 0.14 g KH2PO4, 0.019 g MgCl2, 1 g casamino acids, 5 ml glycerol, and 100 mM of Tris-HCl (pH 7.5) per liter] (Deiwick et al., 1999). When required, the following antibiotics were added: kanamycin (Km), 30 μg ml−1; tetracycline (Tc), 12 μg ml−1, and ampicillin (Ap), 200 μg ml−1.
DNA Manipulations
Plasmid and genomic DNA isolations were carried out according to published protocols (Sambrook et al., 1989). Primers for PCR amplifications were provided by the Oligonucleotide Synthesis Facility at our Institute (Supplementary Table S2). Restriction enzymes, ligase, nucleotides, and polymerases were acquired from New England Biolabs, Invitrogen, or Thermo Scientific. For sequencing, double-stranded DNA was purified with the High Pure Plasmid Isolation Kit (Roche) and sequenced with an automatic Perkin Elmer/Applied Biosystems 377-18 system.
Site-Directed Mutagenesis
The Salmonella mutants were obtained by the one-step non-polar mutagenesis procedure (Datsenko and Wanner, 2000). The target gene was replaced with selectable antibiotic resistance gene markers. The resistance cassette was removed using the pCP20 plasmid. Each mutation was further characterized by sequencing to verify the authenticity of the deletion.
Construction of Transcriptional Reporter Fusions
For transcriptional cat constructs, oligonucleotides (see Supplementary Table S2) were designed to amplify DNA fragments of different lengths from the ompC, ompF, ompS2, and ompR regulatory regions. PCR products were double-digested with BamHI-KpnI and ligated into pKK232-8 or pKK232-9 (Supplementary Table S1), which contain the promoterless cat gene. All constructs were sequenced to verify the correct DNA sequence of the PCR fragments.
CAT Assays
To determine the expression of the cat reporter gene mediated by the S. Typhi promoters, chloramphenicol acetyltransferase (CAT) assays were performed according to a previously published protocol (Martínez-Laguna et al., 1999). Briefly, S. Typhi strains harboring the reporters were grown in N-MM or MA to different optical densities (OD), and the latter medium was supplemented when required with Ap and Km, with or without IPTG (isopropyl-β-D-thiogalactopyranoside; 50 μM). Cells were harvested, centrifuged, washed with 0.8 ml of TDTT buffer (50 mM Tris-HCl, 30 μM DL-dithiothreitol, and pH 7.8), resuspended in 0.5 ml of TDTT, and sonicated on ice for 10-s intervals with 10-s rest periods until the extract was clear. The homogenate was centrifuged at 12,000 g/15 min, and the supernatant used for activity measurement. For CAT assays, 5 μl of each extract were added in duplicate to a 96-well enzyme-linked immunosorbent assay (ELISA) plate, followed by the addition of 0.2 ml of a reaction mixture containing 1 mM DTNB [5,5'-dithiobis (2-nitrobenzoic acid)], 0.1 mM acetyl-coenzyme A (acetyl-CoA), and 0.1 mM chloramphenicol in 0.1 M Tris-HCl, pH 7.8. The absorbance at 412 nm was measured every 5 s for 5 min using a Ceres 900 scanning auto reader and microplate workstation. The protein concentration of the cell extracts was obtained using the bicinchoninic acid (BCA) protein assay reagent (Pierce). Protein values and the mean rate of product formation by CAT were used to determine CAT-specific activity as micromoles per minute per milligram of protein.
Preparation of Crude Cell Extracts for Two-Dimensional Gel Electrophoresis
Salmonella Typhi IMSS-1 and S. Typhi Δcas-CRISPR harboring plasmid pFMTrcleuO-50 were grown in MA medium supplemented with Ap and IPTG (50 μM) to an optical density of 0.6 at 595 nm (OD595). Salmonella cultures (100 ml) were pelleted and washed with 1X phosphate-buffered saline (PBS). Cellular proteins were obtained by sonication at 24 kHz for 1 min in the on position and 1 min in the off position, for five cycles at 4°C using a Vibra Cell (Sonics, United States), in the presence of a protease inhibitor (Complete tablets; Roche Diagnostics GmbH, Mannheim, Germany). To further limit proteolysis, protein isolation was performed using phenol extraction (Hurkman and Tanaka, 1986). To solubilize proteins and to obtain completely denatured and reduced proteins, pellets were dried and resuspended as previously reported (Encarnación et al., 2005). Prior to electrophoresis, samples were mixed with 7 M urea, 2 M thiourea, 4% 3-[(3-choloamidopropyl)-dimethylammonio]-1-propanesulfonate (CHAPS; Roche Diagnostics GmbH, Germany), 2 mM tributylphosphine, 2% ampholytes, and 60 mM dithiothreitol.
Two-Dimensional Gel Electrophoresis
Methods used for sample preparation, analytical two-dimensional gel electrophoresis (2-DGE), image analysis, and preparative 2-DGE were described previously (Encarnación et al., 2003). pH gradients were determined using a two-dimensional sodium dodecyl sulfate-polyacrylamide gel electrophoresis standard (Sigma, United States). For isoelectric focusing, 500 μg of total proteins were loaded. All gel experiments were repeated at least two times.
In-Gel Digestion and Mass Spectrometry-Based Identification of Proteins
Selected spots from Coomassie blue-stained preparative one- or two-dimensional gels were excised manually and frozen at −70°C until use. Samples were prepared for mass spectrum analysis using a slight modification of a previously described procedure (Encarnación et al., 2005). Protein spots were destained, reduced, alkylated, and digested with trypsin (Promega, Madison, WI). Before the mass spectra of the peptide mixtures were obtained, the mixtures were desalted using a C18 Zip Tip (Millipore, Bedford, MA) according to the manufacturer’s recommendations. Mass spectra were determined using a Bruker Daltonics Autoflex (Bruker Daltonics, Billerica, MA) operated in the delayed extraction and reflectron mode. Spectra were externally calibrated using a peptide calibration standard (Bruker Daltonics 206095). Peptide mixtures were analyzed using a saturated solution of alpha-cyano-4-hydroxycinnamic acid in 50% acetonitrile-0.1% trifluoroacetic acid. Peak lists of the tryptic peptide masses were generated and searched against the NCBInr databases using the Mascot search program (Matrix Science, London, United Kingdom).1
Preparation of Outer Membrane Proteins
Outer Membrane Proteins (OMPs) were isolated from S. Typhi IMSS-1 strains grown in N-MM to an OD595 of 0.6 and 1.3 according to previous protocols (Puente et al., 1995). Fifteen milliliter of each culture was harvested and centrifuged at 5,000 g for 10 min at 4°C. Cells were resuspended in 500 μl of 10 mM Na2HPO4 buffer (pH 7.2) and sonicated on ice until the suspensions were clear. Intact cells and debris were eliminated by centrifugation (15,000 g) for 2 min, and the supernatants were transferred to clean microcentrifuge tubes and membrane fractions were pelleted by centrifugation at 12,000 g for 1 h at 4°C. Inner membrane proteins were solubilized by resuspension in 500 μl of 10 mM Na2HPO4 buffer, pH 7.2, containing 2% Triton X-100 for 30 min at 37°C. After incubation, the samples were centrifuged at 12,000 g for 1 h at 4°C. The remaining outer membrane insoluble fraction was washed with 500 μl of 10 mM Na2HPO4, pH 7.2, centrifuged at 12,000 g for 1 h at 4°C, and finally resuspended in 50 μl 1X PBS, pH 7.4. OMP concentrations were determined by BCA assay (Thermo), and 15 μg of each sample was analyzed by SDS-12% polyacrylamide gel electrophoresis. One-dimensional OMP gels were visualized by staining with Coomassie brilliant blue.
Western Blotting
For western blot experiments, S. Typhi wild-type strain and its derivatives were grown in N-MM to OD595 of 1.0 or MA medium to an OD595 of 0.6. The cultures were supplemented, when required, with Ap and IPTG (50 μM). Fifteen milliliter of each culture was harvested and centrifuged at 5,000 g for 8 min. The pellets were resuspended in 600 μl of 1X PBS and sonicated on ice for 12 min at intervals of 10-s with 5-s rest. Total protein concentration was determined by BCA assay (Thermo), and 80 μg of each sample was loaded on a 10% SDS polyacrylamide gel. Following electrophoresis, proteins were transferred to 0.45-μm-pore-size polyvinylidene difluoride membranes (Immobilon; Millipore) using the Trans-Blot SD system (Bio-Rad) according to a previously described procedure (Guadarrama et al., 2014). Membranes were blocked with 10% non-fat milk and incubated with anti-OmpR or anti-GroEL (StressGen) polyclonal antibodies. Then, they were washed with 1X PBS, 0.1% Tween 20. Immunodetection was performed with a 1:10,000 dilution of horseradish peroxidase-conjugated Anti-Rabbit antibody (Pierce) for polyclonal antibodies, and the Western Lightning Plus-ECL Chemiluminescence Reagent Kit (PerkinElmer). The membranes containing the proteins were exposed to Carestream X-OMAT LS films.
Growth Evaluation in 5% Sodium Deoxycholate
Salmonella Typhi wild-type and the different mutant strains were grown 24 h in LB plates at 37°C. A bacterial colony was inoculated in liquid LB broth (5 ml) and grown for 16 h at 37°C/200 rpm. Then, 50 ml of LB broth supplemented with 5% sodium deoxycholate (Sigma Chemical, St. Louis, MO) were inoculated with the pre-inoculum to give an initial OD at 595 nm of 0.02. The cultures were incubated at 37°C/200 rpm during 15 h with OD595 measurements being done every 2 h.
Microtiter Dish Biofilm Formation Assay
The quantification of biofilm formation was performed following a previous established protocol (O’Toole, 2011). Briefly, bacterial cells were grown overnight in LB broth (5 ml) at 37°C/200 rpm. Cells were diluted 1:100 in fresh LB without NaCl for stimulates biofilm production. One hundred microliter of this dilution was added per well in a 96-well polystyrene microtitre plate (Costar Cat. No. 3599, flat bottom with lid). Six replicate wells were prepared for each strain. Microtitre plates were incubated at 30°C for 24 h. Total bacterial growth was measured at OD600, using a GloMax®-Muti Detection System (Promega). The planktonic cells were then discarded, and the plate was washed three times with water. The remaining biofilm was fixed with 200 μl per well of methanol (100%) and stained with a 0.2% solution of crystal violet in water. After incubation at room temperature for 10 min, the plates were rinsed three times with water. The dye was solubilized by adding 125 μl of 33% acetic acid to each well and incubated the microtiter plate at room temperature for 15 min. Finally, the OD560 was determined with the microplate reader. The amount of formed biofilm is reported as the ratio of the OD560/OD600 values (Oropeza et al., 2015).
Results
CRISPR-Cas Is Fundamental for the Synthesis of Major and Quiescent Outer Membrane Proteins in Salmonella Typhi
Studies on the regulation and the signals that induce the CRISPR-cas locus in S. Typhi are available (Hernández-Lucas et al., 2008; Medina-Aparicio et al., 2011, 2017). However, its biological function in this human pathogen remains to be determined. In this regard, previous results in Francisella novicida demonstrated that the CRISPR-Cas system is involved in the synthesis of outer membrane proteins (Sampson et al., 2013, 2014). Therefore, we obtained a strain devoid of cas3, the cas3-cse1 intergenic region, cse1, cse2, cas7, cas5, cas6e, cas1, cas2, and the CRISPR locus (the entire Type I-E CRISPR-Cas system, Figure 1), which was named as ΔCRISPR-cas (Supplementary Table S1). By electrophoretic profiles, the presence of the major outer membrane proteins OmpC, OmpF, and OmpA was detected in the wild-type strain; whereas in the isogenic S. Typhi strain devoid of CRISPR-cas locus, OmpC, and OmpF were not visualized (Figure 2A). To confirm these results, the transcriptional expression of ompC and ompF promoter regions was evaluated. Thus, the reporter plasmids pKK9/ompC-772 + 27 and pKK8/ompF-782 + 184 (Supplementary Table S1) were transformed into S. Typhi IMSS-1 wild type and, in the isogenic ΔCRISPR-cas strain, to perform CAT assays. The experiments showed that the transcriptional activity of the ompC and ompF regulatory regions were of 4,328 and 5,512 CAT units, respectively, in the wild-type strain. However, in the ∆CRISPR-cas strain the ompC and ompF activity decreased by 99 and 73%, respectively (Figure 2B). These data demonstrated that the CRISPR-Cas system is relevant for the expression of the major OmpC and OmpF porins in S. Typhi.
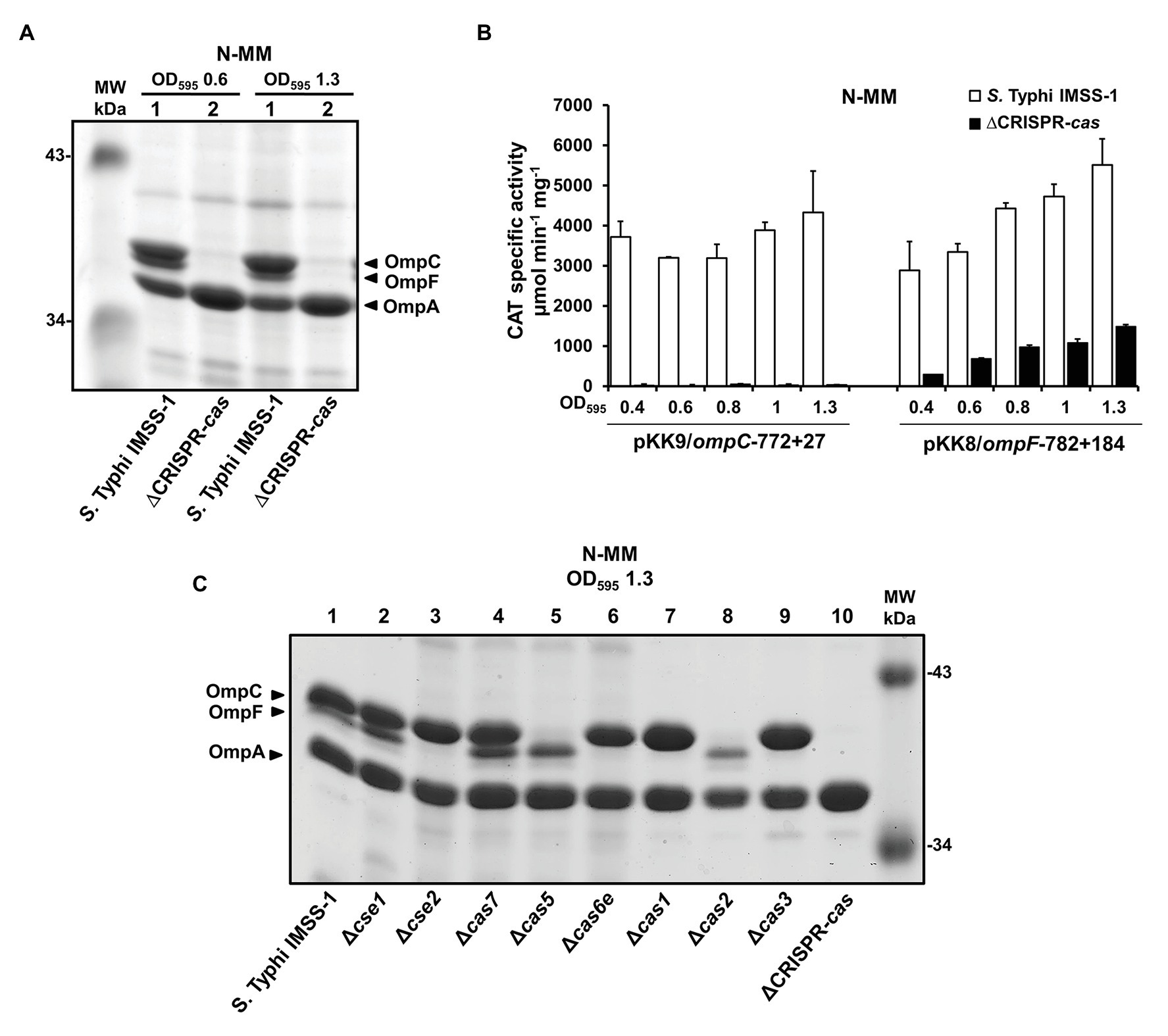
Figure 2. CRISPR-Cas is fundamental for the synthesis of the major outer membrane proteins OmpF and OmpC in Salmonella Typhi. (A) Electrophoretic pattern of Coomassie brilliant blue-stained outer membrane protein preparations, separated by 0.1% SDS-15% PAGE. The bacterial strains Salmonella Typhi IMSS-1 wild type (lane 1) and Salmonella Typhi ΔCRISPR-cas (ΔCRISPR-cas, lane 2) were grown in N-MM to an OD595 of 0.6 and 1.3. The major OMPs, OmpC, OmpF, and OmpA are indicated with a black triangle. (B) Transcriptional profiles of Salmonella Typhi IMSS-1 and Salmonella Typhi ΔCRISPR-cas harboring plasmid pKK9/ompC-772 + 27 or pKK8/ompF-782 + 184 in N-MM. CAT-specific activities were measured at an OD595 of 0.4, 0.6, 0.8, 1.0, and 1.3. The values are the means ± standard deviations for three independent experiments performed in duplicate. (C) Electrophoretic pattern of Coomassie brilliant blue-stained outer membrane protein preparations, separated by 0.1% SDS-15% PAGE from Salmonella Typhi IMSS-1 wild type (lane 1), Δcse1 (lane 2), Δcse2 (lane 3), Δcas7 (lane 4), Δcas5 (lane 5), Δcas6e (lane 6), Δcas1 (lane 7), Δcas2 (lane 8), Δcas3 (lane 9), and Salmonella Typhi ΔCRISPR-cas (ΔCRISPR-cas, lane 10) strains, grown in N-MM at OD595 of 1.3. The OmpC, OmpF, and OmpA porins are indicated with a black triangle. Molecular weight markers (MW) are indicated.
To determine the specific CRISPR-Cas genetic element involved in OmpC and OmpF regulation, a collection of individual cas mutants was generated, and porin profiles of these strains showed that Δcse1, Δcas7, and wild-type S. Typhi present a similar outer membrane protein profile. Nevertheless, in the absence of cas5 and cas2, the amount of OmpC decreased substantially; whereas in the individual cse2, cas6e, cas1 and cas3 mutants the OmpF porin was not observed (Figure 2C). These data support the fundamental role of specific Cas proteins in the regulation of OmpC and OmpF major outer membrane proteins and also are in agreement with the results obtained from the deletion of the entire CRISPR-Cas locus, since this strain lacks cas5, cas2, cse2, cas6e, cas1, and cas3, which resulted in the absence of the two main porins in S. Typhi (Figure 2A).
To continue with the identification of more CRISPR-Cas dependent outer membrane proteins, and since the overexpression of LeuO induces quiescent porins, such as OmpS2 (Fernández-Mora et al., 2004), the induction of this protein was evaluated in the absence of CRISPR-Cas. Salmonella Typhi IMSS-1 harboring plasmid pFMTrcleuO-50 and S. Typhi ∆CRISPR-cas containing pFMTrcleuO-50 were grown to an OD595 of 0.6 in MA medium supplemented with IPTG (50 μM), and 2-DGE profiles were obtained with these cultures. The results showed the presence of OmpS2 in the wild-type strain. However, in the absence of the CRISPR-cas locus, OmpS2 decreased its expression by 99% (Figure 3A). Even more, the expression of a transcriptional fusion of the 5' intergenic region of ompS2 (pKK9/ompS2-482 + 77, Supplementary Table S1), upon induction of the LeuO regulator at various points in the growth curve, was essentially abolished in the ∆CRISPR-cas as compared with the wild-type strain (Figure 3B). Therefore, CRISPR-Cas is also fundamental for OmpS2 expression mediated by LeuO.
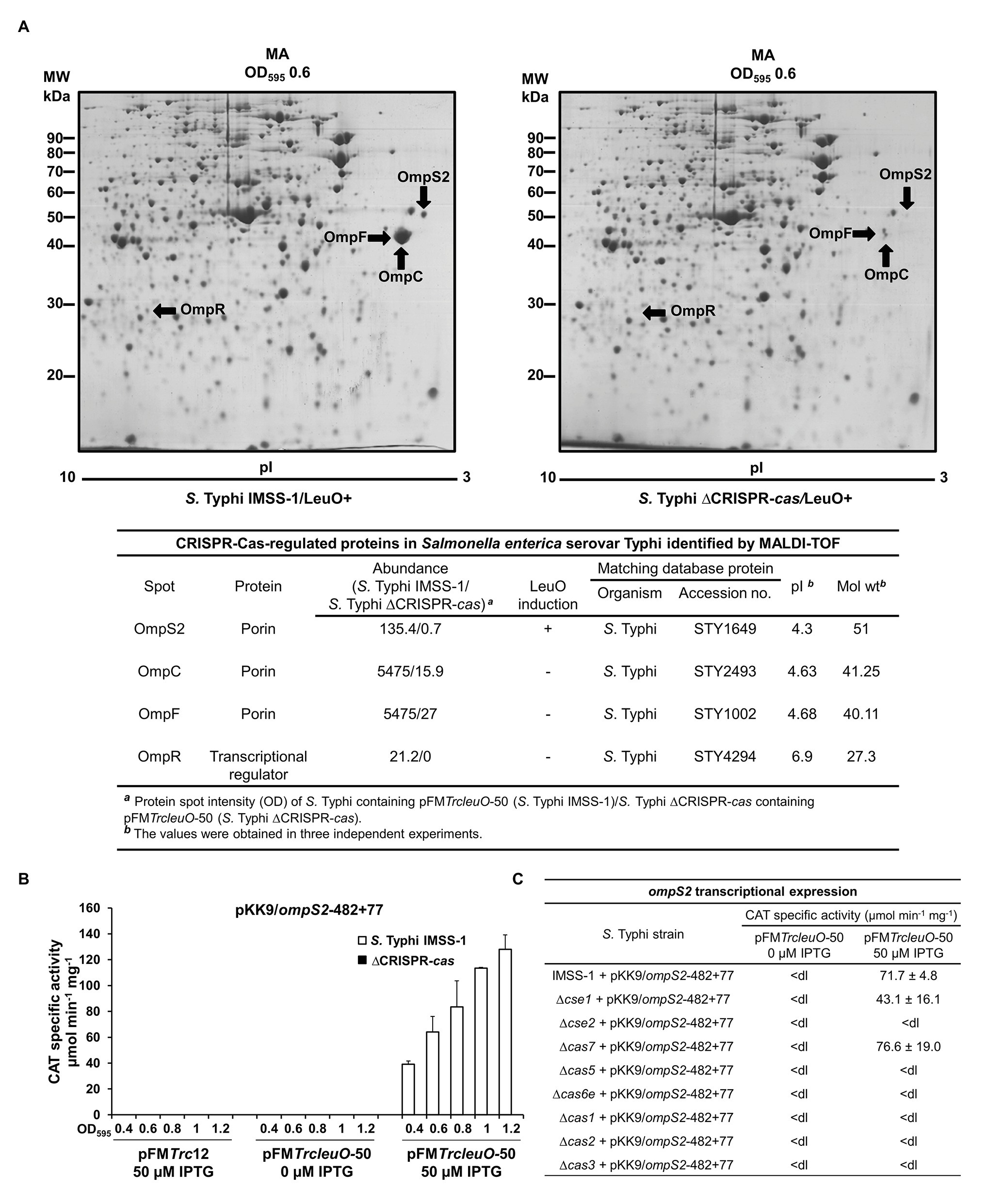
Figure 3. CRISPR-Cas is essential for the synthesis of Omps2, OmpC, and OmpF in Salmonella Typhi. (A) Proteomic profiles of Salmonella Typhi total protein extracts. Salmonella Typhi IMSS-1 wild type and Salmonella Typhi ΔCRISPR-cas containing pFMTrcleuO-50 were grown in MA to an OD595 of 0.6. Cultures were supplemented with 50 μM IPTG. The labeled spots were excised and identified using MALDI-TOF. Below the 2-DGE the CRISPR-Cas regulated proteins in Salmonella Typhi identified by MALDI-TOF are shown. (B) ompS2 transcriptional activities. Salmonella Typhi IMSS-1 (white bars) and Salmonella Typhi ΔCRISPR-cas [ΔCRISPR-cas, black bars (values below the detection limit)] harboring plasmid pFMTrcleuO-50 or pFMTrc12 were independently transformed with pKK9/ompS2-482 + 77. The strains were grown in MA medium and CAT-specific activity was measured at OD595 of 0.4, 0.6, 0.8, 1.0, and 1.2. (C) Expression profiles of Salmonella Typhi IMSS-1, Δcse1, Δcse2, Δcas7, Δcas5, Δcas6e, Δcas1, Δcas2, and Δcas3 strains containing pFMTrcleuO-50 and pKK9/ompS2-482 + 77 plasmids. The strains were grown in MA medium at OD595 of 1.0. The values are the means ± standard deviations for three independent experiments performed in duplicate; <dl (<detection limit) represents values between 0 and 10 CAT units.
To determine the specific cas genetic element involved in OmpS2 regulation, the individual cas mutants were transformed with the transcriptional CAT fusion containing the 5' regulatory region of ompS2 and plasmid pFMTrcleuO-50 for overexpressing LeuO. The expression results showed that ompS2 activity mediated by LeuO depends on cse2, cas5, cas6e, cas1, cas2, and cas3, since in the absence of each of these genetic elements ompS2 was not transcribed (Figure 3C). These results indicated that the majority of Cas proteins, with exception of Cse1 and Cas7, are essential for the synthesis of the quiescent porin OmpS2. Thus, the presence of CRISPR-Cas cluster is essential for the synthesis of major and quiescent porins in S. Typhi.
The Salmonella Typhi CRISPR-Cas System Is Involved in the Expression of the Porin Master Regulator OmpR
The results mentioned above showed that CRISPR-Cas is involved in the synthesis of outer membrane proteins in S. Typhi. Interestingly, in the 2-DGE image shown in Figure 3A, a small spot of 27.3 kDa was absent in the ∆CRISPR-cas, and the mass spectrometry (MS) results of the same spot from S. Typhi IMSS-1, demonstrated that it corresponded to OmpR. To define whether CRISPR-Cas is involved in the control of the gene for this two-component system regulator, we evaluated its transcriptional expression in the S. Typhi wild type and in a ∆CRISPR-cas. The results showed that ompR displayed 941 CAT units in the wild-type strain, and the activity decreased by 60% in the CRISPR-Cas deficient S. Typhi strain (Figure 4A). Previously, it was demonstrated that ompR contains two promoters (Villarreal et al., 2014). To define whether the ompRP1 or ompRP2 promoters are under CRISPR-Cas control, the transcriptional activity of each promoter in the wild-type strain and in the ∆CRISPR-cas was evaluated. The transcriptional results showed ompRP2 activity values of 248 and 279 CAT units in the wild type and in the ∆CRISPR-cas isogenic strain, respectively (Figure 4A). Thus, CRISPR-Cas is not involved in ompRP2 promoter control. However, the activities obtained with ompRP1 were 120 and 37 CAT units in the wild type and in the ∆CRISPR-cas, respectively (Figure 4A). Therefore, CRISPR-Cas is involved in the regulation of the ompRP1 promoter to induce ompR expression.
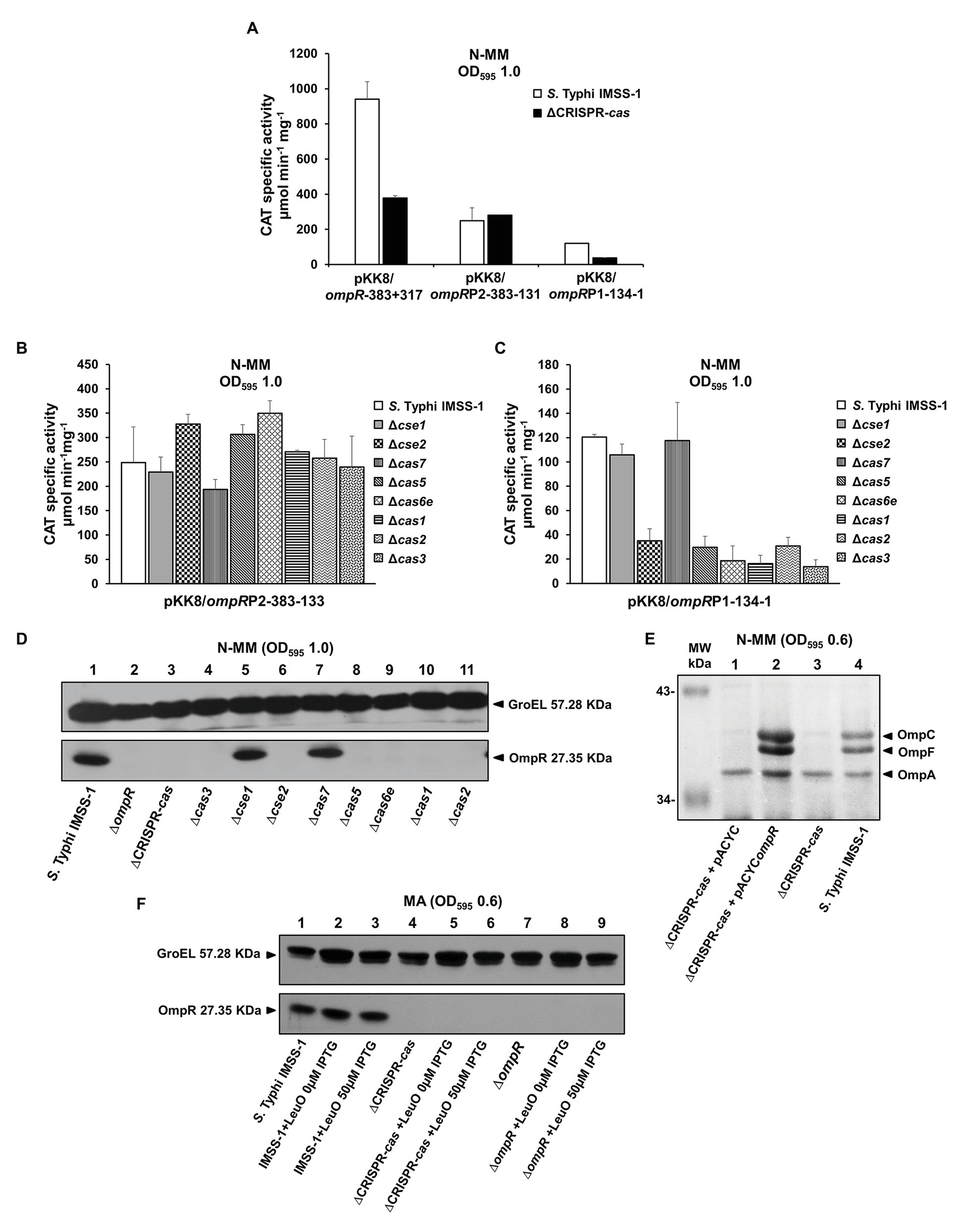
Figure 4. CRISPR-Cas is involved in the genetic control of OmpR. (A) Transcriptional profiles of Salmonella Typhi IMSS-1 and Salmonella Typhi IMSS-1 ΔCRISPR-cas (ΔCRISPR-cas) harboring plasmids pKK8/ompR-383 + 317, pKK8/ompRP2-383-133 (ompRP2) or pKK8/ompRP1-134-1 (ompRP1), grown in N-MM. CAT-specific activity was measured at an OD595 of 1.0. The values are the means ± standard deviations of three independent experiments performed in duplicate. The transcriptional expression of pKK8/ompRP2-383-133 (B) and pKK8/ompRP1-134-1 (C) was also evaluated in Salmonella Typhi IMSS-1, Δcse1, Δcse2, Δcas7, Δcas5, Δcas6e, Δcas1, Δcas2, and Δcas3 strains grown in N-MM. The samples were collected at OD595 of 1.0. The values are the means ± standard deviations for three independent experiments performed in duplicate. (D) Western blot using anti-OmpR polyclonal antibody and total proteins from Salmonella Typhi IMSS-1 wild type (lane 1), Salmonella Typhi ΔompR (ΔompR, lane 2), Salmonella Typhi ΔCRISPR-cas (ΔCRISPR-cas, lane 3), Δcas3 (lane 4), Δcse1 (lane 5), Δcse2 (lane 6), Δcas7 (lane 7), Δcas5 (lane 8), Δcas6e (lane 9), Δcas1 (lane 10), and Δcas2 (lane 11) strains were grown in N-MM at OD595 of 1.0. (E) Electrophoretic pattern of Coomassie brilliant blue-stained outer membrane protein preparations, separated by 0.1% SDS-15% PAGE of Salmonella Typhi ΔCRISPR-cas + pACYC (lane 1), Salmonella Typhi ΔCRISPR-cas + pACYCompR (lane 2), Salmonella Typhi ΔCRISPR-cas (lane 3), and Salmonella Typhi IMSS-1 wild type (lane 4), grown in N-MM to an OD595 of 0.6. The major OMPs: OmpC, OmpF, and OmpA are indicated with a black triangle. (F) Western blot performed with anti-OmpR polyclonal antibody and total proteins from Salmonella Typhi IMSS-1 (lane 1), Salmonella Typhi IMSS-1 + pFMTrcleuO-50 (0 μM IPTG; lane 2), Salmonella Typhi IMSS-1 + pFMTrcleuO-50 (50 μM IPTG; lane 3), Salmonella Typhi IMSS-1 ∆CRISPR-cas (lane 4), Salmonella Typhi IMSS-1 ∆CRISPR-cas + pFMTrcleuO-50 (0 μM IPTG; lane 5), Salmonella Typhi IMSS-1 ∆CRISPR-cas + pFMTrcleuO-50 (50 μM IPTG; lane 6), Salmonella Typhi IMSS-1 ∆ompR (lane 7), Salmonella Typhi IMSS-1 ∆ompR + pFMTrcleuO-50 (0 μM IPTG; lane 8), Salmonella Typhi IMSS-1 ∆ompR + pFMTrcleuO-50 (50 μM IPTG, lane 9). All the Salmonella Typhi bacterial strains were grown in MA to an OD595 of 0.6. GroEL was used as protein loading control. The proteins visualized are indicated with black triangles.
To validate the results obtained and to determine the Cas proteins involved in ompRP1 genetic control, individual cas mutants were transformed with CAT fusions containing either the ompRP2 (pKK8/ompRP2-383-133) or the ompRP1 (pKK8/ompRP1-134-1) promoters. The activity results showed that in the S. Typhi wild type as well as in individual cse1, cse2, cas7, cas5, cas6e, cas1, cas2, and cas3 null mutants, the ompRP2 promoter expression was similar, supporting the notion that the Cas proteins are not implicated in its regulation (Figure 4B). In the case of the ompRP1 promoter, its genetic activity in the individual cse2, cas5, cas6e, cas1, cas2, and cas3 deficient strains was considerably reduced, compared with the CAT units obtained in the wild-type strain and in the cse1 and cas7 mutants (Figure 4C). The data support the proposal that the Cas proteins involved in ompRP1 promoter regulation correspond to Cse2, Cas5, Cas6e, Cas1, Cas2, and Cas3; whereas Cse1 and Cas7 are not implicated in ompRP1 induction.
To determine whether the reduction of ompRP1 promoter activity in the ∆CRISPR-cas, as well as in each cas individual mutant, has an effect on the synthesis of OmpR, western blot experiments were performed. The wild-type S. Typhi, the ∆CRISPR-cas, as well as the individual cas3, cse1, cse2, cas7, cas5, cas6e, cas1, and cas2 deletion mutants were grown in N-MM to an OD595 of 1.0. Total crude cell protein extracts were transferred to membranes and probed using anti-OmpR polyclonal antibody. The western blot results showed a prominent OmpR band of 27.3 KDa in the wild-type strain and in the cse1 and cas7 individual mutants; whereas in the ∆CRISPR-cas strain, and in the individual cse2, cas5, cas6e, cas1, cas2, and cas3 mutants the OmpR protein was absent (Figure 4D).
These results explain the lack of OmpC and OmpF in the corresponding cas deficient strains (Figure 2C), since it is well-known that OmpR binds to their regulatory regions to promote their expression (Yoshida et al., 2006). Therefore, cse2, cas5, cas6e, cas1, cas2, and cas3 genes are fundamental for OmpR expression, whereas cse1 and cas7 are not involved in OmpR regulation, demonstrating that specific cas genes are necessary for OmpR production to control porin synthesis. Moreover, complementation of the S. Typhi ΔCRISPR-cas with the ompR gene on a plasmid restored the presence of OmpC and OmpF porins in this strain (Figure 4E), further supporting the notion that the deletion of the entire CRISPR-cas loci results in the lowering of the expression of the OmpR regulator and thus porin expression.
With respect to the OmpS2 quiescent porin, it is well accepted that LeuO counteracts the negative effect of H-NS on the ompS2 promoter, upon which OmpR binds to its regulatory region promoting ompS2 expression (Fernández-Mora et al., 2004). Thus, it was determined whether the OmpR protein was produced in the S. Typhi strains that overexpress LeuO. Western blot experiments demonstrated the presence of OmpR in S. Typhi IMSS-1 wild-type strain harboring the pFMTrcleuO-50 plasmid. However, OmpR was not detected in the S. Typhi ∆cas-CRISPR mutant overexpressing LeuO (Figure 4F). Thus, OmpS2 was not visualized in the 2-DGE of this strain (Figure 3A) because of the lack of the two-component system regulator OmpR.
In conclusion, the results obtained here showed that the CRISPR-Cas system acts hierarchically on the ompRP1 promoter to induce OmpC, OmpF, or OmpS2 synthesis in S. Typhi.
The Salmonella Typhi cas Genes Are Involved in Sodium Deoxycholate Resistance and Biofilm Formation
In this report, we have shown that the S. Typhi cse2, cas5, cas6e, cas1, cas2, and cas3 genes are involved in porin synthesis through the regulation of the ompR gene which codes for the OmpR transcriptional regulator. In previous studies, it has been demonstrated that ompR is involved in virulence, sodium deoxycholate resistance, biofilm formation, the production of flagella, and curli (Pickard et al., 1994; Shin and Park, 1995; Vidal et al., 1998; Cameron and Dorman, 2012; Villarreal et al., 2014). Therefore, we evaluated whether the cas genes are involved in some of these biological processes. Growth rate experiments of S. Typhi IMSS-1 and the cas individual deleted strains were performed in LB broth supplemented with 5% of the human bile salt sodium deoxycholate. The results showed that the wild-type S. Typhi strain grew in this condition, reaching an OD595 of 0.86 after 15 h. However, growth of the ∆cas5, ∆cas2, and ∆cas-CRISPR mutant strains was impaired in the presence of this bile salt (Figure 5A) since their OD595 were of 0.43, 0.54, and 0.18, respectively, after 15 h of incubation. Remarkably, these strains did not express the OmpC porin, which was previously shown to be determinant for allowing S. Typhi to proliferate in the presence of sodium deoxycholate (Villarreal et al., 2014). The growth rate of ∆cas6e, ∆cas1, and ∆cas3 was similar to that observed with the wild-type strain (Figure 5A), consistent with the presence of the OmpC porin in these mutants. The same experiment was performed with ∆ompR, ∆ompC, ∆ompF, and ∆ompS2 strains. As expected, the ompR and ompC mutants were also impaired in their growth in 5% sodium deoxycholate (OD595 = 0.5 and 0.33, respectively; Figure 5B), as previously reported (Villarreal et al., 2014); whereas the ∆ompF and ∆ompS2 mutant strains grew like the S. Typhi IMSS-1 wild type.
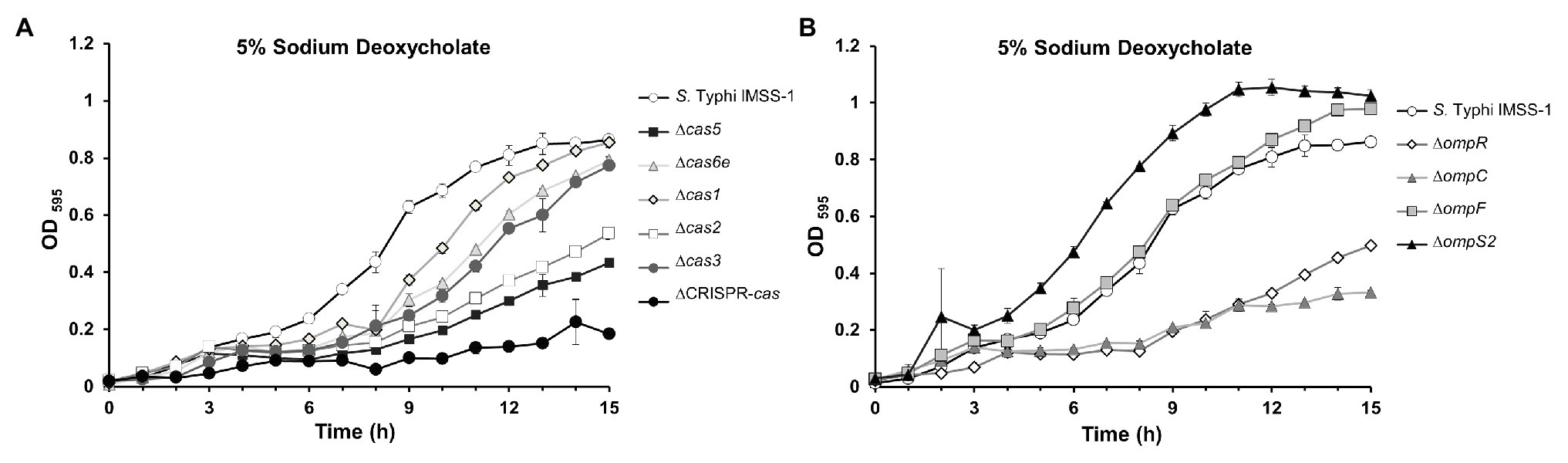
Figure 5. The CRISPR-Cas system is required for resistance to the bile salt sodium deoxycholate in Salmonella Typhi. (A) Growth kinetics in sodium deoxycholate of Salmonella Typhi IMSS-1 wild type (empty circle), Δcas5 (black square), Δcas6e (gray triangle), Δcas1 (gray diamond), Δcas2 (empty square), Δcas3 (gray circle), and ΔCRISPR-cas (black circle). (B) Growth kinetics in sodium deoxycholate of Salmonella Typhi IMSS-1 wild type (empty circle), ΔompR (empty diamond), ΔompC (gray triangle), ΔompF (gray square), and ΔompS2 (black triangle). For both kinetics, the bacterial strains were grown in LB broth supplemented with 5% sodium deoxycholate at 37°C. The growth was monitored by OD595. Three independent experiments were performed in duplicate and representative data are shown.
Additionally, we also evaluated the biofilm formation ability of the S. Typhi IMSS-1 wild type, and of the ∆cse2, ∆cas5, ∆cas6e, ∆cas1, ∆cas2, ∆cas3, ∆cas-CRISPR, ∆ompR, ∆ompC, ∆ompF, and ∆ompS2 mutant strains. The experiments showed that the wild type produced moderate biofilm (0.26 OD560/OD600 ratio). However, the cse2, cas5, cas6e, cas1, cas2, cas3, and cas-CRISPR mutants displayed an increased biofilm formation (OD560/OD600 ratio of 1.06, 1.04, 0.95, 1.04, 1.17, 0.90, and 0.50, respectively; Figure 6). These results suggest that the CRISPR-Cas system negatively regulates genes involved in biofilm production, i.e., that the absence of cas genes allows the expression of factors that increase the ability of S. Typhi to form biofilm.
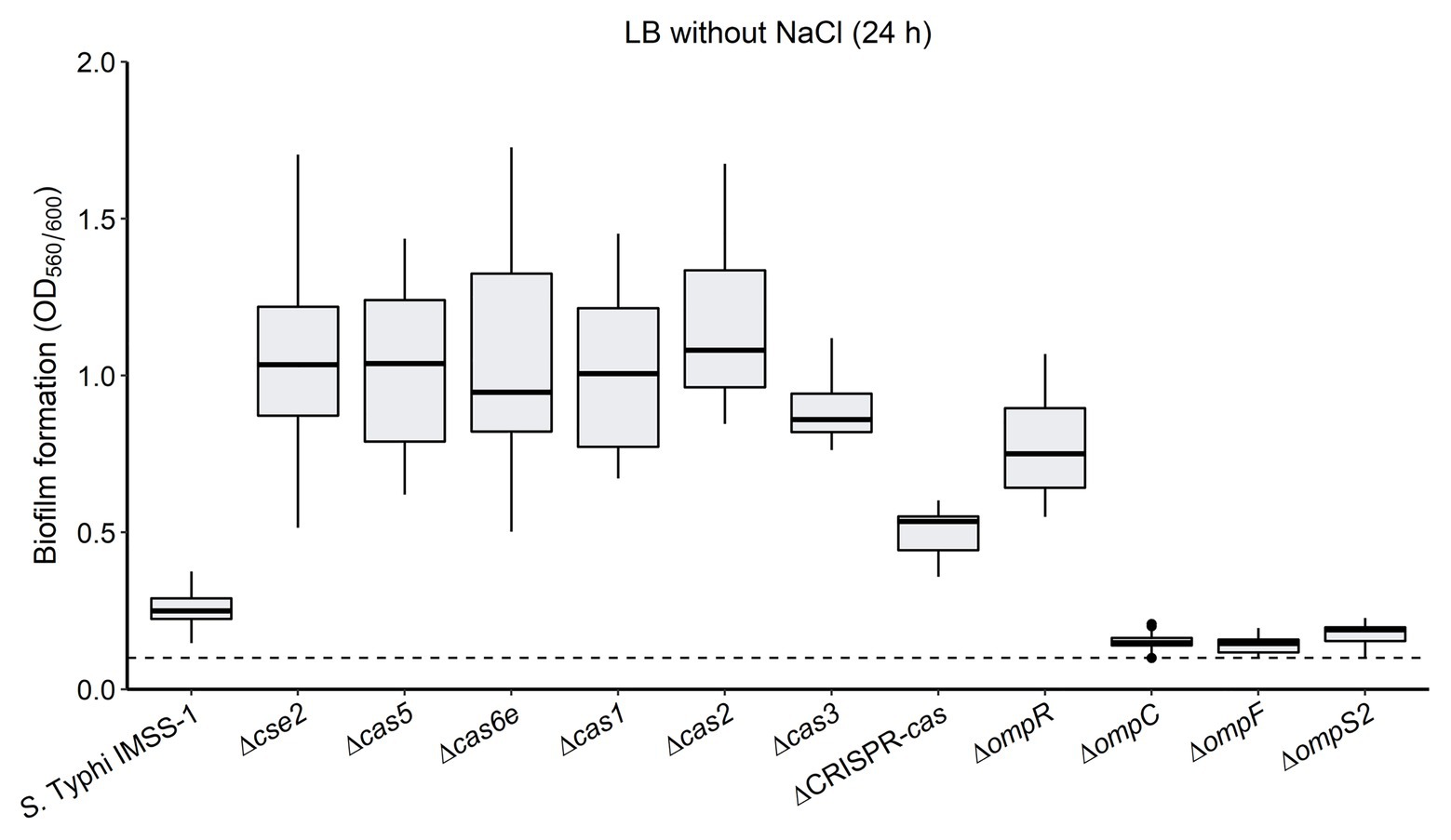
Figure 6. The absence of the CRISPR-Cas system increases biofilm formation in Salmonella Typhi. The Salmonella Typhi IMSS-1 wild type, and Δcse2, Δcas5, Δcas6e, Δcas1, Δcas2, Δcas3, ΔCRISPR-cas, ΔompR, ΔompC, ΔompF, and ΔompS2 mutant strains were grown in LB broth without NaCl and incubated at 30°C for 24 h to evaluate biofilm formation by crystal violet staining in microtitre dishes. The dotted line indicates a 0.1 background value and a bacterial strain produces biofilm when the values obtained are above 0.1. The box plot diagram represents the data obtained from three independent experiments.
The ∆ompR mutant showed an increased biofilm formation (0.77), as compared to the values obtained with the wild type (0.26; Figure 6). However, strains carrying deletions either in the ompC, ompF, or the ompS2 genes presented a slightly decreased biofilm formation (OD560/OD600 ratio of 0.15, 0.14, and 0.18, respectively; Figure 6). Therefore, the biofilm production was independent of the individual absence of the OmpC, OmpF, or OmpS2 porins. Interestingly, it has been demonstrated that ompR mutants in Salmonella enteritidis, Salmonella pullorum, E. coli, and Yersinia enterocolitica presented a decreased biofilm formation ability (Dong et al., 2011; Lu et al., 2012; Samanta et al., 2013; Meng et al., 2019), suggesting that the pathway toward regulating biofilm synthesis is different in S. Typhi.
The data shown are consistent with the notion that the CRISPR-Cas system is relevant for S. Typhi virulence, since this pathogen needs to survive the presence of bile salts in the gut and gallbladder, as well as to persist inside the gallbladder, where the biofilm formation is relevant (Crawford et al., 2010; Gonzalez-Escobedo et al., 2011; Spector and Kenyon, 2012).
Discussion
The results presented here showed that the Cse2, Cas5, Cas6e, Cas1, Cas2, and Cas3 proteins via the positive regulation of the two-component regulator OmpR, have a role in the major and quiescent outer membrane protein synthesis, since they control OmpC, OmpF, and OmpS2. Due to the fact that only a few transcriptional factors have been implicated in the control of ompR in Salmonella, such as LtrR, H-NS, and OmpR (autoregulation; Bang et al., 2002; Villarreal et al., 2014), the data obtained contribute to the understanding of the regulatory network that controls the activity of this master regulator.
The results also support the complex genetic regulation of porins (De la Cruz and Calva, 2010), since in the absence of cas5 and cas2, OmpR becomes undetectable (Figure 4D), as does OmpC (Figure 2C), demonstrating the specific role of these cas genes on ompR regulation to mediate OmpC synthesis. Interestingly, the presence of OmpF was evident in these cas mutants, supporting the notion that OmpF is not only OmpR-dependent, and that other transcriptional factors are able to induce OmpF expression. In this sense, regulators, such as Lrp and CadC, are also involved in its positive control (Ferrario et al., 1995; Lee et al., 2007; De la Cruz and Calva, 2010). In contrast, in the individual cse2, cas6e, cas1, and cas3 mutants the OmpF porin was not visualized (Figure 2C), and OmpR was not detected by western blot (Figure 4D), supporting the role of these genes in the control of ompR to promote OmpF synthesis. In these cas mutants, the presence of OmpC was observed, supporting the proposal that other regulators are able to induce OmpC synthesis. In this respect, the CpxRA and CadC transcriptional factors have been reported to positively regulate ompC (Batchelor et al., 2005; Lee et al., 2007; De la Cruz and Calva, 2010).
In E. coli, it is well-known that ompR-envZ comprises an operon, and a bioinformatic analysis using the Operon-mapper tool suggested that, in S. Typhi, these genes can be also one transcriptional unit (data not shown; Taboada et al., 2018). Therefore, the absence of OmpR in the cas mutants indirectly suggests that EnvZ is not produced by the polar effect of the ompR deletion. However, OmpR is the principal component involved in porin synthesis since the presence of the corresponding porins was reestablished in the CRISPR-cas deleted strain overexpessing OmpR (Figure 4E).
In another report, it has also been shown that a Cas protein, Cas9, negatively regulates the gene coding for a transcriptional regulator of a two-component system: regR. In that case, it was demonstrated that Cas9 was able to degrade the regR mRNA, since the Streptococcus agalactiae CRISPR array contains two homologous sequences to the regR gene (Ma et al., 2018).
Salmonella Typhi contains a Type I-E CRISPR-Cas locus, and in vitro experiments have demonstrated that E. coli components of this genetic system are able to form a complex for recognition and degradation of viral and plasmid DNA (Brouns et al., 2008; Jore et al., 2011). The data obtained in this work showed that two Cas proteins, Cas5 and Cas2, are fundamental for OmpC expression, and other four Cas proteins, Cse2, Cas6e, Cas1, and Cas3, are required for OmpF synthesis. In the case of OmpS2 expression, six Cas proteins are relevant: Cse2, Cas5, Cas6e, Cas1, Cas2, and Cas3. Therefore, it is possible that different combinations of Cas form distinct protein complexes that bind, stabilize, and positively modulate the levels of ompR mRNA, for differentially regulating OmpC, OmpF, or OmpS2. Another possibility for OmpR regulation is that Cse2, Cas6e, Cas1, and Cas2 RNA-nucleases cleave the mRNA of a putative ompR repressor. Thus, when such negative regulator would be degraded, the ompR gene would be able to be expressed for porin synthesis. It is also possible that the function of Cas complexes would be only to bind at DNA to fine-tune ompR expression at specific promoters.
Currently, experiments are being performed in our laboratory to evaluate these hypotheses and to extend these initial observations in order to define how CRISPR-Cas mediate OmpR control. It is evident that much needs to be learned about the mechanisms by which various genetic elements control the expression of the OmpR regulator and thus, the porin phenotype in S. Typhi.
The finding that Cas proteins are able to regulate hierarchically the global two-component regulatory systems present in different proteobacteria, suggesting that the CRISPR-Cas systems could be involved in the regulation of biological processes controlled by two-component regulators, including oxidative stress, low pH, heat shock, bacterial motility, chemotaxis, osmotic changes, resistance to bile salts, and biofilm formation (Groisman, 2016; Pruss, 2017). In this sense, OmpR regulates the expression of hilC, hilD, and ssrAB, the main regulators of pathogenicity islands 1 and 2 of Salmonella Typhimurium, and it also controls the expression of the viaB locus that encodes Vi polysaccharide biosynthesis genes in S. Typhi (Pickard et al., 1994; Lee et al., 2000; Feng et al., 2003; Cameron and Dorman, 2012). Therefore, OmpR is implicated in regulation of virulence.
In the case of the OmpC and OmpF porins, a double mutant of these genes in S. Typhimurium was found to be attenuated for virulence in the mouse model (Chatfield et al., 1991). In addition, it has been observed that OmpC and OmpF induced long-term antibody response with bactericidal capacity and conferred protection against challenge with S. Typhi (Secundino et al., 2006; Pérez-Toledo et al., 2017). Moreover, it has been demonstrated that the immunization of mice with the OmpS2 protein induced the production of specific, long-term antibody titers and conferred protection against S. Typhi challenge. In addition, OmpS2 is a TLR2 and TLR4 agonist. Thus, OmpS2, despite being expressed at low levels under in vitro culture conditions, is a potent protective immunogen with intrinsic adjuvant properties (Moreno-Eutimio et al., 2013). Salmonella Typhimurium mutants with deletions in the ompS2 gene were highly attenuated for virulence in a mouse model, supporting its role in pathogenesis (Rodríguez-Morales et al., 2006).
Thus, a phenotype for the mutants in the genes coding for the S. Typhi Cas was explored. It was found that the cas5 and cas2 genes are necessary for the optimal growth of S. Typhi in the presence of one of the major bile salts found in the human gut, sodium deoxycholate (Figure 5). Most remarkably, the ∆cas5 and ∆cas2 mutant strains lack the OmpC porin (Figure 2C), which was previously shown to be necessary for growth in the presence of this bile salt (Villarreal et al., 2014).
Additionally, the CRISPR-Cas system is implicated in the control of biofilm formation in S. Typhi, since the absence of cse2, cas5, cas6e, cas1, cas2, and cas3 genes resulted in an increase in the biosynthesis of biofilm (Figure 6). Interestingly, the CRISPR-Cas system has also been involved in biofilm formation in Pseudomonas aeruginosa (Zegans et al., 2009). These newfound roles of the S. Typhi CRISPR-Cas system in the resistance to sodium deoxycholate and biofilm production should contribute toward the understanding of the evolutionary conservation of this system in the Salmonella genus, since these biological processes are relevant for the establishment of a successful infection cycle (Gonzalez-Escobedo et al., 2011; Spector and Kenyon, 2012).
Contributions from several other research groups also support the CRISPR-Cas-outer membrane protein association. By gene neighborhood analysis, it has been found that numerous candidate CRISPR-linked genes encode integral membrane proteins in bacterial and archaeal genomes (Shmakov et al., 2018). Furthermore, activation of the CRISPR-Cas system by envelope stress has been suggested in E. coli (Perez-Rodriguez et al., 2011), and a role in regulating the permeability of the bacterial envelope to resist membrane damage caused by antibiotics is suggested for CRISPR-Cas in Francisella novicida (Sampson et al., 2014). In Myxococcus xanthus, the CRISPR-Cas system appears to be involved in fruiting body development and exopolysaccharide production (Viswanathan et al., 2007; Wallace et al., 2014), Moreover, recent microarray experiments performed in our laboratory demonstrated that CRISPR-Cas is able to regulate other outer membrane encoded genes besides ompC, ompF, and ompS2 (data not shown).
Collectively, these data, together with our results suggest a previously unappreciated role for CRISPR-Cas in the formation of bacterial structures and in the maintenance of the cell envelope in different prokaryotic organisms.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
LM-A: methodology, formal analysis, investigation, writing-review, and editing. SR-G: methodology, formal analysis, and validation. JR-F: methodology and validation. AM-B, BM-M, EA-P, and AV: methodology. SE: methodology and resources. EC: writing-review and editing. IH-L: conceptualization, resources, writing-original draft preparation, visualization, supervision, project administration, and funding. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by grants from the Dirección General de Asuntos del Personal Académico, DGAPA/UNAM (IN203618 and IN203621 to IH-L). SR-G was supported by a fellowship from CONACYT, Mexico (No. 887111).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank A. A. Beltrán-Luviano, M. Fernández-Mora, F. J. Santana, L. Perezgasga, I. Serrano-Fujarte, A. Tapia-Rodríguez, P. Gaytan, E. Bustos, S. Becerra, and J. Yañez from IBT-UNAM and S. Dávila from UAEM for their technical help. We also thank S. Mayo-Pérez, I. Olivar-Casique from IBT-UNAM, J. Miranda (IIBO-UNAM), and M. Dunn (CCG-UNAM) for stimulating discussions and critical reading.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2021.657404/full#supplementary-material
Footnotes
References
Bang, I. S., Audia, J. P., Park, Y. K., and Foster, J. W. (2002). Autoinduction of the ompR response regulator by acid shock and control of the Salmonella enterica acid tolerance response. Mol. Microbiol. 44, 1235–1250. doi: 10.1046/j.1365-2958.2002.02937.x
Barrangou, R., Fremaux, C., Deveau, H., Richards, M., Boyaval, P., Moineau, S., et al. (2007). CRISPR provides acquired resistance against viruses in prokaryotes. Science 315, 1709–1712. doi: 10.1126/science.1138140
Batchelor, E., Walthers, D., Kenney, L. J., and Goulian, M. (2005). The Escherichia coli CpxA-CpxR envelope stress response system regulates expression of the porins ompF and ompC. J. Bacteriol. 187, 5723–5731. doi: 10.1128/JB.187.16.5723-5731.2005
Brouns, S. J., Jore, M. M., Lundgren, M., Westra, E. R., Slijkhuis, R. J., Snijders, A. P., et al. (2008). Small CRISPR RNAs guide antiviral defense in prokaryotes. Science 321, 960–964. doi: 10.1126/science.1159689
Cameron, A. D., and Dorman, C. J. (2012). A fundamental regulatory mechanism operating through OmpR and DNA topology controls expression of Salmonella pathogenicity islands SPI-1 and SPI-2. PLoS Genet. 8:e1002615. doi: 10.1371/journal.pgen.1002615
Crawford, R. W., Rosales-Reyes, R., Ramirez-Aguilar, M. L., Chapa-Azuela, O., Alpuche-Aranda, C., and Gunn, J. S. (2010). Gallstones play a significant role in Salmonella spp. gallbladder colonization and carriage. Proc. Natl. Acad. Sci. U. S. A. 107, 4353–4358. doi: 10.1073/pnas.1000862107
Chatfield, S. N., Dorman, C. J., Hayward, C., and Dougan, G. (1991). Role of ompR-dependent genes in Salmonella Typhimurium virulence: mutants deficient in both ompC and ompF are attenuated in vivo. Infect. Immun. 59, 449–452. doi: 10.1128/IAI.59.1.449-452.1991
Datsenko, K. A., and Wanner, B. L. (2000). One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. U. S. A. 97, 6640–6645. doi: 10.1073/pnas.120163297
Deiwick, J., Nikolaus, T., Erdogan, S., and Hensel, M. (1999). Environmental regulation of Salmonella pathogenicity island 2 gene expression. Mol. Microbiol. 31, 1759–1773. doi: 10.1046/j.1365-2958.1999.01312.x
De la Cruz, M. A., and Calva, E. (2010). The complexities of porin genetic regulation. J. Mol. Microbiol. Biotechnol. 18, 24–36. doi: 10.1159/000274309
Deltcheva, E., Chylinski, K., Sharma, C. M., Gonzales, K., Chao, Y., Pirzada, Z. A., et al. (2011). CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III. Nature 471, 602–607. doi: 10.1038/nature09886
Dong, H., Peng, D., Jiao, X., Zhang, X., Chen, S., Lu, Y., et al. (2011). Construction and characterization of an ompR gene deletion mutant from Salmonella enteritidis. Wei Sheng Wu Xue Bao 51, 1256–1262.
Elmore, J. R., Sheppard, N. F., Ramia, N., Deighan, T., Li, H., Terns, R. M., et al. (2016). Bipartite recognition of target RNAs activates DNA cleavage by the type III-B CRISPR-Cas system. Genes Dev. 30, 447–459. doi: 10.1101/gad.272153.115
Encarnación, S., Guzmán, Y., Dunn, M. F., Hernández, M., del Carmen Vargas, M., and Mora, J. (2003). Proteome analysis of aerobic and fermentative metabolism in Rhizobium etli CE3. Proteomics 3, 1077–1085. doi: 10.1002/pmic.200300427
Encarnación, S., Hernández, M., Martínez-Batallar, G., Contreras, S., Vargas, M. C., and Mora, J. (2005). Comparative proteomics using 2-D gel electrophoresis and mass spectrometry as tools to dissect stimulons and regulons in bacteria with sequenced or partially sequenced genomes. Biol. Proced. Online 7, 117–135. doi: 10.1251/bpo110
Feng, X., Oropeza, R., and Kenney, L. J. (2003). Dual regulation by phospho-OmpR of ssrA/B gene expression in Salmonella pathogenicity island 2. Mol. Microbiol. 48, 1131–1143. doi: 10.1046/j.1365-2958.2003.03502.x
Fernández-Mora, M., Puente, J. L., and Calva, E. (2004). OmpR and LeuO positively regulate the Salmonella enterica serovar Typhi ompS2 porin gene. J. Bacteriol. 186, 2909–2920. doi: 10.1128/JB.186.10.2909-2920.2004
Ferrario, M., Ernsting, B. R., Borst, D. W., Wiese, D. E. 2nd., Blumenthal, R. M., and Matthews, R. G. (1995). The leucine-responsive regulatory protein of Escherichia coli negatively regulates transcription of ompC and micF and positively regulates translation of ompF. J. Bacteriol. 177, 103–113. doi: 10.1128/JB.177.1.103-113.1995
Gonzalez-Escobedo, G., Marshall, J. M., and Gunn, J. S. (2011). Chronic and acute infection of the gall bladder by Salmonella Typhi: understanding the carrier state. Nat. Rev. Microbiol. 9, 9–14. doi: 10.1038/nrmicro2490
Groisman, E. A. (2016). Feedback control of two-component regulatory systems. Annu. Rev. Microbiol. 70, 103–124. doi: 10.1146/annurev-micro-102215-095331
Guadarrama, C., Medrano-López, A., Oropeza, R., Hernández-Lucas, I., and Calva, E. (2014). The Salmonella enterica serovar Typhi LeuO global regulator forms tetramers: residues involved in oligomerization, DNA binding, and transcriptional regulation. J. Bacteriol. 196, 2143–2154. doi: 10.1128/JB.01484-14
Hernández-Lucas, I., Gallego-Hernández, A. L., Encarnación, S., Fernández-Mora, M., Martínez-Batallar, A. G., Salgado, H., et al. (2008). The LysR-type transcriptional regulator LeuO controls expression of several genes in Salmonella enterica serovar Typhi. J. Bacteriol. 190, 1658–1670. doi: 10.1128/JB.01649-07
Hurkman, W. J., and Tanaka, C. K. (1986). Solubilization of plant membrane proteins for analysis by two-dimensional gel electrophoresis. Plant Physiol. 81, 802–806. doi: 10.1104/pp.81.3.802
Jore, M. M., Lundgren, M., van Duijn, E., Bultema, J. B., Westra, E. R., Waghmare, S. P., et al. (2011). Structural basis for CRISPR RNA-guided DNA recognition by Cascade. Nat. Struct. Mol. Biol. 18, 529–536. doi: 10.1038/nsmb.2019
Kawaji, H., Mizuno, T., and Mizushima, S. (1979). Influence of molecular size and osmolarity of sugars and dextrans on the synthesis of outer membrane proteins O-8 and O-9 of Escherichia coli K-12. J. Bacteriol. 140, 843–847. doi: 10.1128/JB.140.3.843-847.1979
Lee, A. K., Detweiler, C. S., and Falkow, S. (2000). OmpR regulates the two-component system SsrA-ssrB in Salmonella pathogenicity island 2. J. Bacteriol. 182, 771–781. doi: 10.1128/JB.182.3.771-781.2000
Lee, Y. H., Kim, B. H., Kim, J. H., Yoon, W. S., Bang, S. H., and Park, Y. K. (2007). CadC has a global translational effect during acid adaptation in Salmonella enterica serovar Typhimurium. J. Bacteriol. 189, 2417–2425. doi: 10.1128/JB.01277-06
Lu, Y., Chen, S., Dong, H., Sun, H., Peng, D., and Liu, X. (2012). Identification of genes responsible for biofilm formation or virulence in Salmonella enterica serovar pullorum. Avian Dis. 56, 134–143. doi: 10.1637/9806-052411-Reg.1
Ma, K., Cao, Q., Luo, S., Wang, Z., Liu, G., Lu, C., et al. (2018). cas9 enhances bacterial virulence by repressing the regR transcriptional regulator in Streptococcus agalactiae. Infect. Immun. 86, e00552–e005517. doi: 10.1128/IAI.00552-17
Makarova, K. S., Haft, D. H., Barrangou, R., Brouns, S. J., Charpentier, E., Horvath, P., et al. (2011). Evolution and classification of the CRISPR-Cas systems. Nat. Rev. Microbiol. 9, 467–477. doi: 10.1038/nrmicro2577
Makarova, K. S., Wolf, Y. I., Alkhnbashi, O. S., Costa, F., Shah, S. A., Saunders, S. J., et al. (2015). An updated evolutionary classification of CRISPR-Cas systems. Nat. Rev. Microbiol. 13, 722–736. doi: 10.1038/nrmicro3569
Martínez-Laguna, Y., Calva, E., and Puente, J. L. (1999). Autoactivation and environmental regulation of bfpT expression, the gene coding for the transcriptional activator of bfpA in enteropathogenic Escherichia coli. Mol. Microbiol. 33, 153–166. doi: 10.1046/j.1365-2958.1999.01460.x
Medina-Aparicio, L., Dávila, S., Rebollar-Flores, J. E., Calva, E., and Hernández-Lucas, I. (2018). The CRISPR-Cas system in Enterobacteriaceae. Pathog. Dis. 76. doi: 10.1093/femspd/fty002
Medina-Aparicio, L., Rebollar-Flores, J. E., Beltrán-Luviano, A. A., Vázquez, A., Gutíerrez-Ríos, R. M., Olvera, L., et al. (2017). CRISPR-Cas system presents multiple transcriptional units including antisense RNAs that are expressed in minimal medium and upregulated by pH in Salmonella enterica serovar Typhi. Microbiology 163, 253–265. doi: 10.1099/mic.0.000414
Medina-Aparicio, L., Rebollar-Flores, J. E., Gallego-Hernández, A. L., Vázquez, A., Olvera, L., Gutíerrez-Ríos, R. M., et al. (2011). The CRISPR/Cas immune system is an operon regulated by LeuO, H-NS, and leucine-responsive regulatory protein in Salmonella enterica serovar Typhi. J. Bacteriol. 193, 2396–2407. doi: 10.1128/JB.01480-10
Meng, J., Bai, J., Xu, J., Huang, C., and Chen, J. (2019). Differential regulation of physiological activities by RcsB and OmpR in Yersinia enterocolitica. FEMS Microbiol. Lett. 366:fnz210. doi: 10.1093/femsle/fnz210
Moreno-Eutimio, M. A., Tenorio-Calvo, A., Pastelin-Palacios, R., Pérez-Shibayama, C., Gil-Cruz, C., López-Santiago, R., et al. (2013). Salmonella Typhi OmpS1 and OmpS2 porins are potent protective immunogens with adjuvant properties. Immunology 139, 459–471. doi: 10.1111/imm.12093
Oropeza, R., Salgado-Bravo, R., and Calva, E. (2015). Deletion analysis of RcsC reveals a novel signalling pathway controlling poly-N-acetylglucosamine synthesis and biofilm formation in Escherichia coli. Microbiology 161, 903–913. doi: 10.1099/mic.0.000050
O’Toole, G. A. (2011). Microtiter dish biofilm formation assay. J. Vis. Exp. 47:2437. doi: 10.3791/2437
Perez-Rodriguez, R., Haitjema, C., Huang, Q., Nam, K. H., Bernardis, S., Ke, A., et al. (2011). Envelope stress is a trigger of CRISPR RNA-mediated DNA silencing in Escherichia coli. Mol. Microbiol. 79, 584–599. doi: 10.1111/j.1365-2958.2010.07482.x
Pérez-Toledo, M., Valero-Pacheco, N., Pastelin-Palacios, R., Gil-Cruz, C., Pérez-Shibayama, C., Moreno-Eutimio, M. A., et al. (2017). Salmonella Typhi porins OmpC and OmpF are potent adjuvants for T-dependent and T-independent antigens. Front. Immunol. 8:230. doi: 10.3389/fimmu.2017.00230
Pickard, D., Li, J., Roberts, M., Maskell, D., Hone, D., Levine, M., et al. (1994). Characterization of defined ompR mutants of Salmonella Typhi: ompR is involved in the regulation of Vi polysaccharide expression. Infect. Immun. 62, 3984–3993. doi: 10.1128/IAI.62.9.3984-3993.1994
Pruss, B. M. (2017). Involvement of two-component signaling on bacterial motility and biofilm development. J. Bacteriol. 199, e00259–e002517. doi: 10.1128/JB.00259-17
Puente, J. L., Flores, V., Fernández, M., Fuchs, Y., and Calva, E. (1987). Isolation of an ompC-like outer membrane protein gene from Salmonella Typhi. Gene 61, 75–83. doi: 10.1016/0378-1119(87)90366-0
Puente, J. L., Juárez, D., Bobadilla, M., Arias, C. F., and Calva, E. (1995). The Salmonella ompC gene: structure and use as a carrier for heterologous sequences. Gene 156, 1–9. doi: 10.1016/0378-1119(94)00883-T
Rodríguez-Morales, O., Fernández-Mora, M., Hernández-Lucas, I., Vázquez, A., Puente, J. L., and Calva, E. (2006). Salmonella enterica serovar Typhimurium ompS1 and ompS2 mutants are attenuated for virulence in mice. Infect. Immun. 74, 1398–1402. doi: 10.1128/IAI.74.2.1398-1402.2006
Samai, P., Pyenson, N., Jiang, W., Goldberg, G. W., Hatoum-Aslan, A., and Marraffini, L. A. (2015). Co-transcriptional DNA and RNA cleavage during type III CRISPR-Cas immunity. Cell 161, 1164–1174. doi: 10.1016/j.cell.2015.04.027
Samanta, P., Clark, E. R., Knutson, K., Horne, S. M., and Pruss, B. M. (2013). OmpR and RcsB abolish temporal and spatial changes in expression of flhD in Escherichia coli biofilm. BMC Microbiol. 13:182. doi: 10.1186/1471-2180-13-182
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989). Molecular cloning: A laboratory manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Sampson, T. R., Napier, B. A., Schroeder, M. R., Louwen, R., Zhao, J., Chin, C. Y., et al. (2014). A CRISPR-Cas system enhances envelope integrity mediating antibiotic resistance and inflammasome evasion. Proc. Natl. Acad. Sci. U. S. A. 111, 11163–11168. doi: 10.1073/pnas.1323025111
Sampson, T. R., Saroj, S. D., Llewellyn, A. C., Tzeng, Y. L., and Weiss, D. S. (2013). A CRISPR/Cas system mediates bacterial innate immune evasion and virulence. Nature 497, 254–257. doi: 10.1038/nature12048
Secundino, I., López-Macias, C., Cervantes-Barragan, L., Gil-Cruz, C., Ríos-Sarabia, N., Pastelin-Palacios, R., et al. (2006). Salmonella porins induce a sustained, lifelong specific bactericidal antibody memory response. Immunology 117, 59–70. doi: 10.1111/j.1365-2567.2005.02263.x
Shariat, N., Timme, R. E., Pettengill, J. B., Barrangou, R., and Dudley, E. G. (2015). Characterization and evolution of Salmonella CRISPR-Cas systems. Microbiology 161, 374–386. doi: 10.1099/mic.0.000005
Shin, S., and Park, C. (1995). Modulation of flagellar expression in Escherichia coli by acetyl phosphate and the osmoregulator OmpR. J. Bacteriol. 177, 4696–4702. doi: 10.1128/JB.177.16.4696-4702.1995
Shmakov, S. A., Makarova, K. S., Wolf, Y. I., Severinov, K. V., and Koonin, E. V. (2018). Systematic prediction of genes functionally linked to CRISPR-Cas systems by gene neighborhood analysis. Proc. Natl. Acad. Sci. U. S. A. 115, E5307–E5316. doi: 10.1073/pnas.1803440115
Sinkunas, T., Gasiunas, G., Fremaux, C., Barrangou, R., Horvath, P., and Siksnys, V. (2011). Cas3 is a single-stranded DNA nuclease and ATP-dependent helicase in the CRISPR/Cas immune system. EMBO J. 30, 1335–1342. doi: 10.1038/emboj.2011.41
Spector, M. P., and Kenyon, W. J. (2012). Resistance and survival strategies of Salmonella enterica to environmental stresses. Food Res. Int. 45, 455–481. doi: 10.1016/j.foodres.2011.06.056
Taboada, B., Estrada, K., Ciria, R., and Merino, E. (2018). Operon-mapper: a web server for precise operon identification in bacterial and archaeal genomes. Bioinformatics 34, 4118–4120. doi: 10.1093/bioinformatics/bty496
Touchon, M., and Rocha, E. P. (2010). The small, slow and specialized CRISPR and anti-CRISPR of Escherichia and Salmonella. PLoS One 5:e11126. doi: 10.1371/journal.pone.0011126
Vidal, O., Longin, R., Prigent-Combaret, C., Dorel, C., Hooreman, M., and Lejeune, P. (1998). Isolation of an Escherichia coli K-12 mutant strain able to form biofilms on inert surfaces: involvement of a new ompR allele that increases curli expression. J. Bacteriol. 180, 2442–2449. doi: 10.1128/JB.180.9.2442-2449.1998
Villarreal, J. M., Becerra-Lobato, N., Rebollar-Flores, J. E., Medina-Aparicio, L., Carbajal-Gómez, E., Zavala-García, M. L., et al. (2014). The Salmonella enterica serovar Typhi ltrR-ompR-ompC-ompF genes are involved in resistance to the bile salt sodium deoxycholate and in bacterial transformation. Mol. Microbiol. 92, 1005–1024. doi: 10.1111/mmi.12610
Viswanathan, P., Murphy, K., Julien, B., Garza, A. G., and Kroos, L. (2007). Regulation of dev, an operon that includes genes essential for Myxococcus xanthus development and CRISPR-associated genes and repeats. J. Bacteriol. 189, 3738–3750. doi: 10.1128/JB.00187-07
Wallace, R. A., Black, W. P., Yang, X., and Yang, Z. (2014). A CRISPR with roles in Myxococcus xanthus development and exopolysaccharide production. J. Bacteriol. 196, 4036–4043. doi: 10.1128/JB.02035-14
Westra, E. R., van Erp, P. B., Kunne, T., Wong, S. P., Staals, R. H., Seegers, C. L., et al. (2012). CRISPR immunity relies on the consecutive binding and degradation of negatively supercoiled invader DNA by Cascade and Cas3. Mol. Cell 46, 595–605. doi: 10.1016/j.molcel.2012.03.018
Yoshida, T., Qin, L., Egger, L. A., and Inouye, M. (2006). Transcription regulation of ompF and ompC by a single transcription factor, OmpR. J. Biol. Chem. 281, 17114–17123. doi: 10.1074/jbc.M602112200
Keywords: CRISPR-Cas, porin regulation, Salmonella Typhi, OmpR, outer membrane proteins
Citation: Medina-Aparicio L, Rodriguez-Gutierrez S, Rebollar-Flores JE, Martínez-Batallar ÁG, Mendoza-Mejía BD, Aguirre-Partida ED, Vázquez A, Encarnación S, Calva E and Hernández-Lucas I (2021) The CRISPR-Cas System Is Involved in OmpR Genetic Regulation for Outer Membrane Protein Synthesis in Salmonella Typhi. Front. Microbiol. 12:657404. doi: 10.3389/fmicb.2021.657404
Edited by:
Graciela Castro Escarpulli, Instituto Politécnico Nacional de México (IPN), MexicoReviewed by:
Josep Casadesús, Sevilla University, SpainRolf Dieter Joerger, University of Delaware, United States
Copyright © 2021 Medina-Aparicio, Rodriguez-Gutierrez, Rebollar-Flores, Martínez-Batallar, Mendoza-Mejía, Aguirre-Partida, Vázquez, Encarnación, Calva and Hernández-Lucas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ismael Hernández-Lucas, aXNtYWVsLmhlcm5hbmRlekBpYnQudW5hbS5teA==; aXNtYWVsaEBpYnQudW5hbS5teA==