 Ana Érika Inácio Gomes1†
Ana Érika Inácio Gomes1† Thaisy Pacheco1†Cristiane da Silva dos Santos1José Aires Pereira2
Thaisy Pacheco1†Cristiane da Silva dos Santos1José Aires Pereira2 Marcelo Lima Ribeiro3Michelle Darrieux1
Marcelo Lima Ribeiro3Michelle Darrieux1 Lúcio Fábio Caldas Ferraz1*
Lúcio Fábio Caldas Ferraz1*- 1Laboratório de Biologia Molecular de Microrganismos, Universidade São Francisco, Bragança Paulista, Brazil
- 2Laboratório de Biologia Molecular e Celular de Tumores, Universidade São Francisco, Bragança Paulista, Brazil
- 3Laboratório de Imunofarmacologia e Biologia Molecular, Universidade São Francisco, Bragança Paulista, Brazil
Although originally known as an opportunistic pathogen, Klebsiella pneumoniae has been considered a worldwide health threat nowadays due to the emergence of hypervirulent and antibiotic-resistant strains capable of causing severe infections not only on immunocompromised patients but also on healthy individuals. Fimbriae is an essential virulence factor for K. pneumoniae, especially in urinary tract infections (UTIs), because it allows the pathogen to adhere and invade urothelial cells and to form biofilms on biotic and abiotic surfaces. The importance of fimbriae for K. pneumoniae pathogenicity is highlighted by the large number of fimbrial gene clusters on the bacterium genome, which requires a coordinated and finely adjusted system to control the synthesis of these structures. In this work, we describe KpfR as a new transcriptional repressor of fimbrial expression in K. pneumoniae and discuss its role in the bacterium pathogenicity. K. pneumoniae with disrupted kpfR gene exhibited a hyperfimbriated phenotype with enhanced biofilm formation and greater adhesion to and replication within epithelial host cells. Nonetheless, the mutant strain was attenuated for colonization of the bladder in a murine model of urinary tract infection. These results indicate that KpfR is an important transcriptional repressor that, by negatively controlling the expression of fimbriae, prevents K. pneumoniae from having a hyperfimbriated phenotype and from being recognized and eliminated by the host immune system.
Introduction
Klebsiella pneumoniae is a Gram-negative pathogen responsible for a wide range of healthcare-acquired infections in genitourinary, respiratory, and gastrointestinal tracts, mostly in immunocompromised patients (Podschun and Ullmann, 1998; Paczosa and Mecsas, 2016). Polysaccharides capsule, lipopolysaccharides, siderophore-mediated iron acquisition systems, and fimbrial adhesins are among the virulence determinants of K. pneumoniae (Paczosa and Mecsas, 2016). Currently, this bacterium has become a worldwide public health threat due to the emergence of hypervirulent and antibiotic-resistant strains of K. pneumoniae causing various types of invasive infections (Lee et al., 2017; Rodriguez-Villar et al., 2019; Choby et al., 2020). In addition, the increasing number of severe community-acquired infections in healthy individuals has emphasized the importance of studying the virulence mechanisms that determine the pathogenicity of K. pneumoniae.
Historically known for being responsible for pneumonia, K. pneumoniae has been considered an important causative agent of urinary tract infections (UTIs) (Paczosa and Mecsas, 2016). The successful colonization of the urinary epithelium by K. pneumoniae relies on the expression of fimbriae. These structures mediate adherence to urothelial cells and are also essential for biofilm formation by promoting adhesion of the bacterium to abiotic surfaces, such as urinary catheters (Stahlhut et al., 2012). The establishment of a biofilm structure provides the bacterium more resistant to the host defense system and increases colonization of the host urinary tract (Rosen et al., 2007, 2008a, b; Struve et al., 2008; Hannan et al., 2012; Stahlhut et al., 2012; Murphy et al., 2013). Fimbriae are also required for uropathogens to adhere to specific receptors on urothelial cells and invade the urinary epithelium (Oelschlaeger and Tall, 1997; Rosen et al., 2008b; Struve et al., 2008; Hannan et al., 2012; Murphy et al., 2013). Once in the intracellular environment, the uropathogen replicates to form biofilm-like intracellular bacterial communities (IBCs) (Hunstad and Justice, 2010). Later maturation of IBCs leads to epithelium exfoliation with consequent spread of the bacteria to other sites, initiating a new cycle (Rosen et al., 2007; Hunstad and Justice, 2010). Thus, this intracellular niche allows uropathogens to hide from the host immune system and represents a quiescent intracellular reservoir of bacteria that leads to recurrent episodes of UTI (Rosen et al., 2007; Hunstad and Justice, 2010).
Several types of fimbriae have been identified in K. pneumoniae, with types 1 and 3 fimbriae being the main and best characterized adhesive structures (Paczosa and Mecsas, 2016). Type 1 fimbriae are encoded by the fim gene cluster, composed of eight genes (fimAICDFGHK) (Rosen et al., 2008a,b; Struve et al., 2008). These fimbriae are made up of repeating subunits of the major fimbrial subunit protein FimA, with the adhesin molecule FimH located at the distal end of the structure (Rosen et al., 2008b). FimH has a binding affinity for mannose residues present on the bladder cells surface (Rosen et al., 2008b). Thus, type 1 fimbriae promote the adhesion and invasion of the pathogen in host cells and favor successful colonization (Rosen et al., 2008a,b; Struve et al., 2008). Type 3 fimbriae are encoded by the mrkABCD gene cluster. MrkA subunits make up the major structure of the type 3 fimbriae, with the MrkD adhesin located at the tip. Although the MrkD-specific cell surface receptor has not yet been identified, type 3 fimbriae promote adhesion to tracheal, lung, and kidney epithelial cells (Hornick et al., 1992), and are fundamental in the process of biofilm formation on biotic and abiotic surfaces (Struve et al., 2009). Both type 1 and type 3 fimbriae promote biofilm formation on urinary catheters and, therefore, play a significant role in colonization and persistence in the bladder in catheter-associated UTI (Stahlhut et al., 2012; Murphy et al., 2013).
K. pneumoniae controls the expression of fimbrial genes through transcriptional regulators (Rosen et al., 2008a; Wilksch et al., 2011; Wu et al., 2012; Wang et al., 2013; Ares et al., 2016; Lin et al., 2017), utilizing various environmental stimuli (Schroll et al., 2010; Wu et al., 2012; Lin et al., 2016, 2018) and according to specific anatomic sites (Struve et al., 2008, 2009). Phase variation mediated by invertible DNA elements (Struve et al., 2008; Wu et al., 2010), intracellular levels of cyclic di-GMP (Johnson and Clegg, 2010), iron availability (Wu et al., 2012) and DNA binding regulators (Rosen et al., 2008a; Wilksch et al., 2011; Wu et al., 2012; Wang et al., 2013; Ares et al., 2016; Lin et al., 2017) are among the known mechanisms that regulate expression of fimbriae in K. pneumoniae (Clegg et al., 2011). For instance, the expression of fim gene cluster is regulated by phase variation mediated by an invertible promoter element, named fimS element, whose orientation is switched by FimE- and FimB-recombinases (Struve et al., 2008). Besides this mechanism, the expression of K. pneumoniae fim gene cluster is also regulated by the transcriptional regulator FimK encoded by fimK, a gene found only in fim gene cluster of K. pneumoniae and absent in Escherichia coli (Rosen et al., 2008a; Wang et al., 2013). On the other hand, the expression of type 3 fimbriae has been attributed to extracellular iron levels and ferric uptake regulator (Fur) (Wu et al., 2012). Fur is a transcriptional regulator that modulates gene expression by complexing with ferrous iron and binding to regulatory sequences, named boxes Fur, located in the promoter region of target genes (Escolar et al., 1999).
In its classical mechanism of action, Fur acts as a transcriptional repressor that utilizes ferrous iron as corepressor to bind on Fur box consensus sequence and to block RNA polymerase entry on the promoter region, thus leading to transcription repression of the target genes (Escolar et al., 1999). However, reports in the literature have indicated not only that the Fur-iron complex can also function as a transcriptional activator, but also that both repression and activation of transcription can be achieved by Fur in its holo or apo forms—i.e., complexed or not complexed with ferrous iron, respectively (Carpenter et al., 2009; Troxell and Hassan, 2013; Seo et al., 2014; Deng et al., 2015). This alternative mode of action has been described in some pathogenic bacteria, including Helicobacter pylori (Delany et al., 2001; Gilbreath et al., 2012), Neisseria meningitides (Delany et al., 2004; Yu and Genco, 2012), Salmonella enterica (Teixidó et al., 2011), Campylobacter jejuni (Butcher et al., 2012), Staphylococcus aureus (Deng et al., 2012), and Bacillus subtilis (Pi and Helmann, 2017).
In addition to fim and mrK, the genome of Klebsiella pneumoniae presents at least seven other fimbrial gene clusters still poorly characterized (Wu et al., 2010; Khater et al., 2015). Recently, Alcántar-Curiel et al. (2013) identified and characterized an operon homologous to the Escherichia coli common pilus (ECP) in the K. pneumoniae genome. The fimbriae encoded by the ecp operon, which comprises the ecpABCDE genes, have been associated with epithelial cell infection in vitro and in biofilm-associated bacterial communities (Alcántar-Curiel et al., 2013). A fimbrial gene cluster not yet characterized is kpf gene cluster, first described by Wu et al. (2010) on K. pneumoniae NTUH-K2044. The synthesis of this broad repertoire of fimbrial structures requires a precise and coordinated control that involves specific regulatory proteins. This control guarantees the production of these structures only when necessary, avoiding the uncontrolled expression of fimbriae and unnecessary energy expenditure by bacteria. Besides resulting in unnecessary consumption of energy, the uncontrolled expression of the fimbriae may result in bacterial clearance by the immune cells (Athamna et al., 1991; Przondo-Mordarska et al., 1991).
In the present study, we describe the transcriptional regulator of kpf gene cluster, a poorly characterized cluster of fimbrial genes encoding type 1-like fimbriae. We also demonstrate that KpfR regulator plays an important role in the pathogenicity of K. pneumoniae in murine urinary tract.
Materials and Methods
Bacterial Strains, Eukaryotic Cell Lines, and Growth Conditions
For this study, we used Klebsiella pneumoniae strain #8 (UKP8), a clinical strain isolated from a human patient with urinary tract infection. PCR genotyping using primers described elsewhere (Turton et al., 2008) indicated that the strain UKP8 presents the capsular serotype K2 (data not shown). Bacterial cells were routinely grown in Lysogeny Broth (LB; BD, United States) at 37°C with shaking at 200 rpm, or on LB agar plates in static cultures. Bacterial growth was monitored by measuring the optical density of the cultures at a wavelength of 600 nm (O.D.600 nm), using the GeneQuant Spectrophotometer (GE Healthcare). For growth under iron-replete and iron-limiting conditions the LB medium was supplemented with FeSO4 (Sigma-Aldrich) or the iron chelator Dipyridyl (Sigma-Aldrich), respectively.
For coculture assays, we used the human bladder epithelial cell line T24, purchased from the Bank of Cells of Rio de Janeiro (BCRJ, cell line with code number 0231). T24 cells were cultivated at 37°C in an atmosphere of 95% relative humidity at 5% CO2, in McCoy’s 5A medium (Thermo Fisher Scientific®) with 10% fetal bovine serum (FBS) (Thermo Fisher Scientific®) and subcultivated twice a week at a ratio of 1:8.
Identification of Regulatory Binding Sites on kpfR Gene Promoter
Bioinformatics analyses were performed to identify putative regulatory binding sites on kpfR promoter. These analyses were conducted on the genomic sequence of kpfR on Klebsiella pneumoniae strain ATCC 700721/MGH 78578 (GenBank accession number CP000647.1). To identify Fur-binding sites on kpfR gene we employed a theoretical approach (Ferraz et al., 2011) previously adapted to K. pneumoniae (Gomes et al., 2018). To identify the putative transcription start site and possible Sigma factor positions on the promoter region of kpfR gene we used the web-based programs Neural Network Promoter Prediction (available at https://www.fruitfly.org/seq_tools/promoter.html) and BPROM-Prediction of Bacterial Promoters (available at http://www.softberry.com/berry.phtml?topic=bprom&group=programs&subgroup=gfindb; Solovyev and Solovyev, 2011), respectively. These predictive analyses were performed on 100 nucleotides upstream to the initial codon of kpfR gene.
To determine whether kpfR cotranscribe with the genes from kpf cluster, reverse-transcription-PCRs were performed using cDNAs synthesized from total RNA extracted from K. pneumoniae and primer pairs spanning kpfR to kpfA and kpfA to kpfD genes (see Supplementary Table S1). The resulting amplicons were analyzed by agarose gel electrophoresis. Primers were designed using Primer3 version 0.4.0 web-program (available at http://bioinfo.ut.ee/primer3-0.4.0/; Rozen and Skaletsky, 2000).
Complementary oligonucleotides containing the sequences of the putative Fur boxes were annealed to form double-stranded DNA probes. These DNA probes were cloned into high copy number pGEM®-T Easy vector (Promega) and the recombinant vectors were used in Fur titration assay (FURTA) and DNA Electrophoretic Mobility Shift Assay (EMSA), in order to validate the Fur protein interactions with the putative Fur boxes.
FURTA was performed with Escherichia coli strain H1717, which carries the lacZ reporter gene under the control of the Fur-regulated fhuF gene promoter. When transformed with multicopy plasmids cloned with functional Fur box, H1717 strain will appear red on MacConkey lactose agar plates (Lac+ phenotype) because the high number of newly introduced Fur boxes will titrate the Fur repressor from fhuF:lacZ fusion, thus releasing the transcription of lacZ. On the other hand, if H1717 strain is transformed with a vector cloned with a non-functional Fur box, the Fur repressor will remain bound to the fhuF promoter region, and the lacZ reporter gene is not expressed, rendering colorless E. coli H1717 colonies on MacConkey lactose agar plates (Lac– phenotype). FURTA was conducted as previously described (Johnson and Clegg, 2010). E. coli strain H1717 was transformed with pGEM®-T Easy vector cloned with the DNA probe containing the sequences of the putative Fur box identified on kpfR gene. The transformants were then plated onto MacConkey lactose agar containing 100 μg/mL ampicillin and 100 mM of iron sulfate (FeSO4) (Sigma). After 18 h of incubation at 37°C, the functionality of the putative Fur box on kpfR was assessed according to the Lac ± phenotype (i.e., the color of the H1717 colonies). E. coli strain H1717 transformed with circular pGEM®-T Easy vector alone (i.e., with no insert) was used as negative control, whereas H1717 strain transformed with vector cloned with the previously validated Fur box of the K. pneumoniae entC gene (Carpenter et al., 2009) was used as a positive control.
EMSA was conducted essentially as described by Gomes et al. (2018). DNA probes for EMSA were obtained by PCR amplifying the pGEM®-T Easy vector cloned with the putative Fur identified on kpfR gene, using the universal M13 primers (Gomes et al., 2018). The resulting PCR product of 285 base pairs was then used as probes on the binding reactions, along with the previously purified His-tagged recombinant Fur protein from K. pneumoniae. The negative control consisted of a 254 base pairs DNA fragment without Fur box sequence, obtained by PCR amplification of a pGEM®-T Easy vector without insert (i.e., vector not cloned with the putative Fur boxes) using M13 primers. Reactions were performed with 100, 250, and 500 ηM of His-Fur protein, previously equilibrated for 10 min on ice in 10 μL reaction volume containing 1× binding buffer (10 mM Tris, 50 mM KCl, 1 mM DTT, pH 7.5), 0.5 mM MgCl2, 0.5 mM MnSO4 and 2.5% (v/v) glycerol. Next, 50 ηg of the DNA probes was added and the mixture was incubated for 20 min on ice. EMSA was also performed under divalent cation-free conditions by adding EDTA to a final concentration of 2 mM in the above reaction mixture. Samples were loaded onto a 2% (w/v) agarose gel prepared with 1× Bis-Tris borate buffer containing 0.1 mM MnSO4. After 30 min of electrophoresis, the gels were stained with ethidium bromide solution (0.5 μg/mL) for 15 min, and the DNA bands were visualized and recorded under a digital photodocumentation system (Biorad).
Construction of the kpfR Mutant Strain
Knockout of the kpfR gene from K. pneumoniae UKP8 was constructed using the TargeTron Gene Knockout System (Sigma-Aldrich), according to the manufacturer’s instructions. This system is based on site-specific and not random disruption of the gene of interest by insertion of group II intron harboring a kanamycin resistance gene (kanR).
Firstly, a computer algorithm at TargeTron Design website (Sigma-Aldrich) was used to identify potential target sites for intron insertion on the kpfR coding region. The most efficient target site was selected, based on the lowest E-value predicted by TargeTron Design algorithm (see Supplementary Table S2). The TargeTron Design algorithm also provided the primers used in PCR reactions to mutate (re-target) the RNA segment of the intron (see Supplementary Table S3). The 350 pb PCR product was digested and ligated into chloramphenicol-resistant pACD4K-C vector provided by the manufacturer (Sigma-Aldrich). The resulting recombinant pACD4K-C vector was transformed into Escherichia coli DH5a by heat shock, and clones of the recombinant vector were obtained using Wizard Plus SV Minipreps DNA Purification kit (Promega). The pACD4K-C vector contains a T7 promoter and, therefore, requires T7 RNA Polymerase to express the retargeted intron that will disrupt the target gene. Since Klebsiella pneumonia does not express T7 RNA Polymerase, it was necessary to use the ampicillin-resistant pAR1219 vector (Sigma-Aldrich) in co-transformations because this vector contains the T7 RNA Polymerase gene under the control of the IPTG-inducible lac UV5 promoter. Chemically competent K. pneumoniae UKP8 cells were co-transformed with pAR1219 and the pACD4K-C recombinant vectors and cultured overnight in LB containing 100 μg/mL of ampicillin and 25 μg/mL of chloramphenicol. On the next day, the culture was diluted 1:50 in fresh LB containing ampicillin and chloramphenicol, and cell growth was monitored until they reached O.D.600 nm of 0.2. At this point, to induce intron expression and insertion, 1 mM (final concentration) of IPTG was added to the medium and the culture was incubated at 30°C for 30 min. After induction, cells were harvested by centrifugation at 14,490 g for 1 min, resuspended in antibiotic-free LB broth, and incubated again for 1 h at 30°C. The cells were then plated on LB agar supplemented with 25 μg/mL kanamycin and grown at 30°C for 2–3 days. Knockout bacteria were selected from kanamycin-resistant colonies and the insertion of the intron RNA harboring kanR gene was confirmed by PCR using primers flanking the target insertion site in the coding region of the kpfR gene (Supplementary Table S1). Since the knockout is based on the insertion of kanR gene on kpfR, the K. pneumoniae UKP8 mutant strain was renamed kpfR::kanR.
A complemented strain, named , was obtained by introducing the kpfR gene back into the mutant strain. For this, a DNA fragment comprising the entire coding region of kpfR plus approximately 550 bp of 3’ and 5’ flanking regions (Supplementary Table S4) was inserted on pCR2.1-TOPO vector (Invitrogen) previously cloned with erythromycin-resistance gene. Chemically competent kpfR::kanR was transformed with the recombinant vector and plated on LB agar supplemented with 50 μg/mL erythromycin. Complemented strains were recovery by screening erythromycin-resistant colonies.
Growth Experiments and Phenotypic Assays
Initially, the growth of wild-type UKP8 and mutant kpfR::kanR strains was assessed to investigate whether the lack of KpfR compromises bacterial growth. Strains were separately inoculated into LB medium and grown until saturation (overnight) at 37°C while shaking. The next day, the culture was diluted 1:200 in fresh LB and the bacterial growth was monitored every 15 min by measuring the O.D.600 nm. Growth curves were constructed by plotting the O.D.600 nm values against time.
Ten microliters of saturated cultures of UKP8, kpfR::kanR, and were plated on blood agar (Trypticase Soy Agar plates supplemented with 5% sheep blood) for 18 h at 37°C to check for macroscopic differences in each strain’s morphology.
Motility assays were carried out in triplicate for each K. pneumoniae strains, according to protocol described elsewhere with minor modifications (Losensky et al., 2015). In brief, 10 μL of bacterial cultures standardized at O.D.600 nm of 0.4 was inoculated in the center of soft LB agar plates (prepared with 0.8% bacteriological agar) and incubated at 37°C for 120 h. After this, the extension of the colonies diameter was measured in cm.
Transmission Electron Microscopy (TEM) was carried out for visualization of fimbriae production by the K. pneumoniae strains. Bacterial cells were grown statically on LB agar plates at 37°C for 18 h and resuspended in 500 μL of PBS buffer. A droplet of the cell suspension was spotted on Formvar coated carbon-reinforced copper grid (400 mesh) and incubated at room temperature for 2 min. Grids were negatively stained using a drop of 1.25% phosphotungstic acid pH 6.5 (Sigma-Aldrich) filter sterilized through a 0.22 μm pore-size filter membranes (Millipore). The staining lasted 15 s and then the grids were air-dried on a piece of filter paper. Samples were examined and electron micrographs were obtained with a Zeiss LEO 906 Transmission Electron Microscope operated at 60 kV.
Biofilm formation assays were conducted in polyvinylchloride (PVC) microplates, following a protocol described elsewhere with minor modifications (O’Toole and Kolter, 1998). This protocol is based on the ability of bacteria to form biofilm on PVC microplates and on the ability of crystal violet to stain bacterial cells but not PVC. Strains were grown until saturation at 37°C under shaking. Bacterial cells from saturated cultures were harvested by centrifugation and resuspended in LB broth to a final concentration of 106 cells/mL. 5 μL of this suspension were added on flat-bottom 96-well PVC microplates containing 150 μL of LB and incubated at 37°C for 1, 5, and 10 h under static conditions. After incubation, the culture medium was removed and the wells were gently washed twice with sterile water to remove unattached cells. Next, 25 μL of 1% crystal violet was applied in the wells and the plates were incubated for 15 min at room temperature. After incubation, the excess of dye was removed and the wells were washed with deionized water. Then, the remaining crystal violet staining of the adherent cells was solubilized with absolute ethanol and the absorbance of the ethanol containing the eluted dye was measured at O.D.600 nm in a spectrophotometer. The biofilm formation assays were repeated at least three times.
Yeast agglutination assays were performed to investigate the expression of fimbriae by the K. pneumoniae strains. Assays were carried out in triplicate for each K. pneumoniae strain and were conducted on Kline concavity slides, according to protocol previously described (Roe et al., 2001; Schembri et al., 2004). Bacterial strains grown on LB agar plates up to 72 h were resuspended in PBS buffer and the concentration was standardized at O.D.600 nm of 0.6. Bacteria were then mixed with an equal volume of 5% (wt/vol) suspension of Saccharomyces cerevisiae cells (Sigma-Aldrich) prepared in PBS buffer. The time required for agglutination and its intensity were documented. The agglutination of the yeast cells is specifically mediated by type 1 fimbriae since these fimbriae have great affinity for mannose, a highly abundant residues on yeast cell-surface. Therefore, the assays were also performed in the presence of 5% D-(+)-Mannose (Sigma-Aldrich) to confirm if the agglutination was indeed mediated by the type 1 fimbriae.
To visualize capsular polysaccharide (CPS) produced by UKP8, kpfR::kanR, and , strains were cultured under the same growth conditions, stained with India ink, and analyzed under optical microscopy. The presence of capsule is indicated by a negative staining area around the bacteria. CPS production was quantified by extracting and measuring glucuronic acid, a significant component of the K. pneumoniae capsule, using a colorimetric assay previously described (Lin et al., 2012; Ares et al., 2016). Briefly, 500 μL of K. pneumoniae cultures were mixed with 100 μL of 1% Zwittergent 3–14 detergent (Sigma-Aldrich) in 100 mM citric acid (pH 2.0) and then incubated for 20 min at 50°C. After centrifugation for 2 min at 14,490 g, 250 μL of the supernatants were transferred into new tubes, 1 mL of cold absolute ethanol was added, and the mixtures were incubated for 20 min at 4°C for CPS precipitation. After centrifugation at 14,490 g for 2 min, the supernatants were discarded and the pellets were allowed to dry. The dried pellets were dissolved in 200 μL of distilled water and then 1,200 mL of 12.5 mM tetraborate in concentrated H2SO4 were added. Next, the mixtures were vigorously vortexed, heated in a boiling-water bath for 5 min, and cooled down, before the addition of 20 μL of 0.15% 3-hydroxydiphenol (Sigma-Aldrich) prepared in 0.5% NaOH. Then, the absorbance was measured in a spectrophotometer at a wavelength of 520 nm. The glucuronic acid concentration in each sample was determined from a standard curve of glucuronic acid (Sigma-Aldrich) and expressed in micrograms/109 CFU. Quantification of CPS production was performed from five independent cultures of the strains.
Epithelial Cell Adhesion, Invasion, and Intracellular Replication Assays
Coculture assays were performed as previously described (Elsinghorst, 1994; Oelschlaeger and Tall, 1997), with some modifications. Saturated overnight cultures of UKP8 and kpfR::kanR were inoculated on fresh LB medium and incubated at 37°C with shaking until they reached the mid-logarithmic growth phase (O.D.600 nm of 0.4). Bacteria were added to an 80% confluent T24 cell monolayer seeded into 12-well plates at a multiplicity of infection (MOI) of 200 bacteria per host cell, and the infected monolayers were incubated at 37°C for three different periods, specific for each assay. For adhesion assay, infected T24 cells were incubated for 30 min, washed three times with phosphate-buffered saline (PBS buffer) to remove non-adhered bacteria, and lysed by adding 0.1% Triton X-100 diluted in PBS. Serial dilutions of the lysate were plated on blood agar and incubated overnight at 37°C for counting bacterial colony-forming units (CFUs). For invasion assay, infected T24 cells were initially incubated for 4 h to allow invasion of the bacterial strains into T24 cells. Plates were washed once with PBS buffer and incubated again for an additional 1 h in fresh culture medium containing the antibiotic gentamicin (25 μg/mL) to eliminate extracellular bacteria (both planktonic and adhered). The plasma membrane of host cells is impermeable to gentamicin. Therefore, after treatment with this antibiotic, only extracellular bacteria will be eliminated, while those present in the intracellular environment will be preserved. After treatment with gentamicin, the T24 cells were washed, lysed with Triton X-100 and serial dilutions of the lysate were plated on blood agar for counting of CFUs. For intracellular replication assay, infected T24 cells were initially incubated for 4 h to allow invasion of the bacterial strains. Plates were washed once with PBS buffer and incubated again for 24 h in fresh culture medium containing gentamicin at 10 μg/mL. Then, the T24 cells were washed, lysed with Triton X-100 and serial dilutions of the lysate were plated on blood agar for CFUs counting. The integrity of the T24 cells during the coculture assays was assessed by the standard exclusion protocol of trypan blue 0.4% (Thermo Fisher Scientific®).
Mouse Model UTI
For mouse urinary tract infection analyzes were used 6–8 weeks old female BALB/c mice (CEMIB, UNICAMP, Brazil). Infections were carried out by transurethral inoculation as previously described (Thai et al., 2010; Reis et al., 2011). UKP8 and kpfR::kanR strains were grown on LB agar plates at 37°C for 18 h and resuspended at a concentration of ∼5 × 108 CFU/mL. Fifty microliters of the bacterial suspension were inoculated by transurethral catheterization in mice anesthetized intraperitoneally with ketamine and xylazine. Mice were euthanized at the indicated times, and urine and bladders were aseptically collected and processed for histology and CFU titration. Serial dilutions of urine and homogenized bladders were plated on blood agar and incubated overnight at 37°C for counting of CFUs. Whole bladders were fixed in 10% neutral buffered formalin, embedded in paraffin, and 5 μm-thick sections were stained with hematoxylin and eosin (H&E).
Mice experimental procedures were conducted in accordance with guidelines of the Brazilian College for Animal Experimentation (COBEA) and received prior approval by the Ethics Committee on the Use of Animals in Research of Universidade São Francisco (CIAEP/CONCEA No. 01.0226.2014, protocol number 004.04.2015).
RNA Extraction and RT-qPCR Analyses
Gene expression analysis was performed using Reverse Transcription Quantitative real-time Polymerase Chain Reaction (RT-qPCR). The expression of kpfR under iron-replete and iron-limiting conditions was investigated as previously described (Gomes et al., 2018). Briefly, K. pneumoniae strains were grown in LB broth at 37°C with shaking at 200 rpm until they reached the mid-logarithmic growth phase (O.D.600 nm of 0.4). At this point, ferrous iron (100 μM of FeSO4, final concentration) or an iron chelator (100 μM of 2,2’-Dipyridyl, final concentration) were added and the cells were incubated for 1 h. After 1 h incubation, bacterial cells were harvested by centrifugation and the pellets were resuspended in RNAprotect® Bacteria Reagent for RNA stabilization. All culture conditions were performed at least twice. Control condition consisted of K. pneumoniae cells grown in LB medium without supplements.
The expression of fimbrial genes and galF, the first gene of the capsular polysaccharide synthesis (cps) gene cluster, were also assessed on UKP8 and kpfR::kanR strains grown on blood agar plate. For this, bacterial colonies from each strain were individually collected from the blood agar plates, and the cell pellets were resuspended in RNAprotect® Bacteria Reagent until the moment of the RNA extraction. Total RNA from the cell pellets was extracted using RNeasy Protect Bacteria Mini Kit (Qiagen), following the manufacturer’s protocol. An on-column DNase digestion with the RNase-free DNase Set (Qiagen) was performed to remove genomic DNA contamination in RNA samples.
To assess gene expression of the bacterial strains during coculture with T24 bladder epithelial cells, the adhesion and invasion assays were conducted as previously described in section “Materials and Methods,” the only exception that the T24 cells were seeded in 6-well plates. After the incubation periods, extracellular bacteria were removed by washing with PBS buffer and T24 cells were collected for the extraction of bacterial total RNA. RNA was extracted using MaxTM Bacterial RNA Isolation kit (Ambion), following the manufacturer’s instructions. MICROBEnrichTM kit (Ambion) was applied, in order to promote the selective removal of total RNA from T24 cells, preserving bacterial RNA. Total RNA extracted was further treated with DNAse.
After treatment with DNAse, 0.5–1 μg of total bacterial RNA was used for cDNA synthesis by using the ThermoScriptTM RT-PCR System for First-Strand cDNA Synthesis Kit (Invitrogen) according to the manufacturer’s instructions. The synthesized cDNAs were used in RT-qPCR reactions done in triplicates using the Platinum® SYBR® Green qPCR SuperMix-UDG Kit (Invitrogen) on 7300 Real-Time PCR System equipment (Applied Biosystems). Data were normalized using rho and recA as endogenous genes, which encode transcription termination factor and recombinase A, respectively (Song and Abraham, 2008). The relative expression levels of the selected genes were calculated by the comparative critical threshold (2–ΔΔCT) method (Livak and Schmittgen, 2001). GraphPad Prims 7.00 (GraphPad Software, Inc.) was used for the statistical analyses. Differences on the expression levels were evaluated by Student’s t-test, and differences with p ≤ 0.05 were considered statistically significant. Primers used on RT-qPCR reactions are listed in Supplementary Table S5 and were designed using Primer3 version 4.1.0 web-program.
Results
kpfR Is a Fur-Regulated Gene That Encodes the Transcriptional Regulator of kpf Gene Cluster
The kpf gene cluster comprises 4 genes designated as follow: kpfA, encoding the major pilin subunit, kpfB, encoding a chaperone, kpfC, encoding an usher, and kpfD, encoding an adhesin. The first gene adjacent to kpf cluster at 5’ extremity encodes a putative helix-turn-helix transcriptional regulator. To determine whether this adjacent gene belongs to kpf cluster, reverse-transcription-PCRs were performed using cDNA and primer pairs spanning the entire cluster and confirmed that kpf cluster and the adjacent gene are co-transcribed as a single polycistronic transcript (Figure 1A). Since the adjacent gene encodes a putative helix-turn-helix transcriptional regulator, and following Wu et al. (2010) nomenclature, we designated it kpfR gene.

Figure 1. The kpf gene cluster encoding type 1-like fimbriae is regulated by Fur. (A) kpf cluster comprises kpfR, kpfA, kpfB, kpfC, and kpfD genes and is transcribed as a polycistronic mRNA. cDNA synthesized from total RNA of K. pneumoniae was used in PCR reactions using the primer pairs represented in the scheme. The resulting amplicons were analyzed by agarose gel electrophoresis and confirmed the predicted sizes of 847 and 3,773 base pairs (bp). The red spot on the scheme indicates the putative Fur box sequence identified on kpf cluster. (B) Partial sequence of kpfR showing the initial codon (ATG, double underlined) and the Fur-binding sequence located inside the coding region of kpfR (highlighted in blue). Also indicated on the promoter region of the cluster are the –35 (in green) and –10 (in yellow) domains of the housekeeping Sigma factor 70, the predicted transcription initiation site at position + 1 (cytosine in red), and a putative IHF-binding consensus sequence (in gray; lowercase nucleotides are not identical to the consensus sequence). (C) Fur Titration Assay (FURTA) validated the Fur box on kpfR of K. pneumoniae, as indicated by the E. coli H1717 red colonies on MacConkey plates, similar to the FURTA-positive control (Lac+). (D) DNA Electrophoretic Mobility Shift Assay (EMSA) confirms the direct interaction of the K. pneumoniae Fur protein on the putative Fur box found on kpfR. Fur-DNA probe complexes are formed (mobility shifts, indicated by closed arrowheads) as increased concentrations of purified His-Fur protein from K. pneumoniae were incubated with the probes (DNA fragment containing the Fur box of kpfR). Fur interaction depends on divalent cations, since the addition of EDTA chelator abolished the mobility shift of the probe (open arrowheads). No mobility shift is observed in the EMSA with the negative control (DNA probe without Fur box), indicated by open arrowheads). (E) RT-qPCR analyses showed that Fur represses the expression of the kpfR gene on K. pneumoniae cells cultured under iron-depleted condition when compared to the control condition (bacteria cultured in LB medium only). Since kpfR belongs to the kpf gene cluster, Fur modulates the expression of the entire cluster according to the availability of iron in the culture medium. *p ≤ 0.05.
To better understand the expression regulation of kpf, bioinformatic analyses were employed to identify regulatory domains in the promoter region of the cluster. Thus, a putative Fur box sequence was identified 100 nucleotides downstream to start codon of the kpfR gene (Figure 1B). The long-distance between the Fur box and the promoter region of the kpf cluster led us to investigate additional regulatory elements on the adjacent sequences. As indicated in Figure 1B, a putative binding site of the DNA bending protein IHF (integration host factor) was identified at the intervening region between the promoter and the Fur box. This putative site closely resembles to the consensus recognition sequence of IHF, but its functionality was not validated in the present study.
The putative Fur box identified inside the coding region of kpfR was validated by Fur Titration Assay (FURTA) and DNA Electrophoretic Mobility Shift Assay (EMSA). Figure 1C shows the FURTA-positive control (Lac+, red colonies), the FURTA-negative control (Lac–, colorless colonies) and the validation of the Fur box on kpfR, indicated by the red colonies. This result confirms that Fur regulator from E. coli H1717 was able to bind in vivo to the cloned putative Fur box from kpfR of K. pneumoniae, rendering the colonies with red color. Next, EMSA was performed to verify the direct interaction of K. pneumoniae Fur protein on the putative Fur box found on kpfR. As shown on Figure 1D, a mobility shift of the DNA probes containing the putative Fur box was observed when increasing concentrations of K. pneumoniae purified His-Fur protein was incubated with these probes. Besides, the addition of the divalent cations chelator EDTA abolished the mobility shift of the probe, indicating that divalent cations are required for Fur interaction. No mobility shift is observed when K. pneumoniae Fur protein is incubated with a DNA probe without Fur box sequence.
Taken together, our results confirm that Fur protein recognizes and binds to the Fur box on kpfR of K. pneumoniae, indicating that the expression of kpfR seems to be modulated by Fur transcriptional regulator in an iron-dependent manner. To further investigate how Fur modulates the expression of kpfR, RT-qPCR analyses were performed on K. pneumoniae cells kept under iron-replete and iron-limited conditions. As displayed on Figure 1E, the iron-replete condition presents no changes in the expression pattern of kpfR gene when compared to the control condition. On the other hand, kpfR is repressed under condition of iron scarcity. These results show that Fur modulates the expression of kpfR gene according to the availability of iron in the culture medium. Thus, kpfR belongs to the Fur regulon of K. pneumoniae.
kpfR Disruption Modified the Bacterial Morphology and Enabled a Rudimentary Motility
To evaluate the role of KpfR as regulator of morphology and virulence of Klebsiella pneumoniae, a mutant (kpfR::kanR) was generated (as described in section “Materials and Methods”). The mutant strain exhibits prolonged growth at the lag phase, leading to delayed entry into the logarithmic (log) phase of growth (see Supplementary Figure S1). Nonetheless, once reaching the log phase, the mutant strain displayed a growth rate similar to that observed in the wild-type strain. Slight differences in growth patterns were observed among the UKP8 wild-type strain and the complemented strain; i.e., the mutant strain transformed with a kpfR-carrying vector.
Macroscopic morphologies of wild-type UKP8 and mutant kpfR::kanR strains were investigated on blood agar plates. As shown on Figure 2A, the wild-type strain exhibited circular shape, high elevation, entire margin, and smooth bright surface. On the other hand, the mutant kpfR::kanR strain presented irregular shape, flat elevation, lobed margin, and rough matte surface. In addition, kpfR::kanR colonies exhibited a slight dispersion on the plates. The morphological features observed on the mutant strain can be attributed to the lack of KpfR, because the pattern observed on the wild-type strain was fully reestablished on the complemented mutant strain ().

Figure 2. Bacterial colonies of K. pneumoniae cells depleted of kpfR show altered macroscopic morphologies and rudimentary motility. (A) Colony morphology of wild-type (UKP8), mutant (kpfR::kanR), and complemented () strains were investigated on blood agar plates. The altered pattern on the mutant strain was fully restored in the complemented strain, which exhibited a pattern similar to the wild-type. (B,C) In soft agar plates, kpfR::kanR cells exhibited a pronounced asymmetric dispersion on the surface of the plates when compared to UKP8, resembling a sort of rudimentary motility. Dispersion was only partially restored by complementation. The greater dispersion of the mutant colonies than the wild-type is statistically significant. *p ≤ 0.05.
The spreading behavior of kpfR::kanR colonies on blood agar plates prompted us to perform motility assays on soft agar plates. As displayed on Figure 2B, the wild-type UKP8 strains presented a slight and linear radial migration on soft agar plates. On the other hand, kpfR::kanR cells exhibited a more prominent and asymmetric dispersion on the surface of the plates. The complemented mutant strain was partially restored since it had a reduced migration extension when compared to both wild-type and mutant strains. The extent of the area in which the wild-type, mutant, and complemented strains migrated on the plates was measured, and confirmed that the mutant strain had a greater dispersion when compared to UKP8 and strains (Figure 2C). These results indicate that kpfR disruption provided a sort of rudimentary motility to the mutant bacteria.
Lack of KpfR Triggers the Production of Bacterial Surface Appendages, Enhances Yeast Agglutination and Biofilm Formation but Reduces Capsule Production
Since loss of kpfR rendered K. pneumoniae a prominent dispersion on agar plates, we evaluated the cell surface morphology of the wild-type, mutant, and complemented strains by Transmission Electron Microscopy (TEM). While UKP8 strain appears with no surface appendages, the cell surface of the kpfR::kanR cells was covered by numerous appendages similar to fimbriae or pili that are long and flexible with a varied size larger than 1 μm (Figure 3A). These numerous appendages were no longer seen on the surface of the complemented mutant strain ().
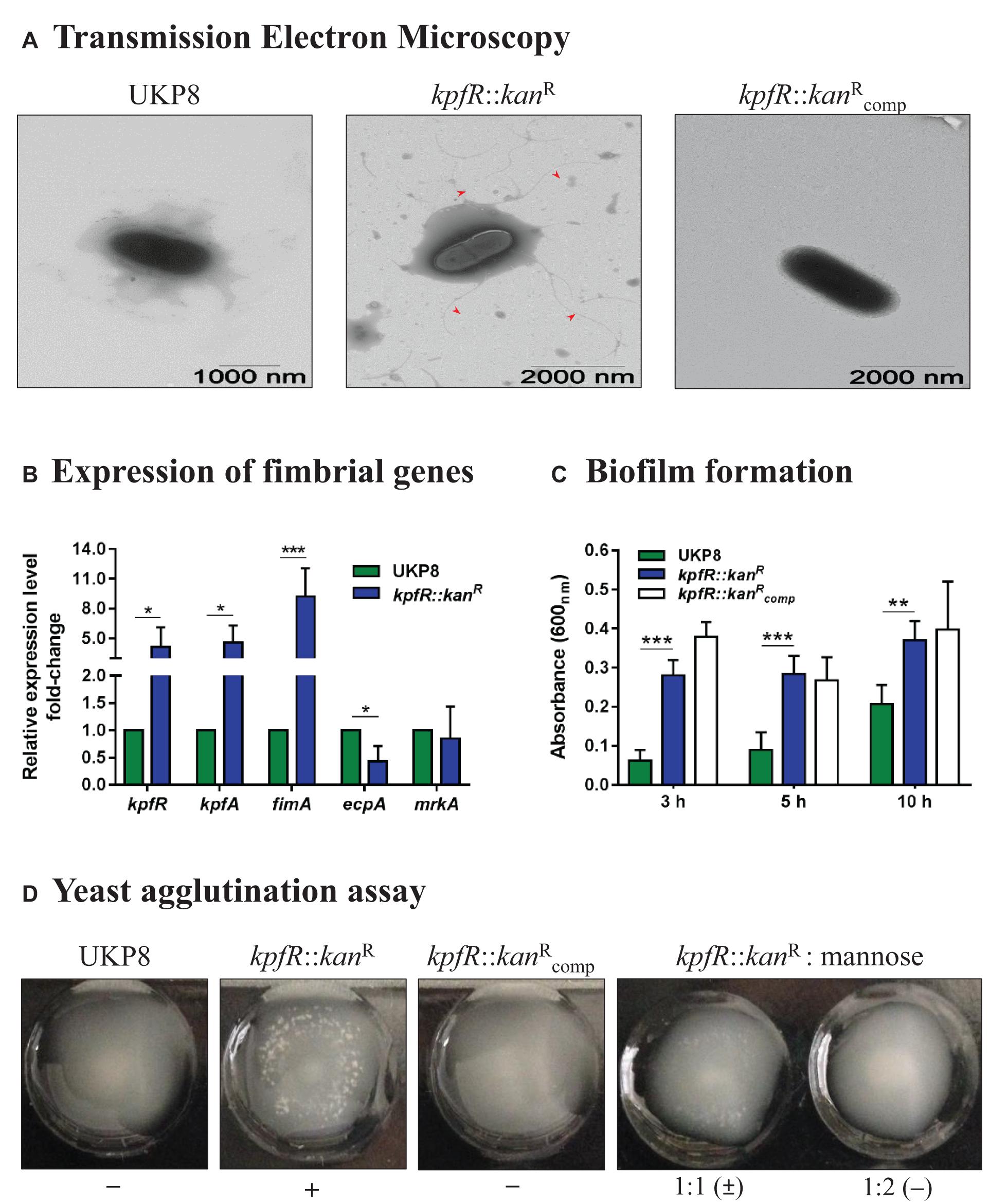
Figure 3. Lack of KpfR triggers the expression of type 1-like fimbriae, increases biofilm formation and yeast cells agglutination, and reduces production of capsule polysaccharide. (A) TEM analyses revealed kpfR::kanR mutant cells covered with numerous fimbriae-like appendages that were absent in wild-type and complemented cells. The images are representative of independent experiments. (B) This hyperfimbriated phenotype is corroborated by the fimbrial genes expression analyses. The ecpA gene, from ecp gene cluster, is downregulated, while fimA (fim gene cluster of type 1 fimbriae) and kpfR and kpfA genes (kpf gene cluster of type 1-like fimbriae) are up-regulated in the kpfR::kanR mutant cells. *p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.005. (C,D) Because of the hyperfimbriated phenotype, the mutant strain forms more biofilm than the wild-type and agglutinates yeast cells. This agglutination is mediated by type 1-like fimbriae because in the presence of mannose the mutant strain loses the ability to agglutinate yeast cells. Complementation failed to restore the biofilm formation exhibited by the wild-type strain, but fully restored the agglutination of yeast cells. The images are representative of independent experiments.
The presence of numerous appendages on kpfR::kanR cells led us to investigate the expression of fimbrial genes on UKP8 and kpfR::kanR cells grown on blood agar plate. Disruption of kpfR resulted in up-regulation of fimA, from type 1 fimbrial gene cluster fim, and kpfR and kpfA genes, from kpf gene cluster (Figure 3B). kpfR and kpfA presented a similar expression pattern, confirming the polycistronic transcription of these genes, as previously shown. Moreover, the up-regulation of kpfR in the cells with disrupted kpfR suggests that KpfR autoregulates its own expression. On the other hand, lack of KpfR regulator resulted in down-regulation of ecpA gene, from the ecp cluster, whereas mrkA gene, from type 3 fimbrial gene cluster mrk, presented unchanged expression pattern between wild-type and mutant strains.
To investigate the phenotypic effects of increased type 1-like fimbriae production due to KpfR absence, biofilm formation, and yeast agglutination assays were performed with the UKP8, kpfR::kanR, and strains. Biofilm formation by kpfR::kanR cells was significantly superior than UKP8 after 3, 5, and 10 h of incubation (Figure 3C). The complemented strain failed to restore the biofilm formation presented by the wild-type UKP8, since exhibited a phenotype similar to the mutant kpfR::kanR strain. Yeast agglutination assays allow specific detection of type 1 fimbriae because type 1 fimbrial adhesins have a great affinity for mannose-containing receptors on the yeast cell surface. UKP8 showed no yeast agglutination, whereas the kpfR::kanR promptly agglutinated the yeast cells (Figure 3D). The lack of agglutination observed on UKP8 was fully restored on the complemented strain. To determine if the agglutination pattern exhibited by the mutant strain was due to type 1 fimbriae expression, the assay was also performed in the presence of mannose prior to the addition of the yeast cells. In the presence of mannose, the mutant strain loses the ability to agglutinate yeast cells. These results reveal that the observed agglutination of yeast cells was indeed mediated by type 1 fimbriae.
As the lack of KpfR regulator resulted in increased expression of type 1-like fimbriae, we decided to investigate the production of capsular polysaccharide, another important virulence factor of the bacterial cell surface. Negative staining with India ink revealed no visible capsule in the kpfR:::kanR mutant strain as compared to the UKP8 wild-type strain (Figure 4A), while this phenotype was partially restored on the complemented strain. In addition, quantification of glucuronic acid revealed that the mutant strain presents 10-fold decrease in capsule production compared to that in the wild-type strain (Figure 4B). Capsule production by the complemented strain was partially restored to the levels observed in the wild-type strain. We next investigated the expression of the cps gene cluster on the wild-type and the mutant strains by determining the expression levels of galF using RT-qPCR analysis. galF is the first gene of the cps cluster and encodes an enzyme dedicated to the synthesis of sugar nucleotides, which are the precursors that will compose the repeating-units of the capsular polysaccharide. Consistent with the less capsule production by the kpfR::kanR mutant strain, RT-qPCR analysis revealed down-regulation of the capsule-producing gene galF on kpfR::kanR in comparison with the expression on UKP8 wild-type strain (Figure 4C).
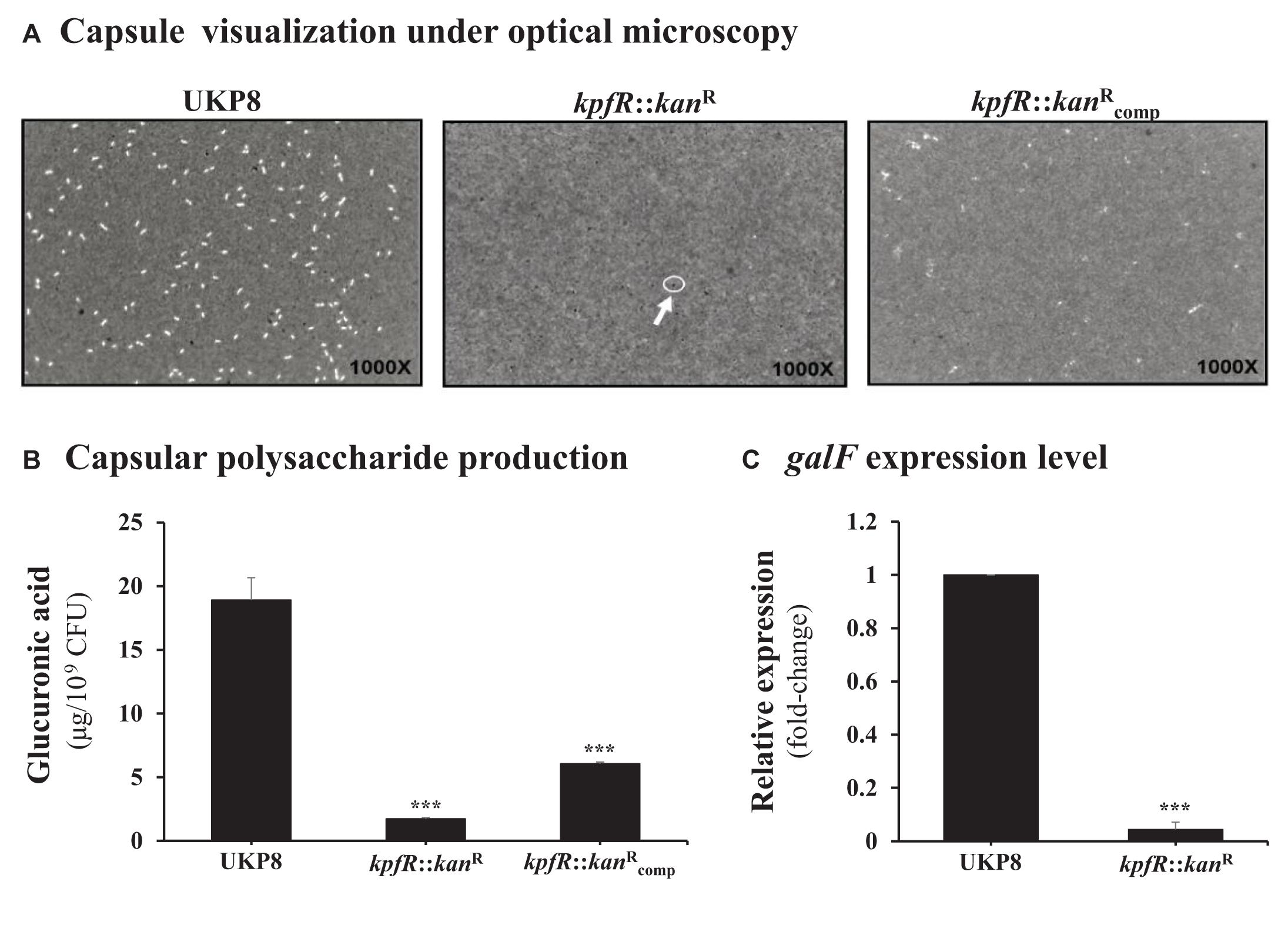
Figure 4. Capsule polysaccharide production is reduced in the kpfR::kanR mutant strain. (A) For visualization of capsule production, wild-type (UKP8), mutant (kpfR::kanR), and complemented () strains were cultured under the same growth conditions, stained with India ink, and analyzed by optical microscopy. The images are representative of independent experiments and the presence of capsule is indicated by a negative staining area around the bacteria. While capsule is observed in the wild-type and partially in the complemented strains, no visible capsule is observed on kpfR::kanR, suggesting a reduction in capsule biosynthesis on the hyperfimbriated mutant strain. (B) For quantification of capsule production, glucuronic acid, an important constituent of the K. pneumoniae capsule, was measured from capsular polysaccharides extracted of 0.5 mL cultures of each strain. The results are indicated as percentages of glucuronic acid production by mutant and complemented strain relative to the production by the wild-type UKP8 strain (set at 100%). The mutant strain presents reduced production of capsule compared to the wild-type strain. Although the complemented strain produces more capsules than the mutant strain, this increased production was not significantly different. Data are means of five independent experiments ± SE. ***p ≤ 0.005 vs. wild-type. (C) The less capsule production by the kpfR::kanR mutant strain correlates with the down-regulation of the capsule-producing gene galF on this strain. ***p ≤ 0.005 vs. wild-type.
kpfR::kanR Colonizes More Efficiently Bladder Epithelial Cells in vitro but Loses Resistance in vivo and Is Unable to Form IBC in Mouse Urinary Bladder
Considering fimbriae important mediators of bacterial adhesion and internalization into host cells, we investigated the role of kpfR gene in host-pathogen interactions by assessing the adhesion, invasion, and intracellular replication of UKP8 and kpfR::kanR strains on human bladder epithelial cell line T24. The data presented on Figure 5A indicate that the kpfR::kanR strain showed greater adhesion to T24 bladder cells than the wild-type UKP8 strain. Moreover, only the mutant strain was able to invade and replicate within T24 cells (Figure 5A). No significant changes in T24 cells viability were observed at a multiplicity of infection (MOI) of 200 bacteria:1 T24 cell and incubation of coculture for 24 h (data not shown). According to the results, the lack of KpfR repressor improved the ability of the mutant strain to adhere, invade and even replicate intracellularly in T24 bladder cells. We next evaluated the expression of kpfR and kpfA (cluster kpf) and fimA (cluster fim) in UKP8 and kpfR::kanR strains during adhesion to the T24 cells. As shown on Figure 5B, all three genes had the expression significantly induced in kpfR::kanR compared to UKP8.
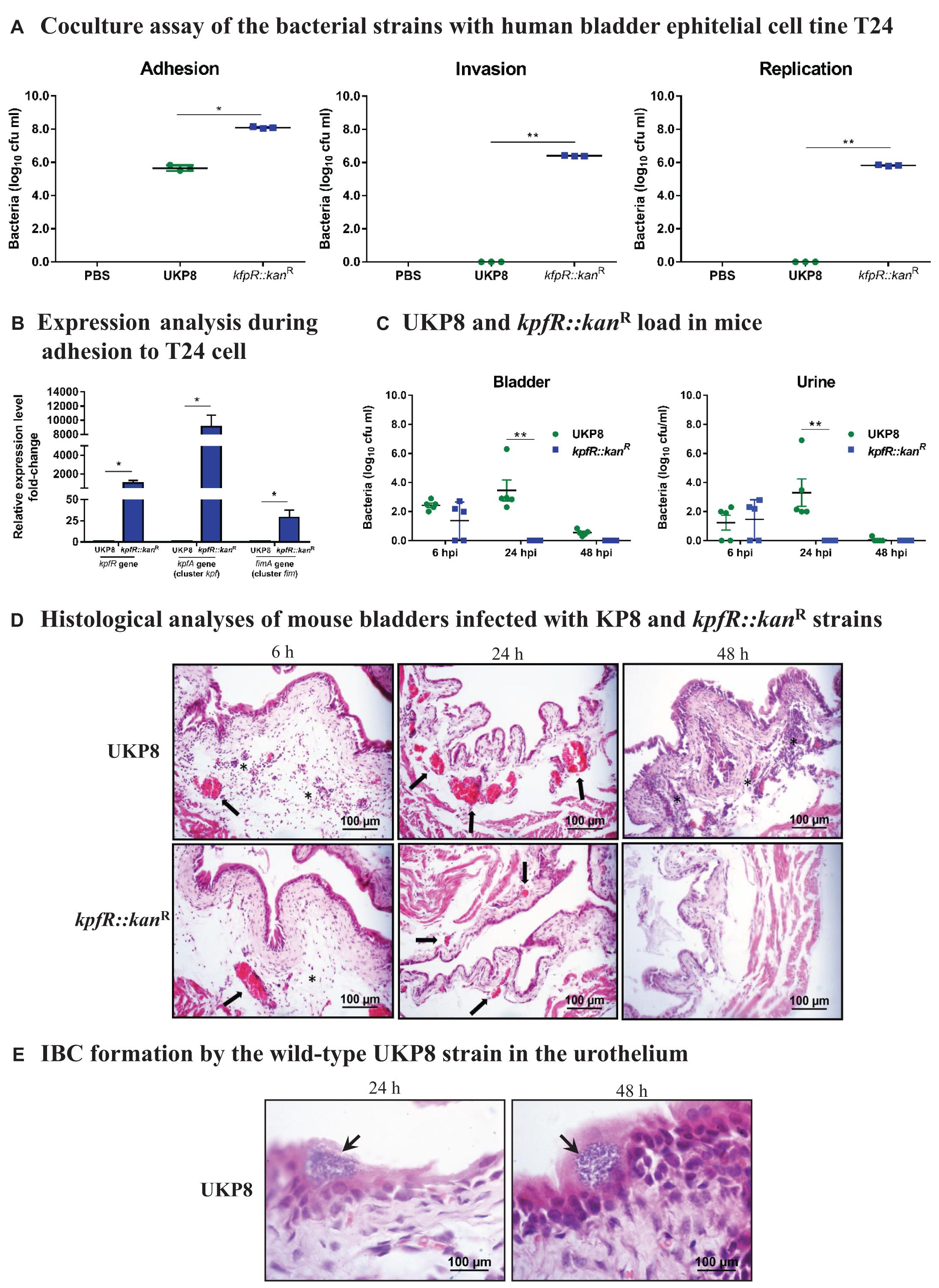
Figure 5. K. pneumoniae mutant for kpfR adheres more efficiently and replicates within bladder epithelial cells, but loses resistance in the mouse model of urinary infection. For the coculture assays, human bladder epithelial cell line T24 were inoculated with the wild-type and kpfR::kanR strains at an MOI of 200 to assess the adhesion, invasion, and intracellular replication of the strains on the host human bladder cell. The incubation periods are described in section “Materials and Methods.” (A) The mutant strain has greater adhesion to T24 bladder cells than the wild-type UKP8 strain and is the only strain able to invade and replicate within T24 cells. (B) During adhesion to T24 bladder cells, the kpfR::kanR mutant strain have increased expression of kpfR and kpfA (cluster kpf) and fimA (cluster fim) than the wild-type UKP8 strain, which may explain the improved ability of the mutant strain to adhere the T24 cells. *p ≤ 0.05; **p ≤ 0.01. For the mouse model of urinary infection, animals were inoculated by transurethral catheterization with 5 × 108 CFU/mL of UKP8 and kpfR::kanR strains. At the indicated times, the animals were euthanized, and urine and bladders were aseptically collected and processed for CFU enumeration and H&E staining. (C) Bacterial CFUs were counted on urine and bladder tissue after 6, 24, and 48 hpi with UKP8 and kpfR::kanR strains. In mice urine, the wild-type strain is recovered at 6 and 24 hpi, while the mutant kpfR::kanR strain is recovered only at 6 hpi. In the bladder tissue, the wild-type is present at 6, 24, and 48 hpi, whereas the mutant strain is also recovered only after 6 hpi. Mice are represented by symbols. Data were statistically analyzed by ANOVA test. **p ≤ 0.01. (D) Histological analyses of bladders infected with the wild-type and kpfR::kanR mutant strains show both strains triggering an inflammatory infiltrate consisting of neutrophils (asterisks) and a hyperemia (arrows) at 6 hpi. At 24 hpi, the hyperemia is more pronounced with the wild-type strain, and inflammatory cells migration is observed at 48 hpi only with UKP8, suggesting that at this period of infection the mutant strain has already been completely eliminated. Images are from an individual representative experiment. (E) Only the wild-type UKP8 is able to form biofilm-like intracellular bacterial communities (IBCs) in the urothelium of mice. Images from histological analyses of bladders infected with the wild-type at 24 and 48 hpi reveal IBCs (arrows) within superficial urothelial cells. Images are from an individual representative experiment.
To further evaluate the in vivo role of KpfR repressor in Klebsiella pneumoniae virulence, a mouse urinary tract infection model was used to compare the ability of UKP8 and kpfR::kanR strains to colonize the bladder of the animals (Figure 5C). At 6 hpi, there were no differences in UKP8 and kpfR::kanR load in mice urine and bladder. At 24 hpi, UKP8 was recovered from bladder tissue and urine, while kpfR::kanR was not present on these samples. At 48 hpi, UKP8 and kpfR::kanR were no longer found in urine, whereas only UKP8 was still present in mice bladder tissue.
Aliquots of urine were analyzed under an optical microscope (data not shown) and revealed red blood cells, leukocytes and scaly epithelium in urine of the animals infected with both UKP8 and kpfR::kanR strains at 6 hpi. At 24 hpi, a larger population of leukocytes and bacteria were observed only in the urine samples of mice infected with the wild-type strain. Urine of mice infected with kpfR::kanR during 24 h presented neither erythrocytes nor leukocytes.
The colonization experiments suggested that the lack of KpfR repressor confers a disadvantage on kpfR::kanR strain. To characterize these effects further, histological analyses were performed to obtain images of mouse bladders infected with the wild-type and kpfR::kanR mutant strains. As shown on Figure 5D, an inflammatory infiltrate consisting mainly of neutrophils and hyperemia of the bladder blood vessels was observed 6 hpi with both strains. At 24 hpi, the hyperemia was more prominent in the bladder tissue infected with UKP8 than kpfR::kanR, and an abundant migration of inflammatory cells was noticed only with UKP8 at 48 hpi (Figure 5D). In addition, only wild-type strain was able to form intracellular bacterial communities (IBCs) in the urothelium at 24 and 48 hpi (Figure 5E). Taken together, the data show successful bladder colonization and formation of IBC in urothelial cells by the wild-type UKP8 strain, while the kpfR::kanR mutant strain loses resistance and ability to survive in the host.
Discussion
Fimbriae are important mediators of UTIs caused by K. pneumoniae (Tinker and Clegg, 2000; Yeh et al., 2002; Rosen et al., 2008a; Stahlhut et al., 2012). Considering the broad number of fimbrial gene clusters on the K. pneumoniae genome, the expression of this range of fimbrial structures demands a coordinated and finely adjusted control. In the current study, we describe the role of KpfR as a transcriptional regulator of fimbrial expression in K. pneumoniae and its role in the pathogenicity of this bacterium.
KpfR is encoded by kpfR, a gene belonging to the kpf gene cluster that encodes type 1-like fimbriae (Wu et al., 2010). We showed that the Fur regulator binds to a Fur box identified on kpfR gene when complexed with iron and that iron scarcity downregulates kpfR. Since the genes of the kpf cluster are cotranscribed as an operon, we propose that Fur exerts a direct regulation of the kpfR gene and kpf gene cluster by acting as a transcriptional activator. This is not the first report of Fur activating the expression of transcriptional regulators of fimbriae. Wu et al. (2012) showed that the Fur controls the expression of type 3 fimbriae by positively regulating mrkHI, which is involved in the regulation of type 3 fimbriae mrk gene cluster.
The location of this Fur box inside the coding region of the kpfR gene is an intriguing finding. Interestingly, a putative IHF binding site that matches the IHF consensus binding sequence was identified at the intervening sequences between the promoter region of the kpf gene cluster and the Fur box inside kpfR. IHF is a nucleoid-associated protein involved in many cellular processes including site-specific recombination, transcription and DNA replication (Freundlich et al., 1992). IHF acts as a transcriptional coactivator that binds to a consensus sequence (WATCARNNNNTTR, where W is A or T, N is any nucleotide, and R is G or A; Freundlich et al., 1992) and bends the DNA into loops that approximate far distant regulatory elements (Clark et al., 2019). Even though the putative IHF binding site found on kpfR has not been functionally validated in the present study, we can hypothesize that IHF induces a bend in the DNA that brings the Fur regulator close to the regulatory elements on the promoter region of kpf, thus allowing Fur to exert its transcriptional activation on the cluster. Nonetheless, this hypothesis deserves a deeper investigation in future research to elucidate the exact mechanism of regulation played by Fur on the kpf cluster.
Historically described as a transcriptional repressor that utilizes ferrous iron as co-repressor to prevent transcription of target genes by binding on the promoter region of these genes and blocking the entry of RNA polymerase (Escolar et al., 1999), recent studies have shown that Fur can also function as an activator of transcription (Carpenter et al., 2009; Troxell and Hassan, 2013; Seo et al., 2014; Deng et al., 2015). According to Troxell and Hassan (2013), Fur may exert transcriptional activation by (i) indirectly regulating small regulatory RNAs, (ii) binding on regulatory elements that enhance recruitment of the RNA polymerase, or (iii) acting as an antirepressor, in which the binding of Fur-iron complexes on the promoter elements blocks the binding of a transcription repressor (Troxell and Hassan, 2013). Fur activation through “RNAP recruitment” mechanism has been described in H. pylori (Delany et al., 2001) and S. enterica (Teixidó et al., 2011), while Fur activation through “antirepressor” mechanism has been described in E. coli (Nandal et al., 2010) and also in H. pylori (Gilbreath et al., 2012). For instance, the Fur-Fe2+ complex releases the transcription of the ftnA gene, which encodes ferritin in E. coli, by binding to the ftnA promoter and displacing the binding of the global repressor H-NS, resulting in the release of access of the RNA polymerase to the promoter and reverting the repression exerted by H-NS (Nandal et al., 2010). Interestingly, our results show Fur binding to the Fur box on kpfR only when complexed with iron, but this binding does not result in expression modulation of kpfR. On the other hand, Fur is no longer bound to the Fur box in conditions of iron scarcity, and the gene is downregulated. This unaltered expression pattern of kpfR under iron-rich conditions despite Fur-Fe complex binding to the kpfR Fur box suggests Fur exerting transcription activation of kpfR through an “antirepressor” mechanism, similarly to that observed in transcription of the E. coli ftnA gene. In this mode of regulation involving an “antirepressor” mechanism, Fur is not required for the transcription of kpfR gene, but it is required to relieve its repression by a transcriptional repressor, possibly the product of kpfR gene itself, the transcriptional repressor KpfR. This hypothesis is corroborated by the upregulation of kpfR and kpfA genes on the mutant strain kpfR::kanR cells, which suggests that KpfR autoregulates its own expression. Additional experiments, such as fimbrial gene expression in a Fur null mutant strain and promoter-reporter gene fusion assays, are needed to better understand the role of Fur as a transcriptional activator of kpfR.
The K. pneumoniae strain mutant for kpfR gene exhibited phenotypic changes, among which macroscopic changes on colony morphology, a slight dispersion on blood agar plates, greater ability to form biofilm, and capacity to agglutinate yeast cells. Also, the mutant strain presented up-regulation of the type 1-like fimbriae gene clusters fim and kpf and exhibited an increased production of cell surface appendages similar to fimbriae, as revealed by TEM analysis. Unfortunately, in the present study we did not conduct further investigations to identify the nature of these appendages, whether they are type 1 or Kpf fimbriae. Nevertheless, given the up-regulation of both fim and kpf gene clusters by the mutant strain and its ability to agglutinate yeast cells, which specifically indicates the production of type 1-like fimbriae, we may infer that these structures revealed by TEM analysis are probably type 1 and Kpf fimbriae. However, further studies are needed to fully and precisely identify the nature of those appendages.
Strikingly, the hyperfimbriated phenotype seems to provide the mutant strain a kind of rudimentary motility. Although K. pneumoniae is recognized as immobile, the finding of a rudimentary motility in this bacterium is not an unprecedented result. Carabarin-Lima and León-Izurieta (2016) reported the presence of flagellar genes on a K. pneumoniae strain isolated from nosocomial infections and described a swim-like motility phenotype mediated by flagella in these clinical isolates. Besides, the K. pneumoniae genome contains the flk gene that encodes a poorly characterized regulator of flagella biosynthesis. It remains to be clarified whether the rudimentary movement observed in the mutant strain is due to the excess of type 1-like fimbriae production, or if the KpfR regulator plays some role in regulating the expression of flagellar genes.
One of the phenotypic changes observed in the kpfR mutant strain was a drastic reduction in the production of capsular polysaccharide as opposed to the higher expression of fimbriae structures. Reports in the literature describe an inverse effect between the production of fimbriae and capsules, and also suggest that capsule production can inhibit the expression of fimbriae and interferes in fimbriae functionality (Matatov et al., 1999; Schembri et al., 2005; Paczosa and Mecsas, 2016). In this respect, it is suggested that a possible coordinated regulation between fimbriae and capsules occurs through an environmental stimulus not yet known. For instance, Schwan et al. (2005) showed that adherence of uropathogenic E. coli to mannose receptors on bladder epithelial cells via type 1 fimbriae triggers a cross-talk that leads to the down-regulation of capsule production. Regarding K. pneumoniae, a possible cross-regulation between the capsule and fimbrial expression has been speculated, although not yet proven (Schembri et al., 2005; Struve et al., 2008; Schroll et al., 2010). A clue to this apparent cross-regulation of capsule and fimbriae expression in K. pneumoniae came from the work of Huang et al. (2014). These authors showed that the deletion of the cps gene cluster, responsible for capsular polysaccharide biosynthesis, inhibits the expression of genes encoding type 1 and type 3 fimbriae. In conformity to these reports, our findings indicate KpfR as a repressor of fimbriae expression which, when knocked out, leads to activation of fimbriae expression and inhibition of the production of capsules in K. pneumoniae.
Most of the phenotypic changes exhibited by the mutant strain were fully or partially restored on the complemented strain. However, complementation tests rendered unexpected results for capsule production and biofilm formation. Although the complemented strain presented a more than threefold increase in capsule production compared to the mutant strain, this increase was not significantly different, and the production did not reach the levels presented by the wild-type strain. Biofilm formation, which is closely related to capsule and fimbriae expression, also rendered unexpected results, since the complemented strain failed to recovery the wild-type phenotype and, instead, presented similar capacity to form biofilm as exhibited by the mutant strain. This phenotype may be related to the capsular polysaccharide production by these strains. In fact, although capsule plays a key role for K. pneumoniae biofilm establishment and maturation (Balestrino et al., 2008), it is well known that the exacerbated production of capsule shields short bacterial adhesins and inhibits fimbriae assembly and activity (Matatov et al., 1999; Schembri et al., 2004, 2005). Besides, unencapsulated K. pneumoniae mutant strains produce greater amounts and more organized biofilms compared to the wild-type strains (Dzul et al., 2011; Huang et al., 2014). Thus, the reduced production of capsule by the kpfR mutant strain and the complemented strain seems to render these strains a greater capacity to form biofilm. Another possible explanation for the biofilm formation by the complemented strain may be related to the construction of the complemented strains. For complementation tests, we cloned the kpfR gene into an ordinary cloning vector, not a simple-copy vector. Thus, some of the unexpected results may likely be due to enhanced expression of kpfR gene from the vector, probably above the native levels. The effects of this unbalanced expression would be further enhanced when the cloned gene under investigation encodes a transcriptional regulator. Finally, the complexity of the mechanisms that regulate the expression of fimbriae and capsular polysaccharides (CPS), the two main surface components of prokaryotic cells, may also explain the enhanced capacity of the complemented strain to form biofilm. In fact, the production of these structures demands a coordinated and tightly regulated expression of numerous gene clusters and it is under the control of a myriad of regulators, such as RmpA (Hsu et al., 2011), Fur (Lin et al., 2011), IscR (Wu et al., 2014), H-NS (Ares et al., 2016), RcsAB (Peng et al., 2018), KvrA and KvrB (Palacios et al., 2018), and RmpC (Walker et al., 2019) for capsule expression, and FimK (Rosen et al., 2008a), MrkH (Wilksch et al., 2011), Fur (Wu et al., 2012), H-NS (Ares et al., 2016), OmpR (Lin et al., 2018), and IscR (Lin et al., 2017) for fimbriae expression. A possible cross-regulation of these processes and the complexity of this fine-tuned regulation may explain the difficulty in recovering the wild-type phenotype in complemented strains. Regarding biofilm, both type 1 and type 3 fimbriae have been related to its formation (Rosen et al., 2008b; Struve et al., 2009; Stahlhut et al., 2012), but the type 3 are considered the predominant fimbriae in biofilm formation in vitro, especially on abiotic surfaces such as urinary catheters and medical devices (Struve et al., 2009). In addition to that, a cross-regulation of both type 1 and type 3 fimbriae during the biofilm formation by K. pneumoniae has been proposed, so that the lack of type 3 fimbriae is compensated by up-regulation of type 1 fimbriae (Schroll et al., 2010). While we cannot ensure that kpfR is being expressed stably in the complemented strain and, therefore, we cannot exclude the effects of this unbalanced expression, we can infer that the unexpected biofilm formation by the complemented strain may be due either to the type 3 fimbriae exerting a prominent role on this strain or that compensation of fimbriae expression may be occurring. Additionally, some authors also reported partial recovery of biofilm formation by K. pneumoniae complemented strains, especially in mutant strains for transcriptional regulators of type 1 fimbriae, such as FimK (Rosen et al., 2008a) or type 3 fimbriae, such as IscR (Lin et al., 2017) and MrkH (Wilksch et al., 2011). These reports illustrate how difficult complementation of the biofilm formation by mutant strains can be, especially when involving mutant genes that encode transcription factors.
Although the hyperfimbriated phenotype yielded advantageous in in vitro coculture assays—as revealed by the greater efficiency of the mutant strain in adhering, invading, and replicating within eukaryotic host cells—, the increased expression of fimbrial structures proved to be detrimental during in vivo colonization in the murine model of urinary tract infection. In fact, our in vivo experiments showed that the kpfR mutant strain was unable to form intracellular bacterial communities (IBCs), exhibited lower titers in the bladder, and was quickly eliminated by the host. Therefore, by negatively controlling the expression of fimbriae and preventing the bacteria from having a hyperfimbriated phenotype, the KpfR regulator protects K. pneumoniae from being recognized and eliminated by the host immune system.
The loss of resistance and ability of the kpfR mutant strain to survive in the host can be attributed to its hyperfimbriated phenotype, which has made it to be readily recognized by the host defenses and becomes more efficiently and rapidly eliminated in vivo. In fact, while adherence is a crucial step for the pathogen to establish contact with the mucosa and invade and colonize the host tissue, pathogen-host interaction activates host antimicrobial defense, and a key event in this process is the bladder epithelium stimulation following bacterial adherence, which results in cytokines production by the epithelial cells through activation of host Toll-like receptor 4 (Ferraz et al., 2011). As stated by Song and Abraham (2008), several Toll-like receptors have been identified on epithelial cells of the bladder which mediate a powerful immune response. Studies have shown that K. pneumoniae type 3 fimbriae can stimulate an oxidative response in neutrophils (Przondo-Mordarska et al., 1991) and that the expression of fimbriae by K. pneumoniae increases their binding to phagocytes and triggers the phagocytosis, thus leading to bacterial killing (Athamna et al., 1991).
Our results with KpfR repressor bear similarities, but also striking differences, with FimK, another transcriptional regulator of fimbriae well characterized in K. pneumoniae (Rosen et al., 2008a, 2015; Wang et al., 2013). Unlike FimK, which has an EAL domain in its carboxy-terminal region (Rosen et al., 2008a; Wang et al., 2013), KpfR does not have such a domain and, therefore, its mechanism of action should not involve the second messenger cyclic-di-GMP. The effects of KpfR and FimK on repressing the expression of type 1-like fimbriae emphasize the role of both regulators as inhibitory factors of biofilm formation. Also, both kpfR and fimK knockouts result in hyperfimbriated mutant bacteria (Rosen et al., 2008a) and reduced capsule production (Rosen et al., 2015). However, contrary to that observed in fimK mutant strain, the kpfR mutant strain did not have its virulence increased in murine UTI model. FimK attenuates virulence on the urinary tract since lack of fimK resulted in higher bacterial titers, increased numbers of IBCs, and enhanced virulence in the murine urinary tract (Rosen et al., 2008a). In contrast, KpfR seems to be essential for virulence in the urinary tract, because the loss of an active KpfR resulted in lower bladder titers, inability to form IBCs, and attenuated effect on urinary tract virulence. A probable explanation for this is that in the absence of KpfR there is an induction of expression of two gene clusters of type 1-like fimbriae (kpf cluster and fim cluster), whereas FimK regulates only the fim gene cluster. Also, the detrimental effect of FimK is restricted to the urinary tract, since in the respiratory tract it has a potentiating effect on virulence (Rosen et al., 2015). Thus, both FimK and KpfR are repressors of fimbriae expression in Klebsiella pneumoniae. But, while FimK seems crucial for virulence in the respiratory tract, the KpfR revealed to be critical for UTI.
In summary, pathogens employ survival strategies when in hostile environments, and to accomplish that they control the expression of virulence factors as needed. The expression of fimbriae represents an important factor of bacterial pathogenicity and, in this sense, it is of fundamental importance to understand the mechanisms that regulate the expression of this virulence factor. Herein, we described the role KpfR, a new transcriptional repressor of fimbrial expression hitherto not characterized, in the K. pneumoniae pathogenicity. By negatively controlling the expression of fimbriae, KpfR prevents K. pneumoniae from having a hyperfimbriated phenotype and from being eliminated in a murine model of urinary tract infection. Further investigations of the kpfR mutant strain on a murine model of respiratory tract infection will help to further elucidate whether loss of KpfR has an opposite effect according to distinct host niches.
Data Availability Statement
The authors acknowledge that the data presented in this study must be deposited and made publicly available in an acceptable repository, prior to publication. Frontiers cannot accept a manuscript that does not adhere to our open data policies.
Ethics Statement
The animal study was reviewed and approved by Ethics Committee on the Use of Animals in Research of Universidade São Francisco (CIAEP/CONCEA No. 01.0226.2014, protocol number 004.04.2015).
Author Contributions
LFCF conceived the experiments and supervised the execution of the study. AÉIG, TP, CSS, and JAP performed the experiments and data analysis. MD and MLR contributed with data analysis. LFCF, AÉIG, and TP wrote the manuscript. MD and MLR assisted with critical revision of the manuscript and LFCF coordinated its revision. All authors contributed to the manuscript revision and approved the submitted version.
Funding
This work was supported by the São Paulo Research Foundation (FAPESP), grants #2008/11365-1 and #2013/06042-7 to LFCF. AÉIG and CSS were supported by scholarships from Coordination for the Improvement of Higher Education Personnel (CAPES, Ministry of Education of Brazil). TP was awarded a scholarship from FAPESP, grant #2013/13949-9. The publication fees of this study were covered by the financial support from FAPESP (grant #2018/26203-9 to LFCF).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Dr. Klaus Hantke from the University of Tübingen (Germany) for providing the Escherichia coli H1717 strain for the Fur Titration Assays (FURTA). We are grateful to Nathália Maria Gonçalves Siqueira for her technical assistance in the DNA Electrophoretic Mobility Shift Assays (EMSA).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.601921/full#supplementary-material
References
Alcántar-Curiel, M. D., Blackburn, D., Saldaña, Z., Gayosso-Vázquez, C., Iovine, N. M., De la Cruz, M. A., et al. (2013). Multi-functional analysis of Klebsiella pneumoniae fimbrial types in adherence and biofilm formation. Virulence 4, 129–138. doi: 10.4161/viru.22974
Ares, M. A., Fernández-Vázquez, J. L., Rosales-Reyes, R., Jarillo-Quijada, M. D., von Bargen, K., Torres, J., et al. (2016). H-NS nucleoid protein controls virulence features of Klebsiella pneumoniae by regulating the expression of type 3 pili and the capsule polysaccharide. Front. Cell. Infect. Microbiol. 6:13. doi: 10.3389/fcimb.2016.00013
Athamna, A., Ofek, I., Keisari, Y., Markowitz, S., Dutton, G. G., and Sharon, N. (1991). Lectinophagocytosis of encapsulated Klebsiella pneumoniae mediated by surface lectins of guinea pig alveolar macrophages and human monocyte-derived macrophages. Infect. Immun. 59, 1673–1682.
Balestrino, D., Ghigo, J.-M., Charbonnel, N., Haagensen, J. A. J., and Forestier, C. (2008). The characterization of functions involved in the establishment and maturation of Klebsiella pneumoniae in vitro biofilm reveals dual roles for surface exopolysaccharides. Environ. Microbiol. 10, 685–701. doi: 10.1111/j.1462-2920.2007.01491.x
Butcher, J., Sarvan, S., Brunzelle, J. S., Couture, J.-F., and Stintzi, A. (2012). Structure and regulon of Campylobacter jejuni ferric uptake regulator Fur define apo-Fur regulation. Proc. Natl. Acad. Sci. U.S.A. 109:10047. doi: 10.1073/pnas.1118321109
Carabarin-Lima, A., and León-Izurieta, L. (2016). Rocha-Gracia RdC, Castañeda-Lucio M, Torres C, Gutiérrez-Cazarez Z, et al. First evidence of polar flagella in Klebsiella pneumoniae isolated from a patient with neonatal sepsis. J. Med. Microbiol. 65, 729–737. doi: 10.1099/jmm.0.000291
Carpenter, B. M., Whitmire, J. M., and Merrell, D. S. (2009). This is not your mother’s repressor: the complex role of fur in pathogenesis. Infect. Immun. 77, 2590–2601. doi: 10.1128/IAI.00116-09
Choby, J. E., Howard-Anderson, J., and Weiss, D. S. (2020). Hypervirulent Klebsiella pneumoniae – clinical and molecular perspectives. J. Internal Med. 287, 283–300. doi: 10.1111/joim.13007
Clark, D. P., Pazdernik, N. J., and McGehee, M. R. (2019). “Chapter 16 – regulation of transcription in Prokaryotes,” in Molecular Biology, 3rd Edn, eds D. P. Clark, N. J. Pazdernik, and M. R. McGehee (Amsterdam: Academic Cell), 522–559.
Clegg, S., Wilson, J., and Johnson, J. (2011). More than one way to control hair growth: regulatory mechanisms in enterobacteria that affect fimbriae assembled by the chaperone/usher pathway. J. Bacteriol. 193:2081. doi: 10.1128/JB.00071-11
Delany, I., Rappuoli, R., and Scarlato, V. (2004). Fur functions as an activator and as a repressor of putative virulence genes in Neisseria meningitidis. Mol. Microbiol. 52, 1081–1090. doi: 10.1111/j.1365-2958.2004.04030.x
Delany, I., Spohn, G., Rappuoli, R., and Scarlato, V. (2001). The Fur repressor controls transcription of iron-activated and -repressed genes in Helicobacter pylori. Mol. Microbiol. 42, 1297–1309. doi: 10.1046/j.1365-2958.2001.02696.x
Deng, X., Sun, F., Ji, Q., Liang, H., Missiakas, D., Lan, L., et al. (2012). Expression of multidrug resistance efflux pump gene nora is iron responsive in Staphylococcus aureus. J. Bacteriol. 194, 1753–1762. doi: 10.1128/JB.06582-11
Deng, Z., Wang, Q., Liu, Z., Zhang, M., Machado, A. C. D., Chiu, T.-P., et al. (2015). Mechanistic insights into metal ion activation and operator recognition by the ferric uptake regulator. Nat. Commun. 6:7642. doi: 10.1038/ncomms8642
Dzul, S. P., Thornton, M. M., Hohne, D. N., Stewart, E. J., Shah, A. A., Bortz, D. M., et al. (2011). Contribution of the Klebsiella pneumoniae capsule to bacterial aggregate and biofilm microstructures. Appl. Environ. Microbiol. 77, 1777–1782. doi: 10.1128/AEM.01752-10
Elsinghorst, E. A. (1994). Measurement of invasion by gentamicin resistance. Methods Enzymol. 236, 405–420.
Escolar, L., Perez-Martin, J., and de Lorenzo, V. (1999). Opening the iron box: transcriptional metalloregulation by the Fur protein. J. Bacteriol. 181, 6223–6229.
Ferraz, L. F. C., Verde, L. C. L., Vicentini, R., Felício, A. P., Ribeiro, M. L., Alexandrino, F., et al. (2011). Ferric iron uptake genes are differentially expressed in the presence of copper sulfides in Acidithiobacillus ferrooxidans strain LR. Antonie van Leeuwenhoek 99, 609–617. doi: 10.1007/s10482-010-9533-2
Freundlich, M., Ramani, N., Mathew, E., Sirko, A., and Tsui, P. (1992). The role of integration host factor in gene expression in Escherichia coli. Mol. Microbiol. 6, 2557–2563. doi: 10.1111/j.1365-2958.1992.tb01432.x
Gilbreath, J. J., West, A. L., Pich, O. Q., Carpenter, B. M., Michel, S., and Merrell, D. S. (2012). Fur activates expression of the 2-oxoglutarate oxidoreductase genes in Helicobacter pylori. J. Bacteriol. 194:6490. doi: 10.1128/JB.01226-12
Gomes, A. E. I., Stuchi, L. P., Siqueira, N. M. G., Henrique, J. B., Vicentini, R., Ribeiro, M. L., et al. (2018). Selection and validation of reference genes for gene expression studies in Klebsiella pneumoniae using Reverse Transcription Quantitative real-time PCR. Sci. Rep. 8:9001. doi: 10.1038/s41598-018-27420-2
Hannan, T. J., Totsika, M., Mansfield, K. J., Moore, K. H., Schembri, M. A., and Hultgren, S. J. (2012). Host-pathogen checkpoints and population bottlenecks in persistent and intracellular uropathogenic Escherichia coli bladder infection. FEMS Microbiol. Rev. 36, 616–648. doi: 10.1111/j.1574-6976.2012.00339.x
Hornick, D. B., Allen, B. L., Horn, M. A., and Clegg, S. (1992). Adherence to respiratory epithelia by recombinant Escherichia coli expressing Klebsiella pneumoniae type 3 fimbrial gene products. Infect. Immun. 60, 1577–1588.
Hsu, C. R., Lin, T. L., Chen, Y. C., Chou, H. C., and Wang, J. T. (2011). The role of Klebsiella pneumoniae rmpA in capsular polysaccharide synthesis and virulence revisited. Microbiology 157(Pt 12), 3446–3457. doi: 10.1099/mic.0.050336-0
Huang, T.-W., Lam, I., Chang, H.-Y., Tsai, S.-F., Palsson, B. O., and Charusanti, P. (2014). Capsule deletion via a λ-Red knockout system perturbs biofilm formation and fimbriae expression in Klebsiella pneumoniae MGH 78578. BMC Res. Notes 7:13. doi: 10.1186/1756-0500-7-13
Hunstad, D. A., and Justice, S. S. (2010). Intracellular lifestyles and immune evasion strategies of uropathogenic Escherichia coli. Ann. Rev. Microbiol. 64, 203–221. doi: 10.1146/annurev.micro.112408.134258
Johnson, J. G., and Clegg, S. (2010). Role of MrkJ, a phosphodiesterase, in type 3 fimbrial expression and biofilm formation in Klebsiella pneumoniae. J. Bacteriol. 192, 3944–3950. doi: 10.1128/jb.00304-10
Khater, F., Balestrino, D., Charbonnel, N., Dufayard, J. F., Brisse, S., and Forestier, C. (2015). In silico analysis of usher encoding genes in Klebsiella pneumoniae and characterization of their role in adhesion and colonization. PLoS One 10:e0116215. doi: 10.1371/journal.pone.0116215
Lee, C.-R., Lee, J. H., Park, K. S., Jeon, J. H., Kim, Y. B., Cha, C.-J., et al. (2017). Antimicrobial resistance of hypervirulent Klebsiella pneumoniae: epidemiology, hypervirulence-associated determinants, and resistance mechanisms. Front. Cell. Infect. Microbiol. 7:483. doi: 10.3389/fcimb.2017.00483
Lin, C.-T., Lin, T.-H., Wu, C.-C., Wan, L., Huang, C.-F., and Peng, H.-L. (2016). CRP-Cyclic AMP regulates the expression of type 3 fimbriae via cyclic di-GMP in Klebsiella pneumoniae. PLoS One 11:e0162884. doi: 10.1371/journal.pone.0162884
Lin, C. T., Wu, C. C., Chen, Y. S., Lai, Y. C., Chi, C., Lin, J. C., et al. (2011). Fur regulation of the capsular polysaccharide biosynthesis and iron-acquisition systems in Klebsiella pneumoniae CG43. Microbiology 157(Pt 2), 419–429. doi: 10.1099/mic.0.044065-0
Lin, T. H., Chen, Y., Kuo, J. T., Lai, Y. C., Wu, C. C., Huang, C. F., et al. (2018). Phosphorylated OmpR is required for type 3 fimbriae expression in Klebsiella pneumoniae under hypertonic conditions. Front. Microbiol. 9:2405. doi: 10.3389/fmicb.2018.02405
Lin, T.-H., Tseng, C.-Y., Lai, Y.-C., Wu, C.-C., Huang, C.-F., and Lin, C.-T. (2017). IscR regulation of type 3 fimbriae expression in Klebsiella pneumoniae CG43. Front. Microbiol. 8:1984. doi: 10.3389/fmicb.2017.01984
Lin, T. L., Yang, F. L., Yang, A. S., Peng, H. P., Li, T. L., Tsai, M. D., et al. (2012). Amino acid substitutions of MagA in Klebsiella pneumoniae affect the biosynthesis of the capsular polysaccharide. PLoS One 7:e46783. doi: 10.1371/journal.pone.0046783
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Losensky, G., Vidakovic, L., Klingl, A., Pfeifer, F., and Fröls, S. (2015). Novel pili-like surface structures of Halobacterium salinarum strain R1 are crucial for surface adhesion. Front. Microbiol. 6:755. doi: 10.3389/fmicb.2014.00755
Matatov, R., Goldhar, J., Skutelsky, E., Sechter, I., Perry, R., Podschun, R., et al. (1999). Inability of encapsulated Klebsiella pneumoniae to assemble functional type 1 fimbriae on their surface. FEMS Microbiol. Lett. 179, 123–130.
Murphy, C. N., Mortensen, M. S., Krogfelt, K. A., and Clegg, S. (2013). Role of Klebsiella pneumoniae type 1 and type 3 fimbriae in colonizing silicone tubes implanted into the bladders of mice as a model of catheter-associated urinary tract infections. Infect. Immun. 81, 3009–3017. doi: 10.1128/IAI.00348-13
Nandal, A., Huggins, C. C. O., Woodhall, M. R., McHugh, J., Rodríguez-Quiñones, F., Quail, M. A., et al. (2010). Induction of the ferritin gene (ftnA) of Escherichia coli by Fe2+–Fur is mediated by reversal of H-NS silencing and is RyhB independent. Mol. Microbiol. 75, 637–657. doi: 10.1111/j.1365-2958.2009.06977.x
Oelschlaeger, T. A., and Tall, B. D. (1997). Invasion of cultured human epithelial cells by Klebsiella pneumoniae isolated from the urinary tract. Infect. Immun. 65, 2950–2958.
O’Toole, G. A., and Kolter, R. (1998). Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signalling pathways: a genetic analysis. Mol. Microbiol. 28, 449–461.
Paczosa, M. K., and Mecsas, J. (2016). Klebsiella pneumoniae: going on the offense with a strong defense. Microbiol. Mol. Biol. Rev. 80, 629–661. doi: 10.1128/MMBR.00078-15
Palacios, M., Miner, T. A., Frederick, D. R., Sepulveda, V. E., Quinn, J. D., Walker, K. A., et al. (2018). Identification of two regulators of virulence that are conserved in Klebsiella pneumoniae classical and hypervirulent strains. mBio 9:e01443-18. doi: 10.1128/mBio.01443-18
Peng, D., Li, X., Liu, P., Zhou, X., Luo, M., Su, K., et al. (2018). Transcriptional regulation of galF by RcsAB affects capsular polysaccharide formation in Klebsiella pneumoniae NTUH-K2044. Microbiol. Res. 216, 70–78. doi: 10.1016/j.micres.2018.08.010
Pi, H., and Helmann, J. D. (2017). Sequential induction of Fur-regulated genes in response to iron limitation in Bacillus subtilis. Proc. Natl. Acad. Sci. U.S.A. 114, 12785–12790. doi: 10.1073/pnas.1713008114
Podschun, R., and Ullmann, U. (1998). Klebsiella spp. as nosocomial pathogens: epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin. Microbiol. Rev. 11, 589–603.
Przondo-Mordarska, A., Ko, H. L., Gamian, A., and Pulverer, G. (1991). Chemiluminescence response of human polymorphonuclear leukocytes induced by purified, latex attached Klebsiella fimbriae. Zentralblatt Bakteriol. 275, 521–529. doi: 10.1016/S0934-8840(11)80174-X
Reis, L. O., Sopena, J. M. G., Fávaro, W. J., Martin, M. C., Simão, A. F. L., dos Reis, R. B., et al. (2011). Anatomical features of the urethra and urinary bladder catheterization in female mice and rats. An essential translational tool. Acta Cirúrgica Brasileira 26, 106–110. doi: 10.1590/S0102-86502011000800019
Rodriguez-Villar, S., Fife, A., Baldwin, C., and Warne, R. R. (2019). Antibiotic-resistant hypervirulent Klebsiella pneumoniae causing community-acquired liver abscess: an emerging disease. Oxford Med. Case Rep. 2019:omz032. doi: 10.1093/omcr/omz032
Roe, A. J., Currie, C., Smith, D. G. E., and Gally, D. L. (2001). Analysis of type 1 fimbriae expression in verotoxigenic Escherichia coli: a comparison between serotypes O157 and O26. Microbiology 147, 145–152. doi: 10.1099/00221287-147-1-145
Rosen, D. A., Hilliard, J. K., Tiemann, K. M., Todd, E. M., Morley, S. C., and Hunstad, D. A. (2015). Klebsiella pneumoniae FimK promotes virulence in murine pneumonia. J. Infect. Dis. 213, 649–658. doi: 10.1093/infdis/jiv440
Rosen, D. A., Hooton, T. M., Stamm, W. E., Humphrey, P. A., and Hultgren, S. J. (2007). Detection of intracellular bacterial communities in human urinary tract infection. PLoS Med. 4:e40329. doi: 10.1371/journal.pmed.0040329
Rosen, D. A., Pinkner, J. S., Jones, J. M., Walker, J. N., Clegg, S., and Hultgren, S. J. (2008a). Utilization of an intracellular bacterial community pathway in Klebsiella pneumoniae urinary tract infection and the effects of FimK on type 1 pilus expression. Infect. Immun. 76, 3337–3345. doi: 10.1128/IAI.00090-08
Rosen, D. A., Pinkner, J. S., Walker, J. N., Elam, J. S., Jones, J. M., and Hultgren, S. J. (2008b). Molecular variations in Klebsiella pneumoniae and Escherichia coli FimH affect function and pathogenesis in the urinary tract. Infect. Immun. 76, 3346–3356. doi: 10.1128/IAI.00340-08
Rozen, S., and Skaletsky, H. (2000). Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 132, 365–386.
Schembri, M. A., Blom, J., Krogfelt, K. A., and Klemm, P. (2005). Capsule and fimbria interaction in Klebsiella pneumoniae. Infect. Immu. 73, 4626–4633. doi: 10.1128/IAI.73.8.4626-4633.2005
Schembri, M. A., Dalsgaard, D., and Klemm, P. (2004). Capsule shields the function of short bacterial adhesins. J. Bacteriol. 186, 1249–1257. doi: 10.1128/jb.186.5.1249-1257.2004
Schroll, C., Barken, K. B., Krogfelt, K. A., and Struve, C. (2010). Role of type 1 and type 3 fimbriae in Klebsiella pneumoniae biofilm formation. BMC Microbiol. 10:179. doi: 10.1186/1471-2180-10-179
Schwan, W. R., Beck, M. T., Hultgren, S. J., Pinkner, J., Woolever, N. L., and Larson, T. (2005). Down-regulation of the kps region 1 capsular assembly operon following attachment of Escherichia coli type 1 fimbriae to D-mannose receptors. Infect. Immun. 73, 1226–1231. doi: 10.1128/IAI.73.2.1226-1231.2005
Seo, S. W., Kim, D., Latif, H., O’Brien, E. J., Szubin, R., and Palsson, B. O. (2014). Deciphering Fur transcriptional regulatory network highlights its complex role beyond iron metabolism in Escherichia coli. Nat. Commun. 5:4910. doi: 10.1038/ncomms5910
Solovyev, V. V., and Solovyev, A. S. (2011). “Automatic annotation of microbial genomes and metagenomic sequences,” in Metagenomics and its Applications in Agriculture, Biomedicine and Environmental Studies, ed. R. W. Li (Hauppauge, NY: Nova Science Publishers), 61–78.
Song, J., and Abraham, S. N. (2008). TLR-mediated immune responses in the urinary tract. Curr. Opin. Microbiol. 11, 66–73. doi: 10.1016/j.mib.2007.12.001
Stahlhut, S. G., Struve, C., Krogfelt, K. A., and Reisner, A. (2012). Biofilm formation of Klebsiella pneumoniae on urethral catheters requires either type 1 or type 3 fimbriae. FEMS Immunol. Med. Microbiol. 65, 350–359. doi: 10.1111/j.1574-695X.2012.00965.x
Struve, C., Bojer, M., and Krogfelt, K. A. (2008). Characterization of Klebsiella pneumoniae type 1 fimbriae by detection of phase variation during colonization and infection and impact on virulence. Infect. Immun. 76, 4055–4065. doi: 10.1128/IAI.00494-08
Struve, C., Bojer, M., and Krogfelt, K. A. (2009). Identification of a conserved chromosomal region encoding Klebsiella pneumoniae type 1 and type 3 fimbriae and assessment of the role of fimbriae in pathogenicity. Infect. Immun. 77, 5016–5024. doi: 10.1128/IAI.00585-09
Teixidó, L., Carrasco, B., Alonso, J. C., Barbé, J., and Campoy, S. (2011). Fur activates the expression of Salmonella enterica pathogenicity island 1 by directly interacting with the hilD operator in vivo and in vitro. PLoS One 6:e19711. doi: 10.1371/journal.pone.0019711
Thai, K. H., Thathireddy, A., and Hsieh, M. H. (2010). Transurethral induction of mouse urinary tract infection. J. Vis. Exp. 42:2070. doi: 10.3791/2070
Tinker, J. K., and Clegg, S. (2000). Characterization of FimY as a coactivator of type 1 fimbrial expression in Salmonella enterica serovar typhimurium. Infect. Immun. 68, 3305–3313. doi: 10.1128/IAI.68.6.3305-3313.2000
Troxell, B., and Hassan, H. M. (2013). Transcriptional regulation by Ferric Uptake Regulator (Fur) in pathogenic bacteria. Front. Cell. Infect. Microbiol. 3:59. doi: 10.3389/fcimb.2013.00059
Turton, J. F., Baklan, H., Siu, L. K., Kaufmann, M. E., and Pitt, T. L. (2008). Evaluation of a multiplex PCR for detection of serotypes K1, K2 and K5 in Klebsiella sp. and comparison of isolates within these serotypes. FEMS Microbiol. Lett. 284, 247–252. doi: 10.1111/j.1574-6968.2008.01208.x
Walker, K. A., Miner, T. A., Palacios, M., Trzilova, D., Frederick, D. R., Broberg, C. A., et al. (2019). A Klebsiella pneumoniae regulatory mutant has reduced capsule expression but retains hypermucoviscosity. mBio 10:e00089-19. doi: 10.1128/mBio.00089-19
Wang, Z. C., Huang, C. J., Huang, Y. J., Wu, C. C., and Peng, H. L. (2013). FimK regulation on the expression of type 1 fimbriae in Klebsiella pneumoniae CG43S3. Microbiology 159(Pt 7), 1402–1415. doi: 10.1099/mic.0.067793-0
Wilksch, J. J., Yang, J., Clements, A., Gabbe, J. L., Short, K. R., Cao, H., et al. (2011). MrkH, a novel c-di-GMP-dependent transcriptional activator, controls Klebsiella pneumoniae biofilm formation by regulating type 3 fimbriae expression. PLoS Pathog. 7:e1002204. doi: 10.1371/journal.ppat.1002204
Wu, C. C., Huang, Y. J., Fung, C. P., and Peng, H. L. (2010). Regulation of the Klebsiella pneumoniae Kpc fimbriae by the site-specific recombinase KpcI. Microbiology 156(Pt 7), 1983–1992. doi: 10.1099/mic.0.038158-0
Wu, C. C., Lin, C. T., Cheng, W. Y., Huang, C. J., Wang, Z. C., and Peng, H. L. (2012). Fur-dependent MrkHI regulation of type 3 fimbriae in Klebsiella pneumoniae CG43. Microbiology 158(Pt 4), 1045–1056. doi: 10.1099/mic.0.053801-0
Wu, C. C., Wang, C. K., Chen, Y. C., Lin, T. H., Jinn, T. R., and Lin, C. T. (2014). IscR regulation of capsular polysaccharide biosynthesis and iron-acquisition systems in Klebsiella pneumoniae CG43. PLoS One 9:e107812. doi: 10.1371/journal.pone.0107812
Yeh, K.-S., Tinker, J. K., and Clegg, S. (2002). FimZ binds the Salmonella typhimurium fimA promoter region and may regulate its own expression with FimY. Microbiol. Immunol. 46, 1–10. doi: 10.1111/j.1348-0421.2002.tb02670.x
Keywords: Klebsiella pneumonia, transcriptional regulation, fimbriae, adherence, biofilms, coculture, urinary tract infection, host-microbe interactions
Citation: Gomes AÉI, Pacheco T, Santos CdSd, Pereira JA, Ribeiro ML, Darrieux M and Ferraz LFC (2021) Functional Insights From KpfR, a New Transcriptional Regulator of Fimbrial Expression That Is Crucial for Klebsiella pneumoniae Pathogenicity. Front. Microbiol. 11:601921. doi: 10.3389/fmicb.2020.601921
Received: 02 September 2020; Accepted: 03 December 2020;
Published: 21 January 2021.
Edited by:
Axel Cloeckaert, Institut National de Recherche pour l’Agriculture, l’Alimentation et l’Environnement (INRAE), FranceReviewed by:
Muhammad Ammar Zafar, Wake Forest School of Medicine, United StatesMiguel A. Ares, Mexican Social Security Institute (IMSS), Mexico
Copyright © 2021 Gomes, Pacheco, Santos, Pereira, Ribeiro, Darrieux and Ferraz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lúcio Fábio Caldas Ferraz, bHVjaW8uZmVycmF6QHVzZi5lZHUuYnI=
†These authors have contributed equally to this work and share first authorship