 Feng-Jie Jin
Feng-Jie Jin Shuang Hu
Shuang Hu Long Jin
Long Jin
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol. , 23 February 2021
Sec. Food Microbiology
Volume 12 - 2021 | https://doi.org/10.3389/fmicb.2021.644404
This article is part of the Research Topic From Traditional to Modern: Progress of Molds and Yeasts in Fermented-Food Production View all 45 articles
The filamentous fungus Aspergillus oryzae is an important strain in the traditional fermentation and food processing industries and is often used in the production of soy sauce, soybean paste, and liquor-making. In addition, A. oryzae has a strong capacity to secrete large amounts of hydrolytic enzymes; therefore, it has also been used in the enzyme industry as a cell factory for the production of numerous native and heterologous enzymes. However, the production and secretion of foreign proteins by A. oryzae are often limited by numerous bottlenecks that occur during transcription, translation, protein folding, translocation, degradation, transport, secretion, etc. The existence of these problems makes it difficult to achieve the desired target in the production of foreign proteins by A. oryzae. In recent years, with the decipherment of the whole genome sequence, basic research and genetic engineering technologies related to the production and utilization of A. oryzae have been well developed, such as the improvement of homologous recombination efficiency, application of selectable marker genes, development of large chromosome deletion technology, utilization of hyphal fusion techniques, and application of CRISPR/Cas9 genome editing systems. The development and establishment of these genetic engineering technologies provided a great deal of technical support for the industrial production and application of A. oryzae. This paper reviews the advances in basic research and genetic engineering technologies of the fermentation strain A. oryzae mentioned above to open up more effective ways and research space for the breeding of A. oryzae production strains in the future.
Aspergillus oryzae is an important filamentous fungus that is widely applied in the traditional fermentation and food processing industries, such as soy sauce, soybean paste, and sake brewing. This long history of its widespread use in the food industry has led to A. oryzae being recognized by the Food and Drug Administration (FDA) of the United States as Generally Recognized as Safe (GRAS) organisms. Its safety is also supported by the World Health Organization (FAO/WHO) (Kobayashi et al., 2007).
In addition, A. oryzae has a strong capacity to secrete large amounts of hydrolytic enzymes (such as amylases and proteases); therefore, it has also been used in the enzyme industry as a cell factory for the production of numerous native and heterologous enzymes. In addition to protein secretion ability, A. oryzae also has a strong synthesis ability, rapid growth, and easy culture, among its advantages. Compared to Escherichia coli and yeast Saccharomyces cerevisiae, A. oryzae has a strong posttranslational modification function; therefore, it is widely used to produce a variety of enzyme preparations (Lissau et al., 1998; Hama et al., 2008; Merz et al., 2015) and in recent years has also been used in the production of secondary metabolites (Yamada et al., 2014; Kan et al., 2019; Liu et al., 2019; Nagamine et al., 2019; Morishita et al., 2020; Oikawa, 2020a).
In the past few decades, research on A. oryzae has focused on breeding techniques and developing methods for brewing. Despite its considerable commercial importance, the study of fundamental biological processes in A. oryzae was delayed in comparison with the yeast S. cerevisiae. The main reasons are that A. oryzae is an imperfect fungus that does not have a sexual life cycle and forms multinucleate conidia, making classical genetic manipulation difficult to implement (Kitamoto, 2015). Therefore, it is difficult to study this organism using conventional genetic methods. The production and secretion of foreign proteins by A. oryzae are often limited by numerous bottlenecks during transcription, translation, protein folding, translocation, degradation, transport, secretion, etc. (Conesa et al., 2001; Ohno et al., 2011). The existence of these problems makes it difficult to achieve the desired target in heterologous protein production by A. oryzae.
In 2005, the genome sequence of A. oryzae RIB40 was deciphered (Machida et al., 2005), which marked the advent of the whole genome era of A. oryzae, and the combination of genetic engineering technology further promoted the development of A. oryzae. Subsequently, many other A. oryzae strains were sequenced, such as 3.042, which is extensively used for the production of both soy sauce and other fermented foods in China (Zhao et al., 2012). Comparative genomics analysis showed some strain-specific genes that encode putative proteins potentially involved in cell growth, salt tolerance, environmental resistance and flavor formation (Zhao et al., 2013). The 37-Mb genome of A. oryzae RIB40 was predicted to contain more than 12,000 genes, which is 7–9 MB larger than the genomes of Aspergillus nidulans and Aspergillus fumigatus (Galagan et al., 2005; Machida et al., 2005; Nierman et al., 2005). The completed genomic studies using expressed sequence tag analyses (Machida, 2002; Akao et al., 2007) combined with the development of genetic engineering technology have made it possible to overcome these deficiencies and promoted the study of heterologous protein production in A. oryzae and the breeding of new strains for industrial production (Vongsangnak et al., 2008; Zhu et al., 2013). Regarding genetic engineering to solve the bottleneck problems in heterologous protein production in A. oryzae, this article elaborated on several aspects, such as optimization of the genetic transformation system and the improvement of transformation efficiency, construction of protease gene-deleted strains, development of fusion expression strategies, optimization of exogenous gene expression systems, and development of other genetic engineering techniques to improve the production of foreign proteins. These advances in basic research and genetic engineering technology have greatly improved the production and application of A. oryzae.
In industrial applications, the establishment of a highly efficient genetic transformation system is a prerequisite for genetic manipulation and recombinant gene expression in A. oryzae. Compared to E. coli and yeast S. cerevisiae, the A. oryzae cell wall is hard and not easily broken, which is an obstacle for gene introduction into host cells in later genetic transformation experiments. Therefore, some rapid and convenient transformation methods, such as the electroporation and gene gun methods, are not suitable for A. oryzae genetic transformation due to their extremely low transformation efficiency. By contrast, another conventional genetic transformation method, which is a polyethylene glycol (PEG)/CaCl2-mediated protoplast transformation system, was widely utilized in A. oryzae.
The application of recombinant DNA methodology in A. oryzae allows advanced molecular genetic analyses, which makes it possible to introduce DNA fragments into the host to produce heterologous proteins (Kitamoto, 2002). For effective molecular genetic analysis and protein production, suitable and stable hosts are needed as genetically engineered strains to transform the selected target genes. In previous research, several auxotrophic marker genes, such as pyrG, argB, niaD, and sC (Gomi et al., 1987; Mattern et al., 1987; Unkles et al., 1989; Yamada et al., 1997), and dominant selectable markers, such as amdS and ptrA (Gomi et al., 1991; Kubodera et al., 2000), have been developed and used in the genetic transformation of A. oryzae. Recently, a new pyrithiamine resistance marker gene, thiI, was also identified for use in genome editing by selecting mutants resistant to pyrithiamine from UV-mutagenized A. oryzae strains (Todokoro et al., 2020). In addition to these, a bleomycin resistance marker gene (blmB) and a carboxin resistance marker gene [AosdhB(cxr)] also were developed for A. oryzae transformation (Shima et al., 2009; Suzuki et al., 2009). However, all these transformation techniques only use a single gene as a selective marker gene; therefore, several multitransformation systems of A. oryzae containing multiple selective marker gene-defective strains were constructed, which could carry out gene engineering operations such as introduction or deletion of multiple genes in the same host. For this purpose, Yamada et al. (1997) further developed a double auxotrophic transformation system (niaD–, sC–) that can conduct two genetic manipulations in a single host strain. On this basis, triple auxotrophic mutants (niaD–, sC–, adeA–/adeB–) were further obtained from A. oryzae by UV mutagenesis, which added a new adenine auxotrophy (Jin et al., 2004a). In addition, a quadruple auxotrophic host transformation system was further developed by argB gene deletion combined with exploiting adenine auxotrophy (Jin et al., 2004b). Among these selection marker genes, the pyrG gene, which encodes an orotidine-5′-phosphate (OMP) decarboxylase, has been used for marker recycling that allows multiple genetic manipulations in the Aspergillus species (Nielsen et al., 2006; Maruyama and Kitamoto, 2008) because pyrG-deficient strains can be positively selected by 5-fluoroorotic acid (5-FOA), which is converted to a toxic compound, to strains expressing pyrG. These selectable marker genes mentioned above are summarized in Table 1. In addition, the Cre-loxP site-specific recombination system, which has been widely applied in a number of organisms, such as bacterial, yeast, plant, and mammalian cells (Sauer, 1987; Gu et al., 1993; Sieburth et al., 1998), was also developed for multiple gene integrations and deletions in A. oryzae (Mizutani et al., 2012; Zhang et al., 2017). This system is used for site-specific recombination between two loxP sites mediated by a Cre recombination protein derived from bacteriophage P1 (Abremski et al., 1986).

Table 1. Selectable marker genes used in Aspergillus oryzae.
One of the major reasons for the lag in basic research on A. oryzae is the low efficiency of homologous recombination transformation, which makes it difficult to obtain gene deletion strains. Takahashi et al. (2006) significantly improved the homologous recombination efficiency of A. oryzae by deleting non-homologous end-joining (NHEJ)-related genes such as ku70 and ku80, thus enhancing the gene deletion efficiency of A. oryzae. Next, some studies also showed that the disruption of another NHEJ-related ligD gene significantly improved gene-targeting efficiency in A. oryzae (Maruyama and Kitamoto, 2008; Mizutani et al., 2008). With the significant improvement in the efficiency of homologous recombination, deletion technology of large chromosomal segments, including loop-out recombination and replacement-type recombination methods, has been developed (Takahashi et al., 2008, 2009), providing technical support for the further deletion of unnecessary and even harmful large chromosomal segments from the chromosomes in A. oryzae. Although A. oryzae is regarded as an industrially important fungus, its genome sequence still contains several gene clusters related to the synthesis of toxins, such as imperfect clustered genes related to aflatoxin biosynthesis and cyclopiazonic acid production. Although these toxic secondary metabolites are not normally synthesized, they are also at a certain risk, and these gene clusters can be deleted through the deletion technology of large chromosomal segments. On this basis, Jin et al. (2010) conducted a chromosome-shortening experiment, and most of the unnecessarily long DNA fragments and potential toxin synthesis-related gene clusters on chromosome 7 of A. oryzae were successfully removed, which not only promoted the production of amylase but also avoided the production of some byproducts that are possibly harmful to human health. By eliminating non-essential chromosomal segments and retaining only the genes that maintain basic physiological functions, we will gain a deeper understanding of the physiological characteristics of this organism and the essential gene functions for its industrial use. The development of these techniques pave a new way for the future production and utilization of A. oryzae.
In addition, A. oryzae was thought to undergo asexual reproduction, and no sexual stage has been reported; therefore, it has certain limitations in production and application. For instance, breeding better production strains cannot be achieved by sexual hybridization. To solve this problem, protoplast fusion technology was developed, and a further reduced chromosome A. oryzae mutant was constructed with a shortened chromosomes 7 and 8, both of which include a large number of non-syntenic blocks by using this protoplast fusion technique (Hara et al., 2012). The large chromosome segment deletion technique combined with the protoplast fusion technique has effectively solved the problem that has puzzled A. oryzae researchers for many years and provided a good technical means for better chromosome processing in the future to obtain the breeding of excellent A. oryzae production strains. In addition, using the large-scale chromosome deletion technique, a member of the basic helix-loop-helix (bHLH) transcription factor family, SclR, was identified and characterized (Jin et al., 2009, 2011b). Studies have shown that SclR cannot only promote sclerotia formation by promoting hyphal fusion but also effectively promote nucleus fusion between different strains (Wada et al., 2014), which provides a more feasible space for breeding excellent A. oryzae strains in the future. These studies indicated that the deletion technique of large chromosome segments is also of great value for the discovery and exploration of unknown functional genes.
Aspergillus oryzae is resistant to most common antifungal antibiotics that are often used as selective agents for transformation experiments of filamentous fungi (Suzuki et al., 2009); therefore, A. oryzae transformation systems were developed mainly based on nutritional selectable markers for complementation of auxotrophic strains (Gomi et al., 1987; Mattern et al., 1987). Currently, PEG-mediated protoplast transformation is the common method utilized for fungal transformation experiments (Liu and Friesen, 2012). The Agrobacterium tumefaciens-mediated transformation (ATMT) system is another method that was developed and used for genetic transformation in the Aspergilli species (Michielse et al., 2005). The approach usually requires a binary vector carrying T-DNA (transfer DNA), an A. tumefaciens strain that harbors a helper plasmid with vir (virulence) supporting the T-DNA transfer process, and a suitable recipient (Michielse et al., 2005; Frandsen, 2011). Using the ATMT method to transfer DNA into the fungal genome has been shown to be more efficient and simpler to implement in different filamentous fungi, while PEG-mediated protoplast transformation is more laborious and time-consuming for protoplast preparation (Idnurm et al., 2017; Weyda et al., 2017). Recently, the efficient approach was used to successfully construct uridine/uracil auxotrophic mutants by deleting the pyrG gene in different industrial fungus A. oryzae wild-type strains, and the genetic transformation efficiency by A. tumefaciens was higher and reached approximately 0.1% using the pyrG selectable marker and these auxotrophic mutants (Nguyen et al., 2016, 2017). These generated auxotrophic mutants were also used to successfully express the DsRed fluorescent reporter gene under the control of the A. oryzae alpha-amylase gene (amyB) promoter by the ATMT method. Furthermore, using the ATMT method, a useful dual selection marker system was developed for genetic manipulations in an industrial A. oryzae 3.042 strain by constructing a uridine/uracil auxotrophic mutant and pyrithiamine (PT) genes as selection markers (Sun et al., 2019). In summary, this ATMT method can simplify transformation procedures and improve the efficiency of genetic manipulations. This strategy can be applied in functional gene research and the production of recombinant proteins by the industrial fungus Aspergillus strains through genetic transformation systems based on nutrition markers.
The increase in the number of whole genomes available for fungal species provided a wider range of possibilities for gene manipulation in filamentous fungi. In recent years, a versatile genomic editing technique, the CRISPR (clustered regularly interspaced short palindromic repeat)–Cas9 (CRISPR-related nuclease 9) system, has been rapidly developed and widely applied in filamentous fungi. The advance of this technology has revolutionized biological research and led to innovative applications in a wide range of fields, showing great prospects in the research and application of filamentous fungi (Song et al., 2019).
The CRISPR/Cas9 system is divided into two categories, including six types and 19 subtypes (Shmakov et al., 2017). Among them, the type II system is a simpler CRISPR system than other systems that have been widely used. The type II CRISPR/Cas9 system contains Cas9 (nuclease), mature crRNA (CRISPR associate RNA), tracrRNA (trans-activating crRNA), and RNaseIII. The crRNA and tracrRNA have been modified and linked into a single-guide RNA (sgRNA) (Jinek et al., 2012) that could efficiently guide the Cas9 nuclease to the target sequences to cut the DNA. Currently, CRISPR/Cas9 technology has been widely used for genome editing in yeasts/fungi, plants, mammalian cells, and others (Cong et al., 2013; van Erp et al., 2015; Schiml and Puchta, 2016). In fungi, the CRISPR/Cas9 genome editing system was first introduced into S. cerevisiae (DiCarlo et al., 2013) and then further applied to Trichoderma reesei, Neurospora crassa, and Aspergillus nidulans (Liu et al., 2015; Matsu-Ura et al., 2015; Nodvig et al., 2018). In addition, the CRISPR/Cas9 system was discovered in bacteria or archaea, and therefore, the Cas9-encoding gene usually needs to be fungal codon-optimized and a nuclear localization signal when the genome editing system is used in fungi (Generoso et al., 2016; Nodvig et al., 2018). In recent years, some studies have shown that the CRISPR/Cas9 technique has been used in the production of recombinant proteins in the Aspergilli species (Dong et al., 2020; Rojas-Sanchez et al., 2020).
To develop the CRISPR/Cas9 genome editing technique in A. oryzae, plasmids that express the Cas9 nuclease gene and sgRNAs were constructed and introduced into an A. oryzae strain for the mutagenesis of target genes (Katayama et al., 2016). In this study, the codon usage of cas9 with a sequence encoding an SV40 nuclear localization signal at its 5′- and 3′-ends was optimized to improve the nuclear localization of Cas9 nuclease. The generated transformants contained mutations in which each target gene showed the expected phenotype. Mutation rates between 10 and 20% with 1 bp of deletion or insertion are the most common induced mutations, and genome editing technology will contribute to the efficient use of targeted mutagenesis in many A. oryzae industrial strains without a clear genetic background. To further improve the targeting efficiency in A. oryzae industrial strains, the ligD gene involved in NHEJ was mutated by employing the CRISPR/Cas9 system (Nakamura et al., 2017). The gene targeting efficiency of the generated ligD mutant was examined using the ecdR gene, which encodes a regulator involved in the early stage of conidiophore development in the A. oryzae (Jin et al., 2011a). The results indicated that the deletion efficiency was significantly improved and reached approximately 60∼80% in the ligD mutants.
In addition to food fermentation and industrial production of recombinant proteins, A. oryzae has also been used for heterologous production of useful secondary metabolites; therefore, multiple steps of genetic manipulation are required.
An improved CRISPR/Cas9 approach that includes an AMA1-based autonomously replicating plasmid harboring the pyrithiamine resistance marker gene ptrA was further developed (Katayama et al., 2019). The conditional expression of the Aoace2 gene in the AMA1-based plasmid significantly repressed the growth of fungi, which made it possible for forced plasmid recycling and allowed repeated genome editing. This study showed that the repeatable marker-free genome editing approach can effectively delete or integrate multiple genes in the industrial A. oryzae strain (Katayama et al., 2019). In the production experiment of human antibody (anti-TNFα antibody adalimumab) by A. oryzae, the CRISPR/Cas9 technique was used for the deletion of the Aooch1 gene, which encodes a key enzyme for the hypermannosylation process, to detect the binding ability of the recombinant antibody with FcγRIIIa (Huynh et al., 2020). In addition, the CRISPR/Cas9 system was also used to delete AoGld3, which is one of 45 glycerol dehydrogenase genes in A. oryzae, to investigate its effect on the production of the secondary metabolite kojic acid. The result showed that the deletion of AoGld3 resulted in the inhibition of kojic acid production by affecting the expression of kojA and kojR, which encode an enzyme and a transcription factor involved in the kojic acid biosynthesis (Fan et al., 2020). In addition, a platinum-fungal TALEN (Transcription activator-like effector nucleases) -based genome editing technique has also been developed for targeted gene mutations in A. oryzae (Mizutani et al., 2017). The platinum-fungal TALEN technique resulted in various different mutation patterns with almost half of deletions larger than 1 kb in the target region. Taken together, these genome editing technologies will greatly enhance the convenience of genetic manipulation in a wide range of industrial production strains and significantly contribute to efficient molecular breeding of these industrial strains.
Aspergillus oryzae has attracted increasing attention due to its wide use as a cell factory in the study of exogenous gene expression and heterologous protein production in recent years. An understanding of how optimize the exogenous gene expression system is of great significance to the industrial application of A. oryzae. By analyzing the advantages and disadvantages of the exogenous gene expression system of A. oryzae and combining genetic engineering technology developed in recent years, the exogenous gene expression of A. oryzae can be improved by making full use of the advantages while avoiding the disadvantages.
Compared with the production and secretion system of prokaryotes such as E. coli, exogenous genes expressed in A. oryzae undergo eukaryotic posttranslational modifications, including glycosylation and protein folding. Therefore, A. oryzae is considered as one of the most advantageous hosts for expressing recombinant proteins from higher eukaryotes (Fleissner and Dersch, 2010). The production of heterologous proteins is limited by many factors, such as the transcriptional level, translation level, secretory process, and extracellular degradation (Jeenes et al., 1991). Therefore, several bottleneck genes related to proteases, secretion pathways, cell polarity establishment, and metabolism must be manipulated to breed favorable hosts for heterologous protein production.
Among them, the strong protease activity of A. oryzae is the most important reason to inhibit the production of heterologous proteins because heterologous proteins are more easily broken down by proteases than the organism’s own proteins. To improve the yield of heterologous proteins, researchers have tried various biological methods. For instance, downstream processing at low temperature, early separation of products from proteases, application of protease inhibitors, etc. can reduce hydrolysis; however, the effect is not optimistic because most heterologous proteins could be degraded in the process of protein production (Jin et al., 2007). Using a quadruple auxotrophic host (Jin et al., 2004b), Jin et al. (2007) expressed the heterologous human lysozyme (HLY) gene by disrupting an intracellular acid protease PepE and a tripeptidyl peptidase TppA genes, and the double disruption led to an increase (63%) in HLY production in comparison to the control. In this study, the HLY gene fused with an A. oryzae α-amylase (AmyB) gene was expressed, the Kex2 cleavage site was inserted between them, and HLY was successfully processed and dissociated from the native AmyB protein and secreted into the medium. In addition, a systematic deletion analysis of five proteinase genes (pepA, pepE, tppA, alpA, and palB) in the HLY-producing strain was performed, and comparative analysis indicated that deletion of the tppA gene resulted in the highest increase (36%) in lysozyme activity of the HLY-producing strain (Jin et al., 2007). On this basis, a quintuple protease gene disrupted strain (tppA, pepE, dppIV, dppV, and nptB) (Yoon et al., 2009) and an additional ten protease gene disrupted strain with five additional deleted genes (alpA, pepA, AopepAa, AopepAd, and cpI) (Yoon et al., 2011) were successively constructed using pyrG marker recycling combined with a highly efficient gene targeting background (△ligD). The results showed that the yield of HLY and bovine chymosin (CHY) protein in the ten protease gene disruptants was significantly enhanced, which was 3.2 and 3.8 times that of the control strains, respectively.
To further solve the bottlenecks limiting the production of heterologous protein by A. oryzae, another tripeptidyl peptidase gene, AosedD, which may be involved in the proteolytic degradation of recombinant proteins, was selected to construct its deletion strain for the production of heterologous proteins. In the AosedD deletion mutant, the production levels of recombinant HLY and CHY improved approximately 1.7- and 2.9-fold, respectively, relative to their control strains (Zhu et al., 2012). The results show that AoSedD is one of the main proteases involved in the degradation of heterologous recombinant proteins in A. oryzae.
Another important bottleneck in the production of heterologous proteins in filamentous fungi is the secretion of the proteins (Conesa et al., 2001). The unfolded protein response (UPR) is an intracellular regulatory pathway that activates the genes involved in folding, quality control, transport, and other functions of secreted proteins. The overexpression of heterologous proteins may result in the overload of unfolded proteins in the endoplasmic reticulum (ER) (Ron and Walter, 2007). The response is accompanied by the transcription activation of ER protein folding- and secretion-related genes. Some studies have shown that heterologous protein production upregulates the expression of ER chaperone genes, such as bipA (a major endoplasmic reticulum chaperone protein gene), prpA (a PDI [protein disulfide isomerase]-related gene), and clxA (a calnexin homologue from Aspergillus niger) (Punt et al., 1998; Wang and Ward, 2000; Wang et al., 2003). To examine the effects of carrier fusion on the UPR and secretion of the target protein, a DNA microarray was used to analyze the transcriptional level of the A. oryzae whole genome using CHY as a model heterologous protein (Ohno et al., 2011). The production level of a carrier-fused CHY with α-amylase increased by approximately twofold in comparison with the non-carrier-fused CHY, and several genes involved in ER protein folding and secretion were significantly upregulated in the carrier fusion CHY strain by DNA microarray analysis. Similarly, the transcripts of hacA, which encodes the UPR transcription factor required for upregulation of the ER chaperone and foldase-encoding genes (Mulder et al., 2006), were effectively spliced in the carrier-fused CHY strain. These results suggest that carrier-fused CHY temporarily accumulated in the ER and remarkably induced UPR in the carrier-fused strain (Ohno et al., 2011). However, recent studies have shown that highly expressed endogenous secretory proteins can also evoke the UPR. This finding raises the possibility that unfolded or misfolded proteins are selectively recognized by quality control mechanisms in filamentous fungi (Yokota et al., 2017).
Protein trafficking through the secretory pathway depends on the production levels of foldases and chaperones, which play crucial roles in protein folding during secretion. Some studies have shown that the overexpression of ER foldase- and molecular chaperone-related genes partially alleviate ER overload. It has been reported that the overexpression of chaperone BipA and PDI genes improve the production of heterologous proteins (Moralejo et al., 2001; Lombrana et al., 2004). Among them, the overexpression of bipA increased thaumatin secretion by 2- to 2.5-fold compared to the control strain; conversely, homologous protein secretion was not influenced by its overexpression in Aspergillus awamori.
In addition, vacuolar protein sorting (VPS), autophagy, and ER-Golgi cargo receptors also affect the heterologous protein production of A. oryzae (Yoon et al., 2013; Hoang et al., 2015). Autophagy is a highly conserved intracellular degradation pathway in eukaryotes that can recycle intracellular components as a survival mechanism under nutritional starvation (Reggiori and Klionsky, 2002). Furthermore, autophagy was confirmed to deliver misfolded secreted proteins accumulated in the ER to vacuoles, which is an important process that adversely affects heterologous protein production in A. oryzae (Kimura et al., 2011). In a previous study, according to the homology analysis with S. cerevisiae, five putative autophagy-related genes: Aoatg1, encoding a kinase involved in autophagy induction; Aoatg4 and Aoatg8, which are essential genes for the formation of autophagosomes and membrane fusion; Aoatg13, encoding a component of the AoAtg1 complex; and Aoatg15, encoding a lipase required for the breakdown of autophagic bodies, were selected and successfully deleted in A. oryzae (Yoon et al., 2013). Using CHY as a reporter heterologous protein, the production level of CHY in the Aoatg1, Aoatg4, and Aoatg8 deletion strains was improved by 2.3-, 3.1-, and 2.5-fold, respectively, compared to the control strain.
In addition, homologs of several VPS genes, which play crucial roles in the secretory pathway, have been identified and characterized (Lemmon and Traub, 2000; Suzuki et al., 2003). Among them, VPS10 has been identified to encode a sorting receptor for the recognition and delivery of some vacuolar proteins in yeast (Cooper and Stevens, 1996). Although it was also found to target aberrant and recombinant proteins for vacuolar degradation, the effect of VPS10 gene deletion on the production of heterologous proteins is unclear. Studies have shown that the vps10 gene deletion resulted in missorting of vacuolar carboxypeptidase CpyA and was then secreted into the medium (Marcusson et al., 1994), indicating that Vps10 is necessary for sorting vacuolar proteins into vacuoles. The analysis of heterologous protein production indicated that the Aovps10 deletion mutation elevated the maximum extracellular production levels of the heterologous proteins HLY and CHY by 2.2- and 3-fold, respectively (Yoon et al., 2010). These results revealed that AoVps10 is involved in the regulation of heterologous protein secretion and vacuolar protein degradation in A. oryzae.
To improve the production of heterologous proteins, in addition to reducing the activity of proteases, a series of strong promoters were developed to induce the expression of heterologous genes (Table 2). In A. oryzae, the starch-inducible promoter of the alpha-amylase gene (PamyB) is one of the most commonly used promoters for the production of foreign proteins (Tsuchiya et al., 1992; Ward et al., 1992; Rey et al., 2003; Hoshida et al., 2005; Yamashita et al., 2011; Yin et al., 2015). The glucoamylase gene promoter (PglaA) and its improved promoter (PglaA142) were often used for the production and function analysis of recombinant proteins, such as Candida antarctica lipase B and A. oryzae recombinant tannase (Tamalampudi et al., 2007; Tokuoka et al., 2008; Ichikawa et al., 2020). The constitutive gpdA gene promoter from A. nidulans (de Ruiter-Jacobs et al., 1989; te Biesebeke et al., 2006; Punya et al., 2013), thiamine-regulatable promoter of the thiamine thiazole synthase gene (PthiA) (Shoji et al., 2005), and the promoter of the translation-elongation factor 1 alpha gene (Ptef1) (Kitamoto et al., 1998) were also used for homologous and heterologous protein production in A. oryzae. In addition, the glucoamylase-encoding gene (glaB) promoter was developed for recombinant protein production in solid-state fermentation of A. oryzae (Ishida et al., 2006), and the promoter of the oxidoreductase gene kojA involved in the kojic acid biosynthesis, was successfully used to induce the expression of the polyketide synthase gene (wA) by A. oryzae (Tamano et al., 2019). The promoters used for A. oryzae recombinant protein production are summarized in Table 2. In addition to these promoters, studies have shown that the alterations in 5′ untranslated region (5′ UTR) importantly improved the translation efficiency of heterologous proteins in A. oryzae (Koda et al., 2004, 2006). The results suggested that the 5′ UTR can be used in combination with various strong promoters to enhance the expression of heterologous proteins. Furthermore, the host endogenous high-secreted protein genes were also further developed for the fusion and expression of exogenous protein genes (Gasser and Mattanovich, 2007; Jin et al., 2007; Hoang et al., 2015). Current studies have shown that the fusion of exogenous protein genes into the end of the highly secreted endogenous protein genes of A. oryzae reduce the degradation of heterologous proteins by the protease of A. oryzae (Ohno et al., 2011). For example, when a reporter heterologous protein CHY was expressed with fused α-amylase (AmyB), which is one of the most abundantly expressed proteins in A. oryzae under starch-induced culture conditions, the production level increased by approximately twofold compared to the non-carrier fused CHY (Ohno et al., 2011). Another study showed that using the A. oryzae glucoamylase gene (glaA) as a carrier protein, carrier-fused recombinant chymosin was secreted at a high level and showed a fivefold higher chymosin production level compared with the control using the glaA promoter alone (Tsuchiya et al., 1994). Furthermore, a mite allergen Der f 7 gene was highly expressed by fusion with the A. oryzae glucoamylase carrier protein, and its expression level was further enhanced by codon optimization which is also an effective method for improving the production levels of heterologous proteins (Tokuoka et al., 2008; Tanaka et al., 2012).
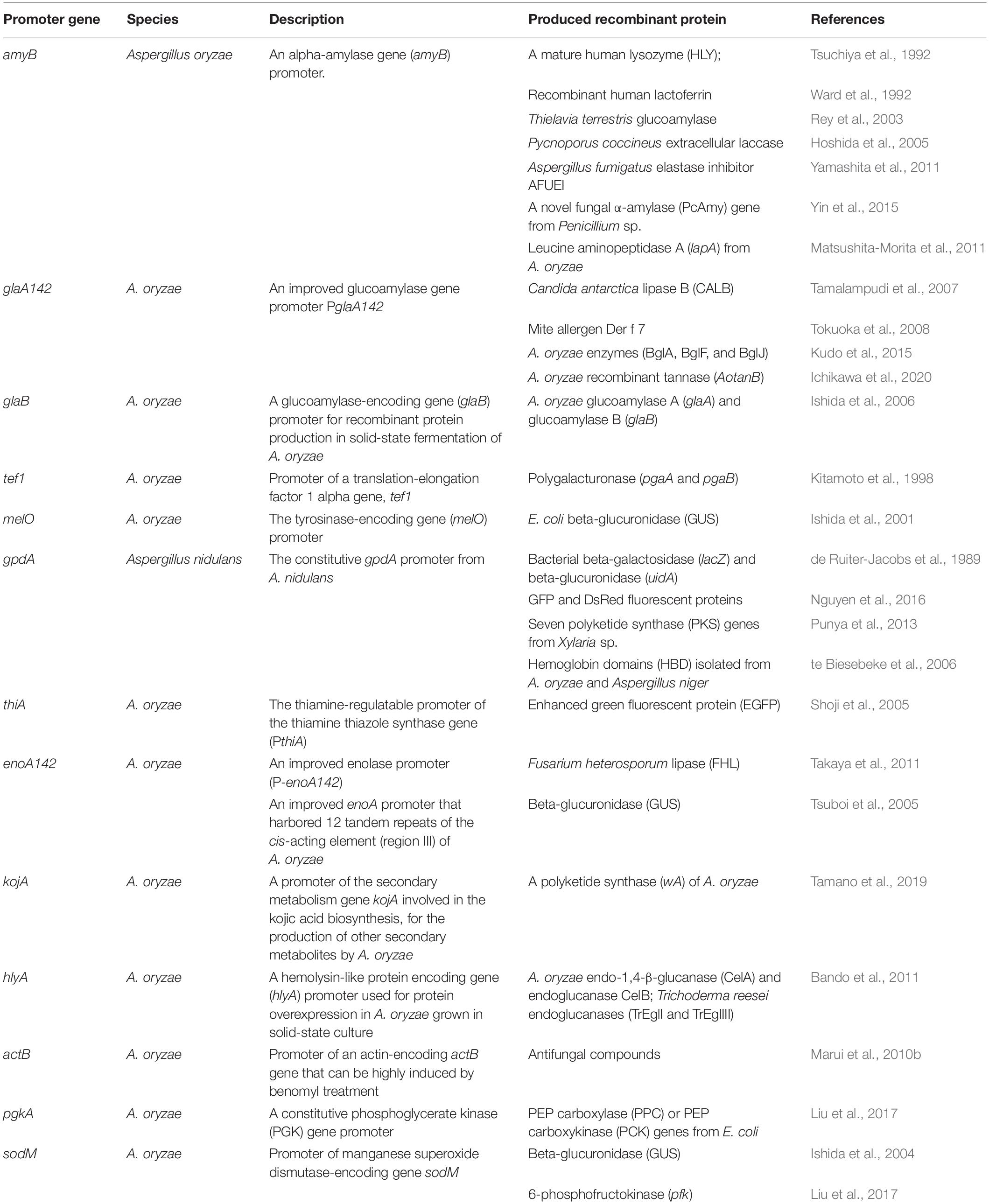
Table 2. Promoters used for the production of recombinant proteins in Aspergillus oryzae.
Random mutations caused by ultraviolet radiation is a traditional method to improve the production capacity of heterologous proteins in filamentous fungi. Through the UV radiation mutation method, an A. oryzae superproductive mutant (AUT1-8) was isolated using HLY as a screening indicator (Nemoto et al., 2009). First, HLY-hyperproducing mutants were screened from the tppA and pepE disruptants through a halo assay based on HLY activity. Then, the plasmid containing the lysozyme gene was removed from the mutants, and the resulting strains were named AUT strains. The AUT1 strain showed production levels of heterologous proteins HLY and CHY that were 2.6 and 3.2 times higher than those of the control strains, respectively (Nemoto et al., 2009). By utilizing next-generation DNA sequencing technology and comparative genomics analysis, some potential protein production-related mutation sites were identified in the A. oryzae mutant (Jin et al., 2016). Among these genes, an autA gene (AO090120000003), which was predicted to encode a cytoplasmic protein with unknown function, including an alpha/beta hydrolase fold domain, was identified for the first time to be responsible for high-level heterologous protein production in the AUT1 strain. The HLY production level of the autA mutant or deletion strains was twofold higher than that of the control strain, especially in the early growth stage of cultivation. To further improve heterologous protein production, using the generated hyperproducing mutant strain of A. oryzae, AUT1, double deletion of the Aovps10 and AosedD genes mentioned above was performed, which increased the yield of HLY and CHY by 2.1- and 1.6-fold, respectively, compared to the parental strain (Zhu et al., 2013). Thus, an excellent fungal host for the production of heterologous recombinant protein was generated by combining the mutation method with molecular breeding techniques.
Through ultraviolet radiation, Murthy and Kusumoto (2015) also obtained an A. oryzae mutant with a high acid protease yield, which was 5.6 times higher than that of the parent strain. In addition, through exposure to different doses of gamma irradiation, two maximum kojic acid overproduction mutants were attained from A. flavus HAk1 and A. oryzae HAk2, and the yield of kojic acid of the two mutants was 1.9- and 2.03-fold higher than that of their wild-type strains under malt extract sucrose culture conditions (Ammar et al., 2017).
The A. oryzae solid-state fermentation has been confirmed to produce high levels of hydrolases critical to the fermentation process. The genomic comparison and transcriptomic analysis revealed that the extracellular hydrolase genes were highly induced during solid-state cultivation in A. oryzae (Machida et al., 2008). Studies have shown that some proteinases and glycoside hydrolases have higher production level in solid-state fermentation than in submerged fermentation by A. oryzae (Zhao et al., 2019; Melnichuk et al., 2020). In addition, a glucoamylase gene glaB expressed exclusively in solid-state culture, therefore, as mentioned above, its promoter has been used for the production of recombinant proteins in solid-state fermentation of A. oryzae (Hata et al., 1998; Ishida et al., 2006). Recent studies have also shown that the transcription factor FlbC is involved in the GlaB production in solid-state culture (Tanaka et al., 2016; Gomi, 2019). These advances in molecular biology are expected to be better applied to the production of homologous and heterologous production in solid-state fermentation of A. oryzae. A transposable element known as a DNA sequence can change its relative position within the genome. According to their different mechanisms of transposition, transposable elements can be divided into two classes: retrotransposons, which replicate through an RNA intermediate and introduce into multiple loci of the genome, and DNA transposons, which transpose directly by a ‘cut-and paste’ mechanism through a DNA form (Daboussi, 1997; Daboussi and Capy, 2003). Although the A. oryzae genome sequence database (Machida et al., 2005) showed that a DNA transposon and some retrotransposons have been identified, their transposition activities have not yet been detected. In recent years, an active class II DNA transposon, crawler, has been identified in an industrial production strain of A. oryzae (OSI1013) under appropriate conditions (Ogasawara et al., 2009). In addition, in a constructed chromosomally reduced mutant of A. oryzae, the copy number of a gene (AO090005001597), which is a part of an LTR retrotransposable element, increased significantly (Jin et al., 2014). The discovery of this phenomenon is expected to improve the production capacity of heterologous recombinant proteins by constructing multicopy exogenous protein production strains in A. oryzae in the future.
As a versatile fermentative strain, A. oryzae cannot only produce a large number of amylases and proteases but also produce useful secondary metabolites; however, the research progress in this area is relatively slow. Moreover, A. oryzae, as the host for fungal secondary metabolite production, has a significant advantage in identifying the function of secondary metabolic genes compared to other filamentous fungi such as A. nidulans, in which almost no production of endogenous secondary metabolites was found. With the development of a large number of Aspergillus genome sequences in recent years, the production of some metabolites can be predicted through a comparative genomics approach, among which only a small part of the related gene clusters of secondary metabolites have been identified (Ehrlich and Mack, 2014; Takeda et al., 2014; Kjaerbolling et al., 2020). For example, genomic comparison analysis has shown that A. oryzae retains a highly intact gene cluster that produces penicillin but has very low yields. By artificially regulating the overexpression of biosynthetic genes by strong promoters, the yield of penicillin was increased by 100 times in A. oryzae (Marui et al., 2010a). In addition to penicillin, A. oryzae also produces useful secondary metabolites such as kojic acid (Marui et al., 2011; Yamada et al., 2014; Ammar et al., 2017). In addition, using the Cre/loxP-mediated marker recycling system, hyperproduction of kojic acid was achieved in A. oryzae by introducing the oxidoreductase gene kojA and transporter gene kojT (Zhang et al., 2017).
Epigenetic manipulation through the gene disruption or drug treatment [e.g., histone deacetylase inhibitors trichostatin A (TSA) and suberoylanilide hydroxamic acid (SAHA)] is one of the trend for waking up silent secondary metabolite genes cluster. It has been reported that the addition of the histone deacetylase inhibitor SAHA markedly improved the diversity of secondary metabolites in filamentous fungi (Zhu et al., 2019). In A. oryzae, through screening a gene-disruption library of transcription factors, Shinohara et al. (2016) found that the production of some secondary metabolites including astellolide F (14-deacetyl astellolide B) were significantly increased by the disruption of cclA, which encodes a component of the histone 3 lysine 4 (H3K4) methyltransferase complex of proteins associated with Set1 complex. Kawauchi et al. (2013) also showed that the fungus-specific sirtuin HstD/AoHst4 could coordinate the production of secondary metabolites by regulating the expression of laeA, which encodes a global regulator importantly involved in the regulation of fungal development and secondary metabolism.
Some studies have shown that A. oryzae can also be used as a host for heterologous expression of biosynthetic gene clusters for useful secondary metabolites, such as diterpene aphidicolin, which is a specific inhibitor of DNA polymerase α, and plant polyketide curcumin (Fujii et al., 2011; Kan et al., 2019). In particularly, a host strain with quadruple selectable markers (Jin et al., 2004b) has also been efficiently used for heterologous production of some fungal natural products (e.g., phytotoxic metabolites, basidiomycete terpenes, and plant hormones) by reconstituting these biosynthetic gene clusters in A. oryzae (Nagamine et al., 2019; Takino et al., 2019; Oikawa, 2020a,b). Moreover, the overexpression of laeA gene was also found to promote the overexpression of two clusters of heterologous biosynthetic genes from A. nidulans, resulting in the production of the corresponding metabolite, monacolin K or terrequinone A (Sakai et al., 2012). In summary, although some progress has been made in the study of secondary metabolites of Aspergillus species in recent years (Sanchez et al., 2012; Amare and Keller, 2014; He et al., 2018; Caesar et al., 2020), there are still many unknown fields awaiting further discovery.
Decoding the A. oryzae genome sequence provides abundant and reliable genetic information for a deep understanding of its genetic background and production performance. Subsequently, the establishment and rapid development of new genetic engineering technology in A. oryzae opened up more ways for its breeding of production strains and the utilization of industrial production. In recent years, although the production capacity of some heterologous proteins and secondary metabolites has been improved to some extent, it is far from the expected goal.
At present, in addition to these bottlenecks in the production of heterologous proteins mentioned above, there are still some problems that need to be further overcome, such as: (1) How to ensure high activity in the enzyme extraction process; (2) In industrial production, some macromolecular substances, such as wheat bran, rice bran, silage, corn, and soybean meal are difficult to utilize by A. oryzae. Therefore, biotechnological improvements are being made to degrade and utilize these substrates, thereby reducing production costs and pollution caused by raw material waste; (3) The large amount of heterologous protein production has a certain adverse effect on A. oryzae itself. (4) In the late growth stage of A. oryzae, the introduction of some strong promoters, such as amyB and glaA promoters, will be inhibited by high-concentration products, such as glucose. A study has shown that deletion of single and double creA/creB genes, which encode regulation factors involved in carbon catabolite repression, significantly improved amylase activity in submerged cultures containing high concentrations of inducing sugars compared to the control strain (Ichinose et al., 2014).
In addition, although the whole genome sequence has been deciphered, the function of most genes in A. oryzae is still unclear, and there remains many restrictions on its production and utilization. Therefore, there are still a large number of unknown functional genes and fields that need to be discovered and explored. Recent advances in omics techniques, such as comparative transcriptomics, proteomics and metabolomics, have opened up a new perspective for future research on functional genes and the exploration of protein secretion pathways. In summary, there remains a great space for further research and exploration on combining genetic engineering technology with A. oryzae production and application to obtain the breeding of highly efficient production strains.
F-JJ wrote the main manuscript text. SH and B-TW contributed to certain sections. LJ contributed to overall editing and proofreading. All the authors approved the submitted version.
This study was supported by the National Natural Science Foundation of China (31570107) and Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abremski, K., Wierzbicki, A., Frommer, B., and Hoess, R. H. (1986). Bacteriophage P1 Cre-loxP site-specific recombination. Site-specific DNA topoisomerase activity of the Cre recombination protein. J. Biol. Chem. 261, 391–396.
Akao, T., Sano, M., Yamada, O., Akeno, T., Fujii, K., Goto, K., et al. (2007). Analysis of expressed sequence tags from the fungus Aspergillus oryzae cultured under different conditions. DNA Res. 14, 47–57. doi: 10.1093/dnares/dsm008
Amare, M. G., and Keller, N. P. (2014). Molecular mechanisms of Aspergillus flavus secondary metabolism and development. Fungal Genet. Biol. 66, 11–18. doi: 10.1016/j.fgb.2014.02.008
Ammar, H. A. M., Ezzat, S. M., and Houseny, A. M. (2017). Improved production of kojic acid by mutagenesis of Aspergillus flavus HAk1 and Aspergillus oryzae HAk2 and their potential antioxidant activity. 3 Biotech. 7:276. doi: 10.1007/s13205-017-0905-4
Bando, H., Hisada, H., Ishida, H., Hata, Y., Katakura, Y., and Kondo, A. (2011). Isolation of a novel promoter for efficient protein expression by Aspergillus oryzae in solid-state culture. Appl. Microbiol. Biotechnol. 92, 561–569. doi: 10.1007/s00253-011-3446-5
Caesar, L. K., Kelleher, N. L., and Keller, N. P. (2020). In the fungus where it happens: history and future propelling Aspergillus nidulans as the archetype of natural products research. Fungal Genet. Biol. 144:103477. doi: 10.1016/j.fgb.2020.103477
Conesa, A., Punt, P. J., van Luijk, N., and van den Hondel, C. A. (2001). The secretion pathway in filamentous fungi: a biotechnological view. Fungal Genet. Biol. 33, 155–171. doi: 10.1006/fgbi.2001.1276
Cong, L., Ran, F. A., Cox, D., Lin, S. L., Barretto, R., Habib, N., et al. (2013). Multiplex genome engineering using CRISPR/Cas systems. Science 339, 819–823. doi: 10.1126/science.1231143
Cooper, A. A., and Stevens, T. H. (1996). Vps10p cycles between the late-Golgi and prevacuolar compartments in its function as the sorting receptor for multiple yeast vacuolar hydrolases. J. Cell Biol. 133, 529–541. doi: 10.1083/jcb.133.3.529
Daboussi, M. J., and Capy, P. (2003). Transposable elements in filamentous fungi. Annu. Rev. Microbiol. 57, 275–299. doi: 10.1146/annurev.micro.57.030502.091029
de Ruiter-Jacobs, Y. M., Broekhuijsen, M., Unkles, S. E., Campbell, E. I., Kinghorn, J. R., Contreras, R., et al. (1989). A gene transfer system based on the homologous pyrG gene and efficient expression of bacterial genes in Aspergillus oryzae. Curr. Genet. 16, 159–163. doi: 10.1007/BF00391472
DiCarlo, J. E., Norville, J. E., Mali, P., Rios, X., Aach, J., and Church, G. M. (2013). Genome engineering in Saccharomyces cerevisiae using CRISPR-Cas systems. Nucleic Acids Res. 41, 4336–4343. doi: 10.1093/nar/gkt135
Dong, L. B., Lin, X. T., Yu, D., Huang, L. G., Wang, B., and Pan, L. (2020). High-level expression of highly active and thermostable trehalase from Myceliophthora thermophila in Aspergillus niger by using the CRISPR/Cas9 tool and its application in ethanol fermentation. J. Ind. Microbiol. Biotechnol. 47, 133–144. doi: 10.1007/s10295-019-02252-9
Ehrlich, K. C., and Mack, B. M. (2014). Comparison of expression of secondary metabolite biosynthesis cluster genes in Aspergillus flavus. A. parasiticus, and A. oryzae. Toxins 6, 1916–1928. doi: 10.3390/toxins6061916
Fan, J., Zhang, Z., Long, C., He, B., Hu, Z., Jiang, C., et al. (2020). Identification and functional characterization of glycerol dehydrogenase reveal the role in kojic acid synthesis in Aspergillus oryzae. World J. Microbiol. Biotechnol. 36:136. doi: 10.1007/s11274-020-02912-4
Fleissner, A., and Dersch, P. (2010). Expression and export: recombinant protein production systems for Aspergillus. Appl. Microbiol. Biotechnol. 87, 1255–1270. doi: 10.1007/s00253-010-2672-6
Frandsen, R. J. (2011). A guide to binary vectors and strategies for targeted genome modification in fungi using Agrobacterium tumefaciens-mediated transformation. J. Microbiol. Methods 87, 247–262. doi: 10.1016/j.mimet.2011.09.004
Fujii, R., Minami, A., Tsukagoshi, T., Sato, N., Sahara, T., Ohgiya, S., et al. (2011). Total biosynthesis of diterpene aphidicolin, a specific inhibitor of DNA polymerase alpha: heterologous expression of four biosynthetic genes in Aspergillus oryzae. Biosci. Biotechnol. Biochem. 75, 1813–1817. doi: 10.1271/bbb.110366
Galagan, J. E., Calvo, S. E., Cuomo, C., Ma, L. J., Wortman, J. R., Batzoglou, S., et al. (2005). Sequencing of Aspergillus nidulans and comparative analysis with A. fumigatus and A. oryzae. Nature 438, 1105–1115. doi: 10.1038/nature04341
Gasser, B., and Mattanovich, D. (2007). Antibody production with yeasts and filamentous fungi: on the road to large scale? Biotechnol. Lett. 29, 201–212. doi: 10.1007/s10529-006-9237-x
Generoso, W. C., Gottardi, M., Oreb, M., and Boles, E. (2016). Simplified CRISPR-Cas genome editing for Saccharomyces cerevisiae. J. Microbiol. Methods 127, 203–205. doi: 10.1016/j.mimet.2016.06.020
Gomi, K. (2019). Regulatory mechanisms for amylolytic gene expression in the koji mold Aspergillus oryzae. Biosci. Biotechnol. Biochem. 83, 1385–1401. doi: 10.1080/09168451.2019.1625265
Gomi, K., Iimura, Y., and Hara, S. (1987). Integrative transformation of Aspergillus oryzae with a plasmid containing the Aspergillus nidulans argB gene. Agric. Biol. Chem. 51, 2549–2555.
Gomi, K., Kitamoto, K., and Kumagai, C. (1991). Cloning and molecular characterization of the acetamidase-encoding gene (amdS) from Aspergillus oryzae. Gene 108, 91–98. doi: 10.1016/0378-1119(91)90491-s
Gu, H., Zou, Y. R., and Rajewsky, K. (1993). Independent control of immunoglobulin switch recombination at individual switch regions evidenced through Cre-loxP-mediated gene targeting. Cell 73, 1155–1164. doi: 10.1016/0092-8674(93)90644-6
Hama, S., Tamalampudi, S., Suzuki, Y., Yoshida, A., Fukuda, H., and Kondo, A. (2008). Preparation and comparative characterization of immobilized Aspergillus oryzae expressing Fusarium heterosporum lipase for enzymatic biodiesel production. Appl. Microbiol. Biotechnol. 81, 637–645. doi: 10.1007/s00253-008-1689-6
Hara, S., Jin, F. J., Takahashi, T., and Koyama, Y. (2012). A further study on chromosome minimization by protoplast fusion in Aspergillus oryzae. Mol. Genet. Genomics 287, 177–187. doi: 10.1007/s00438-011-0669-1
Hata, Y., Ishida, H., Ichikawa, E., Kawato, A., Suginami, K., and Imayasu, S. (1998). Nucleotide sequence of an alternative glucoamylase-encoding gene (glaB) expressed in solid-state culture of Aspergillus oryzae. Gene 207, 127–134. doi: 10.1016/s0378-1119(97)00612-4
He, Y., Wang, B., Chen, W., Cox, R. J., He, J., and Chen, F. (2018). Recent advances in reconstructing microbial secondary metabolites biosynthesis in Aspergillus spp. Biotechnol. Adv. 36, 739–783. doi: 10.1016/j.biotechadv.2018.02.001
Hoang, H. D., Maruyama, J., and Kitamoto, K. (2015). Modulating endoplasmic reticulum-Golgi cargo receptors for improving secretion of carrier-fused heterologous proteins in the filamentous fungus Aspergillus oryzae. Appl. Environ. Microbiol. 81, 533–543. doi: 10.1128/AEM.02133-14
Hoshida, H., Fujita, T., Murata, K., Kubo, K., and Akada, R. (2005). Copper-dependent production of a Pycnoporus coccineus extracellular laccase in Aspergillus oryzae and Saccharomyces cerevisiae. Biosci. Biotechnol. Biochem. 69, 1090–1097. doi: 10.1271/bbb.69.1090
Huynh, H. H., Morita, N., Sakamoto, T., Katayama, T., Miyakawa, T., Tanokura, M., et al. (2020). Functional production of human antibody by the filamentous fungus Aspergillus oryzae. Fungal. Biol. Biotechnol. 7:7. doi: 10.1186/s40694-020-00098-w
Ichikawa, K., Shiono, Y., Shintani, T., Watanabe, A., Kanzaki, H., Gomi, K., et al. (2020). Efficient production of recombinant tannase in Aspergillus oryzae using an improved glucoamylase gene promoter. J. Biosci. Bioeng. 129, 150–154. doi: 10.1016/j.jbiosc.2019.08.002
Ichinose, S., Tanaka, M., Shintani, T., and Gomi, K. (2014). Improved alpha-amylase production by Aspergillus oryzae after a double deletion of genes involved in carbon catabolite repression. Appl. Microbiol. Biotechnol. 98, 335–343. doi: 10.1007/s00253-013-5353-4
Idnurm, A., Bailey, A. M., Cairns, T. C., Elliott, C. E., Foster, G. D., Ianiri, G., et al. (2017). A silver bullet in a golden age of functional genomics: the impact of Agrobacterium-mediated transformation of fungi. Fungal Biol. Biotechnol. 4:6. doi: 10.1186/s40694-017-0035-0
Ishida, H., Hata, Y., Kawato, A., and Abe, Y. (2006). Improvement of the glaB promoter expressed in solid-state fermentation (SSF) of Aspergillus oryzae. Biosci. Biotechnol. Biochem. 70, 1181–1187. doi: 10.1271/bbb.70.1181
Ishida, H., Hata, Y., Kawato, A., Abe, Y., and Kashiwagi, Y. (2004). Isolation of a novel promoter for efficient protein production in Aspergillus oryzae. Biosci. Biotechnol. Biochem. 68, 1849–1857. doi: 10.1271/bbb.68.1849
Ishida, H., Matsumura, K., Hata, Y., Kawato, A., Suginami, K., Abe, Y., et al. (2001). Establishment of a hyper-protein production system in submerged Aspergillus oryzae culture under tyrosinase-encoding gene (melO) promoter control. Appl. Microbiol. Biotechnol. 57, 131–137. doi: 10.1007/s002530100771
Jeenes, D. J., Mackenzie, D. A., Roberts, I. N., and Archer, D. B. (1991). Heterologous protein production by filamentous fungi. Biotechnol. Genet. Eng. Rev. 9, 327–367.
Jin, F. J., Hara, S., Sato, A., and Koyama, Y. (2014). Discovery and analysis of an active long terminal repeat-retrotransposable element in Aspergillus oryzae. J. Gen. Appl. Microbiol. 60, 1–6. doi: 10.2323/jgam.60.1
Jin, F. J., Katayama, T., Maruyama, J. I., and Kitamoto, K. (2016). Comparative genomic analysis identified a mutation related to enhanced heterologous protein production in the filamentous fungus Aspergillus oryzae. Appl. Microbiol. Biotechnol. 100, 9163–9174. doi: 10.1007/s00253-016-7714-2
Jin, F. J., Maruyama, J., Juvvadi, P. R., Arioka, M., and Kitamoto, K. (2004a). Adenine auxotrophic mutants of Aspergillus oryzae: development of a novel transformation system with triple auxotrophic hosts. Biosci. Biotechnol. Biochem. 68, 656–662. doi: 10.1271/bbb.68.656
Jin, F. J., Maruyama, J., Juvvadi, P. R., Arioka, M., and Kitamoto, K. (2004b). Development of a novel quadruple auxotrophic host transformation system by argB gene disruption using adeA gene and exploiting adenine auxotrophy in Aspergillus oryzae. FEMS Microbiol. Lett. 239, 79–85. doi: 10.1016/j.femsle.2004.08.025
Jin, F. J., Nishida, M., Hara, S., and Koyama, Y. (2011a). Identification and characterization of a putative basonidiophore development in Aspergillus oryzae. Fungal Genet. Biol. 48, 1108–1115. doi: 10.1016/j.fgb.2011.10.001
Jin, F. J., Takahashi, T., Matsushima, K., Hara, S., Shinohara, Y., Maruyama, J., et al. (2011b). SclR, a basic helix-loop-helix transcription factor, regulates hyphal morphology and promotes sclerotial formation in Aspergillus oryzae. Eukaryot Cell 10, 945–955. doi: 10.1128/EC.00013-11
Jin, F. J., Takahashi, T., Machida, M., and Koyama, Y. (2009). Identification of a basic helix-loop-helix-type transcription regulator gene in Aspergillus oryzae by systematically deleting large chromosomal segments. Appl. Environ. Microbiol. 75, 5943–5951. doi: 10.1128/AEM.00975-09
Jin, F. J., Takahashi, T., Utsushikawa, M., Furukido, T., Nishida, M., Ogawa, M., et al. (2010). A trial of minimization of chromosome 7 in Aspergillus oryzae by multiple chromosomal deletions. Mol. Genet. Genomics 283, 1–12. doi: 10.1007/s00438-009-0494-y
Jin, F. J., Watanabe, T., Juvvadi, P. R., Maruyama, J., Arioka, M., and Kitamoto, K. (2007). Double disruption of the proteinase genes, tppA and pepE, increases the production level of human lysozyme by Aspergillus oryzae. Appl. Microbiol. Biotechnol. 76, 1059–1068. doi: 10.1007/s00253-007-1088-4
Jinek, M., Chylinski, K., Fonfara, I., Hauer, M., Doudna, J. A., and Charpentier, E. (2012). A programmable Dual-RNA-Guided DNA endonuclease in adaptive bacterial immunity. Science 337, 816–821. doi: 10.1126/science.1225829
Kan, E., Katsuyama, Y., Maruyama, J. I., Tamano, K., Koyama, Y., and Ohnishi, Y. (2019). Production of the plant polyketide curcumin in Aspergillus oryzae: strengthening malonyl-CoA supply for yield improvement. Biosci. Biotechnol. Biochem. 83, 1372–1381. doi: 10.1080/09168451.2019.1606699
Katayama, T., Nakamura, H., Zhang, Y., Pascal, A., Fujii, W., and Maruyama, J. I. (2019). Forced recycling of an AMA1-based genome-editing plasmid allows for efficient multiple gene deletion/integration in the industrial filamentous fungus Aspergillus oryzae. Appl. Environ. Microbiol. 85:e01896-18. doi: 10.1128/AEM.01896-18
Katayama, T., Tanaka, Y., Okabe, T., Nakamura, H., Fujii, W., Kitamoto, K., et al. (2016). Development of a genome editing technique using the CRISPR/Cas9 system in the industrial filamentous fungus Aspergillus oryzae. Biotechnol. Lett. 38, 637–642. doi: 10.1007/s10529-015-2015-x
Kawauchi, M., Nishiura, M., and Iwashita, K. (2013). Fungus-specific sirtuin HstD coordinates secondary metabolism and development through control of LaeA. Eukaryotic Cell 12, 1087–1096. doi: 10.1128/Ec.00003-13
Kimura, S., Maruyama, J., Kikuma, T., Arioka, M., and Kitamoto, K. (2011). Autophagy delivers misfolded secretory proteins accumulated in endoplasmic reticulum to vacuoles in the filamentous fungus Aspergillus oryzae. Biochem. Biophys. Res. Commun. 406, 464–470. doi: 10.1016/j.bbrc.2011.02.075
Kitamoto, K. (2002). Molecular biology of the Koji molds. Adv. Appl. Microbiol. 51, 129–153. doi: 10.1016/s0065-2164(02)51004-2
Kitamoto, K. (2015). Cell biology of the Koji mold Aspergillus oryzae. Biosci. Biotechnol. Biochem. 79, 863–869. doi: 10.1080/09168451.2015.1023249
Kitamoto, N., Matsui, J., Kawai, Y., Kato, A., Yoshino, S., Ohmiya, K., et al. (1998). Utilization of the TEF1-alpha gene (TEF1) promoter for expression of polygalacturonase genes, pgaA and pgaB, in Aspergillus oryzae. Appl. Microbiol. Biotechnol. 50, 85–92. doi: 10.1007/s002530051260
Kjaerbolling, I., Vesth, T., Frisvad, J. C., Nybo, J. L., Theobald, S., Kildgaard, S., et al. (2020). A comparative genomics study of 23 Aspergillus species from section Flavi. Nat. Commun. 11:1106. doi: 10.1038/s41467-019-14051-y
Kobayashi, T., Abe, K., Asai, K., Gomi, K., Juvvadi, P. R., Kato, M., et al. (2007). Genomics of Aspergillus oryzae. Biosci. Biotechnol. Biochem. 71, 646–670. doi: 10.1271/bbb.60550
Koda, A., Bogaki, T., Minetoki, T., and Hirotsune, M. (2006). 5′ Untranslated region of the Hsp12 gene contributes to efficient translation in Aspergillus oryzae. Appl. Microbiol, Biotechnol. 70, 333–336. doi: 10.1007/s00253-005-0083-x
Koda, A., Minetoki, T., Ozeki, K., and Hirotsune, M. (2004). Translation efficiency mediated by the 5′ untranslated region greatly affects protein production in Aspergillus oryzae. Appl. Microbiol. Biotechnol. 66, 291–296. doi: 10.1007/s00253-004-1681-8
Kubodera, T., Yamashita, N., and Nishimura, A. (2000). Pyrithiamine resistance gene (ptrA) of Aspergillus oryzae: cloning, characterization and application as a dominant selectable marker for transformation. Biosci. Biotechnol. Biochem. 64, 1416–1421. doi: 10.1271/bbb.64.1416
Kudo, K., Watanabe, A., Ujiie, S., Shintani, T., and Gomi, K. (2015). Purification and enzymatic characterization of secretory glycoside hydrolase family 3 (GH3) aryl beta-glucosidases screened from Aspergillus oryzae genome. J. Biosci. Bioeng. 120, 614–623. doi: 10.1016/j.jbiosc.2015.03.019
Lemmon, S. K., and Traub, L. M. (2000). Sorting in the endosomal system in yeast and animal cells. Curr. Opin. Cell Biol. 12, 457–466. doi: 10.1016/s0955-0674(00)00117-4
Lissau, B. G., Pedersen, P. B., Petersen, B. R., and Budolfsen, G. (1998). Safety evaluation of a fungal pectinesterase enzyme preparation and its use in food. Food Addit. Contam. 15, 627–636. doi: 10.1080/02652039809374691
Liu, C., Minami, A., Ozaki, T., Wu, J., Kawagishi, H., Maruyama, J. I., et al. (2019). Efficient reconstitution of basidiomycota diterpene erinacine gene cluster in ascomycota host Aspergillus oryzae Based on Genomic DNA sequences. J. Am. Chem. Soc. 141, 15519–15523. doi: 10.1021/jacs.9b08935
Liu, J., Xie, Z., Shin, H. D., Li, J., Du, G., Chen, J., et al. (2017). Rewiring the reductive tricarboxylic acid pathway and L-malate transport pathway of Aspergillus oryzae for overproduction of L-malate. J. Biotechnol. 253, 1–9. doi: 10.1016/j.jbiotec.2017.05.011
Liu, R., Chen, L., Jiang, Y., Zhou, Z., and Zou, G. (2015). Efficient genome editing in filamentous fungus Trichoderma reesei using the CRISPR/Cas9 system. Cell Discov. 1:15007. doi: 10.1038/celldisc.2015.7
Liu, Z., and Friesen, T. L. (2012). Polyethylene glycol (PEG)-mediated transformation in filamentous fungal pathogens. Methods Mol. Biol. 835, 365–375. doi: 10.1007/978-1-61779-501-5_21
Lombrana, M., Moralejo, F. J., Pinto, R., and Martin, J. F. (2004). Modulation of Aspergillus awamori thaumatin secretion by modification of bipA gene expression. Appl. Environ. Microbiol. 70, 5145–5152. doi: 10.1128/AEM.70.9.5145-5152.2004
Machida, M. (2002). Progress of Aspergillus oryzae genomics. Adv. Appl. Microbiol. 51, 81–106. doi: 10.1016/s0065-2164(02)51002-9
Machida, M., Asai, K., Sano, M., Tanaka, T., Kumagai, T., Terai, G., et al. (2005). Genome sequencing and analysis of Aspergillus oryzae. Nature 438, 1157–1161. doi: 10.1038/nature04300
Machida, M., Yamada, O., and Gomi, K. (2008). Genomics of Aspergillus oryzae: learning from the history of Koji mold and exploration of its future. DNA Res. 15, 173–183. doi: 10.1093/dnares/dsn020
Marcusson, E. G., Horazdovsky, B. F., Cereghino, J. L., Gharakhanian, E., and Emr, S. D. (1994). The sorting receptor for yeast vacuolar carboxypeptidase Y is encoded by the VPS10 gene. Cell 77, 579–586. doi: 10.1016/0092-8674(94)90219-4
Marui, J., Ohashi-Kunihiro, S., Ando, T., Nishimura, M., Koike, H., and Machida, M. (2010a). Penicillin biosynthesis in Aspergillus oryzae and its overproduction by genetic engineering. J. Biosci. Bioeng. 110, 8–11. doi: 10.1016/j.jbiosc.2010.01.001
Marui, J., Yoshimi, A., Hagiwara, D., Fujii-Watanabe, Y., Oda, K., Koike, H., et al. (2010b). Use of the Aspergillus oryzae actin gene promoter in a novel reporter system for exploring antifungal compounds and their target genes. Appl. Microbiol. Biotechnol. 87, 1829–1840. doi: 10.1007/s00253-010-2627-y
Marui, J., Yamane, N., Ohashi-Kunihiro, S., Ando, T., Terabayashi, Y., Sano, M., et al. (2011). Kojic acid biosynthesis in Aspergillus oryzae is regulated by a Zn(II)(2)Cys(6) transcriptional activator and induced by kojic acid at the transcriptional level. J. Biosci. Bioeng. 112, 40–43. doi: 10.1016/j.jbiosc.2011.03.010
Maruyama, J., and Kitamoto, K. (2008). Multiple gene disruptions by marker recycling with highly efficient gene-targeting background (DeltaligD) in Aspergillus oryzae. Biotechnol. Lett. 30, 1811–1817. doi: 10.1007/s10529-008-9763-9
Matsushita-Morita, M., Tada, S., Suzuki, S., Hattori, R., Marui, J., Furukawa, I., et al. (2011). Overexpression and characterization of an extracellular leucine aminopeptidase from Aspergillus oryzae. Curr. Microbiol. 62, 557–564. doi: 10.1007/s00284-010-9744-9
Matsu-Ura, T., Baek, M., Kwon, J., and Hong, C. (2015). Efficient gene editing in Neurospora crassa with CRISPR technology. Fungal Biol. Biotechnol. 2:4. doi: 10.1186/s40694-015-0015-1
Mattern, I. E., Unkles, S., Kinghorn, J. R., Pouwels, P. H., and van den Hondel, C. A. (1987). Transformation of Aspergillus oryzae using the A. niger pyrG gene. Mol. Gen. Genet. 210, 460–461. doi: 10.1007/BF00327197
Melnichuk, N., Braia, M. J., Anselmi, P. A., Meini, M. R., and Romanini, D. (2020). Valorization of two agroindustrial wastes to produce alpha-amylase enzyme from Aspergillus oryzae by solid-state fermentation. Waste Manag. 106, 155–161. doi: 10.1016/j.wasman.2020.03.025
Merz, M., Eisele, T., Berends, P., Appel, D., Rabe, S., Blank, I., et al. (2015). Flavourzyme, an enzyme preparation with industrial relevance: automated nine-step purification and partial characterization of eight enzymes. J. Agric. Food Chem. 63, 5682–5693. doi: 10.1021/acs.jafc.5b01665
Michielse, C. B., Arentshorst, M., Ram, A. F., and van den Hondel, C. A. (2005). Agrobacterium-mediated transformation leads to improved gene replacement efficiency in Aspergillus awamori. Fungal Genet. Biol. 42, 9–19. doi: 10.1016/j.fgb.2004.06.009
Mizutani, O., Arazoe, T., Toshida, K., Hayashi, R., Ohsato, S., Sakuma, T., et al. (2017). Detailed analysis of targeted gene mutations caused by the Platinum-Fungal TALENs in Aspergillus oryzae RIB40 strain and a ligD disruptant. J. Biosci. Bioeng, 123, 287–293. doi: 10.1016/j.jbiosc.2016.09.014
Mizutani, O., Kudo, Y., Saito, A., Matsuura, T., Inoue, H., Abe, K., et al. (2008). A defect of LigD (human Lig4 homolog) for nonhomologous end joining significantly improves efficiency of gene-targeting in Aspergillus oryzae. Fungal Genet. Biol. 45, 878–889. doi: 10.1016/j.fgb.2007.12.010
Mizutani, O., Masaki, K., Gomi, K., and Iefuji, H. (2012). Modified Cre-loxP recombination in Aspergillus oryzae by direct introduction of Cre recombinase for marker gene rescue. Appl. Environ. Microbiol. 78, 4126–4133. doi: 10.1128/AEM.00080-12
Moralejo, F. J., Watson, A. J., Jeenes, D. J., Archer, D. B., and Martin, J. F. (2001). A defined level of protein disulfide isomerase expression is required for optimal secretion of thaumatin by Aspegillus awamori. Mol. Genet. Genomics 266, 246–253. doi: 10.1007/s004380100550
Morishita, Y., Sonohara, T., Taniguchi, T., Adachi, K., Fujita, M., and Asai, T. (2020). Synthetic-biology-based discovery of a fungal macrolide from Macrophomina phaseolina. Org. Biomol. Chem. 18, 2813–2816. doi: 10.1039/d0ob00519c
Mulder, H. J., Nikolaev, I., and Madrid, S. M. (2006). HACA, the transcriptional activator of the unfolded protein response (UPR) in Aspergillus niger, binds to partly palindromic UPR elements of the consensus sequence 5′-CAN(G/A)NTGT/GCCT-3’. Fungal Genet. Biol. 43, 560–572. doi: 10.1016/j.fgb.2006.02.005
Murthy, P. S., and Kusumoto, K. I. (2015). Acid protease production by Aspergillus oryzae on potato pulp powder with emphasis on glycine releasing activity: a benefit to the food industry. Food Bioprod. Process. 96, 180–188. doi: 10.1016/j.fbp.2015.07.013
Nagamine, S., Liu, C. W., Nishishita, J., Kozaki, T., Sogahata, K., Sato, Y., et al. (2019). Ascomycete Aspergillus oryzae is an efficient expression host for production of basidiomycete terpenes by using genomic DNA sequences. Appl. Environ. Microbiol. 85:e00409-19. doi: 10.1128/AEM.00409-19
Nakamura, H., Katayama, T., Okabe, T., Iwashita, K., Fujii, W., Kitamoto, K., et al. (2017). Highly efficient gene targeting in Aspergillus oryzae industrial strains under ligD mutation introduced by genome editing: strain-specific differences in the effects of deleting EcdR, the negative regulator of sclerotia formation. J. Gen. Appl. Microbiol. 63, 172–178. doi: 10.2323/jgam.2016.10.002
Nemoto, T., Watanabe, T., Mizogami, Y., Maruyama, J., and Kitamoto, K. (2009). Isolation of Aspergillus oryzae mutants for heterologous protein production from a double proteinase gene disruptant. Appl. Microbiol. Biotechnol. 82, 1105–1114. doi: 10.1007/s00253-008-1851-1
Nguyen, K. T., Ho, Q. N., Do, L. T. B. X., Mai, L. T. D., Pham, D. N., Tran, H. T. T., et al. (2017). A new and efficient approach for construction of uridine/uracil auxotrophic mutants in the filamentous fungus Aspergillus oryzae using Agrobacterium tumefaciens-mediated transformation. World J. Microbiol. Biotechnol. 33:107. doi: 10.1007/s11274-017-2275-9
Nguyen, K. T., Ho, Q. N., Pham, T. H., Phan, T. N., and Tran, V. T. (2016). The construction and use of versatile binary vectors carrying pyrG auxotrophic marker and fluorescent reporter genes for Agrobacterium-mediated transformation of Aspergillus oryzae. World J. Microbiol. Biotechnol. 32:204. doi: 10.1007/s11274-016-2168-3
Nielsen, M. L., Albertsen, L., Lettier, G., Nielsen, J. B., and Mortensen, U. H. (2006). Efficient PCR-based gene targeting with a recyclable marker for Aspergillus nidulans. Fungal Genet. Biol. 43, 54–64. doi: 10.1016/j.fgb.2005.09.005
Nierman, W. C., Pain, A., Anderson, M. J., Wortman, J. R., Kim, H. S., Arroyo, J., et al. (2005). Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus fumigatus. Nature 438, 1151–1156. doi: 10.1038/nature04332
Nodvig, C. S., Hoof, J. B., Kogle, M. E., Jarczynska, Z. D., Lehmbeck, J., Klitgaard, D. K., et al. (2018). Efficient oligo nucleotide mediated CRISPR-Cas9 gene editing in Aspergilli. Fungal Genet. Biol. 115, 78–89. doi: 10.1016/j.fgb.2018.01.004
Ogasawara, H., Obata, H., Hata, Y., Takahashi, S., and Gomi, K. (2009). Crawler, a novel Tc1/mariner-type transposable element in Aspergillus oryzae transposes under stress conditions. Fungal Genet. Biol. 46, 441–449. doi: 10.1016/j.fgb.2009.02.007
Ohno, A., Maruyama, J., Nemoto, T., Arioka, M., and Kitamoto, K. (2011). A carrier fusion significantly induces unfolded protein response in heterologous protein production by Aspergillus oryzae. Appl. Microbiol. Biotechnol. 92, 1197–1206. doi: 10.1007/s00253-011-3487-9
Oikawa, H. (2020a). Heterologous production of fungal natural products: reconstitution of biosynthetic gene clusters in model host Aspergillus oryzae. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 96, 420–430. doi: 10.2183/pjab.96.031
Oikawa, H. (2020b). Reconstitution of biosynthetic machinery of fungal natural products in heterologous hosts. Biosci. Biotechnol. Biochem, 84, 433–444. doi: 10.1080/09168451.2019.1690976
Punt, P. J., van Gemeren, I. A., Drint-Kuijvenhoven, J., Hessing, J. G., van Muijlwijk-Harteveld, G. M., Beijersbergen, A., et al. (1998). Analysis of the role of the gene bipA, encoding the major endoplasmic reticulum chaperone protein in the secretion of homologous and heterologous proteins in black Aspergilli. Appl. Microbiol. Biotechnol. 50, 447–454. doi: 10.1007/s002530051319
Punya, J., Tachaleat, A., Wattanachaisaereekul, S., Haritakun, R., Boonlarppradab, C., and Cheevadhanarak, S. (2013). Functional expression of a foreign gene in Aspergillus oryzae producing new pyrone compounds. Fungal Genet. Biol. 50, 55–62. doi: 10.1016/j.fgb.2012.10.005
Reggiori, F., and Klionsky, D. J. (2002). Autophagy in the eukaryotic cell. Eukaryot Cell 1, 11–21. doi: 10.1128/ec.01.1.11-21.2002
Rey, M. W., Brown, K. M., Golightly, E. J., Fuglsang, C. C., Nielsen, B. R., Hendriksen, H. V., et al. (2003). Cloning, heterologous expression, and characterization of Thielavia terrestris glucoamylase. Appl. Biochem. Biotechnol. 111, 153–166. doi: 10.1385/abab:111:3:153
Rojas-Sanchez, U., Lopez-Calleja, A. C., Millan-Chiu, B. E., Fernandez, F., Loske, A. M., and Gomez-Lim, M. A. (2020). Enhancing the yield of human erythropoietin in Aspergillus niger by introns and CRISPR-Cas9. Protein Exp. Purif. 168:105570. doi: 10.1016/j.pep.2020.105570
Ron, D., and Walter, P. (2007). Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 8, 519–529. doi: 10.1038/nrm2199
Sakai, K., Kinoshita, H., and Nihira, T. (2012). Heterologous expression system in Aspergillus oryzae for fungal biosynthetic gene clusters of secondary metabolites. Appl. Microbiol. Biotechnol. 93, 2011–2022. doi: 10.1007/s00253-011-3657-9
Sanchez, J. F., Somoza, A. D., Keller, N. P., and Wang, C. C. (2012). Advances in Aspergillus secondary metabolite research in the post-genomic era. Nat. Prod. Rep. 29, 351–371. doi: 10.1039/c2np00084a
Sauer, B. (1987). Functional expression of the cre-lox site-specific recombination system in the yeast Saccharomyces cerevisiae. Mol. Cell Biol. 7, 2087–2096. doi: 10.1128/mcb.7.6.2087
Schiml, S., and Puchta, H. (2016). Revolutionizing plant biology: multiple ways of genome engineering by CRISPR/Cas. Plant Methods 12:8. doi: 10.1186/s13007-016-0103-0
Shima, Y., Ito, Y., Kaneko, S., Hatabayashi, H., Watanabe, Y., Adachi, Y., et al. (2009). Identification of three mutant loci conferring carboxin-resistance and development of a novel transformation system in Aspergillus oryzae. Fungal Genet. Biol. 46, 67–76. doi: 10.1016/j.fgb.2008.10.005
Shinohara, Y., Kawatani, M., Futamura, Y., Osada, H., and Koyama, Y. (2016). An overproduction of astellolides induced by genetic disruption of chromatin-remodeling factors in Aspergillus oryzae. J. Antibiot. 69, 4–8. doi: 10.1038/ja.2015.73
Shmakov, S., Smargon, A., Scott, D., Cox, D., Pyzocha, N., Yan, W., et al. (2017). Diversity and evolution of class 2 CRISPR-Cas systems. Nat. Rev. Microbiol. 15, 169–182. doi: 10.1038/nrmicro.2016.184
Shoji, J. Y., Maruyama, J., Arioka, M., and Kitamoto, K. (2005). Development of Aspergillus oryzae thiA promoter as a tool for molecular biological studies. FEMS Microbiol. Lett. 244, 41–46. doi: 10.1016/j.femsle.2005.01.014
Sieburth, L. E., Drews, G. N., and Meyerowitz, E. M. (1998). Non-autonomy of AGAMOUS function in flower development: use of a Cre/loxP method for mosaic analysis in Arabidopsis. Development 125, 4303–4312.
Song, R., Zhai, Q., Sun, L., Huang, E., Zhang, Y., Zhu, Y., et al. (2019). CRISPR/Cas9 genome editing technology in filamentous fungi: progress and perspective. Appl. Microbiol. Biotechnol. 103, 6919–6932. doi: 10.1007/s00253-019-10007-w
Sun, Y. L., Niu, Y. L., He, B., Ma, L., Li, G. H., Tran, V. T., et al. (2019). Dual selection marker transformation system using Agrobacterium tumefaciens for the industrial Aspergillus oryzae 3.042. J. Microbiol. Biotechnol. 29, 230–234. doi: 10.4014/jmb.1811.11027
Suzuki, S., Tada, S., Fukuoka, M., Taketani, H., Tsukakoshi, Y., Matsushita, M., et al. (2009). A novel transformation system using a bleomycin resistance marker with chemosensitizers for Aspergillus oryzae. Biochem. Biophys. Res. Commun. 383, 42–47. doi: 10.1016/j.bbrc.2009.03.108
Suzuki, T., Oiso, N., Gautam, R., Novak, E. K., Panthier, J. J., Suprabha, P. G., et al. (2003). The mouse organellar biogenesis mutant buff results from a mutation in Vps33a, a homologue of yeast vps33 and Drosophila carnation. Proc. Natl. Acad. Sci. U.S.A. 100, 1146–1150. doi: 10.1073/pnas.0237292100
Takahashi, T., Jin, F. J., and Koyama, Y. (2009). Nonhomologous end-joining deficiency allows large chromosomal deletions to be produced by replacement-type recombination in Aspergillus oryzae. Fungal Genet. Biol. 46, 815–824. doi: 10.1016/j.fgb.2009.07.009
Takahashi, T., Jin, F. J., Sunagawa, M., Machida, M., and Koyama, Y. (2008). Generation of large chromosomal deletions in koji molds Aspergillus oryzae and Aspergillus sojae via a loop-out recombination. Appl. Environ. Microbiol. 74, 7684–7693. doi: 10.1128/AEM.00692-08
Takahashi, T., Masuda, T., and Koyama, Y. (2006). Enhanced gene targeting frequency in ku70 and ku80 disruption mutants of Aspergillus sojae and Aspergillus oryzae. Mol. Genet. Genomics 275, 460–470. doi: 10.1007/s00438-006-0104-1
Takaya, T., Koda, R., Adachi, D., Nakashima, K., Wada, J., Bogaki, T., et al. (2011). Highly efficient biodiesel production by a whole-cell biocatalyst employing a system with high lipase expression in Aspergillus oryzae. Appl. Microbiol. Biotechnol. 90, 1171–1177. doi: 10.1007/s00253-011-3186-6
Takeda, I., Umemura, M., Koike, H., Asai, K., and Machida, M. (2014). Motif-independent prediction of a secondary metabolism gene cluster using comparative genomics: application to sequenced genomes of Aspergillus and ten other filamentous fungal species. DNA Res. 21, 447–457. doi: 10.1093/dnares/dsu010
Takino, J., Kozaki, T., Ozaki, T., Liu, C., Minami, A., and Oikawa, H. (2019). Elucidation of biosynthetic pathway of a plant hormone abscisic acid in phytopathogenic fungi. Biosci. Biotechnol. Biochem. 83, 1642–1649. doi: 10.1080/09168451.2019.1618700
Tamalampudi, S., Talukder, M. M., Hama, S., Tanino, T., Suzuki, Y., Kondo, A., et al. (2007). Development of recombinant Aspergillus oryzae whole-cell biocatalyst expressing lipase-encoding gene from Candida antarctica. Appl. Microbiol/Biotechnol. 75, 387–395. doi: 10.1007/s00253-006-0814-7
Tamano, K., Kuninaga, M., Kojima, N., Umemura, M., Machida, M., and Koike, H. (2019). Use of the kojA promoter, involved in kojic acid biosynthesis, for polyketide production in Aspergillus oryzae: implications for long-term production. BMC Biotechnol. 19:70. doi: 10.1186/s12896-019-0567-x
Tanaka, M., Tokuoka, M., Shintani, T., and Gomi, K. (2012). Transcripts of a heterologous gene encoding mite allergen Der f 7 are stabilized by codon optimization in Aspergillus oryzae. Appl. Microbiol. Biotechnol. 96, 1275–1282. doi: 10.1007/s00253-012-4169-y
Tanaka, M., Yoshimura, M., Ogawa, M., Koyama, Y., Shintani, T., and Gomi, K. (2016). The C2H2-type transcription factor, FlbC, is involved in the transcriptional regulation of Aspergillus oryzae glucoamylase and protease genes specifically expressed in solid-state culture. Appl. Microbiol. Biotechnol. 100, 5859–5868. doi: 10.1007/s00253-016-7419-6
te Biesebeke, R., Boussier, A., van Biezen, N., Braaksma, M., van den Hondel, C. A., de Vos, W. M., et al. (2006). Expression of Aspergillus hemoglobin domain activities in Aspergillus oryzae grown on solid substrates improves growth rate and enzyme production. Biotechnol. J. 1, 822–827. doi: 10.1002/biot.200600036
Todokoro, T., Bando, H., Kotaka, A., Tsutsumi, H., Hata, Y., and Ishida, H. (2020). Identification of a novel pyrithiamine resistance marker gene thiI for genome co-editing in Aspergillus oryzae. J. Biosci. Bioeng. 130, 227–232. doi: 10.1016/j.jbiosc.2020.04.013
Tokuoka, M., Tanaka, M., Ono, K., Takagi, S., Shintani, T., and Gomi, K. (2008). Codon optimization increases steady-state mRNA levels in Aspergillus oryzae heterologous gene expression. Appl. Environ. Microbiol. 74, 6538–6546. doi: 10.1128/AEM.01354-08
Tsuboi, H., Koda, A., Toda, T., Minetoki, T., Hirotsune, M., and Machida, M. (2005). Improvement of the Aspergillus oryzae enolase promoter (P-enoA) by the introduction of cis-element repeats. Biosci. Biotechnol. Biochem. 69, 206–208. doi: 10.1271/bbb.69.206
Tsuchiya, K., Nagashima, T., Yamamoto, Y., Gomi, K., Kitamoto, K., Kumagai, C., et al. (1994). High level secretion of calf chymosin using a glucoamylase-prochymosin fusion gene in Aspergillus oryzae. Biosci. Biotechnol. Biochem. 58, 895–899. doi: 10.1271/bbb.58.895
Tsuchiya, K., Tada, S., Gomi, K., Kitamoto, K., Kumagai, C., Jigami, Y., et al. (1992). High level expression of the synthetic human lysozyme gene in Aspergillus oryzae. Appl. Microbiol. Biotechnol. 38, 109–114. doi: 10.1007/BF00169428
Unkles, S. E., Campbell, E. I., de Ruiter-Jacobs, Y. M. J. T., Broekhuijsen, M., Marco, J. A., Carrez, D., et al. (1989). The development of a homologous transformation system for Aspergillus oryzae based on the nitrate assimilation pathway: a convenient and general selection system for ?lamentous fungal transformation. Mol. Gen. Genet. 218, 99–104.
van Erp, P. B. G., Bloomer, G., Wilkinson, R., and Wiedenheft, B. (2015). The history and market impact of CRISPR RNA-guided nucleases. Curr. Opin. Virol. 12, 85–90. doi: 10.1016/j.coviro.2015.03.011
Vongsangnak, W., Olsen, P., Hansen, K., Krogsgaard, S., and Nielsen, J. (2008). Improved annotation through genome-scale metabolic modeling of Aspergillus oryzae. BMC Genomics 9:245. doi: 10.1186/1471-2164-9-245
Wada, R., Jin, F. J., Koyama, Y., Maruyama, J., and Kitamoto, K. (2014). Efficient formation of heterokaryotic sclerotia in the filamentous fungus Aspergillus oryzae. Appl. Microbiol. Biotechnol. 98, 325–334. doi: 10.1007/s00253-013-5314-y
Wang, H., Entwistle, J., Morlon, E., Archer, D. B., Peberdy, J. F., Ward, M., et al. (2003). Isolation and characterisation of a calnexin homologue, clxA, from Aspergillus niger. Mol. Genet. Genomics 268, 684–691. doi: 10.1007/s00438-002-0790-2
Wang, H., and Ward, M. (2000). Molecular characterization of a PDI-related gene prpA in Aspergillus niger var. awamori. Curr. Genet. 37, 57–64. doi: 10.1007/s002940050009
Ward, P. P., Lo, J. Y., Duke, M., May, G. S., Headon, D. R., and Conneely, O. M. (1992). Production of biologically active recombinant human lactoferrin in Aspergillus oryzae. Biotechnology 10, 784–789. doi: 10.1038/nbt0792-784
Weyda, I., Yang, L., Vang, J., Ahring, B. K., Lubeck, M., and Lubeck, P. S. (2017). A comparison of Agrobacterium-mediated transformation and protoplast-mediated transformation with CRISPR-Cas9 and bipartite gene targeting substrates, as effective gene targeting tools for Aspergillus carbonarius. J. Microbiol. Methods 135, 26–34. doi: 10.1016/j.mimet.2017.01.015
Yamada, O., Lee, B. R., and Gomi, K. (1997). Transformation system for Aspergillus oryzae with double auxotrophic mutations, niaD and sC. Biosci. Biotechnol. Biochem, 61, 1367–1369.
Yamada, R., Yoshie, T., Wakai, S., Asai-Nakashima, N., Okazaki, F., Ogino, C., et al. (2014). Aspergillus oryzae-based cell factory for direct kojic acid production from cellulose. Micro.b Cell Fact. 13:71. doi: 10.1186/1475-2859-13-71
Yamashita, N., Komori, Y., Okumura, Y., Uchiya, K., Matsui, T., Nishimura, A., et al. (2011). High-yields heterologous production of the novel Aspergillus fumigatus elastase inhibitor AFUEI in Aspergillus oryzae. J. Biosci. Bioeng. 112, 114–117. doi: 10.1016/j.jbiosc.2011.03.020
Yin, Y., Mao, Y., Yin, X., Gao, B., and Wei, D. (2015). Construction of a shuttle vector for heterologous expression of a novel fungal alpha-amylase gene in Aspergillus oryzae. J. Microbiol. Biotechnol. 25, 988–998. doi: 10.4014/jmb.1410.10022
Yokota, J. I., Shiro, D., Tanaka, M., Onozaki, Y., Mizutani, O., Kakizono, D., et al. (2017). Cellular responses to the expression of unstable secretory proteins in the filamentous fungus Aspergillus oryzae. Appl. Microbiol. Biotechnol. 101, 2437–2446. doi: 10.1007/s00253-016-8086-3
Yoon, J., Aishan, T., Maruyama, J., and Kitamoto, K. (2010). Enhanced production and secretion of heterologous proteins by the filamentous fungus Aspergillus oryzae via disruption of vacuolar protein sorting receptor gene Aovps10. Appl. Environ. Microbiol. 76, 5718–5727. doi: 10.1128/AEM.03087-09
Yoon, J., Kimura, S., Maruyama, J., and Kitamoto, K. (2009). Construction of quintuple protease gene disruptant for heterologous protein production in Aspergillus oryzae. Appl. Microbiol. Biotechnol. 82, 691–701. doi: 10.1007/s00253-008-1815-5
Yoon, J., Kikuma, T., Maruyama, J., and Kitamoto, K. (2013). Enhanced production of bovine chymosin by autophagy deficiency in the filamentous fungus Aspergillus oryzae. PLoS One 8:e62512. doi: 10.1371/journal.pone.0062512
Yoon, J., Maruyama, J., and Kitamoto, K. (2011). Disruption of ten protease genes in the filamentous fungus Aspergillus oryzae highly improves production of heterologous proteins. Appl. Microbiol. Biotechnol, 89, 747–759. doi: 10.1007/s00253-010-2937-0
Zhang, S., Ban, A., Ebara, N., Mizutani, O., Tanaka, M., Shintani, T., et al. (2017). Self-excising Cre/mutant lox marker recycling system for multiple gene integrations and consecutive gene deletions in Aspergillus oryzae. J. Biosci. Bioeng. 123, 403–411. doi: 10.1016/j.jbiosc.2016.11.001
Zhao, G., Ding, L. L., Pan, Z. H., Kong, D. H., Hadiatullah, H., and Fan, Z. C. (2019). Proteinase and glycoside hydrolase production is enhanced in solid-state fermentation by manipulating the carbon and nitrogen fluxes in Aspergillus oryzae. Food Chem. 271, 606–613. doi: 10.1016/j.foodchem.2018.07.199
Zhao, G., Yao, Y., Wang, C., Hou, L., and Cao, X. (2013). Comparative genomic analysis of Aspergillus oryzae strains 3.042 and RIB40 for soy sauce fermentation. Int. J. Food Microbiol. 164, 148–154. doi: 10.1016/j.ijfoodmicro.2013.03.027
Zhao, G. Z., Yao, Y. P., Qi, W., Wang, C. L., Hou, L. H., Zeng, B., et al. (2012). Draft genome sequence of Aspergillus oryzae strain 3.042. Eukaryotic Cell 11, 1178–1178. doi: 10.1128/Ec.00160-12
Zhu, J. X., Ding, L., and He, S. (2019). Discovery of a new biphenyl derivative by epigenetic manipulation of marine-derived fungus Aspergillus versicolor. Nat. Prod. Res. 33, 1191–1195. doi: 10.1080/14786419.2018.1465423
Zhu, L., Maruyama, J., and Kitamoto, K. (2013). Further enhanced production of heterologous proteins by double-gene disruption (DeltaAosedD DeltaAovps10) in a hyper-producing mutant of Aspergillus oryzae. Appl. Microbiol. Biotechnol. 97, 6347–6357. doi: 10.1007/s00253-013-4795-z
Keywords: Aspergillus oryzae, fermentation industry, genetic engineering technology, cell factory, heterologous protein production
Citation: Jin F-J, Hu S, Wang B-T and Jin L (2021) Advances in Genetic Engineering Technology and Its Application in the Industrial Fungus Aspergillus oryzae. Front. Microbiol. 12:644404. doi: 10.3389/fmicb.2021.644404
Received: 21 December 2020; Accepted: 05 February 2021;
Published: 23 February 2021.
Edited by:
Wanping Chen, Georg August University of Göttingen, GermanyReviewed by:
Bin He, Jiangxi Science and Technology Normal University, ChinaCopyright © 2021 Jin, Hu, Wang and Jin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Feng-Jie Jin, amluZmpAbmpmdS5lZHUuY24=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.