 Kyla L. Hooker
Kyla L. Hooker Vitaly V. Ganusov
Vitaly V. Ganusov- 1Genome Science and Technology, University of Tennessee, Knoxville, TN, United States
- 2Department of Microbiology, University of Tennessee, Knoxville, TN, United States
- 3Department of Mathematics, University of Tennessee, Knoxville, TN, United States
Influenza viruses infect millions of humans every year causing an estimated 400,000 deaths globally. Due to continuous virus evolution current vaccines provide only limited protection against the flu. Several antiviral drugs are available to treat influenza infection, and one of the most commonly used drugs is oseltamivir (Tamiflu). While the mechanism of action of oseltamivir as a neuraminidase inhibitor is well-understood, the impact of oseltamivir on influenza virus dynamics in humans has been controversial. Many clinical trials with oseltamivir have been done by pharmaceutical companies such as Roche but the results of these trials until recently have been provided as summary reports or papers. Typically, such reports included median virus shedding curves for placebo and drug-treated influenza virus infected volunteers often indicating high efficacy of the early treatment. However, median shedding curves may be not accurately representing drug impact in individual volunteers. Importantly, due to public pressure clinical trials data testing oseltamivir efficacy has been recently released in the form of redacted PDF documents. We digitized and re-analyzed experimental data on influenza virus shedding in human volunteers from three previously published trials: on influenza A (1 trial) or B viruses (2 trials). Given that not all volunteers exposed to influenza viruses actually start virus shedding we found that impact of oseltamivir on the virus shedding dynamics was dependent on (i) selection of volunteers that were infected with the virus, and (ii) the detection limit in the measurement assay; both of these details were not well-articulated in the published studies. By assuming that any non-zero viral measurement is above the limit of detection we could match previously published data on median influenza A virus (flu A study) shedding but not on influenza B virus shedding (flu B study B) in human volunteers. Additional analyses confirmed that oseltamivir had an impact on the duration of shedding and overall shedding (defined as area under the curve) but this result varied by the trial. Interestingly, treatment had no impact on the rates at which shedding increased or declined with time in individual volunteers. Additional analyses showed that oseltamivir impacted the kinetics of the end of viral shedding, and in about 20–40% of volunteers that shed the virus treatment had no impact on viral shedding duration. Our results suggest an unusual impact of oseltamivir on influenza viruses shedding kinetics and caution about the use of published median data or data from a few individuals for inferences. Furthermore, we call for the need to publish raw data from critical clinical trials that can be independently analyzed.
1. Introduction
Influenza is a respiratory infection caused by different strains of the influenza virus. Influenza A viruses originate from animals such as birds and pigs while influenza B viruses have no known animal origin (Petrova and Russell, 2018). Disease, caused by influenza viruses, commonly known as the flu, typically affects the upper respiratory system such as the sinus cavities, throat, and sometimes the lungs (Moghadami, 2017). The virus spreads from person to person via respiratory droplets when the infected individual coughs or sneezes in close contact with uninfected individuals (Brankston et al., 2007). Symptoms include fever, fatigue, cough, sore throat, and a runny nose. Most individuals recover from the flu (Moghadami, 2017). However, individuals, usually with underlying health conditions, can have serious and even deadly complications. Millions are infected with influenza viruses globally, and 400,000–500,000 people die each year from complications following influenza virus infections (Petrova and Russell, 2018; Paget et al., 2019). The influenza virus has a high mutation rate resulting in new strains (antigenic drift) that are not readily recognized by immunity of individuals who previously experienced influenza infection (Moghadami, 2017). Occasionally, reassortment of viral genes may occur resulting in variants that are markedly different from currently circulating strains (antigenic shift); such process often results in a pandemic (Kim et al., 2018).
A common prevention is the yearly flu vaccine, but it is not very efficient with an estimated efficacy of about 60% that varies with vaccination year and age of vaccinated individuals (Dhakal and Klein, 2019). The influenza virus has a high mutation rate allowing it to escape from vaccine-induced immunity (Moghadami, 2017). Thus, new influenza vaccines need to be created annually. The creation of the annual influenza vaccination takes into consideration both new strains and current strains of influenza viruses that are circulating globally (Ang et al., 2016; Petrova and Russell, 2018).
Luckily there are several antiviral drugs such as oseltamivir, zanamivir, peramivir, and baloxavir that can be used to either treat severely ill influenza-infected patients or household contacts of individuals with confirmed influenza infection (Mifsud et al., 2019; Principi et al., 2019; Yang et al., 2019). Efficacy of such drugs has been extensively evaluated in clinical trials both including infected patients and volunteers that had been infected with a known strain and dose of an influenza virus. In the latter types of experiments treatment start can be well-defined relative to the infection initiation, and for oseltamivir the treatment appears to impact virus dynamics and/or patient's symptoms when the treatment is started only within 24 h of the infection and/or onset of symptoms (Moghadami, 2017). There are also other limitations of oseltamivir including side effects and the appearance of drug resistant variants (Moghadami, 2017; Lampejo, 2020).
While evidence of the efficacy of drugs against influenza infection such as oseltamivir has been well-documented from several clinical trials (e.g., Hayden et al., 1999, 2000) for a long time there has been very limited publicly available data from such clinical trails. In particular, results of clinical trials have been presented as median viral shedding curves or symptoms for placebo and drug-treated volunteers, and side effects of the treatments were barely discussed. Interestingly, initial reviews of such clinical trials data recommended oseltamivir use for treating influenza infection (Jefferson et al., 2000); many governments stockpiled oseltamivir for emergency use in case of a new pandemic virus (Reddy, 2010). However, concerns of whether the clinical trials data were accurately represented in original publications were raised resulting in public release of some of these clinical trial reports. Interestingly, reanalysis of these and other data reduced the initial enthusiasm to recommend oseltamivir for routine treatment of uncomplicated influenza infections (Jefferson et al., 2006, 2014). While the data from several of the early clinical trials are now available, these data are given as pdf scans of redacted reports and not as the actual raw data which precludes more detailed analysis by other investigators. Moreover, as far as we are aware data from most recent clinical trials of other anti-influenza drugs such as baloxavir are not publicly available (Hayden et al., 2018).
Having properly formatted, digitized data from clinical trials could be extremely useful for understanding of the impact of the drug treatment on influenza virus dynamics in humans. Furthermore, such shedding data could be useful to further understand mechanisms that control duration and magnitude of viral shedding in humans. For example, kinetics of influenza A virus shedding in several human volunteers has been analyzed with the use of mathematical models (Baccam et al., 2006; Canini et al., 2014, 2016). Interestingly, one of the earliest modeling studies suggested that the dynamics of influenza A virus shedding in human volunteers can be well-described by a so-called target cell limited model in which virus dynamics is only restricted by the availability of targets for virus replication (Baccam et al., 2006). However, the Baccam et al. (2006) study used data from only a few volunteers that were not treated with drugs; therefore, it remains unclear if the same model can describe more variable data from a larger group of infected volunteers, or if other alternative models of viral control may be also consistent with viral shedding patterns (Le, 2014; Ganusov, 2016).
In this paper we carefully digitized data on influenza virus shedding in human volunteers from three previously published clinical trials and performed basic analysis of these data. The primary goals of the analysis were to reproduce published results on oseltamivir treatment impact on viral shedding and to provide the community with well-curated datasets on viral dynamics that other researchers may utilize further.
2. Materials and Methods
2.1. Experimental Design
Three randomized placebo-controlled clinical trials using influenza A virus (H1N1, 1 trial) or influenza B virus (2 trials) were done using volunteers (Hayden et al., 1999, 2000). In short, volunteers were inoculated through the nose with the influenza virus strain of the trial at time zero. Treatment type varies by trial (Figure 1 and Table 1). The influenza A trial used a placebo group and a group treated with oseltamivir at daily with 20, 100, 200 mg bid (twice daily), or 200 mg od (once daily). Influenza B study A trial used a placebo group and a group treated daily with oseltamivir at 75 or 150 mg. Influenza B study B trial used a placebo group and a group treated daily with 75 mg of oseltamivir. In all three trials treatment began at 24 h after virus inoculation and nasal washings were taken every 12 h until 96 h (4 days) after inoculation. At that point nasal washings were taken every 24 h until 216 h (9 days). The nasal washings were tested for the viral titer (amount of viral shedding in the patient).
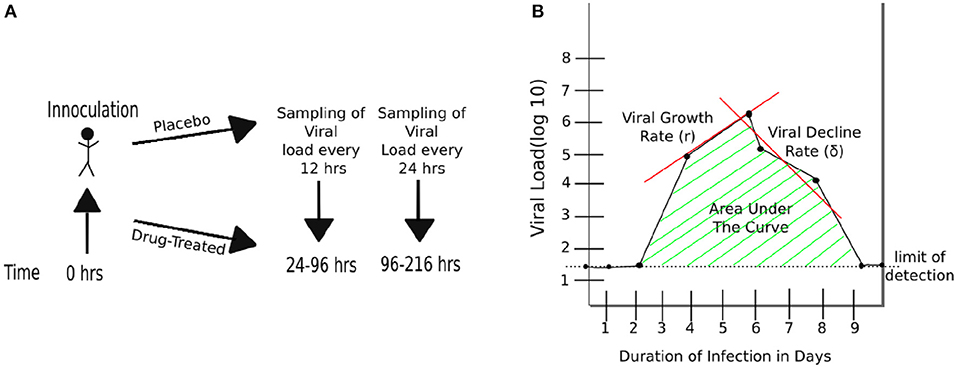
Figure 1. Schematic representation of experimental design and basic characteristics of viral shedding data. (A) volunteers were inoculated with influenza A or B virus at time zero, and treatment with oseltamivir (or placebo) started at 24 h (Hayden et al., 1999, 2000). Nasal washing to measure viral titers were taken every 12 h starting at 24 h until 96 h, and every 24 h thereafter until 216 h (9 days). (B) Basic parameters estimated from kinetics of shedding including the duration of infection (DOI or T; T = 8days in the cartoon), total viral shedding (area under the curve, AUC), viral growth rate (r), and viral decline rate (δ). Area under the curve (AUC) was calculated for viral concentration on the linear scale and then log10-transformed (see Equation 1).
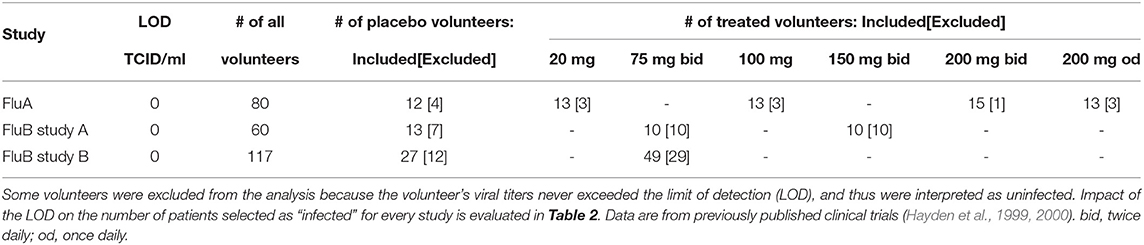
Table 1. Details of the datasets analyzed in the paper.
2.2. Experimental Data
2.2.1. Data Digitization
Redacted PDF files describing in detail clinical trials have been downloaded in 2014 from dryad.org (https://datadryad.org/resource/10.5061/dryad.77471/2). Data from three trials involving treatment of volunteers with oseltamivir (Tamiflu) 24 h after controlled exposure to influenza viruses were chosen for further analysis: study on infection with influenza A virus (“Flu A study,” report PV15616), and two studies on infection with influenza B virus (“Flu B study A,” report NP15717 and “Flu B study B,” report NP15827) (Hayden et al., 1999, 2000). The data in the original PDF files were given in day:hour:min time units for times of virus inoculation or when measurements of viral shedding were taken. Viral titers were measured in tissue-culture infectious doses per ml (TCID50/ml) and are given in log10 units. The data were digitized by KLH into a spreadsheet format with time given in minutes or days since infection. Accuracy of digitization was confirmed by checking the correspondence between pdf files by another student for a set of randomly chosen volunteers.
2.2.2. Defining Uninfected Volunteers
Not all virus-exposed volunteers shed the virus after virus inoculation. Volunteers with no viral titer at any point during the trial (i.e., their viral load measurement never exceeded 0) were excluded from the data analysis (see Table 1 and Supplementary Figures 1–8 for the number and the lists of excluded volunteers).
2.2.3. Calculating Start and Stop of Shedding
To calculate the kinetics at which volunteers started virus shedding in a given cohort (e.g., placebo-treated individuals in Flu A study) we did the following. First, for each volunteer we converted the shedding data into 0 or 1 with 0 values being assigned for times when viral shedding was at or below the LOD and 1 values assigned for all times when viral shedding exceeded LOD or the time was later than the first time point of the positive viral shedding event. Then we used equally spaced time points (0, 0.5, 1, 1.5, etc. days) and we counted the number of 0 or 1 for all volunteers in the cohort. The resulting data are given as the number of volunteers that started shedding the virus at time t = 0, 0.5, 1…days after the virus inoculation. Similarly, to calculate the time by which volunteers stop shedding we converted viral shedding data to 0 or 1 but starting counting time in reverse, starting with the latest time point going backwards. We then calculated the number of individuals in the cohort that were still shedding the virus by time t where t is from sequence 0, 0.5, 1, 1.5, etc. days after virus inoculation. The resulting data are given as the number of volunteers that were still shedding the virus at time t = 0, 0.5, 1…days after the virus inoculation. Generated data for each of the clinical trials are provided as Supplementary Material.
2.2.4. Median Data From Published Studies
To compare median shedding curves calculated in our datasets with published values we digitized data for influenza A virus shedding (from Figure 3 in Hayden et al., 1999) or influenza B virus shedding (Flu B study B from Figure 2 in Hayden et al., 2000). Data were digitized using Engauge Digitizer and are available as Supplementary Material to this paper.
2.3. Statistical Analysis
2.3.1. Duration of Infection (DOI)
Duration of infection was defined as the last time point at which shedding was above the limit of detection.
2.3.2. Area Under the Curve (AUC)
To calculate AUC we converted viral titers which were given in log10 units to the linear scale and then calculated AUC using trapezoid integration method:
where ti is the ith time point of measurement of viral load, where Vi is the viral load in the data, and n is the total number of time points.
2.3.3. Median Shedding Curves
Because measurements of viral shedding by individual volunteers was done at different times after infection to calculate median shedding curves per cohort we did the following. We defined times since infection as t = 0, 0.5, 1, 1.5…9 and calculated median viral titer for all volunteers in the cohort found in the interval (t−0.24, t+0.24). Value 0.24 was chosen so there is no overlap in viral titers used to calculate median for different t.
2.3.4. Viral Shedding Increase (r) or Decline (δ) Rates
To calculate the exponential increase (r) in the viral shedding we selected shedding data which are above the LOD and up to the peak of shedding. The growth rate was calculated as the slope of a linear function fitted to these (log10) viral titer data multiplied by ln(10) [r = slope × ln(10)]. In a similar fashion, for calculating the exponential decline rate (δ) in the viral shedding we selected data after the peak of viral shedding (including the peak value) until the last shedding value above the LOD. The early growth rate was calculated using the last measurement at the LOD and all measurements above the LOD including before the maximum viral titer measurement. In a similar fashion, the late decline rates included all points after the maximum viral titer that were above the LOD and the first measurement at the LOD.
2.3.5. Mathematical Modeling of Viral Shedding Start/End
To quantify the rates at which volunteers start or stop shedding we used a novel mathematical model. In the model we assume that the population of shedders may consist of two sub-populations with fraction f and 1−f and each population either starts or stops shedding at rates s1 and s2 respectively. We assume that progression of a volunteer from “non-shedding” to “shedding” state occurs as a movement via k subcompartments at a rate s1 or s2 (Ganusov and Tomura, 2021). Then the probability that a volunteer starts shedding the virus at time t is given by an incomplete gamma function where k is the shape parameter of the distribution and Γ(k) = Γ(k, 0) = (k−1)!. Assuming that shedding starts (or stops) after a delay τ the proportion of volunteers that start shedding by time t after infection is given by the formula
To describe how volunteers stop shedding the virus at time t after infection we used formula
with Sstart(t) being defined in Equation (2). To characterize the speed at which volunteers start or stop shedding we used average time defined as Ti = k/si. To fit the models to experimental data we used a likelihood approach that had been used previously to describe viral escape from T cell immunity (Ganusov et al., 2013; Ganusov, 2018). Specifically, to describe the fraction of volunteers that start shedding the virus the negative log-likelihood of the model given the data is
where nt is the number of volunteers who have started virus shedding by time t, N is the total number of volunteers in the cohort, Sstart(t) is given in Equation (2), and we ignored the constant terms that are irrelevant when maximizing likelihood. Alternative models in this analysis also included one population model (f = 1 and s2 = 0), no delay (τ = 0), or exponentially distributed shedding times (k = 1). Confidence intervals for the model parameters were estimated using bootstrap approach (Efron and Tibshirani, 1993). For each cohort we resampled volunteers in the cohort with replacement and then followed the same procedure outlined above and calculated the number of individuals that start (or stop) shedding by a particular time after the virus inoculation. The model was then fit to 1,000 of such resampled datasets for each cohort, and 95% confidence intervals were calculated from the distributions of estimated parameters. We used a similar approach to fit the model (Equation 3) to the data on the kinetics of end of viral shedding.
Tests. All major analyses were done in R (version 3.1) or Mathematica 11.3. Fitting the mathematical models to data was done in Mathematica 11.3. Statistical comparisons for various parameters estimated for placebo and treated volunteers were done using non-parameteric unpaired Wilcoxon test (identical to Mann–Whitney test). Nested models were compared using likelihood ratio test (LRT). To compare similarity in kinetics of start or end of virus shedding between placebo- and drug-treated volunteers we fitted the data from two cohorts with either individual parameters per cohort/dataset or with the same parameters for both cohorts/datasets. We then used likelihood ratio test to determine if parameters for virus shedding start/end were different between the two cohorts/datasets.
3. Results
3.1. Impact of Limit of Detection on the Number of Infected Volunteers
In clinical trials testing efficacy of oseltamivir, healthy volunteers were inoculated with a defined dose of the influenza A or B viruses (see section 2 for more detail and Hayden et al., 1999, 2000). Interestingly, many volunteers did not shed any detectable virus for the whole duration of the clinical trial (Table 1 and Supplementary Figures 1–9):
Flu A: placebo 4/16 (25%) and treated 10/64 (15%) or 17.5% overall (, p = 0.38 for placebo vs. treated volunteers);
Flu B study A: placebo 7/20 (35%) and treated 20/40 (50%) or 45% overall (, p = 0.27);
Flu B study B: placebo 12/39 (30%) and treated 29/78 (37%) or 35% overall (, p = 0.49).
These individuals were excluded from further analyses on the kinetics of viral shedding. While most other volunteers showed consistent and high viral shedding, some volunteers showed detectable (above zero) shedding only at one time point. It was unclear from the study descriptions whether such individuals should be counted as infected or if such viral blips are false positives. We investigated how changing the limit of detection (LOD) might impact the number of volunteers classified as infected for these clinical trials, and thus, may influence the estimated percent of infected volunteers.
We reasoned that because treatment started 24 h after infection and the replication cycle of influenza viruses is relatively short (<24 h; Frensing et al., 2016; Laske et al., 2019), oseltamivir should not impact the probability of a person to be infected (Cheung et al., 2015); therefore, by changing the LOD the difference between percent of infected in placebo- or drug-treated groups can be evaluated using two-by-two contingency tables (Table 2). The basic idea was to vary the LOD to several values that viral titers take in a given trial and see at which values the frequency of volunteers that shed the virus at any time point (i.e., have viral titers above the threshold value) is the same between placebo and treatment groups. We tried several different values of the LOD and only in the 3rd trial (Flu B study B) did increasing the LOD to 1.5 log10TCID50/ml significantly impact the frequency of infection between placebo and treated volunteers. Interestingly, changes in the number of volunteers classified as infected at different LOD also impacted the average/median viral shedding curves as well as parameters inferred from such curves (results not shown) suggesting that LOD value is critical for analysis of such data. Because increasing LOD reduced the number of volunteers shedding the virus above LOD (Table 2), we converged to use LOD = 0 in our further calculations; some results were also checked assuming other values for LOD (see below). Importantly, using LOD = 0 we could match the median virus shedding titers for placebo- and drug-treated volunteers in Flu A study suggesting that this was the likely LOD in that study (see below). Taken together, this analysis suggests that the value of LOD is important in quantitative analysis of experimental data; future studies should state clearly the defined LOD assumed in the analyses and provide evidence for why that specific LOD value is appropriate. The best approach to determine the LOD is via specific experiments, e.g., by measuring known amounts of the virus via serial dilutions in a similar fashion as clinical samples to allow for possibility of cross-contamination.
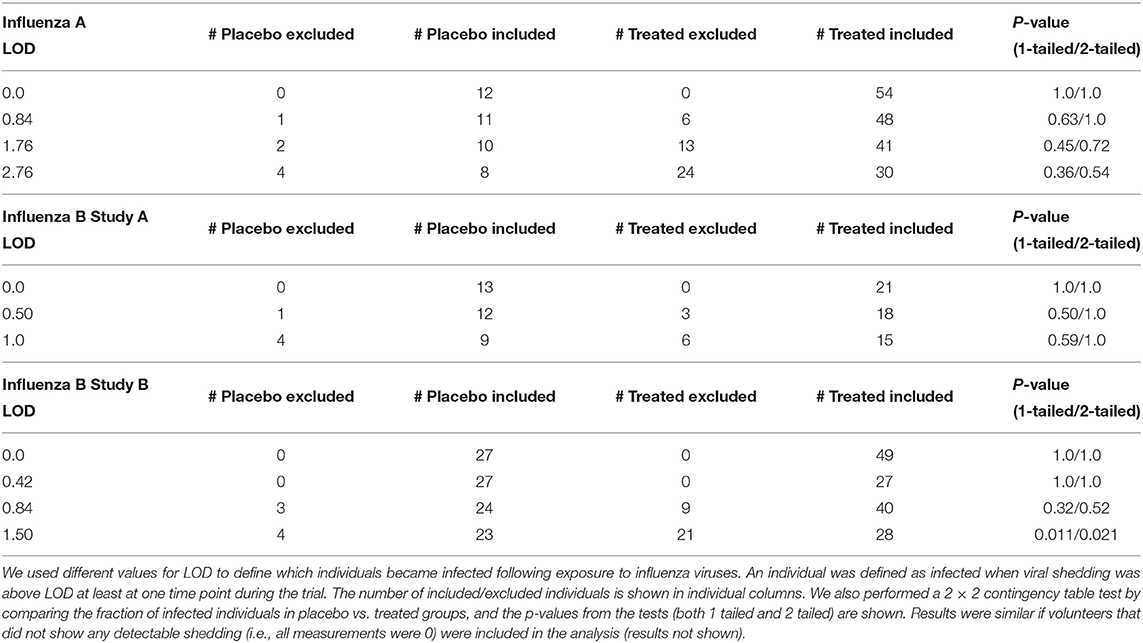
Table 2. Impact of the limit of detection (LOD) on the number of volunteers defined as infected or uninfected in three analyzed clinical trials.
3.2. Impact of Oseltamivir Treatment on Overall Viral Shedding Pattern in Human Volunteers
Having defined the likely LOD for the data we then compared how accurately median shedding curves, generally reported in clinical trials (Hayden et al., 1999, 2000), represent virus shedding patterns in individual volunteers (Figure 2; see section 2 for how median shedding curves were calculated). Interestingly, for flu A-infected volunteers there was a reasonable correspondence between median shedding curves and shedding for individual placebo- and drug-treated volunteers; however, there were clearly examples of individuals that did not follow the median pattern (Figures 2A,D). The match between median shedding and shedding in individual volunteers was poor for both studies with influenza B virus; particularly, median curves poorly represented individuals that continued to shed the virus in both placebo- and drug-treated cohorts (e.g., Figures 2B,E) illustrating a severe limitation of presenting the data by median shedding alone.
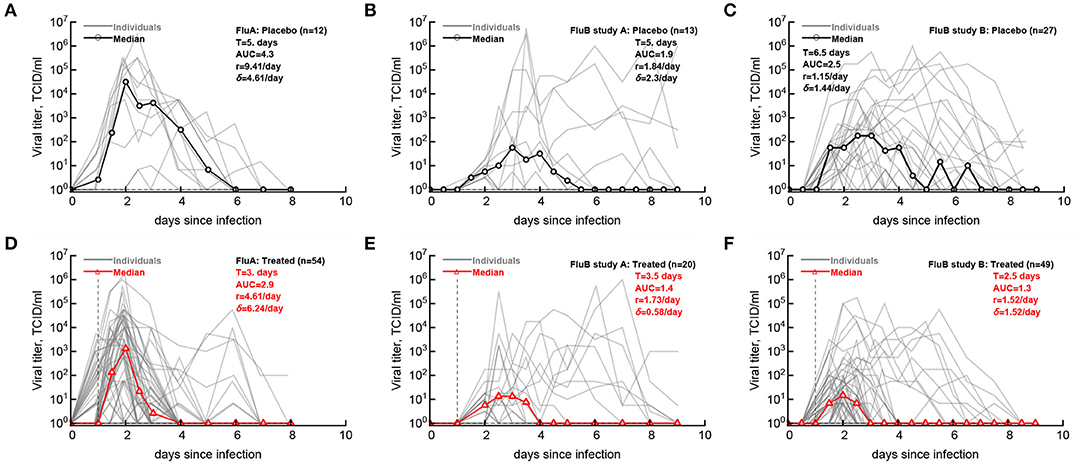
Figure 2. Commonly reported median viral shedding curves do not accurately represent shedding in many patients. We analyzed experimental data from 3 clinical trials of volunteers infected with influenza A (A,D) or influenza B (B,C,E,F) viruses and treated 24 h after viral exposure with placebo (A–C) or oseltamivir (D–F); start of treatment is indicated by the vertical dashed lines. Viral shedding in individual volunteers is shown by gray lines and median viral titers are shown by thick red lines with markers. For median viral titers we also calculated the duration of infection (T), the rate of viral growth (r) and viral decay (δ), and the total area under the curve (AUC, Equation 1). Details of the experiments and basic viral characteristics calculated are given in [Hayden et al. (1999, 2000) see Figure 1]. The limit of detection is represented by the horizontal thin dashed line; the detection limit was 0 log10TCID50/ml for all data. Viral shedding curves in individual volunteers are shown in Supplementary Figures 1–9.
For our median shedding curves we calculated several basic parameters characterizing virus dynamics such as the duration of infection (T), the overall viral shedding (defined as area under the curve, AUC), the virus growth rate prior to peak r, and the virus decline rate after the peak δ. Treatment impacted these parameters for the median shedding curves differently. In particular, oseltamivir treatment reduced the median duration of infection from 5 to 3 days and reduced the overall viral shedding over 10-fold (AUC change from 4.2 to 2.9) in flu A clinical trial as judged by the median shedding curves (Figures 2A,D). Also as judged by the median shedding curves, in influenza A virus-infected individuals treatment reduced the rate of viral growth and increased the rate of viral clearance after the peak (Figures 2A,D). Similarly, based on the median shedding curves in flu B study A olsetamivir reduced the duration of infection from 5 to 3.5 days, and moderately, by three-fold reduced overall shedding (AUC change from 1.9 to 1.4). In contrast, drug treatment resulted in slower viral clearance after the peak (Figures 2B,E). Finally, in flu B study B, treatment did not appear to have an impact on viral growth or decline rates of the median shedding data but did result in shorter infection (by 4 days) and less overall virus being shed (by 16-fold, Figures 2C,F).
Given that median viral shedding curves did not necessarily well represent shedding observed in individual volunteers, impact of the treatment on median shedding curves may be misleading. Therefore, we performed an alternative analysis in which we calculated the same parameters (T, AUC, r, and δ) for individual volunteers and compared these parameters between placebo- and drug-treated cohorts (Figure 3). It is important to emphasize that it was not possible to calculate virus growth and decay rates for all volunteers because this procedure required at least two measurements to be above LOD during viral increase or decline phases which was not available in all volunteers. Interestingly, we found that oseltamivir treatment did reduce the overall duration of infection and the total shedding in flu A study and flu B study B, but there was no difference in these two parameters between placebo- and drug-treated individuals in flu B study A (Figure 3).
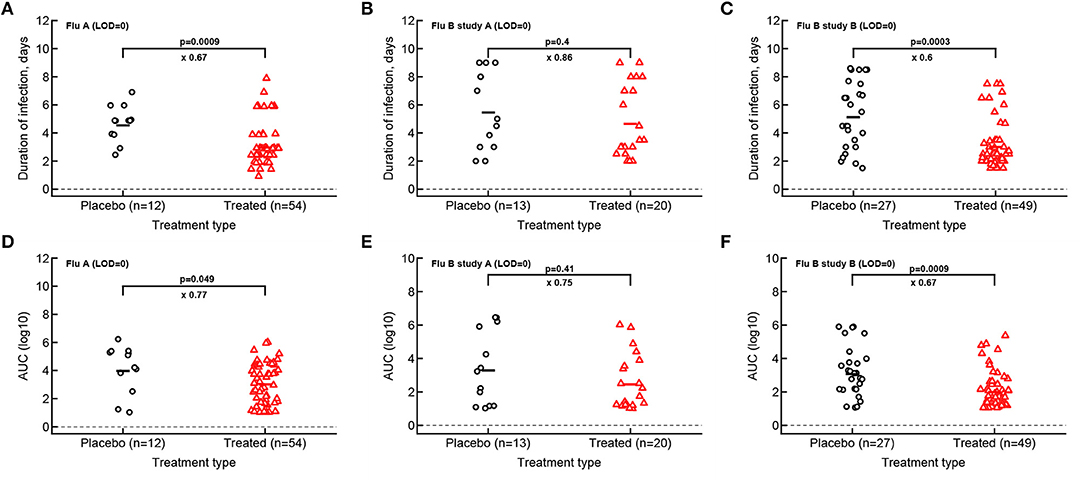
Figure 3. Impact of oseltamivir treatment on the duration of infection and the overall viral shedding depends on the study. For every volunteer in 3 clinical trials we calculated the duration of infection (A–C) or the overall shedding defined as AUC (see section 2 for detail) for Flu A (A,D), Flu B study A (B,E), or Flu B study B (C,F) data. Horizontal lines indicate median values. The number of volunteers, n, analyzed in each of the trial and tests is shown on the x-axis of each graph. Comparisons between groups done using Mann–Whitney test, magnitude of the effect is indicated as × fold change, and p values from the test are shown on individual panels.
Our analysis of the virus kinetics defined by the virus shedding growth rate to the peak and virus decline rate after the peak based on median virus shedding curves suggested that oseltamivir influences these rates albeit in trial-dependent manner (Le, 2014, Figure 2). Specifically, we found that oseltamivir increases the rate of virus decline after the peak in the median data in flu A clinical trial (Le, 2014, Figures 2A,D). In contrast, analysis of individual shedding curves did not reveal impact of oseltamivir on either viral growth or viral decline rates (Figures 4A,D).
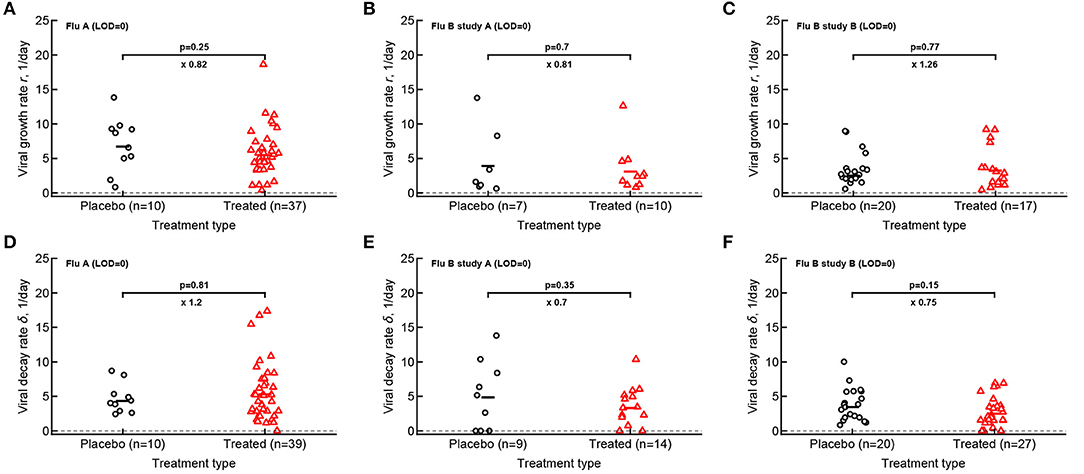
Figure 4. Oseltamivir treatment does not influence median viral growth and decline rates in individual volunteers. For the data from 3 clinical studies for every volunteer we calculated the rate at which viral shedding increased (A–C) or declined (D–F) with time for Flu A (A,D), Flu B study A (B,E), and Flu B study B (C,F) data. Only volunteers with the data above limit of detection (LOD) and at least two data points for virus increase to the peak or virus decline after the peak were included. Horizontal lines denote median values. The number of volunteers, n, analyzed in each of the trial and tests is shown on the x-axis of each graph. Comparisons between groups done using Mann–Whitney test, magnitude of the effect is indicated as × fold change, and p values from the test are shown on individual panels.
In calculating virus growth and decay rates for individual volunteers we only used values that were above the LOD; therefore, this approach is likely to capture the average rates of virus shedding change. We wondered if drug treatment may impact the very early rate of virus shedding increase (or the very late virus shedding decline). Therefore, we also calculated two additional rates - the rate at which virus shedding increases early during the infection (by using the data that included one measurement at the LOD prior to detectable virus but before the viral peak) and the late rate of virus decline (by using the data after the peak shedding until the first measurement at LOD; see section 2 for more detail). Interestingly, there was no evidence that the rate at which virus shedding increased early was changing over time either in placebo- or drug-treated volunteers (Supplementary Figure 11). In contrast, the rate at which virus shedding was declining slowed over time in 5 out of 6 comparisons (Supplementary Figure 12).
Both flu A and flu B study A trials tested the impact of different drug dosing on viral control (see section 2). Additional analysis showed that interestingly the oseltamivir's dose had no measurable impact on the basic parameters for virus shedding kinetics which further justifies considering drug-treated volunteers as a single group (Supplementary Figures 13, 14).
3.3. Matching Median Virus Shedding Curves With Published Studies
Original papers of the analyzed clinical trials included median viral shedding curves for the influenza A trial (Hayden et al., 1999) and the influenza B study B trial (Hayden et al., 2000). We therefore investigated whether median viral shedding curves from our digitized data would accurately match published median viral curves. In our analyses we found that the assumed LOD influenced the resulting median shedding curve (results not shown), and interestingly, by assuming LOD = 0 we found a nearly perfect match between median viral shedding in our data and published values for flu A clinical trial (Figures 5A,C). For the full match the median shedding data found in our analysis had to be shifted by 12h, though. In contrast, we could not fully match median viral shedding curves for flu B study B clinical trial for several values of the LOD (results not shown). Furthermore, even to provide a reasonable match with LOD = 0 the median shedding curve in our analysis had to be shifted by 24 h (Figures 5B,D). We could not identify the reasons for this discrepancy but it is possible that this due to difference of which volunteers are included in the analysis. In particular, in trials with influenza B virus some volunteers were excluded from the analysis because they were found to be seropositive (Hayden et al., 2000). Overall, our result further suggests the need to publish raw data from such clinical trials so that some of the results can be independently verified.
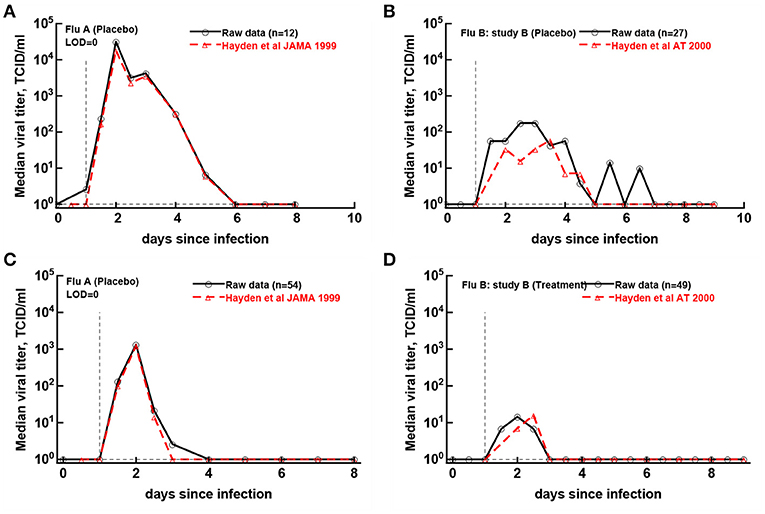
Figure 5. Previously published median virus shedding data could be reproduced for one but not another clinical trial. We compared median shedding data published previously for clinical trial with flu A (Hayden et al., 1999) or flu B study B (Hayden et al., 2000) with median shedding calculated from the raw trial data. The best match was found by assuming LOD = 0. For every panel we also list the number of volunteers used to calculate the median titers. The best match was obtained by shifting the published median shedding curves by 12 h (A,C) or by 24 h (B,D).
3.4. Evaluating Impact of Oseltamivir Treatment on the Overall Kinetics of Start and End of Viral Shedding
Viral shedding dynamics in individual volunteers is relatively asynchronous—the time when individuals start or stop viral shedding (i.e., with viral shedding being above the LOD or reaching the LOD) varied. We therefore investigated if the rate at which volunteers started or stopped shedding was dependent on the virus type and, more importantly, on the oseltamivir treatment. We developed a novel mathematical model predicting start (or end) of shedding in the cohort of patients, based on a mixture of two gamma distributions, and estimated the model parameters by fitting a series of nested models to the data. Specifically, placebo- or drug-treated volunteers were followed as a cohort and the fraction of volunteers that started virus shedding by time t since infection (Figures 6A–E) or stopped shedding by time t (Figures 6B–F) was calculated. The models were fitted using a likelihood method and the best fit model was determined using likelihood ratio test (see section 2 for more detail and Figure 6).
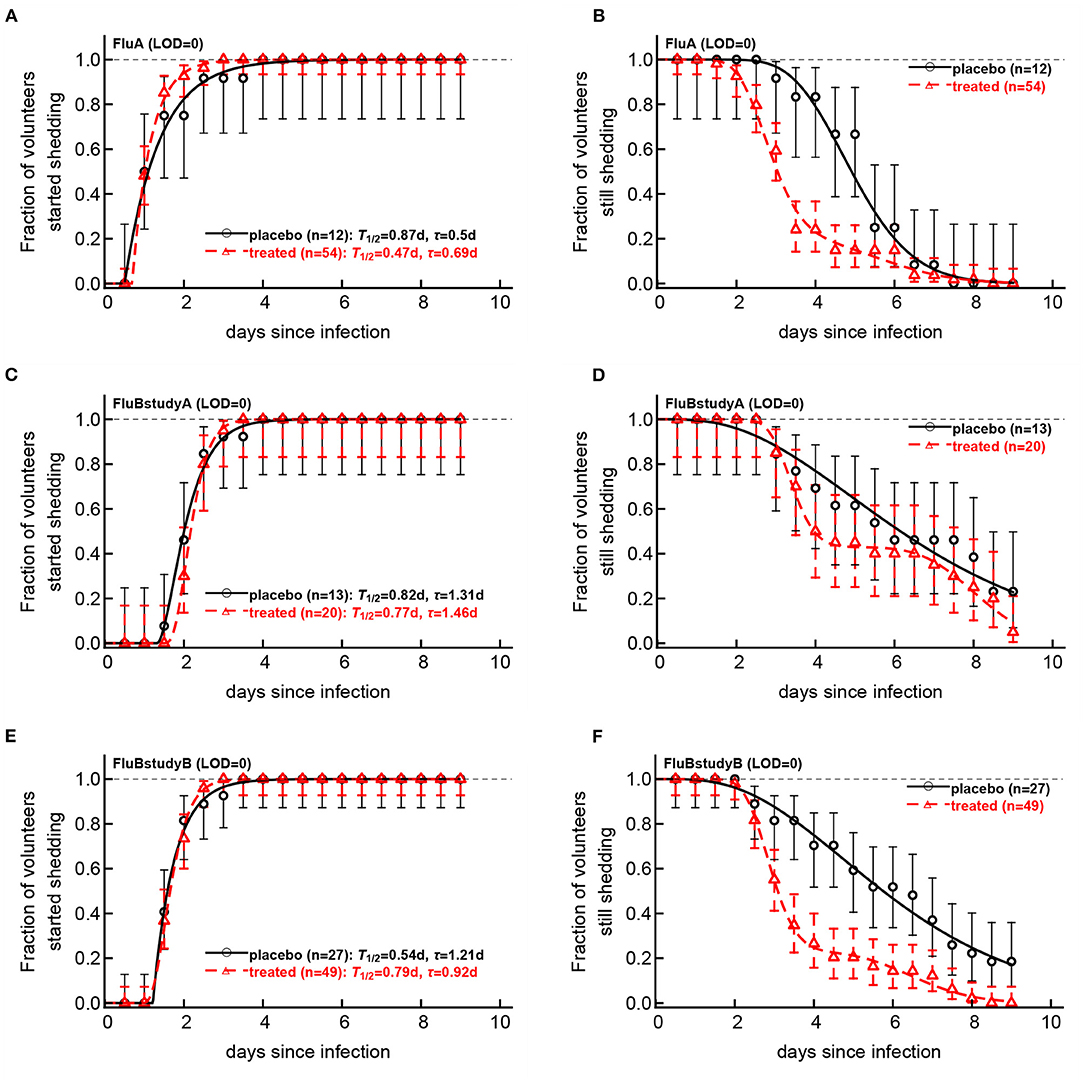
Figure 6. Impact of oseltamivir treatment on kinetics of the start and end of influenza virus shedding. We calculated the cumulative fraction of volunteers that start (A,C,E) or stop (B,D,F) shedding following infection with influenza A (A,B) or influenza B (C–E) viruses. Data are shown by markers for placebo or treated volunteers and in parentheses we show the numbers of volunteers in each cohort. We fitted a general mathematical model (given in Equations 2 or 3) to these data and the predictions of the best fit models are shown by lines. For every fit the minimal number of fitted parameters was selected using a likelihood ratio test. Best fit parameters are shown in Table 3. In panels, T = k/si is the average shedding time and τ is the delay in shedding (see Equation 2 for parameter definition). There was a moderate difference in the kinetics of start of shedding in Flu A study between placebo- and drug-treated volunteers (p = 0.047), but no difference was observed in start of virus shedding two studies with flu B (p>0.2). In FluA study and FluB study B the kinetics of stop of shedding was different between placebo and treated volunteers as judged by the likelihood ratio test (p < 0.001 for both comparisons). Error bars were estimated for binomial proportions using Jefferey's intervals (Brown et al., 2001).
Several interesting results emerged. The kinetics of start of viral shedding was similar for both influenza virus types with half of individuals starting shedding within 1–2 days after infection. The best fit model describing shedding kinetics varied with the study and was dependent on the treatment type; interestingly, oseltamivir-treated volunteers infected with influenza A virus started shedding the virus slightly faster than compared to controls (LRT: , p = 0.047, Figure 6A and Table 3). However, in the other two studies start of shedding kinetics was similar between placebo- and drug-treated volunteers (Figure 6 and Table 3).
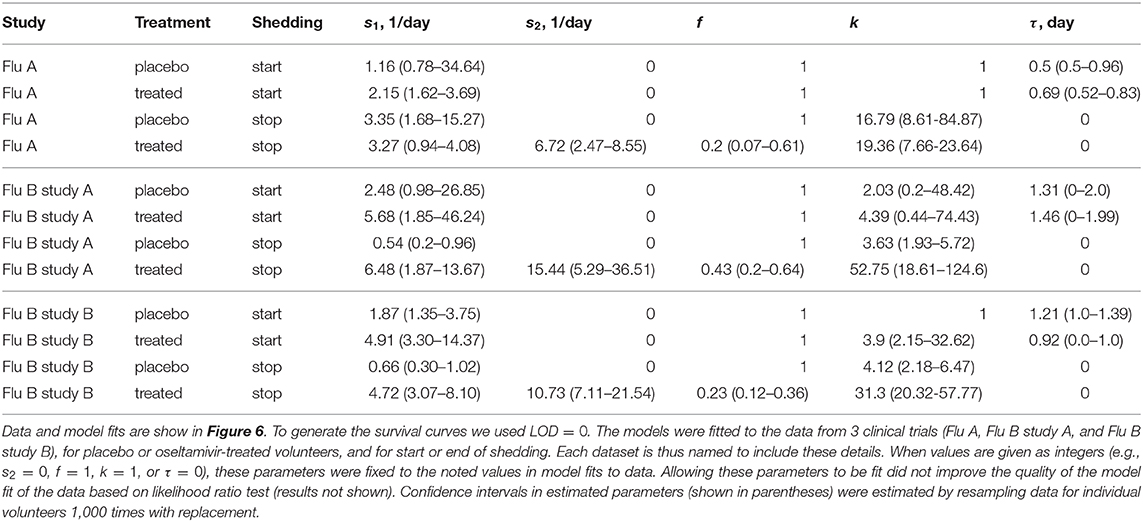
Table 3. Parameters providing the best fit of the model (given by Equations 2 or 3) to the data on start or end of shedding by volunteers in three clinical trials.
The kinetics at which volunteers stopped shedding was even more intriguing. Only for flu B study A we found no differences in the kinetics of viral shedding end between placebo- and oseltamivir-treated volunteers (Figure 6D); however, the model fits predicted that in about 43% of drug-treated individuals in this trial the loss of virus shedding proceeded similarly to that of placebo-treated volunteers (given by parameter f in Table 3). Indeed, the average time to stop shedding was 3.63/0.54 = 6.76days (95% CIs: 4.86−11.03) and 52.75/6.48 = 8.1days (95% CIs: 7.77−10.80) for placebo- and drug-treated volunteers, respectively, suggesting that the drug may have failed in preventing virus replication in many individuals in this trial. There were statistically significant differences in the kinetics of loss of viral shedding between placebo- and drug-treated volunteers in flu A study and flu B study B (Figures 6B,F and Table 3). In these trials, about 20% of drug-treated volunteers stopped shedding the virus with similar timing to that of placebo-treated individuals (Table 3). Thus, our novel analysis identified a variable impact of the oseltamivir treatment on the kinetics at which virus shedding ends in human volunteers and raised an intriguing possibility that oseltamivir treatment may speed up initial viral shedding in influenza A-infected individuals.
4. Discussion
One of the great advantages of testing efficacy of vaccines or drugs against influenza viruses is the ability to perform controlled human challenge studies in which volunteers (placebo and drug-treated or vaccinated) are exposed to well-defined dose and type of virus (Memoli et al., 2015; Balasingam and Wilder-Smith, 2016). However, results of such important trials in general have been presented in succinct manner; e.g., typically median virus shedding curves are presented and public access to data on influenza virus dynamics in individual volunteers is not provided (Hayden et al., 1999, 2000, 2018). In our personal experience, researchers that do have access to such raw data have not been willing to share them. In this study we took advantage of now publicly available pdf reports from three clinical studies testing efficacy of oseltamivir given 24 h after exposure of volunteers to influenza A or B viruses. We have converted scanned pdf pages into spreadsheet format data that can be analyzed further and made it a resource for the community. We also performed several basic analyses of these data.
We found that some of the previously published results could be reproduced, i.e., oseltamivir treatment did reduce the duration of viral shedding and the overall viral shedding in 2 out of 3 trials (Figure 3); however, a closer inspection of the data revealed that many treated volunteers continued shedding the virus suggesting treatment failure. Indeed, by looking at the kinetics at which volunteers stop shedding we found that between 20 and 40% of treated volunteers continued shedding similarly as placebo-treated controls further suggesting treatment failure (Figure 6). Our mathematical model-driven analysis also suggested that treatment with oseltamivir may in fact slightly speed up kinetics of viral shedding as was noted in flu A clinical trial (Figure 6A) but not in flu B trials. While, mechanisms of this minor effect are unclear—we could not find information on the volunteers that shed the virus early—further studies may need to investigate this possibility further.
We could only partially reproduce previously published median shedding curves, specifically for flu A trial but only after shifting the published median shedding curves to allow for a nearly perfect match (Figures 5A,C). Why it was not possible for flu B study B remains unclear. It is also interesting to note that while analysis of the median shedding curves in flu A study suggested a faster viral clearance in oseltamivir-treated volunteers (Le, 2014), virus decay rates were similar when evaluated for individual volunteers (Figure 4). Given that oseltamivir treatment also did not impact the early virus growth rate this may suggest that virus shedding is controlled mainly by innate mechanisms (e.g., target cell limitation, Baccam et al., 2006; Le, 2014). Further studies should investigate in more detail whether alternative models for virus control are consistent with shedding data for individual volunteers infected with influenza A or B viruses (Handel et al., 2010; Le, 2014).
Our work has some limitations. Because original virus shedding data were in PDF format we cannot exclude the possibility that there have been errors in our translation from the given days/times since infection to digital format. We attempted to minimize such errors by having another investigator to check correctness of the digitization. We could not determine why some volunteers responded well to the drug and displayed improved clearance of the virus while others did not. It was not possible to find clear answers in the clinical trial notebooks. Our analysis of virus growth and decay rates depended strongly on the measured viral shedding values (above LOD), and more frequent measurements of viral shedding are likely to provide more accurate estimates of these rates.
Our work generates several recommendations that should be implemented for clinical trial data on human challenge studies with influenza viruses. We believe that original shedding data should be shared following publication of the work. The data can be anonymized and redacted if needed, but the critical numeric data must be provided in the proper digital format (e.g., spreadsheets or similar). For example, recent clinical trial data for baloxavir have not been made available (Hayden et al., 2018). Authors that provide re-analysis of the data from clinical trials should also attempt to provide public access to such data. Data sharing is likely to improve science reproducibility which is likely to ultimately benefit influenza virus-infected patients.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Ethics Statement
Data from redacted documents provided publicly by Roche and published previously.
Author Contributions
KH digitized the data from original publications into spreadsheet format. KH and VG have analyzed the data and wrote the manuscript. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank our research community for pressuring Roche to release primary research data from oseltamivir clinical trials. We also thank several students in the GanusovLab for helping with some primary analyses and checks of the digitized data. This work was in part supported by the NIH grant (R01 GM118553) to VG.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2021.631211/full#supplementary-material
Abbreviations
LOD, limit of detection; DOI, days of infection (also denoted as T); bid, twice daily; od, once daily; AUC, area under the curve; LRT, likelihood ratio test.
References
Ang, L. W., Tien, W. S., Lin, R. T.-P., Cui, L., Cutter, J., James, L., et al. (2016). Characterization of influenza activity based on virological surveillance of influenza-like illness in tropical singapore, 2010-2014. J. Med. Virol. 88, 2069–2077. doi: 10.1002/jmv.24566
Baccam, P., Beauchemin, C., Macken, C. A., Hayden, F. G., and Perelson, A. S. (2006). Kinetics of influenza a virus infection in humans. J. Virol. 80, 7590–7599. doi: 10.1128/JVI.01623-05
Balasingam, S., and Wilder-Smith, A. (2016). Randomized controlled trials for influenza drugs and vaccines: a review of controlled human infection studies. Int. J. Infect. Dis. 49, 18–29. doi: 10.1016/j.ijid.2016.05.013
Brankston, G., Gitterman, L., Hirji, Z., Lemieux, C., and Gardam, M. (2007). Transmission of influenza a in human beings. Lancet Infect. Dis. 7, 257–265. doi: 10.1016/S1473-3099(07)70029-4
Brown, L., Cai, T., DasGupta, A., Agresti, A., Coull, B., Casella, G., et al. (2001). Interval estimation for a binomial proportion. Stat. Sci. 16, 101–133. doi: 10.1214/ss/1009213286
Canini, L., Conway, J. M., Perelson, A. S., and Carrat, F. (2014). Impact of different oseltamivir regimens on treating influenza a virus infection and resistance emergence: insights from a modelling study. PLoS Comput. Biol. 10:e1003568. doi: 10.1371/journal.pcbi.1003568
Canini, L., Woolhouse, M. E. J., Maines, T. R., and Carrat, F. (2016). Heterogeneous shedding of influenza by human subjects and its implications for epidemiology and control. Sci. Rep. 6:38749. doi: 10.1038/srep38749
Cheung, D. H., Tsang, T. K., Fang, V. J., Xu, J., Chan, K.-H., Ip, D. K. M., et al. (2015). Association of oseltamivir treatment with virus shedding, illness, and household transmission of influenza viruses. J. Infect. Dis. 212, 391–396. doi: 10.1093/infdis/jiv058
Dhakal, S., and Klein, S. L. (2019). Host factors impact vaccine efficacy: implications for seasonal and universal influenza vaccine programs. J. Virol. 93:e00797-19. doi: 10.1128/JVI.00797-19
Efron, B., and Tibshirani, R. (1993). An Introduction to the Bootstrap. New York, NY: Chapman and Hall. doi: 10.1007/978-1-4899-4541-9
Frensing, T., Kupke, S. Y., Bachmann, M., Fritzsche, S., Gallo-Ramirez, L. E., and Reichl, U. (2016). Influenza virus intracellular replication dynamics, release kinetics, and particle morphology during propagation in mdck cells. Appl. Microbiol. Biotechnol. 100, 7181–7192. doi: 10.1007/s00253-016-7542-4
Ganusov, V. V. (2016). Strong inference in mathematical modeling: a method for robust science in the twenty-first century. Front. Microbiol. 7:1131. doi: 10.3389/fmicb.2016.01131
Ganusov, V. V. (2018). Time intervals in sequence sampling, not data modifications, have a major impact on estimates of HIV escape rates. Viruses 10:99. doi: 10.3390/v10030099
Ganusov, V. V., Neher, R. A., and Perelson, A. S. (2013). Modeling HIV escape from cytotoxic T lymphocyte responses. J. Stat. Mech. 2013:P01010. doi: 10.1088/1742-5468/2013/01/P01010
Ganusov, V. V., and Tomura, M. (2021). “Experimental and mathematical approaches to quantify recirculation kinetics of lymphocytes,” in Mathematical, Computational and Experimental T Cell Immunology, eds C. Molina-Paris and G. Lythe (Cham: Springer), 151–169.
Handel, A., Longini, I. M., and Antia, R. (2010). Towards a quantitative understanding of the within-host dynamics of influenza a infections. J. R. Soc. Interface 7, 35–47. doi: 10.1098/rsif.2009.0067
Hayden, F. G., Jennings, L., Robson, R., Schiff, G., Jackson, H., Rana, B., et al. (2000). Oral oseltamivir in human experimental influenza b infection. Antiviral Therapy 5, 205–213.
Hayden, F. G., Sugaya, N., Hirotsu, N., Lee, N., de Jong, M. D., Hurt, A. C., et al. (2018). Baloxavir marboxil for uncomplicated influenza in adults and adolescents. N. Engl. J. Med. 379, 913–923. doi: 10.1056/NEJMoa1716197
Hayden, F. G., Treanor, J. J., Fritz, R. S., Lobo, M., Betts, R. F., Miller, M., et al. (1999). Use of the oral neuraminidase inhibitor oseltamivir in experimental human influenza: randomized controlled trials for prevention and treatment. JAMA 282, 1240–1246. doi: 10.1001/jama.282.13.1240
Jefferson, T., Demicheli, V., Deeks, J., and Rivetti, D. (2000). Neuraminidase inhibitors for preventing and treating influenza in healthy adults. Cochrane Datab. Syst. Rev. CD001265. doi: 10.1002/14651858.CD001265
Jefferson, T., Jones, M. A., Doshi, P., Del Mar, C. B., Hama, R., Thompson, M. J., et al. (2014). Neuraminidase inhibitors for preventing and treating influenza in healthy adults and children. Cochrane Datab. Syst. Rev. CD008965. doi: 10.1002/14651858.CD008965.pub4
Jefferson, T. O., Demicheli, V., Di Pietrantonj, C., Jones, M., and Rivetti, D. (2006). Neuraminidase inhibitors for preventing and treating influenza in healthy adults. Cochrane Datab. Syst. Rev. CD001265. doi: 10.1002/14651858.CD001265.pub2
Kim, H., Webster, R. G., and Webby, R. J. (2018). Influenza virus: dealing with a drifting and shifting pathogen. Viral Immunol. 31, 174–183. doi: 10.1089/vim.2017.0141
Lampejo, T. (2020). Influenza and antiviral resistance: an overview. Eur. J. Clin. Microbiol. Infect. Dis. 39, 1201–1208. doi: 10.1007/s10096-020-03840-9
Laske, T., Bachmann, M., Dostert, M., Karlas, A., Wirth, D., Frensing, T., et al. (2019). Model-based analysis of influenza a virus replication in genetically engineered cell lines elucidates the impact of host cell factors on key kinetic parameters of virus growth. PLoS Comput. Biol. 15:e1006944. doi: 10.1371/journal.pcbi.1006944
Le, D. (2014). Mathematical and experimental studies of immune response to viral infection (yellow fever, smallpox, influenza) (Honor's thesis). University of Tennessee, Knoxville, TN, United States.
Memoli, M. J., Czajkowski, L., Reed, S., Athota, R., Bristol, T., Proudfoot, K., et al. (2015). Validation of the wild-type influenza A human challenge model H1N1pdmist: an A(H1N1)pdm09 dose-finding investigational new drug study. Clin. Infect. Dis. 60, 693–702. doi: 10.1093/cid/ciu924
Mifsud, E. J., Hayden, F. G., and Hurt, A. C. (2019). Antivirals targeting the polymerase complex of influenza viruses. Antiviral Res. 169:104545. doi: 10.1016/j.antiviral.2019.104545
Moghadami, M. (2017). A narrative review of influenza: a seasonal and pandemic disease. Iran. J. Med. Sci. 42, 2–13.
Paget, J., Spreeuwenberg, P., Charu, V., Taylor, R. J., Iuliano, A. D., Bresee, J., et al. (2019). Global mortality associated with seasonal influenza epidemics: New burden estimates and predictors from the glamor project. J. Global Health 9:020421. doi: 10.7189/jogh.09.020421
Petrova, V. N., and Russell, C. A. (2018). The evolution of seasonal influenza viruses. Nat. Rev. Microbiol. 16, 47–60. doi: 10.1038/nrmicro.2017.118
Principi, N., Camilloni, B., Alunno, A., Polinori, I., Argentiero, A., and Esposito, S. (2019). Drugs for influenza treatment: is there significant news? Front. Med. 6:109. doi: 10.3389/fmed.2019.00109
Reddy, D. (2010). Responding to pandemic (H1N1) 2009 influenza: the role of oseltamivir. J. Antimicrob. Chemother. 65(Suppl 2), ii35–ii40. doi: 10.1093/jac/dkq014
Keywords: influenza virus, shedding, humans, mathematical modeling, oseltamivir
Citation: Hooker KL and Ganusov VV (2021) Impact of Oseltamivir Treatment on Influenza A and B Virus Dynamics in Human Volunteers. Front. Microbiol. 12:631211. doi: 10.3389/fmicb.2021.631211
Received: 19 November 2020; Accepted: 25 January 2021;
Published: 01 March 2021.
Edited by:
Esteban A. Hernandez-Vargas, Frankfurt Institute for Advanced Studies, GermanyReviewed by:
Hana Maria Dobrovolny, Texas Christian University, United StatesIrene Ramos, Icahn School of Medicine at Mount Sinai, United States
Copyright © 2021 Hooker and Ganusov. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Vitaly V. Ganusov, dml0YWx5LmdhbnVzb3ZAZ21haWwuY29t